

Abstract
Background
Salmonella have potential as anticancer therapeutic because of their innate tumor specificity. In clinical studies, this specificity has been hampered by heterogeneous responses. Understanding the mechanisms that control tumor colonization would enable the design of more robust therapeutic strains. Two mechanisms that could affect tumor colonization are intracellular accumulation and intratumoral motility. Both of these mechanisms have elements that are controlled by the master motility regulator flhDC. We hypothesized that 1) overexpressing flhDC in Salmonella increases intracellular bacterial accumulation in tumor cell masses, and 2) intracellular accumulation of Salmonella drives tumor colonization in vitro.
Methods
To test these hypotheses, we transformed Salmonella with genetic circuits that induce flhDC and express green fluorescent protein after intracellular invasion. The genetically modified Salmonella was perfused into an in vitro tumor-on-a-chip device. Time-lapse fluorescence microscopy was used to quantify intracellular and colonization dynamics within tumor masses. A mathematical model was used to determine how these mechanisms are related to each other.
Results
Overexpression of flhDC increased intracellular accumulation and tumor colonization 2.5 and 5 times more than control Salmonella, respectively (P < 0.05). Non-motile Salmonella accumulated in cancer cells 26 times less than controls (P < 0.001). Minimally invasive, ΔsipB, Salmonella colonized tumor masses 2.5 times less than controls (P < 0.05). When flhDC was selectively induced after penetration into tumor masses, Salmonella both accumulated intracellularly and colonized tumor masses 2 times more than controls (P < 0.05). Mathematical modeling of tumor colonization dynamics demonstrated that intracellular accumulation increased retention of Salmonella in tumors by effectively causing the bacteria to bind to cancer cells and preventing leakage out of the tumors. These results demonstrated that increasing intracellular bacterial density increased overall tumor colonization and that flhDC could be used to control both.
Conclusions
This study demonstrates a mechanistic link between motility, intracellular accumulation and tumor colonization. Based on our results, we envision that therapeutic strains of Salmonella could use inducible flhDC to drive tumor colonization. More intratumoral bacteria would enable delivery of higher therapeutic payloads into tumors and would improve treatment efficacy.
Electronic supplementary material
The online version of this article (10.1186/s40425-018-0490-z) contains supplementary material, which is available to authorized users.
Keywords: Salmonella, Bacterial cancer therapy, Cancer therapy, Intracellular invasion, Intracellular cancer therapy
Introduction
Effective tumor colonization is essential for bacterial anti-cancer therapy. With poor colonization, insufficient treatment is delivered and the tumor response is hampered. For bacterial therapy, the tumor density is controlled more by the rate of colonization than the administered dose [1]. However, the mechanisms that control colonization are poorly understood. It has been well established that after intravenous injection into mice, Salmonella colonizes tumor tissue at ratios greater than 10,000:1 compared to other organs in the body [2]. It is this tumor specificity that makes Salmonella-based therapy particularly attractive as a targeted delivery agent [3]. Unfortunately, clinical trials showed that tumor colonization in humans was not sufficient to induce a lasting response [4]. Therefore, understanding and controlling the mechanisms that drive bacterial tumor colonization could greatly improve bacterial tumor therapy.
Two mechanisms that could affect tumor colonization are intratumoral motility and intracellular accumulation. We have previously shown that bacterial motility plays a critical role in the accumulation of Salmonella in tumors [5–7]. Upregulating motility by swim-plate selection increases distal tumor colonization of the bacteria [6, 8] and altering chemotactic sensing increases bacterial penetration into tumor masses [7–9]. Salmonella motility is controlled by the master regulator flhDC [10–12]. The flhDC protein complex regulates expression of the functional flagellar components [13]. This regulator is one of the most tightly regulated transcription factors within bacteria [14–19]. Flagella-dependent motility is downregulated under nutrient deprivation in Salmonella, which helps Salmonella survive intracellularly where there is limited availability of nutrients [20].
Intracellular invasion and growth are important mechanisms that could also affect Salmonella colonization of tumors. Salmonella have two type three secretion systems, T3SS1 and T3SS2, that promote invasion, survival, and growth inside epithelial cells [21]. Other Salmonella invasion systems include the Rck system, which invades cells by binding to epidermal growth factor receptor [22]. In the gut, Salmonella use these systems to invade and grow inside intestinal cells [23]. Disabling T3SS2 limits the ability of Salmonella to inhibit tumor growth [24]. When T3SS2 genes are deleted by transposon insertion, bacterial accumulation in the spleen is reduced [25]. After serial passaging in mice, Salmonella with increased intracellular invasion had enhanced persistence [26]. We have seen similar effects in tumor cell masses in vitro. Compared to K-12 E. Coli that is T3SS deficient, Salmonella had considerably greater colonization [5].
The two Salmonella secretions systems have distinct functions. T3SS1 initiates invasion into epithelial cells and T3SS2 enables intracellular growth and survival [21]. Both systems are composed of a needle apparatus that spans the inner and outer membranes, and the peptidoglycan layer [27]. Effector proteins are injected into the mammalian cells through the T3SS1 [27, 28]. Secretion of T3SS1 effectors into mammalian cell cytoplasm is required for T3SS dependent intracellular invasion of Salmonella [29]. Once injected, these effectors cause a rearrangement of the mammalian actin cytoskeleton and endocytosis of Salmonella [30, 31]. One essential effector protein is sipB. When knocked out, Salmonella cannot invade using T3SS1 [32]. When Salmonella have internalized, the bacteria modify the endocytic vacuole by secreting T3SS2 effectors [33–35]. These modifications confer protection to the bacteria and enable intracellular growth and survival [36, 37]. The T3SS-dependent intracellular invasion and survival of Salmonella confers protection against extracellular clearance mechanisms, like compliment and attack by macrophages and neutrophils [23, 38]. A non-functional T3SS2 apparatus impairs in vivo colonization and anti-tumor efficacy of Salmonella [24, 25], indicating the importance of intracellular growth for survival of bacteria in vivo.
Flagella-dependent motility and intracellular invasion are not regulated independently. Rather, both of these systems are intertwined and there is a complex feedback between them [39, 40]. Increasing bacterial motility also increases intracellular invasion [41]. The flhDC transcriptional complex controls elements of both motility and cellular invasion. In addition to controlling expression of motility genes, it directly controls the expression of the dual regulatory element, fliZ. FliZ controls both flagellar hook assembly and upregulates the transcription factor hilD [39–41]. HilD expression directly upregulates T3SS1 expression and intracellular invasion [39, 40]. The systems are further connected because flagella can act as physical cell surface sensors to determine the optimal extracellular location to initiate invasion [42]. These systems are connected in part because the T3SS evolved from the flagellar type three secretion system (fT3SS), which is used to assemble functional flagella [43, 44]. The co-regulation of motility and intracellular invasion further supports the idea that both of these phenomena are important for bacterial tumor colonization.
In addition to affecting intracellular invasion, flagella-dependent motility also affects the intracellular lifestyle of Salmonella. Immediately after invasion the majority of Salmonella reside in intracellular vacuoles. A small but significant fraction of the intracellular bacteria escape from the vacuoles into the cytosol [45–47]. Some cytosolic bacteria are degraded by host ubiquitination machinery [48–52]. Those that escape degradation replicate rapidly and are extruded from the cell [45]. The T3SS1 system and functional flagella play important roles in the escape from the vacuole and the hyper-replication [45–47]. After extrusion, the bacteria are primed for reinvasion because of the expression of flagella and SPI-I invasion genes [45, 46].
The goal of this study was to measure the effect of intracellular accumulation on bacterial tumor colonization and quantify the interplay between intracellular accumulation and motility. The interaction of these mechanisms has not been previously studied in relation to using bacteria for cancer therapy. We hypothesized that 1) overexpressing flhDC in Salmonella increases intracellular accumulation in tumor cell masses, and 2) intracellular accumulation of Salmonella drives tumor colonization in vitro. To test these hypotheses, Salmonella were transformed with genetic circuits that induce flhDC and express green fluorescent protein (GFP) after cell invasion. Genetically modified Salmonella were perfused into a microfluidic tumor-on-a-chip device to assess colonization and invasion using time-lapse fluorescence microscopy. The potential to use flhDC as a bispecific switch to increase tumor colonization was determined by inducing expression after initial penetration. A mathematical model was used to investigate why intracellular invasion and growth improved tumor colonization of Salmonella. Controlling Salmonella invasion into cells will increase overall tumor colonization and has the potential to make these therapeutic bacteria more effective in the clinic.
Materials and Methods
Bacterial Strains and Plasmid Construction
Eight strains of Salmonella Enterica serovar Typhimurium were used throughout the experiments (Table 1). The control strain (Sal) was based on an attenuated therapeutic strain of Salmonella (VNP20009) that has three deletions, ΔmsbB, ΔpurI, and Δxyl, that eliminate most toxicities in vivo. The background strain was transformed with a plasmid containing two gene circuits, Plac/DsRed and PSSEJ/GFP, that constitutively express DsRed and express GFP after intracellular invasion (Table 1; Additional file 1: Figure S1-A). The constitutive lac DsRed gene circuit was created by adding the wild-type lac promoter and a ribosomal binding site (AAGGAG) to the 5’ end of the forward DsRed primer. The SSEJ promoter was copied by PCR from VNP20009 genomic DNA using the following primers: forward-ACATGTCACATAAAACACTAGCACTTTAGC and reverse- TCTAGACCTCCTTACTTTATTAAACACGCT. The second strain, F-Sal, was transformed with a plasmid that contains a third gene circuit that enables induction of flhDC with arabinose (Table 1; Additional file 1: Figure S1-B). PCR was used to amplify the flhDC genes from Salmonella genomic DNA using the following primers: forward-AAAAAACCATGGGTTAATAAAAGGAGGAATATATATGCATACATCCGAGTTGCTAAAACA and reverse- AAAAAACTCGAGAAAAATTAAACAGCCTGTTCGATCTGTTCAT. The PCR product and PBAD-his-myc plasmid (Invitrogen, Carlsbad, CA) were digested with NcoI and XhoI and ligated with T4 DNA ligase. The flhDC expression cassette, which includes the AraC regulator and PBAD controlled flhDC, was amplified with PCR and combined with a plasmid containing SSEJ-GFP and Lac-DsRed using Gibson Assembly. Both S-Sal, which has a sipB deletion, and the ΔflgE strain were generated using lambda red recombination [53]. When the flagellar hook (flgE) is deleted, Salmonella are unable to produce functional flagella and are non-motile [54]. The S-Sal strain (strain three) was transformed with the plasmid containing Plac/DsRed and PSSEJ/GFP (Table 1; Additional file 1: Figure S1-A). The fourth strain, FS-Sal, was transformed with a plasmid that contains inducible flhDC (PBAD/flhDC), constitutive DsRed expression (Plac/DsRed) and intracellular GFP expression (PSSEJ/GFP) in a ΔsipB background (Table 1; Additional file 1: Figure S1-B). A second control Salmonella strain (strain five) was transformed with a plasmid containing Plac/GFP to constitutively express GFP (Table 1; Additional file 1: Figure S1-C). The constitutive lac GFP gene circuit was created similarly to the lac DsRed circuit, by adding the wild-type lac promoter and a ribosomal binding site (AAGGAG) to the 5’ end of the forward GFP primer. The sixth strain, Salmonella+pflhDC, expresses GFP constitutively and flhDC upon induction with arabinose (Table 1; Additional file 1: Figure S1-D). The seventh strain, ΔflgE, is non-motile and expresses GFP constitutively (Table 1; Additional file 1: Figure S1-C). The eighth strain, ΔflgE+pflhDC, expresses GFP constitutively and flhDC upon induction with arabinose (Table 1; Additional file 1: Figure S1-D). All cloning was performed with DH5α E. Coli (New England Biolabs, Ipswich, MA) and all plasmids contained a ColE1 origin and either chloramphenicol or ampicillin resistance (Additional file 1: Figure S1). Salmonella were transformed by electroporation. All cloning reagents, buffer reagents, and primers were from New England Biolabs, Fisher Scientific (Hampton, NH), and Invitrogen, (Carlsbad, CA), respectively, unless otherwise noted.
