

Abstract
The use of thermotolerant yeast strains is an important attribute for a cost-effective high temperature biofermentation processes. However, the availability of thermotolerant yeast strains remains a major challenge. Isolation of temperature resistant strains from extreme environments or the improvements of current strains are two major strategies known to date. We hypothesised that bacteria are potential “hurdles” in the life cycle of yeasts, which could influence the evolution of extreme phenotypes, such as thermotolerance. We subjected a wild-type yeast, Lachancea thermotolerans to six species of bacteria sequentially for several generations. After coevolution, we observed that three replicate lines of yeasts grown in the presence of bacteria grew up to 37 °C whereas the controls run in parallel without bacteria could only grow poorly at 35 °C retaining the ancestral mesophilic trait. In addition to improvement of thermotolerance, our results show that the fermentative ability was also elevated, making the strains more ideal for the alcoholic fermentation process because the overall productivity and ethanol titers per unit volume of substrate consumed during the fermentation process was increased. Our unique method is attractive for the development of thermotolerant strains or to augment the available strain development approaches for high temperature industrial biofermentation.
Keywords: Thermotolerance, Experimental evolution, Yeast-bacteria coevolution, Alcoholic fermentation
Introduction
Microbial alcoholic fermentation processes remain at the heart of humankind since the dawn of civilization. The efficiency of this indispensable process is often hampered by the paucity of microbes to withstand harsh environmental stresses [1]. One of the most important environmental factors that affect microbial growth and survival is temperature. Thermotolerant industrial production strains are important for the reduction of cooling costs and protection of fermentation processes from failure due to accidental thermal management faults or increased ambient temperatures [2]. The need for thermotolerant strains, exacerbated by the current global warming effects, which require a more stringent temperature control in large-scale industrial fermentors, is a major concern in modern bioprocesses. Approaches to obtain thermotolerant strains are strongly required. One possible approach is the isolation of naturally thermotolerant yeast species from their native habitats. However, the availability of thermotolerant yeast strains from nature is a major challenge because such characteristics are less commonly found in natural yeasts isolates. This is so, because in nature microorganisms tend to optimize their traits exclusively for survival and reproduction. Such characteristics are mostly irrelevant for modern technical applications, for example, in stressful large-scale industrial conditions [3]. Only a handful of thermotolerant species isolated from tropical environments as well extreme environments around wine production plants and environments with increased levels of solar radiation have been reviewed [2]. Yeasts isolated around the production plants are adaptively evolved and in most cases possess robust abilities to survive other extreme stresses [4]. Another approach to obtain thermotolerant strains is generation of artificial diversity, for example, directed evolutionary engineering. This approach involves the pre-adaptation of yeasts to stressors common in specific industrial conditions. This includes, the pre-exposure of yeasts to mild forms and sub-lethal stressors such as high temperatures, high concentrations of toxic or growth impairing substances, for example, organic compounds, osmolytes, ethanol and weak acids, among others [5–7]. However, such a method is often associated with reduction in productivity and loss of phenotypes or trade-offs in phenotypes important for a specific production process [8, 9]. The reduction in growth rates and inability to utilize the intended source of carbon are an example of major drawbacks associated with this approach reported elsewhere [8, 9]. Another possible approach is genetic engineering. Knowledge-based genetic modification remains less effective as thermotolerance is encoded by multiple quantitative traits [10–12]. Other methods hinged on generation of artificial diversity such as mutagenesis and classical breeding exist albeit with their limitations similar to evolutionary engineering. New methods to develop thermotolerant yeasts retaining fermentative traits are, therefore, of interest.
In this work, we extended the development of thermotolerant yeast strains not only beyond the conventional strategies but also beyond the industrial “workhorse”, Sacharomyces cerevisiae, using non-conventional yeasts. Lachancea thermotolerans, a yeast that diverged from the S. cerevisiae lineage prior to its whole genome duplication [13], was sequentially cocultured with six bacteria of increasing tolerance to ethanol for several generations as reported in our previous studies [14–16]. Our evolution strategy ascertains that microorganisms’ adaptive phenotypes in nature are hinged on the interactions with others [17]. We hypothesised that mimicking the natural habitat where yeasts are sympatric to bacteria may yield isolates with tolerance to extreme stress [14, 16], while keeping their fermentative capabilities as ethanol is key to their survival strategy with respect to bacteria [16].
In this study, we sought to develop strains with elevated thermotolerance. To screen for the emergence of elevated thermotolerant populations of L. thermotolerans after evolution in the presence of bacteria, we investigated their thermal sensitivity and growth between mesophilic, i.e., 30 °C to growth inhibitory temperatures, i.e. ~ 40 °C [18]. Other than thermotolerance, the strain’s ability to produce ethanol is dependent on its tolerance to other stressors that are associated with biofermentation. High ethanol stress, chemical surfactants, inhibitors and ROS exerted by substances found in raw materials as well as others are common examples [19–21]. Thus, we investigated the evolution of cross-protection from other extreme environmental stressors such as high ethanol titers, reactive oxygen species as well as ability to withstand surfactants. Further comparative analyses of molecular changes of the evolved strains were carried out using pulse field gel electrophoresis (PFGE). The fermentative capacity, before and after evolution, was then investigated to ascertain the suitability of the method in developing strains for highly productive high temperature fermentations.
Materials and methods
Strains used in this study
A wild type non-conventional yeast, L. thermotolerans, type strain CBS 6340, was the parental strain used in this study. Bacterial species, Pantoea agglomerans Eh318 (CUCPB 2140), Serattia plymuthica AS9 (CCUG 61396), Bacillus subtilis PS216 (BGSC 3A36), Streptomyces venezuelae (ATCC 10712), Lactococcus lactis (NCDO 2118) and Pseudomonas fluorescens (NCIMB 10462) previously reported [14–16] were sequentially used as a selection pressure to evolve L. thermotolerans. S. cerevisiae (S288c) and Schizosaccharomyces pombe (SJA148) strains were used as standards for karyotyping.
