

Abstract
Background
Insects frequently live in close relationship with symbiotic bacteria that carry out beneficial functions for their host, like protection against parasites and viruses. However, in some cases, the mutualistic nature of such associations is put into question because of detrimental phenotypes caused by the symbiont. One example is the association between the vertically transmitted facultative endosymbiont Spiroplasma poulsonii and its natural host Drosophila melanogaster. Whereas S. poulsonii protects its host against parasitoid wasps and nematodes by the action of toxins from the family of Ribosome Inactivating Proteins (RIPs), the presence of S. poulsonii has been reported to reduce host’s life span and to kill male embryos by a toxin called Spaid. In this work, we investigate the harmful effects of Spiroplasma RIPs on Drosophila in the absence of parasite infection.
Results
We show that only two Spiroplasma RIPs (SpRIP1 and SpRIP2) among the five RIP genes encoded in the S. poulsonii genome are significantly expressed during the whole Drosophila life cycle. Heterologous expression of SpRIP1 and 2 in uninfected flies confirms their toxicity, as indicated by a reduction of Drosophila lifespan and hemocyte number. We also show that RIPs can cause the death of some embryos, including females.
Conclusion
Our results indicate that RIPs released by S. poulsonii contribute to the reduction of host lifespan and embryo mortality. This suggests that SpRIPs may impact the insect-symbiont homeostasis beyond their protective function against parasites.
Electronic supplementary material
The online version of this article (10.1186/s12866-019-1410-1) contains supplementary material, which is available to authorized users.
Keywords: Spiroplasma, Endosymbiosis, Ribosome inactivating protein, Drosophila
Background
Endosymbiosis refers to a persistent interaction between two partners, generally a eukaryotic host and a microbial symbiont that lives within the host’s body. Such interactions are particularly frequent in insects, of which more than half of species are estimated to harbor at least one endosymbiont [1, 2]. Insect endosymbionts can affect their host in multiple ways, including beneficial effects such as metabolic complementation, heat tolerance or protection against viruses and parasites [3–7]. However some endosymbiotic associations can also have detrimental consequences for the insect fitness, such as a decreased lifespan or fertility [8, 9]. Among the most widespread facultative endosymbionts that manipulate insect reproduction are the genera Wolbachia and Spiroplasma [10].
Spiroplasma poulsonii (hereafter Spiroplasma) is a natural symbiont of the fruit fly Drosophila melanogaster [11–13]. It lives extracellularly in the fly hemolymph and is vertically transmitted by trans-ovarial transfer. Spiroplasma colonizes the germline during vitellogenesis by co-opting the yolk transport and internalization machinery [14]. Intriguingly, it completely lacks a cell-wall and thus immunogenic surface molecules, such as peptidoglycan, which renders it invisible for the host immune system [15–19]. In adult flies, Spiroplasma grows over time reaching a titer of 105-106 bacteria per μl of hemolymph [20]. Spiroplasma infection shortens the lifespan of Drosophila, suggesting that either the bacteria causes damages only at high titer, or that the damages take time to kill the host [19]. Interestingly, the growth of S. poulsonii is limited by the availability of host lipids, preventing its overgrowth in condition of nutrient scarcity [19].
One of the most striking phenotypes caused by Spiroplasma is male-killing, whereby infected male embryos die during their development while most infected females survive [12]. As Spiroplasma is only transmitted by female flies, male-killing is thought to favor the spread of the bacteria among host natural populations [21]. Recently, a Spiroplasma toxin containing Ankyrin-repeats, named Spiroplasma Androcidin (Spaid), has been described as a crucial male-killing agent [22, 23]. Heterologous expression of Spaid in uninfected flies is sufficient to kill males. Moreover, its expression during early embryogenesis induces DNA-damage-dependent apoptosis and defective neurogenesis in uninfected male embryos, which fully recapitulates male-killing phenotypes [24–27].
Studies have shown that in some contexts, Spiroplasma can also provide a benefit to its host as they mediate protection against parasitoid wasps and nematodes in several Drosophila species [28–31]. Protection is a major ecological benefit that can lead to a fast spreading of Spiroplasma in wild populations [32]. Two complementary mechanisms have been implicated in Spiroplasma protection against parasites: a metabolic competition for host lipids between Spiroplasma and the parasites, and Spiroplasma production of Ribosome-Inactivating Proteins (RIPs) that damage ribosomes of both wasp eggs and nematodes [28, 30, 31]. RIPs are found in plants and bacteria, where they act as a defense against eukaryotic parasites [30, 31, 33–35]. They recognize a conserved region of the 28S ribosomal RNA called the Sarcin-Ricin Loop (SRL). The secondary structure of the SRL consists in a hairpin loop displaying an adenine that is necessary for protein synthesis [36]. RIPs cleave the central adenine from the SRL in a process called depurination, thus blocking protein synthesis [37].
In this article we investigated the role of RIPs produced by the facultative endosymbiont S. poulsonii (hereafter SpRIPs) in its natural host D. melanogaster. Similarly to the reduced lifespan observed in infected flies, we show that heterologous expression of SpRIPs coding genes in uninfected flies shortens their life span. Furthermore, uninfected-embryos expressing SpRIPs have high mortality rate and a female-biased sex-ratio among the surviving individuals, suggesting that males may be more sensitive to the ectopic expression of this toxin.
Results
SpRIPs depurinate the 28S rRNA of D. melanogaster
S. poulsonii genome contains five genes encoding RIPs (SpRIP1-5) [20, 30, 31, 38]. All of them have a signal peptide, suggesting a secretion of the mature protein, and a conserved N-glycosidase domain in charge of the depurination reaction [30, 31]. All copies are chromosomal, suggesting that they are very stable compared to other endosymbiont toxin coding genes that are located on plasmids or mobile elements such as Spaid in S. poulsonii or the cytoplasmic incompatibility factor of Wolbachia [20, 23, 39]. Transcriptome analysis has shown that only two of them, SpRIP1 and SpRIP2, are significantly expressed in vivo and in vitro, pointing to a possible pseudogenization of SpRIP3, 4 and 5 [20, 30].
