

Abstract
Tsetse flies (Glossina spp.) vector pathogenic trypanosomes (Trypanosoma spp.) in sub-Saharan Africa. These parasites cause human and animal African trypanosomiases, which are debilitating diseases that inflict an enormous socio-economic burden on inhabitants of endemic regions. Current disease control strategies rely primarily on treating infected animals and reducing tsetse population densities. However, relevant programs are costly, labor intensive and difficult to sustain. As such, novel strategies aimed at reducing tsetse vector competence require development. Herein we investigated whether Kosakonia cowanii Zambiae (Kco_Z), which confers Anopheles gambiae with resistance to Plasmodium, is able to colonize tsetse and induce a trypanosome refractory phenotype in the fly. Kco_Z established stable infections in tsetse’s gut and exhibited no adverse effect on the fly’s survival. Flies with established Kco_Z infections in their gut were significantly more refractory to infection with two distinct trypanosome species (T. congolense, 6% infection; T. brucei, 32% infection) than were age-matched flies that did not house the exogenous bacterium (T. congolense, 36% infected; T. brucei, 70% infected). Additionally, 52% of Kco_Z colonized tsetse survived infection with entomopathogenic Serratia marcescens, compared with only 9% of their wild-type counterparts. These parasite and pathogen refractory phenotypes result from the fact that Kco_Z acidifies tsetse’s midgut environment, which inhibits trypanosome and Serratia growth and thus infection establishment. Finally, we determined that Kco_Z infection does not impact the fecundity of male or female tsetse, nor the ability of male flies to compete with their wild-type counterparts for mates. We propose that Kco_Z could be used as one component of an integrated strategy aimed at reducing the ability of tsetse to transmit pathogenic trypanosomes.
Author summary
Tsetse flies transmit pathogenic African trypanosomes, which are the causative agents of socio-economically devastating human and animal African trypanosomiases. These diseases are currently controlled in large part by reducing the population size of tsetse vectors through the use of insecticides, traps and sterile insect technique. However, logistic and monetary hurdles often preclude the prolonged application of procedures necessary to maintain these control programs. Thus, novel strategies, including those aimed at sustainably reducing the ability of tsetse to transmit trypanosomes, are presently under development. Herein we stably colonize tsetse flies with a bacterium (Kosakonia cowanii Zambiae, Kco_Z) that acidifies their midgut, thus rendering the environment inhospitable to infection with two distinct, epidemiologically important trypanosome strains as well as an entomopathogenic bacteria. In addition to inducing a trypanosome refractory phenotype, colonization of tsetse with Kco_Z exerts only a modest fitness cost on the fly. Taken together, these findings suggest that Kco_Z could be applied to enhance the effectiveness of currently employed tsetse control programs.
Introduction
Insects transmit numerous vertebrate pathogens that cause devastating disease throughout tropical and subtropical regions around the globe. The lack of effective and affordable vaccines, coupled with insect and pathogen resistance to pesticides and drug treatments, respectively, severely limits disease control. Many vertebrate pathogens are acquired by insect vectors via the ingestion of an infectious blood meal. The disease causing agents must then establish an infection in the insect’s gut prior to being transmitted to a new vertebrate host during a subsequent bite. In most cases pathogens are eliminated from the insect vector prior to transmission to a new vertebrate host. This outcome reflects the presence of dynamic active and passive immune barriers that function locally in the insect gut and systemically in the hemocoel [1–3].
Although few insect vectors support transmissible infections with vertebrate pathogens, all house symbiotic microorganisms in their gut that influence numerous aspects of their host’s physiological homeostasis. Symbiotic associations between arthropod disease vectors and enteric bacteria have been particularly well-studied in an effort to determine how these microbes influence their host’s ability to transmit disease [4–8]. Tsetse flies, which are the prominent vectors of pathogenic African trypanosomes, house a taxonomically diverse enteric microbiota that includes endosymbiotic Wigglesworthia and Sodalis [9] as well as an assemblage of bacteria obtained from the fly’s environment [10–12]. Both Wigglesworthia and Sodalis are maternally transmitted to developing intrauterine larvae during tsetse’s unique mode of viviparous reproduction [9,13]. Wigglesworthia influences trypanosome infection establishment in tsetse by regulating the production of trypanocidal PGRP-LB [14,15]. Additionally, tsetse that undergo larval development in the absence of this bacterium fail to synthesize a gut-associated peritrophic matrix during adulthood [16]. This structure is an important mediator of tsetse’s vector competence because it serves as a physical barrier that ingested parasites must traverse in order to successfully colonize the fly’s gut [17] and subsequently the salivary glands for transmission in saliva [18]. Sodalis’ impact on tsetse vector competency is less known, although studies suggest that a positive correlation exists between the prevalence and density of this bacterium and trypanosome infection prevalence [19–23]. Like mosquitoes, tsetse’s gut also harbors a diverse population of bacteria obtained from the fly’s environment [10–12]. However, the effect of these bacteria on tsetse vector competency is poorly understood.
Mosquitoes, including Anopheles gambiae and Aedes aegypti, also house bacteria in their gut, and these microbes play a significant role in the ability of their host to transmit vertebrate pathogens. Boissiere et al. [24] discovered a positive correlation between the density of enteric Enterobacteriaceae and Plasmodium infection prevalence in field-captured An. gambiae. These midgut microbes, as well as the enteric microbiota found in Ae. aegypti, indirectly regulate their host’s vector competency by modulating basal expression of genes that encode anti-Plasmodium and anti-dengue effector proteins [5,25,26]. Other members of the mosquito enteric microbiota exert direct effects on their host’s vector competency. Specifically, a Chromobacterium isolated from Ae. aegypti secretes factors that exhibit anti-Plasmodium and anti-Dengue activity [27]. Also, laboratory reared A. gambiae present an abnormal Plasmodium refractory phenotype when their guts are colonized with Kosakonia cowanii Zambiae, (Kco_Z; previously designated Enterobacter sp Zambiae, Esp_Z [28]) that had been isolated from field-captured mosquitoes. Kco_Z was determined to produce reactive oxygen intermediates (ROIs) that exhibit direct anti-Plasmodium properties [29,30].
In this study we investigated whether Kco_Z isolated from A. gambiae is able to successfully colonize tsetse’s gut and induce parasite and pathogen refractory phenotypes in the fly. We found that this bacterium can reside stably in tsetse’s midgut without imparting a detrimental fitness cost on the fly. Kco_Z colonized tsetse present an acidified midgut environment that is inhospitable to both African trypanosomes and entomopathogenic Serratia marcescens. We discuss the potential utility of Kco_Z as a novel component of currently used area wide integrated pest management strategies aimed at controlling tsetse populations and thus transmission of African trypanosomes.
Results
Bacterial infection outcomes in tsetse’s midgut, and subsequent fly survival
We investigated the ability of Kco_Z and Sodalis (as a control) to colonize the gut of both wild-type (hereafter referred to as ‘GmmWT’) and symbiont-free tsetse (aposymbiotic, hereafter referred to as ‘GmmApo’). GmmWT flies were used to interrogate the interaction between Kco_Z and the natural tsetse microbiota, while the use of GmmApo individuals allowed us to correlate the presence of distinct, experimentally introduced bacterial taxa with specific fly phenotypes. We challenged all flies per os with 1x103 CFU of either Kco_Z or Sodalis in their first blood meal and then monitored bacterial proliferation over a 28 day period. By 7 days post-inoculation, midgut bacterial load in GmmWT flies that housed Kco_Z (GmmWT/Kco_Z) and Sodalis (GmmWT/Sgm) was 1.9x107 ± 6.4x106 CFU and 4.5x105 ± 6.4x106 CFU, respectively, and GmmApo/Kco_Z (9.3x106 ± 5.3x105 CFU) and GmmApo/Sgm (1.4x106 ± 3.9x105 CFU) flies harbored a similar bacterial load at the same time point post-inoculation (Fig 1A). The midgut load of Kco_Z and Sodalis did not change significantly in any of the fly groups over the following 21 days (Fig 1A), thus suggesting that the bacteria had achieved stable-state infections within their fly hosts by one week post-acquisition.
Fig 1. Bacterial colonization of tsetse’s midgut, and the effect on fly survival.

Distinct groups of newly emerged adult wild-type (GmmWT) and aposymbiotic (GmmApo) females were colonized with 1x103 CFU of either Kosakonia cowanii Zambiae (Kco_Z) or Sodalis (Sgm), and then bacterial load and fly survival was measured. (A) Average number (±SEM) of bacterial CFUs per tsetse gut per group per time point. n≥5 individuals per group per timepoint. (B) Kaplan-Meier plot depicting survival of GmmWT and GmmApo females colonized with either Kco_Z or Sgm. Infection experiments were performed using three distinct biological replicates (n = 25 flies per replicate). No significant difference in survival was observed between any of the fly groups (p = 0.88; log-rank test).
We next examined whether midgut infections with Kco_Z or Sodalis impacted tsetse survival. We found that 76% of GmmWT/Kco_Z individuals, and 84% of GmmWT/Sgm individuals, survived for 28 days following bacterial inoculation. Similarly, 84% and 80% of GmmApo/Kco_Z and GmmApo/Sgm flies, respectively, survived the duration of the experiment (Fig 1B). Percent survival was not significantly different between any of these groups, indicating that Kco_Z and Sodalis both exhibit commensal phenotypes in wild-type and aposymbiotic tsetse.
Kco_Z is resistant to Peptidoglycan Recognition Protein-LB (PGRP-LB)
The midgut of adult tsetse expresses peptidoglycan recognition protein LB (pgrp-lb), which encodes a pattern recognition receptor that exhibits potent antimicrobial activity [14,15]. Thus, in order to colonize tsetse’s midgut, a microorganism must be resistant to this molecule. We investigated whether innate resistance to PGRP-LB represents one mechanism that allows Kco_Z to colonize tsetse’s gut. We found that 108% (±16) of Kco_Z cells were able to survive 1 h in the presence of recPGRP-LB, while only 2.3% (±1.0) of E. coli cells survived for the same time period. Additionally, 154% (±14) of Sodalis cells survived following a 12 h incubation with recPGRP-LB (Fig 2). These findings suggest that like native Sodalis, Kco_Z is resistant to the antimicrobial properties of PGRP-LB and is able to survive in the presence of this protein (as indicated by a slight increase in bacterial load compared to the initial inoculate). This phenotype may facilitate this bacterium’s ability to successfully colonize tsetse’s gut.