Table 1.
Salmonella strains and plasmids
| Strain | Background | Genetic circuits | Description |
|---|---|---|---|
| 1. Sal |
ΔmsbB, ΔpurI, Δxyl (VNP20009) |
P lac /DsRed P SSEJ /GFP | Constitutive DsRed Intracellularly inducible GFP Additional file 1: Figure S1-A |
| 2. F-Sal | Sal | P lac /DsRed P SSEJ /GFP P BAD /flhDC | Arabinose Inducible flhDC Constitutive DsRed Intracellularly inducible GFP Additional file 1: Figure S1-B |
| 3. S-Sal | ΔsipB Sal | P lac /DsRed P SSEJ /GFP | Minimally Intracellularly Invasive Constitutive DsRed Intracellularly inducible GFP Additional file 1: Figure S1-A |
| 4. FS-Sal | ΔsipB Sal | P lac /DsRed P SSEJ /GFP P BAD /flhDC | Arabinose Inducible flhDC Minimally Intracellularly Invasive Constitutive DsRed Intracellularly inducible GFP Additional file 1: Figure S1-B |
|
5. Salmonella
(control) |
ΔmsbB, ΔpurI, Δxyl | P lac /GFP | Constitutive GFP Additional file 1: Figure S1-C |
| 6. Salmonella + pflhDC | ΔmsbB, ΔpurI, Δxyl |
P
lac
/GFP
P BAD /flhDC |
Arabinose inducible flhDC Constitutive GFP Additional file 1: Figure S1-D |
| 7. ΔflgE | ΔmsbB, ΔpurI, Δxyl ΔflgE | P lac /GFP | Non-motile Constitutive GFP Additional file 1: Figure S1-C |
| 8. ΔflgE+pflhDC | ΔmsbB, ΔpurI, Δxyl ΔflgE |
P
lac
/GFP
P BAD /flhDC |
Arabinose inducible flhDC Non-motile Constitutive GFP Additional file 1: Figure S1-D |
Cell Culture
MCF7 breast carcinoma cells and LS174T colorectal carcinoma cells (ATCC, Manassas, VA) were maintained in DMEM (Dulbecco's Modified Eagle Medium; Sigma Aldrich, St. Louis, MO) with 1 g/L glucose, 3.7 g/L sodium bicarbonate (pH 7.4) and 10% FBS using standard cell culture techniques. Between passages of LS174T cells, single cell suspensions were transferred to PMMA coated cell culture flasks (2 g/L PMMA in 100% ethanol, dried before use) in order to produce spheroids.
Fabrication and Operation of Microfluidic Devices
Photolithography was used to make silicon wafer masters as previously described [55]. Two silicon wafers were made: One silicon wafer was used to make the pneumatic valve layer (layer 1). The other wafer was to make the media perfusion layer (layer 2). The fabrication of multi-layer tumor-on-a-chip devices was based on a previous method [56]. The microfluidic device was fabricated in two parts. Layer 1 was created by mixing 9 parts of Sylgard 184 PDMS (Ellsworth Adhesives, Wilmington, MA) with 1 part of curing agent and poured onto the pneumatic valve layer silicon master wafer. Layer 2 was created by mixing 15 parts of PDMS with 1 part (weight by mass) of curing agent and spun coat onto the media perfusion silicon wafer to a height of 200 μm. Both layers of PDMS were cured at 65 °C for 1.5 h and layer 1 was aligned on top of layer 2. Both layers were cured together at 95 °C for 1 h. Holes were punched in the PDMS layers to receive fluidic and control tubing. The PDMS layers were bonded to a glass slide by plasma treatment (Harrick Plasma Cleaner). The valves were pneumatically actuated before bonding to prevent the valve from sealing. Devices were taped to a microscope stage adaptor and inlet and outlet tubes were inserted. A 10% bleach solution was perfused at 3 μl/min throughout the device for 2 h followed by 70% ethanol for 1 h. The device was prepared for spheroid loading by perfusing for 1 h with DMEM with 1 g/L glucose, 20 mM HEPES (pH 7.4), 10% FBS and 33 μg/ml chloramphenicol (henceforth referred to as DMEM-HEPES-chlor). For all experiments, ~300 μm diameter LS174T spheroids were positioned into a microfluidic device and equilibrated in DMEM-HEPES-chlor for 6 h at a flow rate of 3 μl/min. Some spheroids were damaged in the insertion process and these cell masses were not included in the image analysis.
Quantifying Intracellular Invasion and Colonization of Salmonella in a Tumor-on-a-chip
Four experiments were performed with tumor-on-a-chip device to quantify colonization and intracellular accumulation for (1) induced F-Sal compared to Sal, (2) FS-Sal compared to S-Sal, (3) S-Sal compared to Sal, and (4) for intratumoral induction of F-Sal compared to Sal. Salmonella strains were grown in LB with chloramphenicol (33 μg/ml) to a density of approximately 250 million CFU/ml. Bacteria were resuspended in DMEM-HEPES-chlor at a density of 10 million CFU/ml. The bacterial suspension was perfused into the tumor-on-a-chip device for 1h at a flowrate of 3 μl/min followed by bacteria-free DMEM-HEPES-chlor at the same flowrate for 48 h. In experiments one and two, the F-Sal and FS-Sal conditions contained 0.4% arabinose to induce flhDC. Flowing bacteria-fee medium prevents over growth in the flow channel and mimics clearance in vivo. For experiment four, the procedure was the same (bacterial perfusion for 1 h, followed by perfusion with bacteria-free medium), except that after 11 h, medium containing 0.4% arabinose was perfused into the device to induce flhDC intratumorally.
Transmitted and fluorescent images (480/525 excitation/emission for GFPmut3 and 525/590 for DsRed) of tumor masses were acquired every hour with an Olympus IX71 or a Zeiss Axio Observer Z.1 microscope. Time lapse microscopy images of each tumor mass were cropped using the rectangular cropping tool in ImageJ and were analyzed in Matlab. Each image was background subtracted. Fluorescent intensities of ten spatially equal sections of each tumor mass were averaged to quantify bacterial profiles for each time point. Overall bacterial density as a function of time was determined by averaging fluorescent intensities for entire tumor masses per time point. Red fluorescence was used to calculate total bacterial colonization and green fluorescence was used to calculate intracellular bacterial density. Each experiment was normalized by dividing every calculated average fluorescence intensity by the highest fluorescent intensity observed, which occurred during the last time point.
Quantifying Aqueous Motility of Salmonella
Aqueous motility was determined by growing flhDC inducible Salmonella in 0.4% arabinose. Twenty microliters of 400 million CFU/ml of either flhDC induced or control Salmonella was placed between a coverslip and a glass slide. Transmitted light microscopy images were taken every 0.68 seconds for approximately 30 seconds. The automated particle tracking plugin in ImageJ, Trackmate, was used to analyze bacterial swimming velocity. Aqueous velocity histograms were generated by binning the fraction of total bacteria into three velocity categories: 0-15 μm/s, 15-30 μm/s and >30 μm/s. Motility assays were performed in triplicate.
Quantifying Intracellular Invasion and Growth inside MCF7 Cells in Monolayer
Intracellular invasion of Salmonella was quantified by growing in LB and adding to monolayer cultures of MCF7 cancer cells. Four strains were used to quantify the dependence on flhDC expression and flagella formation: control Salmonella, Salmonella+pflhDC, ΔflgE, ΔflgE+pflhDC. Two strains were used to show the intracellular specificity of the PSSEJ promoter and the dependence on T3SS: Sal and S-Sal, using a modified gentamycin protection assay. Each strain was grown in LB to a density of 5 × 108 CFU/ml and added to 6-well plates of MCF7 cells at a density of 5 × 106 CFU/ml. After two hours of incubation, each well was washed ten times with one milliliter of phosphate buffered saline. DMEM with 20 mM HEPES and 40 μg/ml gentamycin was added to each well to remove residual extracellular bacteria. For two hours following the addition of gentamycin, the cultures were observed microscopically to assess the effectiveness of the PBS washes to remove extracellular bacteria. The few remaining extracellular bacteria were observed over this period to ensure that they were eliminated by the gentamycin treatment. After two hours, intracellular Salmonella were imaged over time at 10X magnification with fluorescence microscopy. After 18 hours, bacterial invasion was quantified by randomly identifying 20 cells in each culture and counting the fraction of cells that contained intracellular Salmonella, as indicated by GFP fluorescence.