To confirm the expression pattern of SpRIPs in infected flies, we performed RT-qPCR analysis on each of the SpRIPs. We confirmed that SpRIP1 and SpRIP2 are strongly expressed by S. poulsonii in D. melanogaster with no significant changes in expression level along the fly life cycle (Fig. 1a and b). Very low levels of SpRIP 3, 4 and 5 transcripts were detected, in accordance with the literature (Additional file 1: Figure S1). We then measured RIP activity using a RT-qPCR assay. This assay relies on the ability of reverse transcriptases to incorporate a thymine in complementary DNA in place of the void position present on the depurinated RNA molecule. It is then possible to design primers that bind specifically to the intact cDNA (containing an adenine) or to the depurinated one (containing a thymine) [31]. Comparisons between infected and uninfected flies confirmed that S. poulsonii depurinates the 28S rRNA of Drosophila, as previously shown for larvae and 1 week old adult flies [30]. Monitoring RIP activity along the whole Drosophila lifecycle revealed particularly high levels of depurination in embryos and old adult flies (Fig. 1c). A control assay using primers amplifying fragments outside of the SRL showed that the total number of 28 rRNA transcripts was the same between infected and uninfected flies (Fig. 1d). As the level of expression of SpRIP in Spiroplasma is constant, the high level of depurination in embryos and old adult flies likely results from the higher Spiroplasma titer in the host at these stages [19].
Fig. 1.

a SpRIP1 expression level in infected flies along Drosophila development stages (One way ANOVA; development stage p = 0.9055). b SpRIP2 expression level in infected flies along Drosophila development stages (One way ANOVA; development stage p = 0.5129). c RIP activity in infected flies compared to uninfected flies (Two way ANOVA; Spiroplasma infection p*** < 0.0001; development stages p*** < 0.0001; interaction p*** < 0.0001). d Intact 28S rRNA quantification in infected versus uninfected flies along Drosophila development stages
SpRIP1 and SpRIP2 expression is toxic for Drosophila melanogaster
We generated four different transgenic fly lines expressing singly SpRIP1, SpRIP2, BiP + SpRIP1 or BiP + SpRIP2 under the control of the GAL4/UAS system [40]. BiP is a signal peptide used to trigger the secretion of proteins in D. melanogaster [41, 42]. The toxicity of these constructs was tested using the “Rough Eye Phenotypes” (REP) assay, which allows to study the activity of a putative toxin driven by an eye-specific driver (ey-GAL4) to observe eventual deleterious effect of the protein on this organ’s structure [43, 44]. The REP assay allows to study toxin activity by monitoring defects including loss of bristles, fusion of ommatidias, necrosis, loss of pigmentation and reduced eye size [43, 45]. All control flies developed a normal eye structure. On the contrary, flies expressing UAS-SpRIPs under ey-GAL4 control developed a reduced eye along with severe abnormalities, and in some cases no eye at all (Fig. 2). This demonstrates that both SpRIP1 and 2 act as toxins on Drosophila cells.
Fig. 2.
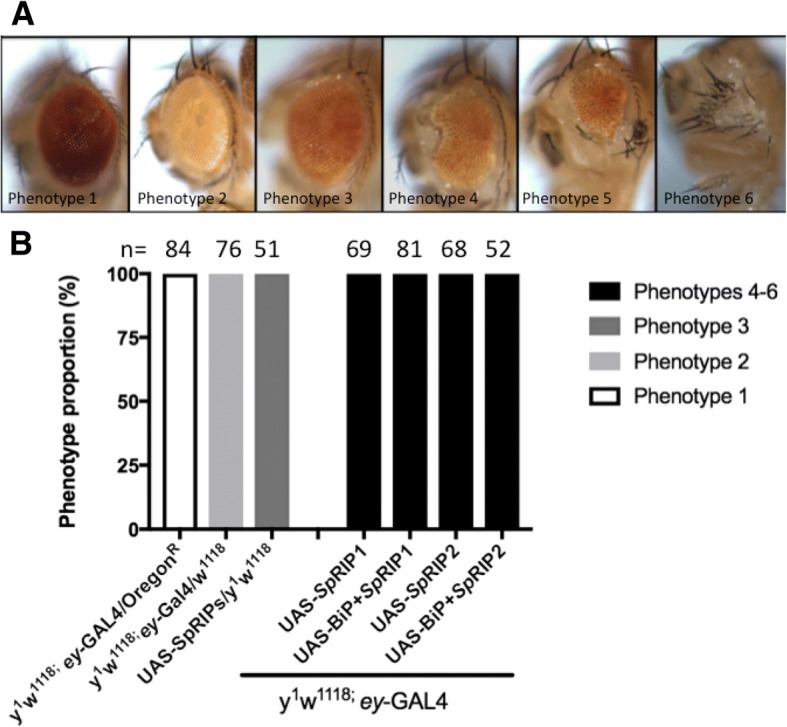
Rough Eye Phenotype assay. a Representative bright-field images of Drosophila eye phenotypes obtained during the assay. b Percentage of each phenotype. n indicates the number of flies obtained for each cross. Each cross has been repeated three independent times
Ectopic expression of SpRIP1 and SpRIP2 decreases uninfected flies life span
Spiroplasma-infected flies have a shorter lifespan compared to uninfected ones [16]. Moreover, old infected flies have been reported to have a decreased climbing activity which suggests neurological damages [19]. We first confirm this phenotype, observing that infected flies have a lifespan reduced by about 20 days (Fig. 3 and Additional file 2: Figure S2). As Spiroplasma resides in the hemolymph, we hypothesized that the impact of Spiroplasma on host lifespan could be due to accumulation of a toxin released in the hemolymph. Accordingly, proteomics analysis of hemolymph of 2 weeks old Spiroplasma-infected flies revealed the presence of SpRIP1 and SpRIP2 (S. Rommelaere, F. Masson, and B. Lemaitre, unpublished data).
Fig. 3.

Effect of SpRIP2 expression on D. melanogaster lifespan. Sp- and Sp + refer to uninfected or Spiroplasma-infected condition respectively. UAS constructs were driven by the ubiquitous Da-GAL4 driver. Sample labels are ordered from the shortest to the longest lifespan. Plain lines represent uninfected stocks and controls. Dashed lines represent either infected flies or expressing SpRIPs. Pairwise comparison of survival fits where analyzed by Log-rank (Mantel-Cox) test
To further address the impact of SpRIPs on Drosophila viability, we tested the effect of ectopic expression of SpRIPs on the life span of uninfected flies. Drosophila expressing SpRIP1 or BiP + SpRIP1 constructs did not develop further than larval instars, preventing the use of these constructs for lifespan analysis. Interestingly, uninfected flies expressing SpRIP2 and BiP + SpRIP2 had a markedly decreased lifespan by about 30 days in average compared to uninfected flies, which live about 75 days (Fig. 3; Logrank test p*** < 0.0001). The lifespan of these transgenic lines was also shorter than the one of infected flies, which live about 45 days (Fig. 3; Logrank test p*** < 0.0001). The lifespans seem to be depending on the expression level and activity of the RIPs (Additional file 3: Figure S3 and Additional file 4: Figure S4).