Adaptive laboratory evolution experiments
A unique experimental evolution strategy involving the sequential introduction of bacteria to compete with yeasts and the subsequent elimination of bacteria through the addition of antibiotics before transferring yeasts into fresh media, previously reported [14–16] was used. In brief, six flasks containing rich medium (YPD: 2% glucose, 0.5% yeast extract and 1% peptone, at a pH of 6.2 in 250 mL baffled-bottom flasks at 25 °C) were inoculated with an isogenic colony of L. thermotolerans (CBS6340 strain). Three flasks with and three without bacteria (controls) were then incubated and refreshed for several passages as reported [14–16]. More specifically, we grew 25 mL of yeast culture (5 ± 0.05 log10 CFUs/mL) in YPD in baffled-bottom flasks in an incubating shaker for 4 h (end of lag phase) at 200 revolutions per minute (r.p.m) at 25 °C before a predetermined amount of bacteria (4 ± 0.05 log10 CFUs/mL) was inoculated into an already adapted yeast population. The co-culture was incubated for 40 h, which was a predetermined time point before diauxic shift, and then bacteria were killed by addition of streptomycin (100 μg/mL). After sufficient time, at least 4 h after addition of the antibiotics, we transferred 50 μL of exponentially growing yeasts (7.7 ± 0.1 log10 CFUs/mL) into 25 mL fresh YPD. We carried out 20 such transfer passages before exchanging the bacterium with another species. Each transfer passage was approximately 8 generations amounting to a total of at least 180 generations, per bacterial species used. We froze 500 μL of the cell culture suspension in 25% glycerol at − 80 °C for analyses before each transfer cycle. This procedure was repeated for several generations allowing yeasts to compete with each bacterial species sequentially in order of increasing tolerance to ethanol (P. agglomerans, S. plymuthica, B. subtilis, S. venezuelae, L. lactis, and P. flourescens) as reported in [14]. The order with which yeast encounter bacteria is significant for selection towards better ethanol producers [14, 16], thus in this study it was intended to select for elevated ethanol producers in addition to thermotolerant strains. We sequenced rDNA D1/D2 region, amplified by universal primers NL1 (forward primer)(5′-GCATATCAATAAGCGGAGGA AAAG-3′) and NL4 (reverse primer) (5′-GGTCCGTGTTTCAAGACGG-3′) of the yeast before proceeding with the next bacterium to examine contamination despite periodic plating and microscopy as reported by [16]. Transfer cycles were carried out before the stationary phase of the yeasts to exclude other possible selection pressures and outcomes that could arise when a substrate (glucose) becomes limiting or is exhausted in the medium as reported elsewhere [22, 23], and thus could mask our bacterial competition selection pressure hypothesis set in the experiment. A detailed description of the method was fully reported elsewhere [15].
Isolation of thermotolerant strains
From each frozen sample stored after every 80 generations (10 passages) we plated out cells (7.7 ± 0.1 log10 CFUs/mL). Single independent colonies were picked and used to initiate cultures for thermal stability investigation. Cultures were grown in 2 mL YPD (2% glucose, 0.5% yeast extract and 1% peptone, at a pH of 6.2) in test tubes and incubated overnight at 25 °C in an orbital shaker (Infors HT). Cells were then pelleted using a microfuge and washed twice with phosphate buffered saline (PBS) and adjusted to OD600nm 0.06, which corresponds to 5.75 ± 0.2 log10 CFUs/mL. 100 μL of each of the independent cultures cells was then transferred to 96-well plates. Cells were then spotted on solid YPD plates using a 6 × 8 array spot replicator (Sigma Aldrich) before incubation at 30 °C, 35 °C, 37 °C, 39 °C, and 40 °C for 5 days. All assays were repeated at least twice, each time starting with an independent colony.
Investigation of tolerance to other stressors
Thermotolerant colonies from each of the six evolution lines (three from control-evolved and three from bacterial competition) were further used to determine their sensitivity to different environmental stresses. Single colonies were used to initiate overnight cultures in 2 mL YPD (2% glucose, 0.5% yeast extract and 1% peptone, at a pH of 6.2) or YNB (Yeast Nitrogen Base: 1.7 g/L; 5 g/L ammonium sulphate supplemented with 2% glucose, pH 6.2), in test tubes and incubated overnight at 25 °C in an orbital shaker (Infors HT). The cultures were then pelleted using a microfuge and washed twice in 1000 μL of PBS and then resuspend in 500 μL. The cultures were then adjusted to an OD600nm of 0.06 before being spotted on plates supplemented with reagents responsible for respective stresses. Effects of stressors were established by scoring growth of the evolved strains in comparisons to the ancestral strain. For record keeping and analyses, spots on petri dishes were scanned after 2 days unless otherwise specified.
Ethanol tolerance assay
Ethanol tolerance was assayed using the same procedures as above except that yeasts were spotted on solid YPD containing different concentrations of ethanol (6–10%) and incubated at 25 °C. Growth was scored for 10 days.
Oxidative stress (menadione and H2O2)
Sensitivity to H2O2 induced oxidative stress was determined using cells at stationary phase. Cells were grown overnight in YNB (Yeast Nitrogen Base: 1.7 g/L; 5 g/L ammonium sulphate supplemented with 2% glucose, pH 6.2), washed and diluted to OD600nm 0.06, which corresponds to 5.75 ± 0.2 log10 CFUs/mL. Cells were then spotted on solid YNB containing different concentrations of H2O2 (0–4 mM) using a spot test replicator. Plates were incubated up to 5 days at 25 °C. The same procedures were repeated with menadione of concentration ranges between 0.14 and 0.18 mM. Experiments were carried out in triplicate and repeated twice.
Sensitivity to sodium dodecyl-sulfate (SDS)
The same procedure was carried out as above except that SDS was used as a stressor at concentration ranges of 0.01–0.02%. Experiments were carried out in triplicate and repeated twice.
Karyotyping of evolved strains
From each frozen sample stored after every 80 generations (10 passages) we plated out cells (7.7 ± 0.1 log10 CFUs/mL) and randomly selected three independent colonies and determined their karyotypes using a CHEF Mapper XA PFGE apparatus (Bio-Rad) as described [24]. To determine the genetic stability of the colonies, 20 sequential transfers of the colonies under non-selective conditions (YPD at 25 °C) were carried out and followed by karyotype analyses.
Fermentation trials
Fermentative capacity of thermotolerant strains under aerobic conditions in flasks was investigated. We used the most thermotolerant and ethanol tolerant strains, i.e., passage 50 from evolution lines B and C. Cells from evolution line A were not included due to their ethanol sensitivity. In addition, those evolved in the absence of bacteria (lines D, E, and F) were not included due to their thermal sensitivity. We revived frozen cultures from evolution lines B as well as C by plating them out (7.7 ± 0.1 log10 CFUs/mL) on solidified YPD agar. The plates were incubated at 25 °C. Three independent colonies were randomly selected for inoculation in YPD as overnight seed cultures. After incubation, the cells were pelleted and washed with sterile water before inoculating 25 mL YPD (2% glucose, 0.5% yeast extract and 1% peptone), at a pH of 6.2 in 250 mL baffled-bottom Erlenmeyer shake flasks. The flasks were incubated at 37 °C at 200 r.p.m in an Infors HT Ecotron shaker unit (Infors HT). The starting OD600nm was set at 1. The ancestral strain was used as a control but was grown at 30 °C as it could not grow at 37 °C. We performed these batch cultures in triplicate. Samples withdrawn at appropriate intervals were used to monitor cell growth and analyze metabolites using an HPLC with specifications as reported [16]. Product yields; maximum specific rates of glucose consumption and ethanol production rates were calculated as reported [16, 25]. The experiments were carried out in triplicate and repeated twice.
