

Abstract
Worldwide use of anticoagulant rodenticides (ARs) for rodents control has frequently led to secondary poisoning of non-target animals, especially raptors. In spite of the occurrence of many incidents of primary or secondary AR-exposure and poisoning of non-target animals, these incidents have been reported only for individual countries, and there has been no comprehensive worldwide study or review. Furthermore, the AR exposure pathway in raptors has not yet been clearly identified. The aim of this review is therefore to comprehensively analyze the global incidence of primary and secondary AR-exposure in non-target animals, and to explore the exposure pathways. We reviewed the published literature, which reported AR residues in the non-target animals between 1998 and 2015, indicated that various raptor species had over 60% AR- detection rate and have a risk of AR poisoning. According to several papers studied on diets of raptor species, although rodents are the most common diets of raptors, some raptor species prey mainly on non-rodents. Therefore, preying on targeted rodents does not necessarily explain all causes of secondary AR-exposure of raptors. Since AR residue-detection was also reported in non-target mammals, birds, reptiles and invertebrates, which are the dominant prey of some raptors, AR residues in these animals, as well as in target rodents, could be the exposure source of ARs to raptors.
Keywords: anticoagulant rodenticide, comprehensive review, non-target animal, raptor, residue
Worldwide use of anticoagulant rodenticides (ARs) for vertebrate pest control has frequently led to the unintentional exposures of non-target animals, especially raptors, to these poisons. Recently, more than 420 birds, including 46 bald eagles (Haliaeetus leucocephalus), died because of a rat-eradication program on an Alaskan island [2]. Reporting that more than 130 dead raptors found in and around Vancouver, Canada, and virtually 100% of the owls and the hawks in this group, had AR residues in their livers, the Nature News article “killing rats is killing birds” had a strong impact on the world [29]. The occurrence of AR poisoning in raptors is related to many factors, such as the exposure pathway, the degree of ARs inhibiting the target molecule (vitamin K 2,3-epoxide reductase, VKOR), and AR metabolism by cytochrome P450 (CYP).
Although it is thought that raptors are sensitive to ARs, the mechanism for this sensitivity has not yet been revealed. Several toxicokinetic of avian species have been studied. Compared to mammals, eastern screech-owls (Megascops asio) have a long elimination half-life of diphacinone in the liver [39]. Furthermore, owls have very low CYP-dependent warfarin metabolic activity compared to rats and other avian species [62]. These facts imply that owls have a limited ability to detoxify ARs. However, toxicokinetics of the other raptor species has been rarely studied.
In spite of frequent incidents of primary or secondary AR-exposure and poisoning in non-target animals, including predatory mammals and birds (especially raptors), these incidents have been reported only in individual countries and there has been no comprehensive worldwide study or review. Therefore, we comprehensively reviewed and analyzed the published literature on AR-exposure occurrence based on the kind of ARs, the country type, and the animal groups. In addition, this review discussed diets of raptors. Some possible exposure pathways in addition to the target rodents were also discussed.
The chemical structures of nine typical anticoagulant rodenticides (ARs) are shown in Fig. 1. ARs are classified into two classes: 4-hydroxycoumarin derivatives (coumarin: warfarin, coumatetralyl, brodifacoum, bromadiolone, difenacoum, difethialone, and flocoumafen), and 1,3-indanedione derivatives (indanedione: chlorophacinone and diphacinone are the most commonly used examples).
Fig. 1.
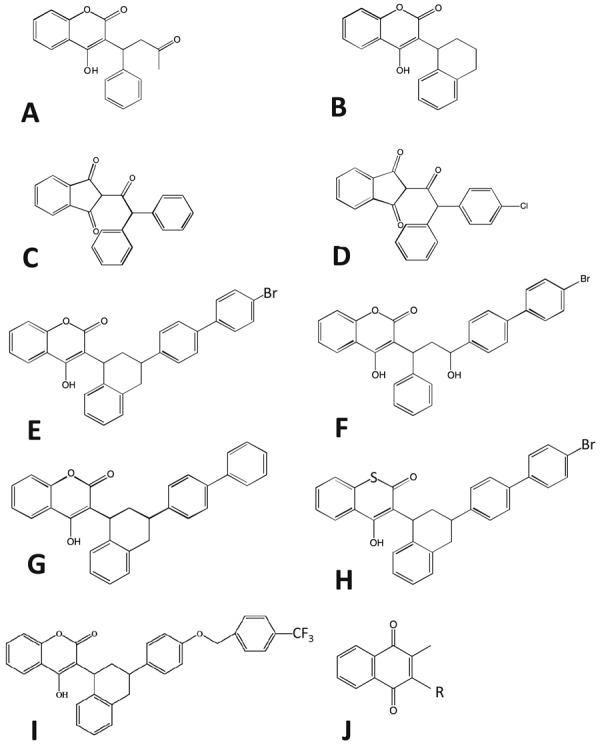
Chemical structure of nine typical ARs (A to I) and vitamin K (J). First-generation anticoagulant rodenticides (FGARs) are represented by the coumarin (warfarin, A, and coumatetralyl, B) and indanedione (diphacinone, C, and chlorophacinone, D) rodenticides. Examples of second-generation anticoagulant rodenticides (SGARs) are brodifacoum (E), bromadiolone (F), difenacoum (G), difethialone (H) and flocoumafen (I). The main chain structures of ARs are similar to that of vitamin K (J).
By the early 1950s, warfarin was being used as a pesticide to control rats and mice [41]. Warfarin and other early ARs, such as coumatetralyl, chlorophacinone, and diphacinone, are called first-generation ARs (FGARs). Multiple ingestions of these compounds are required to cause death in rodents. Since then, FGAR-resistant rats and mice have appeared, so the more potent second-generation ARs (SGARs), such as brodifacoum, bromadiolone, difenacoum, difethialone, and flocoumafen, were developed [41]. SGARs require only a single ingestion to cause death in the targeted rodents. For mice, SGARs have a longer T1/2 than FGARs (Table 1): in the plasma, T1/2 for SGARs is 20.4–91.7 days, while that for FGARs is 0.52–14.9 days; and in the liver, T1/2 for SGARs is 28.1–307.4 days, whereas that for FGARs is 15.8–66.8 days. In addition to longer T1/2, SGARs have a lower LD50 than FGARs (Table 2): LD50 for SGARs and FGARs are 0.4–1.75 and 20.5–1,000 mg/kg in mice; 0.35–0.84 and 11–323 mg/kg in rats; 0.25–8.1 and 0.88–50 mg/kg in dogs; 3.15 and 942 mg/kg in chickens; 0.26–138 and 258–2,150 mg/kg in northern bobwhites; 10 and >100 mg/kg in ring-necked pheasants; and 4.6 and 620–3,158 mg/kg in mallards. In Australian harriers, LD50 for brodifacoum (SGARs) is 10 mg/kg; and in American kestrels, LD50 for diphacinone (FGARs) is 97 mg/kg. These longer T1/2 and lower LD50 for SGARs imply that SGARs are more toxic than FGARs.
Table 1. Eliminated half-life, T1/2 (days) for FGARs (warfarin, coumatetralyl, chlorophacinone and diphacinone) and SGARs (brodifacoum, bromadiolone, difenacoum, difethialone and flocoumafen) in animals. Modified from [12, 18, 20, 39, 40, 57].
| FGARs | SGARs | ||||||||
|---|---|---|---|---|---|---|---|---|---|
| Warfarin | Couma-tetralyl | Dipha-cinone | Chloro-phacinone | Brodi-facoum | Broma-diolone | Difen-acoum | Difeti-alone | Flocou-mafen | |
| T1/2 in the plasma | |||||||||
| Mousea) | 14.9 | 0.52 | - | 11.7 | 91.7 | 33.3 | 20.4 | 38.9 | 26.6 |
| T1/2 in the liver | |||||||||
| Mousea) | 66.8 | 15.8 | - | 35.4 | 307.4 | 28.1 | 61.8 | 28.5 | 93.8 |
| Rat | - | - | 3 | - | - | - | - | - | - |
| Pig | - | - | 5.43 | - | - | - | - | - | - |
| Screech owl | - | - | 11.7 | - | - | - | - | - | - |
a) dose=half of LD50.
Table 2. Median lethal dose, LD50 (mg/kg) for FGARs (warfarin, coumatetralyl, chlorophacinone and diphacinone) and SGARs (brodifacoum, bromadiolone, difenacoum, difethialone and flocoumafen) in animals. Modified from [12, 18, 20, 39, 40, 57].
| FGARs | SGARs | ||||||||
|---|---|---|---|---|---|---|---|---|---|
| Warfarin | Couma-tetralyl | Dipha-cinone | Chloro-phacinone | Brodi-facoum | Broma-diolone | Difen-acoum | Difeti-alone | Flocou-mafen | |
| Mouse | 374 | <1,000 | 141–340 | 20.5 | 0.4 | 1.75 | 0.8 | 1.29 | 0.8 |
| Rat | 14–323 | - | 30 | 11 | 0.35–0.5 | 0.56–0.84 | - | 0.55 | - |
| Dog | 20–50 | - | 0.88–15 | - | 0.25–1.0 | 8.1 | - | - | - |
| Cat | 2.5–20 | - | 5–15 | - | <25 | >25 | - | - | - |
| Chicken | 942 | - | - | - | 3.15 | - | - | - | - |
| Northern bobwhite | >2,150 | - | 2,014 | 258 | - | 138 | - | 0.26 | - |
| Ring-necked pheasant | - | - | - | >100 | 10 | - | - | - | - |
| Mallard | 620 | - | 3,158 | - | 4.6 | - | - | - | - |
| American kestrel | - | - | 97 | - | - | - | - | - | - |
| Australasian harrier | - | - | - | - | 10 | - | - | - | - |
AR EXPOSURE GLOBALLY
From 1998 to 2015, altogether 30 papers were published reporting primary or secondary exposure and poisoning by ARs in non-target animals. Of these, 19 papers report poisoning of raptors. There are six publications from the U.S.A. [9, 42, 45, 48,49,50], three from Canada [1, 18, 55], nine from the U.K. [7, 19, 31, 46, 47, 58,59,60,61], two from France [13, 25], three from Spain [28, 43, 44], two from Denmark [5, 11], one from Norway [26], and four from New Zealand (NZ) [6, 8, 14, 35]. These reports and the proposed exposure pathways are summarized below.
Presence of ARs in non-target animals
According to the literatures published between 1998 and 2015, totally 2,694 out of 4,891 (55%) individual non-target animals have been found to have a residual accumulation of ARs in their livers (Table 3A). Because the kinds of analyzed rodenticides were different depending on the papers, the number of analysis was different for each compound. Brodifacoum was detected in 31% (n=1,465 out of 4,790) of non-target animals, bromadiolone in 30% (n=1,346 out of 4,513), and difenacoum in 26% (n=1,048 out of 4,001). The other compounds were detected in less than 10% of the animals: flocoumafen in 5.7% (n=175 out of 3,077), difethialone in 5.0% (n=101 out of 2,035), chlorofacinone in 5.6% (n=113 out of 2,013), diphacinone in 5.0% (n=99 out of 1,972), coumatetralyl in 4.5%, (n=108 out of 2,391), and warfarin in 1.0% (n=27 out of 2,639). Some animals had more than two types of ARs in their liver.