To further test the implication of SpRIPs in premature adult lethality, we generated a transgenic fly line expressing a 1492 bp fragment of the 28S rRNA under the control of a UAS upstream sequence [40]. This fragment contains the conserved SRL targeted by RIPs and was designed to buffer RIP activity by increasing the number of targets for the toxin, thus working as an antidote. Spiroplasma-infected flies with ubiquitous expression of SRL fragment display an increase in their lifespan by about 5 days compared to infected wild-type flies (Fig. 3; Logrank test p**** < 0.0001). Collectively, these results are consistent with the implication of SpRIP in shortening Drosophila life span.
Spiroplasma-infected flies and uninfected flies expressing SpRIP2 have reduced hemocyte count
As S. poulsonii is found in the fly hemolymph, we hypothesized that hemocytes should be the most affected cell type by RIP toxins. We thus visualized hemocyte in Spiroplasma-infected and uninfected adult flies, carrying the hemocyte marker Hml-GAL4 > UAS-GFP. In uninfected adult flies, sessile hemocytes are found in patches beneath the cuticle in the middle of the dorsal abdomen [46]. Interestingly, Spiroplasma-infected flies have reduced number of sessile patches (Fig. 4a). To confirm this observation, we indirectly estimated the number of hemocytes in adult flies by monitoring the expression of hemolectin (hml), a gene which expression is hemocyte-specific. Consistent with a reduction of the number of hemocytes, the expression of hml was halved in Spiroplasma-infected flies compared to uninfected ones in two different wild type strains (Fig. 4b). We conclude that the presence of Spiroplasma greatly reduces the number of hemocytes. This reduction could reflect the shortening of lifespan as a decreased hemocyte count is one of the hallmark of aging in flies [47]. To test whether SpRIPs could mediate this effect, we monitored the level of hemocytes in adult flies expressing SpRIP2 and BiP + SpRIP2 under the control of two ubiquitous GAL4 drivers. Hml expression quantification revealed a decrease in the number of hemocytes in these flies similar to the decrease observed upon Spiroplasma infection (Fig. 4c and Additional file 5: Figure S5). These results suggest that SpRIPs cause hemocytes death, which in turn could contribute to aging and premature death of flies.
Fig. 4.
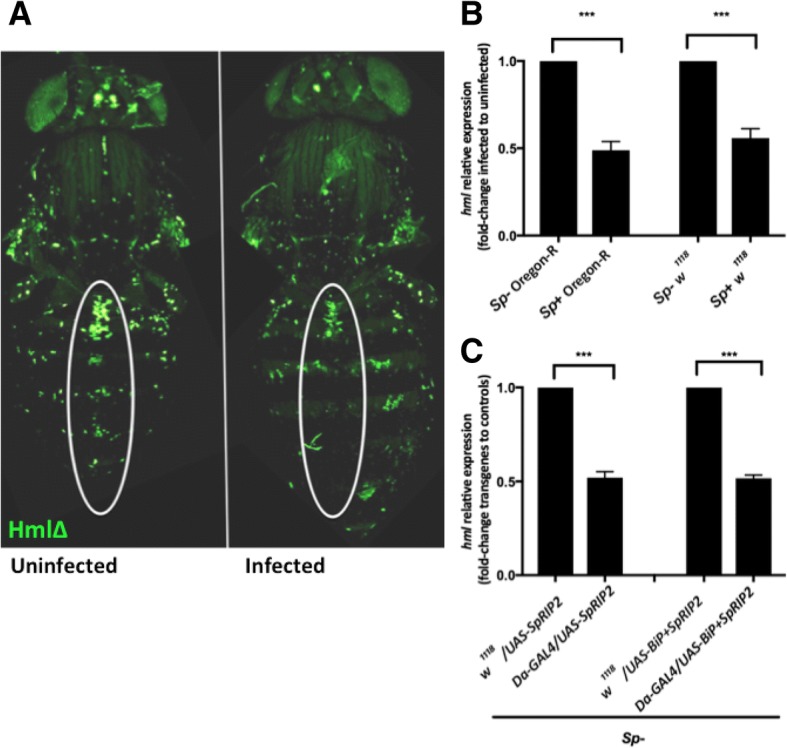
a Confocal image of HmlΔ-GAL4/UAS-GFP adult flies. In uninfected flies, hemocyte patches are mostly located within the white circle, following the antero-posterior axis. In infected flies only a few patches remain and have lower fluorescence intensity. b hml transcription level in infected and uninfected adult wild type flies (Dunnett’s multiple comparisons test p*** < 0.0001). c hml transcription level in lines expressing UAS-SpRIP2 and UAS-BiP + SpRIP2 under Da-GAL4 control. Expression of both constructs lead to a decrease in hemocyte number (Dunnett’s multiple comparisons test p*** < 0.0001). Sp- and Sp + refer to uninfected or Spiroplasma-infected condition respectively. Controls are normalized as 1
SpRIPs ectopic expression causes embryo mortality revealing higher in male embryos compare to female ones
We have shown that RIP activity is particularly high in Spiroplasma-infected embryo compared to other developmental stages (Fig. 1c) raising the possibility that SpRIP1 and SpRIP2 could contribute to embryo mortality. To test this possibility, we first monitored the effect of the ectopic expression of SpRIPs in uninfected individuals by using either the ubiquitous zygotic Da-GAL4 driver in embryos or the maternal driver MTD-GAL4. We monitored embryo mortality as the percent of embryos that do not hatch, which is about 5% in uninfected wild type embryos (Fig. 5a). All uninfected embryos with ectopic expression of SpRIP1 or BiP + SpRIP1 die, reflecting the high toxicity of SpRIP1. However, the expression of UASp-SpRIP2 kills about 70% of the embryos (Dunnett’s multiple comparison test against uninfected w1118 p** < 0.0074). Interestingly, over-expression of UASp-BiP + SpRIP2 shows a lower toxicity with a mortality rate up to 30% (Dunnett’s multiple comparison test against uninfected w1118 p*** < 0.0001) (Fig. 5a). We hypothesized that the secretion of the toxin out of the embryo’s cells reduces its toxicity. To reinforce the hypothesis that RIP activity is indeed responsible for embryo death, we measured RIP activity during embryogenesis for each construct. We observed a correlation between the level of RIP activity and the mortality (Pearson’s correlation test p*** < 0.001) (Additional file 6: Figure S6), suggesting that the mortality indeed results from RIP activity.
