

Abstract
Polyphenol-rich plant extracts might alleviate the negative impact of oxidative stress and inflammation, but careful phytochemical standardisation and evaluation of various mechanisms are required to fully understand their effects. In this context, flower extracts of Sorbus aucuparia L.—a traditional medicinal plant—were investigated in the present work. The LC-MS/MS profiling of the extracts, obtained by fractionated extraction, led to the identification of 66 constituents, mostly flavonols (quercetin and sexangularetin glycosides with dominating isoquercitrin), pseudodepsides of quinic and shikimic acids (prevailing isomers of chlorogenic acid and cynarin), and flavanols (catechins and proanthocyanidins). Minor extract components of possible chemotaxonomic value were flavalignans (cinchonain I isomers) and phenylamides (spermidine derivatives). As assessed by HPLC-PDA and UV-spectrophotometric studies, the extracts were polyphenol-abundant, with the contents up to 597.6 mg/g dry weight (dw), 333.9 mg/g dw, 382.0 mg/g dw, and 169.0 mg/g dw of total phenolics, flavonoids, proanthocyanidins, and caffeoylquinic acids, respectively. Their biological in vitro effects were phenolic-dependent and the strongest for diethyl ether, ethyl acetate, and n-butanol fractions of the methanol-water (7 : 3, v/v) extract. The extracts showed significant, concentration-dependent ability to scavenge in vivo-relevant radical/oxidant agents (O2∙−, OH∙, H2O2, ONOO–, NO∙, and HClO) with the strongest effects towards OH∙, ONOO–, HClO, and O2∙− (compared to ascorbic acid). Moreover, the extracts efficiently inhibited lipoxygenase and hyaluronidase (compared to indomethacin) but were inactive towards xanthine oxidase. At in vivo-relevant levels (1-5 μg/mL), they also effectively protected human plasma components (proteins and lipids) against ONOO–-induced oxidative damage (reduced the levels of 3-nitrotyrosine, lipid hydroperoxides, and thiobarbituric acid-reactive substances) and normalised/enhanced the total nonenzymatic antioxidant capacity of plasma. In cytotoxicity tests, the extracts did not affect the viability of human PBMCs and might be regarded as safe. The results support the application of the extracts in the treatment of oxidative stress-related pathologies cross-linked with inflammatory changes.
1. Introduction
The regular intake of plant products rich in polyphenols is associated with the reduced risk of NCDs, including cardiovascular disease, atherosclerosis, age-related neurodegenerative disorders, diabetes, and some types of cancer [1]. In the treatment of NCDs that are often multicausal, combined therapies are usually most efficient, as they offer the advantage of additive and synergistic effects [2]. Complex chemical composition and relevant synergy of polyphenolic extracts are thus probably behind their ability to modulate multiple NCD-related pathologies. A pivotal role in the initiation and progression of NCDs is ascribed to the interdependent processes of oxidative stress and inflammation [3, 4]. As free radical scavengers, metal chelators, inhibitors of proinflammatory enzymes, and modifiers of cell signalling pathways, polyphenols may protect cells against oxidative stress-related damage and support normal cellular metabolism and functions [1]. Apart from their dietary role as constituents of fruits and vegetables, polyphenols attract increased attention as components of standardised plant extracts applicable in the adjuvant therapy of NCDs. Indeed, some selected extracts, for instance, those of hawthorn fruit/flower, grapevine leaf/seeds, olive leaf, and chokeberry fruit, have already been introduced worldwide as antioxidant nutraceuticals, and extensive research is being conducted to broaden the spectrum of the relevant plant-based products [5].
Sorbus aucuparia L. (rowan, European mountain ash) is a wild rosaceous tree occurring and cultivated across Europe and Asia [6]. Flowers, leaves, and edible fruits (rowanberries) of S. aucuparia are traditionally used for diuretic, antidiabetic, anti-inflammatory, antiatherogenic, vasoprotective, vasorelaxant, and antidiarrheal properties [7, 8]. These activities are commonly linked with polyphenolic components, especially flavonoids (quercetin, kaempferol, and sexangularetin glycosides), anthocyanins (cyanidin glycosides), tannin-type proanthocyanidins, and caffeoylquinic acids (CHA isomers), forming unique and diversified profiles in particular organs and/or plant parts, among which the flowers are the least characterised [9–11]. The accumulating research indicates all rowan tissues as strong antioxidants [9, 11–13] and the flowers as exhibiting the highest total phenolic content (TPC) and superior activity parameters [13]. Our previous screening study revealed that, in terms of TPC values and antioxidant capacity, S. aucuparia flowers are in the top five of the twenty-four most ethnobotanically relevant raw materials in the large genus Sorbus [14]. Moreover, the TPC levels of the dry extracts of rowan flowers and especially their refined fractions of ethyl acetate and n-butanol were comparable with those observed for plant extracts effective in the prevention of oxidative stress-related ailments, such as grape seed, green tea, and green mate [10]. All these activity results, although promising, were obtained in simple chemical tests measuring only basic reducing capacity of the extracts towards transition metal ions and stable synthetic radicals (DPPH, ABTS) of relatively high molecular masses. However, in biological systems, oxidative stress is generated by low-molecular, short-lived ROS such as superoxide (O2∙–), hydrogen peroxide (H2O2), hydroxyl radical (HO∙), nitric oxide (NO∙), peroxynitrite (ONOO–), and hypochlorous acid (HClO) [3]. Moreover, the activity of polyphenols in cells, tissues, and body fluids depends on their affinity to proteins, which may significantly influence the bioavailability, antioxidant effectiveness, and interactions of polyphenols (usually inhibition) with enzymatic systems [15]. As some of the proinflammatory enzymes produce ROS or are secreted in response to ROS-dependent stimuli, their inhibition forms the bridge between antioxidant and anti-inflammatory activities of polyphenols [4]. On the other hand, some polyphenols may be cytotoxic towards mammalian cells, mainly by exhibiting prooxidant effects [16].
Therefore, the objective of this study was to verify the antioxidant activity of the flower dry extract of S. aucuparia in different in vitro models including the chemically based tests towards six radical and nonradical oxidants of physiological significance and the biological model of human plasma exposed to oxidative/nitrative stress generated by ONOO–. Moreover, the potential inhibitory activity towards three proinflammatory and prooxidant enzymes (LOX, HYAL, and XO) and cellular safety of the extracts (cytotoxicity against human peripheral blood mononuclear cells) were also evaluated. All activity studies were performed for extracts standardised by comprehensive phytochemical profiling using complementary UHPLC-PDA-ESI-MS3, HPLC-PDA, and UV-spectrophotometric methods.
2. Materials and Methods
2.1. Plant Material and Extract Preparation
Flowers of Sorbus aucuparia L. were collected and authenticated in May 2015 in the Arboretum (51°49′N, 19°53′E), Forestry Experimental Station of Warsaw University of Life Science (SGGW) in Rogow (Poland). The raw material was dried under normal conditions, powdered with an electric grinder, and subjected to fractionated extraction as previously described [10] to obtain the basic extract MED and its DEF, EAF, BF, and WR fractions. The organic solvent extracts were evaporated in vacuo, and the water-containing fractions were lyophilized using an Alpha 1–2/LD Plus freeze dryer (Christ, Osterode am Harz, Germany) and stored at 4°C. In further analyses, freshly prepared solutions of the extracts in methanol-water (7 : 3, v/v) were used. All quantitative results were calculated per extract dw.
2.2. Phytochemical Standardisation
The qualitative profiling (UHPLC-PDA-ESI-MS3 analysis) of the extracts was performed on a UHPLC-3000 RS system (Dionex, Dreieich, Germany) equipped with a dual low-pressure gradient pump, an autosampler, a column compartment, a diode array detector, and an AmaZon SL ion trap mass spectrometer with an ESI interface (Bruker Daltonik, Bremen, Germany). Separations were carried out on a Kinetex XB-C18 column (1.7 μm, 150 mm × 2.1 mm i.d.; Phenomenex, Torrance, CA, USA) at 25°C. The mobile phase consisted of solvent A (water/acetonitrile/formic acid, 95 : 5 : 0.1, v/v/v), and solvent B (acetonitrile/formic acid, 100 : 0.1, v/v) with the elution profile as follows: 0–45 min, 6%–26% B (v/v); 45–55 min, 26%–95% B; 55–63 min, 95% B; 63–70 min, 95%–6% B; and 70–80 min, 6% B (equilibration). All solvents (Avantor Performance Materials, Gliwice, Poland) were of HPLC-grade purity. The flow rate was 0.3 mL/min. Before injection, sample solutions of the extracts (3.0 mg/mL) were filtered through a PTFE syringe filter (13 mm, 0.2 μm, Whatman, Pittsburgh, PA, USA). UV-Vis spectra were recorded over the range of 200–600 nm. The LC eluate was introduced directly into the ESI interface without splitting and analysed in a negative ion mode. The ESI parameters are as follows: the nebulizer pressure was 40 psi; dry gas flow 9 L/min; dry temperature 300°C; and capillary voltage 4.5 kV. MS2 and MS3 fragmentations were obtained in Auto MS/MS mode for the most abundant ions at the time. Analysis was carried out using scan from m/z 200 to 2200.
The total phenolic contents (TPC) and total proanthocyanidin contents (TPA) were quantified by the Folin-Ciocalteu and n-butanol-HCl methods, respectively, as described previously [10]. Results were expressed as equivalents of gallic acid (GAE) and cyanidin chloride (CYE), respectively.
The quantitative HPLC-PDA-fingerprint assays were performed according to Olszewska et al. [10] using the same equipment and procedure. The phenolic analytes were quantified as equivalents of HPLC-pure external standards (Sigma-Aldrich, Seelze, Germany/St. Louis, MO, USA): flavan-3-ols, flavalignans, and proanthocyanidins as ECA; hydroxybenzoic acids as protocatechuic or p-hydroxybenzoic acids; monocaffeoylquinic acid isomers as CHA; dicaffeoylquinic acid isomers as cynarin; hydroxycinnamic acid derivatives including spermidine isomers as caffeic or p-coumaric acids; flavonoid monoglycosides as IQ; flavonoid diglycosides as RT; and flavonoid aglycones as QU, depending on the PDA spectra.
2.3. Antioxidant Activity Assays against Multiple Oxidants
The antioxidant activity was evaluated in vitro by different spectrophotometric and fluorimetric methods following reported literature and using microplate readers SPECTROstar Nano (BMG Labtech, Ortenberg, Germany) and Synergy HTX (BioTek, Winooski, VT, USA). The scavenging efficacy towards O2∙− was evaluated in a xanthine/xanthine oxidase system with nitrotetrazolium blue chloride (NBT) used for detection according to Michel et al. [17]. The ability to scavenge HO∙ was assayed by the method of Fu et al. [18] with the level of HO∙ (generated in Fenton reaction) monitored in the presence of salicylic acid. The NO∙-scavenging activity was evaluated according to Czerwińska et al. [19] using diaminofluorescein-2 as NO∙ probe. The reducing activity towards H2O2 was determined following the method of Banothu et al. [20] through direct measurement of the oxidant's absorbance. The ability to scavenge ONOO− was determined by the measurement of the inhibition of Evans blue dye oxidation according to Krzyzanowska-Kowalczyk et al. [21]. The HClO-scavenging effect was assayed by the method of Czerwińska et al. [19] with 5-thio-2-nitrobenzoic acid used for detection. The results of triplicate determinations were expressed as SC50 values (defined as the concentration sufficient to obtain 50% of a maximum scavenging capacity), calculated from concentration-inhibition curves, and recalculated into equivalents of AA and TX. All reagents and standards for the assays were obtained from Sigma-Aldrich (Seelze, Germany/St. Louis, MO, USA).