Results
Acquisition of thermotolerance
A type strain, Lachancea thermotolerans CBS6340 isolated from plum jam by Filippov in 1932 [26] was chosen for evolution experiments. New strains of the same species are frequently isolated from drosophilids and other fruit feeding insects [27]. Thermotolerance abilities are variable among the strains of this species [26]. In contrast to the Westerdijk Fungal Biodiversity Institute (CBS-KNAW) strain database, which reports that the strain grows up to 35 °C [28], we report in this study that growth of this strain at this temperature is already impaired (Fig. 1). To establish the evolution of stress tolerance as a measure of adaptation, we investigate thermotolerance of strains evolved in the presence or absence of a bacterial selection pressure. Frozen samples stored after every 80 generations (10 passages) were plated out and used in the investigations. Our results show that isolates evolved in the presence of bacteria grew very well at 35 °C whereas those evolved in the absence of bacteria retained the ancestral phenotype (Fig. 1). These co-culture evolved strains grew well at an even higher temperature, 37 °C, whereas controls were completely inhibited. We noted that strains isolated after 50 passages (400 generations) grew far much better than those isolated from previous passages, suggesting a stepwise evolution of a thermal stress resistant trait. However, a higher temperature, of 39 °C and above, inhibited these evolved strains as well as their ancestor. As the evolution of thermal stress resistance was evident in all triplicate evolution lines (lines A, B and C), we were prompted to investigate if the strains were also tolerant to other environmental stresses.
Fig. 1.

Thermotolerance of evolved strains. Thermotolerance of strains evolved in the presence or absence of bacteria, co-culture evolved and control respectively, with reference to their ancestor, was determined by spotting cells and incubation at different temperatures. Co-culture evolved strains gained an improved thermotolerant phenotype not exhibited by the controls as well as the ancestral strain. The arrangement of the colonies on the plates is shown on the right. Numbers on the column of the illustrative stamp show passage numbers 10 to passage number 80 and continue on the fourth column with strains isolated after 90 passages and 100 passages respectively. The second and third column shows the same orientation. A–B–C were from triplicates of co-culture evolved lines whereas D–E–F in brackets were from controls (evolved without bacteria). For example lines A(D) means that colonies were either from flask A (co-culture evolved) or D (controls). The ancestral strain used as a reference was spotted and denoted as “Anc” on six positions on each plate
Evolution of resistance to other stresses
Ethanol resistance
Thermal stress response exhibits a functional overlap with ethanol stress responses in S. cerevisiae [29]. Thus, we tested the performance of thermotolerant strains on solid media with different concentrations of ethanol. Petri dishes were scanned after 5, 7 and 10 days of incubation. Figure 2 shows a clear distinction of ethanol stress tolerance, as we noted that colonies isolated from the co-culture evolved flasks grew on 6% ethanol after 5 days as compared to absence of growth on the control strain plates. The ancestral strain was also inhibited by 6% ethanol. Co-culture evolved yeasts from passages 10–50 were the most resistant as they appeared on the plates with up to 8% ethanol within 5 days of incubation as compared to the controls, which appeared 2 days later. However, cells plated after 50 passages of evolution, evolution lines B and C stood out both under 7% and 8% ethanol, whereas line A did not grow. Although yeasts evolved in the absence of bacteria grew after 7 days, we noted that they did so poorly. After 10 days, we noted that colonies from the co-culture evolved flasks grew much better on 9% ethanol in comparison to the control evolved strains. As a clear ethanol resistance phenotype, only the co-culture evolved colonies from lines B and lines C survived and grew on 10% ethanol after 10 days.
Fig. 2.
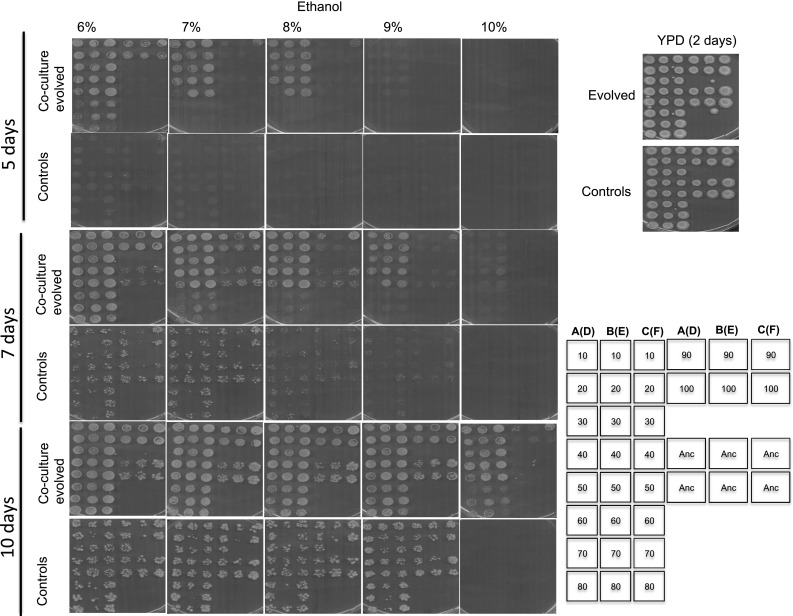
Ethanol tolerance of evolved strains. Ethanol tolerance of strains evolved in the presence or absence of bacteria, co-culture evolved and controls respectively, with reference to their ancestor, was determined by spotting cells from stationary phase cultures onto different concentrations of ethanol. Growth was scored after 5, 7 and 10 days. The strains isolated from co-evolved showed a higher ethanol tolerance than the control and ancestral strains. The arrangement of the colonies on the plates is shown on the right. Numbers on the column of the illustrative stamp represent passage numbers from which the colonies were isolated. Passage numbers 10 to passage numbers 80 and continuing on the fourth column with strains isolated after 90 passages and 100 passages respectively are shown. The second and third column show the same orientation. A–B–C, were from triplicates of co-culture evolved lines whereas D–E–F in brackets were from controls. For example lines A(D) means that colonies were either from flask A (co-culture evolved) or D (controls). “Anc” shows six positions of replicate colonies of the ancestral strain used as a reference
Resistance to surfactants
We further assessed the impacts of a bacterial competition on the cell wall reconstruction and cell membrane stability. We used a known surfactant, sodium dodecyl-sulfate (SDS), which damages cell membrane structures and thereby affects carbon metabolism [30, 31]. Results showed that co-culture evolved yeasts (grown in the presence of bacteria) evolved protection against its effects (Fig. 3). As low as 0.016% of SDS inhibited control strains, whereas the co-culture evolved yeasts were tolerant up to 0.018%. Strains isolated from 10 to 80 passages were the most resistant to 0.018% SDS as compared to their counterparts evolved for more generations suggesting a loss in the trait with increasing time of coevolution. It is noteworthy that none of the cells from either treatments survived 0.02% of SDS. Interestingly, we noted that cells from control-evolved lines were poor in stress resistance as evident in the presence of SDS as well as noted above in the presence of ethanol stress.
Fig. 3.