Fig 2. Kco_Z is resistant to normally bactericidal Peptidoglycan Recognition Protein-LB.
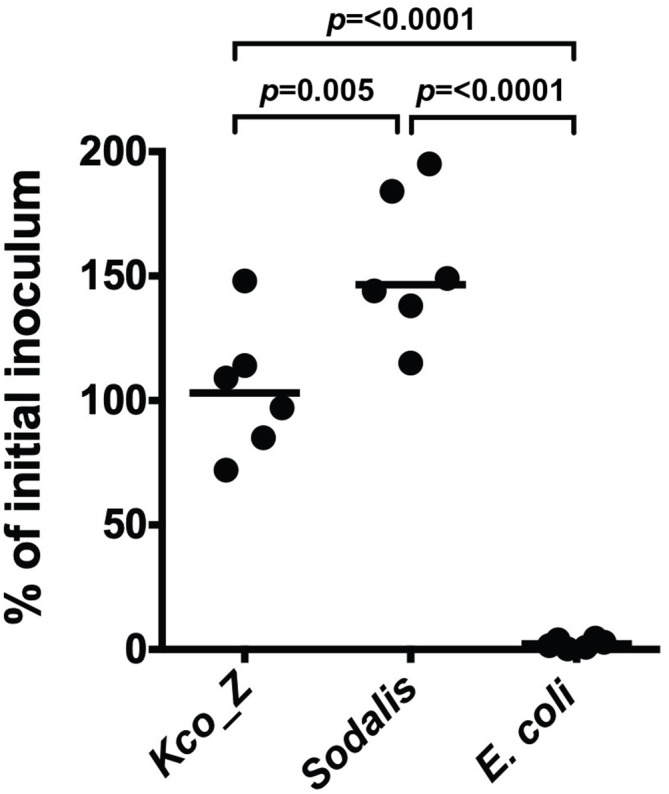
Survival of cultured Kco_Z, Sodalis and E. coli following exposure (1 hr for Kco_Z and E. coli, and 24 hr for Sodalis) to recombinant (rec) PGRP-LB (10 μg/ml). Results are presented as % of initial inoculum, which was determined by dividing the number of bacterial CFU present after treatment with recPGRP-LB by the number of CFU present prior to inoculation. Each point on the graph represents a distinct bacterial culture. Statistical significance was determined using a one-way ANOVA followed by Tukey’s HSD post-hoc analysis.
Kco_Z colonized aposymbiotic tsetse present a trypanosome refractory phenotype
Kco_Z successfully colonizes the gut of GmmApo flies, resides in the niche for at least 28 days, and has no impact on fly survival during that time period. Thus, we next evaluated whether colonization with this bacterium impacts trypanosome infection establishment in tsetse’s midgut. We began by challenging mature GmmApo flies because they are highly susceptible to trypanosome infection (~50%) while their age-matched GmmWT counterparts are refractory (~3%) [16]. Distinct groups of eight day old GmmApo/Sgm and GmmApo/Kco_Z flies, which housed similar numbers of their respective exogenous bacteria (S1A Fig), were administered a meal supplemented with 1x106 blood stream form (BSF) trypanosomes per ml of blood. Thereafter all flies were maintained on regular blood for two weeks, at which point their midguts were dissected and microscopically examined for the presence of parasites. An age-matched control cohort consisted of similarly challenged GmmApo flies. We found that infection prevalence in the GmmApo/Sgm group (57%) was similar to that of GmmApo controls (52%), while infection prevalence in GmmApo/Kco_Z individuals was significantly lower (19%) (Fig 3A). These data indicate that the presence of Kco_Z in tsetse’s gut interferes with the ability of trypanosomes to establish an infection in this niche. This parasite resistant phenotype is similar to that which occurs in the gut of Kco_Z colonized mosquitoes following exposure to malaria parasites [29,30].
Fig 3. The trypanosome refractory phenotype exhibited by GmmApo/Kco_Z flies is not directly caused by reactive oxygen intermediates.

(A) Percentage of GmmApo, GmmApo/Sgm and GmmApo/Kco_Z flies harboring midgut infections with bloodstream form (BSF) YTAT 1.1 trypanosomes. Statistical analysis was performed using a GLM followed by multiple comparisons and Tukey contrasts, and different letters represents statistical significance between treatments and controls. (B) Mid-log phase cultures of Kco_Z and Sodalis synthesize similar quantities of H2O2 (p = 0.6; paired t-test). Measurements were taken from 7 distinct clonal populations of each bacterium. (C) Percentage of GmmApo and GmmApo/Kco_Z flies infected with BSF YTAT 1.1 trypanosomes after being fed blood meals containing antioxidant vitamin C over the course of the 14 day experiment. Despite exposure of both tsetse groups to the ROI-suppressing vitamin, GmmApo/Kco_Z flies were still significantly more refractory to trypanosome infection than were GmmApo individuals (p = 0.002; GLM Wald test). In (A), (B) and (C) different letters represents statistical significance between treatments and treatments and controls.
African trypanosomes are not susceptible to Kco_Z generated reactive oxygen intermediates
In A. gambiae, Kco_Z produces reactive oxygen intermediates (ROIs) that are directly toxic to Plasmodium [29,30]. ROIs have also been implicated as mediators of trypanosome infection outcomes in tsetse. Specifically, tsetse are rendered susceptible to trypanosome infection when the initial infectious blood meal is supplemented with the antioxidants vitamin C or cysteine [18,31]. These antioxidants detoxify ROIs that otherwise induce programmed cell death processes in trypanosomes [32]. In light of this information, we investigated the correlation between Kco_Z generated ROIs and the trypanosome refractory phenotypes we observed in adult GmmApo/Kco_Z individuals. As an indicator of bacterial ROI production, we quantified H2O2 concentrations in supernatants from mid-log phase Kco_Z and Sodalis cultures. Kco_Z and Sodalis supernatants contained 127nM (±15) and 142nM (±13) of H2O2, respectively (Fig 3B).
We next tested whether ROIs produced by Kco_Z inhibit the ability of trypanosome to infect GmmApo/Kco_Z flies. Individual groups of eight day old GmmApo and GmmApo/Kco_Z individuals were offered a blood meal containing infectious trypanosomes together with the antioxidant vitamin C. All trypanosome challenged flies were subsequently maintained on vitamin C supplemented blood for 14 days. Under these conditions, 74% of GmmApo+vitC flies were infected with trypanosomes, while only 11% of their GmmApo/Kco_Z+vitC counterparts housed parasite infections (Fig 3C). These results suggest that ROIs produced by Kco_Z that reside stably in tsetse’s gut are not the sole determinants of the fly’s susceptibility to infection with trypanosomes.
Kco_Z produces acid that is toxic to trypanosomes
We observed that GmmApo/Kco_Z flies are significantly more refractory to infection with trypanosomes than are GmmApo/Sgm individuals, despite the fact that Sodalis and Kco_Z produce similar amounts of H2O2. This outcome implies that Kco_Z modulates trypanosome infection outcomes in tsetse via a mechanism other than ROI production. Members of the genus Kosakonia [33–35], as well as several enteric commensals including Enterobacter spp. [36–38], produce organic acids, and these products can inhibit pathogen growth by creating an acidic environment [39,40]. Because many trypanosomatids, including members of the genera Trypanosoma and Leishmania, are highly sensitive to environmental pH [41,42], we investigated whether Kco_Z creates an acidic environment that prohibits T. brucei growth in vitro. Specifically, we heat killed (HK) Kco_Z (1x106 log-phase in 500 μl of LB media) and added the solution to trypanosome cultures maintained in vitro. This medium includes phenol red, which is a pH-sensitive dye that when in solution turns from red-pink to yellow as the quantity of acid in the environment increases. Addition of this HK Kco_Z extract immediately turned the Beck’s media yellow, and the pH measured at 5.8 (± 0.39). This value was significantly lower than trypanosome cultures that were supplemented with 500 μl of 1x106 log-phase HK trypanosomes (pH 7.3 ± 0.28), HK Sodalis (pH 7.4 ± 0.39) or LB (Kco_Z growth media; pH 7.1 ± 0.28) or MM media (Sodalis growth media; pH 7.2± 0.29) alone (Fig 4A). We subsequently monitored trypanosome growth in cultures that received the above-mentioned supplements. We observed that trypanosomes failed to replicate in Beck’s medium that contained HK Kco_Z extracts, while trypanosomes multiplied in all of the other culture conditions (Fig 4B).
Fig 4. Kco_Z produces a low pH environment that is toxic to trypanosomes.

(A) Early log phase trypanosomes (T. b. brucei YTAT 1.1), cultured in 10ml of Beck’s medium containing the pH sensitive dye phenol red, exposed to 1ml of heat treated LB media (Kco_Z culture medium), 1ml of heat treated BHI media (Sodalis culture medium), heat killed (HK) Kco_Z (5x106 cells) in 1ml of LB media and HK Sodalis (5x106 cells) in 1ml of BHI media. Controls are trypanosomes alone (tryps). All heated treatments and controls were allowed to cool to room temperature prior to adding them to the trypanosome cultures. Two hours post-treatment, culture pH was measured. HK Kco_Z significantly reduced the pH of the trypanosome culture (p<0.001). The experiment was repeated using 6 distinct clonal trypanosome populations (the image represents one of the six replicates). (B) Density of trypanosomes in culture 2h, 5h and 24h after addition of the treatments described in (A) above. At the 24h time point, all trypanosomes exposed to HK Kco_Z extracts were dead while those from the other groups were replicating similarly to controls. (C) Density of trypanosomes cultured in normal (pH 7.2) and artificially produced (via the addition of 0.1N HCl) acidic (pH 5.8) environments. Artificial acidic conditions kill all trypanosomes with 24 h. (D) The density of cultured trypanosomes exposed to HK Kco_Z extracts buffered to pH 7.2 (via the addition of 0.1N NaOH). The buffering treatment rescues parasite growth. In (A), (B) and (C), statistical significance was determined using a one-way ANOVA followed by Tukey’s HSD post-hoc analysis in (A), and a two-way ANOVA followed by Tukey’s HSD post-hoc analysis in (B) and (C). Different letters represent statistical significance between treatments and controls. In (B), (C) and (D), experiments were performed using 5 or 6 distinct clonal trypanosome populations.
