

Abstract
In this issue of Immunity, Graef et al. demonstrate self-renewal and multipotency of a single CD62L+ memory T cell across serial adoptive transfers and infection-driven re-expansions, providing evidence of true stemness within the T cell memory compartment.
Long-term maintenance of tissue homeostasis relies on somatic stem cells, which ensure efficient replacement of short-lived, specialized cells while maintaining themselves through a process of self-renewal (Simons and Clevers, 2011). Analogous to other organ systems, life-long immunological memory is thought to depend on stem cell-like memory cells, as the ability of hematopoietic stem cells (HSCs) to provide for antigen-specific immunity is constrained by stochastic recombination of the T cell receptor and thymic involution in early adulthood (Fearon et al., 2001). Over the past decade, it has become increasingly recognized that memory T cells display core molecular signatures and functional attributes characteristic of stem cells (Gattinoni et al., 2012). For instance, memory T cells share a partially conserved transcriptional profile with HSCs and, similar to stem cells, they can undergo asymmetric division and activate telomerase to maintain telomere length and replicative potential (Gattinoni et al., 2012). Furthermore, several signaling pathways regulating stem cell self-renewal have been found to be active in T cells to promote memory and limit effector T cell differentiation (Gattinoni et al., 2012). However, self-renewal and multipotency, the defining qualities of stem cells, have only been inferred by reconstitution studies analyzing the developmental potential of T cell populations rather than studying individual cells, which is essential to determining the true stemness of a given cell type. These population-based studies have revealed a hierarchical organization of the T cell memory compartment. T memory stem cells (TSCM), a subset of cells displaying a naïve-like phenotype (CD44− CD62L+ in mice; CD45RA+ CD62L+ in human and nonhuman primates) together with the expression of the memory markers interleukin-2 receptor (IL2R) β and the chemokine C-X-C motif receptor 3 (CXCR3) have been shown to be the most undifferentiated memory subset (Gattinoni et al., 2012). In this hierarchical structure, CD62L+ central memory (TCM) cells are located at an intermediate position between TSCM cells and CD62L− effector memory (TEM) cells, which are committed progenitor cells prone to effector (TEFF) differentiation (Gattinoni et al., 2012). While these studies focusing on population analyses helped shape our current understanding of T cell differentiation, they cannot unequivocally establish the developmental potential of individual cells.
In this issue of Immunity, Graef et al. evaluate the stemness of the CD8+ memory T cell compartment by measuring self-renewal and multipotency of a single TCM cell throughout a series of in vivo clonogenic assays. It was an extraordinary undertaking, performing a serial tri-generational single cell transfer, an experiment that was never attempted before for any type of mammalian stem cells. The authors found that following pathogen-stimulation, a single TCM cell could propagate itself while giving rise to a diverse progeny comprising TEM and TEFF cells. Remarkably, progeny size, diversity and long-term maintenance were highly reproducible across serial transfers, demonstrating stem cell behavior of individual TCM cells (Figure 1). In keeping with the stem cell parallel, it was striking to observe that a progeny from single TCM cells could be detected in about 20% of mice, a recovery rate comparable to the reconstitution efficiency originally reported for a single HSC (Osawa et al., 1996). A hallmark of adult stem cells is the ability to confer functional tissue reconstitution. Graef et al. showed that minuscule numbers of tertiary TCM cells, whose ancestors had passed through three generations of single T cells – a naïve T cell, a primary TCM cell and a secondary TCM cell, were capable of reconstituting full immunocompetence and protecting severely immunodeficient hosts from a lethal bacterial challenge (Figure 1). Collectively, these findings further buttress the conclusion that CD62L+ memory T cells functionally behave as adult stem cells.
Figure 1: TCM cells function as adult stem cells.