Table 3. Detection rates and the numbers of non-target animals in which ARs have been detected in their liver, classified based on (A) type of AR, (B) country and (C) animal groups. Altogether, there were 2,694 out of 4,891 individuals (55%) of non-target animals have been reported to have AR residues in their livers between 1981 and 2013 [1, 5,6,7,8,9, 11, 13, 14, 18, 19, 25, 26, 28, 31, 35, 42,43,44,45,46,47,48,49,50, 55, 58,59,60,61].
| (A) | FGARs | SGARs | |||||||
|---|---|---|---|---|---|---|---|---|---|
| Warfarin | Couma-tetralyl | Dipha-cinone | Chloro-phacinone | Brodi-facoum | Broma-diolone | Difen-acoum | Difeti-alone | Flocou-mafen | |
| Detection rate (%) | 1.0 | 4.5 | 5.0 | 5.6 | 31.0 | 30.0 | 26.0 | 5.0 | 5.7 |
| The number of detection | 27 | 108 | 99 | 113 | 1,465 | 1,346 | 1,048 | 101 | 175 |
| The number of analysis | 2,639 | 2,391 | 1,972 | 2,013 | 4,790 | 4,513 | 4,001 | 2,035 | 3,077 |
| (B) | Denmark | Canada | NZ | U.S.A. | Norway | Spain | U.K. | France | |
| Detection rate (%) | 93 | 67 | 59 | 58 | 53 | 51 | 44 | 23 | |
| The number of detection | 523 | 241 | 171 | 474 | 16 | 437 | 790 | 42 | |
| The number of analysis | 560 | 362 | 288 | 812 | 30 | 849 | 1,809 | 181 | |
| (C) | Carnivora | Raptors | Mammals excluding Carnivora | Birds excluding raptors | Reptiles | ||||
| Detection rate (%) | 56.8 | 56.6 | 52.1 | 52.1 | 50.0 | ||||
| The number of detection | 382 | 1,892 | 212 | 207 | 1 | ||||
| The number of analysis | 672 | 3,345 | 407 | 397 | 2 | ||||
High detection-rates of brodifacoum, bromadiolone and difenacoum reflect the relative frequency of use of these SGARs and differences in tissue T1/2 values between compounds. Because brodifacoum has longer T1/2 compared with the other ARs (Table 1), it would be expected that brodifacoum is detected for a long time and is over-reported relative to the amount of use compared with the other compounds. Because of the high toxicity of SGARs, as indicated by their longer T1/2 values in the liver and lower LD50 values relative to FGARs (Tables 1 and 2), the potential adverse effects of SGARs on non-target animals are a particular cause for concern.
AR exposure of non-target animals in each country
In terms of occurrence in each country (Table 3B), residues of rodenticides were detected in 523 out of 560 animals (93%) in Denmark, 241 out of 362 animals (67%) in Canada, 171 out of 288 animals (59%) in NZ, 474 of 812 animals (58%) in the U.S.A., 16 out of 30 animals (53%) in Norway, 437 out of 849 animals (52%) in Spain, 790 out of 1809 animals (44%) in the U.K., and 42 out of 181 animals (23%) in France. Although these rates seem to imply the degree of AR exposure in each country, it is difficult to compare the percentage with each country, because these percentages probably reflect residue detection limits as well as the relative frequency of use. The minimum detectable amounts of individual ARs described in the study of Denmark and Canada were lower than those of France (e.g. 2–5 to 70–80 µg/kg for brodifacoum respectively) [1, 5, 11, 13, 18, 25, 55]. On the other hand, the detection limits for brodifacoum were 10–50, 2–20, 5, 1–6, and 1.4–50 µg/kg of NZ [8, 14, 35], the U.S.A. [9, 42, 45, 48,49,50], Norway [26], Spain [28, 43, 44], and the U.K. [7, 19, 31, 46, 47, 58,59,60], respectively. Therefore, high (or low) sensitivity of detection seemingly not always cause over (or under)-estimation, but in some cases might affect accounting of AR exposure. We would like to note that, although AR detection rates were various in eight countries (from 23 to 93%), AR exposure of non-target animals might certainly occur in all eight countries.
The largest number of species in which AR residues have been found in one country (Table 4) was 39 (Spain), followed by 31 (the U.S.A.), 21 (NZ), 13 (Denmark), 11 (the U.K.), ten (France), five (Canada), and two (Norway). In all of these countries except NZ, raptors constituted the majority of exposed species: 18 in Spain, 13 in the U.S.A., 11 in Denmark, seven in the U.K., four in France and Canada, and two in Norway. In NZ, the majority of exposed species were in the group “birds excluding raptors” (16 species). Various species (totally 94 species), especially raptors (34 species), were exposed to ARs.
Table 4. The number of non-target animal species in which ARs have been detected in the liver. References are given in Table 5.
| U.S.A. | Canada | U.K. | France | Spain | Denmark | Norway | NZ | Totala) | |
|---|---|---|---|---|---|---|---|---|---|
| Raptors | 13 | 4 | 7 | 4 | 18 | 11 | 2 | 2 | 34 |
| Other birds | 9 | 1 | - | 2 | 9 | - | - | 16 | 34 |
| Carnivora | 5 | - | 3 | 4 | 8 | 2 | - | - | 15 |
| Cetartiodactyla | 1 | - | - | - | - | - | - | 2 | 3 |
| Erinaceomorpha | - | - | 1 | - | 2 | - | - | - | 2 |
| Rodentia | 2 | - | - | - | - | - | - | - | 2 |
| Lagomorpha | - | - | - | - | 1 | - | - | - | 1 |
| Chiroptera | - | - | - | - | - | - | - | 1 | 1 |
| Marsupialia | 1 | - | - | - | - | - | - | - | 1 |
| Reptiles | - | - | - | - | 1 | - | - | - | 1 |
| Total | 31 | 5 | 11 | 10 | 39 | 13 | 2 | 21 | 94 |
a) This is not necessarily the sum of the values for each country, because some species were reported in several countries.
AR residues in non-target animals have only been reported in these eight countries. This is probably because this research is implemented only in North America, Europe and NZ. However, ARs have been frequently used worldwide [21]. Global AR market data is difficult to obtain because of confidential business information, but estimates are described as hundreds of millions dollars annually in the U.S.A. and European countries [41]. Primary and secondary AR exposure of non-target animals may indeed occur all over the world, so increased surveys are needed worldwide to determine the extent of the problem.
Classification of animal species exposed to ARs
Table 3C shows that the accumulation of AR residues was detected in 382 out of 672 individuals of Carnivora (56.8%), 1,892 out of 3,345 raptors (56.6%), 212 out of 407 mammals excluding Carnivora (52.1%), 207 out of 397 birds excluding raptors (52.1%), and 1 out of 2 reptiles (50%). Percentages of Carnivora, raptors and reptiles are presumed to be secondary exposure degree, and those of mammals excluding Carnivora and birds excluding raptors seem to be primary exposure degree.
Although secondary exposure seems to occur in Carnivora and raptors at a comparable frequency, secondary poisoning should be considered to occur in raptors frequently relative to Carnivora. Critical liver SGAR concentrations associated with hemorrhaging and mortality have not been defined for most raptor species. However, the potentially lethal range for SGARs in raptors has been described as >100–200 µg/kg [5, 55, 60]. On the other hand, the lethal concentration of SGARs in Carnivora livers have been reported that brodifacoum of 700 µg/kg was detected in stoat and weasel, bromadiolone of 230 µg/kg accumulated in stoat, and difenacoum of 1,400 µg/kg was measured in polecats [31, 46]. Because most of the cited references in the current review did not mention AR concentrations of individual animals, we could not calculate AR-poisoning rate of individual species. However, raptors are presumed to be poisoned by ARs frequently rather than Carnivora, because of the low lethal range for SGAR residues in raptors. Moreover, screech owls have longer T1/2 for diphacinone compared with rats and pigs (Table 1), and owls have very low warfarin metabolic activity relative to rats and other avian species [62]. The adverse effect of AR exposure on raptors is of interest.
The high AR-exposed rate was reported in various raptor species rather than Carnivora species. The number of species ARs detected in more than 60% of individuals was 11 in raptors, and it comprised 48% of 23 raptor species that the number of analysis was more than nine individuals (Note: We did not include the species whose analyzed individual sample size was less than 8. This was to avoid the over-estimation e.g. 100% of detection rate such a case that the one individual detected from the one individual analyzed.). In contrast, 3 species, which had over 60% AR-detection rate, composed 27% of 11 Carnivora species that AR residue was examined in more than nine individuals. AR exposure to extensive raptor species implies that various kinds of raptors have a risk of AR poisoning.
SECONDARY EXPOSURE TO AR IN RAPTORS
Frequently reported raptor species
Of the 39 raptor species analyzed, 34 species were reported to have AR accumulation in their liver, and 17 species had more than 60% detection rate of ARs (Table 5): two out of two turkey vultures (Cathartes aura; 100%), one out of one short toed snake-eagle (Circaetus gallicus; 100%), three out of three marsh harriers (Circus aeruginosus; 100%), five out of five moreporks (Ninox novaeseelandiae; 100%), 15 out of 17 little owls (Athene noctua; 88%), 33 out of 39 Eurasian eagle owls (Bubo bubo; 85%), 20 out of 24 bald eagles (Haliaeetus leucocephalus; 83%), five out of six short-eared owls (Asio flammeus; 83%), 26 out of 32 rough-legged buzzards (Buteo lagopus; 81%), 116 out of 145 sparrowhawks (Accipiter nisus; 80%), 108 out of 138 red kites (Milvus milvus; 78%), 62 out of 80 long-eared owls (Asio otus; 78%), 154 out of 206 kestrels (Falco tinnunculus; 75%), 26 out of 38 barred owls (Strix varia; 68%), 14 out of 22 golden eagles (Aquila chrysaetos; 64%), and 192 out of 308 great horned owls (Bubo virginianus; 62%).
