

Abstract
The present study aimed to investigate the biological function and underlying molecular mechanisms of miR-31 in osteoarthritis (OA). Reverse transcription-quantitative polymerase chain reaction was used to detect miR-31 expression, and it was found that miR-31 was downregulated in the cartilage tissues of OA patients. microRNA.org was used to predict the gene targets of miR-31, and dual luciferase reporter assays were used to verify that C-X-C motif chemokine ligand 12 (CXCL12) was a direct target of miR-31. The human chondrocyte cell line CHON-001 was used to perform MTT and cell migration assays. Western blotting was used to measure the protein expression of CXCL12, type I collagen and aggrecan. The results suggested that CXCL12 was a target of miR-31, and the expression of CXCL12 was negatively regulated by miR-31 in CHON-001 cells. miR-31 increased CHON-001 cell viability and migration, as well as the expression of type I collagen and aggrecan. Furthermore, the overexpression of CXCL12 eliminated the effects of miR-31 mimics on CHON-001 cells. In conclusion, the data indicated that miR-31 promoted chondrocyte viability and migration by directly targeting CXCL12, which provided evidence for CXCL12 as a potential target in OA therapy.
Keywords: microRNA-31, C-X-C motif chemokine ligand 12, osteoarthritis, proliferation, migration
Introduction
There is a poor intrinsic capacity for repair in articular cartilage, and even a minor defect as a result of mechanical damage may fail to heal, thus degenerating to osteoarthritis (OA) (1). OA, one of the most common degenerative joint disorders worldwide, is characterized by slow destruction and loss of articular cartilage, mainly affecting the hips and knees (2). In China specifically, the morbidity rate of symptomatic knee OA is as high as 8.1% (3). Unfortunately, the precise pathogenesis of OA remains unclear.
C-X-C motif chemokine ligand 12 (CXCL12), also termed stromal cell-derived factor 1, is a typical bone marrow-derived chemokine which promotes stem cell homing to injured areas (4,5) and functions in tissue and/or organ repair (4,6) via binding to C-X-C chemokine receptor type 4 (CXCR4), a G protein-coupled receptor. CXCL12 and CXCR4 binding promotes the early osteogenic differentiation of mesenchymal stem cells by regulating the bone morphogenetic protein (BMP)-2/mothers against decapentaplegic homolog (Smad)/Runt-related transcription factor 2 (Runx2)/Osterix axis (7). In addition, CXCL12 from bone marrow-derived mesenchymal stromal cells has been shown to improve muscle regeneration (8). On the other hand, it has been reported that CXCL12 is involved in the destruction of cartilage in OA (9). The present study investigated whether CXCL12 was involved in OA development in chondrocytes.
MicroRNAs (miRNAs/miRs), a group of non-coding small RNAs (~22 nucleotides), are widely expressed in eukaryons and serve important roles in numerous physiological and pathological processes. In OA, a number of miRNAs are aberrantly expressed in tissues, such as miR-33a (3) and miR-155 (4). However the role of miR-31 in OA remains to be elucidated, and was therefore investigated in the present study.
Materials and methods
Clinical specimens
Articular cartilage tissues were collected from 30 OA patients (male, 21; female, 9; age range 52–65 years) undergoing total knee replacement surgery and 30 non-OA patients (male, 19; female, 11; age range 54–67 years) with femoral neck fracture undergoing surgery at the Second People's Hospital of Huai'an (Huai'an, China) between August 2015 and August 2016. The non-OA patients had no known history of OA or rheumatoid arthritis. All patients provided written informed consent and the present study was approved by the ethics committee of the Second People's Hospital of Huai'an.
Cell culture
Human chondrocyte cell line CHON-001 was obtained from American Type Culture Collection (cat no. CRL-2846; Manassas, VA, USA) and incubated in Dulbecco's modified Eagle's medium/nutrient mixture F12 (DMEM/F12; Gibco; Thermo Fisher Scientific, Inc., Waltham, MA, USA) with 10% fetal bovine serum (FBS; Gibco; Thermo Fisher Scientific, Inc.), 1% penicillin-streptomycin solution and 25 µg/ml amphotericin B (Fungizone; Gibco; Thermo Fisher Scientific, Inc.) in an incubator at 37°C with 5% CO2.