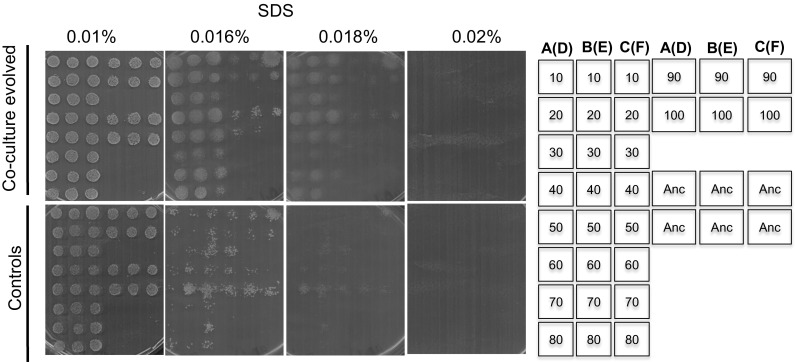
Tolerance of evolved strains to surfactant, SDS. Tolerance to surfactant of strains evolved in the presence or absence of bacteria, co-culture evolved and control respectively, with reference to their ancestor, was determined by spotting cells from stationary phase cultures on different concentrations of SDS. Growth was scored after 5 days. All strains grown in YNB without SDS are included in Fig 4. The results show that competition against bacteria protects the cells against the cell wall and membrane damaging surfactant. The arrangement of the colonies on the plates is shown on the right. Numbers on the column of the illustrative stamp represent passage numbers from which the colonies were isolated. Passage numbers 10 to passage numbers 80 and continuing on the fourth column with strains isolated after 90 passages and 100 passages respectively are shown. The second and third column shows the same orientation. A–B–C are from triplicate co-culture evolved lines whereas D–E–F in brackets are from controls. For example lines A(D) means that colonies were either from flask A (co-culture evolved) or D (controls). “Anc” shows six positions of replicate colonies of the ancestral strain used as a reference.
Oxidative stress
Metabolic processes generate reactive oxygen species, either as peroxides, superoxide, and hydroxyl radicals [32]. Thus, we sought to determine the sensitivity upon exposure to H2O2 as well as to a superoxide generating drug, menadione. In co-culture-evolved strains, there was a clear loss of resistance to H2O2 with increasing exposure to bacteria, for example strains from passages 10–40 were resistant whereas strains isolated 50–100 passages did not survive exposure to 3 mM. Most control strains were more tolerant to 3 mM H2O2 (Fig. 4).
Fig. 4.

Tolerance of evolved strains to oxidative stress in H2O2. Oxidative stress of strains evolved in the presence or absence of bacteria, co-culture evolved and control respectively, with reference to their ancestor, was determined by spotting cells from stationary phase cultures on different concentrations of H2O2. Growth was scored after 3 days. The results show that competition against bacteria led to the evolution of strains sensitive to oxidative stress. The arrangement of the colonies on the plates is shown on the right. Numbers on the column of the illustrative stamp represent passage numbers from which the colonies were isolated. Passage numbers 10 to passage numbers 80 and continuing on the fourth column with strains isolated after 90 passages and 100 passages respectively are shown. The second and third column shows the same orientation. A–B–C are from triplicate co-culture evolved lines whereas D–E–F in brackets are from controls. For example lines A(D) means that colonies were either from flask A (co-culture evolved) or D (controls). “Anc” shows six positions of replicate colonies of the ancestral strain used as a reference.
Stress resistance to menadione was more pronounced in the controls as compared to the co-culture evolved counterparts at 0.18 mM of menadione (Fig. 5). In summary, we noted a decreased resistance to oxidative stress in co-culture-evolved yeasts as compared to their controls. It is common that emergence of adaptive phenotypes in nature may also come at the cost of reduced fitness in other environments [33, 34]. These results suggested that adaptively evolved resistant strategies to thermal and ethanol stress as well as resistance to SDS did not cross protect the cells from oxidative stress.
Fig. 5.

Tolerance of evolved strains to menadione oxidative stress. Oxidative stress of strains evolved in the presence or absence of bacteria, co-culture evolved and control respectively, with reference to their ancestor, was determined by spotting cells from stationary phase cultures on different concentrations of menadione. Growth was scored after 3 days. Strains evolved in the presence of bacteria became sensitive to menadione. All strains grown in YNB without menadione are included in Fig 4. The arrangement of the colonies on the plates is shown on the right. Numbers on the column of the illustrative stamp represent passage numbers from which the colonies were isolated. Passage numbers 10 to passage numbers 80 and continuing on the fourth column with strains isolated after 90 passages and 100 passages respectively are shown. The second and third column shows the same orientation. A–B–C are from triplicate co-culture evolved lines whereas D –E–F in brackets are from controls. For example lines A(D) means that colonies were either from flask A (co-culture evolved) or D (controls). “Anc” shows six positions of replicate colonies of the ancestral strain used as a reference
Fermentative ability
Strains isolated after 50 passages from evolution lines B and C (co-culture evolved) were the best performing under thermal as well as under stressful ethanol conditions. These traits are ideal for a biofuel production strain. We, therefore, thought to investigate if the acquired novel stress resistance did not affect the fermentative capacity of these isolates. Otherwise their use in biofuel production will not be possible. Since these strains grew slowly at 39 °C (Fig. 1), we chose to test these strains at a lower temperature, 37 °C. Our results show that on average, growth rates of isolates from evolution line C were 1.30 times faster than that of the ancestral strain whereas those from evolution lines B were 0.95 slower (Fig. 6a). Isolates from lines C produced on average 1.27 times higher ethanol than the ancestral strain (Fig. 6a). In addition, these isolates (lines C) consumed glucose on average 1.66 times faster and accumulated ethanol at a rate on average 2.22 times faster (Fig. 6b) than the ancestral strain. On the contrary they accumulated glycerol on average 0.47 times lower than the ancestral strain (Fig. 6c). In general, both lines B and C evolved to produce more ethanol at the expense of a lower biomass as compared to their ancestor. The fermentative capacity of these co-culture evolved strains suggests that these strains are applicable as bioethanol production strains as the fermentative ability was retained. In addition, our results show that the fermentative ability was also elevated, making the strains more ideal for the biofuel production process because the overall productivity and ethanol titers per unit volume of substrate consumed during the fermentation process will be elevated.
Fig. 6.