Heat-killed Kco_Z extracts create an acidic environment when added to trypanosome cultures, and trypanosomes fail to replicate in this environment. These findings do not rule out the possibility that trypanosomes are capable of surviving Kco_Z-induced acidic conditions, and instead, some other unknown component of the medium [e.g., a bacterium-derived trypanocidal molecule(s)] exhibits toxic properties. To address this possibility, we monitored trypanosome growth in Beck’s medium, the pH of which was artificially decreased to 5.8 (the same as that achieved by adding HK Kco_Z extracts) via the addition of exogenous acid. Under these conditions trypanosomes failed to replicate (Fig 4C). Furthermore, when we buffered Beck’s medium containing HK Kco_Z extracts back up to pH 7.2, trypanosomes replicated normally (Fig 4D). Taken together, these data indicate that Kco_Z produces an acidic environment that is toxic to trypanosomes, thus impeding their growth in vitro.
Kco_Z acidifies tsetse’s gut
We observed that trypanosomes are unable to multiply when cultured in medium supplemented with acidic Kco_Z extracts. Thus, we next investigated whether Kco_Z produces acid in vivo in tsetse’s gut. To do so we colonized teneral, aposymbiotic flies with either Kco_Z or Sodalis, and 5 days later fed them a meal containing 2.5% sucrose and 0.04% phenol red solubilized in water. Twenty-four hours later, midguts from a sample of flies (n = 8 per group) were excised and plated on solid medium containing phenol red. GmmApo/Kco_Z and GmmApo/Sgm flies housed similar densities of the introduced bacteria (S1B Fig), and their respective mediums changed color to reflect corresponding pH shifts (S1C Fig). The remaining flies were dissected to expose their midgut in situ, and the color of the gut contents was visualized microscopically. We observed that the gut contents of GmmApo/Kco_Z flies were yellow in color (Fig 5), thus indicating that the environment had become acidified. Conversely, the gut contents of GmmApo/Sgm individuals were red, which is similar to the more alkaline environment present in the gut of GmmWT tsetse (Fig 5).
Fig 5. Kco_Z acidifies tsetse’s gut.

Midgut pH of GmmApo/Kco_Z, GmmApo/Sgm, GmmWT/Kco_Z and GmmWT flies. Distinct groups of teneral GmmApo flies were inoculated per os with either Kco_Z or Sodalis, while GmmWT individuals received Kco_Z (all flies received 5x104 CFU of bacteria per ml of blood) or no bacteria. Five days post-inoculation, all individuals were offered a meal containing sucrose (2.5%) and phenol red (0.04%) dissolved in water. Twenty-four hours later, fly abdomens were excised and the color of the solution found within the midgut was observed. Each image represents one of five flies monitored for each treatment.
Finally, we investigated whether Kco_Z also produces acid in the gut GmmWT tsetse by inoculating teneral individuals with 1x103 CFU of the bacterium (these flies were designated GmmWT/Kco_Z). Five days later a cohort of GmmWT/Kco_Z females (these flies housed 1.27x106 ± 8.6x104 Kco_Z at this time point; S1D Fig), as well as age matched GmmWT controls, were fed a sugar meal containing phenol red (as described above) to observe gut pH. Similar to our results noted in GmmApo/Kco_Z flies, we observed that the gut of GmmWT/Kco_Z individuals was yellow, thus indicative of an acidified environment. Conversely, the gut environment of GmmWT flies was red and thus comparatively alkaline (Fig 5). Thus, the presence of indigenous symbionts does not impede the ability of Kco_Z to acidify the gut of wild-type flies.
GmmWT/Kco_Z are highly refractory to infection with trypanosomes and entomopathogenic bacteria
We hypothesized that exogenous microorganisms would be unable to successfully infect GmmWT/Kco_Z due to their acidified midgut environment. To test this hypothesis we first co-inoculated teneral GmmWT males with Kco_Z and T. congolense parasites. Two weeks post-challenge we observed no significant difference in the percentage of GmmWT/Kco_Z (15%) and control GmmWT (23%) flies that harbored trypanosome infections in their midguts (Fig 6A). We next inoculated teneral GmmWT males with Kco_Z and then three days later (5 day old adults) challenged GmmWT/Kco_Z individuals with either T. congolense or T. brucei parasites (both of these parasite species are naturally transmitted by G. m. morsitans; [43,44]). Under these conditions we observed that GmmWT/Kco_Z males were significantly more refractory to infection with both parasite species (T. congolense, 6%; T. brucei, 32%) than were their age-matched GmmWT counterparts (T. congolense, 36%; T. brucei, 70%; Fig 6B). Thus, tsetse must house an established Kco_Z infection in its gut at the time of trypanosome challenge in order to present a refractory phenotype. Finally, we found that trypanosome infected GmmWT/Kco_Z house similar densities of Kco_Z as do age-matched individuals that eliminated their trypanosome infection (S1E Fig), again indicating that exogenous Kco_Z appears to be resistant to tsetse’s trypanocidal immune response. Additionally, the presence of tsetse’s indigenous, enteric microbiota does not interfere with Kco_Z mediated obstruction of trypanosome infection establishment.
Fig 6. GmmWT/Kco_Z flies are significantly more refractory to infection with parasitic trypanosomes and entomopathogenic S. marcescens.

(A) Percentage of GmmWT and GmmWT/Kco_Z flies infected with T. congolense 14 days after they were co-inoculated with Kco_Z and parasites in their first (teneral) blood meal. (B) Percentage of GmmWT and GmmWT/Kco_Z flies infected with T. congolense (left graph) and T. brucei (right graph). For these experiments GmmWT/Kco_Z flies housed the exogenous bacteria for five days prior to challenge with trypanosomes. In (A) and (B) Statistical analysis was performed using a GLM followed by multiple comparisons and Tukey contrasts, and different letters represents statistical significance between treatments and controls. (C) Kaplan-Meier plot depicting survival of GmmWT and GmmWT/Kco_Z flies following challenge (three days after administering Kco_Z or Sodalis) with S. marcescens. Infection experiments were performed using three distinct biological replicates (n = 25 flies per replicate). GmmWT/Kco_Z flies were also significantly more refractory to S. marcescens infection than were their GmmWT counterparts (p = 0.001; log-rank test).
Finally, we investigated whether Kco_Z also protects tsetse against infection with an entomopathogenic bacteria. To do so teneral GmmWT males were fed 1x103 CFU of Kco_Z, and then three days later, the same dose of Serratia marcescens strain db11, which is highly virulent to wild-type tsetse [17,18,45]. Five day old GmmWT males infected with the same dose of S. marcescens were used as controls. Fly survival following Serratia inoculation was monitored over a 14 day period in both fly groups. We observed that 51% and 9% of GmmWT/Kco_Z and GmmWT individuals, respectively, survived their infection with S. marcescens (Fig 6C).
Taken together, our results detailed above indicate that wild-type tsetse present a parasite and entomopathogen refractory phenotype when they house an established Kco_Z infection in their gut. This phenotype like occurs because the acidified nature of the gut environment is inhospitable to the development of exogenous microbes.
Kco_Z infection exerts a minimal fitness cost on tsetse
Kco_Z produces acid in tsetse’s gut such that the environment becomes inhospitable to trypanosomes. To address whether decreased midgut pH adversely impacts tsetse fitness, we quantified several fitness parameters in GmmWT/Kco_Z flies (a sample of these individuals housed 1.39x106 ± 1.1x105 Kco_Z at the time they were used for experimentation; S1F Fig).
We began by measuring midgut weight, which reflects over all digestive health. We observed no significant difference in midgut weight between 8 day old GmmWT/Kco_Z and GmmWT males (3.8 ± 1.1 mg and 3.2 ± 1.2 mg, respectively) and females (12.8 ± 1.8 mg and 13.0 ± 1.8 mg, respectively) 24 hrs after acquiring their last blood meal (Fig 7A). We next measured fecundity parameters in female and male GmmWT/Kco_Z and GmmWT flies to determine if stable infection with this bacterium would alter their reproductive capacity. We began by measuring gonotrophic cycle (GC) duration of GmmWT/Kco_Z and GmmWT females. The length of the 1st GC was not significantly different between GmmWT/Kco_Z (24.0 ± 1.2 days) and GmmWT females (24.0 ± 0.9 days) (Fig 7B). However, the 2nd and 3rd GCs of GmmWT/Kco_Z females (13.0 ± 1.1 and 14.0 ± 1.1 days, respectively) were significantly longer than those of their age-matched WT counterparts (11.7 ± 1.0 and 11.5 ± 1.1 days, respectively) (Fig 7B). We also determined that pupal weight from all three GCs was similar between both fly groups (GC1, GmmWT/Kco_Z = 23.1 ± 1.5 mg, GmmWT = 22.9 ± 1.4 mg; GC2, GmmWT/Kco_Z = 23.4 ± 1.7 mg, GmmWT = 24.2 ± 1.8 mg; GC3, GmmWT/Kco_Z = 24.7 ± 1.7 mg, GmmWT = 24.4 ± 1.6 mg) (Fig 7C). Thus, Kco_Z infection impacts the reproductive physiology of female tsetse by increasing GC duration and hence the number of offspring infected individuals are able to produce over the course of their lifespan. However, despite this effect, infection with this bacterium does not impact pupal weight.
Fig 7. Kco_Z infection impacts specific tsetse fitness parameters.