A similar invasion protocol was used to calculate the intracellular growth rate of Salmonella. Both control Salmonella and Salmonella+pflhDC constitutively expressed GFP (Table 1). Time lapse fluorescence microscopy was used to quantify the fluorescence from Plac/GFP Salmonella inside MCF7 cells over time. Salmonella density was determined by multiplying the average intensity by the area of all intracellular bacteria within a cell, as a function of time. It was assumed that the amount of GFP produced per bacterium was constant over time. Only MCF7 cells containing bacteria and that did not divide for a six hour interval were used. Intracellular growth rate was calculated by fitting an exponential growth function to the intracellular bacterial density.
Mathematical Modeling
A mathematical model was created to interpret the spatiotemporal dynamics of bacterial dispersion, growth and invasion in tumor masses. This model was based on a previous model of bacterial growth in tumor tissue [57].
| 1 |
| 2 |
| 3 |
The coupled PDE model incorporated a balance on extracellular (eq. 1) and intracellular (eq. 2) bacteria. The balance for extracellular bacteria includes the effects of dispersion [], chemotaxis [], growth [μgcex], and invasion [μinvcexθ]. The intracellular balance includes the effects of intracellular growth [μg, incin] and invasion [μinvcexθ]. The initial and boundary conditions (eq. 3) state that (1) there were no intracellular or extracellular bacteria initially within the tumor mass; (2) the flux into or out of the tumor mass was equal to the flux in the supply channel; and (3) there was no flux at the distal (x = 1) boundary. The supply of extracellular bacteria (Cex,0) is a stepwise function that was set to match experimental conditions: 107 CFU/ml of bacteria were administered for 2 h, followed by perfusion of bacteria-free media for the remaining time.
The variables in the model are as follows: Cex and Cin are the normalized extracellular and intracellular densities (a value of one corresponds to 1x1010 CFU/ml), D is the dispersion coefficient, μg and μg,in are the extracellular and intracellular growth rates, μinv is the intracellular invasion rate, θ is the fraction of viable tumor cells, Kaff is the chemotactic affinity to chemokines in the tumor mass, Cchem is the normalized chemokine concentration, Cex,0 is the normalized density of bacteria that was perfused into the microfluidic device as a function of time (1x107 CFU/ml for t ≤ 2 h and 0 for t > 2 h), F0 is the media flow rate in the perfusion channel, V is the volume of the section of the perfusion channel in front of the tumor chamber, and A is the cross-sectional area of the tumor chamber. All intracellular and total bacterial fluorescence values were normalized to the highest cross sectional fluorescence intensity that occurred during the experiment.
Equations were discretized in space and solved in Matlab (The MathWorks, Inc., Natick, MA) using a finite difference method. The spatially discretized coupled ordinary differential equations were solved with the built-in ode15s function in Matlab for all spatial (discretized in ten points in space) and temporal points between 0 and 40 hours in 1 hour intervals. The fraction of viable cancer cells within the tumor mass (θ) was calculated based on previous data [9]. The extracellular growth rate was calculated based on the growth rate in liquid culture.
Two datasets (F-Sal vs. Sal and S-Sal vs. Sal) were used for modelling and normalized to one another to match control (Sal) conditions. The bacterial dispersion coefficient was calculated by fitting the model (eq. 1-3) to the tumor-on-a-chip experimental data of GFP for all spatial and temporal points up to 40 hours. The fminsearch function in Matlab was used to minimize the sum of least squares error between the experimental data and model by adjusting (and calculating) the rates of intracellular invasion and dispersion for both Sal datasets. The intracellular invasion rate of S-Sal was calculated by fixing the dispersion coefficient to be the same as Sal. The dispersion coefficient and intracellular invasion rate of F-Sal were calculated by bounding the dispersion coefficient such that it could not be lower than that of Sal. The intracellular accumulation rate was determined by quantifying the total change in intracellular density between 47 and 48 h.
Data and Statistical Analysis
Image and statistical analysis was performed in Matlab software. Unpaired t-tests with unequal variance were used to determine statistical significance with a level of P < 0.05.
Results
Induction of flhDC increases tumor colonization of Salmonella
Overexpressing flhDC in Salmonella increased intratumoral dispersion and colonization (Fig. 1). When administered to a tumor-on-a-chip device (Fig. 1A), F-Sal (induced flhDC) colonized tumor masses more than Sal (control) Salmonella (Fig. 1B). Both strains contained Plac/DsRed and expressed DsRed constitutively. In these images, red fluorescence indicates overall bacterial density. At 30 h, the size of the colony formed by F-Sal (white arrows) was considerably larger than the one formed by Sal (black arrows, Fig. 1B). The area of both colonies increased in size from 30 to 48 h after bacterial administration. Both colonies were located deep into the tissue, away from the perfusion channel (see Fig. 1A), indicating that both strains actively penetrated the tumor masses as we have described previously [5, 6]. Across multiple cell masses (n = 3 for Sal and n = 5 for F-Sal), the average density of F-Sal was significantly greater than Sal within entire tumor masses between 29 and 45 hours of bacterial colonization (P < 0.05; Fig. 1C). After 48 hours of bacterial colonization, F-Sal colonized both proximal (x ≤ 0.5) and distal (x = 0.9) tumor tissue more than Sal (P < 0.05; Fig. 1D). The density of F-Sal was greater than Sal throughout the middle of tumor masses (0.6 ≤ x 0.8), but was not significant (0.05 < P < 0.08) because of heterogeneous localization of colonies between cell masses (Fig. 1D). Overall, F-Sal colonized tumor tissue five-fold more than Sal (P < 0.05, Fig. 1E).
Fig. 1.

Inducing Salmonella with flhDC increase bacterial tumor colonization and dispersion. a) The microfluidic device contained a media perfusion channel and a chamber that holds tumor cell masses. The perfusion channel mimics tumor vasculature. Masses are formed as spheroids and inserted through tubing and control valves. Prior to insertion, spheroids are approximately 300 μm in diameter. b) Colonization of control Sal (black arrows) and flhDC-induced F-Sal (white arrows) was measured by with red fluorescence (red). Tumor cell masses are shown in the transmitted images under the fluorescence images. Images were background subtracted and shown with the maximum red intensity at the greatest observed value. Scale bar is 100 um. c) Salmonella with induced flhDC (F-Sal) colonized tumors significantly more than Salmonella (Sal) from 29 to 45 hours after bacterial administration (*, P<0.05, n = 3 for Sal and n = 5 for F-Sal). d) F-Sal colonized proximal (x≤0.5) tissue more than control Salmonella (Sal; *, P<0.05). The density was ten-fold greater for F-Sal in distal tumor tissue. e) At 48 hours after administration, F-Sal colonized tumors five-fold more control Sal (*, P<0.05).
Overexpression of flhDC increases intracellular accumulation of Salmonella
Upregulating flhDC in Salmonella increased intracellular accumulation in cells and tumor masses (Fig. 2). After induction with 0.2% arabinose, Salmonella motility increased by 25% (P<0.05, Fig. 2A). The non-motile fraction of bacteria (<15 μm/s) decreased seven-fold (P<0.01) and the motile fraction (>15 μm/s) increased two-fold (P<0.01, Fig. 2B).
Fig. 2.

Induction of flhDC increases intracellular accumulation. a) After flhDC induction, Salmonella (F-Sal) were 33% more motile in aqueous solution than control Salmonella (Sal). b) In aqueous solution, the motile fraction of Salmonella (15-30 μm/s) increased while the non-motile fraction (0-15 um/s) decreased (*, P < 0.05). c) In monolayer culture, Salmonella (green) invaded into MCF7 cells. Salmonella with flagella (control and pflhDC) invaded cells more than non-motile (ΔflgE and ΔflgE+pflhDC) Salmonella. Some ΔflgE+pflhDC Salmonella invaded cells. All Salmonella constitutively expressed GFP. Scale bar is 100 μm. d) Salmonella overexpressing flhDC invaded cells 1.25 times more than control Salmonella (***, P < 0.001). Salmonella with intact flagella (control and pflhDC) invaded cells significantly more than non-flagellated (ΔflgE and ΔflgE+pflhDC) Salmonella (***, P < 0.001). Non-motile ΔflgE+pflhDC Salmonella invaded cells more than ΔflgE Salmonella (**, P <0.01). e) Four strains of Salmonella were transformed with PSSEJ/GFP and Plac/DsRed to identify extracellular (red only) and intracellular (green and red) bacteria. f) The PSSEJ promoter is intracellularly activated. At an early time after invasion (2 hours), Salmonella only express DsRed (top left) and do not express GFP (bottom left). After 18 hours of incubation, intracellular Salmonella express both GFP (bottom right) and DsRed (top right). Scale bar is 100 μm. g) In tumor masses, many of the colonized Salmonella were intracellular. Scale bar is 100 μm. h) Overexpression of flhDC (F-Sal) increased the density of intracellular Salmonella in tumor masses 2.5 fold more than control Salmonella (Sal) at times greater than 29 hours after bacterial administration (*, P < 0.05). i) The average intracellular density of flhDC induced Salmonella was 2.5 fold greater than control Salmonella (*, P < 0.05). j) Induction of flhDC increased intracellular accumulation of F-Sal in medial (0.5 ≤ x ≤ 0.6) and distal (x ≥ 0.8) tumor tissue compared to controls (Sal; *, P < 0.05).
In monolayer culture, Salmonella invaded into MCF7 cells and the extent of invasion was dependent on flagella (Fig. 2C). Overexpression of flhDC increased invasion 1.25 times compared to control Salmonella (P < 0.001, Fig. 2D). Invasion was highly dependent on functional flagella. Control Salmonella invaded cells 26-fold more than non-motile ΔflgE Salmonella (P < 0.001; Fig. 2D). Similarly, functional flagella had a large effect on cell invasion for Salmonella overexpressing flhDC; pflhDC Salmonella invaded 7.2 times more than ΔflgE+pflhDC Salmonella (P < 0.001). Flagella-independent invasion was increased 4.6 times by overexpression of flhDC (P < 0.01).
