

ABSTRACT
Diverse signaling cues and attendant proteins work together during organogenesis, including craniofacial development. Lip and palate formation starts as early as the fourth week of gestation in humans or embryonic day 9.5 in mice. Disruptions in these early events may cause serious consequences, such as orofacial clefts, mainly cleft lip and/or cleft palate. Morphogenetic Wnt signaling, along with other signaling pathways and transcription regulation mechanisms, plays crucial roles during embryonic development, yet the signaling mechanisms and interactions in lip and palate formation and fusion remain poorly understood. Various Wnt signaling and related genes have been associated with orofacial clefts. This Review discusses the role of Wnt signaling and its crosstalk with cell adhesion molecules, transcription factors, epigenetic regulators and other morphogenetic signaling pathways, including the Bmp, Fgf, Tgfβ, Shh and retinoic acid pathways, in orofacial clefts in humans and animal models, which may provide a better understanding of these disorders and could be applied towards prevention and treatments.
KEY WORDS: Orofacial clefts, Cleft lip, Cleft palate, Wnt, Bmp, Fgf, Tgfβ, Shh, Retinoic acid, Epigenetics, Crosstalk
Summary: Investigation of the crosstalk between the Wnt and other signaling pathways will improve understanding of orofacial cleft development and provide opportunities for treatment and prevention.
Introduction
Orofacial clefts, mainly cleft lip and/or cleft palate, are among the commonest structural birth defects (Tolarova and Cervenka, 1998; Mossey et al., 2009; Shkoukani et al., 2014; Roosenboom et al., 2015; Kousa and Schutte, 2016). The occurrence of orofacial clefts varies with geographic and ethnic background and with socioeconomic status, with an average rate of 1 in 700 newborns or a range of 0.5-2.6 per 1000 live births (Vanderas, 1987; Croen et al., 1998; Carmichael et al., 2003; Panamonta et al., 2015). Children born with orofacial clefts have severe feeding problems, speech difficulties, frequent middle ear infections and dental defects (Mossey et al., 2009). The long-term and multidisciplinary treatments for these problems are a heavy burden for patients and the healthcare system. Orofacial clefts can either be syndromic or non-syndromic, sporadic or familial (see Glossary, Box 1), and their etiology involves a combination of genetic and environmental risk factors (Tolarova and Cervenka, 1998; Mossey et al., 2009). To date, more than 100 genes have been associated with orofacial clefts (Gritli-Linde, 2007; Juriloff and Harris, 2008; Bush and Jiang, 2012; Iwata et al., 2012), but the underlying mechanisms of these associations remain poorly understood. Mutant mouse models have provided a powerful tool to examine the roles of various genes in contributing to orofacial clefts.
Box 1. Glossary.
C6 motif: the six-amino-acid C-terminal domain of Axin proteins. It is implicated in JNK activation, but has no effect on Wnt signaling.
Epithelial-mesenchymal transition (EMT): the induction of adhesive epithelial cells to become migratory and proliferative cells during developmental processes, including in palatogenesis.
Goltz-Gorlin syndrome: a rare genetic disorder, also known as focal dermal hypoplasia (FDH), characterized by distinctive skin abnormalities, including CLP in some cases, and other defects that affect eyes, teeth, and the skeletal, urinary, gastrointestinal, cardiovascular and central nervous systems. Mutations in PORCN, an upstream regulator of Wnt signaling, are associated with FDH.
Maxillary prominences: a pair of developmental structures at the lateral edges of the oral cavity that give rise to the upper jaw elements, including the maxillary bone.
Medial edge epithelium (MEE): the distalmost edge of the palatal epithelium that surrounds the proliferating mesenchyme. On each palatal shelf, this layer will meet and fuse during secondary palatogenesis.
Midline epithelial seam (MES): the layer of epithelial cells that separates the two lateral pools of the mesenchyme after palatal shelf fusion. MES cells undergo apoptosis to allow the formation of a continuous mesenchyme layer across the secondary palate.
Nasal prominences: two pairs of medial and lateral extensions derived from the unpaired frontonasal prominence during early craniofacial development, which fuse on either side with the maxillary prominences to form the primary palate and nostrils, and separate the nasal cavity from the oral cavity.
Neurulation: a stage in vertebrate embryogenesis in which the neural plate folds to form the neural tube.
Palatal shelf: a pair of palatal structures elongated from the maxillary prominences between the nasal prominence and the first branchial arch/mandibular process, which eventually fuse to separate the oral and nasal cavities.
Primary palate: a rostralmost palatal structure formed by the fusion of the nasal and maxillary prominences to separate the nasal pits from the oral cavity.
Regulator of G protein signaling (RGS) domain: a motif required for the protein's activity in accelerating the GTPase activity of G-proteins. The RGS domain in Axin proteins is required for binding APC in Wnt signaling.
Robinow syndrome: congenital syndrome characterized by craniofacial, skeletal and urogenital defects, which frequently includes orofacial clefts and has been associated with mutations in noncanonical Wnt signaling genes, including WNT5A and ROR2.
Rugae: the series of ridges produced by folding of the anterior wall of the palate behind the incisive papillae.
Secondary palate: a roof structure of the oral cavity that arises from the fusion of the left and right palatal shelves posterior to the primary palate.
Sporadic or familial CLP: occurrence of CLP within families or close relatives is referred to as familial CLP, whereas appearance of the phenotype without apparent genetic predisposition is termed sporadic CLP.
Submucous cleft palate: a form of cleft palate in which the two palatal shelves incompletely fuse such that the oral and nasal cavities are separated from each other by soft tissue, but a continuous layer of bone has not developed across the midline.
Syndromic or non-syndromic CLP: CLP patients carrying additional dysmorphic or clinical features are syndromic; non-syndromic CLP is not associated with other phenotypes.
Tetra-amelia syndrome: a congenital syndrome characterized by limb malformation, often coupled with craniofacial and urogenital defects, associated with WNT3 mutations.
Unilateral and bilateral CLP: a cleft can occur either at one (unilateral) or both sides (bilateral) of the face.
Van der Woude syndrome: a congenital syndrome characterized by craniofacial, limb, and cardiac defects, associated with mutations in the transcription factors downstream of canonical Wnt signaling.
Murine and human facial formation follow a similar developmental trajectory, and facial structures arise from several primordial tissues as described below (Francis-West et al., 1998; Schutte and Murray, 1999; Jiang et al., 2006; Szabo-Rogers et al., 2010; Suzuki et al., 2016). Facial primordia begin to form as early as the fourth week of gestation in humans or embryonic day (E) 9.5 in mice, following the migration of cranial neural crest cells into the frontonasal prominence, paired maxillary prominences (Box 1) and paired mandibular prominences (Cordero et al., 2011). By the fifth week, the medial and lateral nasal prominences (Box 1) outgrow rapidly on either side of the nasal pit. At the ventral junction region, these nasal prominences will subsequently fuse with the maxillary prominence to establish the upper jaw structures, including the upper lip, primary palate (Box 1) and nose. Disruption of any of these early craniofaciogenic processes may result in cleft lip with or without cleft palate (CLP). Secondary palate (Box 1) formation is a multifaceted process involving a shift in growth orientation by the palatal shelves (Box 1) (Lough et al., 2017).
In mice, the palatal shelves first emerge from the maxillary prominences at E11.5 and continue to proliferate, elongating ventrally between E12 and E14 (Bush and Jiang, 2012). The elongating palatal shelves consist of mesenchymal tissue with an external epithelial layer. Epithelial-mesenchymal interactions (EMIs) allow communication between the two layers and are important for cell growth and differentiation during many craniofacial developmental processes, including facilitating epithelial-mesenchymal transition (EMT; Box 1) within the palatal shelves during palatogenesis (Sun et al., 1998; Lan and Jiang, 2009; Levi et al., 2011; Santosh and Jones, 2014). The palatal shelves then elevate and continue to grow horizontally toward the midline, which entails significant extracellular matrix remodeling (Bush and Jiang, 2012), until they fuse along the medial edge epithelium (MEE; Box 1) at E14.5-E15. The palatal shelves at the midline fuse both anteriorly and posteriorly from the initial point of contact in a zipper-like manner to form a midline epithelial seam (MES; Box 1). Disintegration of the MES, which may involve apoptosis, EMT and cell migration, is required to establish palatal confluence (Bush and Jiang, 2012). At E15.5-E16.5, the palatal shelves fuse with the nasal septum and the primary palate, separating the nasal and oral cavities, which are required for breathing and feeding after birth (Gritli-Linde, 2007). Disruptions during any stage of palatogenesis can result in a cleft palate (Dixon et al., 2011). Although the mechanisms that drive palatogenesis are believed to be conserved among mammals, differences in the morphological structures, and in the interactions that occur during palatal closure, exist between species (Yu et al., 2017). An extensive list of different mouse models for cleft lip and/or cleft palate has been previously reviewed elsewhere (Gritli-Linde, 2007; Gritli-Linde, 2008; Juriloff and Harris, 2008; Funato et al., 2015). However, mutations in specific genes do not always produce the same phenotype in humans and mouse models (Gritli-Linde, 2008).
Wingless-type MMTV integration site (Wnt) signaling is required for body axis patterning, cell fate specification, cell proliferation and cell migration during embryonic development (Kimura-Yoshida et al., 2005; Komiya and Habas, 2008; Basson, 2012; Clevers and Nusse, 2012; Perrimon et al., 2012; Hikasa and Sokol, 2013; Clevers et al., 2014; Nusse and Clevers, 2017). Wnt signaling (see Box 2) is active in most tissues during craniofacial development (Mani et al., 2010), and includes multiple distinct pathways that are activated by the binding of the secreted Wnt ligand proteins to a complex receptor system. Wnts bind to the frizzled (Fzd) receptors along with the co-receptors, such as members of the low-density lipoprotein receptor-related protein (Lrp) or receptor tyrosine kinase-like orphan receptor (Ror) families, at the surface of the Wnt-responding cells (Fig. 1, Box 2). The ligand-receptor complex interacts with cytoplasmic proteins, such as the axis inhibition (Axin) and disheveled (Dvl) proteins, to trigger intracellular signaling (Wallingford and Habas, 2005; Niehrs, 2012; Stamos and Weis, 2013; Bernatik et al., 2011) (Fig. 1, Box 2). Wnt pathways are broadly classified as β-catenin-dependent canonical and β-catenin-independent non-canonical pathways, such as the planar cell polarity (PCP) pathway (Box 2) and the Wnt/Ca2+ pathway (Komiya and Habas, 2008; Gao and Chen, 2010). This Review discusses the role of Wnt signaling and its crosstalk with other signaling pathways in orofacial cleft etiology and related developmental processes, which may provide a better understanding of basic mechanisms and future translational applications.
Box 2. Wnt signaling.
Wnts are secreted lipid-modified signaling proteins that are evolutionally conserved and play vital roles in development, homeostasis and disease. Nineteen Wnt ligand proteins encoded by respective genes in mammalian genomes act through a variety of receptors and co-receptors, including ten seven-transmembrane frizzled (Fzd) receptors, two single-transmembrane co-receptors Lrp5/6, and the receptor tyrosine kinase-like receptors Ror1/2 and Ryk.
In the Wnt-producing cells, the nascent Wnt proteins are palmitoylated by porcupine O-acyltransferase (Porcn), followed by their secretion in the extracellular matrix via Wnt ligand secretion mediator (Wls; Fig. 1). Wnt signaling is initiated when a secreted Wnt ligand binds to a Fzd receptor along with a Lrp co-receptor in the canonical pathway, or to a tyrosine kinase-like Ror or Ryk receptor in the non-canonical pathway. The ligand-receptor interaction at the surface of the Wnt-responding cell is modulated by various positive or negative regulatory factors and is transmitted through numerous intracelluar molecules. Three major signaling pathways have been demonstrated downstream of the initial Wnt ligand-receptor interaction: the canonical Wnt/β-catenin signaling pathway, the non-canonical planar cell polarity (PCP) pathway, and the Wnt/Ca2+ pathway, which is less understood.
Canonical Wnt/β-catenin pathway: when Wnts are absent, intracellular β-catenin is constantly phosphorylated for degradation by the glycogen synthase kinase Gsk3β in the β-catenin destruction complex, which also includes the tumor suppressing Axin proteins and adenomatous polyposis coli (APC), the casein kinase CK1, the protein phosphatase 2A (PP2A) and the E3-ubiquitin ligase β-TrCP. Upon the binding of a Wnt ligand to a Fzd receptor, the Fzd recruits a Dvl cytoplasmic phosphoprotein and a Lrp co-receptor recruits an Axin, which inhibits the destruction complex. This stabilizes cytoplasmic β-catenin, resulting in its accumulation and translocation into the nucleus. There it binds to Tcf/Lef1 transcription factors to regulate the transcriptional activation of critical Wnt target genes in various cells/tissues, such as the orofacial cleft-associated genes Msx1/Msx2 in orofacial primordia (Fig. 1).
Non-canonical PCP pathway: The binding of a Wnt ligand to Ror or Ryk receptors promotes the interaction of a Dvl with disheveled-associated activator of morphogenesis 1 (Daam1), which activates several downstream GTPases, including the Rac proteins and ras homolog family member A (RhoA). This results in the restructuring of actin to change cell shape, polarity and movement. Dvl can also activate phospholipase C to generate inositol triphosphate, which activates the release of Ca2+ to trigger a number of downstream effects, such as cell migration and proliferation.
Fig. 1.

Key components and potential pharmacological targets of Wnt signaling implicated in orofacial clefts. Wnt proteins are lipid modified by Porcn and secreted in the extracellular matrix by Wls from the Wnt-producing cells. Wnt9b and Wnt5a are representative orofacial cleft-associated ligands in the canonical and non-canonical Wnt signaling pathways, respectively. In the canonical pathway, Wnt9b may bind to receptor Fzd1/Fzd2 and co-receptor Lrp6, which respectively recruit Dvl and Axin proteins, disrupting the β-catenin destruction complex that includes the glycogen synthase kinase Gsk3β in the Wnt-responding cells. This leads to intracellular accumulation of β-catenin and its translocation to the nucleus, where, together with the Tcf/Lef1 transcription factors, it regulates the expression of downstream target genes that are critical during lip and palate formation, such as Msx1 and Msx2. The Lrp6 co-receptor can be regulated positively by Rspo2 and negatively by Dkk proteins. In the non-canonical pathway, Wnt5a binds to co-receptor Ror2 and/or Ryk to suppress Prickle1, which in turn activates Dvl proteins (involved in both canonical and non-canonical pathways) and facilitates the cytoskeletal rearrangements during palatogenesis. It is unclear whether Wnt5a also binds to Fzd/Lrp proteins to regulate canonical Wnt signaling during orofacial development. The key components of the signaling cascade that are underlined in the diagram could be targeted by small molecules to modulate Wnt signaling. Orofacial cleft-associated signaling molecules are marked with symbols according to their resulting phenotype (see figure key).