2.4. Antioxidant Activity in Human Plasma Model
2.4.1. Isolation of Blood Plasma and Sample Preparation
Blood from healthy, nonsmoking volunteers declaring balanced diet free of antioxidant supplements was obtained from the Regional Centre of Blood Donation and Blood Treatment in Lodz (Poland), collected on CPD (citrate/phosphate/dextrose) solution in the Fresenius-Kabi Compoflex bags, and plasma was isolated by differential centrifugation of the blood [22]. The study was conducted in accordance with the Declaration of Helsinki and all experiments were approved by the committee on the Ethics of Research at the University of Lodz (8/KBBN-UŁ/II/2015). For the FRAP assay and measurements of 3-NT, plasma samples were diluted with a (Ca2+)-free phosphate-buffered saline (PBS; 1 : 4, v/v) purchased from Biomed (Lublin, Poland), whereas for LOOH and TBARS assays, plasma was diluted with (Ca2+)-free PBS in a volume ratio 1 : 1. All samples were preincubated for 5 min at 37°C with the examined extracts added to the final concentration range of 1-50 μg/mL and then exposed to 150 μM (the FRAP assay) or 100 μM (the remaining experiments on blood plasma) of ONOO–. Control samples were prepared with plasma untreated with the extracts and/or ONOO–. In the experiments with blood plasma and the extracts only (without adding ONOO–), no significant differences (p > 0.05) were found between the levels of the tested biomarkers in the plasma samples incubated with the extracts and control (untreated) serum. Protein concentration in blood plasma was estimated using the bicinchoninic acid (BCA) assay with the Pierce BCA Protein Assay Kit (Thermo Scientific, Waltham, MA, USA) according to the manufacturer protocol. ONOO– was synthesised as reported earlier [21]. All tests in blood plasma were conducted using 96-well plates and a microplate reader, SPECTROstar Nano (BMG Labtech, Ortenberg, Germany).
2.4.2. Determination of 3-NT in Plasma Proteins
Detection of 3-NT-containing proteins by the competitive ELISA (C-ELISA) method in plasma samples (control or antioxidants and ONOO–-treated plasma) was performed according to Kolodziejczyk-Czepas et al. [22] using immunoreagents purchased from Abcam (Cambridge, UK). The concentrations of nitrated proteins were estimated from the standard curve of nitrated fibrinogen (Fg) and expressed as the 3NT-Fg equivalents (in nmol/mg of plasma protein).
2.4.3. Determination of LOOH in Plasma Lipids
Concentration of hydroperoxides in plasma samples (control or antioxidants and ONOO–-treated plasma) was determined by the ferric-xylenol orange (FOX-1) protocol according to Kolodziejczyk-Czepas et al. [22]. The FOX-1 reagent contained 125 μM xylenol orange and 100 mM sorbitol in 25 mM sulphuric acid and was freshly prepared each time before use by the addition of ammonium ferrous sulphate to the final concentration of 250 μM. All reagents were purchased from Sigma-Aldrich (Seelze, Germany/St. Louis, MO, USA). To perform the assay, blood plasma samples were mixed with the reagent in a volume ratio 1 : 9 and incubated for 30 min in the dark (25°C). Absorbance of the samples was measured at 560 nm against blank (water instead of the plasma). The amount of lipid hydroperoxides was calculated from the standard curve of hydrogen peroxide and expressed in nmol/mg of plasma proteins.
2.4.4. TBARS Test
Determination of TBARS in plasma samples (control or antioxidants and ONOO–-treated plasma) was performed according to Kolodziejczyk-Czepas et al. [23]. 2-Thiobarbituric acid and other reagents were purchased from Sigma-Aldrich (Seelze, Germany/St. Louis, MO, USA). Results were expressed in μmol TBARS/mL of plasma.
2.4.5. FRAP Assay
The influence of the extracts on the NEAC of plasma was determined according to Kolodziejczyk-Czepas et al. [24] with some modifications. The plasma samples prepared as described above were added to the reagent mixture in a volume ratio of 1 : 10 : 1 : 1 for plasma, acetate buffer (300 mM, pH 3.6), TPTZ (2,4,6-tris-(2-pyridyl)-s-triazine; 10 mM, in 0.04 M hydrochloric acid), and ferric chloride (20 mM), respectively. After incubation for 15 min at 37°C, the measured FRAP of plasma samples (control or antioxidants and ONOO–-treated plasma) was expressed in millimolar equivalents of ferrous (Fe2+) ions calculated from the calibration curve of ferrous sulphate. All reagents were purchased from Sigma-Aldrich (Seelze, Germany/St. Louis, MO, USA).
2.5. Inhibition of Proinflammatory Enzymes
The ability of the extracts to inhibit LOX and HYAL was examined according to Matczak et al. [25]. The inhibitory activity towards XO was analysed according to Michel et al. [17]. All reagents and standards for the study including IND, bovine testis HYAL, LOX from soybean, and XO were purchased from Sigma-Aldrich (Seelze, Germany/St. Louis, MO, USA). Results were expressed as IC50 values calculated from concentration-inhibition curves.
2.6. Cellular Safety Studies
Cytotoxicity of the examined extracts was determined in an experimental model of PBMCs. Cells were isolated from human blood using the Histopaque®-1077 medium purchased from Sigma-Aldrich (Seelze, Germany/St. Louis, MO, USA) as a sterile solution of polysucrose (57 g/L) and sodium diatrizoate (90 g/L) with a density of 1.077 g/mL. From each of the six donors, two independent PBMC isolations and incubations with the extracts were performed. Blood was carefully layered (in a volume ratio of 1 : 1) onto the medium and centrifuged for 30 min (400×g, at room temperature). Then, the pellet was washed two times with 0.02 M PBS buffer. The obtained fraction of PBMCs was suspended in PBS. Cell suspensions (1 × 106 PBMCs/mL) were incubated for 1 and 2 h with plant extracts added to the final concentration of 5 and 50 μg/mL (at 37°C). Cell viability (%) was determined during a spectrofluorimetric analysis, involving the use of propidium iodide as a fluorescent dye. Measurements were conducted using a microchip-type automatic cell counter Adam-MC DigitalBio (NanoEnTek Inc., Seoul, Korea) according to the manufacturer's protocol. Additionally, the PBMCs, isolated as described above, were suspended in the RPMI-1640 medium (3 × 106 PBMCs/mL) and incubated with the extracts (5-50 μg/mL) for 24 h (in 96-well microplates, at 37°C, in a humidified atmosphere). Measurements were carried out analogously as described above.
2.7. Data Analysis
The results were reported as means ± SD (standard deviation) or ± SE (standard error) for the indicated number of experiments. The significance of differences between samples and controls was determined with one-way ANOVA (for chemical tests) or one-way ANOVA for repeated measures (for human plasma model), followed by post hoc Tukey's test for multiple comparisons. The correlations were evaluated using an F-test. All calculations were performed using the Statistica12Pl software for Windows (StatSoft Inc., Krakow, Poland) with p values less than 0.05 regarded as significant.
3. Results
3.1. Phytochemical Profiling
The LC-MS analysis revealed significant, extraction solvent-dependent differences in the chemical composition of the extracts (Figure 1, Table 1). The assay enabled the detection of 72 phenolic components (UHPLC peaks 1–72), the structures of 66 of which were fully or tentatively characterised based on the comparison of their chromatographic behaviour and ESI-MS3 fragmentation patterns (in a negative ionisation mode) with the literature data [11, 26–33] or reference standards, both commercial and isolated previously in our laboratory from various Sorbus species [34, 35]. The analytes represented a wide range of polyphenolic classes, including flavonols, pseudodepsides of quinic acid and shikimic acid, flavan-3-ol derivatives (catechins and proanthocyanidins), simple phenolic acids, phenylamides, flavanones, and flavalignans. The greatest chemical diversity was observed for MED, while its fractions (DEF, EAF, BF, and WR) obtained after sequential liquid-liquid partitioning were enriched in selected constituents, depending on the fractionation solvent. Among the analytes, only CHA (7) was found in all extracts. The fractionation has influenced also the quantitative profiles of the extracts (Table 2). The TPC levels determined by the Folin-Ciocalteu assay varied in a wide range of 111.7-597.6 mg GAE/g dw with the highest value observed for EAF. The primary constituents of the basic extract MED were mono- and dicaffeoylquinic acids with the dominating CHA and flavonol mono- and diglycosides with the prevailing IQ (35). In accordance with their polarity, flavonol monoglycosides were concentrated mainly in EAF and DEF, flavonol diglycosides in BF, and monocaffeoylquinic acids (CHA isomers) in BF and WR, while dicaffeoylquinic acids (cynarin isomers) and simple phenolic acids in DEF. The total contents of phenolics (TPH), calculated as a sum of individual analytes quantified by RP-HPLC-PDA, were in the range of 82.7-554.0 mg/g dw, with the highest concentration observed still for EAF. The TPH values were, however, significantly lower than the TPC levels. This difference might be related to the presence of proanthocyanidins—the compounds occurring in plants at various degrees of polymerisation and measureable by RP-HPLC only in the form of oligomers built from less than four flavan-3-ol monomers. Within this group, only low levels of flavan-3-ols (peaks: 6, 12, 32, 38) and B-type proanthocyanidin dimers (peaks: 2, 4, 10, and 37) were observed in the extracts. In consequence, the total contents of proanthocyanidins (TPA) determined by the n-butanol/HCl assay (11.4-380.2 mg CYE/g dw) might be considered as a measure of the content of higher flavan-3-ol oligomers.
Figure 1.

Representative UHPLC chromatograms at 280 nm of (a) DEF and (b) EAF extracts of S. aucuparia flowers. Peak numbers refer to those implemented in Table 1.
Table 1.