Cell transfection
For cell transfection, 5×104 CHON-001 cells/well were seeded into a 24-well plate. miRNA mimic and inhibitor were obtained from Shanghai GenePharma Co., Ltd., (Shanghai, China). The next day, cells were transfected with 50 nM miR-31 mimic (forward: 5′AGGCAAGAUGCUGGCAUAGCU3′, reverse: 5′CUAUGCCAGCAUCUUGCCUUU3′), 50 nM miR-31 inhibitor (5′AGCUAUGCCAGCAUCUUGCCU3′), miR-control (forward: 5′UUCUCCGAACGUGUCACGUTT3′, reverse: 5′ACGUGACACGUUCGGAGAATT3′), 2 µl control plasmid (cat. no. sc-108083; Santa Cruz Biotechnology, Inc., Santa Cruz, CA, USA), 2 µl CXCL12-plasmid (cat. no. sc-422854-ACT; Santa Cruz Biotechnology, Inc.), miR-31 mimic + control plasmid or miR-31 mimic+CXCL12-plasmid using Lipofectamine® 3000 (Thermo Fisher Scientific, Inc.). After 48 h, the cells were harvested for subsequent experimentation.
Reverse transcription-quantitative polymerase chain reaction (RT-qPCR)
Total RNA was extracted by TRIzol® (Invitrogen; Thermo Fisher Scientific, Inc.). RNA was reverse transcribed into cDNA using a Reverse Transcription system (Promega Corporation, Madison, WI, USA). The qPCR reaction mixture was as follows: SYBR Premix Ex Taq™ II (Bio-Rad Laboratories, Inc., Hercules, CA, USA), 2 µl cDNA, 5 µl 2X master mix, 0.5 µl forward/reverse primer and 2 µl nanopure water. qPCR was performed on an ABI 7900HT machine (Applied Biosystems; Thermo Fisher Scientific, Inc.). Amplification conditions were as follows: 95°C for 5 min, followed by 40 cycles of 95°C for 15 sec and 60°C for 1 min. The primer sequences were obtained from Jinsite Science and Technology (Nanjing) Co., Ltd. (Nanjing, China) and listed in Table I. Data were calculated by 2−ΔΔCq method (10).
Table I.
Primer sequences used for quantitative polymerase chain reaction analysis.
| Gene | Direction | Sequence (5′-3′) |
|---|---|---|
| C-X-C motif chemokine ligand 12 | F | ATGCCCATGCCGATTCTT |
| R | GCCGGGCTACAATCTGAAGG | |
| Type I collagen | F | CCCTGAGTGGAAGAGTGGAG |
| R | GAGGCGTGAGGTCTTCTGTG | |
| Aggrecan | F | CTAGAGATCAGTGGACTGCCT |
| R | TCTGGAGCTGTGCAGTCTAGTGG | |
| microRNA-31 | F | CTCGGATCCTGTGCATAACTGCCTTCA |
| R | CACAAGCTTGAAGTCAGGGCGAGACAGAC | |
| U6 | F | Universal_RNU6B_Primer |
| R | Uni-miR qPCR Primer | |
| GAPDH | F | CTTTGGTATCGTGGAAGGACTC |
| R | GTAGAGGCAGGGATGATGTTCT |
F, forward; R, reverse.
Western blot analysis
CHON-001 cell lysates were washed with PBS and homogenized in radioimmunoprecipitation assay buffer (Roche Diagnostics, Basel, Switzerland). Proteins (30 µg per lane) were separated by 10% SDS-PAGE and transferred to polyvinylidene fluoride membranes, which were blocked with 5% milk in Tris buffered saline with 0.1% Tween-20 for 2 h at room temperature. Next, they were incubated with primary antibodies against β-actin (cat no. 4970; 1:1,000; Cell Signaling Technology, Inc., Danvers, MA, USA), CXCL12 (cat no. 3530; 1:1,000; Cell Signaling Technology, Inc.), type I collagen (cat no. ab34710; 1:1,000; Abcam) and aggrecan (cat no. ab36861; 1:1,000; Abcam) at 4°C overnight. Membranes were subsequently incubated with horseradish peroxidase-conjugated anti-rabbit IgG secondary antibody (cat no. 7074; 1:5,000; Cell Signaling Technology, Inc.) for 1 h at room temperature. Bands were visualized by SuperSignal® West Pico Chemiluminescent Substrate (Thermo Fisher Scientific, Inc.). Band density was normalized to β-actin.