WNT signaling genes associated with orofacial clefts in humans
Both syndromic and non-syndromic orofacial clefts have been attributed to mutations of various WNT signaling component genes (Table 1). Nascent WNT proteins are lipid modified by the enzyme porcupine O-acyltransferase (PORCN) within the endoplasmic reticulum of the WNT-producing cell and subsequently transported by WNT ligand secretion mediator (WLS, also known as GPR177) through the Golgi apparatus to the cell surface for secretion (Port and Basler, 2010; Barrott et al., 2011) (Fig. 1, Table 1, Box 2). An extensive number of mutations throughout the coding region and large gene rearrangements of PORCN have been identified in focal dermal hypoplasia or Goltz–Gorlin syndrome (Box 1), which includes orofacial clefts (Table 1) (Lombardi et al., 2011). However, a role for PORCN in non-syndromic cleft lip and palate (NSCLP), or a role for WLS in human orofacial clefts, has not been demonstrated. A homozygous nonsense mutation in WNT3 has been correlated with orofacial clefts and tetra–amelia syndrome (Box 1) (Niemann et al., 2004). Meanwhile, multiple non-coding single-nucleotide polymorphisms (SNPs) in WNT3 have been associated with NSCLP in a wide range of populations, including Latin American, European and Chinese (Chiquet et al., 2008; Nikopensius et al., 2010; Nikopensius et al., 2011; Mostowska et al., 2012; Lu et al., 2015). Yet, in some populations, such as Caucasian Brazilian, the relationship between WNT3 variants and NSCLP remains unclear (Fontoura et al., 2015; Machado et al., 2016). Intriguingly, Nikopensius et al. (2010) reported a potential epistatic interaction between WNT3 and collagen, type II, alpha 1 (COL2A1), an important gene in the production of collagen. Mutations in either gene are associated with NSCLP (Nikopensius et al., 2010; Nikopensius et al., 2011), and COL2A1 mutations also cause Stickler Syndrome, which frequently includes a cleft palate only (CPO) phenotype (Hoornaert et al., 2010), suggesting a relationship between canonical WNT signaling and the extracellular matrix during palatogenesis. WNT3 is clustered at 17q21.31-17q21.32 with WNT9B, variants of which were also associated with a predisposition to NSCLP (Nikopensius et al., 2011; Fontoura et al., 2015). Several SNPs near the WNT6-WNT10A cluster at the 2q35 region of chromosome 2 were associated with either CLP or CPO (Beaty et al., 2006), while more recently, a missense mutation within WNT10A was identified in a Chinese NSCLP cohort (Feng et al., 2014). A WNT7A variant containing a missense SNP was identified as a contributor to NSCLP in several heterozygous members of a multi-case family (Pengelly et al., 2016).
Table 1.
Summary of WNT signaling genes associated with orofacial clefts in humans and animal models

Intronic and 3′UTR SNPs in the prototypical canonical WNT ligand gene WNT3A have also been identified in patients with NSCLP within European American, Hispanic and Chinese populations, with homozygosity presenting an increased risk over heterozygosity for each allele identified in the Chinese cohort (Chiquet et al., 2008; Yao et al., 2011) (Table 1). Association analyses within the European American cohort also implicated WNT11, while an association between SNPs in the non-coding regions downstream of WNT3A and the non-canonical WNT5A was reported in the Hispanic population (Chiquet et al., 2008) (Table 1). An intronic WNT5A SNP was associated with unilateral CLP (Box 1) with marginal significance in a Caucasian Brazilian population (Menezes et al., 2010). Robinow syndrome (Box 1), which frequently includes cleft palate, is associated with mutations in WNT5A (Person et al., 2010), along with mutations in the co-receptor gene receptor tyrosine kinase-like orphan receptor 2 (ROR2) (Afzal et al., 2000; van Bokhoven et al., 2000) and the signal transducer DVL1 (Bunn et al., 2015; White et al., 2015), indicating the importance of a non-canonical WNT5A/ROR2/DVL1 signaling cascade in human palatogenesis. A missense mutation and rare haplotypes of another non-canonical co-receptor gene, receptor-like tyrosine kinase (RYK), have also been linked to NSCLP in Vietnamese and Japanese patients (Watanabe et al., 2006). Additionally, several non-coding variants and one missense SNP of PRICKLE1 have demonstrated association with non-canonical WNT signaling and human NSCLP (Yang et al., 2014).
Analysis of an African American family with 11 members displaying NSCLP identified a variant of the WNT receptor gene FZD6 with an intronic mutation that creates a protein-binding site, resulting in decreased expression and contributing to CLP (Cvjetkovic et al., 2015). Among other FZD genes, a nonsense mutation of FZD2 was identified in a family with omodysplasia that includes CLP (Saal et al., 2015). By contrast, frameshift, nonsense and missense mutations in the WNT co-receptor gene LRP6 have been associated with orofacial clefts and tooth agenesis (Basha et al., 2018; Ockeloen et al., 2016), suggesting that deficient LRP6-mediated canonical WNT signaling has a crucial role in CLP pathogenesis. However, de novo nonsense and frameshift mutations in the key canonical WNT signaling mediator gene catenin beta 1 (CTNNB1; encoding β-catenin) were linked with abnormal craniofacial features, but not with orofacial clefts (Tucci et al., 2014). Conversely, analysis of variants of the β-catenin destruction complex genes AXIN2 and glycogen synthase kinase 3 beta (GSK3B) in NSCLP families across multiple populations identified intronic SNPs that contribute to orofacial clefts (Letra et al., 2009; Letra et al., 2012; Vijayan et al., 2018) (Table 1), suggesting that excessive WNT/β-catenin signaling also contributes to CLP pathogenesis. Nevertheless, gene association studies in humans with orofacial clefts have proven challenging, complicated by the fact that the same variants can be associated with orofacial clefts in one population but not in others. Therefore, animal models, especially the mutant mouse model, have a crucial role in investigating the genetic mechanisms of orofacial clefts in mammals.
Wnt signaling genes as the cause of orofacial clefts in animal models
Mutations in various Wnt signaling genes cause orofacial clefts in animal models (He and Chen, 2012) (Table 1). The following discussion highlights animal models of orofacial clefts involving mutations in both canonical and non-canonical Wnt signaling components, from ligand secretion through signal transduction, focusing predominantly on mouse models.
Mouse models with mutations in regulatory genes upstream of Wnt
Absence of Porcn or Wls from Wnt-producing cells (Fig. 1) results in Wnt protein retention, which leads to Wnt signaling failure (Barrott et al., 2011). Conditional ablation of Porcn in neural crest cells results in defective facial formation in mice, including CLP (Bankhead et al., 2015) (Fig. 1). Previous studies suggest that Wls is required for Wnt/β–catenin signaling during craniofacial development (Fu et al., 2011). Facial ectodermal/epithelial ablation of either β-catenin or Wls arrests the formation of orofacial primordia (Wang et al., 2011; Zhu et al., 2016), and conditional knockout of Wls in craniofacial neural crest cells with a Wnt1-driven Cre recombinase causes cleft palate (Liu et al., 2015) (Table 1). Additionally, Rspo2, a member of the R-spondin family, is a well-known enhancer of canonical Wnt signaling (Fig. 1). Rspo2 loss-of-function mice exhibit cleft palate with a partially penetrant cleft lip, along with mandibular hypoplasia and maxillary and mandibular skeletal deformation, which are caused by defective patterning and morphogenesis of the first pharyngeal arch due to altered EMI (Yamada et al., 2009; Jin et al., 2011). However, cleft palate in Rspo2-null mice is likely caused by delayed palatal shelf (Box 1) elevation, a possible secondary effect of aberrant mandible and tongue morphogenesis (Jin et al., 2011). Nevertheless, Wnt signaling may mediate Tgfβ signaling to regulate EMI in muscle development of the soft palate (Iwata et al., 2014).
Mouse models with mutations in canonical Wnt signaling genes
Among the 19 Wnt ligands, Wnt2, Wnt2b, Wnt4, Wnt5a, Wnt5b, Wnt6, Wnt7b, Wnt9a, Wnt10a, Wnt10b, Wnt11 and Wnt16 are expressed in the palatal primordia during palatogenesis (Warner et al., 2009). Five Wnts appear to be temporally regulated in embryonic palatal tissue, showing more than 2.0-fold changes in expression levels, either between E12.5 and E13.5, or between E13.5 and E14.5. Of these five ligands, Wnt4, Wnt10a and Wnt10b are expressed in epithelial tissues, while Wnt2 and Wnt16 are expressed in the mesenchyme (Warner et al., 2009). However, the roles of these temporarily expressed Wnts in palatogenesis remain unclear. By contrast, Wnt6 has been demonstrated to play a role in palatal shelf elongation and elevation through the activation of the β-catenin pathway, promoting cell proliferation in the palatal mesenchyme (Jiang et al., 2017) (Table 1).
Wnt9b might activate the canonical Wnt signaling pathway during midfacial development (Lan et al., 2006). Wnt9b-null mice die perinatally, exhibiting incompletely penetrant CLP (Carroll et al., 2005; Juriloff et al., 2006; Ferretti et al., 2011) (Table 1, Fig. 1), while ablation of Wnt9b in the facial ectoderm also causes CLP (Jin et al., 2012). These findings suggest a key role of facial ectodermal and epithelial Wnt/β-catenin signaling in primary lip and palate formation and fusion. In addition, Wnt3 may also regulate midfacial development, as well as lip fusion, through the canonical Wnt pathway, with both Wnt9b and Wnt3 playing distinct roles during midfacial morphogenesis (Lan et al., 2006). Wnt3-null embryos do not survive beyond E10.5, while morphological differences from wild-type embryos become apparent from E6.5 onward (Liu et al., 1999). In vitro experiments further suggest that Wnt3 and Wnt9b may activate canonical Wnt signaling during palatogenesis through the receptors Fzd1 and Fzd2 (Lan et al., 2006; Yu et al., 2010). Palatal shelves fail to close in doubly homozygous Fzd1 and Fzd2 knockout mice with complete penetrance (Yu et al., 2010) (Table 1, Fig. 1), while Fzd7 is highly redundant with Fzd2 during palatogenesis (Yu et al., 2012). Canonical Wnt signaling through the co-receptor Lrp6 plays an indispensable role in primary lip and palate formation and fusion (Song et al., 2009; Zhou et al., 2010). Lrp6-deficient mutant mouse embryos exhibit fully penetrant CLP as a consequence of diminished Wnt signaling and disrupted expression of downstream target genes in the orofacial primordia (Song et al., 2009) (Fig. 1).
While conditional loss of function of β-catenin in palatal epithelial cells leads to cleft palate, conditional gain of function of β-catenin in the epithelium also leads to cleft palate and aberrant fusion between the palate shelf and mandible (He et al., 2011), suggesting crucial roles of epithelial Wnt signaling in palatal shelf fusion. Moreover, homozygous knockout of Gsk3b, which encodes a β-catenin-degrading enzyme in the canonical Wnt signaling pathway, results in mice displaying full cleft palate (Liu et al., 2007) (Table 1, Fig. 1), suggesting that excessive β-catenin signaling also causes cleft palate in these mouse models.
Axin1 is another component of the β-catenin destruction complex and therefore a negative regulator of Wnt signaling (Fig. 1). Early embryonic lethality is observed in homozygous Axin1 mutant mouse embryos carrying alleles with deletions in either the regulator of G protein signaling (RGS) domain (Box 1) or the C6 motif (Box 1) that encodes the six C-terminal amino acids (Axin1ΔC6) (Chia et al., 2009). Intriguingly, many mouse embryos with compound mutant alleles of Axin1ΔC6/ΔC6 and Ctnnb1+/− can survive to term but develop craniofacial defects, including CLP (Chia et al., 2009) (Table 1, Fig. 1). This suggests that diminished Wnt/β-catenin signaling can partially rescue the early lethality that is likely caused by excessive β-catenin signaling, but it cannot rescue the CLP phenotype that may be caused by both excessive β-catenin and defective JNK signaling (Chia et al., 2009). Together, these findings highlight the importance of appropriate spatiotemporal control of Wnt/β-catenin signaling and the complexity of the regulatory processes in lip and palate development.
Mouse models with mutations in non-canonical Wnt signaling genes
Wnt5a acts through the non-canonical Wnt pathway to alter directional cell movements (Liu et al., 2015). Wnt5a-null mouse embryos exhibit cleft palate (Table 1, Fig. 1), along with other phenotypes, such as defective outgrowth of the snout, tongue, mandible, limb, tail and other skeletal defects, leading to perinatal lethality (Yamaguchi et al., 1999; Li et al., 2002; Yang et al., 2003; Cervantes et al., 2009; Tai et al., 2009; Buttler et al., 2013; Okamoto et al., 2014). Wnt5a plays a key role in the migration of mesenchymal cells during palatogenesis (Xiao et al., 2005; He et al., 2008), possibly acting through Ror2, which is expressed in the mesenchyme of the secondary palate (Schwabe et al., 2004). Studies suggested that Wnt5a binds to the cysteine-rich domain of Ror2 to activate the non-canonical Wnt pathway, interacting both physically and functionally (Oishi et al., 2003). In mesenchymal cell culture, cell migration seems to be driven by the Wnt5a-Ror2-Kif26b signaling cascade (Susman et al., 2017), further suggesting the significance of this non-canonical Wnt signaling cascade in palatogenesis. Furthermore, phosphorylation of the Wnt signal transducer Dvl2 seems to be triggered by the Wnt5a-Ror2 pathway, and Dvl2 may be the molecular switch that allows Wnt5a to activate both non-canonical and canonical Wnt pathways (Ho et al., 2012).
Ror2 knockout mice display craniofacial defects, including cleft palate, further implicating this cascade in the etiology of non-canonical Wnt-signaling-caused orofacial clefts (Schwabe et al., 2004). It has also been suggested that the Ryk receptor may interact with Ror2 to bind Wnt5a (Oishi et al., 2003), and mutations in Ryk also cause cleft palate in mice (Halford et al., 2000) (Table 1, Fig. 1). In addition, ablation of the non-canonical Wnt signaling molecule Prickle1 causes cleft palate and limb defects (Yang et al., 2014) (Table 1, Fig. 1), which are similar to those of Wnt5a mutants (He et al., 2008). However, Prickle1 mutants present less severe limb defects than Wnt5a mutants, implying that the transduction of Wnt5a signaling might not act through Prickle1 alone. Similarly to Wnt5a mutants, Prickle1 knockout mice present with improper sonic hedgehog (Shh) expression during palatogenesis (Yang et al., 2014). Furthermore, Prickle1 has been shown to act downstream of Wnt5a and interact with Dvl2, and Prickle1 mutants display characteristics that resemble Robinow syndrome (Liu et al., 2014a) (Fig. 1). Thus, a signaling cascade of Wnt5a-Ror2-Prickle1/Dvl2 might be crucial for proper tissue growth and morphogenesis during palatogenesis in mice.