UHPLC-PDA-ESI-MS3 identification data of polyphenols detected in the dry extracts from S. aucuparia flowers.
| No. | Analyte | R t (min) | UV λmax (nm) | [M–H]– (m/z) | MS2/MS3 fragmentation | Extract | References |
|---|---|---|---|---|---|---|---|
| 1 | Protocatechuic acida | 3.0 | 259, 294 | 153 | DEF, EAF | ||
| 2 | (Epi)catechin-B-(epi)catechinb,c | 3.4 | 280 | 577 | 451 (46), 425 (100), 407 (33) | DEF, EAF | [29] |
| 3 | 3-O-Caffeoylquinic acid (neochlorogenic acid)a | 3.5 | 216, 324 | 353 | 191 (100), 179 (30), 135 (3) | ME, BF, WR | [9, 11, 26] |
| 4 | (Epi)catechin-B-(epi)catechinb,c | 4.2 | 279 | 577 | 451 (60), 425 (100), 407 (28) | ME, DEF, EAF | [29] |
| 5 | p-Hydroxybenzoic acida | 4.6 | 255 | 137 | DEF | ||
| 6 | (+)-Catechina,b | 4.9 | 278 | 289 | 245 (100), 205 (21) | ME, DEF, EAF | |
| 7 | 5-O-Caffeoylquinic acid (chlorogenic acid, CHA)a | 5.4 | 216, 325 | 353 | 191 (100), 179 (3) | ME, DEF, EAF, BF, WR | [9, 11, 26] |
| 8 | 4-O-Caffeoylquinic acid (cryptochlorogenic acid)a | 6.3 | 216, 325 | 353 | 191 (50), 179 (49), 173 (100) | ME, EAF, BF, WR | [26] |
| 9 | Caffeic acida | 6.4 | 215, 323 | 179 | DEF | ||
| 10 | Procyanidin B2 (PB2)b | 6.8 | 280 | 577 | 451 (44), 425 (100), 407 (23) | DEF, EAF, BF | [29] |
| 11 | Unidentified | 7.8 | 261 | 360 | 313 (100), 151 (49) | EAF, BF | |
| 12 | (–)-Epicatechin (ECA)a,b,c | 8.1 | 279 | 289 | 245 (100), 205 (32) | ME, DEF, EAF | |
| 13 | Eriodictyol O-hexosideb,c | 8.2 | 279, 330sh | 449 | 287 (100), 269 (25), 259 (31) | BF | [31] |
| 14 | Unidentified | 8.4 | 249, 297 | 641 | 461 (17), 317 (100) | ME, BF | |
| 15 | 5-O-p-Coumaroylquinic acidb,c | 9.0 | 207, 311 | 337 | 191 (100), 163 (13) | EAF, BF | [26] |
| 16 | 5-O-Caffeoylshikimic acidb,c | 9.2 | 211, 325 | 335 | 291 (25), 179 (100), 135 (11) | ME, DEF, EAF, BF | [30] |
| 17 | Vanillic acida,b,c | 9.6 | 260, 293 | 167 | DEF | ||
| 18 | Sexangularetin di-O-hexosideb,c | 9.7 | 272, 326 | 639 | 519 (25), 477 (100), 315 (21) | ME, BF | [31] |
| 19 | p-Coumaric acida | 10.5 | 222, 309 | 163 | DEF | ||
| 20 | Quercetin 3-O-β-sophorosidea | 11.6 | 255, 353 | 625 | 463 (23), 445 (39), 301 (100) | ME, EAF, BF | |
| 21 | Coumaric acid isomerb,c | 11.9 | 293 | 163 | DEF | ||
| 22 | Quercetin O-dihexosideb,c | 12.3 | 255, 352 | 625 | 463 (35), 445 (60), 301 (100) | ME, EAF, BF | [31] |
| 23 | 4-O-Feruloylquinic acidb,c | 12.7 | 214, 323 | 367 | 191 (37), 179 (100), 135 (35) | DEF, EAF | [26] |
| 24 | Sexangularetin O-dihexosideb,c | 14.2 | 270, 337 | 639 | 477 (16), 459 (86), 315 (100) | BF | [31] |
| 25 | Quercetin O-pentosylhexosideb,c | 14.3 | 270, 340 | 595 | 463 (12), 445 (25), 301 (100) | EAF, BF | [31] |
| 26 | Kaempferol O-dihexosideb,c | 15.0 | 267, 340 | 609 | 447 (6), 429 (27), 285 (100) | BF | [31] |
| 27 | Kaempferol O-dihexosideb,c | 15.5 | 266, 343 | 609 | 447 (13), 429 (86), 285 (100) | BF | [31] |
| 28 | Quercetin O-hexosylpentosideb,c | 16.2 | 269, 342 | 595 | 433 (10), 415 (35), 301 (100) | ME, BF | [31] |
| 29 | Sexangularetin O-rhamosylhexosideb,c | 16.3 | 273, 333 | 623 | 477 (15), 459 (61), 315 (100) | ME, EAF, BF | [31] |
| 30 | Quercetin O-rhamosylhexosideb,c | 16.9 | 255, 353 | 609 | 301 (100) | ME, EAF, BF | [31] |
| 31 | Cinchonain I isomerb,c | 17.0 | 280 | 451 | 341 (100) | DEF | [32] |
| 32 | (Epi)catechin derivativeb,c | 17.3 | 279 | 483 | 451 (45), 341 (17), 289 (100) | DEF | |
| 33 | Quercetin 3-O-(6″-O-α-l-rhamnosyl)-β-d-glucoside (rutin, RT)a | 17.5 | 256, 353 | 609 | 301 (100) | ME, BF | [11] |
| 34 | Quercetin 3-O-β-d-galactoside (hyperoside)a | 17.6 | 255, 354 | 463 | 301 (100) | ME, DEF, EAF, BF | [11] |
| 35 | Quercetin 3-O-β-d-glucoside (isoquercitrin, IQ)a | 18.6 | 256, 353 | 463 | 301 (100) | ME, DEF, EAF, BF | [11] |
| 36 | Unidentified | 19.0 | 312 | 193 | DEF | ||
| 37 | (Epi)catechin-B-(epi)catechinb,c | 19.2 | 279 | 577 | 451 (60), 425 (100), 407 (28) | EAF | [29] |
| 38 | (Epi)catechin derivativeb,c | 19.4 | 256 | 483 | 451 (77), 341 (37), 289 (100) | DEF | |
| 39 | Unidentified | 19.6 | 280, 333sh | 597 | 477 (75), 387 (75), 357 (100) | BF | |
| 40 | Kaempferol O-rhamosylhexosideb,c | 19.8 | 277, 335 | 593 | 447 (11), 285 (100) | BF | [31] |
| 41 | Unidentifiedb,c | 19.9 | 282 | 519 | 309 (100) | DEF | |
| 42 | Ferulic acida,b,c | 20.2 | 217, 321 | 193 | DEF | ||
| 43 | Kaempferol O-hexosideb,c | 21.0 | 264, 344 | 447 | 285 (100) | DEF, EAF | [31] |
| 44 | Eriodictyol O-glucuronideb,c | 21.5 | 283 | 463 | 287 (100) | ME, BF | [9] |
| 45 | Sexangularetin 3-O-β-d-glucoside (sorbaroside)a | 21.8 | 270, 350 | 477 | 315 (100) | ME, DEF, EAF, BF | |
| 46 | Kaempferol 3-O-β-d-glucoside (astragalin)a,b,c | 22.9 | 265, 343 | 447 | 285 (100) | ME, DEF, EAF, BF | |
| 47 | Unidentified | 23.2 | 273, 350 | 507 | 491 (100), 345 (40), 329 (36) | EAF, BF | |
| 48 | Cinchonain I isomerb,c | 23.2 | 279 | 451 | 341 (100), 299 (23) | ME, DEF | [32] |
| 49 | Cinchonain I isomerb,c | 23.3 | 279 | 451 | 341 (100) | ME, DEF | [32] |
| 50 | Quercetin acetylhexosideb,c | 24.3 | 255, 350 | 505 | 463 (21), 301 (100) | ME, DEF, EAF, BF | [28] |
| 51 | 3,5-O-Dicaffeoylquinic acidb,c | 25.4 | 217, 326 | 515 | 353 (100), 191 (100)d, 179 (47)d | ME, DEF, EAF, BF | [27] |
| 52 | Caffeic acid derivativeb,c | 25.9 | 218, 328 | 437 | 377 (30), 275 (100), 179 (5) | ME, DEF, EAF | |
| 53 | Cinchonain I isomerb,c | 27.4 | 281 | 451 | 341 (100) | DEF | [32] |
| 54 | Sexangularetin O-acetylhexosideb,c | 27.9 | 271, 348 | 519 | 315 (100), 301 (60) | ME, BF | [28] |
| 55 | Quercetin O-acetylhexosideb,c | 28.3 | 260, 348 | 505 | 463 (50), 301 (100) | DEF | [28] |
| 56 | Sexangularetin O-acetylhexosideb,c | 28.5 | 271, 333 | 519 | 505 (32), 315 (100) | ME, DEF | [28] |
| 57 | 4,5-O-Dicaffeoylquinic acidb,c | 28.9 | 218, 325 | 515 | 353 (100), 179 (60)d, 173 (100)d | ME, DEF, EAF, BF | [27] |
| 58 | Kaempferol O-acetylhexosideb,c | 30.0 | 265, 335 | 489 | 327 (12), 285 (100) | DEF | [28] |
| 59 | Cinchonain I isomerb,c | 31.4 | 280 | 451 | 341 (100) | DEF | [32] |
| 60 | 3-O-Caffeoyl-5-O-feruloylquinic acidb,c | 31.7 | 308 | 529 | 367 (100), 191 (100)d, 179 (70)d | DEF | [27, 30] |
| 61 | 3-O-Feruloyl-5-O-caffeoylquinic acidb,c | 32.8 | 322 | 529 | 367 (100), 353 (84), 193 (100)d | DEF | [27, 30] |
| 62 | 3-O-Caffeoyl-4-O-p-coumaroilquinic acidb,c | 34.4 | 324 | 499 | 337 (100), 179 (100)d, 173 (97)d | DEF | [27, 30] |
| 63 | Caffeic acid derivativeb,c | 34.9 | 218, 328 | 437 | 377 (30), 275 (100), 179 (5) | BF | |
| 64 | Quercetina | 35.0 | 268, 364 | 301 | DEF | ||
| 65 | Tricoumaroyl spermidine isomerb,c | 39.0 | 285 | 582 | 462 (100), 342 (12) | DEF | [33] |
| 66 | Dicoumaroyl-caffeoyl spermidine isomerb,c | 39.7 | 288 | 598 | 478 (70), 462 (100), 342 (26) | DEF | [33] |
| 67 | Dicoumaroyl-caffeoyl spermidine isomerb,c | 41.1 | 313 | 598 | 478 (47), 462 (100), 342 (30) | ME, DEF, EAF | [33] |
| 68 | Tricoumaroyl spermidine isomerb,c | 41.6 | 293 | 582 | 462 (100), 342 (14) | DEF | [33] |
| 69 | Tricoumaroyl spermidine isomerb,c | 43.0 | 289 | 582 | 462 (100), 342 (9) | DEF, EAF | |
| 70 | Kaempferola | 43.6 | 272, 373 | 285 | DEF | ||
| 71 | Tricoumaroyl spermidine isomerb,c | 44.1 | 291 | 582 | 462 (100), 342 (12) | DEF | [33] |
| 72 | Tricoumaroyl spermidine isomerb,c | 45.2 | 293 | 582 | 462 (100), 342 (24) | ME, DEF, EAF | [33] |
R t: retention time. UV λmax: absorbance maxima in PDA spectra. [M–H]–: pseudomolecular ions in MS spectra recorded in a negative mode. MS2: secondary ions (the most abundant ions were subjected to MS3 fragmentation). Intensities of particular ions are given in parentheses. The nomenclature of the pseudodepsides of quinic acid and shikimic acid is given according to IUPAC [26, 27, 30]. aCompounds identified with authentic standards. bCompounds detected for the first time in S. aucuparia flowers. cCompounds detected for the first time in S. aucuparia. dMS3 ions.
Table 2.