(A) Midgut weight, as an indicator of blood meal digestion proficiency, in six day old GmmWT/Kco_Z and GmmWT males and females (guts were weighed 24 h after the flies had consumed their last blood meal). Each point on the graph represents one individual, and statistical significance was determined via multiple t-tests. (B). Gonotrophic cycle (GC) length of GmmWT/Kco_Z and GmmWT females. Age-matched, pregnant females from each group (n = 35 per group) were housed in individuals cages and monitored daily to observe frequency of pupal deposition. Statistical significance was determined via log-rank test. (C) Weight of pupae from GmmWT/Kco_Z and GmmWT females over three GCs. Each point on the graph represents one individual, and statistical significance was determined via multiple t-tests. (D) Mating competitiveness of GmmWT/Kco_Z compared to GmmWT males. Matings were setup in individual cages (n = 80). Each cage housed a virgin female, to which one virgin GmmWT/Kco_Z and GmmWT male was added. Males and females were age-matched, and each had fed twice prior to exposure. Statistical significance was determined via Chi-squared test. (E) Sperm abundance in the reproductive tracts of three and 14 day old (fed twice) virgin GmmWT/Kco_Z and GmmWT males. Sperm quantity is a reflection of sperm-specific dynein intermediate chain (sdic) transcript abundance. Absolute sdic transcript abundance was determined by comparing experimental sample cycle threshold (Ct) values to those derived from an sdic internal standard curve. Each point on the graph represents one individual, and statistical significance was determined via multiple t-tests. (F) Quantitation of Wigglesworthia and Sodalis in GmmWT/Kco_Z and GmmWT male and female tsetse. Abundance of symbiont specific gene transcripts (Wigglesworthia, thiC; Sodalis, fliC) was used as a proxy to quantify bacterial load. This was performed by comparing thiC and fliC cycle threshold (Ct) values in GmmWT/Kco_Z and GmmWT flies to those derived from symbiont gene-specific internal standard curves. Wigglesworthia and Sodalis can be polyploid (Rio et al., 2006; Weiss et al., 2006), and as such, we normalized symbiont genome copy number to constitutively expressed tsetse gapdh copy number. Each point on the graph represents one individual, and statistical significance was determined via multiple t-tests.
We also investigated the effect of Kco_Z infection on the reproductive fitness of male tsetse by comparing the mating competitiveness of GmmWT/Kco_Z and GmmWT individuals. To do so we set up 80 individual cages, each of which contained one sexually mature virgin female. We subsequently placed one sexually mature GmmWT/Kco_Z and GmmWT male in each cage and monitored the arena to determine which of the two males successfully mated with the female. We observed that 47.5% of matings occurred between GmmWT/Kco_Z males and females (neither male mated with the female in two of the cages) (Fig 7D), thus indicating that Kco_Z infection does not significantly alter male mating competitiveness (χ2 = 0.1025641, df = 1, p = 0.748774). Next we compared the number of sperm present in three and 14 day old GmmWT/Kco_Z and GmmWT males by quantifying transcript abundance of sperm-specific dynein intermediate chain (sdic). The Drosophila homologue of this gene is transcribed exclusively in sperm cells [46] and is used to quantify sperm abundance [47]. We observed no significant difference in sdic transcript abundance between three day old or 14 day old GmmWT/Kco_Z and GmmWT males (Fig 7E).
Finally, we examined whether the presence of Kco_Z impacts the load of endosymbiotic Wigglesworthia and Sodalis. These measurements, which were taken at 14 days post-inoculation with Kco_Z, are important because tsetse’s microbiota impact many aspects of their host’s fitness, including fecundity and immune system development and function [48,49]. We observed that infection with Kco_Z did not significantly alter the load of tsetse’s midgut (bacteriome) population of obligate Wigglesworthia in GmmWT/Kco_Z males or females (Fig 7F). Conversely, midguts from GmmWT/Kco_Z males and females housed significantly fewer Sodalis than did midguts from their GmmWT counterparts (Fig 7F). Taken together, these data indicate that Kco_Z significantly impacts specific fitness parameters in both female and male flies.
Discussion
Morbidity and mortality caused by vector-borne diseases currently inflicts a devastating socioeconomic burden on a significant percentage of the global population. To reduce this burden, novel disease control strategies that inhibit pathogen maturation within arthropod disease vectors require development. The enteric microbiota is being increasingly studied for use in this context, and one such novel strategy could employ the use of ‘probiotic’ bacteria, the presence of which would alter the physiology of the vector’s gut to make the environment inhospitable to pathogens. Herein we use the tsetse fly model system to highlight how an exogenous bacterium can be employed in this capacity to impede infection establishment of two pathogens in this insect disease vector. Specifically, we determined that Kco_Z, which is a bacterium found naturally in the gut of some An. gambiae populations, and directly kills Plasmodium by producing anti-parasitic ROIs [29,30], can stably colonize tsetse’s gut for at least 28 days. When the bacterium is present in this niche, tsetse are significantly more refractory to infection with parasitic African trypanosomes and entomopathogenic S. marcescens than are flies that house only their indigenous microbiota. Kco_Z creates this inimical environment by acidifying tsetse’s gut such that trypanosomes and S. marcescens, which are sensitive to these conditions, are no longer able to successfully infect the fly. While infection with Kco_Z exerts only a negligible effect on tsetse’s reproductive fitness, the bacterium’s presence does reduce the load of endosymbiotic Sodalis. Cumulatively, our findings suggest that Kco_Z could be used in a natural setting to artificially reduce disease transmission by this arthropod vector.
Herein we demonstrate that Kco_Z is able to stably colonize tsetse’s gut, which is an outcome that likely results at least in part from the bacterium’s resistance to antimicrobial PGRP-LB. This protein is constitutively produced in the fly’s midgut and directly kills trypanosomes and E. coli K12 (Fig 2 in this study, and [16]). Kco_Z resistance to tsetse PGRP-LB may reflect one of many bacterial adaptations that result from residing within the immunologically hostile environment of the insect midgut. While the specific physiological mechanism(s) that Kco_Z uses to facilitate its colonization of tsetse’s midgut are currently unknown, the bacterium survives for prolonged periods within the gut of An. gambiae in part by increasing expression of genes that encode a type III secretion system apparatus protein as well as glutathione S-transferase and oxidoreductase [30]. Type III secretion system proteins can facilitate bacterial penetrance into host cells [50] and be involved in subversion of host immunity [51], while the latter two proteins are antioxidant pathway components that mediate redox homeostasis in oxidatively stressful environments such as the insect midgut [52,53]. Kco_Z may employ similar mechanisms to survive in tsetse’s immunologically hostile gut. Sodalis is also resistant to tsetse antimicrobial immune response [9,54], which may be the result of structural adaptations present in exposed bacterial surface coat molecules, including lipopolysaccharide [55] and outer membrane protein A [56]. Furthermore, Sodalis enters into host cells through the use of a type III secretion system [57], which may further protect the bacterium from tsetse’s immunologically hostile midgut environment. Likewise, similar mechanisms may facilitate Kco_Z survival in this niche. Additionally, Kco_Z, like Sodalis [58], may reside extracellularly in the endoperitrophic space of tsetse’s midgut. In this position, the fly’s peritrophic matrix barrier would physically separate the bacteria from immunocompetent epithelial cells, thus reducing their exposure to harmful antimicrobial responses.
Microbes can alter their environment such that it either favors or hinders its own prosperity as well as the prosperity of other resident organisms [59]. Depending on specific physiological circumstances, these effects can reflect the consumption of resources and/or the production of beneficial or harmful metabolic byproducts [60,61]. Tsetse’s sole energy source, vertebrate blood, is rich in glucose. Many bacterial taxa, including members of the Enterobacteriaceae (in which Kosakonia and Enterobacter spp. cluster), ferment this sugar, thus producing hydrogen that acidifies their environment (38,61]. The acidic environment present in tsetse’s gut when Kco_Z resides stably in the tissue likely results at least in part from the bacterium’s utilization of blood glucose as an energy source. The pH in the gut of insect vectors, including tsetse flies [62], sand flies [63] and mosquitoes [64] is normally alkaline, and the parasites they transmit, as well as other enteric microbes, are adapted to survive in this environment. Correspondingly, our results indicate that Kco_Z induced conditions in tsetse’s gut detrimentally impact not only trypanosomes but also other enteric microbes including entomopathogenic S. marcescens and symbiotic Sodalis. Kco_Z mediated suppression of S. marcescens, or any other pathogen, would have obvious benefits to the fly. However, dysbiosis of tsetse’s facultative and commensal enteric microbiota could impact the fly’s overall fitness and/or vector competency. For example, reducing Sodalis density significantly decreases tsetse longevity [20]. This may prove beneficial because flies with a reduced life span could perish before trypanosomes are able to complete their 20–30 extrinsic incubation period [65]. A reduction in Sodalis density could be further beneficial because tsetse that house relatively low densities of the bacterium are less likely to be infected with trypanosomes than are individuals that house more of the symbiont [19–23]. Thus, the trypanosome refractory phenotype presented by Kco_Z colonized tsetse may result in part from, or be enhanced by, the fact that they contain fewer Sodalis. Finally, the midgut of wild tsetse is colonized by a transient population of environmentally acquired bacteria [9]. The contribution of these bacteria to tsetse’s physiology has not been characterized, and as such, interference with this microbial population could further alter the fly’s physiological homeostasis. To the contrary, the environmentally acquired microbiota could out-compete Kco_Z that reside in tsetse’s gut, or could prevent the bacterium from acidifying the environment. Future studies are required to elucidate microbe-microbe interactions in the gut Kco_Z colonized flies after their release into the field.