Zebrafish models
Although mouse models have vastly contributed to our understanding of Wnt signaling in palatogenesis, other models, such as the zebrafish, provide unique insight into craniofacial formation and the basic requirements for palate formation (Duncan et al., 2017). Canonical Wnt signaling through Lrp5 is required for appropriate cranial neural crest cell migration, but not their induction, and for craniofacial morphogenesis in zebrafish (Willems et al., 2015). Wnt9a is expressed in the zebrafish pharyngeal arch, implicating its role during craniofacial development (Curtin et al., 2011). Interestingly, Wnt9a has been shown to play a role in palatogenesis in fish, but not in mammals (Dougherty et al., 2013; Rochard et al., 2016), suggesting taxon-specific Wnt signaling functions in palatogenesis. Wnt5b is thought to assume a similar craniofacial role in zebrafish that Wnt5a plays in mammals (Topczewski et al., 2011). Non-canonical Wnt signaling mediated by epithelial Wnt5b and Wnt9b was demonstrated to stimulate the PCP pathway in chondrocytes, facilitated by Secreted frizzled-related protein 3 (Sfrp3, also known as Frzb) and Glypican 4 (Gpc4) activity during palate extension (Rochard et al., 2016). Additionally, morpholino-based knockdown of Wnt3a and Tubulointerstitial nephritis antigen-like 1 (Tinagl1), a Wnt-interacting extracellular matrix protein, results in defects of the pharyngeal arch and ethmoid plate, which corresponds to the mammalian palate (Neiswender et al., 2017). Loss of function of the Wnt modulator Sfrp3 in zebrafish results in the failure of anterior palate extension, further highlighting the role of Wnt signaling in palatal extension and convergence in zebrafish (Kamel et al., 2013).
Chick models
Orofacial clefts have also been observed in chick embryos (Abramyan and Richman, 2018), where Wnt signaling similarly mediates the growth of primordial facial processes and the developing palate, in which six epithelial and three mesenchymal Wnt ligands, as well as several other pathway components, are expressed (Geetha-Loganathan et al., 2009). Wnt11 was shown to activate the non-canonical Wnt/PCP pathway and inhibit canonical Wnt/β-catenin signaling in the maxillary prominence, and its ectopic expression results in a notched beak/cleft lip phenotype (Geetha-Loganathan et al., 2014). Similarly, overexpression of Wnt2b leading to ectopic expression of msh homeobox 1 (Msx1) results in a foreshortened rostrum/upper beak, corresponding with a mammalian CLP phenotype (Medio et al., 2012).
Frog models
Recently, the suitability of Xenopus embryos for transplanting tissue and local chemical perturbation have provided a suitable clefting model (Dickinson, 2016). Although several studies have assessed the involvement of various biochemical pathways and factors in frog palatal clefts, including retinoic acid and folate metabolism, few studies have probed Wnt signaling during orofacial development in this organism (Dickinson and Sive, 2009; Kennedy and Dickinson, 2012; Wahl et al., 2015).
In vitro models
Another means by which investigators study secondary palate fusion is by culturing palatal shelf explants and assaying their ability to complete the final stages of palate fusion in vitro, such as adherence and formation of the MES and subsequent apoptosis to establish mesenchymal confluence (Ibrahim et al., 2015). Although not directly analogous to in vivo palatogenesis, this approach has helped examine the roles of many factors and processes that are important for the fusion process specifically, including Wnt11 and its dependence on Fgf signaling in palatal closure (Lee et al., 2008).
Crosstalk between Wnt signaling, cell adhesion molecules and transcription factors in orofacial clefts
Because β-catenin has dual roles in Wnt signaling and in cell adhesion, it remains unclear which functions of β-catenin are required for which stages of orofacial development. The roles of other cell-cell adhesion proteins, such as E-cadherin (Cdh1), during palatogenesis remain to be elucidated (reviewed in Lough et al., 2017). Mutations in CDH1 have been associated with an increased risk for non-syndromic orofacial clefts in humans (Rafighdoost et al., 2013; Vogelaar et al., 2013; Bureau et al., 2014; Hozyasz et al., 2014; Brito et al., 2015; Ittiwut et al., 2016; Song et al., 2017). In mouse models, Cdh1 knockout is embryonic lethal and mutant embryos do not develop beyond E10.5 (Garcia-Higuera et al., 2008). Conditional Cdh1 knockout in neural crest cells results in craniofacial defects related to bone development, including a shortened snout, abnormal teeth and twisted nasal bones (Shao et al., 2016). However, these mutants did not develop orofacial clefts. A possible interaction between CDH1 and the WNT signaling pathway has been suggested in human endometrial epithelial cells, where ablation of CDH1 enhances canonical WNT signaling (Zhu et al., 2018) (Fig. 2). Furthermore, increased expression of Cdh1 in mouse maxillary mesenchymal cells during palatogenesis results in a reduction of cytosolic β-catenin (Warner et al., 2016) (Table 2). These studies suggest that Cdh1 may negatively regulate the canonical Wnt/β-catenin signaling pathway in humans and mice (Table 2, Fig. 2).
Fig. 2.
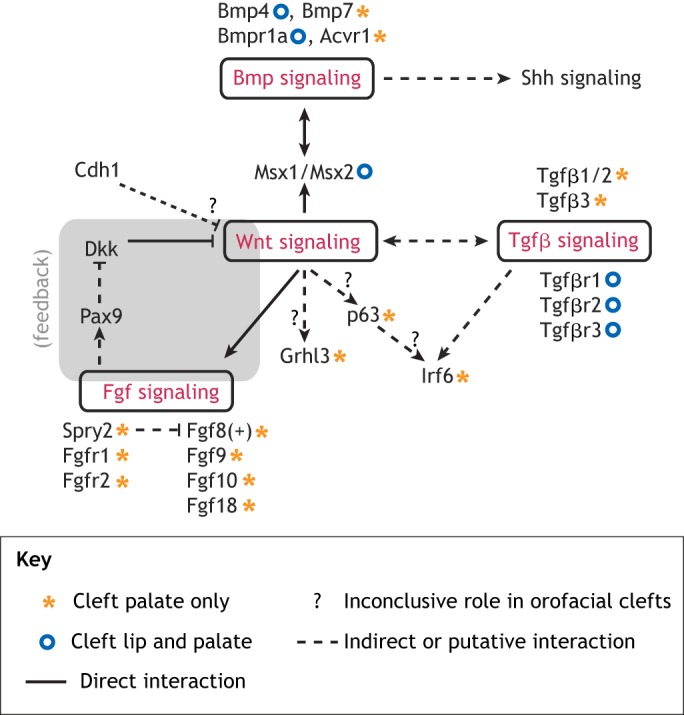
Crosstalk between Wnt, Cdh1, Bmp, Tgfβ and Fgf signaling pathways in orofacial clefts. In lip and palate primordia, the cell adhesion molecule Cdh1 may negatively regulate canonical Wnt signaling, which may interact with Bmp signaling through the common targets Msx1/Msx2. Wnt signaling also regulates Tgfβ signaling in palatogenesis. Tgfβ signaling modulates the orofacial cleft-causing gene Irf6, which may also be regulated by Wnt signaling through p63. Moreover, Wnt signaling can activate Fgf signaling, which modulates Pax9 to repress Dkk protein, an inhibitory ligand of Lrp6 in the canonical Wnt pathway, forming a positive-feedback regulatory loop during orofacial development. The Spry protein family is known to inhibit Fgf signaling. Grhl3 and Irf6 are well-known candidates for Van der Woude syndrome, a syndromic form of CLP. However, their regulation by Wnt has yet to be elucidated. The phenotypic outcomes of the key signaling components, demonstrated in mutant mouse models, are marked as per the figure key.
Table 2.
Wnt pathway-interacting signaling components and transcription factors linked to orofacial clefts

Individuals with mutations in either of the epithelial transcription factors grainyhead-like transcription factor 3 (GRHL3) and interferon regulatory factor 6 (IRF6), detected in families with Van der Woude syndrome (Box 1), tend to present with CLP (de Lima et al., 2009; Peyrard-Janvid et al., 2014). Further investigation in mice suggested that there is no epistatic interaction between these two transcription factors during palatogenesis (Peyrard-Janvid et al., 2014). However, both Grhl3 and Irf6 can be activated in mouse epithelial cells by Wnt/β-catenin signaling (Fig. 2), either directly or indirectly (Ferretti et al., 2011; Kimura-Yoshida et al., 2015). Therefore, Grhl3 and Irf6 may have essential roles during palatogenesis, acting in pathways that diverge from each other downstream of the canonical Wnt pathway and converge again during later stages of palatal development. Irf6 can directly activate Grhl3 (de la Garza et al., 2013), and Grhl3 expression is significantly reduced during embryonic development of Irf6-null mice (Fakhouri et al., 2017) (Table 2). Grhl3-null embryos die shortly after birth, presenting defects in skull morphology (Ting et al., 2005; Goldie et al., 2016). During early embryogenesis, Grhl3 expression is restricted to ectodermal and epidermal lineages (Wang and Samakovlis, 2012; Peyrard-Janvid et al., 2014). The importance of the grainyhead-like gene family, which includes Grhl3, is emphasized by its high level of conservation between fungi and animals, and its conserved role in epithelial specification (Paré et al., 2012; Miles et al., 2017). Exome sequencing-based association studies have revealed several variations in human GRHL3 associated with non-syndromic CPO in almost all populations studied (Leslie et al., 2016; Mangold et al., 2016; Hoebel et al., 2017; Eshete et al., 2018) (Table 2), except in the Han Chinese (He and Bian, 2016), suggesting GRHL3 as a strong candidate gene for non–syndromic CPO. Thus, proper Grhl3 expression in the epithelium during palatogenesis seems to be crucial for normal palate formation (Carpinelli et al., 2017). In mice, a cooperative interaction has been suggested between Grhl2 and Grhl3 during primary neurulation (Box 1) (Rifat et al., 2010), and Grhl3 might act downstream of canonical Wnt signaling during neural tube closure (Kimura-Yoshida et al., 2015). Nevertheless, it remains unclear whether Grhl3 is a direct downstream target of the canonical Wnt/β-catenin signaling, or whether Grhl2 and Grhl3 act cooperatively during palate development.
Analysis of Irf6 expression and function in mouse and chick developmental models suggests that Irf6 might play a role in tissue fusion events during palatogenesis (Knight et al., 2006; Velazquez-Aragon et al., 2016). The Wnt target p63 (also known as TP63), a key regulator of proliferation and differentiation (Truong et al., 2006), inhibits the Wnt signaling output by repressing Wnt/β-catenin responsive elements in target genes (Katoh et al., 2016). p63 may directly activate Irf6 in the facial ectoderm, and a defective pre-B cell leukemia homeobox (Pbx)-Wnt-p63-Irf6 signaling cascade has been suggested in cleft lip formation (Ferretti et al., 2011). Irf6 interacts upstream of the cleft-associated Rho GTPase-activating protein 29 (Arhgap29), which regulates Rho activity downstream of Wnt5a in the PCP pathway (Leslie et al., 2012). Furthermore, several genome-wide association studies have uncovered interactions between IRF6 and other factors linked with orofacial cleft susceptibility. Li and colleagues have identified a three-way gene interaction of SNPs in IRF6, WNT5A and C1orf107 (also known as UTP25, a nucleolar protein), and a separate interaction between IRF6 and WNT2, in association with NSCLP (Li et al., 2015). IRF6 mutations were also associated with SNPs in the actin-binding protein tropomysin (TPM1) and in the axon guidance signaling molecule netrin 1 (NTN1) (Velazquez-Aragon et al., 2016).
Wnt signaling crosstalk with other morphogenetic signaling pathways in orofacial clefts
Wnt signaling does not act in isolation during lip/palate development. Several other signaling pathways are involved in orofacial development, including the fibroblast growth factor (Fgf), bone morphogenic protein (Bmp), transforming growth factor beta (Tgfβ), sonic hedgehog (Shh) and retinoic acid (RA) signaling pathways (Iwata et al., 2011; Bush and Jiang, 2012; Cobourne and Green, 2012; Parada and Chai, 2012; Stanier and Pauws, 2012; Wang et al., 2013; Kurosaka et al., 2014; Okano et al., 2014; Yan et al., 2018; Graf et al., 2016;). This section discusses how these pathways interact with each other and with Wnt signaling.
Wnt-Fgf signaling crosstalk
Wnt/β-catenin signaling activates Fgf8 expression in early facial patterning (Wang et al., 2011), while Fgf8 induces paired box 9 (Pax9) expression during palatogenesis (Neübuser et al., 1997) (Fig. 2). Fgf8 overexpression in mice results in cleft palate, while Fgf10, a loss of which also results in cleft palate, seems to function in cooperation with Wnt signaling (Alappat et al., 2005; Wu et al., 2015). Knockout of other Fgf genes (Fgf9 and Fgf18) and their receptors (Fgfr1 and Fgfr2) has also been associated with a cleft palate phenotype (Liu et al., 2002; Trokovic et al., 2003; Rice et al., 2004) (Fig. 2). Pax9 may feed back into and regulate canonical Wnt/β-catenin signaling in the anterior palatal mesenchyme during palatogenesis. Pax9 ablation causes an increase of the Wnt signaling modulators dickkopf Wnt signaling inhibitor 1 and 2 (Dkk1 and Dkk2), and intravenous delivery of small-molecule Dkk inhibitors can rescue the cleft palate phenotype in utero in Pax9-null mouse embryos (Jia et al., 2017b; Li et al., 2017) (Fig. 2). Likewise, Pax9 seems to target Wnt signaling through the downregulation of β-catenin and Axin2 in the canonical Wnt signaling pathway (Li et al., 2017). Knockout of the human cleft palate-associated transcription factor Tbx22 in mice results in submucous CPO and ankyloglossia (Marçano et al., 2004; Pauws et al., 2009). Tbx22 is at the interface between Bmp and Fgf signaling, being repressed by the former and activated by the latter, downstream of canonical Wnt signaling in the developing palate (Fuchs et al., 2010). Sprouty 2 (Spry2) is also a candidate Wnt/β-catenin target gene (Ordόñez-Morán et al., 2014), and Spry family members inhibit Fgf signaling. Spry2 knockout mice display aberrant growth and movement of palatal shelves, and cleft palate occurs due to a failure of palatal shelf elevation (Welsh et al., 2007; Matsumura et al., 2011) (Fig. 2). These animals show altered expression of the Bmp target and cleft palate gene Msx1, implying a possible link between Fgf and Bmp signaling in the palate (Welsh et al., 2007). Moreover, Spry4 signaling interacts with the Irf6 pathway, which, as we discuss above, is a crucial player in orofacial development (Kousa et al., 2017).