Table 5. Presence of ARs in animal species reported in various countries. AR concentrations (µg/kg) given are in the form minimum–maximum, or mean ± S.D. (values marled with an asterisk are medians, with a sharp are standard error, S. E. and † means prevalence of any ARs); N gives the number of analysis; n+ gives the number of individuals with detectable residues; % means detection rate; and n is the total number of individuals in which each AR has been detected.
| Species | Year | Country | N | n+ | % | Warfa- rin |
Couma- tetralyl |
Dipha- cinone |
Chloro- phacinone |
Brodi- facoum |
Broma- diolone |
Difen- acoum |
Difeti- alone |
Flocou- mafen |
References |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Raptors (39 species analyzed) | |||||||||||||||
| Turkey vulture Cathartes aura |
1998–2001 | U.S.A. | 2 | 2 | 100 | n=0 | n=0 | n=1 | n=0 | n=1 | n=1 | n=0 | n=0 | - | [50] |
| Short toed snake-eagle Circaetus gallicus |
2005–2010 | Spain | 1 | 1 | 100 | n=0 | n=0 | n=0 | n=0 | n=1 | n=1 | n=1 | [44] | ||
| 9 | 10 | n=0 | n=0 | 2 | |||||||||||
| Marsh harrier Circus aeruginosus |
2000–2009 | Denmark | 3 | 3 | 100 | - | n=† | - | - | n=† | n=† | n=† | - | n=† | [5] |
| Morepork Ninox novaeseelandiae | 1994–1999 | NZ | 5 | 5 | 100 | - | - | - | - | n=5 | - | - | - | - | [8, 35] |
| 610–3,440 | |||||||||||||||
| Little owl Athene noctua |
17 | 15 | 88 | n=5 | n=1 | n=2 | n=1 | ||||||||
| 2005–2013 | Spain | 8 | 6 | 75 | n=0 | - | - | - | 62–574 | 79.5 | 2, 56 | n=0 | 33 | [28, 44] | |
| 2000–2009 | Denmark | 9 | 9 | 100 | - | n=† | - | - | n=† | n=† | n=† | - | n=† | [5] | |
| Eagle owl Bubo bubo |
39 | 33 | 85 | n=20 | n=13 | n=11 | n=3 | n=7 | |||||||
| 2005–2013 | Spain | 21 | 18 | 86 | n=0 | n=0 | n=0 | n=0 | 10–2,008 | 2–208 | 1–281 | 35–200 | 3–90 | [28, 44] | |
| 2000–2009 | Denmark | 10 | 10 | 100 | - | n=† | - | - | n=† | n=† | n=† | - | n=† | [5] | |
| 2009–2011 | Norway | 8 | 5 | 63 | - | - | - | - | 74–158 | n=0 | 39, 181 | - | 13 | [26] | |
| Bald eagle Haliaeetus leucocephalus |
1995–2009 | U.S.A. | 24 | 20 | 83 | n=1 | n=0 | n=0 | n=0 | n=18 | n=0 | n=1 | n=0 | - | [9, 49, 50] |
| 1,400 | 429–2,599 | ||||||||||||||
| Short-eared owl Asio flammeus |
6 | 5 | 83 | ||||||||||||
| 1998–2001 | U.S.A. | 1 | 0 | 0 | n=0 | n=0 | n=0 | n=0 | n=0 | n=0 | n=0 | n=0 | - | [50] | |
| 2000–2009 | Denmark | 5 | 5 | 100 | - | n=† | - | - | n=† | n=† | n=† | - | n=† | [5] | |
| Rough-legged buzzard Buteo lagopus |
32 | 26 | 81 | n=3 | n=19 | n=5 | n=23 | ||||||||
| 1998–2001 | U.S.A. | 1 | 0 | 0 | n=0 | n=0 | n=0 | n=0 | n=0 | n=0 | n=0 | n=0 | - | [50] | |
| 2000–2009 | Denmark | 31 | 26 | 84 | - | 1–3 | - | - | 3*–34 | 0–130 | 14*–105 | - | n=0 | [5] | |
| Sparrowhawk Accipiter nisus |
145 | 116 | 80 | n=3 | n=53 | n=62 | n=90 | n=1 | |||||||
| 2000–2013 | U.K. | 131 | 104 | 79 | n=0 | n=0 | n=0 | n=0 | n=48 | n=57 | n=85 | 3.3 | n=0 | [19, 59] | |
| 2009–2012 | Spain | 14 | 12 | 86 | n=0 | n=0 | - | 0.34 ± 0.57 | 13.2 ± 8.1 | 31.9 ± 22.6 | 3.6 ± 2.0 | n=0 | - | [43] | |
| Red kite Milvus milvus |
138 | 108 | 78 | n=40 | n=74 | n=78 | n=1 | n=5 | |||||||
| 1994–2011 | U.K. | 127 | 98 | 77 | n=0 | n=0 | n=0 | n=0 | 71–222 | 56–94 | 40–67 | n=1 | 15 | [19, 58, 60] | |
| 2005–2010 | Spain | 8 | 7 | 88 | n=0 | n=0 | n=0 | n=0 | 129, 210 | 5–490 | 1 | n=0 | 53, 400 | [44] | |
| 2000–2009 | Denmark | 3 | 3 | 100 | - | n=† | - | - | n=† | n=† | n=† | - | n=† | [5] | |
| Long-eared owl Asio otus |
80 | 62 | 78 | n=4 | n=1 | n=1 | n=39 | n=22 | n=36 | n=3 | |||||
| 1998–2001 | U.S.A. | 7 | 2 | 29 | n=0 | n=0 | n=1 | n=0 | n=1 | n=1 | n=0 | n=0 | - | [50] | |
| 2009–2013 | Spain | 35 | 24 | 69 | n=0 | n=0 | - | 0.5 ± 0.4 | 12–42 | 77.2 ± 29.6 | 1–53 | n=0 | n=0 | [28, 43] | |
| 2000–2009 | Denmark | 38 | 36 | 95 | - | 0*–29 | - | - | 3*–40 | 0*–33 | 7*–52 | - | 0*–2 | [5] | |
| Kestrel Falco tinnunculus |
206 | 154 | 75 | n=12 | n=3 | n=71 | n=94 | n=106 | n=1 | n=18 | |||||
| 2000–2011 | U.K. | 115 | 78 | 68 | - | - | - | - | n=21 | n=52 | n=58 | n=1 | n=0 | [19, 58, 60] | |
| 2003 | France | 4 | 3 | 75 | n=0 | n=0 | - | - | 80–250 | 80–250 | n=0 | - | - | [25] | |
| 2009–2012 | Spain | 21 | 14 | 67 | n=0 | n=0 | - | 0.6 ± 1.8 | 57.4 ± 34.6 | 79.8 ± 34.4 | 8.2 ± 6.9 | n=0 | - | [43] | |
| 2000–2009 | Denmark | 66 | 59 | 89 | - | 0*–64 | - | - | 2*–298 | 0*–679 | 6.5*–450 | - | 0*–20 | [5] | |
| Barred owl Strix varia |
38 | 26 | 68 | n=1 | n=2 | n=4 | n=18 | n=20 | n=1 | ||||||
| 1998–2001 | U.S.A. | 13 | 3 | 23 | n=0 | n=0 | n=1 | n=0 | n=1 | n=1 | n=0 | n=0 | - | [50] | |
| 1988–2003 | Canada | 25 | 23 | 92 | 2.6 | - | 10 | 2.5–15 | 1–927 | 2–1,012 | - | 3 | - | [1] | |
| Golden eagle Aquila chrysaetos |
22 | 14 | 64 | n=1 | n=9 | n=7 | n=3 | ||||||||
| 1996–2001 | U.S.A. | 2 | 2 | 100 | n=0 | n=0 | n=1 | n=0 | 30 | n=0 | n=0 | n=0 | - | [49, 50] | |
| 2005–2010 | Spain | 4 | 1 | 25 | n=0 | n=0 | n=0 | n=0 | n=0 | n=0 | n=0 | n=0 | 6 | [44] | |
| 2009–2011 | Norway | 16 | 11 | 69 | - | - | - | - | 11–110 | 13–154 | n=0 | - | 15, 117 | [26] | |
| Great horned owl Bubo virginianus |
308 | 192 | 62 | n=4 | n=4 | n=3 | n=157 | n=111 | n=1 | n=7 | |||||
| 1994–2012 | U.S.A. | 136 | 74 | 54 | 730 | n=0 | n=0 | n=0 | 7–970 | 28–1,080 | 22 | n=0 | - | [48, 49, 50] | |
| 1988–2003 | Canada | 172 | 118 | 69 | 2.5–720 | - | 8–12 | 2.5–14 | 1–610 | 1–570 | - | 3–30 | - | [1, 55] | |
| Black kite Milvus migrans |
2005–2010 | Spain | 5 | 3 | 60 | n=0 | n=0 | n=0 | n=0 | n=1 | n=0 | n=0 | n=0 | n=2 | [44] |
| 25 | 55, 84 | ||||||||||||||
| Common buzzard Buteo buteo |
639 | 380 | 60 | n=4 | n=31 | n=2 | n=130 | n=238 | n=236 | n=3 | n=35 | ||||
| 2000–2010 | U.K. | 407 | 195 | 48 | n=0 | n=0 | n=0 | n=0 | n=24 | n=122 | n=110 | - | - | [19] | |
| 2003 | France | 11 | 10 | 91 | 350–2,000 | n=0 | - | - | 80–250 | 80–290 | 80–250 | - | - | [25] | |
| 2005–2013 | Spain | 80 | 43 | 54 | n=0 | n=0 | n=0 | 0.3, 120 | 4.9–1,356 | 1–586 | 2.9–1,921 | 85–539 | 1–175 | [28, 43, 44] | |
| 2000–2009 | Denmark | 141 | 132 | 94 | - | 0*–435 | - | - | 2*–613 | 7.5*–282 | 10*–170 | - | 0*–115 | [5] | |
| Australasian harrier Circus approximans |
1994–1999 | NZ | 4 | 2 | 50 | - | - | - | - | n=2 | - | - | - | - | [8, 35] |
| 610, 660 | |||||||||||||||
| Screech owl Otus asio |
1997–2001 | U.S.A. | 24 | 12 | 50 | n=0 | n=0 | n=0 | n=0 | n=10 | n=3 | n=0 | n=0 | - | [49, 50] |
| 7–800 | 50–500 | ||||||||||||||
| Red-tailed hawk Buteo jamaicensis |
362 | 176 | 49 | n=1 | n=1 | n=155 | n=59 | ||||||||
| 1994–2010 | U.S.A. | 297 | 137 | 46 | n=0 | n=0 | 340 | 180 | 6–1,600 | 31–543 | n=0 | n=0 | - | [48, 49, 50] | |
| 2011 | Canada | 65 | 39 | 60 | - | - | - | - | 1–170 | 1–64 | - | n=0 | - | [55] | |
| Scops owl Otus scops |
2011–2013 | Spain | 33 | 16 | 49 | n=0 | n=0 | n=0 | n=0 | n=9 | n=5 | n=8 | n=0 | n=2 | [28] |
| 3–158.4 | 2–44 | 1–10 | 3, 10 | ||||||||||||
| Barn owl Tyto alba |
769 | 370 | 48 | n=1 | n=13 | n=3 | n=3 | n=149 | n=235 | n=203 | n=22 | n=25 | |||
| 1988–2003 | Canada | 78 | 48 | 62 | 2.5 | - | 10–20 | n=0 | 10–470 | 5–720 | - | 2.5–720 | - | [1] | |
| 2000–2011 | U.K. | 535 | 193 | 36 | - | - | - | - | n=33 | n=111 | n=115 | n=4 | n=9 | [19, 58, 60] | |
| 2003 | France | 10 | 7 | 70 | n=0 | 640 | - | - | n=0 | 80–260 | 80–260 | - | - | [25] | |
| 2005–2013 | Spain | 66 | 47 | 71 | n=0 | n=0 | - | 1.2 ± 1.0 | 2–839 | 7.1–180 | 1–198 | 45–4,463 | 14–299 | [28, 43, 44] | |