Quantitative profile of the S. aucuparia flower dry extracts (mg/g dw).
| MED | DEF | EAF | BF | WR | |
|---|---|---|---|---|---|
| TPC (GAE) | 221.9 ± 6.7d | 533.3 ± 3.7b | 597.6 ± 4.6a | 485.0 ± 12.7c | 111.7 ± 4.5e |
| TPH | 137.5 ± 2.3d | 300.6 ± 6.8b | 559.6 ± 8.5a | 248.7 ± 6.7c | 82.7 ± 1.3e |
| TFL | 40.0 ± 1.4d | 130.7 ± 1.2b | 333.9 ± 4.4a | 115.2 ± 1.8c | 2.9 ± 0.2e |
| SQ | 4.4 ± 0.1b | n.d. | 1.1 ± 0.1c | 21.6 ± 1.2a | 0.66 ± 0.04d |
| HY | 6.2 ± 0.2d | 24.2 ± 1.1b | 79.1 ± 3.7a | 12.2 ± 0.5c | 0.11 ± 0.01e |
| IQ | 11.7 ± 0.5d | 56.2 ± 1.8b | 155.2 ± 4.4a | 21.6 ± 1.7c | 0.07 ± 0.01e |
| GS | 4.6 ± 0.2d | 18.1 ± 0.7b | 57.6 ± 2.1a | 7.5 ± 0.3c | n.d. |
| RT | 3.5 ± 0.1b | n.d. | 3.4 ± 0.1b | 18.1 ± 0.3a | 0.34 ± 0.01c |
| TCFA (TCHA+CFA) | 88.7 ± 3.3c | 91.2 ± 4.1c | 181.9 ± 5.5a | 126.3 ± 4.4b | 77.4 ± 3.1d |
| TCHA | 86.7 ± 4.8c | 57.5 ± 3.3e | 169.0 ± 3.6a | 108.3 ± 4.2b | 76.3 ± 2.9d |
| NCHA | 14.6 ± 0.5b | 1.4 ± 0.1d | 2.3 ± 0.1c | 15.1 ± 0.6a,b | 16.5 ± 0.8a |
| CHA | 49.7 ± 1.4b | 5.5 ± 0.2d | 19.3 ± 0.8c | 79.1 ± 2.8a | 47.7 ± 1.9b |
| CCHA | 8.6 ± 0.7b | 1.2 ± 0.1d | 2.9 ± 0.1c | 13.9 ± 0.8a | 10.2 ± 0.6b |
| 1-CHA | 2.2 ± 0.1b | n.d. | n.d. | 4.9 ± 0.2a | 2.0 ± 0.2b |
| CNE | 11.6 ± 0.6c | 49.3 ± 2.7b | 144.5 ± 6.9a | n.d. | n.d. |
| CFA | 1.9 ± 0.1d | 33.7 ± 1.4a | 12.9 ± 0.5c | 18.1 ± 0.4b | 1.0 ± 0.1e |
| HCA | 3.5 ± 0.2c | 8.9 ± 0.5a | 6.4 ± 0.2b | 4.3 ± 0.2c | 1.2 ± 0.1d |
| HBA | 1.1 ± 0.1c,d | 38.2 ± 1.3a | 2.2 ± 0.1b,c | 0.62 ± 0.04d | 0.71 ± 0.06d |
| TPA (CYE) | 110.9 ± 2.2b,c | 11.4 ± 0.3e | 103.0 ± 2.8c | 382.0 ± 4.3a | 52.8 ± 1.1d |
| TLPA | 2.1 ± 0.1c | 9.1 ± 0.4b | 22.5 ± 1.3a | 2.3 ± 0.1c | 0.59 ± 0.04d |
| LG | n.d. | 2.7 ± 0.1a | 0.76 ± 0.05b | n.d. | n.d. |
| SP | 2.3 ± 0.2c | 19.9 ± 1.2a | 5.6 ± 0.2b | n.d. | n.d. |
Results are presented as means ± SD (n = 3). For each parameter, different superscript letters indicate significant differences (p < 0.05). Additional abbreviations: n.d.: not detected; SQ: quercetin 3-O-sophoroside; HY: hyperoside; GS: sexangularetin 3-O-glucoside; NCHA: neochlorogenic acid (3-O-caffeoylquinic acid); CCHA: cryptochlorogenic acid (4-O-caffeoylquinic acid); 1-CHA: 1-O-caffeoylquinic acid; CNE: total content of dicaffeoylquinic acids (cynarin isomers); CFA: total content of caffeic acid derivatives other than TCHA; HCA: total content of simple hydroxycinnamic acids; HBA: total content of simple hydroxybenzoic acids; LG: total content of flavalignans; SP: total content of phenolic amides (spermidine derivatives). The highest levels for each parameter are printed in bold.
3.2. Antioxidant Activity Assays against Multiple Oxidants
All of the investigated extracts showed significant and concentration-dependent ability to scavenge the six most common in vivo-relevant ROS (Figure 2). In terms of AA (primary antioxidant in human plasma) equivalents, the strongest effects were revealed towards OH∙, ONOO–, and HClO, while with the reference to TX (a synthetic analogue of vitamin E) towards OH∙, HClO, and O2∙−. In five of the tests (except the HClO-scavenging), the extract activity decreased in the same order, i.e., EAF>DEF>BF>MED>WR and significantly correlated (p < 0.05) with the amounts of polyphenols, i.e., TPC (r > 0.86) and/or TPH (r > 0.88) values (Table 3). In the O2∙−, H2O2, NO∙, and ONOO–-scavenging assays, the significant influence of total flavonoids (TFL; r > 0.88), total low-molecular-weight proanthocyanidins (TLPA; r > 0.83), and/or total caffeic acid derivatives (TCFA; r > 0.91) was also evidenced. In the case of HClO, the activity order was slightly different, i.e., DEF>BF≥EAF>MED>WR, but the correlation with the TPC levels was still significant (r = 0.96). As no correlation was found between the antioxidant activity parameters and TPA values, the extract effects might be attributed mainly to low-molecular-weight polyphenols, detectable by RP-HPLC. Indeed, the simultaneous analysis of model extract constituents, representing the main groups of Sorbus polyphenols (QU, RT, ECA, PB2, and CHA), revealed their high scavenging efficiency towards the analysed ROS, similar in terms of the order of magnitude to the most active extracts (Figure 2).
Figure 2.

Scavenging activity of the S. aucuparia flower extracts, model polyphenols, and standard antioxidants on six different biologically relevant oxidants, expressed in equivalents of AA (mmol AAE/g dw) and TX (mmol TE/g dw). Values on particular charts labelled with the same italics (a-i) did not differ significantly at α = 0.05.
Table 3.
Correlation (r) coefficients and probability (p) values of linear relationships between phenolic contents of the extracts and their activity parameters towards multiple oxidants.
| r (p) for | TPC | TPH | TFL | TCFA | TCHA | TPA | TLPA |
|---|---|---|---|---|---|---|---|
| AAE (O2∙) | 0.8784 (0.050) ∗ | 0.9992 (0.000) ∗∗∗ | 0.9976 (0.000) ∗∗∗ | 0.9106 (0.032) ∗ | 0.4419 (0.456) | 0.0423 (0.946) | 0.9534 (0.012) ∗ |
| AAE (OH∙) | 0.9885 (0.001) ∗∗∗ | 0.8864 (0.045) ∗ | 0.8569 (0.064) | 0.7528 (0.142) | 0.4567 (0.439) | 0.1515 (0.808) | 0.7677 (0.130) |
| AAE (H2O2) | 0.9796 (0.003) ∗∗ | 0.9161 (0.029) ∗ | 0.8847 (0.046) ∗ | 0.7326 (0.159) | 0.4455 (0.452) | 0.0192 (0.976) | 0.8314 (0.081) ∗ |
| AAE (NO∙) | 0.8666 (0.057) | 0.9176 (0.028) ∗ | 0.8863 (0.045) ∗ | 0.6486 (0.236) | 0.4001 (0.504) | 0.2778 (0.651) | 0.9183 (0.028) ∗ |
| AAE (ONOO–) | 0.9773 (0.004) ∗∗ | 0.9170 (0.028) ∗ | 0.8853 (0.046) ∗ | 0.7308 (0.161) | 0.4419 (0.456) | 0.0105 (0.987) | 0.8343 (0.079) |
| AAE (HClO) | 0.9627 (0.009) ∗∗ | 0.7162 (0.174) | 0.6722 (0.214) | 0.5734 (0.312) | 0.2302 (0.709) | 0.2290 (0.711) | 0.5573 (0.329) |
Activity and concentration parameters according to Figure 2 and Tables 2 and 3. Asterisks mean statistical significance of the estimated linear relationships (∗p < 0.05, ∗∗p < 0.01, and ∗∗∗p < 0.001). All statistically significant relationships are printed in bold. The (r) and (p) values for TE towards individual oxidants were the same as given for AAE.
3.3. Protective Effects on Human Plasma Components
The addition of ONOO– (100-150 μM) to the plasma samples induced oxidative stress resulting in a considerable (p < 0.001) oxidative and nitrative damage to blood plasma components, confirmed by the measurements of specific biomarkers of protein nitration (3-NT, Figure 3(a)) and lipid peroxidation (LOOH and TBARS, Figures 3(b) and 3(c), respectively), as well as in a slight but statistically significant (p < 0.05) impairment of the NEAC of the plasma measured by the FRAP assay (Figure 3(d)). In comparison to the control plasma, in the ONOO–-treated samples, the NEAC was diminished by about 15%, and the oxidative damage was evidenced by an approximately 3.5-fold, 11-fold, and 2-fold increase in the levels of 3-NT, LOOH, and TBARS, respectively. In plasma samples incubated with ONOO– in the presence of the analysed extracts (at 1-50 μg/mL), the rate of oxidative and nitrative damage was significantly reduced (Figures 3(a)–3(c); p < 0.05). The antinitrative activity of the extracts was solvent-dependent, and the strongest effects were observed for MED, DEF, and EAF, with the highest inhibitory percentage (49%) found for EAF at 50 μg/mL (Figure 3(a)). However, even at the lowest concentration (1 μg/mL), these three extracts were able to diminish effectively (by about 24-33%) the ONOO–-induced protein nitration. Moreover, the antinitrative effects of the extracts were dose-dependent (except those of MED) and phenolic-dependent, which was evidenced by significant (p < 0.05) correlations between percentage inhibition of tyrosine nitration and phenolic contents (Table 4), especially the TCFA levels (r = 0.8220). All of the tested extracts protected also the plasma lipids against ONOO–-caused peroxidation (Figures 3(b) and 3(c); p < 0.05), regardless of the concentration levels. The strongest inhibitory effects—in the range of 21-35% at 1 μg/mL—were revealed for the formation of LOOH that reflected the first stage of the peroxidation process. The influence of the extracts on the generation of TBARS, which measured the final stadium of the process, was slightly lower (11-32% of inhibition at 1 μg/mL). The dose dependence was evident only for some of the extracts (e.g., for MED and DEF in the LOOH test), and the impact of phenolics was statistically proved (p > 0.05) only for the LOOH levels. In contrast, the ability of the extracts to normalise and/or enhance the NEAC status of plasma (Figure 3(d)) was strongly dose-dependent, and thus strong and statistically significant (p < 0.05) relationships were found between the percentage increase in the FRAP values of the oxidised plasma and the phenolic contents (Table 4), especially the TPC levels (r = 0.9864).