Wnt-Bmp-Shh signaling crosstalk
The homeobox-containing Msx transcription factors function as downstream effectors of Bmp signaling in many developmental processes, including in palatogenesis (Cheng et al., 2003; Tribulo et al., 2003; Hayashi et al., 2006; Parada and Chai, 2012). Bmp signaling directly activates Msx genes at early stages of ectodermal patterning in order to specify the neural crest (Graham et al., 1994; Tribulo et al., 2003). Lrp6-mediated Wnt signaling also regulates Msx1 and Msx2 expression in the orofacial primordia, but Lrp6 ablation does not affect Bmp4 during primary lip/palate formation and fusion, suggesting that the Bmp and Wnt pathways may converge by way of common activation of Msx1/Msx2 (Song et al., 2009) (Fig. 2). Mice with knockouts of either Msx1 or the Bmp receptor Bmpr1a display cleft palate and downregulated Shh signaling. Bmp4 expression in the anterior palate mesenchyme is lost in Msx1-null mice, while transgenic ectopically expressed human BMP4 is able to rescue both Shh activity and the cleft palate phenotype in these animals (Zhang et al., 2002; Baek et al., 2011). This implies that Bmp proteins are the primary effectors of Msx1 activity, and that an Msx1-Bmp-Shh cascade may act downsteam of Lrp6-mediated Wnt signaling to regulate palatogenesis. This link between Bmp and Shh signaling in the palate primordia appears to be mediated by the epithelial transcription factor heart and neural crest derivatives expressed 2 (Hand2) (Xiong et al., 2009).
Many Bmp receptors are expressed in differing patterns along the anterior-posterior (A-P) axis of the palatal shelves during palatogenesis. Submucous cleft palate (Box 1) results from the overexpression of the Bmp receptor activin A receptor-type I (Acvr1) (Noda et al., 2016), so appropriate levels and localization of Bmp pathway activity appear critical for correct tissue responses to palatogenic signaling. Conditional deletion of the Bmp signaling receptors Bmpr1a or Acvr1 in neural crest cells results in multiple craniofacial abnormalities, including submucous cleft palate (Dudas et al., 2004; Saito et al., 2012) (Fig. 2). Interestingly, Wnt9b may regulate Bmp4 during lip fusion (Lan et al., 2006), but this interaction has not been demonstrated during palate fusion. Wnt5a, however, is expressed in a descending gradient from the anterior to the posterior developing palate, and it can act as a negative regulator of Bmp4 in a concentration-dependent manner across the palate (He et al., 2008). Bmp2 seems unaffected by Wnt5a, thus Bmp2 activation may occur in a separate pathway from that of Bmp4 (He et al., 2008). Bmp7 is also expressed in the developing palate and during rugae (Box 1) formation, where it acts downstream of canonical Wnt signaling (Lin et al., 2011), and has been linked with cleft palate in both humans and mice (Kouskoura et al., 2013; Yu et al., 2015). Bmp signaling is upregulated in homeobox A2 (Hoxa2)-null embryos, and Hoxa2 may inhibit palatal osteogenic differentiation from mesenchymal cells via its modulation of Bmp signaling (Iyyanar and Nazarali, 2017). In addition, homeobox protein sine oculis-related homeobox 2 (Six2) likely acts as a downstream effector of Hoxa2 in regulating mesenchymal cell proliferation during secondary palate formation (Okello et al., 2017), and palatal shelves fail to extend to the midline in Six2 knockout mice (Sweat et al., 2018). It remains unclear whether this activity is related to the interaction of Hoxa2 with Bmp. Six2 is known to repress Wnt/β-catenin by binding to T-cell factor/lymphoid enhancer binding factor 1 (Tcf/Lef1) family members during nephrogenesis (Self et al., 2006; Park et al., 2012), but it remains unclear whether Six2 does so during palatogenesis.
Wnt-Tgfβ signaling crosstalk
Epithelial Wnt/β-catenin signaling also regulates Tgfβ signaling. Wnt-mediated Tgfβ3 activation is required for MEE cell apoptosis during palatal shelf closure (He et al., 2011) (Fig. 2), and knockout of all three isoforms of Tgfβ has been associated with cleft palate in mice, in either single or doubly mutant lines (Kaartinen et al., 1995; Sanford et al., 1997; Jin and Ding, 2014) (Fig. 2). Tgfβ1 and Tgfβ3 are semi-redundant, and overexpression of Tgfβ1 can partially rescue the cleft phenotype observed in Tgfβ3-null mice (Yang and Kaartinen, 2007). Mutations in transforming growth factor beta receptor 3 (Tgfβr3, also known as betaglycan), which binds Tgfβ ligands without transducing the signal, cause cleft palate due to reduced cell proliferation and increased apoptosis (Hill et al., 2015) (Fig. 2). By contrast, conditional Tgfβr1 and Tgfβr2 knockout in neural crest cells also causes cleft palate and skull defects due to insufficient cell proliferation (Ito et al., 2003; Dudas et al., 2006) (Table 2, Fig. 2). Tgfβ signaling through epithelial Tgfβr2 feeds back into the Wnt pathway by repressing Dkk1 and Dkk4 to enhance mesenchymal Wnt signaling activity (Iwata et al., 2014). The forkhead box transcription factor Foxf2, which represses Wnt signaling in the gastrointestinal system (Ormestad et al., 2006), may effect Tgfβ signaling during palate development, and has been linked to orofacial clefts in both mice and humans (Bu et al., 2015; Nik et al., 2016) (Table 2, Fig. 3). Foxf2 ablation downregulates Tgfβ2 during palatogenesis, causing a decrease in mesenchymal cell proliferation and aberrant collagen accumulation (Nik et al., 2016), resulting in cleft palate in mice (Wang et al., 2003) (Table 2, Fig. 3). Repression of Wnt signaling by Foxf2 has been demonstrated in intestinal fibroblasts (Nik et al., 2013), although a direct relationship between Foxf2 and Wnt signaling during palatogenesis remains undemonstrated.
Fig. 3.
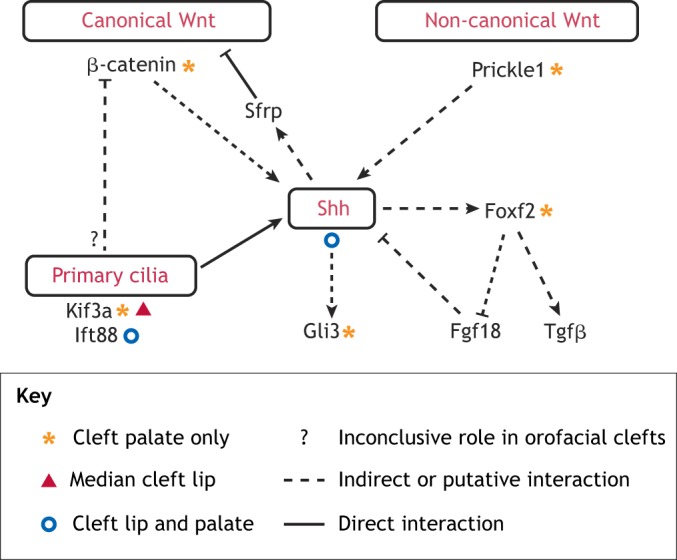
Crosstalk between Wnt, Shh, primary cilia and other related signaling components in orofacial clefts. Shh signaling, which is regulated by primary cilia or intraflagellar transport proteins, represses canonical Wnt signaling through the negative Wnt regulator Sfrp, while β-catenin may regulate Shh signaling, suggesting a negative-feedback loop between the Wnt and Shh pathways in lip/palate formation. The non-canonical Wnt signaling molecule Prickle1 also activates Shh signaling, which may subsequently inhibit the canonical Wnt pathway. There is a complex interplay between the Fox, Fgf, Shh and Tgfβ family members during lip/palate development. The phenotypic outcomes of the key signaling components highlighted in this figure were demonstrated in the respective mutant mouse models.
Wnt-Shh-cilia crosstalk
Hedgehog signaling during embryogenesis depends on primary cilia function and intraflagellar transport (Huangfu et al., 2003; Huangfu and Anderson, 2005) (Fig. 3). Individuals with ciliopathies resulting from defects of the primary cilia often have CLP, and tissue-specific deletion of the intraflagellar transport genes intraflagellar transport 88 (Ift88) or kinesin family member 3A (Kif3a) in mice causes CLP (Liu et al., 2014b; Schock et al., 2017; Tian et al., 2017) (Fig. 3). Mouse pups with conditional deletion of Ift88 in cranial neural crest cells with Wnt1-driven Cre die at birth due to severe craniofacial defects, including bilateral CLP (Box 1), whereas elimination of Ift88 specifically in the palatal mesenchyme results in CPO (Tian et al., 2017). Loss of Ift88 results in a downregulation of Shh signaling in the palatal mesenchyme (Tian et al., 2017). In addition, a novel missense mutation in IFT88 has been reported in a family affected by isolated CLP, suggesting it as a candidate gene for orofacial clefts (Tian et al., 2017). Both Ift88 and Kif3a may repress canonical Wnt signaling (Corbit et al., 2008; Chang and Serra, 2013) (Fig. 3). Combined, these results underscore the significance of intraflagellar proteins in craniofacial development, which involves Shh signaling, and the role of Shh signaling in feeding back to negatively regulate Wnt signaling.
The ventral anterior homeobox (Vax) transcription factors are important for neural patterning, and they mediate signaling between Shh and Wnt (Vacik et al., 2011). Sfrp and ventral anterior homeobox 1 (Vax1) are the downstream effectors of Shh signaling, and Shh, in turn, inhibits Wnt/β-catenin signaling (Kurosaka et al., 2014) (Fig. 3). Furthermore, VAX1 is a candidate human NSCLP gene (Mangold et al., 2010), and Vax1-null mouse embryos exhibit cell proliferation problems during cranial development around E10.5, possibly due to a downregulation of Shh. These embryos do not present with a CLP phenotype, suggesting that Vax1 does not play a direct role in palatogenesis (Geoghegan et al., 2017). Palatal rugae are established by Shh expression, which is opposed by Fgf signaling at the inter-rugal regions of the epithelium (Economou et al., 2012). Wnt/β-catenin signaling is also required for Shh induction in the palatal rugae (Lin et al., 2011; Kawasaki et al., 2018).
Wnt5a/Ror2 may act upstream of the non-canonical Wnt signaling molecule Prickle1 (Liu et al., 2014a; Yang et al., 2014) (Fig. 1), and Prickle1 itself may act upstream of Shh during palatogenesis (Yang et al., 2014) (Table 2, Fig. 3). In osteoblast-lineage cells, the non-canonical and canonical Wnt pathways have a positive relationship, and Wnt5a upregulates Wnt/β-catenin signaling, while its ablation inhibits canonical signaling by reducing Lrp5/Lrp6 expression (Okamoto et al., 2014). Wnt5a may also act upstream of Msx1, Bmp2, Bmp4 and Shh during palatogenesis, placing Wnt5a as a promising candidate for signaling pathway crosstalk during palate development (He et al., 2008; Smith et al., 2012). Although Msx1 expression was downregulated in Wnt5a knockout mouse palates, an Msx1-binding enhancer was identified upstream of Wnt5a, implying a possible synergistic relationship between these two factors (He et al., 2008; Nishihara et al., 2016).
Shh has been reported to activate Fox genes during lip and palate development (Jeong et al., 2004; Nik et al., 2016; Everson et al., 2017). Foxf2 also represses Fgf18 signaling from the palatal mesenchyme, which itself negatively regulates Shh expression in the palatal epithelium, leading to reduced Shh expression in Foxf2 knockout mice (Xu et al., 2016), which suggests a positive-feedback loop (Fig. 3). Gli3 acts as an activator of hedgehog pathway targets in the presence of Shh signaling and becomes a repressor when Shh signaling is absent (Wang et al., 2000). Moreover, GLI3 has been associated with NSCLP in human patients (Wang et al., 2017b), and Gli3-null mouse embryos exhibit cleft palate and tongue abnormalities due to improper tongue morphogenesis and failure of palatal shelf elevation and fusion (Table 2, Fig. 3) (Huang et al., 2008). The repressor form of Gli3 modulates Wnt signaling and physically interacts with β-catenin, linking the Shh and Wnt pathways (Ulloa et al., 2007).
Wnt-RA-Fgf signaling crosstalk
RA plays an important role in normal palatogenesis (Okano et al., 2014), and excess RA exposure in human and murine embryos can cause orofacial clefts (Abbott and Pratt, 1987; Abbott et al., 1989). Several aldehyde dehydrogenases are involved in the synthesis of RA from retinaldehyde, with aldehyde dehydrogenase family 1, subfamily A3 (Aldh1a3) being largely responsible for RA production in the oral epithelium (Kato et al., 2013). RA signaling interacts with the Wnt/β-catenin pathway (Fig. 4) (Kumar and Duester, 2010; Yasuhara et al., 2010; von Gise et al., 2011; Zhao and Duester, 2009; Osei-Sarfo and Gudas, 2014), and alters cellular proliferation and apoptosis in the craniofacial mesenchyme and epithelium through its repression of Wnt signaling in palatogenesis (Hu et al., 2013). Canonical Wnt signaling appears to feed back into and inhibit RA signaling, as Aldh1a3 is ectopically expressed in the upper lip primordia of Lrp6-deficient embryos (Song et al., 2009) (Fig. 4). Cytochrome P450, family 26, subfamily b, polypeptide 1 (Cyp26b1), the enzyme that degrades RA and therefore regulates endogenous RA levels, is required for proper elevation of palatal shelves, and Cyp26b1 knockout mice display cleft palate due to excess RA (Okano et al., 2012). Cyp26b1 enhances T-box 1 (Tbx1) and Fgf10 expression in the oral epithelium, while an excess of RA represses both. Fgf10 expression is lost in Cyp26b1-null mice, and palatal Tbx1 expression was downregulated when murine fetuses were treated with exogenous RA (Okano et al., 2008; Okano et al., 2012) (Fig. 4). Both Tbx1-null and Fgf10-null mice display cleft palate (Alappat et al., 2005; Funato et al., 2012), suggesting that these important regulators of palatal shelf elevation act downstream of Cyp26b1, and their expression is likely modulated by RA levels. As researchers continue to identify the factors that connect these different pathways, it becomes increasingly important to understand how they are regulated.