Reducing the incidence of African trypanosomiases has to date been achieved largely by controlling the size of tsetse populations. This process is currently accomplished by employing area wide integrated pest management (AW-IPM) strategies that make use of insecticides, traps and sterile insect technique (SIT) [66,67]. SIT involves sequentially releasing a large number of sterilized males (achieved by irradiating pupae or adults) into the target environment. These males reproductively outcompete wild males for female mates, and the population size drops significantly, or the fly is completely eradicated [68]. The efficacy of SIT as a means of controlling tsetse populations is well exemplified on Unguja Island (the large island of the Zanzibar archipelago), where the technique was used to eradicate G. austeni, the main vector of trypanosomes that cause animal African trypanosomiasis in that locale [69]. One shortcoming of this procedure is that releasing large numbers of sterile males significantly increases the population of potential disease vectors in that environment (male tsetse also feed exclusively on vertebrate blood). One way to overcome this obstacle is to release sterilized males that present enhanced refractoriness to parasite infection. This outcome is currently achieved by feeding the sterilized males twice with the drug isometamidium chloride prior to their release [70]. However, treated flies are not 100% resistant to infection [70], and the risk exists that the parasite will eventually develop resistance to the drug. Our data presented herein indicate that inoculating sterilized males with Kco_Z prior to their release would serve as an alternative, or supplemental, means of making the flies resistant to infection. Specifically, GmmWT/Kco_Z males (and females) are significantly more refractory to infection with trypanosomes than are their wild-type counterparts. This finding implies that sterilized, GmmWT/Kco_Z individuals would be relatively poor vectors of disease-causing trypanosomes and thus safer to release than sterilized males that do not house this bacterium. Furthermore, our preliminary analyses suggest that Kco_Z infection does not compromise the mating competitiveness nor sperm abundance of male tsetse, thus suggesting that GmmWT/Kco_Z individuals would be as successful as their wild counterparts at locating females and engaging in viable matings. Finally, male tsetse could be irradiated as pupae or teneral adults [71], prior to colonization with Kco_Z, thus eliminating the possibility that this treatment could detrimentally impact the bacterium’s fitness and thus its effect on fly vector competency. These characteristics provide preliminary evidence that releasing sterilized, Kco_Z infected male tsetse as part of an AW-IPM program would significantly reduce the capacity of these flies to transmit disease.
In conclusion, data presented in this study indicates that Kco_Z could effectively complement currently used AW-IPM programs aimed at reducing or eliminating tsetse populations by inhibiting trypanosome infection establishment in the fly’s gut. However, the complex relationship between tsetse, its indigenous (endosymbionts) and exogenous (trypanosomes and environmentally acquired microorganisms) microbiota, and Kco_Z must be studied in more detail before the bacterium is used in this capacity. Of particular importance are studies aimed at determining whether Kco_Z presents trypanocidal activity in other epidemiologically important tsetse species (e.g., G. fuscipes). Furthermore, field-based studies would shed light on how the ecology of tsetse’s natural environment influences the overall efficacy of the system.
Materials and methods
Ethical consideration
This work was carried out in strict accordance with the recommendations in the Office of Laboratory Animal Welfare at the National Institutes of Health and the Yale University Institutional Animal Care and Use Committee. The experimental protocol was reviewed and approved by the Yale University Institutional Animal Care and Use Committee (Protocol 2011–07266).
Tsetse, bacteria and trypanosomes
Tsetse flies (Glossina morsitans morsitans) were maintained in Yale University’s insectary at 24°C with 55% relative humidity. Flies received defibrinated (via mechanical agitation for 20 minutes) bovine blood (Quad Five, Ryegate, MT, USA) through an artificial membrane feeding system every 48 h [72]. Aposymbiotic tsetse (GmmApo) were generated and maintained as described previously [73]. Throughout the manuscript, flies referred to as ‘teneral’ were unfed adults recently eclosed (≤ 24h) from their pupal case. All tsetse lines used in this study are described in S1 Table.
Sodalis were isolated from tsetse pupae as described previously [74], and subsequently maintained in liquid brain heart infusion (BHI) media (Becton Dickinson). When necessary, Sodalis were plated on either Brain Heart Infusion agar supplemented with 10% defibrinated bovine blood (BHIB) or Mitsuhashi-Maramorosch (MM)-agar plates. Kosakonia cowanii Zambiae [previously designated Enterobacter sp Zambiae (Esp_Z); GenBank accession number CP022690.1; 28], isolated from the gut of the mosquito, Anopheles gambiae [25]), and Serratia marcescens (strain db11; isolated from a moribund Drosophila sp.) [75] were grown in liquid LB media or on LB-agar plates at 30°C.
In vivo Trypanosoma congolense and T. brucei brucei (YTAT 1.1) were expanded in rats and harvested from infected blood at peak parasitemia. Rat blood containing blood stream form (BSF) parasites was aliquoted and cryopreserved (in the liquid nitrogen vapor phase, approximately -150°C) for subsequent tsetse challenge experiments.
Recombinant PGRP-LB antibacterial assays
Antibacterial activity of recombinant (rec) PGRP-LB was determined as described previously [15], with minor modification. Specifically, recPGRP-LB was added (10 μg/ml of media) to early log-phase (OD = 0.2–0.4) cultures of Kco_Z, Sodalis and E. coli. Controls consisted of bacterial cultures exposed to bovine serum albumin. Using a plate-based quantification assay [58], E. coli and Kco_Z load was subsequently measured 1 hr. later, while Sodalis load was measured 24 hr. later. Results are presented as % of initial inoculum, which was determined by dividing the number of bacterial CFU present after treatment with recPGRP-LB by the number of CFU present prior to inoculation.
Microbial infection assays
Per os bacterial challenge of wild-type (GmmWT) and GmmApo flies was performed by feeding teneral adults a heat inactivated (HI; 56°C for 1 hr) blood meal (to inactivate the vertebrate complement system) inoculated with 5x104 colony forming units (CFU) of each bacterial strain per ml of blood. Because tsetse flies consume approximately 20 μl of blood per feeding, each fly is inoculated with 1x103 bacterial cells. GmmApo flies colonized with either Sodalis or Kco_Z are designated GmmApo/Sgm and GmmApo/Kco_Z, respectively, and GmmWT flies colonized with Kco_Z are designated GmmWT/Kco_Z. For all experiments that employed tsetse flies inoculated with either Sodalis or Kco_Z, bacterial midgut load was determined by homogenizing microscopically dissected gut tissue in 0.85% NaCl and serially diluting and plating the samples on LB-agar (E. coli, Kco_Z and Serratia) or BHIB or MM (Sodalis) plates supplemented with antibiotics [58]. CFU per plate were counted manually, and counts are presented in the corresponding Results subsections.
For trypanosome infections, all flies received infectious blood meals containing 1x106/mL BSF T. congolense or T. b. brucei parasites. GmmApo, GmmApo/Sgm and GmmApo/Kco_Z were challenged as eight day old adults (3rd blood meal), while GmmWT and GmmWT/Kco_Z flies were challenged as five day old adults (2nd blood meals). For Kco_Z/trypanosome co-infection experiments, distinct groups of mature GmmWT individuals were inoculated with 1x106/mL BSF T. congolense parasites and 5x104 CFU/ml of Kco_Z. Two weeks post-trypanosome challenge, all flies were dissected and their midguts microscopically examined to determine parasite infection status.
Detection and inhibition of tsetse reactive oxygen intermediates
Kco_Z and Sodalis cultures were grown to mid-log phase (OD = 0.25), and cell-free supernatants were generated via centrifugation. Hydrogen peroxide (H2O2) concentrations in bacterial culture supernatants were determined using an Amplex Red Hydrogen Peroxide/Peroxidase assay kit according to the manufacturer’s (Invitrogen) protocol. In brief, supernatants were incubated for 30 min. with the assay reagent, and resulting fluorescence units were quantified using a Bio-Tek plate reader.
Antioxidants were used to inhibit tsetse ROI activity in vivo. The assay used was similar to those described previously [18,29,30]. In brief, treated flies were offered a blood meal inoculated with trypanosomes [1x106/mL BSF T. b. brucei (YTAT 1.1)] and supplemented with vitamin C (10mM) or cysteine (10μM). All subsequent meals also contained antioxidant supplements.
Determination of bacterial acid production in vitro
Sodalis and Kco_Z were grown in their respective liquid media to an O.D. of 1.0. Subsequently, 5x106 cells (this value represents the approximate maximum load to which these bacteria grow in tsetse’s gut; see Fig 1A) were diluted to a volume of 1 ml (again in respective liquid media) and heat-killed (80°C for 1.5 hr). Conditioned media containing dead cells was added to early log growth phase T. b. brucei YTAT 1.1 grown in a Beck’s medium (GE Hyclone), which contains phenol red. When in solution this compound serves as a pH-sensitive colorimetric indicator that changes from pink-red to yellow as environmental pH drops. Other treatment groups were inoculated with 1 ml of heated, clean LB (Kco_Z growth medium) or clean BHI (Sodalis growth medium), while the control group consisted of trypanosomes alone. Two hours after exposing T. b. brucei to treatment conditions, cultures were assayed to determine pH using a Mettler Toledo pH meter. The pH of trypanosome containing Beck’s medium was experimentally reduced (to pH 5.8) via the addition of 0.1N HCl, while HK Kco_Z extracts were buffered to pH 7.2 via the addition of 0.1N NaOH. Trypanosome load in all treatment and control groups was determined at 2, 5 and 24 hour time points by counting live parasites using a Brite-Line hemocytometer.
Determination of bacterial acid production in vivo
Microbial regulation of pH in tsetse’s midgut was determined by feeding teneral GmmApo flies a HI blood meal inoculated with either Sodalis or Kco_Z (5x104 CFU/ml of blood). Additionally, teneral GmmWT flies received the same quantity of Kco_Z. Five days post-bacterial challenge, colonized individuals were administered a meal composed of sucrose (2.5%) and phenol red (0.04%) solubilized in water. Twenty-four hours later, the color of the solution contained in the midgut was determined by incising the fly abdomen and observing the intact gut using a dissecting microscope (Zeiss Discovery) equipped with a digital camera (Zeiss AxioCam MRc 5). Remaining flies were dissected and their guts were harvested, homogenized in 0.85% NaCl, serially diluted and plated onto MM-agar plates (prepared as described in [56]) supplemented with phenol red (0.025 g/L) and sucrose (a 2.5% sucrose solution was spread onto plates immediately prior to applying tsetse gut extracts). CFU per plate was counted manually, and the growth medium was monitored to observe pH-induced changes in color.