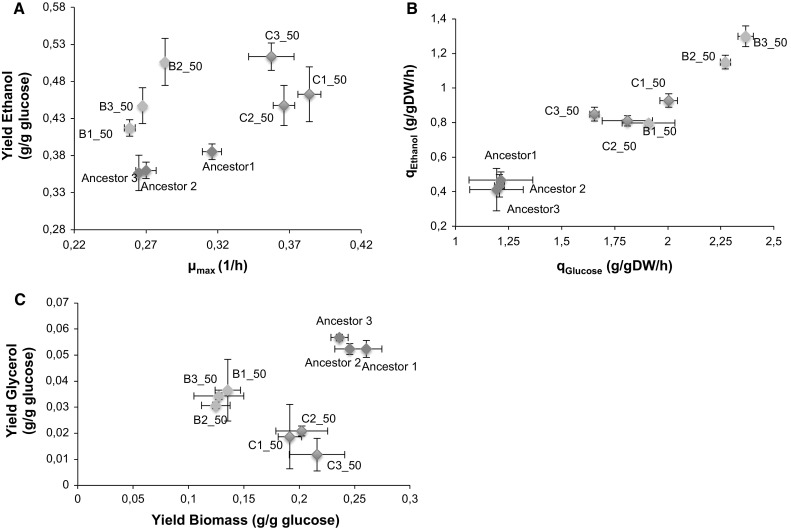
Fermentative capacity of the thermotolerant strains. The plots show that A growth rates of isolates from evolution line C (blue marker style) were faster than the ancestral strain but those from line B (green marker style) were not different from that of the ancestor (red marker style). Both evolved lines produced more ethanol that their ancestor, B the maximum glucose consumption rate and ethanol production rates were higher than the ancestral strain but C accumulated lower glycerol and biomass
Large-scale chromosomal rearrangements
Although we had established the evolution of stress tolerance traits important for bioethanol production, as the consequence of a cross-kingdom competition experiment, the molecular mechanisms behind the observed phenotypes warranted further investigation. We, therefore, studied the molecular polymorphisms by examining the electrophoretic karyotypes of derived strains using PFGE. The publicly available ancestral strain used in this study, L. thermotolerans CBS6340, S. pombe and S. cerevisiae strains were used as references to estimate chromosomes size. It is noteworthy that the type strain, CBS6340 karyotype reported elsewhere [35] does not have the same karyotype with our CBS6340 parental strain used in this study as the size of the biggest chromosomes is about 2.7 Mb in the former whereas in the latter we found it to be approximately 2.1 Mb (Fig. 7). In addition, the numbers and size of chromosomes reported elsewhere [36] do not tally among these studies probably accounting for the difference in phenotypes among natural L. thermotolerans strains. Our results show that the control strains did not exhibit any change in karyotypes during the evolution experiment. Their co-culture evolved counterparts, however, lost the biggest chromosomal band evident in the ancestral lane (about 2.1 Mb) in the strain isolated after 10 passages in all three parallel lines. The biggest band in these strains now corresponds to about 1.8 Mb in size. The evolved karyotype did not change thereafter until the end of the evolution experiment (Fig. 7).
Fig. 7.

Investigation of molecular changes based on electrophoretic karyotypes. Karyotypes of strains isolated after every 80 generations from co-culture evolved lines (Line A–Line C), and control lines (D–F) are shown. Extrapolation from the migration of marker strains of S. pombe (SJA148) and S. cerevisiae (S288C) was carried out to estimate the size of the novel bands at approximately 1.8 Mb (red arrow)
Discussion
In nature, yeasts like any other organisms compete for resources amongst either themselves or other yeasts of different species as well as against co-habitants from across the kingdom, such as bacteria. Competition is a crucial driver of evolution [37]. Interspecific competition is one such ecological force that reduces population density [37, 38], which subsequently reduces the accumulation of adaptive mutations increasing the risks of extinction [38, 39]. On the other hand, such an increased strength of selection pressure increases the rate of evolution [40–42].
Based on the fact that organisms adapt to environments they live in and may as well cope with extreme environments they may encounter in their evolutionary life history [43], we created an ecological battlefield by subjecting yeasts to the competition against bacteria with a goal to develop thermotolerant yeasts without compromising their fermentative capabilities. We hypothesized that probing yeasts would force them to define mechanisms to adapt as well as to out-grow competitors; such as efficient resource utilization, ability to tolerate a variety of stresses associated with such a habitat, and the ability to mount a refined niche defense strategy in quest to dominate habitats. A comprehensive review of how microorganisms aggressively deploy niche-dominating strategies is available elsewhere [44].
Competition is known to increase rates of evolution by increasing a selective pressure that pushes organisms to explore multiple paths to survive [40, 41, 45]. In our study, we forced yeasts to evolve survival responses against the presence and activities of bacteria. A basic principle of this selection pressures is based on the relatively faster growth rates of bacteria when compared to that of yeasts coupled to their faster consumption of the available carbon sources, endowing them with a competitive advantage. Among other studied selection pressures exerted by bacteria is the alteration of environmental pH, secretion of antifungals and chitinases and other growth inhibitory molecules [46, 47].
We show that in response to an increasing bacterial selection pressure, yeasts evolved elevated thermotolerance as well as ethanol tolerance and resistance to surfactants. There is an increase in ethanol production accompanied by an increased ethanol tolerance, however, based on our data available this is an observation is not necessarily a causative factor. We also show that adaptive evolution experiments using poorly studied and less exploited non-conventional yeasts may yield insights in the study of stress resistance and development of stress resistant strains of other species.
Conventional industrial yeast production strains are exposed to several stressful environments such as low and high temperatures, exposure to ethanol, osmotic stress, and oxidative stress during the preparation as starter cultures as well as during fermentation reviewed in [48]. Strain robustness in such stressful environments is essential for increasing productivity of fermentation, which often culminates in an economically viable bioprocess. There has been extensive research going on to ameliorate the negative effects of sensitivity to stress in conventional yeasts [49, 50]. This research could benefit from the knowledge derived from the studies of non-conventional yeasts, which could point towards novelties in stress resistance strategies when compared to conventional yeasts.
The evolution of a phenotype protecting against thermal stress when yeasts were co-cultured with bacteria suggests that our method is an attractive, inexpensive and a simple alternative to develop yeast strains with robust resistance to a number of stressful conditions. The emergence of stress tolerance phenotypes in all three independent evolution lines, which was not evident in the control strains, is suggestive of an adaptive and not transient phenotype. This is a key factor for reproducibility and adoption of the method. A more pronounced thermotolerance with increasing selection pressure as evident in the terminally evolved strains, suggestive of a stepwise evolution of the mechanism involved, makes this method more attractive because there is a probable room for further improvement thermotolerance improvement.
The exposure to a bacterial selection pressure cross-protected the evolved strains from ethanol and surfactants. These results are in agreement with reported cases, where stressors are known to induce cross-protection against an impending stressor [51–53]. A functional overlap of responses behind thermotolerance and ethanol tolerance observed, is not surprising as they are in agreement with the use of similar stress response mechanisms under these two scenarios in yeast [29, 54]. Both stressors cause similar plasma membrane protein changes. In addition, heat-shock proteins involved in mounting stress responses against these two stressors also overlap [29, 54]. In addition, we noted that in addition co-culture evolved strains were protected against surfactant, SDS, whose response mechanism is based on membrane structures [30, 31].
One of the drawbacks of experimental evolution is the emergence of trade-offs, which negatively impact the robustness and applicability of the developed strains [55]. There was a loss of resistance to oxidative stress, which may be considered as an undesirable trade-off when the method is transferred for the development of S. cerevisiae strains for biofuels production. This is because cells are in constant exposure to oxygen during the propagation stages that induce oxidative stress [56]. Current strategies to reduce the toxicity of the lignocellulosic feedstocks reviewed in [20] may also mitigate the possible effects.