| 2000–2009 | Denmark | 80 | 75 | 94 | - | 0*–18 | - | - | 4*–957 | 16*–252 | 11*–223 | - | 0*–34 | [5] | |
| Tawny owl Strix aluco |
276 | 109 | 40 | n=6 | n=53 | n=65 | n=51 | n=4 | n=12 | ||||||
| 1990–2010 | U.K. | 200 | 45 | 28 | - | - | - | - | n=12 | n=26 | n=13 | - | n=0 | [19, 60] | |
| 2003 | France | 5 | 2 | 40 | n=0 | n=0 | - | - | n=0 | 80, 250 | n=0 | - | - | [25] | |
| 2011–2013 | Spain | 27 | 21 | 78 | n=0 | - | - | - | 2–1582 | 2–77 | 1–84 | 93–430 | 0–118 | [28] | |
| 2000–2009 | Denmark | 44 | 41 | 93 | 0*–39 | - | - | 3*–220 | 8*–496 | 7*–90 | - | 0*–42 | [5] | ||
| Cooper’s hawk Accipiter cooperii |
1998–2001 | U.S.A. | 50 | 18 | 36 | n=1 | n=0 | n=1 | n=0 | n=12 | n=5 | n=0 | n=0 | - | [50] |
| 100 | 100 | 8–220 | 40–600 | ||||||||||||
| Peregrine falcon Falco peregrinus |
30 | 10 | 33 | n=1 | n=1 | n=3 | n=8 | ||||||||
| 1986–2009 | U.S.A. | 4 | 3 | 75 | 1,480 | n=0 | n=1 | n=0 | n=2 | n=1 | n=0 | n=0 | - | [9, 49, 50] | |
| 2000–2010 | U.K. | 24 | 7 | 29 | n=0 | n=0 | n=0 | n=0 | n=1 | n=7 | n=0 | - | n=0 | [19] | |
| 2009–2011 | Norway | 2 | 0 | 0 | - | - | - | - | n=0 | n=0 | n=0 | - | n=0 | [43] | |
| Northern goshawk Accipiter gentilis |
3 | 1 | 33 | n=1 | |||||||||||
| 1998–2001 | U.S.A. | 1 | 0 | 0 | n=0 | n=0 | n=0 | n=0 | n=0 | n=0 | n=0 | n=0 | - | [50] | |
| 2005–2010 | Spain | 2 | 1 | 50 | n=0 | n=0 | n=0 | n=0 | 38 | n=0 | n=0 | n=0 | n=0 | [44] | |
| Saw-whet owl Aegolius acadicus |
1998–2001 | U.S.A. | 3 | 1 | 33 | n=0 | n=0 | n=1 | n=0 | n=1 | n=1 | n=0 | n=0 | - | [50] |
| Bearded vulture Gypaetus barbatus |
2005–2010 | Spain | 3 | 1 | 33 | n=0 | n=0 | n=0 | n=0 | n=0 | n=1 | n=0 | n=0 | n=0 | [44] |
| 1 | |||||||||||||||
| Snowy owl Nyctea scandiaca |
1993, 1998–2001 | U.S.A. | 3 | 1 | 33 | n=0 | n=0 | n=1 | n=0 | n=0 | n=0 | n=0 | n=0 | - | [49, 50] |
| 260 | |||||||||||||||
| Barbary falcon Falco peregrinoides |
2009–2012 | Spain | 16 | 5 | 31 | n=0 | n=0 | - | n=1 | n=1 | n=3 | n=1 | n=0 | - | [43] |
| 0.1 | 0.8 | 26.2 ± 18.6 | 1.4 | ||||||||||||
| Eurasian griffon Gyps fulvus |
2005–2010 | Spain | 23 | 3 | 13 | n=0 | n=0 | n=0 | n=1 | n=0 | n=1 | n=1 | n=0 | n=0 | [44] |
| 4 | 208 | 1 | |||||||||||||
| Spanish imperial eagle Aquila adalberti |
2005–2010 | Spain | 8 | 1 | 13 | n=0 | n=0 | n=0 | n=0 | n=0 | n=0 | n=1 | n=0 | n=0 | [44] |
| 8 | |||||||||||||||
| Sharp-shinned hawk Accipiter striatus |
1998–2001 | U.S.A. | 11 | 1 | 9 | n=0 | n=0 | n=1 | n=0 | n=1 | n=1 | n=0 | n=0 | - | [50] |
| Broad-winged hawk Buteo platypterus |
1998–2001 | U.S.A. | 11 | 0 | 0 | n=0 | n=0 | n=0 | n=0 | n=0 | n=0 | n=0 | n=0 | - | [50] |
| Black vulture Coragyps atratus |
1998–2001 | U.S.A. | 1 | 0 | 0 | n=0 | n=0 | n=0 | n=0 | n=0 | n=0 | n=0 | n=0 | - | [50] |
| Merlin Falco columbarius |
1998–2001 | U.S.A. | 1 | 0 | 0 | n=0 | n=0 | n=0 | n=0 | n=0 | n=0 | n=0 | n=0 | - | [50] |
| Gyrfalcon Falco rusticolus |
2009–2011 | Norway | 1 | 0 | 0 | - | - | - | - | n=0 | n=0 | n=0 | - | n=0 | [43] |
| Osprey Pandion haliaetus |
2009–2011 | Norway | 3 | 0 | 0 | - | - | - | - | n=0 | n=0 | n=0 | - | n=0 | [43] |
| Birds excluding raptors (40 species analyzed) | |||||||||||||||
| Common myna Acridotheres tristis |
1994–1999 | NZ | 3 | 3 | 100 | - | - | - | - | n=3 | - | - | - | - | [8] |
| 540–1,270 | |||||||||||||||
| Gray duck Anas superciliosa |
1994–1999 | NZ | 1 | 1 | 100 | - | - | - | - | n=1 | - | - | - | - | [8] |
| 910 | |||||||||||||||
| Gray heron Ardea cinerea |
2005–2010 | Spain | 1 | 1 | 100 | n=0 | n=0 | n=0 | n=0 | n=1 | n=0 | n=0 | n=0 | [44] | |
| n=0 | 10 | ||||||||||||||
| Lapland longspur Calcarius lapponicus |
2009 | U.S.A. | 2 | 2 | 100 | n=0 | n=0 | n=0 | n=0 | n=2 | n=0 | n=0 | n=0 | - | [9] |
| 560, 2,989 | |||||||||||||||
| Rock sandpiper Calidris ptilocnemis |
2009 | U.S.A. | 1 | 1 | 100 | n=0 | n=0 | n=0 | n=0 | n=1 | n=0 | n=0 | n=0 | - | [9] |
| 43 | |||||||||||||||
| Emperor goose Chen canagica |
2009 | U.S.A. | 1 | 1 | 100 | n=0 | n=0 | n=0 | n=0 | n=1 | n=0 | n=0 | n=0 | - | [9] |
| 27 | |||||||||||||||
| Great spotted cuckoo Clamator glandarius |
2005–2010 | Spain | 1 | 1 | 100 | n=0 | n=0 | n=0 | n=1 | n=0 | n=0 | n=0 | n=0 | n=0 | [44] |
| 6 | |||||||||||||||
| Common crow Corvus brachyrhynchos |
1997 | U.S.A. | 1 | 1 | 100 | n=0 | n=0 | n=0 | n=0 | n=1 | n=0 | n=0 | - | - | [49] |
| 1,340 | |||||||||||||||
| Raven Corvus corax |
14 | 14 | 100 | n=14 | |||||||||||
| 1996 | U.S.A. | 1 | 1 | 100 | n=0 | n=0 | n=0 | n=0 | 1,040 | n=0 | n=0 | - | - | [49] | |
| 1995 | Canada | 13 | 13 | 100 | - | - | - | - | 980–2,520 | - | - | - | - | [18] | |
| Chaffinch Fringilla coelebs |
1994–1999 | NZ | 3 | 3 | 100 | - | - | - | - | n=3 | - | - | - | - | [8] |
| 120–2,310 | |||||||||||||||
| Southern black-backed gull Larus dominicanus |
1994–1999 | NZ | 1 | 1 | 100 | - | - | - | - | n=1 | - | - | - | - | [8] |
| 580 | |||||||||||||||
| Gray-crowned rosy finch Leucosticte tephrocotis |
2009 | U.S.A. | 1 | 1 | 100 | n=0 | n=0 | n=0 | n=0 | n=1 | n=0 | n=0 | n=0 | - | [9] |
| 1,219 | |||||||||||||||
| Kaka Nestor meridionalis |
1994–1999 | NZ | 3 | 3 | 100 | - | - | - | - | n=3 | - | - | - | - | [8] |
| 1,200–4,100 | |||||||||||||||
| Pukeko Porphyrio porphyrio |
1994–1999 | NZ | 8 | 8 | 100 | - | - | - | - | n=8 | - | - | - | - | [8] |
| 520–1,350 | |||||||||||||||
| Spotless crake Porzana tabuensis |
1994–1999 | NZ | 1 | 1 | 100 | - | - | - | - | n=1 | - | - | - | - | [8] |
| 40 | |||||||||||||||
| Paradise shelduck Tadorna variegata |
1994–1999 | NZ | 4 | 4 | 100 | - | - | - | - | n=4 | - | - | - | - | [8] |
| 240–800 | |||||||||||||||
| Black bird Turdus merula |
1994–1999 | NZ | 13 | 13 | 100 | - | - | - | - | n=13 | - | - | - | - | [8] |
| 10–1,100 | |||||||||||||||
| Northern fulmar Fulmarus glacialis |
2009 | U.S.A. | 1 | 1 | 100 | n=0 | n=0 | n=0 | n=0 | n=1 | n=0 | n=0 | n=0 | - | [9] |
| 57 | |||||||||||||||
| Pelagic cormorant Phalacrocorax pelagicus |
2009 | U.S.A. | 1 | 1 | 100 | n=0 | n=0 | n=0 | n=0 | n=1 | n=0 | n=0 | n=0 | - | [9] |
| 44 | |||||||||||||||
| Glaucous-winged gull Larus glaucescens |
2009 | U.S.A. | 10 | 9 | 90 | n=0 | n=0 | n=0 | n=0 | n=9 | n=1 | n=2 | n=0 | - | [9] |
| 709–4,189 | n=1 | n=2 | |||||||||||||
| Rock dove Columba livia |
2005–2010 | Spain | 97 | 64 | 66 | n=0 | n=0 | n=0 | n=64 | n=0 | n=0 | n=0 | n=0 | n=0 | [44] |
| 550–55,100 | |||||||||||||||
| Weka Gallirallus australis |
1994–1999 | NZ | 55 | 31 | 56 | - | - | - | - | n=31 | - | - | - | - | [8] |
| 10–2,300 | |||||||||||||||
| Kakariki Cyanoramphus sp. |
1994–1999 | NZ | 2 | 1 | 50 | - | - | - | - | n=1 | - | - | - | - | [8] |
| 30 | |||||||||||||||
| Saddleback Philesturnus carunculatus |
1994–1999 | NZ | 4 | 2 | 50 | - | - | - | - | n=2 | - | - | - | - | [8] |
| 50, 600 | |||||||||||||||
| Brown kiwi Apteryx australis |
1994–1999 | NZ | 29 | 14 | 48 | - | - | - | - | n=14 | - | - | - | - | [8] |
| 10–690 | |||||||||||||||
| Lesser black-backed gull Larus fuscus |
2005–2010 | Spain | 8 | 3 | 38 | n=0 | n=0 | n=0 | n=0 | n=0 | n=3 | n=0 | n=0 | n=0 | [44] |
| 2–5 | |||||||||||||||
| Common starling Sturnus vulgaris |
2005–2010 | Spain | 3 | 1 | 33 | n=0 | n=0 | n=0 | n=0 | n=0 | n=1 | n=0 | n=0 | n=0 | [44] |
| 15 | |||||||||||||||
| Calandra lark Melanocorypha calandra |
2005–2010 | Spain | 7 | 2 | 29 | n=0 | n=0 | n=0 | n=2 | n=0 | n=0 | n=0 | n=0 | n=0 | [44] |
| 1,040, 2,090 | |||||||||||||||
| Mallard Anas platyrhynchos |