Fitness assays
For all fitness assays, GmmWT teneral females and males were infected with Kco_Z during their first blood meal. To determine midgut weight, midguts were dissected from 9 day old GmmWT/Kco_Z and GmmWT females and males (24 h after consuming their last blood meal) and weighed using a Mettler Toledo (AL104) balance. The effect of Kco_Z infection on female fecundity was measured by quantifying the length of three gonotrophic cycles (GC) and by weighing pupal offspring. To measure GC length, GmmWT/Kco_Z and GmmWT females were mated as 5 day old adults and thereafter maintained in individual cages. All females were monitored daily to determine when they deposited larvae, and all deposited larvae were weighed.
The effect of Kco_Z infection on male reproductive fitness was measured by quantifying the mating competitiveness and sperm abundance of individuals that housed the bacterium versus those that did not. Mating competitiveness assays were performed in individual tubular cages (height, 6 cm; diameter 12.7 cm), each of which housed one 5 day old virgin female (fed twice). Subsequently, one age-matched GmmWT/Kco_Z and GmmWT male (also fed twice) was added to each cage. These males were distinguished from one another by removing the proximal tarsus of the right foreleg from one of the individuals. The arena was observed until one of the males had successfully mounted the female, at which point the cage was submerged in ice and the free male identified. To eliminate any bias associated with removal of the foreleg tarsus, the experiment was repeated twice (n = 40 cages per experiment), each time with either GmmWT/Kco_Z or GmmWT males receiving the distinguishing procedure. Sperm abundance was measured by RT-qPCR quantification of sperm-specific dynein intermediate chain (sdic) expression in the reproductive tracts of three and 14 day old (fed twice) virgin GmmWT/Kco_Z and GmmWT males. Absolute sdic transcript abundance was determined by comparing experimental sample cycle threshold (Ct) values to those derived from an sdic internal standard curve.
Sodalis fliC and Wigglesworthia thiC gene specific primers were used to quantify the absolute abundance of these bacteria. This was performed by comparing Sodalis fliC and Wigglesworthia thiC cycle threshold (Ct) values in GmmWT/Kco_Z and GmmWT females and males to those derived from symbiont gene-specific internal standard curves. Because Wigglesworthia and Sodalis can be polyploid [76,77], we normalized symbiont genome copy number to constitutively expressed tsetse gapdh copy number. All RT-qPCR primers are listed in S2 Table. All RT-qPCR assays were carried out in duplicate, and replicates were averaged for each sample. Negative controls were included in all amplification reactions.
Statistical analyses
For trypanosome infection experiments, statistical analyses were carried out using the R software for macOS (version 3.3.2) or GraphPad Prism (v.6). A generalized linear model (GLM) was generated using binomial distribution with a logit transformation of the data. The binary infection status (infected or recovered) was analyzed as a function of the bacterium used to colonize the insects (or its absence). For experiments requiring a pairwise comparison, we performed a Wald test on the individual regression parameter (nature of the bacterium used to colonize) to test its statistical difference. For experiments requiring multiple comparisons, multiple pairwise tests were generated using Tukey contrasts on the generalized linear model (GLM) using glht function of "multcomp" package in R. For the mating assay, under the null hypothesis "Kco_Z does not change the attractiveness of the male", the probability that a female will chose either of the males is p = 0.5. We tested the validity of the null hypothesis by using a Chi-square goodness of fit test. Details of the statistical tests described above are indicated in S1 Dataset. All statistical tests used, and statistical significance between treatments, and treatments and controls, are indicated on the figures or in their corresponding legends. All samples sizes are provided in corresponding figure legends or are indicated graphically as points on dot plots. Biological replication implies distinct groups of flies were collected on different days, and where applicable, treated with distinct treatments (e.g., different bacterial cultures or individual aliquots of trypanosomes).
Supporting information
Kco_Z and Sodalis CFU/gut in 8 day old GmmApo/Kco_Z (Kco_Z) and GmmApo/Sgm (Sodalis) flies prior to (A) challenge with 1x106 blood stream form (BSF) trypanosomes per ml of blood and (B) measuring gut pH in vivo. (C) Guts from GmmApo/Kco_Z (Kco_Z) and GmmApo/Sgm (Sodalis) flies homogenized and plated onto MM-agar plates supplemented with phenol red (0.025 g/L) and sucrose (a 2.5% sucrose solution was spread onto plates immediately prior to applying gut extracts). Plate color reflects bacteria induced changes in pH relative to the empty control. (D) Kco_Z density in the gut of GmmWT/Kco_Z (Kco_Z) flies prior to measuring gut pH in vivo. (E) Kco_Z density in the gut of trypanosome infected (Kco_Z TI) and trypanosome refractory (Kco_Z TR) GmmWT/Kco_Z flies. Measurements were taken at the time infection status was determined (14 days post-challenge). (F) Kco_Z density in the gut of a random sample of GmmWT/Kco_Z (Kco_Z) flies used to determine the bacterium’s impact of tsetse fitness parameters. In panels (A), (B) and (D-F) bacterial load (CFU/gut) was determined via a plating assay described in the Materials and Methods (subsection Microbial infection assays) and in reference [58].
(TIF)
(XLSX)
(XLSX)
(XLSX)
Acknowledgments
We thank Dr. George Dimopoulos (Department of Molecular Microbiology and Immunology, Johns Hopkins Bloomberg School of Public Health) for generously sharing Kco_Z. We thank members of the Aksoy lab for providing critical review of the manuscript. We thank the International Atomic Energy Association (IAEA), under the auspices of a Coordinated Research Project entitled ‘Enhancing tsetse fly refractoriness to trypanosome infection’, and Dr. Peter Takac (Institute of Zoology,Slovak Academy of Science), for providing G. morsitans pupae used in this study.
Data Availability
All relevant data are within the manuscript and its Supporting Information files.
Funding Statement
This work was supported by NIH awards UO1AI115648 (SA), RO1AI051584 (SA), D43TW007391 (SA) and R21AI1011456 (BW), and by the Ambrose-Monell and Bill and Melinda Gates Foundations (SA). The funders had no role in study design, data collection and analysis, decision to publish, or preparation of the manuscript.
References
- 1.Baxter RH, Contet A, Krueger K. Arthropod innate immune systems and vector-borne diseases. Biochemistry 2017;56: 907–918. 10.1021/acs.biochem.6b00870 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 2.Saraiva RG, Kang S, Simões ML, Angleró-Rodríguez YI, Dimopoulos G. Mosquito gut antiparasitic and antiviral immunity. Dev Comp Immunol. 2016;64: 53–64. 10.1016/j.dci.2016.01.015 [DOI] [PubMed] [Google Scholar]
- 3.Aksoy S, Weiss BL, Attardo GM. Trypanosome transmission dynamics in tsetse. Curr Opin Insect Sci. 2014;3: 43–49. 10.1016/j.cois.2014.07.003 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 4.Weiss B, Aksoy S. Microbiome influences on insect host vector competence. Trends Parasitol. 2011;27: 514–522. 10.1016/j.pt.2011.05.001 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 5.Cirimotich CM, Ramirez JL, Dimopoulos G. Native microbiota shape insect vector competence for human pathogens. Cell Host Microbe 2011;10: 307–310. 10.1016/j.chom.2011.09.006 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 6.Narasimhan S, Fikrig E. Tick microbiome: the force within. Trends Parasitol. 2015;31: 315–323. 10.1016/j.pt.2015.03.010 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 7.Song X, Wang M, Dong L, Zhu H, Wang J. PGRP-LD mediates A. stephensi vector competency by regulating homeostasis of microbiota-induced peritrophic matrix synthesis. PLoS Pathog. 2018;14: e1006899 10.1371/journal.ppat.1006899 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 8.Dey R, Joshi AB, Oliveira F, Pereira L, Guimarães-Costa AB, Serafim TD, et al. Gut microbes egested during bites of infected sand flies augment severity of Leishmaniasis via inflammasome-derived IL-1β. Cell Host Microbe 2018;23: 134–143. 10.1016/j.chom.2017.12.002 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 9.Wang J, Weiss BL, Aksoy S. Tsetse fly microbiota: form and function. Front Cell Infect Microbiol. 2013;3: 69 10.3389/fcimb.2013.00069 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 10.Aksoy E, Telleria EL, Echodu R, Wu Y, Okedi LM, Weiss BL, et al. Analysis of multiple tsetse fly populations in Uganda reveals limited diversity and species-specific gut microbiota. Appl Environ Microbiol. 2014;80: 4301–4312. 10.1128/AEM.00079-14 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 11.Geiger A, Fardeau ML, Njiokou F, Joseph M, Asonganyi T, Ollivier B, et al. Bacterial diversity associated with populations of Glossina spp. from Cameroon and distribution within the Campo sleeping sickness focus. Microb Ecol. 2011;62: 632–643. 10.1007/s00248-011-9830-y [DOI] [PubMed] [Google Scholar]
- 12.Lindh JM, Lehane MJ. The tsetse fly Glossina fuscipes fuscipes (Diptera: Glossina) harbours a surprising diversity of bacteria other than symbionts. Antonie Van Leeuwenhoek 2011;99: 711–720. 10.1007/s10482-010-9546-x [DOI] [PubMed] [Google Scholar]