We speculate that the presence and activity of bacteria could have induced adaptive changes either in the plasma membrane protein structures and permeability or an altered expression of either plasma membrane associated genes and heat shock protein family of genes. An investigation of the molecular karyotypes of evolved strains revealed a significant polymorphism. Although changes involving large-scale genomic rearrangements allow a swift mechanism of evolutionary adaptation [57], by regulating the expression of several genes in a metabolic pathway as well as gene dosage [57–59], the link between rearrangements and the observed phenotypes remain to be proven. In our previous studies, we noted the emergence of new metabolic traits due to a large-scale genomic rearrangement following a segmental duplication and translocation event. Whole genome sequencing revealed the duplication of vital genes involved in osmotolerance, DNA replication stress, and thermotolerance [16]. In addition, several other yeasts with large-scale rearrangements were reported [14, 60]. Our method does not impair growth and fermentation of the yeasts, and thus it is suitable for industrial strain development. In conclusion, our study highlights the importance of ecological battlefields for the induction of adaptive phenotypes. Interspecific competitions in nature are often aggressive [44], so microorganisms reshape molecular mechanisms to avoid being annihilated or develop new strategies to dominate the niches. The deployment of such versatile strategies may also entail the protection against other types of stress not tested or contemplated in this work.
We propose the use of this method as a valuable approach to develop robust stress resistant industrial strains. Our tool mimicks the natural environment characteristic of cross-kingdom competition for sugars in nature when fruits ripen, every autumn, availing excess sugars for a short period of time. Although complex interactions among a multitudes of microorganism exist in nature, here we created a less complex competitive environment in which yeasts were allowed to sequentially encounter each of the six bacterial species used in this study, one at a time, for a time long enough to effect an evolutionary change. The order with which yeasts encountered bacteria was assumed to be a gradient of strength of selection, from weak (ethanol sensitive bacteria) to strong (ethanol tolerant bacteria). A weak selection pressure during the early stages of evolution was ascertained to allow the emergence of more genetically diverse variants, an attribute important for organisms to explore a multiple paths to adaptation as well as avoiding populations being driven to extinction when the solution becomes inaccessible; associated with strong selection pressure as reviewed elsewhere [22]. Nutrient limitation, exhaustion and end product toxicity characteristic of the stationary phase of growth have been shown to force yeasts to adaptively evolve mechanisms to either utilise alternative or limiting nutrients reviewed in [22]. To exclude such possible selection pressures and outcomes that could mask our bacterial competition selection pressure hypothesis set in our development tool, transfer of cultures was carried out before reaching the stationary phases. This allowed us to explore the evolvability of yeasts only during the exponential phases, which is a competitive growth against bacteria in the presence of a carbon source depreciating over the course of time. The use of antibiotic drugs to kill the surviving bacteria before transfer into fresh media was an important step to avoid emergence of antibiotic resistant bacteria. Antibiotics were added even in the absence of a bacterial challenge (controls) so that the consequence of a bacterial selection pressure on evolving yeast populations instead of unknown selection pressures could be investigated. This is due to the fact that it is characteristic of yeasts to undergo metabolic shifts even in a simple laboratory environment, as they tend to maximize their survival and reproduction.
Until now, thermal stress tolerant yeasts have been developed through the exposure to a mild form and gradually increasing thermal stress, which unfortunately, negatively affects the productivity of the production strains due to major trade-offs. Our tool suggests that improvement of the production capacity as well as cross-protection from other environmental stresses encountered during production is possible. Both of these attributes add up to the most desirable phenotypes of industrial yeasts. It should be noted that for a specific application of thermotolerance strains, such as bioethanol production from inexpensive lignocellulosic feedstocks, further studies are need to investigate if the acquisition of thermotolerance affects the utilisation of alternative carbon sources (xylose, arabinose, galactose and mannitol), abundant in lignocellulosic hydrolysates.
Conclusion
Our novel two-species cross-kingdom competition strategy is an attractive tool to develop stress tolerant industrial strains.
Acknowledgements
We would like to dedicate this work to our late friend, teacher and mentor, Professor Jure Piskur. Our continued respect and admiration of him is the reason why this study was continued.
Author contributions
Conceptualization: JP CC. Funding acquisition: JP CC WK. Investigation: NZ OPI. Methodology: NZ OPI. Project administration: WK. Resources: JP WK. Supervision: JP CC WK. Writing—original draft: NZ. Writing—review and editing: NZ CC OPI WK.
Funding
This work was funded by the Marie Curie, Cornucopia Project (264717), Fysiografen and Lindström Foundations.
Compliance with ethical standards
Conflict of interest
None declared.
References
- 1.Steensels J, Snoek T, Meersman E, Nicolino MP, Voordeckers K, Verstrepen KJ. Improving industrial yeast strains: exploiting natural and artificial diversity. FEMS Microbiol Rev. 2014;38(5):947–995. doi: 10.1111/1574-6976.12073. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 2.Matsushita K, Azuma Y, Kosaka T, Yakushi T, Hoshida H, Akada R, et al. Genomic analyses of thermotolerant microorganisms used for high-temperature fermentations. Biosci Biotechnol Biochem. 2016;80(4):655–668. doi: 10.1080/09168451.2015.1104235. [DOI] [PubMed] [Google Scholar]
- 3.Steensels J, Meersman E, Snoek T, Saels V, Verstrepen KJ. Large-scale selection and breeding to generate industrial yeasts with superior aroma production. Appl Environ Microbiol. 2014;80(22):6965–6975. doi: 10.1128/AEM.02235-14. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 4.Querol A, Fernández-Espinar MT, del Olmo M, Barrio E. Adaptive evolution of wine yeast. Int J Food Microbiol. 2003;86(1–2):3–10. doi: 10.1016/s0168-1605(03)00244-7. [DOI] [PubMed] [Google Scholar]