Four of the Salmonella strains (Sal, F-Sal, S-Sal and FS-Sal; Table 1) were transformed with PSSEJ/GFP (intracellular GFP) and Plac/DsRed (constitutive DsRed) to identify and differentiate total (red only) and intracellular (red and green) Salmonella (Fig. 2E). This genetic circuit is necessary in tumor cell masses, because constitutive fluorescence would not differentiate intracellular and extracellular bacteria. A gentamycin protection assay was used to show that PSSEJ is a specific intracellular promoter. After applying control Salmonella (Sal) to a monolayer of cancer cells, all extracellular bacteria were removed with gentamycin. At early time points (2 h after gentamycin addition), GFP had yet to be translated (Fig. 2F, lower left) and all bacteria expressed DsRed (Fig. 2F, upper left). By 18 h, all intracellular bacteria (Fig. 2F, upper right) expressed both DsRed (Fig. 2F, upper right) and GFP (Fig. 2F, lower right), showing that the genetic circuits functioned as expected. In tumor-on-a-chip devices, overexpressing flhDC increased intracellular bacterial density (green, Fig. 2G). The high expression of GFP throughout the tumor masses (Fig. 2G) indicates that many of the Salmonella (both Sal and F-Sal) were intracellular (Additional file 2: Figure S2). Across all cell masses, the intracellular density of flhDC-induced F-Sal was significantly greater than control Sal from 29 to 45 h after administration (P < 0.05; Fig. 2H). Forty-eight hours after bacterial administration, the intracellular colonization of F-Sal was 2.5 fold more than Sal (P<0.05, Fig. 2I). In the middle of cell masses (0.5 < x < 0.6), induced F-Sal accumulated in cells 2.5 times more than control Sal (P < 0.05, Fig. 2J). Highly motile F-Sal also accumulated in distal tumor tissue (x ≥ 0.8) ten-fold more than Sal (P<0.05, Fig. 2J). These results demonstrate that flhDC induced Salmonella to accumulate in tumor cells.
Induction of flhDC does not increase tumor colonization in the absence of intracellular accumulation
To investigate the effect of flhDC induction in the absence of T3SS-based invasion, ∆sipB Salmonella (S-Sal) were administered to a tumor-on-a-chip device (Fig. 3). No difference was seen in the colonization pattern of extracellular (red) or intracellular (green) Salmonella (Fig. 3A). Across multiple tumor cell masses (n = 3), no differences were observed in the location of Salmonella colonization after flhDC induction, based on DsRed expression (Fig. 3B), and there was no effect on total bacterial density (Fig. 3C). Similarly, flhDC induction did not affect the location of intracellular Salmonella based on GFP expression (Fig. 3D) or overall density of intracellular Salmonella (Fig. 3E). The lack of difference between FS-Sal and S-Sal indicates that flhDC-mediated intracellular accumulation requires a functional T3SS-I.
Fig. 3.
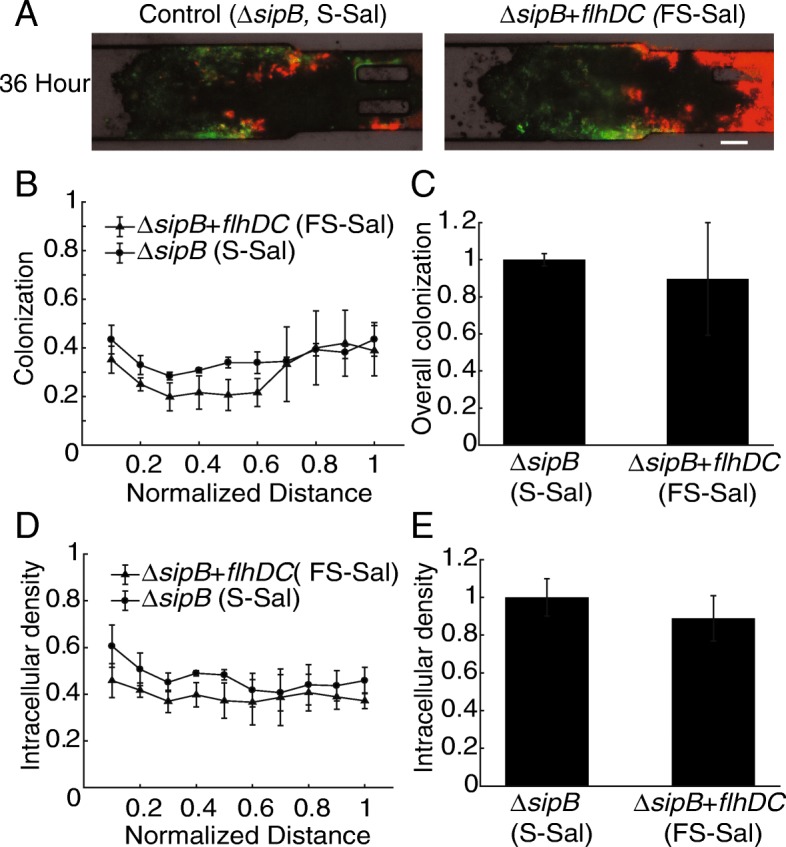
Induction of flhDC does not increase tumor colonization in the absence of T3SS1. a) In the absence of T3SS, extracellular (red only) and intracellular colonization (green and red) was minimal and uneven for flhDC-induced (FS-Sal) and control (S-Sal) Salmonella. Images were acquired 36 h after bacterial administration. Scale bar is 100 μm. b-e) When compared to control ΔsipB Salmonella (S-Sal), flhDC-induced ΔsipB Salmonella (FS-Sal) did not affect (b) the location of colonization, (c) the overall bacterial density, (d) the location of intracellular invasion, or (e) the overall extent of intracellular accumulation. Data (n = 3) were acquired 36 h after bacterial administration.
Intracellular accumulation of Salmonella increases tumor colonization in vitro
Minimally invasive, ΔsipB Salmonella (S-Sal) colonized tumor tissue less than control Salmonella (Sal, Fig. 4). Both S-Sal and control Sal expressed GFP after intracellular invasion and constitutively expressed DsRed (Table 1). Without sipB, Salmonella invaded cancer cells considerably less than controls, as indicated by diminished GFP fluorescence (Fig. 4A). S-Sal invaded MCF-7 cells six-fold less than the Sal control (P < 0.05, Fig. 4B). When, S-Sal were administered to tumor-on-a-chip devices the amount of intracellular bacteria (green) was considerably less than for control Sal (Fig. 4C). The number of intracellular Sal increased from 30 to 48 hours as indicated by the increase in GFP intensity, but little increase was observed for S-Sal (Fig. 4C). Over multiple devices (n = 6), S-Sal accumulated within tumor masses 2.5 fold less than the Sal control (P<0.05, Fig. 4D) and the rate of GFP fluorescence increase of S-Sal was four fold less than Sal (P<0.05; Fig. 4E). Total tumor colonization was quantified through constitutive DsRed fluorescence. Thirty hours after administration, more control Sal bacteria were present in devices than S-Sal (Fig. 4F). The difference between Sal and S-Sal was due to the increase in intracellular invasion because knocking out sipB did not affect the growth rates of the strains (Additional file 3: Figure S3-A). Over multiple masses, S-Sal colonized tumor tissue four fold less (P<0.05, Fig. 4G) and grew four fold slower than the Sal control (P<0.05; Fig. 4H). Sal visibly grew between 30 and 48 hours after bacterial administration, while the S-Sal density remained relatively unchanged during the same time period (Fig. 4F). These results demonstrated that intracellular accumulation is an essential component of Salmonella tumor colonization in vitro.
Fig. 4.

Tumor colonization of Salmonella depends on intracellular accumulation in tumor masses. a) Control Salmonella (Sal) intracellularly invaded MCF7 cells more than the minimally invasive ΔsipB Salmonella (S-Sal). Green fluorescence indicates induction of GFP expression by the PSSEJ promoter, which is activated intracellularly. Scale bar is 100 μm. b) The ΔsipB mutant (S-Sal) intracellularly invaded tumor cells ten-fold less than control Salmonella in monolayer (*, P<0.05). c) The sipB knockout reduced the amount of intracellular Salmonella (green) in devices at 30 and 48 h after administration. Scale bar is 100 μm. d, e) Compared to control Sal, S-Sal (d) accumulated in tumor cells in devices 2.5 fold less (*, P<0.05, n = 6) and (e) had a four-fold slower rate of fluorescence increase (*, P<0.05). f) The sipB knockout also reduced the total density of colonized Salmonella (red) in devices at 30 and 48 h after administration. Scale bar is 100 μm. g, h) Compared to control (Sal), S-Sal (G) colonized tumors 2.5 fold less (*, P<0.05) and (h) grew in tumors four-fold slower (*, P<0.05).
Intratumoral induction of flhDC improves colonization and intracellular accumulation of Salmonella
To determine if flhDC could be induced intratumorally, F-Sal was grown without arabinose and administered to tumor-on-a-chip devices. After induction with arabinose, F-Sal were 1.2 times faster in aqueous media compared to uninduced F-Sal (P<0.05; Fig. 5A). To test intratumoral induction, F-Sal were administered to devices for one hour in arabinose free medium (Fig. 5B). Twelve hours after administration, 0.4% arabinose added to the medium delivered in the flow channel to induce flhDC (Fig. 5B). Twelve hours was chosen as the time to induce, because this was the time when bacterial colonies could first be seen in the tumor cell masses (red arrows, Fig. 5C). At 47 h after administration, colonies grew in both uninduced and induced devices, but the induced colonies were visibly larger and located farther from the flow channel (Fig. 5C). Over multiple devices (n = 5 for uninduced and n = 6 for induced), intratumorally induced F-Sal colonized distal tumor tissue (0.8 ≤ x ≤ 1) five-fold more than the Sal control after 47 hours (P<0.05, Fig. 5D). The total amount of intratumorally induced F-Sal was two-fold greater than Sal (P <0.05, Fig. 5E).
Fig. 5.