Fig. 5.
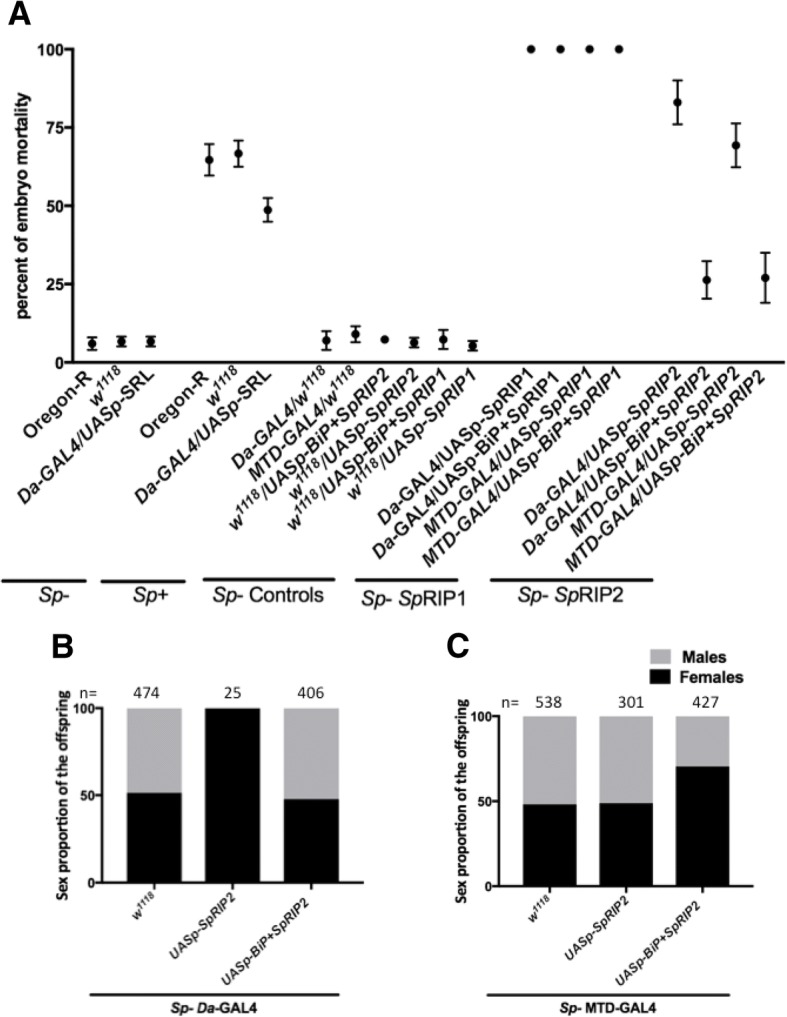
a Effect of SpRIP expression on embryo mortality. b Sex ratio of uninfected flies expressing UASp-SpRIP2 under Da-GAL4 control (ubiquitous). c Sex ratio of uninfected flies expressing UASp-SpRIP2 under MTD-GAL4 control (maternal specific). n indicates the number of adult flies counted for the assay. Sp- and Sp + refer to uninfected or Spiroplasma-infected condition respectively
To further test the possible implication of SpRIPs in embryo mortality, we took advantage of the UASp-SRL construct by analyzing whether buffering RIP activity with additional SRL target could rescue Spiroplasma-infected embryos from dying. We first observed that embryonic lethality reaches about 65% in Spiroplasma-infected flies, well above the expected 50% if males only were dying. This suggests that not only does Spiroplasma kill males, but also a small fraction of the female progeny, roughly estimated at 12.5% (Dunnett’s multiple comparison test p*** < 0.0001). Interestingly, ectopic expression of SRL slightly decreases mortality of infected embryos by 10% (Dunnett’s multiple comparison test p*** < 0.0001) (Fig. 5a). The sex ratio of the surviving embryo was still 100% females, suggesting that the buffering of RIP activity by the UASp-SRL construct is sufficient to rescue females but not males (Fig. 5a).
While the sex-ratio of uninfected hatching flies is of 50% females and 50% males (Fig. 5b, c and Additional file 7: Figure S7), the sex-ratio of uninfected D. melanogaster hatching flies from embryos expressing SpRIP2 were biased toward female. It ranged from 70% when the construct is under the control of the maternal driver MTD (Fisher’s exact test p*** < 0.001) to 100% when the ubiquitous driver Da-GAL4 was used (Fisher’s exact test p*** < 0.001) (Fig. 5b and c), suggesting that males are more sensitive to RIP activity.
Discussion
S. poulsonii protects its host against macro-parasites such as nematodes and parasitoid wasps and RIP toxins has been shown to play a major role in this protection [30, 31]. In this study, we provide evidence that Spiroplasma RIPs could affect symbiosis beyond their implication in endosymbiont-mediated protection by harming the host and contributing to lifespan shortening and embryo mortality.
We show that only two out of the five putative RIP genes contained in Spiroplasma genome are expressed all along the life cycle of Drosophila with peaks during embryogenesis and late adulthood. As S. poulsonii titer increases with time [19], we hypothesize that the peak in old adults is a consequence of the high density of Spiroplasma in the host hemolymph, rather than a change in the gene expression in the bacteria. Infected embryos also reveal particularly high RIP activity, likely due the transmission of RIPs and of already depurinated ribosomes from the mother. Our over-expression studies were carried out with the ubiquitous Da-GAL4 driver as it revealed the closest expression level to natural infection. Such studies confirm that SpRIP1 and SpRIP2 target the 28S rRNA of its host as previously shown [30]. Transgenic fly lines expressing SpRIP1 display a higher mortality rate for embryos and larva than those expressing SpRIP2, which may result from a higher RIP1 transgene expression (Additional file 3: Figure S3). Last, the addition of a Drosophila secretion signal to the protein sequence tends to reduce its toxicity, which is consistent with SpRIPs targeting 28S rRNA within the cells.