23 | 6 | 26 | n=3 | n=2 | n=1 | |||||||||
| 2003 | France | 15 | 1 | 7 | n=0 | n=0 | - | - | - | 80–250 | - | - | - | [25] | |
| 2005–2010 | Spain | 6 | 3 | 50 | n=0 | n=0 | n=0 | 710–2,170 | - | n=0 | n=0 | n=0 | n=0 | [44] | |
| 1994–1999 | NZ | 2 | 2 | 100 | - | - | - | 900, 1,230 | - | - | - | - | [8] | ||
| Black coot Fulica atra |
2003 | France | 13 | 3 | 23 | n=1 | n=0 | - | - | n=1 | n=1 | n=0 | - | - | [25] |
| 23,520 | 80–250 | 80–250 | |||||||||||||
| Magpie Gymnorhina tibicen |
1994–1999 | NZ | 30 | 6 | 20 | - | - | - | - | n=6 | - | - | - | - | [8, 35] |
| 80–990 | |||||||||||||||
| Robin Petroica australis |
1994–1999 | NZ | 10 | 2 | 20 | - | - | - | - | n=2 | - | - | - | - | [8, 35] |
| 350, 580 | |||||||||||||||
| Red-legged partridge Alectoris rufa |
2005–2010 | Spain | 7 | 1 | 14 | n=0 | n=0 | n=0 | n=0 | n=0 | n=0 | n=0 | n=0 | n=1 | [44] |
| 143 | |||||||||||||||
| Eurasian collared-dove Streptopelia decaocto |
2005–2010 | Spain | 8 | 1 | 13 | n=0 | n=0 | n=0 | n=0 | n=0 | n=1 | n=0 | n=0 | n=0 | [44] |
| 127 | |||||||||||||||
| Bellbird Anthornis melanura |
1994–1999 | NZ | 2 | 0 | 0 | - | - | - | - | n=0 | - | - | - | - | [8, 35] |
| Northwestern crow Corvus caurinus |
1995 | Canada | 9 | 0 | 0 | - | - | - | - | n=0 | - | - | - | - | [18] |
| Common moorhen Gallinula chloropus |
2003 | France | 1 | 0 | 0 | n=0 | n=0 | - | - | n=0 | n=0 | n=0 | - | - | [25] |
| Whitehead Mohoua albicilla |
1994–1999 | NZ | 12 | 0 | 0 | - | - | - | - | n=0 | - | - | - | - | [8, 35] |
| Tomtit Petroica macrocephala |
1994–1999 | NZ | 5 | 0 | 0 | - | - | - | - | n=0 | - | - | - | - | [35] |
| Fantail Rhipidura fuliginosa |
1994–1999 | NZ | 2 | 0 | 0 | - | - | - | - | n=0 | - | - | - | - | [8, 35] |
| Carnivola (15 species analyzed) | |||||||||||||||
| Skunk Mephitis mephitis |
1996 | U.S.A. | 3 | 3 | 100 | n=0 | n=0 | n=0 | n=0 | n=0 | n=3 | n=0 | - | - | [49] |
| 20–280 | |||||||||||||||
| Mountain lion Puma concolor |
1997–2006 | U.S.A. | 4 | 4 | 100 | n=0 | - | n=1 | n=0 | n=4 | n=4 | - | n=1 | ||
| <250 | 310–570 | 370–1,270 | <250 | ||||||||||||
| Bobcat Lynx rufus |
1988–2012 | U.S.A. | 169 | 148 | 88 | n=11 | - | n=67 | n=10 | n=135 | n=133 | - | n=50 | - | [42, 45] |
| n=11 | 30 ± 120 | n=10 | 140 ± 200 | 380 ± 550 | 40 ± 310 | ||||||||||
| Weasel Mustela nivalis |
80 | 69 | 86 | n=15 | n=39 | n=17 | n=60 | n=20 | |||||||
| 1996–1997 | U.K. | 10 | 3 | 30 | n=0 | 8.5–60 | - | - | n=0 | 250 | n=0 | - | n=0 | [31] | |
| 2005–2010 | Spain | 1 | 1 | 100 | n=0 | n=0 | n=0 | n=0 | 2,930 | n=0 | n=0 | n=0 | n=0 | [44] | |
| 1984–2008 | Denmark | 69 | 65 | 94 | - | 4–45 | - | - | 4–159 | 7–1,610 | 5–292 | - | 3–49 | [11] | |
| Feral cat Felis catus |
2005–2010 | Spain | 4 | 3 | 75 | n=0 | n=0 | n=0 | n=0 | n=2 | n=1 | n=1 | - | n=1 | [44] |
| 34, 350 | 52 | 70 | 72 | ||||||||||||
| Stoat Mustela erminea |
101 | 68 | 67 | n=22 | n=31 | n=34 | n=49 | n=25 | |||||||
| 1996–1997 | U.K. | 40 | 9 | 23 | n=0 | 4.6–9.7 | - | - | 120 | 40–380 | n=0 | - | n=0 | [31] | |
| 1993–2007 | Denmark | 61 | 59 | 97 | - | 2–61 | - | - | 2–317 | 3–1,290 | 2–280 | - | 1–86 | [11] | |
| Stone marten Martes foina |
2005–2010 | Spain | 19 | 11 | 58 | n=0 | n=0 | n=0 | n=0 | n=5 | n=6 | n=3 | n=1 | n=5 | [44] |
| 19–390 | 7–17,900 | 7–520 | 926 | 8–230 | |||||||||||
| Raccoon Procyon lotor |
16 | 8 | 50 | n=6 | n=2 | ||||||||||
| 1992–1997 | U.S.A. | 6 | 6 | 100 | n=0 | n=0 | n=0 | n=0 | 320–5,300 | n=0 | n=0 | - | - | [49] | |
| 2005–2010 | Spain | 10 | 2 | 20 | n=0 | n=0 | n=0 | n=0 | n=0 | 1,090, 6,800 | n=0 | n=0 | n=0 | [44] | |
| Red fox Vulpes vulpes |
33 | 14 | 42 | n=7 | n=8 | n=1 | |||||||||
| 1996 | U.S.A. | 2 | 2 | 100 | n=0 | n=0 | n=0 | n=0 | 1,320, 4,010 | n=0 | n=0 | - | - | [46] | |
| 2005–2010 | Spain | 31 | 12 | 39 | n=0 | n=0 | n=0 | n=0 | 5–4,500 | 5–12,300 | 78 | n=0 | n=0 | [44] | |
| Domestic dog Canis familiaris |
2005–2010 | Spain | 11 | 4 | 36 | n=0 | n=0 | n=0 | n=0 | n=0 | n=3 | n=1 | n=0 | n=0 | [44] |
| 6–308 | 4 | ||||||||||||||
| Common genet Genetta genetta |
2005–2010 | Spain | 7 | 2 | 29 | n=0 | n=0 | n=0 | n=0 | n=2 | n=2 | n=1 | n=0 | n=1 | [44] |
| 16, 2,020 | 1, 350 | 12 | 60 | ||||||||||||
| European otter Lutra lutra |
14 | 4 | 29 | n=1 | n=2 | n=1 | |||||||||
| 1990–2002 | France | 11 | 3 | 27 | n=0 | n=0 | - | 5,000 | n=0 | 6,00, 7,100 | n=0 | n=0 | - | [13] | |
| 2005–2010 | Spain | 3 | 1 | 33 | n=0 | n=0 | n=0 | n=0 | n=0 | n=0 | n=0 | n=0 | 353 | [44] | |
| Polecat Mustela putorius |
133 | 36 | 27 | n=3 | n=17 | n=22 | |||||||||
| 1992–1999 | U.K. | 100 | 31 | 31 | - | - | - | - | 8–70 | 16–217 | 5–917 | - | n=0 | [49, 50] | |
| 1990–2002 | France | 33 | 5 | 15 | n=0 | n=0 | - | n=0 | n=0 | 600–9,000 | n=0 | n=0 | - | [13] | |
| American mink Mustela vison |
1990–2002 | France | 47 | 7 | 15 | n=0 | n=0 | - | n=4 | n=0 | n=3 | n=0 | n=0 | - | [13] |
| 3,400–8,500 | 1,900–4,200 | ||||||||||||||
| European mink Mustela lutreola |
1990–2002 | France | 31 | 1 | 3 | n=0 | n=0 | - | n=0 | n=0 | n=1 | n=0 | n=0 | - | [13] |
| 5,000 | |||||||||||||||
| Cetartiodactyla (3 species analyzed) | |||||||||||||||
| White-tailed deer Odocoileus virginianus |
1994–1997 | U.S.A. | 7 | 7 | 100 | n=0 | n=1 | n=2 | n=0 | n=5 | n=0 | n=0 | - | - | [49] |
| 500 | 200, 930 | 120–410 | |||||||||||||
| Red deer Cervus elaphus |
1994–1999 | NZ | 37 | 33 | 89 | - | - | - | - | n=33 | - | - | - | - | [8, 35] |
| <30 | |||||||||||||||
| Domestic pig Sus scrofa |
1994–1999 | NZ | 40 | 26 | 65 | - | - | - | - | n=26 | - | - | - | - | [8, 35] |
| 7–1,780 | |||||||||||||||
| Erinaceomorpha (2 species analyzed) | |||||||||||||||
| Algerian hedgehog Atelerix algirus |
2011–2013 | Spain | 106 | 61 | 58 | n=1 | - | - | - | n=18 | n=35 | n=29 | n=4 | n=0 | [28] |
| 611.8 | 5–1,533 | 6–2,548 | 1–659 | 71–256 | |||||||||||
| European hedgehog Erinaceus europaeus |
170 | 57 | 34 | n=29 | n=28 | n=28 | n=2 | n=6 | |||||||
| 2004–2006 | U.K. | 120 | 27 | 23 | - | - | - | - | 50 ± 10# | 590 ± 240# | 100 ± 30# | - | n=0 | [7] | |
| 2005–2013 | Spain | 50 | 30 | 60 | n=0 | - | - | - | 3–1,390 | 2–1,110 | 0–672 | 4, 142 | 1–29 | [28, 44] | |
| Rodentia (2 species analyzed) | |||||||||||||||
| Gray squirrel Sciurus carolinensi |
1981–1997 | U.S.A. | 7 | 7 | 100 | n=1 | n=0 | n=1 | n=1 | n=5 | n=0 | n=0 | - | - | [50] |
| 228 | 2,000 | 620 | 530–4,100 | ||||||||||||
| Eastern chipmunk Tamias striatus |
1992 | U.S.A. | 1 | 1 | 100 | n=0 | n=0 | n=0 | n=0 | n=1 | n=0 | n=0 | - | - | [50] |
| 3,800 | |||||||||||||||
| Lagomorpha (1 species analyzed) | |||||||||||||||
| Iberian hare Lepus granatensis |
2005–2010 | Spain | 25 | 8 | 32 | n=0 | n=0 | n=0 | n=7 | n=2 | n=0 | n=1 | n=0 | n=0 | [44] |
| 580–9,520 | 130, 270 | 15 | |||||||||||||
| Chiroptera (1 species analyzed) | |||||||||||||||
| NZ lesser short-tailed bats Mystacina tuberculata |
2009 | NZ | 12 | 10 | 83 | - | - | n=10 | - | - | - | - | - | - | [6] |
| 190–680 | |||||||||||||||
| Marsupialia (1 species analyzed) | |||||||||||||||
| Opossum Didelphis virginiana |
1996, 1997 | U.S.A. | 2 | 2 | 100 | n=0 | n=0 | n=0 | n=0 | n=1 | n=1 | n=0 | - | - | [49] |
| 180 | 800 | ||||||||||||||
| Reptile (1 species analyzed) | |||||||||||||||
| Horseshoe whip snake Hemorrhois hippocrepis |
2005–2010 | Spain | 1 | 1 | 100 | n=0 | n=0 | n=0 | n=0 | n=0 | n=0 | n=0 | n=0 | n=1 | [44] |
| 540 | |||||||||||||||