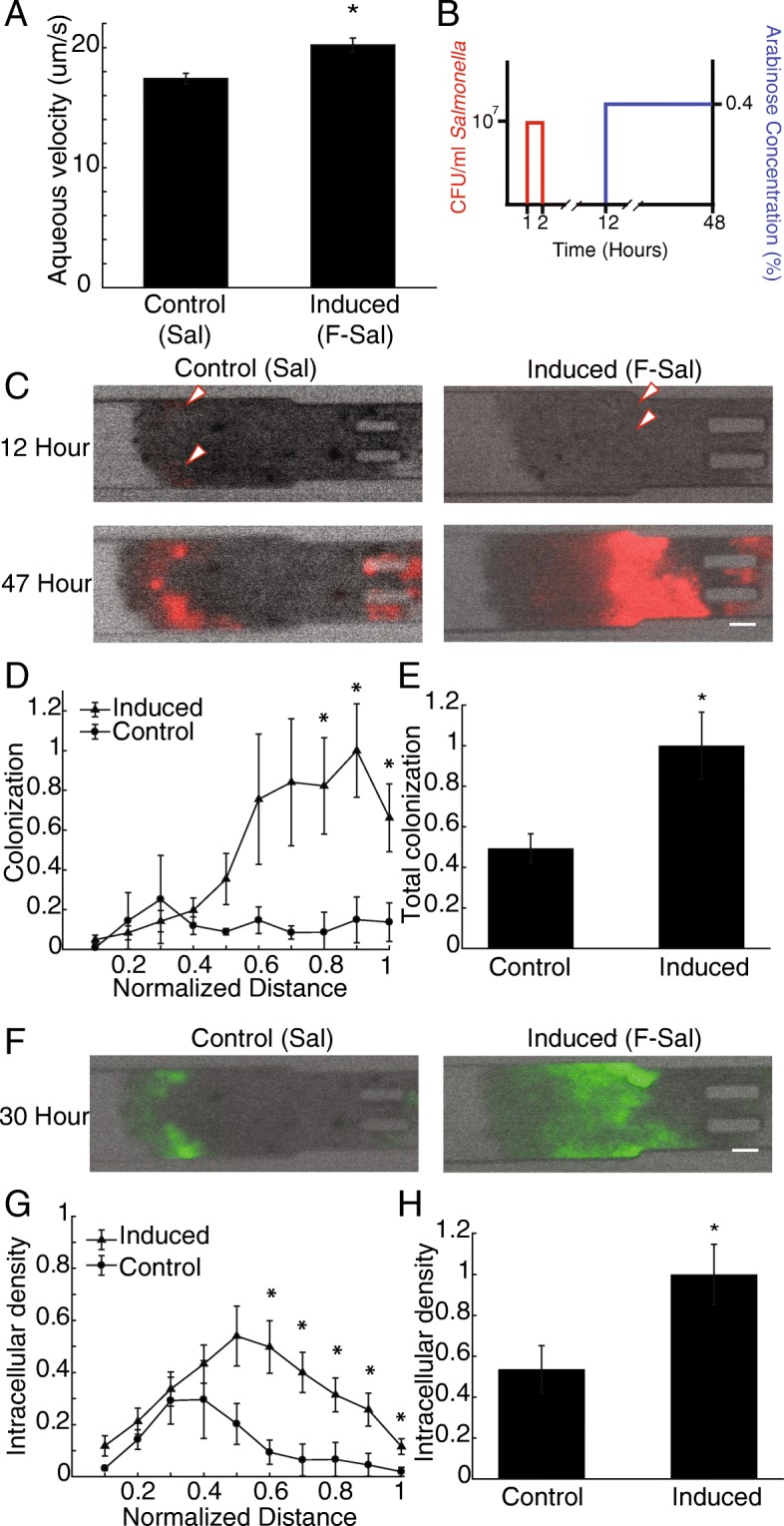
Intratumoral flhDC induction increases colonization, dispersion and intracellular accumulation of Salmonella. a) When flhDC was induced in Salmonella, aqueous motility increased by 18% compared to uninducedSalmonella containing the same pBAD-flhDC construct (*, P<0.05). b) Graphical depiction of the dosing scheme. One hour after tumors were placed into devices, Salmonella was administered for 1 hour. Eleven hours after bacterial administration, media with 0.4% (w/v) arabinose was administered to the devices to induce bacterial flhDC expression. c) When F-Sal was administered to devices, bacteria colonies (red arrows) were first detected at 12 hours. At 47 h, colonies formed by F-Sal with intratumorally induced flhDC were larger than control Salmonella (Sal). Scale bar is 100 μm. d) Spatial distribution of intratumoral bacteria. Intratumoral induction of flhDC increased the level of distal bacterial colonization in tumor masses after 47 hours (*, P<0.05). e) Intratumoral induction of flhDC increased overall tumor colonization (*, P<0.05). f) Intratumorally induction of flhDC increased the number of intracellular Salmonella (green). Scale bar is 100 μm. g) Intratumoral flhDC expression increased intracellular accumulation in the distal region (0.6 < x < 1) of tumor masses (*, P < 0.05). h) Induction of flhDC increased intracellular accumulation within entire tumor masses after 36 hours (*, P < 0.05).
Similar to overall density, induction increased the amount of intracellular F-Sal (Fig. 5F). Intracellular accumulation of intratumorally induced F-Sal was five-fold greater (P< 0.5) in intermediate tumor tissue (0.6 ≤ x ≤ 0.7) and two-fold greater (P< 0.5) in distal tumor tissue (0.8 ≤ x ≤ 1) compared to Sal (Fig. 5G). Total intracellular colonization of F-Sal was 1.8 fold greater than Sal after 30 hours (P <0.05, Fig. 5H). Intratumoral flhDC induction in Salmonella improved both distal colonization and intracellular accumulation when compared to Salmonella control, demonstrating that flhDC could be induced within tumors.
Intracellular accumulation improves bacterial retention in tumors
A model of bacterial dispersion, growth and intracellular invasion was used to determine how modulating intracellular accumulation affected tumor colonization. The model includes balances on extracellular and intracellular bacteria (eq. 1-2). Extracellular bacteria (eq. 1) could accumulate, disperse, chemotax, invade cells, or be convectively transferred into the perfusion channel at the x = 0 boundary (eq. 3 middle). The number of intracellular bacteria increase because of either growth or cell invasion (eq. 2).
The model was used to calculate rates of intracellular accumulation and the bacterial dispersion coefficient in tumor masses. The model was fit to the spatiotemporal profiles of intracellular bacterial density for S-Sal, Sal and F-Sal (Fig. 6A-C). The dispersion coefficient (D) was calculated to be 23.5 μm2/s, by fitting to the Sal data set. The dispersion coefficient did not increase when the mathematical model was fit to the F-Sal dataset. The rate of intracellular accumulation for F-Sal was 4.47 times greater than Sal, and the accumulation rate of S-Sal was 2.39 times less than Sal (Table 2).
Fig. 6.

Intracellular accumulation increases retention of bacteria by preventing flux out of tumors. a-c) The mathematical model of intratumoral dispersion and invasion (eq 1-3) was fit to (a) ΔsipB Salmonella (S-Sal), (b) Salmonella (Sal), and (c) pflhDC+Salmonella (F-Sal) to determine the intracellular accumulation rate of the three strains. The model was fit to all time points; images show the data and model fit at 31 h. d) The mathematical model fits experimental data and predicts that increasing intracellular accumulation would increase overall tumor colonization. e) The model predicts that increasing the rate of intracellular accumulation would increase overall tumor colonization, especially in intermediate tumor tissue (0.4 < x < 0.7). f, g) When the extracellular bacteria density is higher (compare S-Sal to F-Sal), there is a larger gradient at the front edge of the tumor (f), which causes more bacteria to leak out of tumors (g).
Table 2.
Calculated Intracellular accumulation rates
| Strain | Intracellular Accumulation Rate |
|---|---|
| S-Sal | 1.8x107 CFU ∗ hr−1 |
| Sal | 4.3x107 CFU ∗ hr−1 |
| F-Sal | 19.2x107 CFU ∗ hr−1 |
The model prediction of overall colonization as a function of the intracellular accumulation closely matched experimental data (Fig. 6D). When intracellular accumulation increased, overall tumor colonization increased. Theoretically extrapolating to bacteria that neither invade nor grow intracellularly suggests that they would not colonize tumors (Fig. 6D). Based on the model, the increase in bacterial density with higher rates of intracellular accumulation occurred primarily in intermediate regions of the cell masses (0.4 ≤ x ≤ 0.6; Fig. 6E). The calculated amounts of extracellular bacteria was greater for bacteria with lower rates of intracellular accumulation (i.e. S-Sal and Sal compared to F-Sal; Fig. 6F). Based on the model, this higher extracellular density (Fig. 6F) lead to greater leakage from the tumor and a lower overall density (Fig. 6G).
Discussion
The results of this study demonstrate key mechanisms that control Salmonella colonization of tumors. Using in vitro tumors that can be monitored for bacterial infiltration and proliferation in real time, we demonstrated that overexpressing the master motility regulator, flhDC, increased tumor colonization (Fig. 1). As expected, induction of flhDC increased the motility of Salmonella, but it also increased the accumulation inside cancer cells (Fig. 2). In Salmonella with impaired invasiveness, flhDC induction did not affect colonization (Fig. 3) showing that flhDC enhances colonization by increasing the number of intracellular bacteria. Similarly, when Salmonella were modified to impair their invasiveness, tumor colonization was dramatically reduced (Fig. 4), showing that intracellular invasion and growth is important for Salmonella colonization of tumors, independent of flhDC overexpression. Integrating the spatial and temporal tumor penetration data into a mathematical model enabled calculation of the intracellular accumulation rate and showed that invasion promotes colonization by increasing bacterial retention in tumors (Fig. 6). These mechanisms could be used to improve therapeutic efficacy by enhancing bacterial tumor colonization. When flhDC was induced after initial penetration, intracellular accumulation and tumor colonization both increased (Fig. 5).
Overexpression of flhDC increased intracellular accumulation through a T3SS-dependent mechanism. When flhDC was upregulated in T3SS-deficient Salmonella (FS-Sal), neither intracellular accumulation nor colonization increased (Fig. 3B-E). Induction of flhDC increased T3SS-dependent intracellular accumulation primarily through flagella production and moderately through increased synthesis of T3SS components (Figs. 2 and 3). Salmonella that were incapable of producing flagella (ΔflgE and ΔflgE+pflhDC) accumulated significantly less than those able to assemble flagella (Fig. 2C, D). Overexpressing flhDC in ΔflgE Salmonella only marginally improved intracellular accumulation (Fig. 2D). The difference between these effects shows that the major contribution of flhDC was to produce flagella, which in turn improved accumulation. The increase in accumulation of non-motile ΔflgE+pflhDC Salmonella, however, shows that flhDC control of T3SS synthesis does play a role in controlling accumulation.
Two primary mechanisms could have increased intracellular accumulation after flhDC induction: cell invasion and intracellular growth. The T3SS1 system and functional flagella are important for both. The injection of T3SS1 effectors into mammalian cells is critical for cell invasion [29]. Similarly, T3SS1 plays an important role in the escape of Salmonella from intracellular vacuoles and hyper-replication in the cellular cytoplasm [45–47]. In addition to T3SS, invasion could have been mediated by alternate mechanisms, such as the EGFR-dependent Rck system. The contribution of alternate mechanisms was considerably less than the T3SS system (Fig. 4B). T3SS-deficient Salmonella (S-Sal) colonized tumor masses three-fold less than T3SS-competent control bacteria (Sal; Fig. 4G), although residual intracellular accumulation (Fig. 4D) and colonization (Fig. 4G) was observed.