MTT assay
Cell viability was assessed by MTT assays at 24, 48 and 72 h after cell transfection. Cells (5×103 cells/well) were seeded in 96-well plates and incubated for 24 h at 37°C. Next, MTT (20 µl; 5 mg/ml) was added to each well and incubated for another 4 h. Wells were then washed in PBS, dried and 150 µl dimethyl sulfoxide was added. Data were measured by a microtiter plate reader at 490 nm (Thermo Fisher Scientific, Inc.).
Cell migration assay
The migratory ability of CHON-001 cells was detected using 24-well Transwell chambers. Cells (2×104/well) were plated into the upper chambers and incubated in 200 µl serum-free DMEM (Gibco; Thermo Fisher Scientific, Inc.). The bottom chamber contained DMEM (0.5 ml) with 20% FBS (Gibco; Thermo Fisher Scientific, Inc.). Following a 48 h incubation, cells on the upper chambers were removed, and the migrated cells on the lower chambers were fixed with 10% formalin for 30 min at room temperature and stained with hematoxylin and eosin (H&E) for 15 min at 37°C. Cell migratory ability was assessed by counting the number of H&E-stained cells of five fields of view under a light microscope (Olympus Corporation, Tokyo, Japan).
Bioinformatic analysis
Prediction of miRNA target sites was performed using microRNA.org (www.microrna.org).
Luciferase reporter assays
A luciferase reporter (Promega Corporation) for the wild-type (WT) CXCL12 3′untranslated region (UTR) and mutant (MUT) CXCL12 3′UTR was constructed by site-directed mutagenesis with Phusion™ High-Fidelity DNA Polymerase (Thermo Fisher Scientific, Inc.). CHON-001 cells (5×104) were seeded into each well of a 24-well plate. The next day, cells were co-transfected with 50 nM CXCL12-3′UTR-WT or CXCL12-3′UTR-MUT and miR-31 mimics or control using Lipofectamine® 3000 (Thermo Fisher Scientific, Inc.). After 72 h, cells were harvested to determine luciferase activity with the Dual-Luciferase Reporter Assay system (Beyotime Institute of Biotechnology, Haimen, China) and a luminometer. Firefly luciferase activity was normalized to Renilla.
Statistical analysis
Data were analyzed by Student's t-test or one-way analysis of variance followed by Tukey's test, using SPSS version 21 (IBM Corp., Armonk, NY, USA). Data were expressed as the mean ± standard deviation. P<0.05 was considered to indicate a statistically significant difference.
Results
miR-31 is downregulated in OA patients
The expression of miR-31 in the articular cartilage tissues of patients with and without OA was determined by RT-qPCR. It was found that that miR-31 expression was significantly decreased in OA patients, compared with the non-OA patients (Fig. 1).
Figure 1.
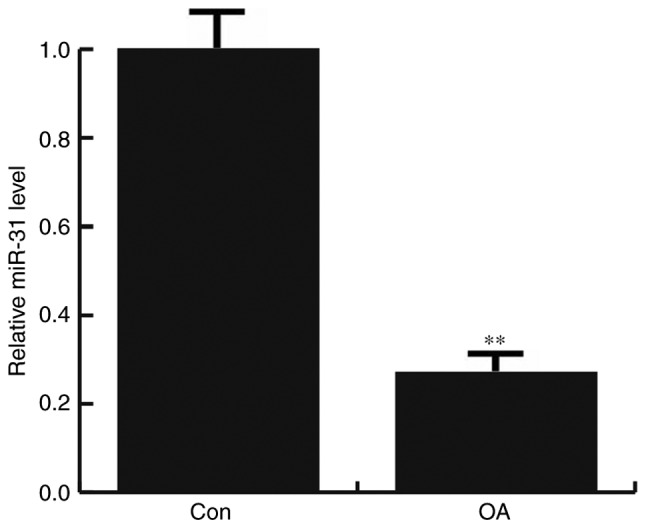
Relative expression of miR-31 in patients with OA. Expression was detected in 30 paired tissue samples by reverse transcription-quantitative polymerase chain reaction. **P<0.01 vs. Con. miR, microRNA; OA, osteoarthritis; Con, control.