Fig. 4.
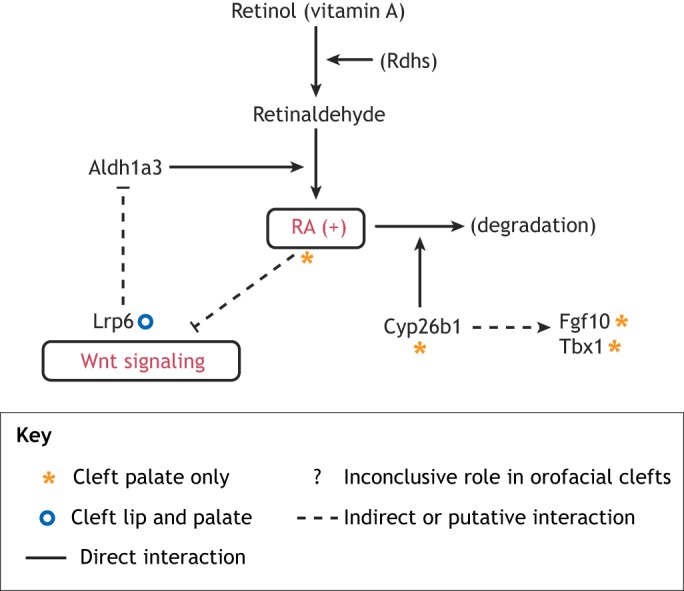
Crosstalk between Wnt, RA and related signaling pathways in orofacial clefts. A surplus (+) of RA induces cleft palate, and RA is known to interact directly with the canonical Wnt pathway. In orofacial primordia, retinol dehydrogenases (Rdhs) convert vitamin A (retinol) to retinaldehyde, which subsequently produces RA mainly via the aldehyde dehydrogenase Aldh1a3. RA represses Wnt signaling, while Lrp6-mediated canonical Wnt signaling represses Aldh1a3 expression in the orofacial epithelium, suggesting a reciprocal negative regulation between the Wnt and RA pathways. The cytochrome P450 family member Cyp26b1 is responsible for RA degradation, thereby regulating the endogenous levels of RA. Fgf10 and Tbx1 may act downstream of Cyp26b1 during palatogenesis. The phenotypic outcomes of the key signaling components highlighted in this figure were demonstrated in the respective mutant mouse models.
Crosstalk of Wnt signaling with epigenetic regulators in orofacial clefts
Sequence-independent gene regulatory mechanisms, such as histone modification, DNA methylation and microRNA (miRNA) transcript regulation, have garnered increasing attention in recent years. These epigenetic mechanisms play a role in regulating many Wnt pathway components (Wils and Bijlsma, 2018). Studies suggest that miRNAs are involved in regulating Wnt signaling during palatogenesis; in mice, conditional deletion of Dicer1, the key effector of RNA interference (RNAi)-mediated mRNA cleavage, leads to craniofacial defects, including cleft palate (Zehir et al., 2010). A 2016 study of plasma miRNAs expressed in human NSCLP patients suggests that many key targets of dysregulated miRNAs share functional relationships with Wnt, Notch, hedgehog and lipid signaling pathways (Li et al., 2016). The miRNAs hsa-miR-24-3p, hsa-miR-1260b and hsa-miR-205-5p have been identified in a human transcriptome screen as candidates for NSCLP, and were computationally predicted to target several Wnt signaling pathway components (Wang et al., 2017a). Another miRNA, miR-544a, has been associated with downregulation of CDH1 during EMT in cancer cells, in turn activating the Wnt signaling pathway (Yanaka et al., 2015). The miR-17-92 cluster reportedly targets transcripts of NSCLP-associated Wnt target genes Tbx1 and Tbx3, and is itself a target of Bmp signaling and of the craniofacial pioneer factor AP-2α. miR-17-92 knockout in mouse embryos results in severe craniofacial defects, including CLP, the severity and penetrance of which are increased in miR-17-92;miR-106b-25 compound mutants (Wang et al., 2013a). In zebrafish, platelet-derived growth factor (Pdgf) signaling is an important regulator of palatogenesis, and it is modulated by miR-140 during palatogenesis (Eberhart et al., 2008). Further studies discerning whether the described roles of miRNAs in other cellular processes resemble their roles in palatogenesis may contribute to our understanding of the mechanisms by which cleft palate arises.
Several studies examined epigenetic modifications in NSCLP, including DNA methylation (Alvizi et al., 2017; Sharp et al., 2017). The transcription factor and human CLP candidate SOX4, the targets of which include FZD5 (Scharer et al., 2009), has been implicated in studies of genomic regions that are differentially methylated during palatogenesis (Seelan et al., 2012). Changes in CDH1 promoter methylation levels in human blood and lip tissue have been correlated with NSCLP, as well as with differences in NSCLP penetrance in susceptible families, implying that DNA methylation patterns may account for the variable penetrance of CLP phenotypes (Alvizi et al., 2017). A stochastic deficiency in DNA methylation of a retrotransposon near the coding region of Wnt9b in the A/WySn mouse impaired its transcription and contributed to an incompletely penetrant CLP phenotype (Juriloff et al., 2014). The role of histone modifications in craniofacial development is less understood, but recent studies suggest that histone H3 acetylation can play a role in the formation of cleft palate in mouse due to dysregulation of Tgfβ signaling, although how this process affects Wnt signaling has not been demonstrated (Yuan et al., 2016). However, the histone acetyltransferase p300 (also known as Ep300) is important for gene regulation, and its ablation in mouse palatal mesenchyme cells results in altered Wnt signaling, as well as in aberrant Wnt-dependent proliferation and migration (Warner et al., 2016). An improved understanding of epigenetic regulation of Wnt signaling and related pathways may hold the key to addressing the impact of environmental and non-genetic factors on the presentation of orofacial clefts.
Translational perspectives
To date, the most prevalent treatment for orofacial clefts is surgical repair coupled with nasoalveolar molding to direct postnatal tissue growth and subsequent orthodontic treatment (Chen et al., 2005). Protocols and procedures have varied widely, not only in developing areas of the world, but within developed countries as well (Mossey et al., 2009). A better understanding of the complex interactions between components of the Wnt and other signaling pathways that govern lip/palate formation will provide better opportunities for treatment and prevention of orofacial clefts through cellular- and molecular-based methods, reducing the need for surgical intervention (Panetta et al., 2008). Research in animal models has identified several altered pathways that, when targeted, reversed orofacial clefts. Direct modulation of Wnt signaling by chemically stabilizing a catalytically inactive allele of the canonical Wnt pathway factor Gsk3 has shown therapeutic potential in mice, where its timely reactivation could reverse a cleft palate phenotype in Gsk3β-deficient mice (Liu et al., 2007). Additionally, ectopic expression of Wnt in the ectoderm rescued the orofacial cleft phenotype in Pbx-deficient mouse models (Feretti et al., 2011). Modulation of Shh signaling has also been shown to rescue cleft palate in the Msx1-null mouse model, both through ectopic Bmp expression and through downregulation of distal-less homeobox 5 (Dlx5) (Zhang et al., 2002; Han et al., 2009). Reynolds and colleagues have shown that administration of either 3-4 mg/kg folic acid or 140-187 mg/kg methionine to pregnant mice that were previously treated with intraperitoneal RA to induce CLP reduces the frequency of cleft palate to 6%, compared with 76% in RA-treated controls. Interestingly, the combined folic acid and methionine treatment completely rescued the RA-induced aberrant palatogenesis (Reynolds et al., 2003).
Utilizing controlled intravenous delivery of the small-molecule Wnt agonists WAY-262611 and IIIc3a (both acting as Dkk inhibitors) into Pax9 mutant mice rescues the growth and fusion of palatal shelves by restoring Wnt signaling (Jia et al., 2017a; Li et al., 2017). In addition to small-molecule modulation, synthetic ligand analogs have also shown potential to stimulate Wnt signaling (Andersson et al., 2015; Zhan et al., 2017) and could lead to the development of future treatments for orofacial clefts. Additionally, genetic inactivation of Wise (also known as Sostdc1), a canonical Wnt antagonist, in Pax9-deficient mouse embryos rescued the palatal shelf elevation, mainly through restoring hyaluronic acid accumulation in the palatal mesenchyme (Li et al., 2017). Wnt5a analogs, such as Foxy-5 or Box-5, currently used in cancer research (Andersson et al., 2015; Zhan et al., 2017), may also serve as a treatment approach for orofacial clefts by targeting non-canonical Wnt signaling, warranting future research. Taken together, these reports indicate that Wnt signaling modulators could contribute to an effective molecular treatment regime for orofacial clefts.
Given the heterogeneous causes of orofacial clefts and the variability in the genotypes of affected individuals, it is unlikely that we will see a ‘one-size’ approach to non-surgical orofacial cleft treatment any time soon. However, successful studies using mutant mouse models are promising for the prospect of pathway-specific treatments to allow for prenatal intervention when a fetal genotype renders human embryos at risk of orofacial cleft development. Implementation of promising therapies to human patients would impinge on the timing of application and on the accurate detection of improper palatogenesis. Even if parents carrying alleles linked to orofacial clefts were to commit to prenatal genotyping, many palatal development processes occur early in gestation. Even with improved protocols for correcting the levels of a target signaling factor in patients, implementing postconception measures at such an early stage of pregnancy, before many mothers know that they are pregnant, is challenging. Further challenges arise from the high variability in the manifestation and penetrance of orofacial cleft phenotypes. The complexity of the molecular processes that govern orofacial development makes it difficult to predict the therapeutic requirement. Moreover, a particular intervention may not be applicable to more than a minor subset of cases, even in individuals with defects in a particular gene or pathway. A deeper understanding of the pathways that govern palatogenesis may allow in vitro fertilization with selected gametes that do not possess the risk-imparting allele.
Interactions between Wnt/β-catenin signaling and other morphogenetic signaling pathways are widely employed in many different developmental programs. The nature of embryonic development creates significant potential for off-target effects and disruption of other essential developmental processes in both mother and child if molecular treatments to correct palatogenesis errors are applied systemically. Before treatments targeting signaling pathways could be considered for clinical trials with human patients, such risks would need to be thoroughly explored and addressed, and likely require new delivery techniques more advanced than those currently available. Additionally, because NSCLP is not life threatening, many parents may be unwilling to attempt untested and potentially dangerous approaches, despite the burden and difficulty of current treatments. However, in the future, a more complete understanding of morphogenetic pathway crosstalk and the systemic impact of perturbations to them may eventually allow the progression of molecular clinical approaches to a point at which they are considered safe. A robust understanding of how signaling pathways function in all systems and processes during development will be able to not only inform studies related to orofacial clefts, but also contribute to the development of treatments for other syndromes and disorders with Wnt pathway etiologies. Despite the many barriers that still remain, knowledge of developmental mechanisms is helping to, and will continue to, facilitate the refinement of techniques for the application of that knowledge to develop the means to safely and effectively treat congenital disorders like orofacial clefts.
Acknowledgements
We appreciate reviewers' insightful comments and Dr Julija Hmeljak's constructive editorial work during manuscript preparation and revision. We apologize to colleagues whose important work we were unable to cite due to space constraints or inadvertently overlooking.
Footnotes
Competing interests
The authors declare no conflict of interest. The funding sponsors had no role in the preparation of the manuscript and in the decision to publish this review.
Author contributions
K.R., P.K., L.S.R., and R.G. collected and analyzed the references and wrote the manuscript; Y.J. and S.K. assisted with manuscript preparation; C.J.Z. conceptualized, advised, edited and approved the manuscript.
Funding
This work is supported by research funds from the National Institutes of Health (R01DE026737, R01DE021696 and R01NS102261 to C.J.Z.) and the Shriners Hospitals for Children (85105 to C.J.Z.; postdoctoral fellowship 84551 to R.G.).