- 13.Benoit JB, Attardo GM, Baumann AA, Michalkova V, Aksoy S. Adenotrophic viviparity in tsetse flies: potential for population control and as an insect model for lactation. Annu Rev Entomol. 2015;60: 351–371. 10.1146/annurev-ento-010814-020834 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 14.Wang J, Wu Y, Yang G, Aksoy S. Interactions between mutualist Wigglesworthia and tsetse peptidoglycan recognition protein (PGRP-LB) influence trypanosome transmission. Proc Natl Acad Sci USA. 2009;106: 12133–12138. 10.1073/pnas.0901226106 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 15.Wang J, Aksoy S. PGRP-LB is a maternally transmitted immune milk protein that influences symbiosis and parasitism in tsetse’s offspring. Proc Natl Acad Sci USA. 2012;109: 10552–10557. 10.1073/pnas.1116431109 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 16.Weiss BL, Wang J, Maltz MA, Wu Y, Aksoy S. Trypanosome infection establishment in the tsetse fly gut is influenced by microbiome-regulated host immune barriers. PLoS Pathog. 2013;9: e1003318 10.1371/journal.ppat.1003318 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 17.Weiss BL, Savage AF, Griffith BC, Wu Y, Aksoy S. The peritrophic matrix mediates differential infection outcomes in the tsetse fly gut following challenge with commensal, pathogenic, and parasitic microbes. J Immunol. 2014;193: 773–782. 10.4049/jimmunol.1400163 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 18.Vigneron A, Aksoy E, Weiss BL, Bing X, Zhao X, Awuoche EO, et al. A fine-tuned vector-parasite dialogue in tsetse’s cardia determines peritrophic matrix integrity and trypanosome transmission success. PLoS Pathog. 2018;14: e1006972 10.1371/journal.ppat.1006972 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 19.Welburn SC, Arnold K, Maudlin I, Gooday GW. Rickettsia-like organisms and chitinase production in relation to transmission of trypanosomes by tsetse flies. Parasitology 1993;107: 141–145. [DOI] [PubMed] [Google Scholar]
- 20.Dale C, Welburn SC. The endosymbionts of tsetse flies: manipulating host-parasite interactions. Int J Parasitol. 2001;31: 628–631. [DOI] [PubMed] [Google Scholar]
- 21.Farikou O, Njiokou F, Mbida Mbida JA, Njitchouang GR, Djeunga HN, Asonganyi T, et al. Tripartite interactions between tsetse flies, Sodalis glossinidius and trypanosomes-an epidemiological approach in two historical human African trypanosomiasis foci in Cameroon. Infect Genet Evol. 2010;10: 115–121. 10.1016/j.meegid.2009.10.008 [DOI] [PubMed] [Google Scholar]
- 22.Soumana IH, Simo G, Njiokou F, Tchicaya B, Abd-Alla AM, Cuny G, et al. The bacterial flora of tsetse fly midgut and its effect on trypanosome transmission. J Invertebr Pathol. 2013;112 Suppl: S89–93. [DOI] [PubMed] [Google Scholar]
- 23.Griffith BC, Weiss BL, Aksoy E, Mireji PO, Auma JE, Wamwiri FN, et al. Analysis of the gut-specific microbiome of field-captured tsetse flies, and its potential relevance to host trypanosome vector competence. 2018;18(Suppl 1): 146. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 24.Boissière A, Tchioffo MT, Bachar D, Abate L, Marie A, Nsango SE, et al. Midgut microbiota of the malaria mosquito vector Anopheles gambiae and interactions with Plasmodium falciparum infection. PLoS Pathog. 2012;8: e1002742 10.1371/journal.ppat.1002742 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 25.Dennison NJ, Jupatanakul N, Dimopoulos G. The mosquito microbiota influences vector competence for human pathogens. Curr Opin Insect Sci. 2014;3: 6–13. 10.1016/j.cois.2014.07.004 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 26.Bahia AC, Dong Y, Blumberg BJ, Mlambo G, Tripathi A, BenMarzouk-Hidalgo OJ, et al. Exploring Anopheles gut bacteria for Plasmodium blocking activity. Environ Microbiol. 2014;16: 2980–2994. 10.1111/1462-2920.12381 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 27.Ramirez JL, Short SM, Bahia AC, Saraiva RG, Dong Y, Kang S, et al. Chromobacterium Csp_P reduces malaria and dengue infection in vector mosquitoes and has entomopathogenic and in vitro anti-pathogen activities. PLoS Pathog. 2014;10: e1004398 10.1371/journal.ppat.1004398 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 28.Brady C, Cleenwerck I, Venter S, Coutinho T, De Vos P. Taxonomic evaluation of the genus Enterobacter based on multilocus sequence analysis (MLSA): proposal to reclassify E. nimipressuralis and E. amnigenus into Lelliottia gen. nov. as Lelliottia nimipressuralis comb. nov. and Lelliottia amnigena comb. nov., respectively, E. gergoviae and E. pyrinus into Pluralibacter gen. nov. as Pluralibacter gergoviae comb. nov. and Pluralibacter pyrinus comb. nov., respectively, E. cowanii, E. radicincitans, E. oryzae and E. arachidis into Kosakonia gen. nov. as Kosakonia cowanii comb. nov., Kosakonia radicincitans comb. nov., Kosakonia oryzae comb. nov. and Kosakonia arachidis comb. nov., respectively, and E. turicensis, E. helveticus and E. pulveris into Cronobacter as Cronobacter zurichensis nom. nov., Cronobacter helveticus comb. nov. and Cronobacter pulveris comb. nov., respectively, and emended description of the genera Enterobacter and Cronobacter. Syst Appl Microbiol. 2013;36: 309–319. 10.1016/j.syapm.2013.03.005 [DOI] [PubMed] [Google Scholar]
- 29.Cirimotich CM, Dong Y, Clayton AM, Sandiford SL, Souza-Neto JA, Mulenga M, Dimopoulos G. Natural microbe-mediated refractoriness to Plasmodium infection in Anopheles gambiae. Science 2011;332: 855–858. 10.1126/science.1201618 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 30.Dennison NJ, Saraiva RG, Cirimotich CM, Mlambo G, Mongodin EF, Dimopoulos G. Functional genomic analyses of Enterobacter, Anopheles and Plasmodium reciprocal interactions that impact vector competence. Malar J. 2016;15: 425 10.1186/s12936-016-1468-2 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 31.MacLeod ET, Maudlin I, Darby AC, Welburn SC. Antioxidants promote establishment of trypanosome infections in tsetse. Parasitology 2007;134: 827–831. 10.1017/S0031182007002247 [DOI] [PubMed] [Google Scholar]
- 32.Ridgley EL, Xiong ZH, Ruben L. Reactive oxygen species activate a Ca2+-dependent cell death pathway in the unicellular organism Trypanosoma brucei brucei. Biochem J. 1999;340: 33–40. [PMC free article] [PubMed] [Google Scholar]
- 33.Witzel K, Üstün S, Schreiner M, Grosch R, Börnke F, Ruppel S. A proteomic approach suggests unbalanced proteasome functioning induced by the growth-promoting bacterium Kosakonia radicincitans in Arabidopsis. Front Plant Sci. 2017;8: 661 10.3389/fpls.2017.00661 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 34.Chin CFS, Furuya Y, Zainudin MHM, Ramli N, Hassan MA, Tashiro Y, et al. Novel multifunctional plant growth-promoting bacteria in co-compost of palm oil industry waste. J Biosci Bioeng. 2017;124: 506–513. 10.1016/j.jbiosc.2017.05.016 [DOI] [PubMed] [Google Scholar]
- 35.Chakdar H, Dastager SG, Khire JM, Rane D, Dharne MS. Characterization of mineral phosphate solubilizing and plant growth promoting bacteria from termite soil of arid region. 3 Biotech. 2018;8: 463 10.1007/s13205-018-1488-4 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 36.Podleśny M, Jarocki P, Wyrostek J, Czernecki T, Kucharska J, Nowak A, et al. Enterobacter sp. LU1 as a novel succinic acid producer—co-utilization of glycerol and lactose. Microb Biotechnol. 2017;10: 492–501. 10.1111/1751-7915.12458 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 37.Fischer CN, Trautman EP, Crawford JM, Stabb EV, Handelsman J, Broderick NA. Metabolite exchange between microbiome members produces compounds that influence Drosophila behavior. Elife 2017;6. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 38.Goldford JE, Lu N, Bajić D, Estrela S, Tikhonov M, Sanchez-Gorostiaga A, et al. Emergent simplicity in microbial community assembly. Science 2018;361: 469–474. 10.1126/science.aat1168 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 39.Buffie CG, Pamer EG. Microbiota-mediated colonization resistance against intestinal pathogens. Nat Rev Immunol. 2013;13: 790–801. 10.1038/nri3535 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 40.Neal-McKinney JM, Lu X, Duong T, Larson CL, Call DR, Shah DH, et al. Production of organic acids by probiotic lactobacilli can be used to reduce pathogen load in poultry. PLoS One 2012;7: e43928 10.1371/journal.pone.0043928 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 41.Nolan DP, Voorheis HP. Hydrogen ion gradients across the mitochondrial, endosomal and plasma membranes in bloodstream forms of Trypanosoma brucei solving the three-compartment problem. Eur J Biochem. 2000;267: 4601–4614. [DOI] [PubMed] [Google Scholar]
- 42.Zilberstein D, Shapira M. The role of pH and temperature in the development of Leishmania parasites. Annu Rev Microbiol. 1994;48: 449–470. 10.1146/annurev.mi.48.100194.002313 [DOI] [PubMed] [Google Scholar]
- 43.Harley JM, Wilson AJ. Comparison between Glossina morsitans, G. pallidipes and G. fuscipes as vectors of trypanosomes of the Trypanosoma congolense group: the proportions infected experimentally and the numbers of infective organisms extruded during feeding. Ann Trop Med Parasitol. 1968;62: 178–187. [DOI] [PubMed] [Google Scholar]