Of these 17 species, ten species included some individuals, which had liver SGAR concentrations more than 100 µg/kg (Table 5). The potentially lethal level for brodifacoum residues were reported in some individuals of all ten species: at least five moreporks (610–3,440 µg/kg), one little owl (574 µg/kg), four Eurasian eagle owls (133–2,008 µg/kg), 18 bald eagles (429–2,599 µg/kg), one sparrowhawk (112 µg/kg), three red kites (129–222 µg/kg), two kestrels (240 and 298 µg/kg), one barred owl (927 µg/kg), one golden eagle (110 µg/kg), and 15 great horned owls (100–970 µg/kg). High bromadiolone concentrations were reported in six species: at least one Eurasian eagle owl (208 µg/kg), one red kite (490 µg/kg), one kestrel (679 µg/kg), one barred owl (1,012 µg/kg), one golden eagle (154 µg/kg), and four great horned owls (226–1,080 µg/kg). High difenacoum accumulations were reported in two Eurasian eagle owls (181 and 281 µg/kg) and one kestrel (450 µg/kg), at least. High flocoumafen accumulations were reported in one red kite (400 µg/kg) and one golden eagle (117 µg/kg). High difethialone accumulations were reported in one Eurasian eagle owl (200 µg/kg). Four sparrowhawks were reported to have sum SGAR concentrations of 100–157 µg/kg. Because of the high AR exposure rates and including some individuals that have high liver SGAR concentrations, these ten species are seemingly affected by ARs more severely than other raptor species.
Threatened raptors
A wide variety of raptor species was found to have been exposed to ARs. Furthermore, these reported raptors include species with special conservation status, such as red kites (Near Threatened, IUCN Red List [54]) and the Spanish imperial eagle (Aquila adalberti, Vulnerable, IUCN Red List [51]).
Although there are no reports of AR residues in the livers of raptors from Russia or Mongolia, it has been reported that the numbers of breeding pairs of Eastern imperial eagles (Aquila heliaca, Vulnerable, IUCN Red List [52]) and Saker falcons (Falco cherrug milvipes, Endangered, IUCN Red List [53]) decreased following bromadiolone application [15, 36]. In Japan, diphacinone was used on Ogasawara Island and it was reported that at least three Eastern buzzards (Buteo japonicus toyoshimai), classified as locally Endangered (Ministry of the Environment, Government of Japan [33]), might have been poisoned as a result [4, 16]. These incidents imply that poisoning by ARs could have occurred worldwide, despite the lack of studies on the existence of AR residues in the livers of raptors from these areas.
EXPOSURE PATHWAYS
Diets of raptors
Although it is widely thought that preying on the target rodents is the dominant pathway by which raptors are exposed to ARs, there have been a few studies on the relationship between the diets of raptors and the incidence of AR-exposure. This study is the first to discuss the exposure pathways for ARs based on both a comprehensive analysis of primary and secondary AR-exposure worldwide and the diets of raptors.
The most affected raptor species have commonly been thought to prey predominantly on mammals, especially the targeted rodent species. Figure 2 shows the diet composition of raptors whose AR-exposure are frequently reported. Because mammals constitute 60–80% of the diets of great horned owls [3], barred owls [27], kestrels [24], Eurasian eagle owls [37], and red kites [34], targeted rodents can be the source of ARs in some cases. Moreover, ARs were detected in barn owls (Tyto alba) pellets that contained rat fur [10]. However, the Eurasian eagle owls occasionally prey on larger mammals (e.g., hares, foxes, and deer) [37], and red kites sometimes feed on hares, rabbits, and as carrion, foxes, stoats, polecats and deer [34]. Therefore, a wide range of mammals can be the source of ARs as well as target rodents. On the other hand, raptors also prey on other animals, such as birds, reptiles, amphibians, fish, and invertebrates. Birds constitute even 99 and 73% of the diets in sparrowhawks [63] and golden eagles [22], respectively; and 26, 21 and 14% in bald eagles [32], red kites [34] and great horned owls [3] following, respectively. In addition, the Eurasian eagle owls occasionally prey on other raptors (e.g., buzzards, falcons, tawny owls, and long-eared owls), and the diets of Eurasian eagle owls also consist of amphibians (28%) [37]. Diets of bald eagles consist of fish (70%) [32]. Invertebrates compose 99, 72 and 25% of prey of moreporks [17], little owls [56] and kestrels [24], respectively. Therefore, preying on targeted rodents does not necessarily explain all causes of secondary AR-exposure in raptors.
Fig. 2.
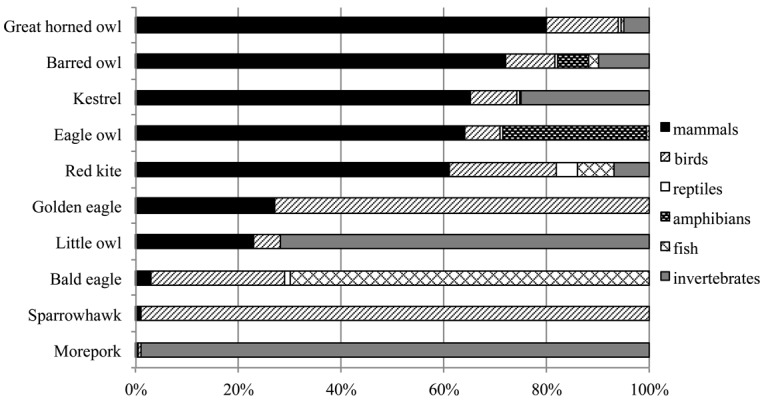
Diet composition of ten raptor species, which frequently reported AR-exposure. Predominant prey of raptors is mammal in great horned owls (80%), barred owls (72%), kestrels (65%), Eurasian eagle owls (64%), and red kites (61%), whereas it is bird in sparrowhawks (99%) and golden eagles (73%), fish in bald eagles (70%), and invertebrate in moreporks (99%) and little owls (72%) [3, 17, 22, 24, 27, 32, 34, 37, 56, 63].
AR residues in the prey of raptors
Relatively high concentrations of AR residues have been detected in the livers of mallards (chlorophacinone, 710–2,170 µg/kg, and brodifacoum, 1,230 µg/kg), rock doves (chlorophacinone, 550–55,100 µg/kg), chaffinches (brodifacoum, 120–2,310 µg/kg), Algerian hedgehogs (brodifacoum, 5–1,533 µg/kg and bromadiolone, 6–2,548 µg/kg), European hedgehogs (brodifacoum, 3–1,390 µg/kg and bromadiolone, 2–1,110 µg/kg), Iberian hares (chlorophacinone, 580–9,520 µg/kg), and a horseshoe whip snake (flocoumafen, 540 µg/kg) (Table 5). Because all of these animals are the prey of raptors, non-target animals (i.e., non-target rodents, birds, hedgehogs, hares and snakes) could also be a source of exposure to ARs.
Some studies have also reported AR residues in other reptiles (geckos) and invertebrates such as ants, cockroaches, beetles, slugs, and snails [10, 23, 38]. In addition, recent studies have shown that ARs can be detected not only in shorebirds and seabirds, but also in marine biota, including fish, crabs, sea urchins, and shellfish [30, 38]. These papers suggest exposure pathways for ARs from marine biota to shorebirds and/or seabirds, and from shorebirds and seabirds to raptors. In the case of Rat Island in Alaska, when gulls died after eating bait containing brodifacoum, their carcasses attracted bald eagles [9]. During the study period, 46 bald eagle carcasses were collected, and the brodifacoum residues were detected in the livers of all the carcasses tested (18 bald eagles).
In conclusion, primary AR exposure has occurred in non-target mammals, reptiles, invertebrates, marine biota, and birds, including land birds, shorebirds and seabirds. It is possible that AR residues in these animals constitute exposure pathways for raptors, resulting in the secondary exposure (Fig. 3). In some cases, these exposure pathways are presumed to cause secondary poisoning in raptors. More studies focusing on the dose-response relations between hepatic AR concentration and both hemorrhage and mortality of various raptor species are necessary to understand incidents of secondary AR poisoning in raptors accurately. Furthermore, toxicokinetics studies of raptors (e.g. AR metabolism, and degree of ARs inhibiting the target molecule) are also necessary to reveal the risk of raptors poisoned by ARs.