References
- Abbott B. D. and Pratt R. M. (1987). Retinoids and epidermal growth factor alter embryonic mouse palatal epithelial and mesenchymal cell differentiation in organ culture. J. Craniofac. Genet. Dev. Biol. 7, 219-240. [PubMed] [Google Scholar]
- Abbott B. D., Harris M. W. and Birnbaum L. S. (1989). Etiology of retinoic acid-induced cleft palate varies with the embryonic stage. Teratology 40, 533-553. 10.1002/tera.1420400602 [DOI] [PubMed] [Google Scholar]
- Abramyan J. and Richman J. M. (2018). Craniofacial development: discoveries made in the chicken embryo. Int. J. Dev. Biol. 62, 97-107. 10.1387/ijdb.170321ja [DOI] [PubMed] [Google Scholar]
- Afzal A. R., Rajab A., Fenske C. D., Oldridge M., Elanko N., Ternes-Pereira E., Tüysüz B., Murday V. A., Patton M. A., Wilkie A. O. M. et al. (2000). Recessive Robinow syndrome, allelic to dominant brachydactyly type B, is caused by mutation of ROR2. Nat. Genet. 25, 419 10.1038/78107 [DOI] [PubMed] [Google Scholar]
- Alappat S. R., Zhang Z., Suzuki K., Zhang X., Liu H., Jiang R., Yamada G. and Chen Y. P. (2005). The cellular and molecular etiology of the cleft secondary palate in Fgf10 mutant mice. Dev. Biol. 277, 102-113. 10.1016/j.ydbio.2004.09.010 [DOI] [PubMed] [Google Scholar]
- Alexander C., Piloto S., Le Pabic P. and Schilling T. F. (2014). Wnt signalling interacts with bmp and edn1 to regulate dorsal-ventral patterning and growth of the craniofacial skeleton. PLoS Genet. 10, e1004479 10.1371/journal.pgen.1004479 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Alkuraya F. S., Saadi I., Lund J. J., Turbe-Doan A., Morton C. C. and Maas R. L. (2006). SUMO1 haploinsufficiency leads to cleft lip and palate. Science 313, 1751 10.1126/science.1128406 [DOI] [PubMed] [Google Scholar]
- Alvizi L., Ke X., Brito L. A., Seselgyte R., Moore G. E., Stanier P. and Passos-Bueno M. R. (2017). Differential methylation is associated with non-syndromic cleft lip and palate and contributes to penetrance effects. Sci. Rep. 7, 2441 10.1038/s41598-017-02721-0 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Andersson T., Axelsson L., Mohapatra P., Prasad C., Soerensen P. G., Mau-Soerensen M., Lassen U., Molvadgaard T., Buhl U., Brünner N. et al. (2015). Abstract A116: Targeting the Wnt-5a signaling pathway as a novel anti-metastatic therapy. Mol. Cancer Ther. 14 12 Suppl. 2, A116 10.1158/1535-7163.TARG-15-A116 [DOI] [Google Scholar]
- Baek J. A., Lan Y., Liu H., Maltby K. M., Mishina Y. and Jiang R. (2011). Bmpr1a signaling plays critical roles in palatal shelf growth and palatal bone formation. Dev. Biol. 350, 520-531. 10.1016/j.ydbio.2010.12.028 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Bankhead E. J., Colasanto M. P., Dyorich K. M., Jamrich M., Murtaugh L. C. and Fuhrmann S. (2015). Multiple requirements of the focal dermal hypoplasia gene porcupine during ocular morphogenesis. Am. J. Pathol. 185, 197-213. 10.1016/j.ajpath.2014.09.002 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Barrott J. J., Cash G. M., Smith A. P., Barrow J. R. and Murtaugh L. C. (2011). Deletion of mouse Porcn blocks Wnt ligand secretion and reveals an ectodermal etiology of human focal dermal hypoplasia/Goltz syndrome. Proc. Natl. Acad. Sci. USA 108, 12752-12757. 10.1073/pnas.1006437108 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Basha M., Demeer B., Revencu N., Helaers R., Theys S., Bou Saba S., Boute O., Vevauchelle B., Francois G., Bayet B. et al. (2018). Whole exome sequencing identifies mutations in 10% of patients with familial non-syndromic cleft lip and/or palate in genes mutated in well-known syndromes. J. Med. Genet. 55, 449-458. 10.1136/jmedgenet-2017-105110 [DOI] [PubMed] [Google Scholar]
- Basson M. A. (2012). Signaling in cell differentiation and morphogenesis. Cold Spring Harb. Perspect. Biol. 4, a008151 10.1101/cshperspect.a008151 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Basu M. and Roy S. S. (2013). Wnt/beta-catenin pathway is regulated by PITX2 homeodomain protein and thus contributes to the proliferation of human ovarian adenocarcinoma cell, SKOV-3. J. Biol. Chem. 288, 4355-4367. 10.1074/jbc.M112.409102 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Beaty T. H., Hetmanski J. B., Fallin M. D., Park J. W., Sull J. W., McIntosh I., Liang K. Y., Vanderkolk C. A., Redett R. J., Boyadjiev S. A. et al. (2006). Analysis of candidate genes on chromosome 2 in oral cleft case-parent trios from three populations. Hum. Genet. 120, 501-518. 10.1007/s00439-006-0235-9 [DOI] [PubMed] [Google Scholar]
- Bernatik O., Ganji R. S., Dijksterhuis J. P., Konik P., Cervenka I., Polonio T., Krejci P., Schulte G. and Bryja V. (2011). Sequential activation and inactivation of dishevelled in the Wnt/β-catenin pathway by casein kinases. J. Biol. Chem. 286, 10396-10410. 10.1074/jbc.M110.169870 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Borycki A., Brown A. M. and Emerson C. P. Jr. (2000). Shh and Wnt signaling pathways converge to control Gli gene activation in avian somites. Development 127, 2075-2087. [DOI] [PubMed] [Google Scholar]
- Brito L. A., Yamamoto G. L., Melo S., Malcher C., Ferreira S. G., Figueiredo J., Alvizi L., Kobayashi G. S., Naslavsky M. S., Alonso N. et al. (2015). Rare variants in the epithelial cadherin gene underlying the genetic etiology of nonsyndromic cleft lip with or without cleft palate. Hum. Mutat. 36, 1029-1033. 10.1002/humu.22827 [DOI] [PubMed] [Google Scholar]
- Bu L., Chen Q., Wang H., Zhang T., Hetmanski J. B., Schwender H., Parker M., Chou Y. H., Yeow V., Chong S. S. et al. (2015). Novel evidence of association with nonsyndromic cleft lip with or without cleft palate was shown for single nucleotide polymorphisms in FOXF2 gene in an Asian population. Birth Defects Res. A Clin. Mol. Teratol 103, 857-862. 10.1002/bdra.23413 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Bunn K. J., Daniel P., Rösken H. S., O'Neill A. C., Cameron-Christie S. R., Morgan T., Brunner H. G., Lai A., Kunst H. P., Markei D. M. et al. (2015). Mutations in DVL1 cause an osteosclerotic form of Robinow syndrome. Am. J. Hum. Genet. 96, 623-630. 10.1016/j.ajhg.2015.02.010 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Bureau A., Parker M. M., Ruczinski I., Taub M. A., Marazita M. L., Murray J. C., Mangold E., Noethen M. M., Ludwig K. U., Hetmanski J. B. et al. (2014). Whole exome sequencing of distant relatives in multiplex families implicates rare variants in candidate genes for oral clefts. Genetics 197, 1039-1044. 10.1534/genetics.114.165225 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Bush J. O. and Jiang R. (2012). Palatogenesis: morphogenetic and molecular mechanisms of secondary palate development. Development 139, 231-243. 10.1242/dev.067082 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Buttler K., Becker J., Pukrop T. and Wilting J. (2013). Maldevelopment of dermal lymphatics in Wnt5a-knockout-mice. Dev. Biol. 381, 365-376. 10.1016/j.ydbio.2013.06.028 [DOI] [PubMed] [Google Scholar]
- Byrd N. A. and Meyers E. N. (2005). Loss of Gbx2 results in neural crest cell patterning and pharyngeal arch artery defects in the mouse embryo. Dev. Biol. 284, 233-245. 10.1016/j.ydbio.2005.05.023 [DOI] [PubMed] [Google Scholar]
- Carmichael S. L., Nelson V., Shaw G. M., Wasserman C. R. and Croen L. A. (2003). Socio-economic status and risk of conotruncal heart defects and orofacial clefts. Paediatr. Perinat. Epidemiol. 17, 264-271. 10.1046/j.1365-3016.2003.00498.x [DOI] [PubMed] [Google Scholar]
- Carpinelli M. R., De Vries M. E., Jane S. M. and Dworkin S. (2017). Grainyhead-like transcription factors in craniofacial development. J. Dent. Res. 96, 1200-1209. 10.1177/0022034517719264 [DOI] [PubMed] [Google Scholar]
- Carroll T. J., Park J. S., Hayashi S., Majumdar A. and McMahon A. P. (2005). Wnt9b plays a central role in the regulation of mesenchymal to epithelial transitions underlying organogenesis of the mammalian urogenital system. Dev. Cell 9, 283-292. 10.1016/j.devcel.2005.05.016 [DOI] [PubMed] [Google Scholar]
- Castro Colabianchi A. M., Revinski D. R., Encinas P. I., Baez M. V., Monti R. J., Rodríguez Abinal M., Kodjabachian L., Franchini L. F. and López S.L. (2018). Notch1 is asymmetrically distributed from the beginning of embryogenesis and controls the ventral center. Development 145, dev159368 10.1242/dev.159368 [DOI] [PubMed] [Google Scholar]
- Cervantes S., Yamaguchi T. P. and Hebrok M. (2009). Wnt5a is essential for intestinal elongation in mice. Dev. Biol. 326, 285-294. 10.1016/j.ydbio.2008.11.020 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Chang C. F. and Serra R. (2013). Ift88 regulates Hedgehog signaling, Sfrp5 expression, and ß-catenin activity in post-natal growth plate. J. Orthop. Res. 31, 350-356. 10.1002/jor.22237 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Chen P. K.-T., Noordhoff M. S. and Liou E. J. W. (2005). Treatment of complete bilateral cleft lip-nasal deformity. Semin. Plast. Surg. 19, 329-342. 10.1055/s-2005-925905 [DOI] [Google Scholar]
- Chen Q., Wang H., Hetmanski J. B., Zhang T., Ruczinski I., Schwender H., Liang K. Y., Fallin M. D., Redett R. J., Raymond G. V. et al. (2012). BMP4 was associated with NSCL/P in an Asian population. PLoS One 7, e35347 10.1371/journal.pone.0035347 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Cheng S.-L., Shao J.-S., Charlton-Kachigian N., Loewy A. P. and Towler D. A. (2003). MSX2 promotes osteogenesis and suppresses adipogenic differentiation of multipotent mesenchymal progenitors. J. Biol. Chem. 278, 45969-45977. 10.1074/jbc.M306972200 [DOI] [PubMed] [Google Scholar]
- Chia I. V., Kim M. J., Itoh K., SokolS Y. and Costantini F. (2009). Both the RGS domain and the six C-terminal amino acids of mouse Axin are required for normal embryogenesis. Genetics 181, 1359-1368. 10.1534/genetics.109.101055 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Chiquet B. T., Blanton S. H., Burt A., Ma D., Stal S., Mulliken J. B. and Hecht J. T. (2008). Variation in WNT genes is associated with non-syndromic cleft lip with or without cleft palate. Hum. Mol. Genet. 17, 2212-2218. 10.1093/hmg/ddn121 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Clevers H. and Nusse R. (2012). Wnt/beta-catenin signaling and disease. Cell 149, 1192-1205. 10.1016/j.cell.2012.05.012 [DOI] [PubMed] [Google Scholar]
- Clevers H., Loh K. M. and Nusse R. (2014). Stem cell signaling. An integral program for tissue renewal and regeneration: Wnt signaling and stem cell control. Science 346, 1248012 10.1126/science.1248012 [DOI] [PubMed] [Google Scholar]
- Cobourne M. T. and Green J. B. (2012). Hedgehog signaling in development of the secondary palate. Front. Oral Biol. 16, 52-59. 10.1159/000337543 [DOI] [PubMed] [Google Scholar]
- Cong W., Liu B., Liu S., Sun M., Liu H., Yang Y., Wang R. and Xiao J. (2014). Implications of the Wnt5a/CaMKII pathway in retinoic acid-induced myogenic tongue abnormalities of developing mice. Sci. Rep. 4, 6082 10.1038/srep06082 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Cooper S. C., Flaitz C. M., Johnston D. A., Lee B. and Hecht J. T. (2001). A natural history of cleidocranial dysplasia. Am. J. Med. Genet. 104, 1-6. 10.1002/ajmg.10024 [DOI] [PubMed] [Google Scholar]
- Cordero D. R., Brugmann S., Chu Y., Bajpai R., Jame M. and Helms J. A. (2011). Cranial neural crest cells on the move: their roles in craniofacial development. Am. J. Med. Genet. Part A 155A, 270-279. 10.1002/ajmg.a.33702 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Corbit K. C., Shyer A. E., Dowdle W. E., Gaulden J., Singla V., Chen M. H., Chuang P. T. and Reiter J. F. (2008). Kif3a constrains beta-catenin-dependent Wnt signalling through dual ciliary and non-ciliary mechanisms. Nat. Cell Biol. 10, 70-76. 10.1038/ncb1670 [DOI] [PubMed] [Google Scholar]
- Croen L. A., Shaw G. M., Wasserman C. R. and Tolarova M. M. (1998). Racial and ethnic variations in the prevalence of orofacial clefts in California, 1983–1992. Am. J. Med. Genet. 79, 42-47. [DOI] [PubMed] [Google Scholar]
- Curtin E., Hickey G., Kamel G., Davidson A. J. and Liao E. C. (2011). Zebrafish wnt9a is expressed in pharyngeal ectoderm and is required for palate and lower jaw development. Mech. Dev. 128, 104-115. 10.1016/j.mod.2010.11.003 [DOI] [PubMed] [Google Scholar]
- Cvjetkovic N., Maili L., Weymouth K. S., Hashmi S. S., Mulliken J. B., Topczewski J., Letra A., Yuan Q. P., Blanton S. H., Swindell E. C. et al. (2015). Regulatory variant in FZD6 gene contributes to nonsyndromic cleft lip and palate in an African-American family. Mol. Genet. Genomic. Med. 3, 440-451. 10.1002/mgg3.155 [DOI] [PMC free article] [PubMed] [Google Scholar]
- de la Garza G., Schleiffarth J. R., Dunnwald M., Mankad A., Weirather J. L., Bonde G., Butcher S., Mansour T. A., Kousa Y. A., Fukazawa C. F. et al. (2013). Interferon regulatory factor 6 promotes differentiation of the periderm by activating expression of grainyhead-like 3. J. Invest. Dermatol. 133, 68-77. 10.1038/jid.2012.269 [DOI] [PMC free article] [PubMed] [Google Scholar]
- de Lima R. L., Hoper S. A., Ghassibe M., Cooper M. E., Rorick N. K., Kondo S., Katz L., Marazita M. L., Compton J., Bale S. et al. (2009). Prevalence and nonrandom distribution of exonic mutations in interferon regulatory factor 6 in 307 families with Van der Woude syndrome and 37 families with popliteal pterygium syndrome. Genet. Med. 11, 241-247. 10.1097/GIM.0b013e318197a49a [DOI] [PMC free article] [PubMed] [Google Scholar]