- 5.Caspeta L, Chen Y, Ghiaci P, Feizi A, Buskov S, Hallström BM, et al. Altered sterol composition renders yeast thermotolerant. Science. 2014;346(6205):75–78. doi: 10.1126/science.1258137. [DOI] [PubMed] [Google Scholar]
- 6.Trollmo C, André L, Blomberg A, Adler L. Physiological overlap between osmotolerance and thermotolerance in Saccharomyces cerevisiae. FEMS Microbiol Lett. 1988;56(3):321–325. [Google Scholar]
- 7.Vriesekoop F, Pamment NB. Acetaldehyde addition and pre-adaptation to the stressor together virtually eliminate the ethanol-induced lag phase in Saccharomyces cerevisiae. Lett Appl Microbiol. 2005;41(5):424–427. doi: 10.1111/j.1472-765X.2005.01777.x. [DOI] [PubMed] [Google Scholar]
- 8.Caspeta L, Nielsen J. Thermotolerant yeast strains adapted by laboratory evolution show trade-off at ancestral temperatures and preadaptation to other stresses. mBio. 2015;6(4):e00431. doi: 10.1128/mBio.00431-15. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 9.Lindquist S. The heat-shock response. Annu Rev Biochem. 1986;55:1151–1191. doi: 10.1146/annurev.bi.55.070186.005443. [DOI] [PubMed] [Google Scholar]
- 10.Ishchuk OP, Voronovsky AY, Abbas CA, Sibirny AA. Construction of Hansenula polymorpha strains with improved thermotolerance. Biotechnol Bioeng. 2009;104(5):911–919. doi: 10.1002/bit.22457. [DOI] [PubMed] [Google Scholar]
- 11.Jeffries TW, Jin YS. Ethanol and thermotolerance in the bioconversion of xylose by yeasts. Adv Appl Microbiol. 2000;47:221–268. doi: 10.1016/s0065-2164(00)47006-1. [DOI] [PubMed] [Google Scholar]
- 12.Steinmetz LM, Sinha H, Richards DR, Spiegelman JI, Oefner PJ, McCusker JH, et al. Dissecting the architecture of a quantitative trait locus in yeast. Nature. 2002;416(6878):326–330. doi: 10.1038/416326a. [DOI] [PubMed] [Google Scholar]
- 13.Genolevures C, Souciet JL, Dujon B, Gaillardin C, Johnston M, Baret PV, et al. Comparative genomics of protoploid Saccharomycetaceae. Genome Res. 2009;19(10):1696–1709. doi: 10.1101/gr.091546.109. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 14.Zhou N, Bottagisi S, Katz M, Schacherer J, Friedrich A, Gojkovic Z, et al. Yeast-bacteria competition induced new metabolic traits through large-scale genomic rearrangements in Lachancea kluyveri. FEMS Yeast Res. 2017 doi: 10.1093/femsyr/fox060. [DOI] [PubMed] [Google Scholar]
- 15.Zhou N, Katz M, Knecht W, Compagno C, Piskur J. Genome dynamics and evolution in yeasts: a long-term yeast-bacteria competition experiment. PLoS One. 2018;13(4):e0194911. doi: 10.1371/journal.pone.0194911. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 16.Zhou N, Swamy KB, Leu J, McDonald MJ, Galafassi S, Compagno C, et al. Coevolution with bacteria drives the evolution of aerobic fermentation in Lachancea kluyveri. PLoS One. 2017;12(3):e0173318. doi: 10.1371/journal.pone.0173318. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 17.Zhang QG, Buckling A. Antagonistic coevolution limits population persistence of a virus in a thermally deteriorating environment. Ecol Lett. 2011;14(3):282–288. doi: 10.1111/j.1461-0248.2010.01586.x. [DOI] [PubMed] [Google Scholar]
- 18.Hottiger T, Schmutz P, Wiemken A. Heat-induced accumulation and futile cycling of trehalose in Saccharomyces cerevisiae. J Bacteriol. 1987;169(12):5518–5522. doi: 10.1128/jb.169.12.5518-5522.1987. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 19.Souza CS, Oliveira KF, Trevisan HC, Laluce C (2007) A strategy to compare yeast strains and improve cell survival in ethanol production processes above 30 °C. Commun Curr Res Educ Top Trends Appl Microbiol 410–417. http://www.formatex.org/microbio/pdf/Pages410-417.pdf
- 20.Parawira W, Tekere M. Biotechnological strategies to overcome inhibitors in lignocellulose hydrolysates for ethanol production: review. Crit Rev Biotechnol. 2010;31(1):20–31. doi: 10.3109/07388551003757816. [DOI] [PubMed] [Google Scholar]
- 21.Hahn-Hagerdal B, Karhumaa K, Fonseca C, Spencer-Martins I, Gorwa-Grauslund MF. Towards industrial pentose-fermenting yeast strains. Appl Microbiol Biotechnol. 2007;74(5):937–953. doi: 10.1007/s00253-006-0827-2. [DOI] [PubMed] [Google Scholar]
- 22.Elena SF, Lenski RE. Evolution experiments with microorganisms: the dynamics and genetic bases of adaptation. Nat Rev Genet. 2003;4(6):457–469. doi: 10.1038/nrg1088. [DOI] [PubMed] [Google Scholar]
- 23.Zeyl C. Experimental evolution with yeast. FEMS Yeast Res. 2006;6(5):685–691. doi: 10.1111/j.1567-1364.2006.00061.x. [DOI] [PubMed] [Google Scholar]
- 24.Peterson CL, Herskowitz I. Characterization of the yeast SWI1, SWI2, and SWI3 genes, which encode a global activator of transcription. Cell. 1992;68(3):573–583. doi: 10.1016/0092-8674(92)90192-f. [DOI] [PubMed] [Google Scholar]
- 25.van Hoek P, van Dijken JP, Pronk JT. Regulation of fermentative capacity and levels of glycolytic enzymes in chemostat cultures of Saccharomyces cerevisiae. Enzyme Microb Technol. 2000;26(9–10):724–736. doi: 10.1016/s0141-0229(00)00164-2. [DOI] [PubMed] [Google Scholar]
- 26.Kurtzman CP. Phylogenetic circumscription of Saccharomyces, Kluyveromyces and other members of the Saccharomycetaceae, and the proposal of the new genera Lachancea, Nakaseomyces, Naumovia, Vanderwaltozyma and Zygotorulaspora. FEMS Yeast Res. 2003;4(3):233–245. doi: 10.1016/S1567-1356(03)00175-2. [DOI] [PubMed] [Google Scholar]
- 27.Ganter P. Yeast and invertebrate associations. In: Rosa C, Péter G, editors. Biodiversity and ecophysiology of yeasts. The yeast handbook. Berlin: Springer; 2006. pp. 303–370. [Google Scholar]
- 28.Institute WFB. Westerdijk Fungal Biodiversity Institute; (13 December 2016). http://www.westerdijkinstitute.nl/Collections/Biolomics.aspx?Table=CBS%20strain%20database. Accessed 10 June 2018
- 29.Piper PW. The heat shock and ethanol stress responses of yeast exhibit extensive similarity and functional overlap. FEMS Microbiol Lett. 1995;134(2–3):121–127. doi: 10.1111/j.1574-6968.1995.tb07925.x. [DOI] [PubMed] [Google Scholar]
- 30.Sirisattha S, Momose Y, Kitagawa E, Iwahashi H. Toxicity of anionic detergents determined by Saccharomyces cerevisiae microarray analysis. Water Res. 2004;38(1):61–70. doi: 10.1016/j.watres.2003.08.027. [DOI] [PubMed] [Google Scholar]
- 31.Igual JC, Johnson AL, Johnston LH. Coordinated regulation of gene expression by the cell cycle transcription factor Swi4 and the protein kinase C MAP kinase pathway for yeast cell integrity. EMBO J. 1996;15(18):5001–5013. [PMC free article] [PubMed] [Google Scholar]