Fig. 3.
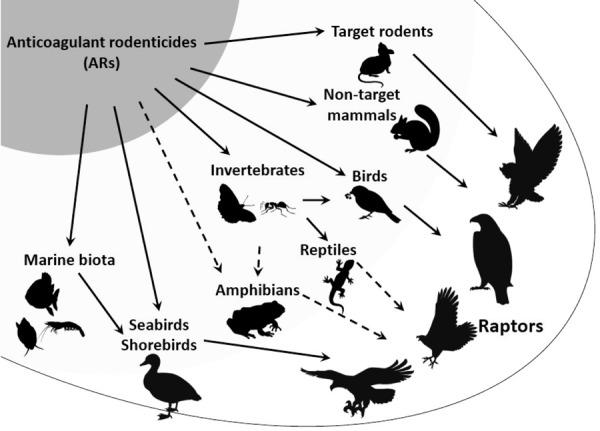
Proposed pathways by which raptors are exposed to ARs. Primary poisoning occurs in target rodents, non-target mammals, reptiles, invertebrates, marine biota, and birds, including land birds, shorebirds and seabirds. It is possible that AR residues in these animals are transferred to raptors, causing secondary poisoning.
CONFLICTS OF INTEREST
The authors declare no conflict of interest.
Acknowledgments
This study was supported by Grants-in-Aid for Scientific Research from the Ministry of Education, Culture, Sports, Science and Technology of Japan awarded to M. Ishizuka (No. 16H0177906) and Y. Ikenaka (No. 26304043, 15H0282505, 15K1221305), and S.M.M. Nakayama (No. 16K16197, 17KK0009), and the foundation of JSPS Core to Core Program (AA Science Platforms) and Bilateral Joint Research Project (PG36150002 and PG36150003), as well as Ministry of Environment, Japan awarded to S.M.M. Nakayama (4RF-1802). We also acknowledge financial support from The Soroptimist Japan Foundation, The Nakajima Foundation, The Sumitomo foundation, The Nihon Seimei Foundation and The Japan Prize Foundation. This research was supported by JST/JICA, SATREPS (Science and Technology Research Partnership for Sustainable Development). The data analyses were partially supported by Mr. Takahiro Ichise, Ms. Mio Yagihashi and Ms. Nagisa Hirano.
REFERENCES
- 1.Albert C. A., Wilson L. K., Mineau P., Trudeau S., Elliott J. E.2010. Anticoagulant rodenticides in three owl species from Western Canada, 1988-2003. Arch. Environ. Contam. Toxicol. 58: 451–459. doi: 10.1007/s00244-009-9402-z [DOI] [PubMed] [Google Scholar]
- 2.Borrell B.2011. Where eagles die. NATNEWS 2011: . [Google Scholar]
- 3.Carl D. M., Michael N. K.1995. Are red-tailed hawks and great horned owls diurnal-nocturnal dietary counterparts? Wilson Bull. 107: 615–628. [Google Scholar]
- 4.Chiba Y., Chiba H.2014. Ogasawara Buzzard (Buteo buteo toyoshimai) utilizes small islands of the Bonin (Ogasawara) Islands during the rodenticides application season. Tokyo Metrop. Univ. Institutional Repos.: 67–79 (The title in Japanese is translated into English by the authors).
- 5.Christensen T. K., Lassen P., Elmeros M.2012. High exposure rates of anticoagulant rodenticides in predatory bird species in intensively managed landscapes in Denmark. Arch. Environ. Contam. Toxicol. 63: 437–444. doi: 10.1007/s00244-012-9771-6 [DOI] [PubMed] [Google Scholar]
- 6.Dennis G. C., Gartrell B. D.2015. Nontarget mortality of New Zealand lesser short-tailed bats (Mystacina tuberculata) caused by diphacinone. J. Wildl. Dis. 51: 177–186. doi: 10.7589/2013-07-160 [DOI] [PubMed] [Google Scholar]
- 7.Dowding C. V., Shore R. F., Worgan A., Baker P. J., Harris S.2010. Accumulation of anticoagulant rodenticides in a non-target insectivore, the European hedgehog (Erinaceus europaeus). Environ. Pollut. 158: 161–166. doi: 10.1016/j.envpol.2009.07.017 [DOI] [PubMed] [Google Scholar]
- 8.Eason C. T., Murphy E. C., Wright G. R. G., Spurr E. B.2002. Assessment of risks of brodifacoum to non-target birds and mammals in New Zealand. Ecotoxicology 11: 35–48. doi: 10.1023/A:1013793029831 [DOI] [PubMed] [Google Scholar]
- 9.Ebbert S., Burek-Huntington K.2010. Anticoagulant residual concentration and poisoning in birds following a large-scale aerial broadcast of 25-ppm brodifacoum bait for rat eradication on Rat Island, Alaska. Proc. 24th Vertebr. Pest Conf. Publ. Univ. Calif., Davis: 153–160.
- 10.Elliott J. E., Hindmarch S., Albert C. A., Emery J., Mineau P., Maisonneuve F.2014. Exposure pathways of anticoagulant rodenticides to nontarget wildlife. Environ. Monit. Assess. 186: 895–906. doi: 10.1007/s10661-013-3422-x [DOI] [PubMed] [Google Scholar]
- 11.Elmeros M., Christensen T. K., Lassen P.2011. Concentrations of anticoagulant rodenticides in stoats Mustela erminea and weasels Mustela nivalis from Denmark. Sci. Total Environ. 409: 2373–2378. doi: 10.1016/j.scitotenv.2011.03.006 [DOI] [PubMed] [Google Scholar]
- 12.Erickson W., Urban D.2004. Potential Risks of Nine Rodenticides to Birds and Non Target Mammals: A Comparative Approach. United States Environ. Prot. Agency, Washington, D.C. [Google Scholar]
- 13.Fournier-Chambrillon C., Berny P. J., Coiffier O., Barbedienne P., Dassé B., Delas G., Galineau H., Mazet A., Pouzenc P., Rosoux R., Fournier P.2004. Evidence of secondary poisoning of free-ranging riparian mustelids by anticoagulant rodenticides in France: implications for conservation of European mink (Mustela lutreola). J. Wildl. Dis. 40: 688–695. doi: 10.7589/0090-3558-40.4.688 [DOI] [PubMed] [Google Scholar]
- 14.Gillies C. A., Pierce R. J.1999. Secondary poisoning of mammalian predators during possum and rodent control operations at Trounson Kauri Park, Northland, New Zealand. N. Z. J. Ecol. 23: 183–192. [Google Scholar]
- 15.Gombobaatar S., Sumiya D., Shagdarsuren O., Potapov E. R., Fox N.2004. Saker falcon (Falco cherrug milvipes Jerdon) mortality in central Mongolia and population threats. Mong. J. Biol. Sci. 2: 13–21. [Google Scholar]
- 16.Hashimoto T.2009. Eradication and Ecosystem Impact of rats in the Ogasawara Islands. pp. 153–159. In: Restoring the Oceanic Island Ecosystem, Springer, Basel. [Google Scholar]
- 17.Haw J. M., Clout M. N., Powlesland R. G.2001. Diet of moreporks (Ninox novaeseelandiae) in Pureora Forest determined from prey remains in regurgitated pellets. N. Z. J. Ecol. 25: 61–67. [Google Scholar]
- 18.Howald G. R., Mineau P., Elliott J. E., Cheng K. M.1999. Brodifacoum poisoning of avian scavengers during rat control on a seabird colony. Ecotoxicology 8: 431–447. doi: 10.1023/A:1008951701780 [DOI] [Google Scholar]
- 19.Hughes J., Sharp E., Taylor M. J., Melton L., Hartley G.2013. Monitoring agricultural rodenticide use and secondary exposure of raptors in Scotland. Ecotoxicology 22: 974–984. doi: 10.1007/s10646-013-1074-9 [DOI] [PubMed] [Google Scholar]
- 20.Jackson W. R., Ashton A. D.1992. A review of available anticoagulants and their use in the United States. Proc. Fifteenth Vertebr. Pest Conf.:156–160.
- 21.Jacob J., Buckle A.2018. Use of anticoagulant rodenticides in different applications around the world. pp. 11–43. In: Anticoagulant Rodenticides and Wildlife. Emerging Topics in Ecotoxicology (Principles, Approaches and Perspectives) (van den Brink, N., Elliott, J., Shore, R. and Rattner, B. eds.), Springer, Cham. [Google Scholar]
- 22.Johnsen T. V., Systad G. H., Jacobsen K. O., Nygard T., Bustnes J. O.2007. The occurrence of reindeer calves in the diet of nesting Golden Eagles in Finnmark, northern Norway. Ornis Fenn. 84: 112–118. [Google Scholar]
- 23.Johnston J. J., Pitt W. C., Sugihara R. T., Eisemann J. D., Primus T. M., Holmes M. J., Crocker J., Hart A.2005. Probabilistic risk assessment for snails, slugs, and endangered honeycreepers in diphacinone rodenticide baited areas on Hawaii, U.S.A. Environ. Toxicol. Chem. 24: 1557–1567. doi: 10.1897/04-255R.1 [DOI] [PubMed] [Google Scholar]
- 24.Korpimaki E.1985. Diet of the Kestrel Falco tinnunculus in the breeding season. Ornis Fenn. 62: 130–137. [Google Scholar]
- 25.Lambert O., Pouliquen H., Larhantec M., Thorin C., L’Hostis M.2007. Exposure of raptors and waterbirds to anticoagulant rodenticides (difenacoum, bromadiolone, coumatetralyl, coumafen, brodifacoum): epidemiological survey in Loire Atlantique (France). Bull. Environ. Contam. Toxicol. 79: 91–94. doi: 10.1007/s00128-007-9134-6 [DOI] [PubMed] [Google Scholar]
- 26.Langford K. H., Reid M., Thomas K. V.2013. The occurrence of second generation anticoagulant rodenticides in non-target raptor species in Norway. Sci. Total Environ. 450-451: 205–208. doi: 10.1016/j.scitotenv.2013.01.100 [DOI] [PubMed] [Google Scholar]
- 27.Livezey K. C.2007. Barred owl habitat and prey: a review and synthesis of the literature. J. Raptor Res. 41: 177–201. doi: 10.3356/0892-1016(2007)41[177:BOHAPA]2.0.CO;2 [DOI] [Google Scholar]
- 28.López-Perea J. J., Camarero P. R., Molina-López R. A., Parpal L., Obón E., Solá J., Mateo R.2015. Interspecific and geographical differences in anticoagulant rodenticide residues of predatory wildlife from the Mediterranean region of Spain. Sci. Total Environ. 511: 259–267. doi: 10.1016/j.scitotenv.2014.12.042 [DOI] [PubMed] [Google Scholar]
- 29.Lovett R. A.2012. Killing rats is killing birds. NATNEWS 2012: . [Google Scholar]
- 30.Masuda B. M., Fisher P., Beaven B.2015. Residue profiles of brodifacoum in coastal marine species following an island rodent eradication. Ecotoxicol. Environ. Saf. 113: 1–8. doi: 10.1016/j.ecoenv.2014.11.013 [DOI] [PubMed] [Google Scholar]
- 31.McDonald R. A., Harris S., Turnbull G., Brown P., Fletcher M.1998. Anticoagulant rodenticides in stoats (Mustela erminea) and weasels (Mustela nivalis) in England. Environ. Pollut. 103: 17–23. doi: 10.1016/S0269-7491(98)00141-9 [DOI] [Google Scholar]
- 32.McEwan L. C., Hirth D. H.1980. Food habitats of bald eagles in north-central Florida. Condor 82: 229–231. doi: 10.2307/1367485 [DOI] [Google Scholar]
- 33.Ministry of the Environment, Government of Japan 2018. Third revision to the 4th Version of the Japanese Red Lists. https://www.env.go.jp/press/files/jp/109278.pdf (in Japanese) [accessed on November 30, 2018].