CXCL12 is targeted by miR-31
To investigate the role of miR-31 in OA progression, bioinformatics software (www.microrna.org) was used to predict the potential targets of miR-31. A total of 6,829 targets were identified, including CXCL12 (Fig. 2A). As it has been previously reported that CXCL12 has critical function in the development of OA (11,12), this gene was selected for further analysis.
Figure 2.

CXCL12 is a direct target of miR-31. (A) microRNA.org was used to predict the interaction between miR-31 and the 3′UTR of CXCL12. (B) Luciferase activity was detected by dual luciferase reporter assays. (C) The protein expression of CXCL12 was measured in OA (OA1 and OA2) and control patients (Con1 and Con2) by western blotting. (D) The relative expression of CXCL12 in OA patients and control individuals was determined by reverse transcription-quantitative polymerase chain reaction. **P<0.01 vs. Con. CXCL12, C-X-C motif chemokine ligand 12; miR, microRNA; UTR, untranslated region; OA, osteoarthritis; Con, control.
Next, the correlation between miR-31 and CXCL12 was verified by dual-luciferase reporter assays, which showed that the luciferase activity of the CXCL12-WT group significantly declined following miR-31 mimic transfection compared with the miR-control group, with no evident alteration in the CXCL12-MUT groups (Fig. 2B).
In addition, the expression of CXCL12 in the articular cartilage tissues of patients was determined by RT-qPCR and western blotting. It was demonstrated that CXCL12 protein (Fig. 2C) and mRNA (Fig. 2D) expression was significantly increased in OA patients compared with non-OA patients.
miR-31 inhibits CXCL12 expression in CHON-001 cells
To further investigated whether miR-31 regulated CXCL12 expression in CHON-001 cells, they were transfected with miR-31 mimics, inhibitor or control mimics. Transfection efficiency was determined 48 h after transfection by RT-qPCR (Fig. 3A). It was revealed that CXCL12 expression was markedly decreased by miR-31 mimic transfection, and increased by miR-31 inhibitor transfection, compared with the control group (Fig. 3B and C).
Figure 3.

Expression of CXCL12 and miR-31 in CHON-001 cells. Expression was detected by reverse transcription-quantitative polymerase chain reaction and western blotting 48 h after transfection. (A) Relative mRNA expression level of miR-31 and (B) CXCL 12. (C) Protein expression of CXCL12. *P<0.05, **P<0.01 vs. Con. CXCL12, C-X-C motif chemokine ligand 12; miR, microRNA; Con, control.
miR-31 increases CHON-001 cell viability and migration
Compared with the control group, CHON-001 cells transfected with miR-31 mimics exhibited a significant increase in cell viability, while miR-31 inhibitor transection results in a significant decrease in cell viability (Fig. 4A). In addition, it was found that the number of migrated cells in the miR-31 mimic group was increased compared with the control group, whereas miR-31 inhibitor transfection significantly prevented CHON-001 cell migration (Fig. 4B and C).
Figure 4.

Effects of miR-31 on CHON-001 cell viability and migration. CHON-001 cell viability and migration was determined by MTT and Transwell migration, respectively, assays 48 h after transfection. (A) CHON-001 cell viability. (B) Migrated CHON-001 cell number and (C) representative images of migrated cells in each group. *P<0.05, **P<0.01 vs. Con. miR, microRNA; Con, control; OD, optical density.
miR-31 promotes the expression of type I collagen and aggrecan
During the progression of OA, the collagen matrix undergoes a transitional degradation (13,14). Therefore, the expression of type I collagen and aggrecan was detected in the present study. The findings indicated that the protein and mRNA expression of type I collagen and aggrecan was increased by miR-31 mimic transfection, and miR-31 inhibitor had the opposite effect in CHON-001 cells (Fig. 5).
Figure 5.