Figure 3.

Effects of S. aucuparia flower extracts on human plasma exposed to oxidative stress: (a) effects on the nitration of plasma proteins and formation of 3-NT; (b, c) effects on the peroxidation of plasma lipids and formation of (b) LOOH and (c) TBARS; (d) effects on NEAC of plasma (measured by FRAP). Results presented as means ± SE (n = 12). Statistical differences: ###p < 0.001 for control plasma versus ONOO–-treated plasma (without the extracts); ∗p < 0.05, ∗∗p < 0.01, and ∗∗∗p < 0.001 for ONOO–-treated plasma in the presence of the extracts (1, 5, or 50 μg/mL) or standards (5 μg/mL) versus ONOO–-treated plasma in the absence of the extracts.
Table 4.
Correlation (r) coefficients and probability (p) values of linear relationships between phenolic contents of the extracts and their antioxidant activity parameters in human plasma (n = 15).
| r (p) for | TPC | TPH | TFL | TCFA | TCHA | TPA | TLPA |
|---|---|---|---|---|---|---|---|
| 3-NT | 0.7833 (0.001) ∗∗∗ | 0.7729 (0.001) ∗∗∗ | 0.7153 (0.003) ∗∗ | 0.8220 (0.000) ∗∗∗ | 0.8068 (0.000) ∗∗∗ | 0.5766 (0.024) ∗ | 0.6393 (0.010) ∗∗ |
| LOOH | 0.5439 (0.036) ∗ | 0.4574 (0.086) | 0.3429 (0.211) | 0.5697 (0.027) ∗ | 0.5477 (0.035) ∗ | 0.3945 (0.146) | 0.2948 (0.286) |
| TBARS | 0.2978 (0.281) | 0.3596 (0.188) | 0.3072 (0.265) | 0.4111 (0.128) | 0.4323 (0.108) | 0.0375 (0.895) | 0.3525 (0.198) |
| FRAP | 0.9864 (0.000) ∗∗∗ | 0.9367 (0.007) ∗∗∗ | 0.8572 (0.000) ∗∗∗ | 0.9295 (0.000) ∗∗∗ | 0.8717 (0.000) ∗∗∗ | 0.5394 (0.038) ∗ | 0.7963 (0.000) ∗∗∗ |
The observed effects of the extracts were in general comparable to (p > 0.05) or higher (p < 0.05) than those of positive phenolic standards—RT, CHA, and TX (Figures 3(a)–3(d)) applied at the same concentration. Even the activity of QU, one of the most powerful natural antioxidants, did not differ significantly (p > 0.05) in the majority of the tests from that observed for the most effective extracts (Figures 3(a)–3(c)). Only in the FRAP assay, two of the standards—QU and CHA—enhanced the NEAC of plasma more strongly than the extracts at the corresponding concentration (Figure 3(d); p < 0.05).
3.4. Inhibitory Effects on Proinflammatory Enzymes
The extracts inhibited the activity of LOX and HYAL in a concentration-dependent manner, but with different responses towards particular enzymes (Table 5). Considering the IC50 values expressed in μg/U, most extracts and all standards were stronger inhibitors of LOX than of HYAL. The exceptions were BF and WR, showing similar inhibitory potential towards both enzymes. Different orders of potency were also observed in both tests, with BF being the most active towards HYAL and three extracts (DEF, EAF, and BF, not differing significantly in activity) the most effective towards LOX. Moreover, BF was a stronger HYAL inhibitor than all standards including IND, a potent nonsteroidal anti-inflammatory drug. In the LOX test, the activity of the most effective extracts was intermediate between that of IND and that of most of the phenolic standards except QU. The responses in this test were strongly TPC-dependent (r = –0.9733, p < 0.01) with some effects demonstrated for the main groups of phenolics, including TFL (r = −0.7379, p = 0.16), TCFA (r = –0.6844, p = 0.20), and TPH (r = –0.7694, p = 0.13). On the other hand, the activity of the extracts in the HYAL test was only slightly related to the TPA (r = –0.7927, p = 0.11) and not connected with the TPC levels (r = –0.0565, p = 0.93). Nevertheless, similar IC50 values obtained for the extracts and most of the model polyphenols (p < 0.05) might indicate strong synergic effects between the extract components. In contrast to their inhibitory potential towards LOX and HYAL, the extracts did not influence XO, the activity of which, in the presence and absence of the extracts in a wide range of concentrations, did not differ significantly (p > 0.05; results not shown).
Table 5.
Inhibition of the proinflammatory enzymes.
| Analyte | LOX | HYAL | ||
|---|---|---|---|---|
| IC50 (μg/mL)∗ | IC50 (μg/U)∗∗ | IC50 (μg/mL)∗ | IC50 (μg/U)∗∗ | |
| MED | 188.1 ± 6.8e | 6.94 | 18.3 ± 0.7f | 16.0 |
| DEF | 91.6 ± 3.6b | 3.38 | 25.3 ± 0.9h | 22.2 |
| EAF | 89.8 ± 4.3b | 3.31 | 12.4 ± 0.5c | 10.9 |
| BF | 96.3 ± 3.7b,c | 3.55 | 4.1 ± 0.2a | 3.62 |
| WR | 265.3 ± 7.2f | 9.79 | 11.3 ± 0.5c | 9.94 |
| QU | 58.3 ± 2.3a | 2.15 | 15.6 ± 0.9d,e | 13.7 |
| RT | 104.8 ± 4.1c | 3.86 | 23.2 ± 1.6g | 20.4 |
| CHA | 114.3 ± 5.2d | 4.21 | 16.5 ± 0.7e | 14.5 |
| ECA | 90.6 ± 3.1b | 3.34 | 14.3 ± 0.7d | 12.5 |
| PB2 | 77.0 ± 1.8b | 2.84 | 12.8 ± 0.6d | 11.2 |
| IND | 63.0 ± 2.7a | 2.32 | 8.5 ± 0.4b | 7.46 |
Results are presented as means ± SD (n = 3) calculated per dry weight of the extract or standard. For extract codes, see Table 1. Different superscripts in each column indicate significant differences in the means at p < 0.05. ∗,∗∗Inhibition concentration (amount of analyte needed for 50% inhibition of enzyme activity) expressed as follows: ∗in μg of the dry extract or standard/mL of the enzyme solution; ∗∗in μg of the extracts/enzyme unit (U).
3.5. Influence of the Extracts on Cell Viability
The potential cytotoxicity of the extracts was evaluated in a model of PBMCs after 1-24-hour incubation with the extracts at 5-50 μg/mL. Regardless of the incubation time and extract concentration, the viability of the PBMCs treated with the extracts constituted 93.5-100.5% of that of the control (untreated) samples. Cellular safety of the extracts was evidenced by the lack of significant differences (p > 0.05) between the respective results (Figure 4).
Figure 4.
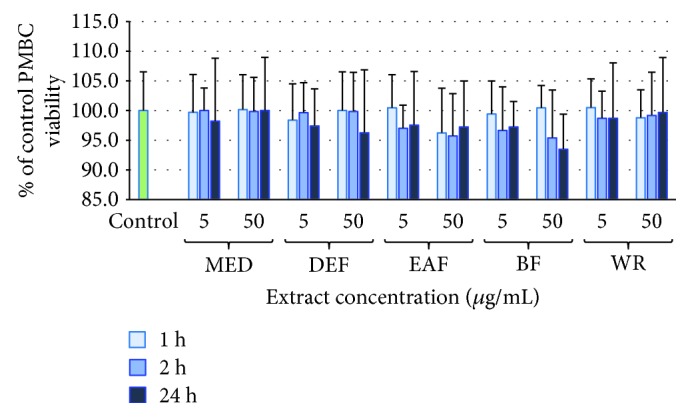
Viability of PBMCs after 1, 2, and 24 h of incubation with S. aucuparia flower extracts at 5 and 50 μg/mL. All values are not statistically different (p > 0.05).
4. Discussion
Standardised dry extracts from dietary and medicinal plants constitute the basis of modern phytotherapy as they contain concentrated active components and provide higher therapeutic effectiveness than unprocessed plant materials. Our previous studies had demonstrated that S. aucuparia flower is a promising candidate for cost-effective production of dry extracts rich in polyphenols [10, 14], but their phytochemical profiles and biological effects have remained inadequately recognised. In the present work, after fractionated extraction (which enabled enrichment of phenolic subfractions in active constituents) and subsequent LC-MS/MS profiling, we were able to detect and fully or tentatively identify 66 flower components, among which 52 and 50 structures were found for the first time in the analysed plant material and in the species S. aucuparia, respectively (Table 1). In contrast, only 17 constituents were observed earlier in the leaves [11] and 24 in the fruits of rowan [9]. A distinctive feature of the flower extracts was a vast diversity of the flavonol fraction (24 peaks) and high abundance of sexangularetin (8-methoxykaempferol) glycosides. As the derivatives of this flavonol are typical of bee pollen of rosaceous plants [36], they might be considered as a valuable marker for the identification of the origin of the extracts from the flowers. Flower-specific are also phenylamides (tricoumaroyl- and dicoumaroyl-caffeoyl spermidine isomers). This type of N-acylated biogenic amines, described here for the first time in the genus Sorbus, is usually reported in floral buds and reproductive organs with the suggested role in plant growth, flower development, and antimicrobial defence [33]. High analytical and chemotaxonomic value might be also ascribed to other new Sorbus constituents—flavalignans (three cinchonain I isomers). These compounds are phenylpropanoid-substituted flavanols in which two phenolic units are coupled by a two-bond lignin-like linkage [37]. Flavalignans are rare in nature, having been previously found, among others, in some species of Acer, Cinchona, and Trichilia [32, 37]. Although in the analysed Sorbus extracts flavalignans occurred at relatively low levels, the accumulated knowledge on their bioactivity suggests that by the additive and/or synergistic effects, they might influence the biological capacity of the extracts. According to Tang et al. [37], cinchonain I isomers are strong antioxidants, and their antiradical properties (towards DPPH) are up to four times stronger than those of (+)-catechin—one of the most effective antioxidants in vitro and in vivo [38]. Indeed, the cinchonains were concentrated in the most active fractions DEF and EAF (Figure 1, Table 2).
The quantitative survey of the Sorbus extracts revealed remarkable constancy of their polyphenolic profiles, reflected in similar levels of the main active components in flowers harvested in different years, i.e., 2009 [10] and 2015 (the present work). For example, the levels of TPC, TPA, TFA, and TCHA in the 2009 sample were 211.7, 111.6, 58.3, and 65.6 mg/g dw [10], while those reported here for the 2015 sample were 221.9, 110.9, 40.0, and 86.7 mg/g dw, respectively (Table 2). The intersample variations might be caused in part by different environmental (climatic) conditions in the year of harvest and in part by differences in the applied analytical protocols. For instance, the use of MS/MS detection in the present work enabled the identification of dicaffeoylquinic acid isomers, which then could have been accurately quantified using an authentic standard of cynarin (Figure 1, Table 2) instead of caffeic acid applied earlier [10]. Nevertheless, the relative stability of the chemical composition of the extracts indicates their great potential for industrial and phytotherapeutic applications.