- 34.Mougeot F., Garcia J. T., Viñuela J.2011. Breeding biology, behaviour, diet and conservation of the red kite (Milvus milvus), with particular emphasis on Mediterranean populations. Ecol. Conserv. Eur. Dwell. For. raptors owls.:190–204. http://www.eeza.csic.es/Documentos/Publicaciones/2011-Red kite book chapter.pdf [accessed on November 30, 2018].
- 35.Murphy E. C., Clapperton B. K., Bradfield P. M. F., Speed H. J.1998. Brodifacoum residues in target and non-target animals following large-scale poison operations in New Zealand podocarp-hardwood forests. New Zeal. J. Zool. (Lond.) 25: 307–314. [Google Scholar]
- 36.Novgorod N., ЦeHTp, K.И.B, HOBrOpOд, H. 2010. The Imperial Eagle is a Vanishing Species in the Tyva Republic, Russia. Rap. Res. Raptors Conserv. 20: 177–185. [Google Scholar]
- 37.Obuch J., Karaska D.2010. The Eurasian eagle-owl (Bubo bubo) diet in the Orava Region (N Slovakia). Slovak Raptor J. 4: 83–98. doi: 10.2478/v10262-012-0048-9 [DOI] [Google Scholar]
- 38.Pitt W. C., Berentsen A. R., Shiels A. B., Volker S. F., Eisemann J. D., Wegmann A. S., Howald G. R.2015. Non-target species mortality and the measurement of brodifacoum rodenticide residues after a rat (Rattus rattus) eradication on Palmyra Atoll, tropical Pacific. Biol. Conserv. 185: 36–46. doi: 10.1016/j.biocon.2015.01.008 [DOI] [Google Scholar]
- 39.Rattner B. A., Horak K. E., Lazarus R. S., Goldade D. A., Johnston J. J.2014. Toxicokinetics and coagulopathy threshold of the rodenticide diphacinone in eastern screech-owls (Megascops asio). Environ. Toxicol. Chem. 33: 74–81. doi: 10.1002/etc.2390 [DOI] [PubMed] [Google Scholar]
- 40.Rattner B. A., Horak K. E., Warner S. E., Day D. D., Meteyer C. U., Volker S. F., Eisemann J. D., Johnston J. J.2011. Acute toxicity, histopathology, and coagulopathy in American kestrels (Falco sparverius) following administration of the rodenticide diphacinone. Environ. Toxicol. Chem. 30: 1213–1222. doi: 10.1002/etc.490 [DOI] [PubMed] [Google Scholar]
- 41.Rattner B. A., Lazarus R. S., Elliott J. E., Shore R. F., van den Brink N.2014. Adverse outcome pathway and risks of anticoagulant rodenticides to predatory wildlife. Environ. Sci. Technol. 48: 8433–8445. doi: 10.1021/es501740n [DOI] [PubMed] [Google Scholar]
- 42.Riley S. P. D., Bromley C., Poppenga R. H., Uzal F. A., Whited L., Sauvajot R. M.2007. Anticoagulant exposure and notoedric mange in bobcats and mountain lions in urban southern California. J. Wildl. Manage. 71: 1874–1884. doi: 10.2193/2005-615 [DOI] [Google Scholar]
- 43.Ruiz-Suárez N., Henríquez-Hernández L. A., Valerón P. F., Boada L. D., Zumbado M., Camacho M., Almeida-González M., Luzardo O. P.2014. Assessment of anticoagulant rodenticide exposure in six raptor species from the Canary Islands (Spain). Sci. Total Environ. 485-486: 371–376. doi: 10.1016/j.scitotenv.2014.03.094 [DOI] [PubMed] [Google Scholar]
- 44.Sánchez-Barbudo I. S., Camarero P. R., Mateo R.2012. Primary and secondary poisoning by anticoagulant rodenticides of non-target animals in Spain. Sci. Total Environ. 420: 280–288. doi: 10.1016/j.scitotenv.2012.01.028 [DOI] [PubMed] [Google Scholar]
- 45.Serieys L. E. K., Armenta T. C., Moriarty J. G., Boydston E. E., Lyren L. M., Poppenga R. H., Crooks K. R., Wayne R. K., Riley S. P. D.2015. Anticoagulant rodenticides in urban bobcats: exposure, risk factors and potential effects based on a 16-year study. Ecotoxicology 24: 844–862. doi: 10.1007/s10646-015-1429-5 [DOI] [PubMed] [Google Scholar]
- 46.Shore R. F., Birks J. D. S., Afsar A., Wienburg C. L., Kitchener A. C.2003. Spatial and temporal analysis of second-generation anticoagulant rodenticide residues in polecats (Mustela putorius) from throughout their range in Britain, 1992-1999. Environ. Pollut. 122: 183–193. doi: 10.1016/S0269-7491(02)00297-X [DOI] [PubMed] [Google Scholar]
- 47.Shore R. F., Birks J. D. S., Freestone P.1999. Exposure of non-target vertebrates to second-generation rodenticides in Britain, with particular reference to the polecat Mustela putorius. N. Z. J. Ecol. 23: 199–206. [Google Scholar]
- 48.Stansley W., Cummings M., Vudathala D., Murphy L. A.2014. Anticoagulant rodenticides in red-tailed hawks, Buteo jamaicensis, and great horned owls, Bubo virginianus, from New Jersey, U.S.A., 2008–2010. Bull. Environ. Contam. Toxicol. 92: 6–9. doi: 10.1007/s00128-013-1135-z [DOI] [PubMed] [Google Scholar]
- 49.Stone W. B., Okoniewski J. C., Stedelin J. R.1999. Poisoning of wildlife with anticoagulant rodenticides in New York. J. Wildl. Dis. 35: 187–193. doi: 10.7589/0090-3558-35.2.187 [DOI] [PubMed] [Google Scholar]
- 50.Stone W. B., Okoniewski J. C., Stedelin J. R.2003. Anticoagulant rodenticides and raptors: recent findings from New York, 1998-2001. Bull. Environ. Contam. Toxicol. 70: 34–40. doi: 10.1007/s00128-002-0152-0 [DOI] [PubMed] [Google Scholar]
- 51.The IUCN Red List of Threatened SpeciesTM, 2017. Aquila adalberti, Spanish Imperial Eagle. Available from https://www.iucnredlist.org/species/22696042/110743671 [accessed on November 30, 2018].
- 52.The IUCN Red List of Threatened SpeciesTM, 2017. Aquila heliaca, Eastern Imperial Eagle. https://www.iucnredlist.org/species/22696048/117070289 [accessed on November 30, 2018].
- 53.The IUCN Red List of Threatened SpeciesTM, 2017. Falco cherrug, Saker Falcon. https://www.iucnredlist.org/species/22696495/110525916 [accessed on November 30, 2018].
- 54.The IUCN Red List of Threatened SpeciesTM, 2017. Milvus milvus, Red Kite. https://www.iucnredlist.org/species/22695072/131877336 [accessed on November 30, 2018].
- 55.Thomas P. J., Mineau P., Shore R. F., Champoux L., Martin P. A., Wilson L. K., Fitzgerald G., Elliott J. E.2011. Second generation anticoagulant rodenticides in predatory birds: Probabilistic characterisation of toxic liver concentrations and implications for predatory bird populations in Canada. Environ. Int. 37: 914–920. doi: 10.1016/j.envint.2011.03.010 [DOI] [PubMed] [Google Scholar]
- 56.Tome R., Catry P., Bloise C., Korpimaki E.2008. Breeding density and success, and diet composition of Little Owls Athene noctua in steppe-like habitats in Portugal. Ornis Fenn. 85: 22–32. [Google Scholar]
- 57.Vandenbroucke V., Bousquet-Melou A., De Backer P., Croubels S.2008. Pharmacokinetics of eight anticoagulant rodenticides in mice after single oral administration. J. Vet. Pharmacol. Ther. 31: 437–445. doi: 10.1111/j.1365-2885.2008.00979.x [DOI] [PubMed] [Google Scholar]
- 58.Walker L. A., Llewellyn N. R., Pereira M. G., Potter E. D., Sainsbury A. W., Shore R. F.2013. Anticoagulant rodenticides in predatory birds 2011: a Predatory Bird Monitoring Scheme (PBMS) report.:1–29. http://nora.nerc.ac.uk/id/eprint/500093/ [accessed on November 30, 2018].
- 59.Walker L. A., Llewellyn N. R., Pereira M. G., Potter E. D., Sainsbury A. W., Shore R. F.2015. Anticoagulant rodenticides in sparrowhawks: a Predatory Bird Monitoring Scheme (PBMS) report.:1–29. http://nora.nerc.ac.uk/id/eprint/511023/ [accessed on November 30, 2018].
- 60.Walker L. A., Shore R. F., Turk A., Pereira M. G., Best J.2008. The Predatory Bird Monitoring Scheme: identifying chemical risks to top predators in Britain. Ambio 37: 466–471. doi: 10.1579/0044-7447(2008)37[469:TPBMSI]2.0.CO;2 [DOI] [PubMed] [Google Scholar]
- 61.Walker L. A., Turk A., Long S. M., Wienburg C. L., Best J., Shore R. F.2008. Second generation anticoagulant rodenticides in tawny owls (Strix aluco) from Great Britain. Sci. Total Environ. 392: 93–98. doi: 10.1016/j.scitotenv.2007.10.061 [DOI] [PubMed] [Google Scholar]
- 62.Watanabe K. P., Saengtienchai A., Tanaka K. D., Ikenaka Y., Ishizuka M.2010. Comparison of warfarin sensitivity between rat and bird species. Comp. Biochem. Physiol. Part C Toxicol. Pharmacol. 152: 114–119. [DOI] [PubMed] [Google Scholar]
- 63.Zawadzka D., Zawadzki J.2001. Breeding Populations and Diets of the Sparrowhawk Accipiter nisus and the Hobby Falco subbuteo in the Wigry National Park (Ne Poland). Acta Ornithol. 36: 25–31. doi: 10.3161/068.036.0111 [DOI] [Google Scholar]