The intracellular niche provides Salmonella with an environment to proliferate (Additional file 3: Figure S3B-C) and that is protected from convective clearance (Fig. 6G). In MCF7 cells in monolayers, Salmonella grew with a doubling time of 3.6 h (Additional file 3: Figure S3C), which is considerably faster than the doubling time within tumors in mice (16.8 h) [58]. Overexpressing flhDC increased bacterial density inside cells (Fig. 2D) and in distal tumor tissue (Fig. 1D). The fact that T3SS-deficient Salmonella accumulated far less in tumor masses than control Salmonella (Sal, Fig. 4F, G) suggests that intracellular and distal tumor tissue are protected from convective clearance (Fig. 6E, F).
The mathematical model of bacterial invasion and colonization shows how intracellular accumulation would improve bacterial retention (Fig. 6). Convection continuously clears bacteria from tumor tissue located near the perfusion channel (Fig. 6F). This mechanism is analogous to convective clearance of bacteria from tumors by the bloodstream. By invading tumor cells, fewer bacteria would reside extracellularly (Fig. 6F) and fewer would be cleared (Fig. 6G). As the rate of intracellular accumulation increases, more bacteria are retained within the tumor (Fig. 6D), a mechanism similar to the ‘binding’ of small-molecule drugs to cancer cells [59]. With small molecule drugs, it has been shown that drug/receptor binding improves retention within tumors once the drug clears from the blood [59]. By ‘binding’ to cancer cells, the model suggested that Salmonella with higher rates of intracellular accumulation are less prone to leaking out of tumors (Fig. 6G).
A distally located reservoir of extracellular bacteria could serve as a continuous source for intracellular invasion and colonization of tumors. Within in vitro tumor masses, there is a considerable amount of bacterial colonization in necrotic and quiescent tissue, which is located between necrotic and actively dividing tumor tissue [7]. Of the total population of colonized bacteria, the majority of extracellular bacteria were located in necrosis (Fig. 6F). Neither intracellular nor extracellular bacteria resided in tissue near the channel because of the high rate of convective clearance (Fig. 6E, F). Due to the high dispersion coefficient, extracellular bacteria would rapidly clear out of proximal tissue close to the perfusion channel. However, extracellular bacteria residing in necrosis could grow faster than the rate of dispersion (Fig. 6F) allowing for high regional accumulation and migration to viable tissue to invade cells.
Controlling intracellular accumulation by inducing flhDC would increase tumor colonization. It would be beneficial to suppress flagellar expression outside of tumors. Flagella biosynthesis is an energetically costly process and can consume as much as 2% of bacterial energy [10, 60]. In addition , Salmonella flagellin is an immunogenic agonist that facilitates accelerated bacterial clearance [61]. Inducing flhDC selectively after initial penetration into tumors would improve fitness prior to administration, while promoting invasion and colonization within tumors (Fig. 7).
Fig. 7.
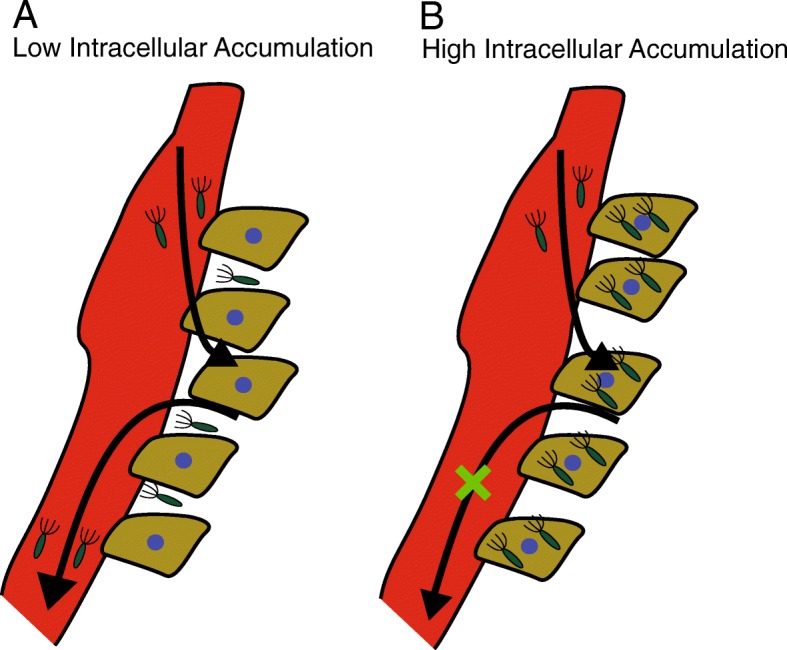
Graphical depiction of how intracellular accumulation could mechanistically improve tumor colonization. a) When Salmonella have a low intracellular accumulation rate, the rate of dispersion back into tumor vasculature is high, thus reducing bacterial tumor colonization due to a lack of “binding” to cancer cells. b) When Salmonella have a high intracellular accumulation rate, more bacteria would be retained in the tumor and not leak back into tumor microvasculature, thus increasing overall tumor colonization.
Conclusion
This study demonstrates that overexpressing flhDC increases intracellular accumulation within tumor cell masses, which drives tumor colonization. Robust tumor colonization is necessary for Salmonella to be an effective drug delivery vehicle. Intracellular accumulation increased colonization by causing Salmonella to ‘bind’ to tumor cells. This binding prevented bacteria from being convectively cleared from tumor masses. Selectively inducing flhDC expression within tumor masses would promote fitness prior to administration and enhance colonization after initial penetration. We envision that therapeutic strains of Salmonella will utilize inducible flhDC to drive colonization in human tumors. After intravenous administration and a period of initial penetration, an inducer would be provided to activate the flhDC regulator. Intracellular invasion enables Salmonella to deliver a wide range therapies directly into the intracellular space of tumors. Measuring the mechanisms of intracellular bacterial accumulation and tumor colonization has identified a key regulator, flhDC, that could be used to amplify colonization and make Salmonella an effective anticancer therapeutic.
Additional files
Figure S1. The four plasmids used in this study. A) The control plasmid contains the PSSEJ/GFP and Plac/DsRed genetic circuits as well as chloramphenicol resistance and the ColE1 origin of replication. It was transformed into the Sal and S-Sal (ΔsipB) strains. B) The motility induction plasmid contains all of the components of the control plasmid (panel A) in addition to an arabinose inducible PBAD/flhDC genetic circuit. This plasmid was transformed into the F-Sal and FS-Sal strains. C) The constitutive GFP control plasmid contains the Plac/GFP genetic circuit, ampicillin resistance, and the ColE1 origin of replication. This plasmid was transformed into the control Salmonella and ΔflgE strains for measurement of cell invasion and intracellular growth. D) The motility induction, constitutive GFP plasmid contains all of the components of the constitutive GFP plasmid (panel C) in addition to an arabinose inducible PBAD/flhDC genetic circuit. This plasmid was transformed into the Salmonella+pflhDC and ΔflgE+pflhDC strains for measurement of cell invasion and intracellular growth. (PDF 955 kb)
Figure S2. Merged fluorescent images of intratumoral Salmonella. Merged fluorescent images of intratumoral Salmonella. DsRed indicates the presence of all bacteria while GFP indicates the presence of intracellular bacteria. DsRed images have been enhanced to visualize all intratumoral bacteria. (PDF 3750 kb)
Figure S1. Growth Rates of Salmonella. A) Growth rate of Salmonella in liquid media (LB). All three strains grew at about the same rate (Sal, 1.313 hr-1; F-Sal, 1.273 hr-1; S-Sal; 1.26 hr-1), although F-Sal grew at a significantly slower rate than Sal (*, P < 0.05). There was no difference in the growth rates of ΔsipB (S-Sal) and control (Sal). B) Constitutive GFP fluorescence of intracellular Salmonella within MCF7 cells. The increase in intensity from one to five hours indicates the increase in the number of bacteria. Scale bar is 10 μm. C) Intracellular bacteria grew exponentially at a rate of 0.19 hr-1. (PDF 950 kb)
Acknowledgements
We would like to thank Shuo Sui and Sarah Perry in the Department of Chemical Engineering at the University of Massachusetts, Amherst for helping design the tumor-on-a-chip devices used in this study. We would like to thank James Fargnoli for assistance in optimizing tumor-on-a-chip operation protocols. We would also like to thank Poonam Phalak, also in the Chemical Engineering Department for help with mathematical modeling.
Funding
This work was funded by The National Institutes of Health (R01CA188382) and Baystate Health Foundation-Rays of Hope.
Availability of data and materials
Experimental data is available upon request
List of abbreviations
- flhDC
Salmonella master motility regulator
- T3SS1
Type three secretion system-1
- T3SS2
Type three secretion system-2
- fT3SS
Flagellar type three secretion system
- sipB
Type three secretion system cap protein
- GFP
Green fluorescent protein
- DsRed
A red fluorescent protein
- SSEJ-GFP
Intracellular GFP expression genetic circuit
- Lac-DsRed
Constitutive red fluorescent protein expression
- F-Sal
Salmonella transformed with SSEJ-GFP and Lac-DsRed
- Sal
Salmonella transformed with SSEJ-GFP and Lac-DsRed
- S-Sal
ΔsipB Salmonella transformed with Lac-DsRed
- FS-Sal
ΔsipB Salmonella transformed with SSEJ-GFP
- Lac
DsRed and PBAD-flhDC
- DMEM
Dulbecco’s minimal eagle medium
- FBS
Fetal bovine serum
- PMMA
Poly-(methyl)-methacrylate
- PDMS
Poly-(dimethyl)-siloxane
- HEPES
(4-(2-hydroxyethyl)-1-piperazineethanesulfonic acid)
- CFU
Colony forming unit
- LB
Luria Bertani broth
Authors’ contributions
VR designed, performed all experiments and wrote the manuscript. NVD and OMO assisted in performing experiments. NSF designed the experiments and wrote the manuscript.
Ethics approval and consent to participate
The research (protocol 2015-004) was approved by the UMass Institutional Animal Care and Use Committee on August 26, 2017.
Consent for publication
All listed authors consent to the publication of this research article.
Competing interests
Vishnu Raman and Owen O’Connor do not have any competing interests. Neil Forbes and Nele Van Dessel share ownership of Ernest Pharmaceuticals.