- 32.Jamieson DJ. Saccharomyces cerevisiae has distinct adaptive responses to both hydrogen peroxide and menadione. J Bacteriol. 1992;174(20):6678–6681. doi: 10.1128/jb.174.20.6678-6681.1992. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 33.Wenger JW, Piotrowski J, Nagarajan S, Chiotti K, Sherlock G, Rosenzweig F. Hunger artists: yeast adapted to carbon limitation show trade-offs under carbon sufficiency. PLoS Genet. 2011;7(8):e1002202. doi: 10.1371/journal.pgen.1002202. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 34.Cooper VS. The study of microbial adaptation by long-term experimental evolution. Washington: Evolution of Microbial Pathogens ASM Press; 2006. [Google Scholar]
- 35.Naumova ES, Serpova EV, Naumov GI. Molecular systematics of Lachancea yeasts. Biochem Biokhimiia. 2007;72(12):1356–1362. doi: 10.1134/s0006297907120097. [DOI] [PubMed] [Google Scholar]
- 36.Souciet J-L, Aigle M, Artiguenave F, Blandin G, Bolotin-Fukuhara M, Bon E, et al. Genomic exploration of the hemiascomycetous yeasts: 1. A set of yeast species for molecular evolution studies1. FEBS Lett. 2000;487(1):3–12. doi: 10.1016/s0014-5793(00)02272-9. [DOI] [PubMed] [Google Scholar]
- 37.Little AE, Robinson CJ, Peterson SB, Raffa KF, Handelsman J. Rules of engagement: interspecies interactions that regulate microbial communities. Annu Rev Microbiol. 2008;62:375–401. doi: 10.1146/annurev.micro.030608.101423. [DOI] [PubMed] [Google Scholar]
- 38.Gause GF, Witt AA. Behavior of mixed populations and the problem of natural selection. Am Nat. 1935;69(725):596–609. [Google Scholar]
- 39.Martin PR, Martin TE. Ecological and fitness consequences of species coexistence: a removal experiment with wood warblers. Ecology. 2001;82(1):189–206. [Google Scholar]
- 40.Martin MJ, Perez-Tome JM, Toro MA. Competition and genotypic variability in Drosophila melanogaster. Heredity. 1988;60(Pt 1):119–123. doi: 10.1038/hdy.1988.17. [DOI] [PubMed] [Google Scholar]
- 41.Birch LC. Selection in Drosophila pseudoobscura in relation to crowding. Evolution. 1955;9(4):389–399. [Google Scholar]
- 42.Barrick JE, Lenski RE. Genome-wide mutational diversity in an evolving population of Escherichia coli. Cold Spring Harb Symp Quant Biol. 2009;74:119–129. doi: 10.1101/sqb.2009.74.018. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 43.Hereford J. A quantitative survey of local adaptation and fitness trade-offs. Am Nat. 2009;173(5):579–588. doi: 10.1086/597611. [DOI] [PubMed] [Google Scholar]
- 44.Cray JA, Bell ANW, Bhaganna P, Mswaka AY, Timson DJ, Hallsworth JE. The biology of habitat dominance; can microbes behave as weeds? Microb Biotechnol. 2013;6(5):453–492. doi: 10.1111/1751-7915.12027. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 45.Barrick JE, Lenski RE. Genome dynamics during experimental evolution. Nat Rev Genet. 2013;14(12):827–839. doi: 10.1038/nrg3564. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 46.Chernin LS, De la Fuente L, Sobolev V, Haran S, Vorgias CE, Oppenheim AB, et al. Molecular cloning, structural analysis, and expression in Escherichia coli of a chitinase gene from Enterobacter agglomerans. Appl Environ Microbiol. 1997;63(3):834–839. doi: 10.1128/aem.63.3.834-839.1997. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 47.Beckner M, Ivey ML, Phister TG. Microbial contamination of fuel ethanol fermentations. Lett Appl Microbiol. 2011;53(4):387–394. doi: 10.1111/j.1472-765X.2011.03124.x. [DOI] [PubMed] [Google Scholar]
- 48.Attfield PV. Stress tolerance: the key to effective strains of industrial baker’s yeast. Nat Biotechnol. 1997;15(13):1351–1357. doi: 10.1038/nbt1297-1351. [DOI] [PubMed] [Google Scholar]
- 49.Zhao XQ, Bai FW. Mechanisms of yeast stress tolerance and its manipulation for efficient fuel ethanol production. J Biotechnol. 2009;144(1):23–30. doi: 10.1016/j.jbiotec.2009.05.001. [DOI] [PubMed] [Google Scholar]
- 50.Watanabe T, Srichuwong S, Arakane M, Tamiya S, Yoshinaga M, Watanabe I, et al. Selection of stress-tolerant yeasts for simultaneous saccharification and fermentation (SSF) of very high gravity (VHG) potato mash to ethanol. Biores Technol. 2010;101(24):9710–9714. doi: 10.1016/j.biortech.2010.07.079. [DOI] [PubMed] [Google Scholar]
- 51.Berry DB, Gasch AP. Stress-activated genomic expression changes serve a preparative role for impending stress in yeast. Mol Biol Cell. 2008;19(11):4580–4587. doi: 10.1091/mbc.E07-07-0680. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 52.Costa V, Reis E, Quintanilha A, Moradas-Ferreira P. Acquisition of ethanol tolerance in Saccharomyces cerevisiae: the key role of the mitochondrial superoxide dismutase. Arch Biochem Biophys. 1993;300(2):608–614. doi: 10.1006/abbi.1993.1084. [DOI] [PubMed] [Google Scholar]
- 53.McAlister L, Finkelstein DB. Heat shock proteins and thermal resistance in yeast. Biochem Biophys Res Commun. 1980;93(3):819–824. doi: 10.1016/0006-291x(80)91150-x. [DOI] [PubMed] [Google Scholar]
- 54.Auesukaree C, Damnernsawad A, Kruatrachue M, Pokethitiyook P, Boonchird C, Kaneko Y, et al. Genome-wide identification of genes involved in tolerance to various environmental stresses in Saccharomyces cerevisiae. J Appl Genet. 2009;50(3):301–310. doi: 10.1007/BF03195688. [DOI] [PubMed] [Google Scholar]
- 55.Bennett AF, Dao KM, Lenski RE. Rapid evolution in response to high-temperature selection. Nature. 1990;346(6279):79–81. doi: 10.1038/346079a0. [DOI] [PubMed] [Google Scholar]
- 56.Spencer J, Phister TG, Smart KA, Greetham D. Tolerance of pentose utilising yeast to hydrogen peroxide-induced oxidative stress. BMC Res Notes. 2014;7:151. doi: 10.1186/1756-0500-7-151. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 57.Hewitt SK, Donaldson IJ, Lovell SC, Delneri D. Sequencing and characterisation of rearrangements in three S. pastorianus strains reveals the presence of chimeric genes and gives evidence of breakpoint reuse. PLoS One. 2014;9(3):e92203. doi: 10.1371/journal.pone.0092203. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 58.Chang SL, Lai HY, Tung SY, Leu JY. Dynamic large-scale chromosomal rearrangements fuel rapid adaptation in yeast populations. PLoS Genet. 2013;9(1):e1003232. doi: 10.1371/journal.pgen.1003232. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 59.Koszul R, Dujon B, Fischer G. Stability of large segmental duplications in the yeast genome. Genetics. 2006;172(4):2211–2222. doi: 10.1534/genetics.105.048058. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 60.Zhou N (2015) Carbon metabolism in non-conventional yeasts: biodiversity, origins of aerobic fermentation and industrial applications. (Doctoral Thesis)