The cooccurrence of high levels of flavonols and caffeic acid pseudodepsides in the investigated extracts suggests their possible significant biological effects in NCDs. Flavonols, especially QU glycosides, show a wide range of biological activities and are considered the most active compounds among flavonoids [39]. Their beneficial effects in NCDs depend largely on the nature of sugar units attached to the aglycones and on the other components of the plant matrix (emulsifiers) which may affect the solubility and bioavailability of the individual glycosides. One of the most effective flavonols is IQ, the bioavailability of which was up to 2.4-fold higher than that of QU and up to 6.4-fold higher than that of RT—the most abundant flavonoid in nature [40]. In consequence, ingestion of IQ as a pure compound (a 150 mg dose in humans) or in the plant matrix (a 62 mg/kg dose in rats) resulted in the plasma levels up to 5 μM (1.5 μg/mL) or 9.3 μM (2.8 μg/mL) of QU equivalents, respectively [40, 41]. Due to the advantageous bioavailability, IQ has been attracting increased attention, and its antioxidant, anti-inflammatory, anticarcinogenic, cardioprotective, antidiabetic, antiallergic, and neuropharmacological activity have been demonstrated ex vivo and in vivo [40]. Although IQ is widely distributed, its content in plant extracts is usually low; e.g., in the leaf extract of Morus alba, it amounts to 38.1 mg/g dw [40]. With the IQ levels up to 155.2 mg/g dw in EAF, the flower dry extracts of S. aucuparia appear thus promising sources of this valuable compound. Worth noting is also the abovementioned abundance of sexangularetin 3-glucoside in the analysed extracts (Table 2). Methoxylated flavonoids display improved intestinal absorption and enhanced resistance to the intestinal and hepatic metabolism and can be accumulated in mammalian tissues at up to 3.5-fold higher levels than their hydroxylated counterparts [42]. The relatively high bioavailability is also reported for caffeoylquinic acids, the dominant constituents of the Sorbus extracts. The plasma level of total monocaffeoylquinica acids and CHA after ingestion of green coffee bean extract may reach up to 14.8 μM (5.2 μg/mL) and 5.9 μM (2.1 μg/mL), respectively [43]. The best known source of these compounds is green coffee. The green coffee bean extract, standardised for total caffeoylquinic acids not less than 250 mg/g dw, is indicated mainly in the treatment of NCDs as antiatherogenic, antihypercholesteraemic, choleretic, antiobesity, hepatoprotective, and cardioprotective agent [44]. With the TCHA levels up to 169.0 mg/g dw (Table 2), the dry Sorbus extracts might be thus expected an attractive alternative for similar applications. The dominant groups of flavonols and caffeoylquinic acids were accompanied in the analysed extracts by flavanols and proanthocyanidins (Table 2), known antioxidant and anti-inflammatory plant constituents acting predominantly within the vascular system or the gastrointestinal tract, depending on the molecular mass and bioavailability [45]. The beneficial effects of proanthocyanidins in NCDs are attributed mainly to low-molecular-weight compounds and their ability to inhibit lipid peroxidation, reduce serum concentrations of proinflammatory cytokines and related inflammation of blood vessel walls, diminish capillary fragility and permeability, and decrease blood pressure [45]. As this group occurs in the analysed Sorbus extracts at relatively low levels, its possible influence on the extract activity in vivo is expected to be inferior to that of the dominant flavonols and caffeoylquinic acids. Nevertheless, considering possible additive and/or synergistic effects, the composition of the leaf extracts appears to be promising in the context of their potential application as antioxidant and anti-inflammatory agents, especially in the prevention of NCDs.
In our previous works, we had reported strong antiradical activity of the flower extracts of S. aucuparia towards synthetic free radicals: DPPH and ABTS, as well as their FRAP reactivity and ability to inhibit linoleic acid peroxidation in nonbiological systems [10, 14]. The present study investigated the interactions of the extracts with six radical and nonradical ROS involved in generation of oxidative stress in vivo. The primary ROS expressed by the human cells, e.g., by stimulated neutrophils during the inflammatory process, is O2∙– [3]. The ROS-mediated oxidative damage to biomolecules in vivo may result from the overproduction of O2∙– itself or from the oxidants derived in the downstream reaction cascade of O2∙–, such as H2O2, HClO, OH∙, and ONOO–. The most stable and diffusive form of ROS is H2O2, the harmful effects of which are connected with its selective reactivity with cysteine residues in proteins and ability to generate OH∙, the most reactive species in chemistry, able to attack and damage almost every molecule in the living cell [3, 46]. Highly destructive and nonselective oxidants are also HClO, the key antimicrobial agent involved, however, in the pathogenesis of many diseases, and ONOO–. The latter one, formed in the reaction between O2∙– and NO∙, causes protein nitration—one of the most dangerous processes in the cells [47]. The case of NO∙ is a good example of a dual nature of ROS. When produced by endothelial NO synthase (eNOS) in the vessel endothelium, it plays an important role in vessel dilatation and inflammatory protection (inhibition of leukocyte adhesion), but if synthesised in stimulated macrophages by inducible NO synthase (iNOS), it makes contact with O2∙– and rapidly forms the toxic ONOO– [48]. We found that the Sorbus extracts were able to inactivate all of the abovementioned ROS of physiological importance, and the effectiveness of the most active analytes was comparable to that of AA—the primary antioxidant of human plasma (Table 3). The correlation studies and experiments with the model compounds proved that the observed effects are determined by polyphenols (Table 4). The polyphenol-rich Sorbus extracts might be therefore expected to reduce the consequences of oxidative stress in biological systems.
This hypothesis was verified in an in vitro model of human plasma exposed to oxidative stress. The stress conditions were induced by ONOO–, one of the strongest oxidative and nitrative agents operating in vivo, involved in the pathophysiology of various inflammatory, neurodegenerative, metabolic, and especially cardiovascular disorders [49]. Although it is a short-lived oxidant, it easily crosses biological membranes and generates or decomposes into highly reactive ROS, such as OH∙, CO3∙–, NO∙, or ∙NO2, forming an aggressive cocktail of oxidants able to interact with the most critical biomolecules including plasma proteins and lipids [47]. The destructive effect of ONOO– is reflected, among others, in the increased plasma levels of nitrative/oxidative stress biomarkers such as 3-NT, LOOH, or TBARS and in the decrease of the NEAC of plasma [50]. The levels of these parameters have been connected with the progress or worse prognosis of many NCDs, including cardiovascular events, hypertension, diabetes, rheumatoid arthritis, chronic hepatitis, multiple sclerosis, and Alzheimer's disease [51].
In our experimental model, the plasma samples were treated with 100-150 μM ONOO– (depending on the test), which was sufficient to induce measurable changes in the levels of the oxidative stress biomarkers. Moreover, the applied concentrations correspond to the levels of ONOO– that can be reached in vivo in local compartments, e.g., during a serious inflammation of blood vessels [47]. The results indicated that the extracts might indeed have a beneficial impact on the plasma antioxidant status and protect its components against the harmful effect of oxidative stress (Figure 2). The significant effects obtained for the model extract constituents and the results of correlation studies (Table 5) suggest that low-molecular-weight polyphenols play a crucial role in the observed activity. On the other hand, worse correlation parameters and weaker dose dependency than observed in nonbiological models might indicate the impact of some other factors, such as the interactions between endogenous plasma constituents and the Sorbus phenolics. For example, the binding of polyphenols to proteins may suppress their antioxidant properties, and these masking effects may depend both on proteins and on polyphenols [15]. It is symptomatic that the relatively high correlation was observed by us in the 3-NT and FRAP tests. We suppose that the formation of the polyphenolic adducts may protect proteins against nitration on a mechanistic way, while the suppression of the direct reactivity of the adducts with oxidants may be less pronounced in the case of small molecules such as ferric ions (FRAP assay) than lipid-derived radicals formed in chain reactions during lipid peroxidation (LOOH and TBARS tests).
Nevertheless, the mechanism behind the observed protective effects was probably a direct and polyphenol-related scavenging of various ROS operating in plasma under the applied experimental conditions. It is consistent with the noticeable scavenging potential of the extracts towards ONOO– and some of the ONOO–-derived secondary radicals (OH∙, NO∙), as well as with the accumulating evidence of antiradical effects of polyphenols against CO3∙– and ∙NO2, formed in the fundamental reaction of ONOO– (with CO2) in biological systems [52]. The protective effects of the extracts in plasma were statistically significant (p < 0.05) at the concentrations as low as 1-5 μg/mL, equivalent to 0.11-3.0 μg GAE/mL, depending on the TPC value. Taking into account the results of the bioavailability studies that reported plasma levels of the model polyphenols (IQ, CHA, and ECA) in the range of 5-15 μM (1.5-5.2 μg/mL) [40, 41, 43], these concentrations seem possible to be achieved in vivo after ingestion of the analysed extracts. At these concentrations, their polyphenolic constituents might support the endogenous nonenzymatic antioxidant system by, among others, additive or sparing effects on the primary plasma antioxidant—ascorbic acid. It is well established that in various pathologic conditions, the plasma ascorbate level is below the optimal value of 50 μM (8.8 μg/mL) adequate to retain the redox homeostasis, and this decrease is one of the preconditions for the development of oxidative stress-related NCDs [53]. For instance, Deicher et al. [54] indicated that the ascorbate levels less than 32 μM (5.6 μg/mL) are associated with the increased risk of adverse cardiovascular events including myocardial infarction and death. Considering the protective effects of the Sorbus extracts in plasma at physiological levels as well as their antioxidant capacity towards the in vivo-relevant oxidants comparable to that of AA, the extracts might be expected to reduce the negative consequences of the disturbed redox homeostasis in vivo at the appropriate oral doses. In the context of future in vivo applications, it is also of note that at the wide range of concentrations (1-50 μg/mL), the extracts did not exhibit any prooxidant effects, did not deteriorate the viability of PBMCs, and thus may be regarded as safe (Figure 4).
Oxidative stress in vivo is closely linked with inflammation, and simultaneous influence on both processes is crucial for the effectiveness of antioxidant therapies [3]. In the present study, we investigated the anti-inflammatory potential of Sorbus extracts by determining their inhibitory activity towards three model enzymes involved in inflammation, which are some of the targets proposed for the treatment of inflammatory-related complaints [55, 56]. The first enzyme—LOX—is a prooxidant agent belonging to the family of lipoxygenases, enzymes catalysing the incorporation of dioxygen molecules into polyunsaturated fatty acids and formation of key chemokines and ROS, such as leukotrienes and O2∙–, associated with the development of numerous NCDs, e.g., atherosclerosis, myocardial infarction/reperfusion injury, rheumatoid arthritis, and cancer [56]. The second enzyme, HYAL, known as a spreading factor, hydrolyses hyaluronic acid, an important constituent of, i.e., endothelial surface layer, the disruption of which causes endothelium dysfunction and increases the instability of the atherosclerotic plate [55]. Our study also included XO, the prooxidant, O2∙–-generating enzyme that plays an important role in various ischemic and inflammatory diseases [57]. The results indicated that the extracts are inactive towards XO but are potent inhibitors of LOX and HYAL (Table 5). The inhibitory potential of the most active extracts was at most 1.5-fold lower than that of indomethacin—a commercial nonsteroidal anti-inflammatory drug. The activity towards LOX was found to be dependent on low-molecular-weight polyphenols (relatively well bioavailable), which is promising in the context of possible future application of the extracts. The anti-HYAL capacity was in turn related to poorly bioavailable condensed proanthocyanidins (TPA); the systemic effects of the TPA-rich extracts, especially BF, are thus less likely.