Publisher’s Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
References
- 1.Kocijancic D, Felgner S, Schauer T, Frahm M, Heise U, Zimmermann K, Erhardt M, Weiss S. Local application of bacteria improves safety of Salmonella-mediated tumor therapy and retains advantages of systemic infection. Oncotarget. 2017;8:49988–50001. doi: 10.18632/oncotarget.18392. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 2.Forbes NS, Munn LL, Fukumura D, Jain RK. Sparse initial entrapment of systemically injected Salmonella typhimurium leads to heterogeneous accumulation within tumors. Cancer Res. 2003;63:5188–5193. [PubMed] [Google Scholar]
- 3.Forbes NS. Engineering the perfect (bacterial) cancer therapy. Nature Reviews Cancer. 2010;10:785–794. doi: 10.1038/nrc2934. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 4.Toso JF, Gill VJ, Hwu P, Marincola FM, Restifo NP, Schwartzentruber DJ, Sherry RM, Topalian SL, Yang JC, Stock F, et al. Phase I study of the intravenous administration of attenuated Salmonella typhimurium to patients with metastatic melanoma. J Clin Oncol. 2002;20:142–152. doi: 10.1200/JCO.2002.20.1.142. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 5.Toley BJ, Forbes NS. Motility is critical for effective distribution and accumulation of bacteria in tumor tissue. Integr Biol (Camb) 2012;4:165–176. doi: 10.1039/c2ib00091a. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 6.Thornlow DN, Brackett EL, Gigas JM, Van Dessel N, Forbes NS. Persistent enhancement of bacterial motility increases tumor penetration. Biotechnol Bioeng. 2015;112:2397–2405. doi: 10.1002/bit.25645. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 7.Zhang M, Forbes NS. Trg-deficient Salmonella colonize quiescent tumor regions by exclusively penetrating or proliferating. J Control Release. 2015;199:180–189. doi: 10.1016/j.jconrel.2014.12.014. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 8.Silva-Valenzuela CA, Desai PT, Molina-Quiroz RC, Pezoa D, Zhang Y, Porwollik S, Zhao M, Hoffman RM, Contreras I, Santiviago CA, McClelland M. Solid tumors provide niche-specific conditions that lead to preferential growth of Salmonella. Oncotarget. 2016;7:35169–35180. doi: 10.18632/oncotarget.9071. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 9.Kasinskas RW, Forbes NS. Salmonella typhimurium lacking ribose chemoreceptors localize in tumor quiescence and induce apoptosis. Cancer Res. 2007;67:3201–3209. doi: 10.1158/0008-5472.CAN-06-2618. [DOI] [PubMed] [Google Scholar]
- 10.Gauger EJ, Leatham MP, Mercado-Lubo R, Laux DC, Conway T, Cohen PS. Role of motility and the flhDC Operon in Escherichia coli MG1655 colonization of the mouse intestine. Infect Immun. 2007;75:3315–3324. doi: 10.1128/IAI.00052-07. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 11.Wang X, Wood TK. IS5 inserts upstream of the master motility operon flhDC in a quasi-Lamarckian way. ISME J. 2011;5:1517–1525. doi: 10.1038/ismej.2011.27. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 12.Macnab RM. Genetics and biogenesis of bacterial flagella. Annu Rev Genet. 1992;26:131–158. doi: 10.1146/annurev.ge.26.120192.001023. [DOI] [PubMed] [Google Scholar]
- 13.Liu X, Matsumura P. The FlhD/FlhC complex, a transcriptional activator of the Escherichia coli flagellar class II operons. J Bacteriol. 1994;176:7345–7351. doi: 10.1128/jb.176.23.7345-7351.1994. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 14.Clarke MB, Sperandio V. Transcriptional regulation of flhDC by QseBC and sigma (FliA) in enterohaemorrhagic Escherichia coli. Mol Microbiol. 2005;57:1734–1749. doi: 10.1111/j.1365-2958.2005.04792.x. [DOI] [PubMed] [Google Scholar]
- 15.Singer HM, Kuhne C, Deditius JA, Hughes KT, Erhardt M. The Salmonella Spi1 virulence regulatory protein HilD directly activates transcription of the flagellar master operon flhDC. J Bacteriol. 2014;196:1448–1457. doi: 10.1128/JB.01438-13. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 16.Soutourina O, Kolb A, Krin E, Laurent-Winter C, Rimsky S, Danchin A, Bertin P. Multiple control of flagellum biosynthesis in Escherichia coli: role of H-NS protein and the cyclic AMP-catabolite activator protein complex in transcription of the flhDC master operon. J Bacteriol. 1999;181:7500–7508. doi: 10.1128/jb.181.24.7500-7508.1999. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 17.Sperandio V, Torres AG, Kaper JB. Quorum sensing Escherichia coli regulators B and C (QseBC): a novel two-component regulatory system involved in the regulation of flagella and motility by quorum sensing in E. coli. Mol Microbiol. 2002;43:809–821. doi: 10.1046/j.1365-2958.2002.02803.x. [DOI] [PubMed] [Google Scholar]
- 18.Wei BL, Brun-Zinkernagel AM, Simecka JW, Pruss BM, Babitzke P, Romeo T. Positive regulation of motility and flhDC expression by the RNA-binding protein CsrA of Escherichia coli. Mol Microbiol. 2001;40:245–256. doi: 10.1046/j.1365-2958.2001.02380.x. [DOI] [PubMed] [Google Scholar]
- 19.Yakhnin AV, Baker CS, Vakulskas CA, Yakhnin H, Berezin I, Romeo T, Babitzke P. CsrA activates flhDC expression by protecting flhDC mRNA from RNase E-mediated cleavage. Mol Microbiol. 2013;87:851–866. doi: 10.1111/mmi.12136. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 20.Wada T, Hatamoto Y, Kutsukake K. Functional and expressional analyses of the anti-FlhD4C2 factor gene ydiV in Escherichia coli. Microbiology. 2012;158:1533–1542. doi: 10.1099/mic.0.056036-0. [DOI] [PubMed] [Google Scholar]
- 21.Schlumberger MC, Hardt WD. Salmonella type III secretion effectors: pulling the host cell's strings. Curr Opin Microbiol. 2006;9:46–54. doi: 10.1016/j.mib.2005.12.006. [DOI] [PubMed] [Google Scholar]
- 22.Wiedemann A, Mijouin L, Ayoub MA, Barilleau E, Canepa S, Teixeira-Gomes AP, Le Vern Y, Rosselin M, Reiter E, Velge P. Identification of the epidermal growth factor receptor as the receptor for Salmonella Rck-dependent invasion. Faseb Journal. 2016;30:4180–4191. doi: 10.1096/fj.201600701R. [DOI] [PubMed] [Google Scholar]
- 23.Hapfelmeier S, Stecher B, Barthel M, Kremer M, Muller AJ, Heikenwalder M, Stallmach T, Hensel M, Pfeffer K, Akira S, Hardt WD. The Salmonella pathogenicity island (SPI)-2 and SPI-1 type III secretion systems allow Salmonella serovar typhimurium to trigger colitis via MyD88-dependent and MyD88-independent mechanisms. J Immunol. 2005;174:1675–1685. doi: 10.4049/jimmunol.174.3.1675. [DOI] [PubMed] [Google Scholar]
- 24.Pawelek JM, Sodi S, Chakraborty AK, Platt JT, Miller S, Holden DW, Hensel M, Low KB. Salmonella pathogenicity island-2 and anticancer activity in mice. Cancer Gene Ther. 2002;9:813–818. doi: 10.1038/sj.cgt.7700501. [DOI] [PubMed] [Google Scholar]
- 25.Arrach N, Cheng P, Zhao M, Santiviago CA, Hoffman RM, McClelland M. High-throughput screening for salmonella avirulent mutants that retain targeting of solid tumors. Cancer Res. 2010;70:2165–2170. doi: 10.1158/0008-5472.CAN-09-4005. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 26.Koskiniemi S, Gibbons HS, Sandegren L, Anwar N, Ouellette G, Broomall S, Karavis M, McGregor P, Liem A, Fochler E, et al. Pathoadaptive mutations in Salmonella enterica isolated after serial passage in mice. PLoS One. 2013;8:e70147. doi: 10.1371/journal.pone.0070147. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 27.Dumoux M, Nans A, Saibil HR, Hayward RD. Making connections: snapshots of chlamydial type III secretion systems in contact with host membranes. Curr Opin Microbiol. 2015;23:1–7. doi: 10.1016/j.mib.2014.09.019. [DOI] [PubMed] [Google Scholar]
- 28.Lucas RL, Lee CA. Roles of hilC and hilD in regulation of hilA expression in Salmonella enterica serovar Typhimurium. J Bacteriol. 2001;183:2733–2745. doi: 10.1128/JB.183.9.2733-2745.2001. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 29.Myeni SK, Wang L, Zhou D. SipB-SipC complex is essential for translocon formation. PLoS One. 2013;8:e60499. doi: 10.1371/journal.pone.0060499. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 30.Van Engelenburg SB, Palmer AE. Imaging type-III secretion reveals dynamics and spatial segregation of Salmonella effectors. Nat Methods. 2010;7:325–330. doi: 10.1038/nmeth.1437. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 31.Galan JE, Wolf-Watz H. Protein delivery into eukaryotic cells by type III secretion machines. Nature. 2006;444:567–573. doi: 10.1038/nature05272. [DOI] [PubMed] [Google Scholar]
- 32.Hayward RD, McGhie EJ, Koronakis V. Membrane fusion activity of purified SipB, a Salmonella surface protein essential for mammalian cell invasion. Mol Microbiol. 2000;37:727–739. doi: 10.1046/j.1365-2958.2000.02027.x. [DOI] [PubMed] [Google Scholar]
- 33.Knodler LA, Vallance BA, Hensel M, Jackel D, Finlay BB, Steele-Mortimer O. Salmonella type III effectors PipB and PipB2 are targeted to detergent-resistant microdomains on internal host cell membranes. Mol Microbiol. 2003;49:685–704. doi: 10.1046/j.1365-2958.2003.03598.x. [DOI] [PubMed] [Google Scholar]
- 34.Uchiya K, Barbieri MA, Funato K, Shah AH, Stahl PD, Groisman EA. A Salmonella virulence protein that inhibits cellular trafficking. EMBO J. 1999;18:3924–3933. doi: 10.1093/emboj/18.14.3924. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 35.Meresse S, Unsworth KE, Habermann A, Griffiths G, Fang F, Martinez-Lorenzo MJ, Waterman SR, Gorvel JP, Holden DW. Remodelling of the actin cytoskeleton is essential for replication of intravacuolar Salmonella. Cell Microbiol. 2001;3:567–577. doi: 10.1046/j.1462-5822.2001.00141.x. [DOI] [PubMed] [Google Scholar]