Previous studies have shown that Spiroplasma shortens the fly lifespan but the underlying mechanism was unknown, although the synthesis of cardiolipins by the bacteria has been proposed as a cause [19]. Our results suggest that Spiroplasma RIPs contribute to the premature death of infected flies. According to this model, the increasing Spiroplasma titer in aging flies is accompanied by an increase of SpRIP release in the fly hemolymph, which eventually damages the host tissues. Ectopic expression of SpRIPs within cells can be more toxic than natural bacterial expression, as ribosomes are then more exposed to the toxin. This situation is however happening naturally only during the first 3 h of embryogenesis (before the cellularization), after which Spiroplasma is extracellular. The addition of a Drosophila secretion signal to the toxin thus better mimics the natural situation. However, Drosophila life span remained shortened even upon SpRIP secretion, suggesting that the toxin is able to enter the cells and depurinate ribosomes efficiently.
Similarly to the phenotype of Spiroplasma-infected flies, we show that over-expression of SpRIP1 and SpRIP2 are associated with an increase in embryo lethality, a shorter lifespan and a decrease in hemocytes number. While SpRIPs contribute to the protection against Drosophila’s parasites, our study suggests that these toxins have also a strong detrimental effect in the host with a tangible impact in late adulthood. This suggests that Spiroplasma has not developed any mechanism to shut down RIP expression at the adult stage or in absence or parasite infections. Maintaining a constitutive SpRIP production could be a way to react as quickly as possible to parasite infections at a low cost for the host. It is indeed likely that the fitness cost associated with lifespan reduction in Drosophila is minimal, as most eggs are laid during the first 2 weeks [48].
Conclusion
Insect endosymbioses encompass a continuum of interactions ranging from mutualism to parasitism. In some cases however, assessing the beneficial or detrimental nature of the interaction for the host can be delicate. The Spiroplasma/Drosophila symbiosis is a prime example of such versatile ecological outcome: the bacteria protects its host against widespread parasites, conferring a major ecological benefit, but also kills male progeny and drastically reduces the adults lifespan, reflecting a pathogenic interaction. SpRIPs are involved in these two different faces of Spiroplasma endosymbiosis. They are directly involved in host protection against parasites, but can also cause strong damage to the host in absence of parasite infection, making them the first described endosymbiont-encoded toxins to directly harm its adult host.
Methods
Fly stocks and handling
Infected lines were generated in 2011 by injection of Spiroplasma-infected hemolymph in Oregon-R females. [17]. Infected lines have been maintained in the laboratory establishing genetically identical lines of Oregon-R Spiroplasma infected and uninfected. Hemocytes were observed on 4 weeks old females w1118; HmlΔGAL-4 > UAS-GFP [49]. For all the experiments, flies were maintained at 25 °C on standard cornmeal medium. Embryos were collected from 5 to 7 days old flies by using cages and yeasted grape juice plates. Lifespan experiments were done as described in [17]. The driver for REP assay (ey-GAL4) was obtained from Bloomington stock center (#8221). All experiments have been repeated three independent times.
RNA, DNA extractions and RT-qPCR
RNA, DNA extractions, and RT-qPCR were performed as described in [17, 19, 28]. Reverse transcription was done using 500 ng of RNA per sample, which was isolated from 3 adult flies, 3 larvae, 3 pupae, or 100 to 300 embryos. SpRIPs expression and activity were measured along the whole life cycle by RT-qPCR. Expression for each SpRIP was analyzed individually except for SpRIP3, SpRIP4 & SpRIP5 that were measured with a single pair of primers because of their high sequence identity. RT-qPCR calculations for the expression level of SpRIPs was done following the ΔCT method normalizing by dnaA expression level. Primers for dnaA are described in [17]. RT-qPCR calculations to compare the expression level of SpRIPs between the transgenic fly lines and Spiroplasma-infected flies was done following the ΔCT method normalizing by rps17 expression level. Primers for UAS-SpRIP1 expression are Forward: 5′- CGT AGC AGGTGGTGTTGTTC-3’ Reverse: 5′- GCTTCACCCACATCAGCAAG-3′ (efficiency = 1.81). Primers for UAS-SpRIP2 expression are Forward: 5′- CGT AGC TCGATACCAGCGTGACCATC-3’ Reverse: 5′- CGTTCTGCAGGTTGTACTCG-3′ (efficiency = 1.94). RIP activity assay was performed as described in [31]. All calculations for RIP activity and hemocyte count have been done following the ΔΔCT method and these figures represent the fold change between the experimental condition samples and the controls which are valued as 1 [50]. Primers for hml are: Forward: 5’-GAGCACTGCATACCCCTACC-3’ Reverse: 5’-CCGTGCTGGTTACACTCCTT-3′ (efficiency = 1.88). Gene expression levels were normalized to rps17. Figures and statistical results were obtained using GraphPad Prism 7.0b software. All experiments have been repeated three independent times.
Design and construction of UAS-SpRIP1 and UAS-SpRIP2 constructs
Spiroplasma has an alternative genetic code and a strong codon bias compared to Drosophila [38]. SpRIP1 and SpRIP2 gene sequences were codon optimized for insect translation using Geneious v8.1.9. The secretion signal from the sequence of BiP (Hsc70-3) was added at the 5′ end of the RIP genes flanked by two BglII restriction sites. The optimized BiP-SpRIP1 and 2 were fully synthesized and cloned in a pDONR221 vector for Gateway cloning by Invitrogen GeneArt gene synthesis services. Optimized SpRIP1 and 2 were obtained from BiP + SpRIP1 and 2 by digestion of the BiP sequence by BglII and re-ligation of the plasmid on itself. The fragment of 28S rRNA was amplified from Oregon-R flies and also cloned in pDONR221. All transgenes were cloned into a UASp and a UASt vector by Gateway LR reaction and injected in D. melanogaster w1118 embryos by Bestgene Inc., Chino Hills, USA.
Embryo mortality assay
A total of 100 embryos were collected per genotype on grape juice plates 15-20 h after egg laying. After ten more hours, the remaining embryos that did not hatch (dead) were counted. Experiments were done simultaneously with two different drivers, the ubiquitous Da-GAL4, and the maternal MTD-GAL4. All experiments have been repeated three independent times.