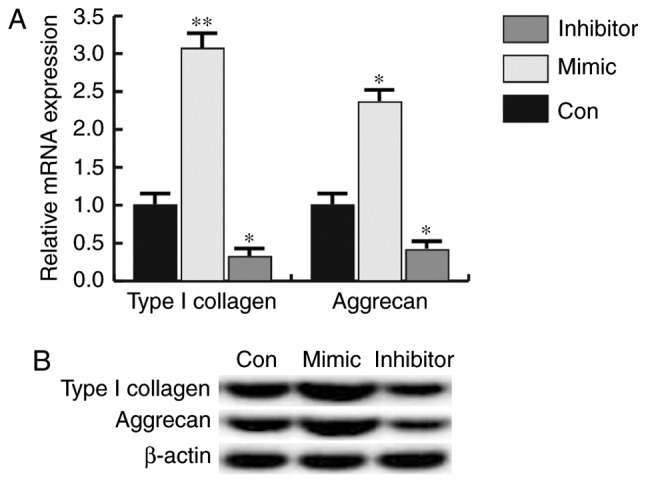
Effects of microRNA-31 on type I collagen and aggrecan expression. Expression was detected by reverse transcription-quantitative polymerase chain reaction and western blotting. (A) Relative mRNA and (B) protein expression level of type I collagen and aggrecan. *P<0.05, **P<0.01 vs. Con. Con, control.
CXCL12 overexpression prevents the alterations induced by miR-31
To examine whether miR-31 affected CHON-001 cells by directly targeting CXCL12, control or CXCL12-plasmid was initially transfected into CHON-001 cells to assess transfection efficiency by western blotting (Fig. 6A) and RT-qPCR (Fig. 6B). Following this, miR-31 mimics + control plasmid or miR-31 mimics+CXCL12-plasmid was transfected into CHON-001 cells. The protein and mRNA expression of CXCL12, type I collagen and aggrecan was detected by western blot and RT-PCR, 48 h after transfection. It was found that the protein (Fig. 6C) and mRNA expression (Fig. 6D and E) of type I collagen and aggrecan was notably enhanced by miR-31 mimics, whereas CXCL12 overexpression eliminated these effects.
Figure 6.

Effects of CXCL12 plasmid on CXCL12, type I collagen and aggrecan expression. The protein and mRNA expression level of CXCL12, type I collagen and aggrecan in each group was determined 48 h after transfection by western blot analysis and reverse transcription-quantitative polymerase chain reaction, respectively. Transfection efficiency of CXCL12 was confirmed at the (A) protein and (B) mRNA level. (C) CXCL12, type I collagen and aggrecan protein and (D and E) mRNA expression in cells transfected with mimics+control plasmid or mimics+CXCL12 plasmid. *P<0.05, **P<0.01 vs. control; #P<0.05, ##P<0.01 vs. m+plasmid-c Con, control; plasmid-c, control-plasmid; CXCL-p, CXCL 12-plasmid; m, microRNA-31 mimics; CXCL12, C-X-C motif chemokine ligand 12.
Furthermore, compared with the cells transfected with miR-31 mimics + control plasmid, cell viability (Fig. 7A) and migration (Fig. 7B and C) significantly decreased in the miR-31 mimic+CXCL12-plasmid transfected cells, compared with the mimics + control plasmid transfected cells (Fig. 7). Taken together, these data indicated that miR-31 promoted CHON-001 cell proliferation and migration, as well as type I collagen and aggrecan expression by directly targeting CXCL12.
Figure 7.

Effects of CXCL 12 plasmid on CHON-001 cell viability and migration. Cell viability and migration was determined by MTT and Transwell migration assays, respectively. (A) Effects of CXCL12 plasmid on CHON-001 cell viability. (B) Migrated CHON-001 cell number and (C) representative images of migrated cells in each group. *P<0.05, **P<0.01 vs. control; #P<0.05, ##P<0.01 vs. m+plasmid-c. Con, control; plasmid-c, control-plasmid; CXCL-p, CXCL 12-plasmid; m, microRNA-31 mimics; CXCL12, C-X-C motif chemokine ligand 12.
Discussion
In the current study, it was demonstrated that miR-31 was downregulated in patients with OA, compared with controls. miR-31 promoted human chondrocyte CHON-001 cell viability and migration, as well as and the expression of type I collagen and aggrecan, which provided evidence for miR-31 as a potential target in cartilage defection therapy.