- 44.Moloo SK, Sabwa CL, Kabata JM. Vector competence of Glossina pallidipes and G. morsitans centralis for Trypanosoma vivax, T. congolense and T. b. brucei. Acta Trop. 1992;51: 271–280. [DOI] [PubMed] [Google Scholar]
- 45.Aksoy E, Vigneron A, Bing X, Zhao X, O’Neill M, Wu YN, et al. Mammalian African trypanosome VSG coat enhances tsetse’s vector competence. Proc Natl Acad Sci USA. 2016;113: 6961–6966. 10.1073/pnas.1600304113 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 46.Nurminsky DI, Nurminskaya MV, De Aguiar D, Hartl DL. Selective sweep of a newly evolved sperm-specific gene in Drosophila. Nature 1998;396: 572–575. 10.1038/25126 [DOI] [PubMed] [Google Scholar]
- 47.Yeh SD, Do T, Chan C, Cordova A, Carranza F, Yamamoto EA, et al. Functional evidence that a recently evolved Drosophila sperm-specific gene boosts sperm competition. Proc Natl Acad Sci USA. 2012;109: 2043–2048. 10.1073/pnas.1121327109 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 48.Michalkova V, Benoit JB, Weiss BL, Attardo GM, Aksoy S. Obligate symbiont-generated vitamin B6 is critical to maintain proline homeostasis and fecundity in tsetse flies. Appl Environ Microbiol. 2014;80: 5844–5853. 10.1128/AEM.01150-14 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 49.Vigneron A, Weiss BL. Role of the microbiota during development of the arthropod vector immune system In: Wikel S, editor. Arthropod vector: controller of disease transmission, vol. 1: vector microbiome and innate immunity of arthropods. Elsevier; 2017. pp. 161–169. [Google Scholar]
- 50.Dale C, Moran NA. Molecular interactions between bacterial symbionts and their hosts. Cell. 2006;126: 453–465. 10.1016/j.cell.2006.07.014 [DOI] [PubMed] [Google Scholar]
- 51.Raymond B, Young JC, Pallett M, Endres RG, Clements A, Frankel G. Subversion of trafficking, apoptosis, and innate immunity by type III secretion system effectors. Trends Microbiol. 2013;21: 430–441. 10.1016/j.tim.2013.06.008 [DOI] [PubMed] [Google Scholar]
- 52.Ketterman AJ, Saisawang C, Wongsantichon J. Insect glutathione transferases. Drug Metab Rev. 2011;43: 253–265 10.3109/03602532.2011.552911 [DOI] [PubMed] [Google Scholar]
- 53.Pedrini N, Ortiz-Urquiza A, Huarte-Bonnet C, Fan Y, Juárez MP, Keyhani NO. Tenebrionid secretions and a fungal benzoquinone oxidoreductase form competing components of an arms race between a host and pathogen. Proc Natl Acad Sci USA. 2015;112: E3651–3660. 10.1073/pnas.1504552112 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 54.Hu Y, Aksoy S. An antimicrobial peptide with trypanocidal activity characterized from Glossina morsitans morsitans. Insect Biochem Mol Biol. 2005;35: 105–115. 10.1016/j.ibmb.2004.10.007 [DOI] [PubMed] [Google Scholar]
- 55.Toh H, Weiss BL, Perkin SA, Yamashita A, Oshima K, Hattori M, et al. Massive genome erosion and functional adaptations provide insights into the symbiotic lifestyle of Sodalis glossinidius in the tsetse host. Genome Res. 2006;16: 149–156. 10.1101/gr.4106106 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 56.Weiss BL, Wu Y, Schwank JJ, Tolwinski NS, Aksoy S. An insect symbiosis is influenced by bacterium-specific polymorphisms in outer-membrane protein A. Proc Natl Acad Sci USA. 2008;105): 15088–15093. 10.1073/pnas.0805666105 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 57.Dale C, Young SA, Haydon DT, Welburn SC. The insect endosymbiont Sodalis glossinidius utilizes a type III secretion system for cell invasion. Proc Natl Acad Sci USA. 2001;98: 1883–1888. 10.1073/pnas.98.4.1883 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 58.Maltz MA, Weiss BL, O’Neill M, Wu Y, Aksoy S. OmpA-mediated biofilm formation is essential for the commensal bacterium Sodalis glossinidius to colonize the tsetse fly gut. Appl Environ Microbiol. 2012;78: 7760–7768. 10.1128/AEM.01858-12 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 59.Ratzke C, Denk J, Gore J. Ecological suicide in microbes. Nat Ecol Evol. 2018;2: 867–872. 10.1038/s41559-018-0535-1 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 60.Celiker H, Gore J. Cellular cooperation: insights from microbes. Trends Cell Biol. 2013;23: 9–15. 10.1016/j.tcb.2012.08.010 [DOI] [PubMed] [Google Scholar]
- 61.Ratzke C, Gore J. Modifying and reacting to the environmental pH can drive bacterial interactions. PLoS Biol. 2018b;16: e2004248. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 62.Liniger M, Acosta-Serrano A, Van Den Abbeele J, Kunz Renggli C, Brun R, Englund PT, et al. Cleavage of trypanosome surface glycoproteins by alkaline trypsin-like enzyme(s) in the midgut of Glossina morsitans. Int J Parasitol. 2003;33: 1319–1328. [DOI] [PubMed] [Google Scholar]
- 63.Rosenzweig D, Smith D, Opperdoes F, Stern S, Olafson RW, Zilberstein D. Retooling Leishmania metabolism: from sand fly gut to human macrophage. FASEB J. 2008;22: 590–602. 10.1096/fj.07-9254com [DOI] [PubMed] [Google Scholar]
- 64.del Pilar Corena M, VanEkeris L, Salazar MI, Bowers D, Fiedler MM, Silverman D, et al. Carbonic anhydrase in the adult mosquito midgut. J Exp Biol. 2005;208: 3263–3273. 10.1242/jeb.01739 [DOI] [PubMed] [Google Scholar]
- 65.Aksoy S O’Neill SL, Maudlin I, Dale C, Robinson AS. Prospects for control of African trypanosomiasis by tsetse vector manipulation. Trends Parasitol. 2001;17: 29–35. [DOI] [PubMed] [Google Scholar]
- 66.Vreysen MJ, Seck MT, Sall B, Bouyer J. Tsetse flies: their biology and control using area-wide integrated pest management approaches. J Invertebr Pathol. 2013;112 Suppl: S15–25. [DOI] [PubMed] [Google Scholar]
- 67.Percoma L, Sow A, Pagabeleguem S, Dicko AH, Serdebéogo O, Ouédraogo M, et al. Impact of an integrated control campaign on tsetse populations in Burkina Faso. Parasit Vectors. 2018;11: 270 10.1186/s13071-017-2609-3 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 68.McGraw EA O’Neill SL. Beyond insecticides: new thinking on an ancient problem. Nat Rev Microbiol. 2013;11: 181–193. 10.1038/nrmicro2968 [DOI] [PubMed] [Google Scholar]
- 69.Vreysen MJ, Saleh KM, Ali MY, Abdulla AM, Zhu ZR, Juma KG, et al. Glossina austeni (Diptera: Glossinidae) eradicated on the island of Unguja, Zanzibar, using the sterile insect technique. J Econ Entomol. 2000;93: 123–135. [DOI] [PubMed] [Google Scholar]
- 70.Bouyer J. Does isometamidium chloride treatment protect tsetse flies from trypanosome infections during SIT campaigns? Med Vet Entomol. 2008;22: 140–143. 10.1111/j.1365-2915.2008.00727.x [DOI] [PubMed] [Google Scholar]
- 71.de Beer CJ, Moyaba P, Boikanyo SN, Majatladi D, Yamada H, Venter GJ, et al. Evaluation of radiation sensitivity and mating performance of Glossina brevipalpis males. PLoS Negl Trop Dis. 2017;11: e0005473 10.1371/journal.pntd.0005473 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 72.Moloo SK. An artificial feeding technique for Glossina. Parasitology. 1971;63: 507–512. [DOI] [PubMed] [Google Scholar]
- 73.Weiss BL, Maltz M, Aksoy S. Obligate symbionts activate immune system development in the tsetse fly. J Immunol. 2012;188: 3395–3403. 10.4049/jimmunol.1103691 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 74.Dale C, Maudlin I. Sodalis gen. nov. and Sodalis glossinidius sp. nov., a microaerophilic secondary endosymbiont of the tsetse fly Glossina morsitans morsitans. Int J Syst Bacteriol. 1999;49 Pt 1: 267–275. [DOI] [PubMed] [Google Scholar]
- 75.Nehme NT, Liégeois S, Kele B, Giammarinaro P, Pradel E, Hoffmann JA, et al. A model of bacterial intestinal infections in Drosophila melanogaster. PLoS Pathog. 2007;3: e173 10.1371/journal.ppat.0030173 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 76.Rio RV, Wu YN, Filardo G, Aksoy S. Dynamics of multiple symbiont density regulation during host development: tsetse fly and its microbial flora. Proc Biol Sci. 2006;273: 805–814. 10.1098/rspb.2005.3399 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 77.Weiss BL, Mouchotte R, Rio RV, Wu YN, Wu Z, Heddi A, Aksoy S. Interspecific transfer of bacterial endosymbionts between tsetse fly species: infection establishment and effect on host fitness. Appl Environ Microbiol. 2006;72: 7013–7021. 10.1128/AEM.01507-06 [DOI] [PMC free article] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.
Supplementary Materials
Kco_Z and Sodalis CFU/gut in 8 day old GmmApo/Kco_Z (Kco_Z) and GmmApo/Sgm (Sodalis) flies prior to (A) challenge with 1x106 blood stream form (BSF) trypanosomes per ml of blood and (B) measuring gut pH in vivo. (C) Guts from GmmApo/Kco_Z (Kco_Z) and GmmApo/Sgm (Sodalis) flies homogenized and plated onto MM-agar plates supplemented with phenol red (0.025 g/L) and sucrose (a 2.5% sucrose solution was spread onto plates immediately prior to applying gut extracts). Plate color reflects bacteria induced changes in pH relative to the empty control. (D) Kco_Z density in the gut of GmmWT/Kco_Z (Kco_Z) flies prior to measuring gut pH in vivo. (E) Kco_Z density in the gut of trypanosome infected (Kco_Z TI) and trypanosome refractory (Kco_Z TR) GmmWT/Kco_Z flies. Measurements were taken at the time infection status was determined (14 days post-challenge). (F) Kco_Z density in the gut of a random sample of GmmWT/Kco_Z (Kco_Z) flies used to determine the bacterium’s impact of tsetse fitness parameters. In panels (A), (B) and (D-F) bacterial load (CFU/gut) was determined via a plating assay described in the Materials and Methods (subsection Microbial infection assays) and in reference [58].
(TIF)
(XLSX)
(XLSX)
(XLSX)
Data Availability Statement
All relevant data are within the manuscript and its Supporting Information files.