- Dickinson A. J. G. (2016). Using frogs faces to dissect the mechanisms underlying human orofacial defects. Semin. Cell Dev. Biol. 51, 54-63. 10.1016/j.semcdb.2016.01.016 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Dickinson A. J. G. and Sive H. L. (2009). The Wnt antagonists Frzb-1 and Crescent locally regulate basement membrane dissolution in the developing primary mouth. Development 136, 1071-1081. 10.1242/dev.032912 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Dixon M. J., Marazita M. L., Beaty T. H. and Murray J. C. (2011). Cleft lip and palate: understanding genetic and environmental influences. Nat. Rev. Genet. 12, 167-178. 10.1038/nrg2933 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Dodé C., Levilliers J., Dupont J. M., De Paepe A., Le Dû N., Soussi-Yanicostas N., Coimbra R. S., Delmaghani S., Compain-Nouaille S. and Baverel F. (2003). Loss-of-function mutations in FGFR1 cause autosomal dominant Kallmann syndrome. Nat. Genet. 33, 463-465. 10.1038/ng1122 [DOI] [PubMed] [Google Scholar]
- Donaldson I. J., Amin S., Hensman J. J., Kutejova E., Rattray M., Lawrence N., Hayes A., Ward C. M. and Bobola N. (2012). Genome-wide occupancy links Hoxa2 to Wnt-β-catenin signaling in mouse embryonic development. Nucleic Acids Res. 40, 3990-4001. 10.1093/nar/gkr1240 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Dougherty M., Kamel G., Grimaldi M., Gfrerer L., Shubinets V., Ethier R., Hickey G., Cornell R. A. and Liao E. C. (2013). Distinct requirements for wnt9a and irf6 in extension and integration mechanisms during zebrafish palate morphogenesis. Development 140, 76-81. 10.1242/dev.080473 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Dudas M., Sridurongrit S., Nagy A., Okazaki K. and Kaartinen V. (2004). Craniofacial defects in mice lacking BMP typr I receptor for Alk2 in neural crest cells. Mech. Dev. 121, 173-182. 10.1016/j.mod.2003.12.003 [DOI] [PubMed] [Google Scholar]
- Dudas M., Kim J., Li W. Y., Nagy A., Larsson J., Karlsson S., Chai Y. and Kaartinen V. (2006). Epithelial and ectomesenchymal role of the type I TGF-beta receptor ALK5 during facial morphogenesis and palatal fusion. Dev. Biol. 296, 298-314. 10.1016/j.ydbio.2006.05.030 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Duncan K. M., Mukherjee K., Cornell R. A. and Liao E. C. (2017). Zebrafish models of orofacial clefts. Dev. Dyn.. 246, 897-914. 10.1002/dvdy.24566 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Dupé V., Matt N., Garnier J. M., Chambon P., Mark M. and Ghyselinck N. B. (2003). A newborn lethal defect due to inactivation of retinaldehyde dehydrogenase type 3 is prevented by maternal retinoic acid treatment. Proc. Natl. Acad. Sci. USA 100, 14036-14041. 10.1073/pnas.2336223100 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Eberhart J. K., He X., Swartz M. E., Yan Y. L., Song H., Boling T. C., Kunerth A. K., Walker M. B., Kimmel C. B. and Postlethwait J. H. (2008). MicroRNA Mirn140 modulates Pdgf signaling during palatogenesis. Nat. Genet. 40, 290-298. 10.1038/ng.82 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Economou A. D., Ohazama A., Porntaveetus T., Sharpe P. T, Kondo S., Basson M. A., Gritli-Linde A., Cobourne M. T. and Green J. B. A. (2012). Periodic stripe formation by a Turing mechanism operating at growth zones in the mammalian palate. Nat. Genet. 44, 348-351. 10.1038/ng.1090 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Eshete M. A., Liu H., Li M., Adeyemo W. L., Gowans L. J. J., Mossey P. A., Busch T., Deressa W., Donkor P., Olaitan P. B. et al. (2018). Loss-of-function GRHL3 variants detected in african patients with isolated cleft palate. J. Dent. Res. 97, 41-48. 10.1177/0022034517729819 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Everson J. L., Fink D. M., Yoon J. W., Leslie E. J., Kietzman H. W., Ansen-Wilson L. J., Chung H. M., Walterhouse D. O., Marazita M. L. and Lipinski R. J. (2017). Sonic hedgehog regulation of Foxf2 promotes cranial neural crest mesenchyme proliferation and is disrupted in cleft lip morphogenesis. Development 144, 2082 10.1242/dev.149930 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Fakhouri W. D., Metwalli K., Naji A., Bakhiet S., Quispe-Salcedo A., Nitschke L., Kousa Y. A. and Schutte B. C. (2017). Intercellular genetic interaction between Irf6 and Twist1 during craniofacial development. Sci. Rep. 7, 7129 10.1038/s41598-017-06310-z [DOI] [PMC free article] [PubMed] [Google Scholar]
- Feng C., Xu Z., Li Z., Zhang D., Liu Q. and Lu L. (2013). Down-regulation of Wnt10a by RNA interference inhibits proliferation and promotes apoptosis in mouse embryonic palatal mesenchymal cells through Wnt/ß-catenin signaling pathway. J. Physiol. Biochem. 69, 855-863. 10.1007/s13105-013-0262-7 [DOI] [PubMed] [Google Scholar]
- Feng C., Duan W., Zhang D., Zhang E., Xu Z. and Lu L. (2014). C392T polymorphism of the Wnt10a gene in non-syndromic oral cleft in a northeastern Chinese population. Br. J. Oral Maxillofac. Surg. 52, 751-755. 10.1016/j.bjoms.2014.06.001 [DOI] [PubMed] [Google Scholar]
- Ferretti E., Li B., Zewdu R., Wells V., Hebert J. M., Karner C., Anderson M. J., Williams T., Dixon J., Dixon M. J. et al. (2011). A conserved Pbx-Wnt-p63-Irf6 regulatory module controls face morphogenesis by promoting epithelial apoptosis. Dev. Cell 21, 627-641. 10.1016/j.devcel.2011.08.005 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Fontoura C., Silva R. M., Granjeiro J. M. and Letra A. (2015). Association of WNT9B gene polymorphisms with nonsyndromic cleft lip with or without cleft palate in Brazilian nuclear families. Cleft Palate Craniofac. J. 52, 44-48. 10.1597/13-146 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Francis-West P., Ladher R., Barlow A. and Graveson A. (1998). Signalling interactions during facial development. Mech. Dev. 75, 3-28. 10.1016/S0925-4773(98)00082-3 [DOI] [PubMed] [Google Scholar]
- Fu J., Yu H.-M. I., Maruyama T., Mirando A. J. and Hsu W. (2011). Gpr177/mouse Wntless is essential for Wnt-mediated craniofacial and brain development. Dev. Dyn. 240, 365-371. 10.1002/dvdy.22541 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Fu X., Xu J., Chaturvedi P., Liu H., Jiang R. and Lan Y. (2017). Identification of Osr2 transcriptional target genes in palate development. J. Dent. Res. 96, 1451-1458. 10.1177/0022034517719749 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Fuchs A., Inthal A., Herrmann D., Cheng S., Nakatomi M., Peters H. and Neubüser A. (2010). Regulation of Tbx22 during facial and palatal development. Dev. Dyn. 239, 2860-2874. 10.1002/dvdy.22421 [DOI] [PubMed] [Google Scholar]
- Funato N., Nakamura M., Richardson J. A., Srivastava D. and Yanagisawa H. (2012). Tbx1 regulates oral epithelial adhesion and palatal development. Hum. Mol. Genet. 21, 2524-2537. 10.1093/hmg/dds071 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Funato N., Nakamura M. and Yanagisawa H. (2015). Molecular basis of cleft palates in mice. World J. Biol. Chem. 6, 121-138. 10.4331/wjbc.v6.i3.121 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Garcia-Higuera I., Manchado E., Dubus P., Canamero M., Mendez J., Moreno S. and Malumbres M. (2008). Genomic stability and tumour suppression by the APC/C cofactor Cdh1. Nat. Cell Biol. 10, 802-811. 10.1038/ncb1742 [DOI] [PubMed] [Google Scholar]
- Gao C. and Chen Y.-G. (2010). Dishevelled: the hub of Wnt signaling. Cell. Signal. 22, 717-727. 10.1016/j.cellsig.2009.11.021 [DOI] [PubMed] [Google Scholar]
- Gaur T., Lengner C. J., Hovhannisyan H., Bhat R. A., Bodine P. V., Komm B. S., Javed A., van Wijnen A. J., Stein J. L., Stein G. S. et al. (2005). Canonical WNT signaling promotes osteogenesis by directly stimulating Runx2 gene expression. J. Biol. Chem. 280, 33132-33140. 10.1074/jbc.M500608200 [DOI] [PubMed] [Google Scholar]
- Geetha-Loganathan P., Nimmagadda S., Antoni L., Fu K., Whiting C. J., Francis-West P. and Richman J. M. (2009). Expression of WNT signalling pathway genes during chicken craniofacial development. Dev. Dyn. 238, 1150-1165. 10.1002/dvdy.21934 [DOI] [PubMed] [Google Scholar]
- Geetha-Loganathan P., Nimmagadda S., Fu K. and Richman J. M. (2014). Avian facial morphogenesis is regulated by c-Jun N-terminal kinase/planar cell polarity)JNK/PCP) wingless-related (WNT) signaling. J. Biol. Chem. 289, 24153-24167. 10.1074/jbc.M113.522003 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Geoghegan F., Xavier G. M., Birjandi A. A., Seppala M. and Cobourne M. T. (2017). Vax1 plays an indirect role in the etiology of murine cleft palate. J. Dent. Res. 96, 1555-1562. 10.1177/0022034517724145 [DOI] [PubMed] [Google Scholar]
- Goldie S. J., Arhatari B. D., Anderson P., Auden A., Partridge D. D., Jane S. M. and Dworkin S. (2016). Mice lacking the conserved transcription factor Grainyhead-like 3 (Grhl3) display increased apposition of the frontal and parietal bones during embryonic development. BMC Dev. Biol. 16, 37 10.1186/s12861-016-0136-7 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Graf D., Malik Z., Hayano S. and Mishina Y. (2016). Common mechanisms in development and disease: BMP signaling in craniofacial development. Cytokine Growth Factor Rev. 27, 129-139. 10.1016/j.cytogfr.2015.11.004 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Graham A., Francis-West P., Brickell P. and Lumsden A. (1994). The signalling molecule BMP4 mediates apoptosis in the rhombencephalic neural crest. Nature 372, 684 10.1038/372684a0 [DOI] [PubMed] [Google Scholar]
- Green R. M., Feng W., Phang T., Fish J. L., Li H., Spritz R. A., Marcucio R. S., Hooper J., Jamniczky H., Hallgrímsson B. et al. (2015). Tfap2a-dependent changes in mouse facial morphology result in clefting that can be ameliorated by a reduction in Fgf8 gene dosage. Dis. Model Mech. 8, 31-43. 10.1242/dmm.017616 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Gritli-Linde A. (2007). Molecular control of secondary palate development. Dev. Biol. 301, 309-326. 10.1016/j.ydbio.2006.07.042 [DOI] [PubMed] [Google Scholar]
- Gritli-Linde A. (2008). The etiopathogenesis of cleft lip and cleft palate: usefulness and caveats of mouse models. Curr. Top. Dev. Biol. 84, 37-138. 10.1016/S0070-2153(08)00602-9 [DOI] [PubMed] [Google Scholar]
- Halford M. M., Armes J., Buchert M., Meskenaite V., Grail D., Hibbs M. L., Wilks A. F., Farlie P. G., Newgreen D. F., Hovens C. M. et al. (2000). Ryk-deficient mice exhibit craniofacial defects associated with perturbed Eph receptor crosstalk. Nat. Genet. 25, 414 10.1038/78099 [DOI] [PubMed] [Google Scholar]
- Hallonet M., Hollemann T., Pieler T. and Gruss P. (1999). Vax1, a novel homeobox-containing gene, directs development of the basal forebrain and visual system. Genes Dev. 13, 3106-3114. 10.1101/gad.13.23.3106 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Han J., Mayo J., Xu X., Li J., Bringas P. Jr., Maas, R. L., Rubenstein J. L. and Chai Y. (2009). Indirect modulation of Shh signaling by Dlx5 affects the oral-nasal patterning of palate and rescues cleft palate in Msx1-null mice. Development 136, 4225-4233. 10.1242/dev.036723 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Han Y., Zhou L., Ma L., Li D., Xu M., Yuan H., Ma J., Zhang W., Jiang H., Wu Y. et al. (2014). The axis inhibition protein 2 polymorphisms and non-syndromic orofacial clefts susceptibility in a Chinese Han population. J. Oral Pathol. Med. 43, 554-560. 10.1111/jop.12162 [DOI] [PubMed] [Google Scholar]
- Hayashi K. I., Nakamura S., Nishida W. and Sobue K. (2006). Bone morphogenetic protein-induced Msx1 and Msx2 inhibit myocardin-dependent smooth muscle gene transcription. Mol. Cell. Biol. 26, 9456-9470. 10.1128/MCB.00759-06 [DOI] [PMC free article] [PubMed] [Google Scholar]
- He M. and Bian Z. (2016). Lack of association between missense variants in GRHL3 (rs2486668 and rs545809) and susceptibility to non-syndromic orofacial clefts in a Han Chinese Population. PLoS ONE 11, e0159940 10.1371/journal.pone.0159940 [DOI] [PMC free article] [PubMed] [Google Scholar]
- He F. and Chen Y. (2012). Wnt signaling in lip and palate development. Front Oral Biol. 16, 81-90. 10.1159/000337619 [DOI] [PubMed] [Google Scholar]
- He F., Xiong W., Yu X., Espinoza-Lewis R., Liu C., Gu S., Nishita M., Suzuki K., Yamada G., Minami Y. et al. (2008). Wnt5a regulates directional cell migration and cell proliferation via Ror2-mediated non-canonical pathway in mammalian palate development. Development 135, 3871-3879. 10.1242/dev.025767 [DOI] [PMC free article] [PubMed] [Google Scholar]
- He F., Xiong W., Wang Y., Matsui M., Yu X., Chai Y., Klingensmith J. and Chen Y. (2010). Modulation of BMP signaling by Noggin is required for the maintenance of palatal epithelial integrity during palatogenesis. Dev. Biol. 347, 109-121. 10.1016/j.ydbio.2010.08.014 [DOI] [PMC free article] [PubMed] [Google Scholar]
- He F., Xiong W., Wang Y., Li L., Liu C., Yamagami T., Taketo M. M., Zhou C. and Chen Y. P. (2011). Epithelial Wnt/beta-catenin signaling regulates palatal shelf fusion through regulation of Tgfbeta3 expression. Dev. Biol. 350, 511-519. 10.1016/j.ydbio.2010.12.021 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Herr P. and Basler K. (2012). Porcupine-mediated lipidation is required for Wnt recognition by Wls. Dev. Biol. 361, 392-402. 10.1016/j.ydbio.2011.11.003 [DOI] [PubMed] [Google Scholar]