OA is characterized by slow destruction and loss of articular cartilage, which predominantly affects the hips and knees (2). Unfortunately, the precise pathogenesis of OA remains unclear. miRNAs have received increasing attention for their contribution to signaling network disruption in OA (15). Many specific miRNAs have been identified to be critical during OA pathogenesis (16). For example, miR-384-5p downregulation alleviates OA through inhibiting cartilage cell apoptosis via the NF-κB signaling pathway by targeting SOX9 (17). MiR-34a has been identified to facilitate the development of OA by enhancing chondrocyte apoptosis and senescence (18). MiR-449a upregulation was found to promote chondrocyte extracellular matrix degradation in OA (19). It has been reported that downregulation of miR-31 enhances the osteogenic differentiation of human mesenchymal stem cells by targeting SATB2 (20). miR-31-5p expression is reduced in cartilage-derived mesenchymal stem cells from OA degraded cartilage compared to normal cartilage, and may regulate osteogenic differentiation (5). During bone mesenchymal stem cell osteogenic differentiation, miR-31 expression is markedly decreased, and inhibition of miR-31 enhances osteogenic differentiation (21). Furthermore, miR-31 has been reported to enhance cell proliferation during differentiation by targeting CCAAT enhancer binding protein α (C/EBP-α) (22). However, to the best of our knowledge, the role of miR-31 in the development of OA remained largely unclear. In the present study, it was found that miR-31 was significantly downregulated in OA patients. In addition, the 3′UTR of CXCL12 was targeted by miR-31, which was predicted via microRNA.org and verified with dual-luciferase reporter assays. Additionally, it was indicated that miR-31 negatively regulated CXCL12 expression in human chondrocyte cells. CXCL12 has been revealed to take part in tissues and organ repair (4,6). Inhibition of the CXCL12/CXCR4 signaling axis prevents aggrecanase expression and reduces cartilage degeneration (23,24). Furthermore, CXCL12 expression in the plasma and synovial fluid of OA patients may serve as an effective biomarker for the severity of OA (25).
CHON-001 cells were subsequently transfected with miR-31 mimics, miR-31 inhibitor or mimic control, and cell viability was detected by MTT assays 24, 48 and 72 h post-transfection. CHON-001 cell migration assays were performed and the expression of type I collagen and aggrecan was also determined. The results suggested that miR-31 enhanced the cell viability and migration of CHON-001 cells, and the expression of type I collagen and aggrecan was increased. Furthermore, to explore whether miR-31 affected CHON-001 cells by directly targeting CXCL12, cells were transfected with miR-31 mimics + control plasmid or miR-31 mimics+CXCL12-plasmid, and cell viability, migration, and type I collagen and aggrecan expression were subsequently determined. The findings revealed that the effects of miR-31 on CHON-001 cell viability and migration, as well as type I collagen and aggrecan expression were eliminated by CXCL12 overexpression.
Taken together, the results indicated that miR-31 was down-regulated in OA, and miR-31 promoted the growth and migration of chondrocytes. The data suggested a crucial role of miR-31 in OA, suggesting that miR-31 may serve as a potential target for OA treatment. However, whether there is a correlation between the expression of miR-31 and its target gene CXCL12 in OA patients, as well as the clinical significance of miR-21 has not been elucidated. Furthermore, the specific mechanisms by which miR-31 functions in OA development remain unclear. Therefore, the authors of the present study will perform more in-depth study on the role of miR-31 in OA in the near future.
Acknowledgements
The authors thank the Life Science College of Huaiyin Normal University for providing technical guidance in the experiments, and the reviewers for their invaluable help in improving the manuscript.
Funding
No funding was received.
Availability of data and materials
The datasets used and/or analyzed during the current study are available from the corresponding author on reasonable request.
Authors' contributions
YD and SL contributed to the concept and design of the study; YD, SL, XX and MD accessed and analyzed the data; QZ and XZ contributed to data analysis and prepared the paper. All authors collaborated to interpret the results and develop the manuscript.
Ethics approval and consent to participate
All patients provided written informed consent and the present study was approved by the ethics committee of the Second People's Hospital of Huai'an.
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing interests.