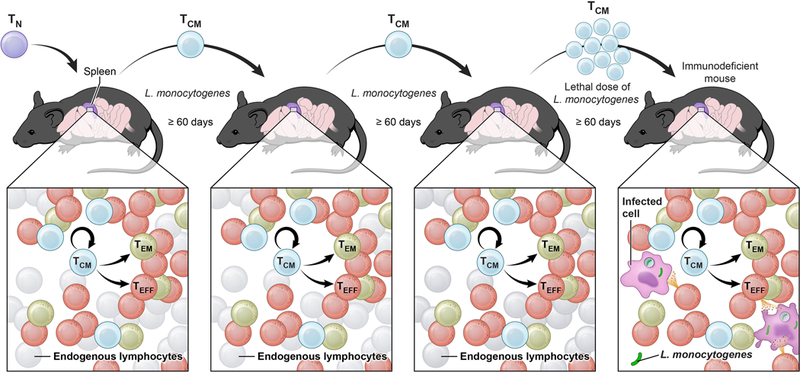
Self-renewal and multipotency of a single central memory T cell (TCM) across serial adoptive transfers and infection-driven re-expansions ensure full immunocompetence. TN, naïve T cell; TEM, effector memory T cell; TEFF, effector T cell.
It is currently unclear whether TCM cells have a persistent broad developmental potential or if they accumulate inherited restrictions as a result of their replicative history. The authors concluded in favor of the first hypothesis, as they observed no correlation between the degree of expansion that had originated from a single ancestor and the proliferation of its TCM daughter cells. However, it should be noted that the degree of expansion of a single cell progeny does not necessarily reflect the replicative history of TCM daughters, as the size of large colonies is primarily driven by TEM and TEFF cells. Conversely, paralleling other types of adult stem cells, whose stemness is often preserved by enforcing cellular quiescence (Simons and Clevers, 2011), the reconstitution potential of individual TCM cells might also be conditioned by their past proliferation. Indeed, the authors observed that about 20% of single TCM-derived progenies that were detectable at the peak of expansion failed to persist for the long-term, indicating a stochastic loss of stemness, possibly reflecting heterogeneity of replicative history in the TCM compartment. These results underscore that, akin to other types of tissue stem cells (Simons and Clevers, 2011), T cell memory stemness is maintained not only at single cell level but also through the “robustness” of the CD62L+ memory pool.
It is unfortunate that TSCM cell stemness could not be evaluated in this study, as the experimental conditions employed by Graef et al. did not support the formation of this memory subset. Pathogen-specific TSCM cells have yet to be reported in mice but have been described in human and nonhuman primates (Gattinoni et al., 2011; Lugli et al., 2013). More importantly, it is becoming clear that these cells are fundamental for the maintenance of immune homeostasis as perturbation of the TSCM cell compartment has been linked to the pathogenesis of simian immunodeficiency virus infection in rhesus macaques (Cartwright et al., 2014). The authors contended that since naïve T cells and TCM were virtually indistinguishable in their capacity for immune reconstitution, intermediates are unlikely to do better. It should be stressed that naïve and TCM cells are not functionally identical. Like two paths leading to the same destination, naïve and TCM cells might exhibit comparable efficiencies for immune reconstitution as a result of different functional properties. For example, a single naïve T cell might be outcompeted for antigen access by increased numbers of endogenous naïve T cells specific for the same epitope, impairing its reconstitution efficiency. TCM cells, instead, might have a defective reconstitution potential compared to naïve T cells that could be compensated by CXCR3-mediated migratory advantages facilitating their encounter with pathogens (Sung et al., 2012). Thus, it is possible to envision that an intermediate subset possessing the “best of both worlds” could outperform naïve and TCM cell ability for reconstitution. In fact, while there were not significant differences in the engraftment and expansion of human naïve and TCM cells adoptively transferred into highly immunodeficient NOD.Cg-PrkdcscidIl2rgtm1Wjl/SzJ mice, TSCM cells outpaced both subsets generating 10- to 100-fold more progenies in both lymphoid and peripheral tissues (Gattinoni et al., 2011). Nevertheless, the data presented here by Graef et al. clearly indicated that TCM cells are sufficient to propagate T cell memory and reconstitute epitope-specific immunocompetence of the host.