5. Conclusion
The present paper is the first detailed study of flower extracts of S. aucuparia and provides new insights into their phytochemical composition, standardisation, biological activity, and cellular safety. The results revealed that the extracts accumulate a vast diversity of bioactive polyphenols with the levels and structures promising for the use in the prophylaxis or adjunctive therapy of oxidative stress- and inflammation-related diseases. Some of the phenolic components, such as sexangularetin glycosides, phenylamides, and flavalignans, might be of chemotaxonomic importance and serve as analytical markers for the authentication of the extract origin. The main constituents, such as flavonols and phenolic pseudodepsides of quinic and shikimic acids, which were found responsible for antioxidant and anti-inflammatory activity of the extracts, might be in turn recommended as standardisation targets for routine quality control. Among the extracts, the defatted methanol-water (7 : 3, v/v) extract and its diethyl ether and ethyl acetate fractions appear to be the most advantageous for biological applications, considering both the yield and the activity in comparison to AA and IND. Some of their expected health benefits might be associated with the ability to neutralise multiple oxidants operating in human plasma, protect the plasma components (both proteins and lipids) against oxidative and nitrative damage, and increase the NEAC of plasma, as well as inhibit proinflammatory enzymes, especially LOX. However, although significant antioxidant protection was showed at physiological levels, the real extract effects should be verified in vivo in animal and clinical studies. As the observed effects might be related to possible antiatherogenic, anticoagulant, and antiplatelet functions of the extracts or their ability to influence endothelium, these issues should be first addressed in future research.
Acknowledgments
This work was financially supported by the Medical University of Lodz (grant nos. 503/3-022-01/503-31-001 and 503/3-022-01/503-36-001) and the University of Lodz (grant no. 506/1136). The authors would like also to express their gratitude to the Department of Pharmacognosy and Molecular Basis of Phytotherapy, Faculty of Pharmacy, Medical University of Warsaw, for enabling access to the UHPLC-MS equipment.
Abbreviations
- dw:
Dry weight
- MED:
Defatted methanol-water (7 : 3, v/v) extract
- DEF:
Diethyl ether fraction
- EAF:
Ethyl acetate fraction
- BF:
n-Butanol fraction
- WR:
Water residue
- NCDs:
Noncommunicable (chronic) diseases
- ROS:
Reactive oxygen species
- LOX:
Lipoxygenase
- HYAL:
Hyaluronidase
- XO:
Xanthine oxidase
- 3-NT:
3-Nitrotyrosine
- LOOH:
Lipid hydroperoxides
- TBARS:
Thiobarbituric acid-reactive substances
- FRAP:
Ferric reducing antioxidant power
- NEAC:
Nonenzymatic antioxidant capacity of plasma
- ONOO–:
Peroxynitrite
- PBMCs:
Peripheral blood mononuclear cells
- AA:
Ascorbic acid
- TX:
(±)-6-Hydroxy-2,2,7,8-tetramethylchroman-2-carboxylic acid (Trolox®)
- AAE:
Ascorbic acid equivalents
- TE:
Trolox® equivalents
- ECA:
(–)-Epicatechin
- PB2:
Procyanidin B2
- CHA:
Chlorogenic acid
- IQ:
Isoquercitrin
- RT:
Rutin
- QU:
Quercetin
- IND:
Indomethacin
- TPC:
Total phenolic content (Folin-Ciocalteu assay)
- GAE:
Gallic acid equivalents
- TPH:
Total phenolic content (HPLC)
- TFL:
Total flavonoid content
- TCFA:
Total content of caffeic acid derivatives
- TCHA:
Total content of CHA isomers
- TPA:
Total proanthocyanidin content (n-butanol/HCl assay)
- CYE:
Cyanidin chloride equivalents
- TLPA:
Total content of low-molecular-mass flavanols and proanthocyanidins (HPLC).
Data Availability
The source data used to support the findings of this study are available from the corresponding author upon request.
Conflicts of Interest
The authors declare that there is no conflict of interests regarding the publication of this paper.
References
- 1.Pandey K. B., Rizvi S. I. Plant polyphenols as dietary antioxidants in human health and disease. Oxidative Medicine and Cellular Longevity. 2009;2(5):270–278. doi: 10.4161/oxim.2.5.9498. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 2.Barrajón-Catalán E., Herranz-lópez M., Joven J., et al. Molecular promiscuity of plant polyphenols in the management of age-related diseases: far beyond their antioxidant properties. In: Camps J., editor. Oxidative Stress and Inflammation in Non-communicable Diseases - Molecular Mechanisms and Perspectives in Therapeutics. Vol. 824. Springer; 2014. pp. 141–159. (Advances in Experimental Medicine and Biology). [DOI] [PubMed] [Google Scholar]
- 3.Biswas S. K. Does the interdependence between oxidative stress and inflammation explain the antioxidant paradox? Oxidative Medicine and Cellular Longevity. 2016;2016:9. doi: 10.1155/2016/5698931.5698931 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 4.Hussain T., Tan B., Yin Y., Blachier F., Tossou M. C. B., Rahu N. Oxidative stress and inflammation: what polyphenols can do for us? Oxidative Medicine and Cellular Longevity. 2016;2016:9. doi: 10.1155/2016/7432797.7432797 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 5.Franz C., Chizzola R., Novak J., Sponza S. Botanical species being used for manufacturing plant food supplements (PFS) and related products in the EU member states and selected third countries. Food and Function. 2011;2(12):720–730. doi: 10.1039/c1fo10130g. [DOI] [PubMed] [Google Scholar]
- 6.McAllister H. A. The Genus Sorbus: Mountain Ash and Other Rowans. Kew, Richmond, Surrey, UK: Royal Botanic Gardens; 2005. [Google Scholar]
- 7.Kültür S. Medicinal plants used in Kirklareli Province (Turkey) Journal of Ethnopharmacology. 2007;111(2):341–364. doi: 10.1016/j.jep.2006.11.035. [DOI] [PubMed] [Google Scholar]
- 8.Shikov A. N., Pozharitskaya O. N., Makarov V. G., Wagner H., Verpoorte R., Heinrich M. Medicinal plants of the Russian Pharmacopoeia; their history and applications. Journal of Ethnopharmacology. 2014;154(3):481–536. doi: 10.1016/j.jep.2014.04.007. [DOI] [PubMed] [Google Scholar]
- 9.Kylli P., Nohynek L., Puupponen-Pimiä R., et al. Rowanberry phenolics: compositional analysis and bioactivities. Journal of Agricultural and Food Chemistry. 2010;58(22):11985–11992. doi: 10.1021/jf102739v. [DOI] [PubMed] [Google Scholar]
- 10.Olszewska M. A., Presler A., Michel P. Profiling of phenolic compounds and antioxidant activity of dry extracts from the selected Sorbus species. Molecules. 2012;17(3):3093–3113. doi: 10.3390/molecules17033093. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 11.Raudonė L., Raudonis R., Gaivelytė K., et al. Phytochemical and antioxidant profiles of leaves from different Sorbus L. species. Natural Product Research. 2015;29(3):281–285. doi: 10.1080/14786419.2014.950577. [DOI] [PubMed] [Google Scholar]
- 12.Hukkanen A. T., Pölönen S. S., Kärenlampi S. O., Kokko H. I. Antioxidant capacity and phenolic content of sweet rowanberries. Journal of Agricultural and Food Chemistry. 2006;54(1):112–119. doi: 10.1021/jf051697g. [DOI] [PubMed] [Google Scholar]
- 13.Olszewska M. A., Michel P. Antioxidant activity of inflorescences, leaves and fruits of three Sorbus species in relation to their polyphenolic composition. Natural Product Research. 2009;23(16):1507–1521. doi: 10.1080/14786410802636177. [DOI] [PubMed] [Google Scholar]
- 14.Olszewska M. A., Nowak S., Michel P., Banaszczak P., Kicel A. Assessment of the content of phenolics and antioxidant action of inflorescences and leaves of selected species from the genus Sorbus sensu stricto. Molecules. 2010;15(12):8769–8783. doi: 10.3390/molecules15128769. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 15.Ozdal T., Capanoglu E., Altay F. A review on protein-phenolic interactions and associated changes. Food Research International. 2013;51(2):954–970. doi: 10.1016/j.foodres.2013.02.009. [DOI] [Google Scholar]
- 16.Eghbaliferiz S., Iranshahi M. Prooxidant activity of polyphenols, flavonoids, anthocyanins and carotenoids: updated review of mechanisms and catalyzing metals. Phytotherapy Research. 2016;30(9):1379–1391. doi: 10.1002/ptr.5643. [DOI] [PubMed] [Google Scholar]
- 17.Michel P., Dobrowolska A., Kicel A., et al. Polyphenolic profile, antioxidant and anti-inflammatory activity of eastern teaberry (Gaultheria procumbens L.) leaf extracts. Molecules. 2014;19(12):20498–20520. doi: 10.3390/molecules191220498. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 18.Fu R., Zhang Y., Guo Y., Liu F., Chen F. Determination of phenolic contents and antioxidant activities of extracts of Jatropha curcas L. seed shell, a by-product, a new source of natural antioxidant. Industrial Crops and Products. 2014;58(24):265–270. doi: 10.1016/j.indcrop.2014.04.031. [DOI] [Google Scholar]
- 19.Czerwińska M., Kiss A. K., Naruszewicz M. A comparison of antioxidant activities of oleuropein and its dialdehydic derivative from olive oil, oleacein. Food Chemistry. 2012;131(3):940–947. doi: 10.1016/j.foodchem.2011.09.082. [DOI] [Google Scholar]
- 20.Banothu V., Neelagiri C., Adepally U., Lingam J., Bommareddy K. Phytochemical screening and evaluation of in vitro antioxidant and antimicrobial activities of the indigenous medicinal plant Albizia odoratissima. Pharmaceutical Biology. 2017;55(1):1155–1161. doi: 10.1080/13880209.2017.1291694. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 21.Krzyzanowska-Kowalczyk J., Kolodziejczyk-Czepas J., Kowalczyk M., Pecio Ł., Nowak P., Stochmal A. Yunnaneic acid B, a component of Pulmonaria officinalis extract, prevents peroxynitrite-induced oxidative stress in vitro. Journal of Agricultural and Food Chemistry. 2017;65(19):3827–3834. doi: 10.1021/acs.jafc.7b00718. [DOI] [PubMed] [Google Scholar]