References
- 1.Redman SN, Oldfield SF, Archer CW. Current strategies for articular cartilage repair. Eur Cell Mater. 2005;9:23–32. doi: 10.22203/eCM.v009a04. [DOI] [PubMed] [Google Scholar]
- 2.Tsezou A. Osteoarthritis year in review 2014: Genetics and genomics. Osteoarthritis Cartilage. 2014;22:2017–2024. doi: 10.1016/j.joca.2014.07.024. [DOI] [PubMed] [Google Scholar]
- 3.Tang X, Wang S, Zhang Y, Niu J, Tao K, Zhang Y, Lin J. The prevalence of symptomatic knee osteoarthritis in China: Results from China health and retirement longitudinal study. Arthritis Rheumatol. 2016;68:648–653. doi: 10.1002/art.39465. [DOI] [PubMed] [Google Scholar]
- 4.Dowthwaite GP, Bishop JC, Redman SN, Khan IM, Rooney P, Evans DJ, Haughton L, Bayram Z, Boyer S, Thomson B, et al. The surface of articular cartilage contains a progenitor cell population. J Cell Sci. 2004;117:889–897. doi: 10.1242/jcs.00912. [DOI] [PubMed] [Google Scholar]
- 5.Xian CJ, Foster BK. Repair of injured articular and growth plate cartilage using mesenchymal stem cells and chondrogenic gene therapy. Curr Stem Cell Res Ther. 2006;1:213–229. doi: 10.2174/157488806776956904. [DOI] [PubMed] [Google Scholar]
- 6.Ji YH, Ji JL, Sun FY, Zeng YY, He XH, Zhao JX, Yu Y, Yu SH, Wu W. Quantitative proteomics analysis of chondrogenic differentiation of C3H10T1/2 mesenchymal stem cells by iTRAQ labeling coupled with on-line two-dimensional LC/MS/MS. Mol Cell Proteomics. 2010;9:550–564. doi: 10.1074/mcp.M900243-MCP200. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 7.Fox JM, Chamberlain G, Ashton BA, Middleton J. Recent advances into the understanding of mesenchymal stem cell trafficking. Br J Haematol. 2007;137:491–502. doi: 10.1111/j.1365-2141.2007.06610.x. [DOI] [PubMed] [Google Scholar]
- 8.Marquez-Curtis LA, Janowska-Wieczorek A. Enhancing the migration ability of mesenchymal stromal cells by targeting the SDF-1/CXCR4 axis. Biomed Res Int 2013. 2013:561098. doi: 10.1155/2013/561098. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 9.Li Q, Guo Y, Chen F, Liu J, Jin P. Stromal cell derived factor-1 promotes human adipose tissue-derived stem cell survival and chronic wound healing. Exp Ther Med. 2016;12:45–50. doi: 10.3892/etm.2016.3309. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 10.Livak KJ, Schmittgen TD. Analysis of relative gene expression data using real-time quantitative PCR and the 2(-Delta Delta C(T)) method. Methods. 2001;25:402–408. doi: 10.1006/meth.2001.1262. [DOI] [PubMed] [Google Scholar]
- 11.Wei F, Moore DC, Wei L, Li Y, Zhang G, Wei X, Lee JK, Chen Q. Attenuation of osteoarthritis via blockade of the SDF-1/CXCR4 signaling pathway. Arthritis Res Ther. 2012;14:R177. doi: 10.1186/ar3930. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 12.Lisignoli G, Toneguzzi S, Piacentini A, Cristino S, Grassi F, Cavallo C, Facchini A. CXCL12 (SDF-1) and CXCL13 (BCA-1) chemokines significantly induce proliferation and collagen type I expression in osteoblasts from osteoarthritis patients. J Cell Physiol. 2006;206:78–85. doi: 10.1002/jcp.20435. [DOI] [PubMed] [Google Scholar]
- 13.Maroudas AI. Balance between swelling pressure and collagen tension in normal and degenerate cartilage. Nature. 1976;260:808–809. doi: 10.1038/260808a0. [DOI] [PubMed] [Google Scholar]
- 14.Chou MC, Tsai PH, Huang GS, Lee HS, Lee CH, Lin MH, Lin CY, Chung HW. Correlation between the MR T2 value at 4.7 T and relative water content in articular cartilage in experimental osteoarthritis induced by ACL transection. Osteoarthritis Cartilage. 2009;17:441–447. doi: 10.1016/j.joca.2008.09.009. [DOI] [PubMed] [Google Scholar]