Graef et al. also provide new experimental evidence that helps resolve the ongoing debate regarding the ontogeny of memory cells. The findings that even 100 TEM cells could not be propagated across serial adoptive transfer strongly argue against a linear model of differentiation where TCM cells develop from TEFF cells through maturational transition into TEM cells (Wherry et al., 2003). Instead, these results are consistent with a developmental model of differentiation where cells progressively lose their proliferative and developmental potential as they differentiate from CD62L+ T cells to TEM and TEFF cells.
The findings reported here by Graef et al. have important therapeutic implications and add to the mounting evidence that less-differentiated CD62L+ T cells are the ideal cell population to use in cellular therapies targeting intracellular pathogens and cancer (Gattinoni et al., 2012). Long-term persistence of adoptively transferred T cells have been shown to correlate with objective tumor responses across multiple clinical trials employing either naturally occurring or gene-engineered tumor-reactive T cells (Gattinoni et al., 2012). However, cell products currently employed in adoptive immunotherapy studies predominantly comprise TEFF and TEM cells, which have a limited life span. Adoptive transfer of long-lived, multipotent CD62L+ memory T cells might significantly improve persistence and potentiate the therapeutic efficacy of adoptive immunotherapies. New clinical trials employing TCM or CD62L+-derived T cells have been initiated and hopefully will translate into increased tumor response rates. Finally, the experimental demonstration that tiny numbers of CD62L+ memory cells can fully reconstitute immunocompetence emphasizes the idea that large numbers of cells are not necessary for therapeutic success if memory stem cell populations are employed. The use of small numbers of CD62L+ memory cells might reduce the cost and complexity of the treatment and, ultimately, allow the widespread application of adoptive immunotherapies.
Footnotes
Conflict-of-interest disclosure: The author declares no competing financial interests.
REFERENCES
- Cartwright EK, McGary CS, Cervasi B, Micci L, Lawson B, Elliott ST, Collman RG, Bosinger SE, Paiardini M, Vanderford TH, et al. (2014). Divergent CD4+ T Memory Stem Cell Dynamics in Pathogenic and Nonpathogenic Simian Immunodeficiency Virus Infections. J Immunol [DOI] [PMC free article] [PubMed]
- Fearon DT, Manders P, and Wagner SD (2001). Arrested differentiation, the self-renewing memory lymphocyte, and vaccination. Science 293, 248–250. [DOI] [PubMed] [Google Scholar]
- Gattinoni L, Klebanoff CA, and Restifo NP (2012). Paths to stemness: building the ultimate antitumour T cell. Nature reviews. Cancer 12, 671–684. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Gattinoni L, Lugli E, Ji Y, Pos Z, Paulos CM, Quigley MF, Almeida JR, Gostick E, Yu Z, Carpenito C, et al. (2011). A human memory T cell subset with stem cell-like properties. Nature medicine 17, 1290–1297. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Lugli E, Dominguez MH, Gattinoni L, Chattopadhyay PK, Bolton DL, Song K, Klatt NR, Brenchley JM, Vaccari M, Gostick E, et al. (2013). Superior T memory stem cell persistence supports long-lived T cell memory. The Journal of clinical investigation [DOI] [PMC free article] [PubMed]
- Osawa M, Hanada K, Hamada H, and Nakauchi H (1996). Long-term lymphohematopoietic reconstitution by a single CD34-low/negative hematopoietic stem cell. Science 273, 242–245. [DOI] [PubMed] [Google Scholar]
- Simons BD, and Clevers H (2011). Strategies for homeostatic stem cell self-renewal in adult tissues. Cell 145, 851–862. [DOI] [PubMed] [Google Scholar]
- Sung JH, Zhang H, Moseman EA, Alvarez D, Iannacone M, Henrickson SE, de la Torre JC, Groom JR, Luster AD, and von Andrian UH (2012). Chemokine guidance of central memory T cells is critical for antiviral recall responses in lymph nodes. Cell 150, 1249–1263. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Wherry EJ, Teichgraber V, Becker TC, Masopust D, Kaech SM, Antia R, von Andrian UH, and Ahmed R (2003). Lineage relationship and protective immunity of memory CD8 T cell subsets. Nature immunology 4, 225–234. [DOI] [PubMed] [Google Scholar]