- 36.Birmingham CL, Jiang X, Ohlson MB, Miller SI, Brumell JH. Salmonella-induced filament formation is a dynamic phenotype induced by rapidly replicating Salmonella enterica serovar typhimurium in epithelial cells. Infect Immun. 2005;73:1204–1208. doi: 10.1128/IAI.73.2.1204-1208.2005. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 37.Guignot J, Caron E, Beuzon C, Bucci C, Kagan J, Roy C, Holden DW. Microtubule motors control membrane dynamics of Salmonella-containing vacuoles. J Cell Sci. 2004;117:1033–1045. doi: 10.1242/jcs.00949. [DOI] [PubMed] [Google Scholar]
- 38.Ribet D, Cossart P. How bacterial pathogens colonize their hosts and invade deeper tissues. Microbes Infect. 2015;17:173–183. doi: 10.1016/j.micinf.2015.01.004. [DOI] [PubMed] [Google Scholar]
- 39.Chubiz JE, Golubeva YA, Lin D, Miller LD, Slauch JM. FliZ regulates expression of the Salmonella pathogenicity island 1 invasion locus by controlling HilD protein activity in Salmonella enterica serovar typhimurium. J Bacteriol. 2010;192:6261–6270. doi: 10.1128/JB.00635-10. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 40.Mouslim C, Hughes KT. The effect of cell growth phase on the regulatory cross-talk between flagellar and Spi1 virulence gene expression. PLoS Pathog. 2014;10:e1003987. doi: 10.1371/journal.ppat.1003987. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 41.Elhadad D, Desai P, Rahav G, McClelland M, Gal-Mor O. Flagellin Is Required for Host Cell Invasion and Normal Salmonella Pathogenicity Island 1 Expression by Salmonella enterica Serovar Paratyphi A. Infect Immun. 2015;83:3355–3368. doi: 10.1128/IAI.00468-15. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 42.Misselwitz B, Barrett N, Kreibich S, Vonaesch P, Andritschke D, Rout S, Weidner K, Sormaz M, Songhet P, Horvath P, et al. Near surface swimming of Salmonella Typhimurium explains target-site selection and cooperative invasion. PLoS Pathog. 2012;8:e1002810. doi: 10.1371/journal.ppat.1002810. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 43.Abby SS, Rocha EP. The non-flagellar type III secretion system evolved from the bacterial flagellum and diversified into host-cell adapted systems. PLoS Genet. 2012;8:e1002983. doi: 10.1371/journal.pgen.1002983. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 44.Lee SH, Galan JE. Salmonella type III secretion-associated chaperones confer secretion-pathway specificity. Mol Microbiol. 2004;51:483–495. doi: 10.1046/j.1365-2958.2003.03840.x. [DOI] [PubMed] [Google Scholar]
- 45.Knodler LA, Vallance BA, Celli J, Winfree S, Hansen B, Montero M, Steele-Mortimer O. Dissemination of invasive Salmonella via bacterial-induced extrusion of mucosal epithelia. Proc Natl Acad Sci U S A. 2010;107:17733–17738. doi: 10.1073/pnas.1006098107. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 46.Wrande M, Andrews-Polymenis H, Twedt DJ, Steele-Mortimer O, Porwollik S, McClelland M, Knodler LA. Genetic Determinants of Salmonella enterica Serovar Typhimurium Proliferation in the Cytosol of Epithelial Cells. Infect Immun. 2016;84:3517–3526. doi: 10.1128/IAI.00734-16. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 47.Knodler LA, Nair V, Steele-Mortimer O. Quantitative assessment of cytosolic Salmonella in epithelial cells. PLoS One. 2014;9:e84681. doi: 10.1371/journal.pone.0084681. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 48.van Wijk SJL, Fricke F, Herhaus L, Gupta J, Hotte K, Pampaloni F, Grumati P, Kaulich M, Sou YS, Komatsu M, et al. Linear ubiquitination of cytosolic Salmonella Typhimurium activates NF-kappaB and restricts bacterial proliferation. Nat Microbiol. 2017;2:17066. doi: 10.1038/nmicrobiol.2017.66. [DOI] [PubMed] [Google Scholar]
- 49.Fiskin E, Bionda T, Dikic I, Behrends C. Global Analysis of Host and Bacterial Ubiquitinome in Response to Salmonella Typhimurium Infection. Mol Cell. 2016;62:967–981. doi: 10.1016/j.molcel.2016.04.015. [DOI] [PubMed] [Google Scholar]
- 50.Wild P, Farhan H, McEwan DG, Wagner S, Rogov VV, Brady NR, Richter B, Korac J, Waidmann O, Choudhary C, et al. Phosphorylation of the autophagy receptor optineurin restricts Salmonella growth. Science. 2011;333:228–233. doi: 10.1126/science.1205405. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 51.Thurston TL, Ryzhakov G, Bloor S, von Muhlinen N, Randow F. The TBK1 adaptor and autophagy receptor NDP52 restricts the proliferation of ubiquitin-coated bacteria. Nat Immunol. 2009;10:1215–1221. doi: 10.1038/ni.1800. [DOI] [PubMed] [Google Scholar]
- 52.Cemma M, Kim PK, Brumell JH. The ubiquitin-binding adaptor proteins p62/SQSTM1 and NDP52 are recruited independently to bacteria-associated microdomains to target Salmonella to the autophagy pathway. Autophagy. 2011;7:341–345. doi: 10.4161/auto.7.3.14046. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 53.Datsenko KA, Wanner BL. One-step inactivation of chromosomal genes in Escherichia coli K-12 using PCR products. Proc Natl Acad Sci U S A. 2000;97:6640–6645. doi: 10.1073/pnas.120163297. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 54.Deditius JA, Felgner S, Sporing I, Kuhne C, Frahm M, Rohde M, Weiss S, Erhardt M. Characterization of Novel Factors Involved in Swimming and Swarming Motility in Salmonella enterica Serovar Typhimurium. PLoS One. 2015;10:e0135351. doi: 10.1371/journal.pone.0135351. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 55.Walsh CL, Babin BM, Kasinskas RW, Foster JA, McGarry MJ, Forbes NS. A multipurpose microfluidic device designed to mimic microenvironment gradients and develop targeted cancer therapeutics. Lab Chip. 2009;9:545–554. doi: 10.1039/B810571E. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 56.Mohan R, Schudel BR, Desai AV, Yearsley JD, Apblett CA, Kenis PJA. Design considerations for elastomeric normally closed microfluidic valves. Sensors and Actuators B-Chemical. 2011;160:1216–1223. doi: 10.1016/j.snb.2011.09.051. [DOI] [Google Scholar]
- 57.Kasinskas RW, Forbes NS. Salmonella typhimurium specifically chemotax and proliferate in heterogeneous tumor tissue in vitro. Biotechnol Bioeng. 2006;94:710–721. doi: 10.1002/bit.20883. [DOI] [PubMed] [Google Scholar]
- 58.Ganai S, Arenas RB, Sauer JP, Bentley B, Forbes NS. In tumors Salmonella migrate away from vasculature toward the transition zone and induce apoptosis. Cancer Gene Ther. 2011;18:457–466. doi: 10.1038/cgt.2011.10. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 59.Toley BJ, Tropeano Lovatt ZG, Harrington JL, Forbes NS. Microfluidic technique to measure intratumoral transport and calculate drug efficacy shows that binding is essential for doxorubicin and release hampers Doxil. Integr Biol (Camb) 2013;5:1184–1196. doi: 10.1039/c3ib40021b. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 60.Soutourina OA, Bertin PN. Regulation cascade of flagellar expression in Gram-negative bacteria. FEMS Microbiol Rev. 2003;27:505–523. doi: 10.1016/S0168-6445(03)00064-0. [DOI] [PubMed] [Google Scholar]
- 61.Yang X, Thornburg T, Suo Z, Jun S, Robison A, Li J, Lim T, Cao L, Hoyt T, Avci R, Pascual DW. Flagella overexpression attenuates Salmonella pathogenesis. PLoS One. 2012;7:e46828. doi: 10.1371/journal.pone.0046828. [DOI] [PMC free article] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.
Supplementary Materials
Figure S1. The four plasmids used in this study. A) The control plasmid contains the PSSEJ/GFP and Plac/DsRed genetic circuits as well as chloramphenicol resistance and the ColE1 origin of replication. It was transformed into the Sal and S-Sal (ΔsipB) strains. B) The motility induction plasmid contains all of the components of the control plasmid (panel A) in addition to an arabinose inducible PBAD/flhDC genetic circuit. This plasmid was transformed into the F-Sal and FS-Sal strains. C) The constitutive GFP control plasmid contains the Plac/GFP genetic circuit, ampicillin resistance, and the ColE1 origin of replication. This plasmid was transformed into the control Salmonella and ΔflgE strains for measurement of cell invasion and intracellular growth. D) The motility induction, constitutive GFP plasmid contains all of the components of the constitutive GFP plasmid (panel C) in addition to an arabinose inducible PBAD/flhDC genetic circuit. This plasmid was transformed into the Salmonella+pflhDC and ΔflgE+pflhDC strains for measurement of cell invasion and intracellular growth. (PDF 955 kb)
Figure S2. Merged fluorescent images of intratumoral Salmonella. Merged fluorescent images of intratumoral Salmonella. DsRed indicates the presence of all bacteria while GFP indicates the presence of intracellular bacteria. DsRed images have been enhanced to visualize all intratumoral bacteria. (PDF 3750 kb)
Figure S1. Growth Rates of Salmonella. A) Growth rate of Salmonella in liquid media (LB). All three strains grew at about the same rate (Sal, 1.313 hr-1; F-Sal, 1.273 hr-1; S-Sal; 1.26 hr-1), although F-Sal grew at a significantly slower rate than Sal (*, P < 0.05). There was no difference in the growth rates of ΔsipB (S-Sal) and control (Sal). B) Constitutive GFP fluorescence of intracellular Salmonella within MCF7 cells. The increase in intensity from one to five hours indicates the increase in the number of bacteria. Scale bar is 10 μm. C) Intracellular bacteria grew exponentially at a rate of 0.19 hr-1. (PDF 950 kb)
Data Availability Statement
Experimental data is available upon request