- 15.Iliopoulos D, Malizos KN, Oikonomou P, Tsezou A. Integrative microRNA and proteomic approsteoarthritisches identify novel osteoarthritis genes and their collaborative metabolic and inflammatory networks. PLoS One. 2008;3:e3740. doi: 10.1371/journal.pone.0003740. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 16.Sumiyoshi K, Kubota S, Ohgawara T, Kawata K, Nishida T, Shimo T, Yamashiro T, Takigawa M. Identification of miR-1 as a micro RNA that supports late-stage differentiation of growth cartilage cells. Biochem Biophys Res Commun. 2010;402:286–290. doi: 10.1016/j.bbrc.2010.10.016. [DOI] [PubMed] [Google Scholar]
- 17.Zhang W, Cheng P, Hu W, Yin W, Guo F, Chen A, Huang H. Inhibition of microRNA-384-5p alleviates osteoarthritis through its effects on inhibiting apoptosis of cartilage cells via the NF-κB signaling pathway by targeting SOX9. Cancer Gene Ther. 2018;25:326–338. doi: 10.1038/s41417-018-0029-y. [DOI] [PubMed] [Google Scholar]
- 18.Zhang W, Hsu P, Zhong B, Guo S, Zhang C, Wang Y, Luo C, Zhan Y, Zhang C. MiR-34a enhances chondrocyte apoptosis, senescence and facilitates development of osteoarthritis by targeting DLL1 and regulating PI3K/AKT pathway. Cell Physiol Biochem. 2018;48:1304–1316. doi: 10.1159/000492090. [DOI] [PubMed] [Google Scholar]
- 19.Wu J, Zou M, Ping A, Deng Z, Cai L. MicroRNA-449a upregulation promotes chondrocyte extracellular matrix degradation in osteoarthritis. Biomed Pharmacother. 2018;105:940–946. doi: 10.1016/j.biopha.2018.06.074. [DOI] [PubMed] [Google Scholar]
- 20.Xie Q, Wang Z, Bi X, Zhou H, Wang Y, Gu P, Fan X. Effects of miR-31 on the osteogenesis of human mesenchymal stem cells. Biochem Biophys Res Commun. 2014;446:98–104. doi: 10.1016/j.bbrc.2014.02.058. [DOI] [PubMed] [Google Scholar]
- 21.Deng Y, Wu S, Zhou H, Bi X, Wang Y, Hu Y, Gu P, Fan X. Effects of a miR-31, Runx2, and Satb2 regulatory loop on the osteogenic differentiation of bone mesenchymal stem cells. Stem Cells Dev. 2013;22:2278–2286. doi: 10.1089/scd.2012.0686. [DOI] [PubMed] [Google Scholar]
- 22.Liu Y, Wang Y, He X, Zhang S, Wang K, Wu H, Chen L. LncRNA TINCR/miR-31-5p/C/EBP-α feedback loop modulates the adipogenic differentiation process in human adipose tissue-derived mesenchymal stem cells. Stem Cell Res. 2018;32:35–42. doi: 10.1016/j.scr.2018.08.016. [DOI] [PubMed] [Google Scholar]
- 23.Lu W, Shi J, Zhang J, Lv Z, Guo F, Huang H, Zhu W, Chen A. CXCL12/CXCR4 axis regulates aggrecanase activation and cartilage degradation in a post-traumatic osteoarthritis rat model. Int J Mol Sci. 2016;17:E1522. doi: 10.3390/ijms17101522. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 24.Dong Y, Liu H, Zhang X, Xu F, Qin L, Cheng P, Huang H, Guo F, Yang Q, Chen A. Inhibition of CXCL12/CXCR4 signalling in subchondral bone attenuates post-traumatic osteoarthritis. Int J Mol Sci. 2016;17:E943. doi: 10.3390/ijms17060943. [DOI] [PMC free article] [PubMed] [Google Scholar] [Retracted]
- 25.He W, Wang M, Wang Y, Wang Q, Luo B. Plasma and synovial fluid CXCL12 levels are correlated with disease severity in patients with knee osteoarthritis. J Arthroplasty. 2016;31:373–377. doi: 10.1016/j.arth.2015.08.018. [DOI] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.
Data Availability Statement
The datasets used and/or analyzed during the current study are available from the corresponding author on reasonable request.