- 22.Kolodziejczyk-Czepas J., Wachowicz B., Moniuszko-Szajwaj B., Kowalska I., Oleszek W., Stochmal A. Antioxidative effects of extracts from Trifolium species on blood platelets exposed to oxidative stress. Journal of Physiology and Biochemistry. 2013;69(4):879–887. doi: 10.1007/s13105-013-0264-5. [DOI] [PubMed] [Google Scholar]
- 23.Kolodziejczyk J., Olas B., Wachowicz B., Szajwaj B., Stochmal A., Oleszek W. Clovamide-rich extract from Trifolium pallidum reduces oxidative stress-induced damage to blood platelets and plasma. Journal of Physiology and Biochemistry. 2011;67(3):391–399. doi: 10.1007/s13105-011-0089-z. [DOI] [PubMed] [Google Scholar]
- 24.Kolodziejczyk-Czepas J., Nowak P., Kowalska I., Stochmal A. Biological activity of clovers-free radical scavenging ability and antioxidant action of six Trifolium species. Pharmaceutical Biology. 2014;52(10):1308–1314. doi: 10.3109/13880209.2014.891042. [DOI] [PubMed] [Google Scholar]
- 25.Matczak M., Marchelak A., Michel P., et al. Sorbus domestica L. leaf extracts as functional products: phytochemical profiling, cellular safety, pro-inflammatory enzymes inhibition and protective effects against oxidative stress in vitro. Journal of Functional Foods. 2018;40:207–218. doi: 10.1016/j.jff.2017.10.046. [DOI] [Google Scholar]
- 26.Clifford M. N., Johnston K. L., Knight S., Kuhnert N. Hierarchical scheme for LC-MSn identification of chlorogenic acids. Journal of Agricultural and Food Chemistry. 2003;51(10):2900–2911. doi: 10.1021/jf026187q. [DOI] [PubMed] [Google Scholar]
- 27.Clifford M. N., Knight S., Kuhnert N. Discriminating between the six isomers of dicaffeoylquinic acid by LC-MSn. Journal of Agricultural and Food Chemistry. 2005;53(10):3821–3832. doi: 10.1021/jf050046h. [DOI] [PubMed] [Google Scholar]
- 28.Cuyckens F., Claeys M. Mass spectrometry in the structural analysis of flavonoids. Journal of Mass Spectrometry. 2004;39(1):1–15. doi: 10.1002/jms.585. [DOI] [PubMed] [Google Scholar]
- 29.Hamed A. I., Al-Ayed A. S., Moldoch J., Piacente S., Oleszek W., Stochmal A. Profiles analysis of proanthocyanidins in the argun nut (Medemia argun - an ancient Egyptian palm) by LC-ESI-MS/MS. Journal of Mass Spectrometry. 2014;49(4):306–315. doi: 10.1002/jms.3344. [DOI] [PubMed] [Google Scholar]
- 30.Jaiswal R., Sovdat T., Vivan F., Kuhnert N. Profiling and characterization by LC-MSn of the chlorogenic acids and hydroxycinnamoylshikimate esters in maté (Ilex paraguariensis) Journal of Agricultural and Food Chemistry. 2010;58(9):5471–5484. doi: 10.1021/jf904537z. [DOI] [PubMed] [Google Scholar]
- 31.Vukics V., Guttman A. Structural characterization of flavonoid glycosides by multi-stage mass spectrometry. Mass Spectrometry Reviews. 2010;29(1):1–16. doi: 10.1002/mas.20212. [DOI] [PubMed] [Google Scholar]
- 32.Zhang L., Tu Z. C., Xie X., et al. Antihyperglycemic, antioxidant activities of two Acer palmatum cultivars, and identification of phenolics profile by UPLC-QTOF-MS/MS: new natural sources of functional constituents. Industrial Crops and Products. 2016;89:522–532. doi: 10.1016/j.indcrop.2016.06.005. [DOI] [Google Scholar]
- 33.Elejalde-Palmett C., de Bernonville T. D., Glevarec G., et al. Characterization of a spermidine hydroxycinnamoyltransferase in Malus domestica highlights the evolutionary conservation of trihydroxycinnamoyl spermidines in pollen coat of core eudicotyledons. Journal of Experimental Botany. 2015;66(22):7271–7285. doi: 10.1093/jxb/erv423. [DOI] [PubMed] [Google Scholar]
- 34.Olszewska M. A., Roj J. M. Phenolic constituents of the inflorescences of Sorbus torminalis (L.) Crantz. Phytochemistry Letters. 2011;4(2):151–157. doi: 10.1016/j.phytol.2011.02.005. [DOI] [Google Scholar]
- 35.Olszewska M. A., Michel P. Activity-guided isolation and identification of free radical-scavenging components from various leaf extracts of Sorbus aria (L.) Crantz. Natural Product Research. 2012;26(3):243–254. doi: 10.1080/14786419.2010.537271. [DOI] [PubMed] [Google Scholar]
- 36.Dauguet J. C., Bert M., Dolley J., Bekaert A., Lewin G. 8-Methoxykaempferol 3-neohesperidoside and other flavonoids from bee pollen of Crataegus monogyna. Phytochemistry. 1993;33(6):1503–1505. doi: 10.1016/0031-9422(93)85121-7. [DOI] [Google Scholar]
- 37.Tang W., Hioki H., Harada K., Kubo M., Fukuyama Y. Antioxidant phenylpropanoid-substituted epicatechins from Trichilia catigua. Journal of Natural Products. 2007;70(12):2010–2013. doi: 10.1021/np0703895. [DOI] [PubMed] [Google Scholar]
- 38.Grzesik M., Naparło K., Bartosz G., Sadowska-Bartosz I. Antioxidant properties of catechins: comparison with other antioxidants. Food Chemistry. 2018;241:480–492. doi: 10.1016/j.foodchem.2017.08.117. [DOI] [PubMed] [Google Scholar]
- 39.Perez-Vizcaino F., Duarte J. Flavonols and cardiovascular disease. Molecular Aspects of Medicine. 2010;31(6):478–494. doi: 10.1016/j.mam.2010.09.002. [DOI] [PubMed] [Google Scholar]
- 40.Valentová K., Vrba J., Bancířová M., Ulrichová J., Křen V. Isoquercitrin: pharmacology, toxicology, and metabolism. Food and Chemical Toxicology. 2014;68:267–282. doi: 10.1016/j.fct.2014.03.018. [DOI] [PubMed] [Google Scholar]
- 41.Manach C., Williamson G., Morand C., Scalbert A., Rémésy C. Bioavailability and bioefficacy of polyphenols in humans. I. Review of 97 bioavailability studies. The American Journal of Clinical Nutrition. 2005;81(1):230S–242S. doi: 10.1093/ajcn/81.1.230S. [DOI] [PubMed] [Google Scholar]
- 42.Cai H., Boocock D. J., Steward W. P., Gescher A. J. Tissue distribution in mice and metabolism in murine and human liver of apigenin and tricin, flavones with putative cancer chemopreventive properties. Cancer Chemotherapy and Pharmacology. 2007;60(2):257–266. doi: 10.1007/s00280-006-0368-5. [DOI] [PubMed] [Google Scholar]
- 43.Farah A., Monteiro M., Donangelo C. M., Lafay S. Chlorogenic acids from green coffee extract are highly bioavailable in humans. Journal of Nutrition. 2008;138(12):2309–2315. doi: 10.3945/jn.108.095554. [DOI] [PubMed] [Google Scholar]
- 44.Gil M., Wianowska D. Chlorogenic acids – their properties, occurrence and analysis. Annales Universitatis Mariae Curie-Sklodowska Lublin-Polonica. 2017;72(1):61–104. [Google Scholar]
- 45.Williamson G., Manach C. Bioavailability and bioefficacy of polyphenols in humans. II. Review of 93 intervention studies. The American Journal of Clinical Nutrition. 2005;81(1):243S–255S. doi: 10.1093/ajcn/81.1.243S. [DOI] [PubMed] [Google Scholar]
- 46.Marengo B., Nitti M., Furfaro A. L., et al. Redox homeostasis and cellular antioxidant systems: crucial players in cancer growth and therapy. Oxidative Medicine and Cellular Longevity. 2016;2016:16. doi: 10.1155/2016/6235641.6235641 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 47.Szabó C., Ischiropoulos H., Radi R. Peroxynitrite: biochemistry, pathophysiology and development of therapeutics. Nature Reviews Drug Discovery. 2007;6(8):662–680. doi: 10.1038/nrd2222. [DOI] [PubMed] [Google Scholar]
- 48.Cheng Y.-C., Sheen J.-M., Hu W. L., Hung Y.-C. Polyphenols and oxidative stress in atherosclerosis-related ischemic heart disease and stroke. Oxidative Medicine and Cellular Longevity. 2017;2017:16. doi: 10.1155/2017/8526438.8526438 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 49.Pacher P., Beckman J. S., Liaudet L. Nitric oxide and peroxynitrite in health and disease. Physiological Reviews. 2007;87(1):315–424. doi: 10.1152/physrev.00029.2006. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 50.Marrocco I., Altieri F., Peluso I. Measurement and clinical significance of biomarkers of oxidative stress in humans. Oxidative Medicine and Cellular Longevity. 2017;2017:32. doi: 10.1155/2017/6501046.6501046 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 51.Frijhoff J., Winyard P. G., Zarkovic N., et al. Clinical relevance of biomarkers of oxidative stress. Antioxidants & Redox Signaling. 2015;23(14):1144–1170. doi: 10.1089/ars.2015.6317. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 52.Miao J.-L., Wang W.-F., Pan J.-X., Lu C.-Y., Li R.-Q., Yao S.-D. The scavenging reactions of nitrogen dioxide radical and carbonate radical by tea polyphenol derivatives: a pulse radiolysis study. Radiation Physics and Chemistry. 2001;60(3):163–168. doi: 10.1016/S0969-806X(00)00387-X. [DOI] [Google Scholar]
- 53.German Nutrition Society (DGE) New reference values for vitamin C intake. Annals of Nutrition and Metabolism. 2015;67(1):13–20. doi: 10.1159/000434757. [DOI] [PubMed] [Google Scholar]
- 54.Deicher R., Ziai F., Bieglmayer C., Schillinger M., Hörl W. H. Low total vitamin C plasma level is a risk factor for cardiovascular morbidity and mortality in hemodialysis patients. Journal of the American Society of Nephrology. 2005;16(6):1811–1818. doi: 10.1681/ASN.2004100850. [DOI] [PubMed] [Google Scholar]
- 55.Girish K. S., Kemparaju K., Nagaraju S., Vishwanath B. S. Hyaluronidase inhibitors: a biological and therapeutic perspective. Current Medicinal Chemistry. 2009;16(18):2261–2288. doi: 10.2174/092986709788453078. [DOI] [PubMed] [Google Scholar]
- 56.Schneider I., Bucar F. Lipoxygenase inhibitors from natural plant sources. Part 2: medicinal plants with inhibitory activity on arachidonate 12-lipoxygenase, 15-lipoxygenase and leukotriene receptor antagonists. Phytotherapy Research. 2005;19(4):263–272. doi: 10.1002/ptr.1604. [DOI] [PubMed] [Google Scholar]
- 57.Pacher P., Nivorozhkin A., Szabó C. Therapeutic effects of xanthine oxidase inhibitors: renaissance half a century after the discovery of allopurinol. Pharmacological Reviews. 2006;58(1):87–114. doi: 10.1124/pr.58.1.6. [DOI] [PMC free article] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.
Data Availability Statement
The source data used to support the findings of this study are available from the corresponding author upon request.