Additional files
Figure S1. SpRIPs 3,4 and 5 expression level in infected flies along Drosophila development stages (One way ANOVA; development stage p = 0.9992). (TIF 1384 kb)
Figure S2. Lifespan of infected and uninfected wild types flies (controls for Fig. 3). Sp- and Sp + refer to uninfected or Spiroplasma-infected condition respectively. (TIF 1983 kb)
Figure S3. Expression level of (A) SpRIP1 and (B) SpRIP2 in embryos from uninfected, infected and transgenic fly lines expressing UAS-RIP normalized by host rsp17 transcript level (One way ANOVA; p** = 0.0031 for SpRIP1 and p** = 0.0049 for SpRIP2). (C) Expression level of SpRIP2 in adults from uninfected, infected and transgenic fly lines expressing UAS-RIP normalized by host rsp17 transcript level (One way ANOVA; p** < 0.0081). (TIF 5534 kb)
Figure S4. (A) Comparison of RIP activity in Spiroplasma-infected embryos with uninfected transgenic embryos expressing SpRIPS (One way ANOVA; SpRIP1 p*** = 0.001; One way ANOVA; SpRIP2 p** = 0.0021). (B) Intact 28S rRNA quantification in infected embryos and uninfected transgenic fly lines (C) Comparison of RIP activity in Spiroplasma-infected adult flies with uninfected transgenic adult fly (One way ANOVA; SpRIP2 p*** = 0.001) (D) Intact 28S rRNA quantification in infected adult flies and uninfected transgenic adult flies. (TIF 6378 kb)
Figure S5. hml transcription level in lines expressing UAS-SpRIP2 and BiP + SpRIP2 under actin-GAL4 control. Expression of both constructs also leads to a decrease in hemocyte number (Dunnett’s multiple comparisons test p*** < 0.0001). Controls are normalized as 1. (TIF 1147 kb)
Figure S6. Correlation between RIP activity in embryos aged 0 to 24 h after egg laying and embryo mortality (Pearson’s test p < 0.0001). RIP activity in infected wild types was normalized by uninfected samples. Transgenic fly lines were normalized by Da-GAL4/w1118. Controls are normalized as 1. (TIF 3499 kb)
Figure S7. Sex ratio of the control fly lines for Fig. 5. Sp- and Sp + refer to uninfected or Spiroplasma-infected condition respectively. (TIF 4019 kb)
Acknowledgments
We sincerely thank Steve J. Perlman (Department of Biology, University of Victoria, Canada) along with two people from his laboratory, Matthew J. Ballinger and Phineas T. Hamilton for the constructive discussions and the sharing of information. We also thank Maroun Bou Sleiman, Samuel Rommelaere and Toshiyuki Harumoto (School of Life Science, École Polytechnique Fédérale de Lausanne) for their comments and discussion about this project.
Funding
This work was funded by ERC (European Research Council) advanced grant no. 339970 and the SNF (Swiss National Science Foundation) Sinergia grant no. CRSII3_154396.
Availability of data and materials
All data generated or analyzed during this study are included in this published article (and its additional files).
Abbreviations
- ARP
Adhesion Related Proteins
- qPCR
Quantitative Polymerase Chain Reaction
- REP
Rough Eye Phenotypes
- RIP
Ribosome Inactivating Protein
- RT
Reverse Transcription
- Spaid
S. poulsonii androcidin
- SpRIP
S. poulsonii Ribosome Inactivating Protein
Authors’ contributions
GGA performed fly handling assays and RT-qPCR experiments and drafted the manuscript. FM collaborated in the creation of transgenic fly lines. FM, JCP and BL supervised the project and contributed to the manuscript. All authors read and approved the final manuscript.
Ethics approval and consent to participate
Not applicable.
Consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing interests.
Publisher’s Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Contributor Information
Mario Gonzalo Garcia-Arraez, Phone: +41216931857, Email: mario.garciaarraez@epfl.ch.
Florent Masson, Email: florent.masson@epfl.ch.
Juan Camilo Paredes Escobar, Email: jparedes@icipe.org.
Bruno Lemaitre, Phone: +41 21 693 18 31, Email: bruno.lemaitre@epfl.ch.
References
- 1.Kikuchi Y. Endosymbiotic Bacteria in insects: their diversity and Culturability. Microbes Environ. 2009;24:195–204. doi: 10.1264/jsme2.ME09140S. [DOI] [PubMed] [Google Scholar]
- 2.Ishikawa H. Insect Symbiosis. An introduction. 2003. [Google Scholar]
- 3.Ferrari J, Vavre F. Bacterial symbionts in insects or the story of communities affecting communities. Philos Trans R Soc B. 2011;366:1389–1400. doi: 10.1098/rstb.2010.0226. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 4.Teixeira L, Ferreira Á, Ashburner M. The bacterial symbiont Wolbachia induces resistance to RNA viral infections in Drosophila melanogaster. PLoS Biol. 2008;6:e1000002. doi: 10.1371/journal.pbio.1000002. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 5.Vigneron A, Masson F, Vallier A, Balmand S, Rey M, Vincent-Monégat C, et al. Insects recycle endosymbionts when the benefit is over. Curr Biol. 2018;24:2267–2273. doi: 10.1016/j.cub.2014.07.065. [DOI] [PubMed] [Google Scholar]
- 6.Montllor CB, Maxmen A, Purcell AH. Facultative bacterial endosymbionts benefit pea aphids Acyrthosiphon pisum under heat stress. Ecol Entomol. 2002;27:189–195. doi: 10.1046/j.1365-2311.2002.00393.x. [DOI] [Google Scholar]
- 7.Dunbar HE, Wilson ACC, Ferguson NR, Moran NA. Aphid thermal tolerance is governed by a point mutation in bacterial symbionts. PLoS Biol. 2007;5:e96. doi: 10.1371/journal.pbio.0050096. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 8.Ciche TA, Darby C, Ehlers R-U, Forst S, Goodrich-Blair H. Dangerous liaisons: the symbiosis of entomopathogenic nematodes and bacteria. Biol Control. 2006;38:22–46. doi: 10.1016/j.biocontrol.2005.11.016. [DOI] [Google Scholar]
- 9.Loreto RG, Hughes DP. Genetics and molecular biology of Entomopathogenic Fungi. 2016. Chapter eight - disease dynamics in ants: a critical review of the ecological relevance of using generalist Fungi to study infections in insect societies; pp. 287–306. [DOI] [PubMed] [Google Scholar]
- 10.Hamilton PT, Perlman SJ. Host defense via Symbiosis in Drosophila. PLoS Pathog. 2013;9:e1003808. doi: 10.1371/journal.ppat.1003808. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 11.Mateos M, Castrezana SJ, Nankivell BJ, Estes AM, Markow TA, Moran NA. Heritable endosymbionts of Drosophila. Genetics. 2006;174:363–376. doi: 10.1534/genetics.106.058818. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 12.Counce SJ, Poulson DF. Developmental effects of the sex-ratio agent in embryos of Drosophila willistoni. J Exp Zool. 1962;151:17–31. doi: 10.1002/jez.1401510103. [DOI] [PubMed] [Google Scholar]
- 13.Anbutsu H, Fukatsu T. Spiroplasma as a model insect endosymbiont. Environ Microbiol Rep. 2011;3:144–153. doi: 10.1111/j.1758-2229.2010.00240.x. [DOI] [PubMed] [Google Scholar]
- 14.Herren JK, Paredes JC, Schüpfer F, Lemaitre B. Vertical transmission of a Drosophila endosymbiont via cooption of the yolk transport and internalization machinery. MBio. 2013;4:e00532. doi: 10.1128/mBio.00532-12. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 15.Lemaitre B, Nicolas E, Michaut L, Reichhart J-M, Hoffmann JA. The Dorsoventral regulatory gene cassette spätzle/toll/cactus controls the potent antifungal response in Drosophila adults. Cell. 1996;86:973–983. doi: 10.1016/S0092-8674(00)80172-5. [DOI] [PubMed] [Google Scholar]
- 16.Lemaitre B, Hoffmann J. The host defense of Drosophila melanogaster. Annu Rev Immunol. 2007;25:697–743. doi: 10.1146/annurev.immunol.25.022106.141615. [DOI] [PubMed] [Google Scholar]
- 17.Herren JK, Lemaitre B. Spiroplasma and host immunity: activation of humoral immune responses increases endosymbiont load and susceptibility to certain gram-negative bacterial pathogens in Drosophila melanogaster. Cell Microbiol. 2011;13:1385–1396. doi: 10.1111/j.1462-5822.2011.01627.x. [DOI] [PubMed] [Google Scholar]
- 18.Hurst GDD, Anbutsu H, Kutsukake M, Fukatsu T. Hidden from the host: Spiroplasma bacteria infecting Drosophila do not cause an immune response , but are suppressed by ectopic immune activation. Insect Mol Biol. 2003;12:93–97. doi: 10.1046/j.1365-2583.2003.00380.x. [DOI] [PubMed] [Google Scholar]
- 19.Herren JK, Paredes JC, Schüpfer F, Arafah K, Bulet P, Lemaitre B. Insect endosymbiont proliferation is limited by lipid availability. elife. 2014;3:e02964. doi: 10.7554/eLife.02964. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 20.Masson F, Copete Sandra C, Schüpfer F, Garcia-Arraez G, Lemaitre B. In vitro culture of the insect endosymbiont Spiroplasma poulsonii highlights bacterial genes involved in HostSymbiont interaction. MBio. 2018;9:1–11. doi: 10.1128/mBio.00024-18. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 21.Hurst GDD, Crystal L. Reproductive parasitism: maternally lnherited symbionts in a Biparental world. Cold Spring Harb Perspect Biol. 2015;7:1–20. doi: 10.1101/cshperspect.a017699. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 22.Oishi K. Spirochaete-mediated abnormal sex-ratio (SR) condition in Drosophila: a second virus associated with spirochaetes and its use in the study of the SR condition. Genet Res. 1971;18:45–56. doi: 10.1017/S0016672300012404. [DOI] [PubMed] [Google Scholar]
- 23.Harumoto T, Lemaitre B. Male-killing toxin in a bacterial symbiont of Drosophila. Nature. 2018;557:252–255. doi: 10.1038/s41586-018-0086-2. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 24.Martin J, Chong T, Ferree PM. Male killing Spiroplasma preferentially disrupts neural development in the Drosophila melanogaster embryo. PLoS One. 2013;8:1–8. doi: 10.1371/annotation/1c88ae20-07d1-4fda-8cd8-058d95af057d. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 25.Harumoto T, Anbutsu H, Lemaitre B, Fukatsu T. Male-killing symbiont damages host’s dosage-compensated sex chromosome to induce embryonic apoptosis. Nat Commun. 2016;7:12781. doi: 10.1038/ncomms12781. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 26.Cheng B, Kuppanda N, Aldrich JC, Akbari OS, Ferree PM. Male-killing Spiroplasma alters behavior of the dosage compensation complex during Drosophila melanogaster embryogenesis. Curr Biol. 2016;26:1339–1345. doi: 10.1016/j.cub.2016.03.050. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 27.Harumoto T, Anbutsu H, Fukatsu T. Male-killing Spiroplasma induces sex-specific cell death via host apoptotic pathway. PLoS Pathog. 2014;10:1–10. doi: 10.1371/journal.ppat.1003956. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 28.Paredes JC, Herren JK, Schüpfer F, Lemaitre B. The role of lipid competition for endosymbiont-mediated protection against parasitoid wasps in Drosophila. MBio. 2016;7:1–8. doi: 10.1128/mBio.01006-16. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 29.Xie J, Tiner B, Vilchez I, Mateos M. Effect of the Drosophila endosymbiont Spiroplasma on parasitoid wasp development and on the reproductive fitness of wasp-attacked fly survivors. Evol Ecol. 2011;53:1065–1079. doi: 10.1007/s10682-010-9453-7. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 30.Ballinger MJ, Perlman SJ. Generality of toxins in defensive symbiosis: ribosome-inactivating proteins and defense against parasitic wasps in Drosophila. PLoS Pathog. 2017;13:1–19. doi: 10.1371/journal.ppat.1006431. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 31.Hamilton PT, Peng F, Boulanger MJ, Perlman SJ. A ribosome-inactivating protein in a Drosophila defensive symbiont. Proc Natl Acad Sci U S A. 2016;113:350–355. doi: 10.1073/pnas.1518648113. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 32.Jaenike J, Unckless R, Cockburn SN, Boelio LM, Perlman SJ. Adaptation via Symbiosis: recent spread of a Drosophila defensive symbiont. Science. 2010;329:212–215. doi: 10.1126/science.1188235. [DOI] [PubMed] [Google Scholar]
- 33.Endo Y, Tsurugi K. Mechanism of action of ricin and related toxic lectins on eukaryotic ribosomes. Nucleic Acids Symp Ser. 1986;262:187–190. [PubMed] [Google Scholar]
- 34.Virgilio M, Lombardi A, Caliandro R, Fabbrini Maria S. Ribosome-inactivating proteins: from plant defense to tumor attack. Toxins. 2010;2:2700–2737. doi: 10.3390/toxins2112699. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 35.Lainhart W, Stolfa G, Koudelka GB. Shiga toxin as a bacterial defense against a eukaryotic predator, Tetrahymena thermophila. J Bacteriol. 2009;191:5116–5122. doi: 10.1128/JB.00508-09. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 36.Bergan J, Dyve L, Anne B, Simm R, Skotland T, Sandvig K. Shiga toxins. Toxicon. 2012;60:1085–1107. doi: 10.1016/j.toxicon.2012.07.016. [DOI] [PubMed] [Google Scholar]
- 37.Szewczak AA, Moore PB. The Sarcin/ricin loop, a modular RNA. J Mol Biol. 1995;247:81–98. doi: 10.1006/jmbi.1994.0124. [DOI] [PubMed] [Google Scholar]
- 38.Paredes JC, Herren JK, Schüpfer F, Marin R, Claverol S, Kuo C-H, et al. Genome sequence of the Drosophila melanogaster male-killing Spiroplasma strain MSRO endosymbiont. MBio. 2015;6:1–12. doi: 10.1128/mBio.02437-14. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 39.Le Page DP, Metcalf JA, Bordenstein SR, On J, Perlmutter JI, Shropshire JD, et al. Prophage WO genes recapitulate and enhance Wolbachia-induced cytoplasmic incompatibility. Nature. 2017;543:243–247. doi: 10.1038/nature21391. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 40.Duffy JB. GAL4 system indrosophila: a fly geneticist’s swiss army knife. Genesis. 2002;34:1–15. doi: 10.1002/gene.10150. [DOI] [PubMed] [Google Scholar]
- 41.Soejima Y, Lee, Jae M, Nagata Y, Mon H, Iiyama K, Kitano H, et al. Comparison of signal peptides for efficient protein secretion in the baculovirus-silkworm system. Cent Eur J Biol. 2013;8:1–7. [Google Scholar]
- 42.Robert BK, Subinay G, Monica A, Sandra G, Allan S, Carol S, et al. Heavy chain dimers as well as complete antibodies are efficiently formed and secreted from Drosophila via a BiP-mediated pathway. J Biol Chem. 1998;273:20533–20538. doi: 10.1074/jbc.273.46.30372. [DOI] [PubMed] [Google Scholar]
- 43.Iyer J, Wang Q, Le T, Pizzo L, Grönke S, Ambegaokar SS, et al. Quantitative assessment of eye phenotypes for functional genetic studies using Drosophila melanogaster. G3: genes|genomes|. Genetics. 2016;6:1427–1437. doi: 10.1534/g3.116.027060. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 44.Halder G, Callaerts P, Flister S, Walldorf U, Kloter U, Gehring WJ. Eyeless initiates the expression of both sine oculis and eyes absent during Drosophila compound eye development. Development. 1998;125:2181–2191. doi: 10.1242/dev.125.12.2181. [DOI] [PubMed] [Google Scholar]
- 45.Van-Vactor DL, Cagan RL, Krämer H, Zipursky SL. Induction in the developing compound eye of Drosophila: multiple mechanisms restrict R7 induction to a single retinal precursor cell. Cell. 1991;67:1145–1155. doi: 10.1016/0092-8674(91)90291-6. [DOI] [PubMed] [Google Scholar]
- 46.Ramond E, Meister M, Lemaitre B. From embryo to adult: hematopoiesis along the Drosophila life cycle. Dev Cell. 2015;33:367–368. doi: 10.1016/j.devcel.2015.05.002. [DOI] [PubMed] [Google Scholar]
- 47.Horn L, Leips J, Starz-Gaiano M. Phagocytic ability declines with age in adult Drosophila hemocytes. Aging Cell. 2014;13:719–728. doi: 10.1111/acel.12227. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 48.Ashburner M. Drosophila: a laboratory handbook and manual. 1989. [Google Scholar]
- 49.Bretscher AJ, Honti V, Binggeli O, Burri O, Poidevin M, Kurucz É, et al. The nimrod transmembrane receptor eater is required for hemocyte attachment to the sessile compartment in Drosophila melanogaster. Biology Open. 2015;4:355–363. doi: 10.1242/bio.201410595. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 50.Livak KJ, Schmittgen TD. Analysis of relative gene expression data using real-time quantitative PCR and the 2−ΔΔCT method. Methods. 2001;25:402–408. doi: 10.1006/meth.2001.1262. [DOI] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.
Supplementary Materials
Figure S1. SpRIPs 3,4 and 5 expression level in infected flies along Drosophila development stages (One way ANOVA; development stage p = 0.9992). (TIF 1384 kb)
Figure S2. Lifespan of infected and uninfected wild types flies (controls for Fig. 3). Sp- and Sp + refer to uninfected or Spiroplasma-infected condition respectively. (TIF 1983 kb)
Figure S3. Expression level of (A) SpRIP1 and (B) SpRIP2 in embryos from uninfected, infected and transgenic fly lines expressing UAS-RIP normalized by host rsp17 transcript level (One way ANOVA; p** = 0.0031 for SpRIP1 and p** = 0.0049 for SpRIP2). (C) Expression level of SpRIP2 in adults from uninfected, infected and transgenic fly lines expressing UAS-RIP normalized by host rsp17 transcript level (One way ANOVA; p** < 0.0081). (TIF 5534 kb)
Figure S4. (A) Comparison of RIP activity in Spiroplasma-infected embryos with uninfected transgenic embryos expressing SpRIPS (One way ANOVA; SpRIP1 p*** = 0.001; One way ANOVA; SpRIP2 p** = 0.0021). (B) Intact 28S rRNA quantification in infected embryos and uninfected transgenic fly lines (C) Comparison of RIP activity in Spiroplasma-infected adult flies with uninfected transgenic adult fly (One way ANOVA; SpRIP2 p*** = 0.001) (D) Intact 28S rRNA quantification in infected adult flies and uninfected transgenic adult flies. (TIF 6378 kb)
Figure S5. hml transcription level in lines expressing UAS-SpRIP2 and BiP + SpRIP2 under actin-GAL4 control. Expression of both constructs also leads to a decrease in hemocyte number (Dunnett’s multiple comparisons test p*** < 0.0001). Controls are normalized as 1. (TIF 1147 kb)
Figure S6. Correlation between RIP activity in embryos aged 0 to 24 h after egg laying and embryo mortality (Pearson’s test p < 0.0001). RIP activity in infected wild types was normalized by uninfected samples. Transgenic fly lines were normalized by Da-GAL4/w1118. Controls are normalized as 1. (TIF 3499 kb)
Figure S7. Sex ratio of the control fly lines for Fig. 5. Sp- and Sp + refer to uninfected or Spiroplasma-infected condition respectively. (TIF 4019 kb)
Data Availability Statement
All data generated or analyzed during this study are included in this published article (and its additional files).