- Hill C. R., Jacobs B. H., Brown C. B., Barnett J. V. and Goudy S. L. (2015). Type III transforming growth factor beta receptor regulates vascular and osteoblast development during palatogenesis. Dev. Dyn. 244, 122-133. 10.1002/dvdy.24225 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Hikasa H. and Sokol S. Y. (2013). Wnt signaling in vertebrate axis specification. Cold Spring Harb. Perspect. Biol. 5, a007955 10.1101/cshperspect.a007955 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Ho H. Y., Susman M. W., Bikoff J. B., Ryu Y. K., Jonas A. M., Hu L., Kuruvilla R. and Greenberg M. E. (2012). Wnt5a-Ror-Dishevelled signaling constitutes a core developmental pathway that controls tissue morphogenesis. Proc. Natl. Acad. Sci. USA 109, 4044-4051. 10.1073/pnas.1200421109 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Hoebel A. K., Drichel D., van de Vorst M., Bohmer A. C., Sivalingam S., Ishorst N., Klamt J., Golz L., Alblas M., Maaser A. et al. (2017). Candidate genes for nonsyndromic cleft palate detected by exome sequencing. J. Dent. Res. 96, 1314-1321. 10.1177/0022034517722761 [DOI] [PubMed] [Google Scholar]
- Hoornaert K. P., Vereecke I., Dewinter C., Rosenberg T., Beemer F. A., Leroy J. G., Bendix L., Björck E., Bonduelle M., Boute O. et al. (2010). Stickler syndrome caused by COL2A1 mutations: genotype-phenotype correlation in a series of 100 patients. Eur. J. Hum. Genet. 18, 872-880. 10.1038/ejhg.2010.23 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Hosokawa R., Deng X., Takamori K., Xu X., Urata M., Bringas P. and Chai Y. (2009). Epithelial-specific requirement of FGFR2 signaling during tooth and palate development. J. Exp. Zool. B. Mol. Dev. Evol. 312B, 343-350. 10.1002/jez.b.21274 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Hozyasz K. K., Mostowska A., Wojcicki P., Lasota A., Offert B., Balcerek A., Dunin-Wilczynska I. and Jagodzinski P. P. (2014). Nucleotide variants of the cancer predisposing gene CDH1 and the risk of non-syndromic cleft lip with or without cleft palate. Fam. Cancer 13, 415-421. 10.1007/s10689-014-9727-2 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Hu X., Gao J., Liao Y., Tang S. and Lu F. (2013). Retinoic acid alters the proliferation and survival of the epithelium and mesenchyme and suppresses Wnt/β-catenin signaling in developing cleft palate. Cell Death Dis. 4, e898 10.1038/cddis.2013.424 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Huang X., Litingtung Y. and Chiang C. (2007). Ectopic sonic hedgehog signaling impairs telencephalic dorsal midlinedevelopment: implication for human holoprosencephaly. Hum. Mol. Genet. 16, 1454-1468. 10.1093/hmg/ddm096 [DOI] [PubMed] [Google Scholar]
- Huang X., Goudy S. L., Ketova T., Litingtung Y. and Chiang C. (2008). Gli3-deficient mice exhibit cleft palate associated with abnormal tongue development. Dev. Dyn 237, 3079-3087. 10.1002/dvdy.21714 [DOI] [PubMed] [Google Scholar]
- Huang H. J., Zhou L. L., Fu W. J., Zhang C. Y., Jiang H., Du J. and Hou J. (2015). β-catenin SUMOylation is involved in the dysregulated proliferation of myeloma cells. Am. J. Cancer Res. 5, 309-320. [PMC free article] [PubMed] [Google Scholar]
- Huangfu D. and Anderson K. V (2005). Cilia and Hedgehog responsiveness in the mouse. Proc. Natl. Acad. Sci. USA 102, 11325-11330. 10.1073/pnas.0505328102 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Huangfu D., Liu A., Rakeman A. S., Murcia N. S., Niswander L. and Anderson K. V. (2003). Hedgehog signaling in the mouse requires intraflagellar transport proteins. Nature 426, 83-87. 10.1038/nature02061 [DOI] [PubMed] [Google Scholar]
- Ibrahim I., Serrano M. J. and Svoboda K. K. H. (2015). Method of studying palatal fusion using static organ culture. J. Vis. Exp. e53063, 10.3791/53063 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Ichikawa E., Watanabe A., Nakano Y., Akita S., Hirano A., Kinoshita A., Kondo S., Kishino T., Uchiyama T., Niikawa N. et al. (2006). PAX9 and TGFB3 are linked to susceptibility to nonsyndromic cleft lip with or without cleft palate in the Japanese: population-based and family-based candidate gene analyses. J. Hum. Genet. 51, 38-46. 10.1007/s10038-005-0319-8 [DOI] [PubMed] [Google Scholar]
- Ishii M., Merrill A. E., Chan Y. S., Gitelman I., Rice D. P., Sucov H. M. and Maxson R. E. (2003). Msx2 and Twist cooperatively control the development of the neural crest-derived skeletogenic mesenchyme of the murine skull vault. Development 130, 6131-6142. 10.1242/dev.00793 [DOI] [PubMed] [Google Scholar]
- Ishii M., Han J., Yen H. Y., Sucov H. M., Chai Y. and Maxson R. E. (2005). Combined deficiencies of Msx1 and Msx2 cause impaired patterning and survival of the cranial neural crest. Development 132, 4937-4950. 10.1242/dev.02072 [DOI] [PubMed] [Google Scholar]
- Ito Y., Yeo J. Y., Chytil A., Han J., Bringas P. Jr, Nakajima A., Shuler C. F., Moses H. L. and Chai Y. (2003). Conditional inactivation of Tgfbr2 in cranial neural crest causes cleft palate and calvaria defects. Development 130, 5269-5280. 10.1242/dev.00708 [DOI] [PubMed] [Google Scholar]
- Ittiwut R., Ittiwut C., Siriwan P., Chichareon V., Suphapeetiporn K. and Shotelersuk V. (2016). Variants of the CDH1 (E-Cadherin) gene associated with oral clefts in the Thai population. Genet. Test Mol. Biomarkers 20, 406-409. 10.1089/gtmb.2015.0325 [DOI] [PubMed] [Google Scholar]
- Iwata J., Parada C. and Chai Y. (2011). The mechanism of TGF-ß signaling during palate development. Oral Dis. 17, 733-744. 10.1111/j.1601-0825.2011.01806.x [DOI] [PMC free article] [PubMed] [Google Scholar]
- Iwata J., Tung L., Urata M., Hacia J. G., Pelikan R., Suzuki A., Ramenzoni L., Chaudhry O., Parada C., Sanchez-Lara P. A. et al. (2012). Fibroblast growth factor 9 (FGF9)-pituitary homeobox 2 (PITX2) pathway mediates transforming growth factor beta (TGFbeta) signaling to regulate cell proliferation in palatal mesenchyme during mouse palatogenesis. J. Biol. Chem. 287, 2353-2363. 10.1074/jbc.M111.280974 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Iwata J., Suzuki A., Yokota T., Ho T. V., Pelikan R., Urata M., Sanchez-Lara P. A. and Chai Y. (2014). TGFβ regulates epithelial-mesenchymal interactions through WNT signaling activity to control muscle development in the soft palate. Development 141, 909-917. 10.1242/dev.103093 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Iyyanar P. P. R. and Nazarali A. J. (2017). Hoxa2 inhibits bone morphogenetic protein signaling during osteogenic differentiation of the palatal mesenchyme. Front Physiol. 8, 929 10.3389/fphys.2017.00929 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Jeong J., Mao J., Tenzen T., Kottmann A. H. and McMahon A. P. (2004). Hedgehog signaling in the neural crest cells regulates the patterning and growth of facial primordia. Genes Dev. 18, 937-951. 10.1101/gad.1190304 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Jia S., Kwon H. E., Lan Y., Zhou J., Liu H. and Jiang R. (2016). Bmp4-Msx1 signaling and Osr2 control tooth organogenesis through antagonistic regulation of secreted Wnt antagonists. Dev. Biol. 420, 110-119. 10.1016/j.ydbio.2016.10.001 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Jia S., Zhou J., Fanelli C., Wee Y., Bonds J., Schneider P., Mues G. and D'Souza R. N. (2017a). Small-molecule Wnt agonists correct cleft palates in Pax9 mutant mice in utero. Development 144, 3819-3828. 10.1242/dev.157750 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Jia S., Zhou J., Wee Y., Mikkola M. L., Schneider P. and D'Souza R. N. (2017b). Anti-EDAR agonist antibody therapy resolves palate defects in Pax9-/- mice. J. Dent. Res. 96, 1282-1289. 10.1177/0022034517726073 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Jiang R., Bush J. O. and Lidral A. C. (2006). Development of the upper lip: morphogenetic and molecular mechanisms. Dev. Dyn. 235, 1152-1166. 10.1002/dvdy.20646 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Jiang Z., Pan L., Chen X., Chen Z. and Xu D. (2017). Wnt6 influences the viability of mouse embryonic palatal mesenchymal cells via the β-catenin pathway. Exp. Ther. Med. 14, 5339-5344. 10.3892/etm.2017.5240 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Jin J.-Z. and Ding J. (2014). Strain-dependent effects of transforming growth factor-beta1 and 2 during mouse secondary palate development. Reprod. Toxicol. 50, 129-133. 10.1016/j.reprotox.2014.10.018 [DOI] [PubMed] [Google Scholar]
- Jin Y. R., Turcotte T. J., Crocker A. L., Han X. H. and Yoon J. K. (2011). The canonical Wnt signaling activator, R-spondin2, regulates craniofacial patterning and morphogeneis within the branchial arch through ectodermal-mesenchymal interaction. Dev. Biol. 352, 1-13. 10.1016/j.ydbio.2011.01.004 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Jin Y. R., Han X. H., Taketo M. M. and Yoon J. K. (2012). Wnt9b-dependent FGF signaling is crucial for outgrowth of the nasal and maxillary processes during upper jaw and lip development. Development 139, 1821-1830. 10.1242/dev.075796 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Juriloff D. M. and Harris M. J. (2008). Mouse genetic models of cleft lip with or without cleft palate. Birth Defects Res. A Clin. Mol. Teratol 82, 63-77. 10.1002/bdra.20430 [DOI] [PubMed] [Google Scholar]
- Juriloff D. M., Harris M. J., McMahon A. P., Carroll T. J. and Lidral A. C. (2006). Wnt9b is the mutated gene involved in multifactorial nonsyndromic cleft lip with or without cleft palate in A/WySn mice, as confirmed by a genetic complementation test. Birth Defects Res. A Clin. Mol. Teratol 76, 574-579. 10.1002/bdra.20302 [DOI] [PubMed] [Google Scholar]
- Juriloff D. M., Harris M. J., Mager D. L. and Gagnier L. (2014). Epigenetic mechanism causes Wnt9b deficiency and nonsyndromic cleft lip and palate in the A/WySn mouse strain. Birth Defects Res. A Clin. Mol. Teratol 100, 772-788. 10.1002/bdra.23320 [DOI] [PubMed] [Google Scholar]
- Kaartinen V., Voncken J. W., Shuler C., Warburton D., Bu D., Heisterkamp N. and Groffen J. (1995). Abnormal lung development and cleft palate in mice lacking TGF-beta 3 indicates defects of epithelial-mesenchymal interaction. Nat. Genet. 11, 415-421. 10.1038/ng1295-415 [DOI] [PubMed] [Google Scholar]
- Kaartinen V., Cui X. M., Heisterkamp N., Groffen J. and Shuler C. F. (1997). Transforming growth factor-β3 regulates transdifferentiation of medial edge epithelium during palatal fusion and associated degradation of the basement membrane. Dev. Dyn. 209, 255-260. [DOI] [PubMed] [Google Scholar]
- Kamel G., Hoyos T., Rochard L., Dougherty M., Kong Y., Tse W, Shubinets V., Grimaldi M. and Liao E. C. (2013). Requirement for frzb and fzd7a in cranial neural crest convergence and extension mechanisms during zebrafish palate and jaw morphogenesis. Dev. Biol. 381, 423-433. 10.1016/j.ydbio.2013.06.012 [DOI] [PubMed] [Google Scholar]
- Kato H., Izumi K., Saito T., Ohnuki H., Terada M., Kawano Y., Nozawa-Inoue K., Saito C. and Maeda T. (2013). Distinct expression patterns and roles of aldehyde dehydrogenases in normal oral mucosa keratinocytes: differential inhibitory effects of a pharmacological inhibitor and RNAi-mediated knockdown on cellular phenotype and epithelial morphology. Histochem. Cell Biol. 139, 847-862. 10.1007/s00418-012-1064-7 [DOI] [PubMed] [Google Scholar]
- Katoh I., Fukunishi N., Fujimuro M., Kasai H., Moriishi K., Hata R. and Kurata S. (2016). Repression of Wnt/β-catenin response elements by p63 (TP63). Cell Cycle 15, 699-710. 10.1080/15384101.2016.1148837 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kawasaki M., Kawasaki K., Meguro F., Yamada A., Ishikawa R., Porntaveetus T., Blackburn J., Otsuka-Tanaka Y., Saito N., Ota . S. et al. (2018). Lrp4/Wise regulates palatal rugae development through Turing-type reaction-diffusion mechanisms. PLoS One 13, e0204126 10.1371/journal.pone.0204126 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kennedy A. E. and Dickinson A. J. (2012). Median facial clefts in Xenopus laevis: roles of retinoic acid signaling and homeobox genes. Dev. Biol. 365, 229-240. 10.1016/j.ydbio.2012.02.033 [DOI] [PubMed] [Google Scholar]
- Kimura-Yoshida C., Nakano H., Okamura D., Nakao K., Yonemura S., Belo J. A., Aizawa S., Matsui Y. and Matsuo I. (2005). Canonical Wnt signaling and its antagonist regulate anterior-posterior axis polarization by guiding cell migration in mouse visceral endoderm. Dev. Cell 9, 639-650. 10.1016/j.devcel.2005.09.011 [DOI] [PubMed] [Google Scholar]
- Kimura-Yoshida C., Mochida K., Ellwanger K., Niehrs C. and Matsuo I. (2015). Fate specification of neural plate border by canonical Wnt signaling and Grhl3 is crucial for neural tube closure. EBioMedicine 2, 513-527. 10.1016/j.ebiom.2015.04.012 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kioussi C., Briata P., Baek S. H., Rose D. W., Hamblet N. S., Herman T., Ohgi K. A., Lin C., Gleiberman A., Wang J. et al. (2002). Identification of a Wnt/Dvl/β-catenin→Pitx2 pathway mediating cell-type-specific proliferation during development. Cell 111, 673-685. 10.1016/S0092-8674(02)01084-X [DOI] [PubMed] [Google Scholar]
- Knight A. S., Schutte B. C., Jiang R. and Dixon M. J. (2006). Developmental expression analysis of the mouse and chick orthologues of IRF6: the gene mutated in Van der Woude syndrome. Dev. Dyn. 235, 1441-1447. 10.1002/dvdy.20598 [DOI] [PubMed] [Google Scholar]
- Komiya Y. and Habas R. (2008). Wnt signal transduction pathways. Organogenesis 4, 68-75. 10.4161/org.4.2.5851 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Koneczny I., Schulenburg A., Hudec X., Knöfler M., Holzmann K., Piazza G., Reynolds R., Valent P. and Marian B. (2015). Autocrine fibroblast growth factor 18 signaling mediates Wnt-dependent stimulation of CD44-positive human colorectal adenoma cells. Mol. Carcinog. 54, 789-799. 10.1002/mc.22146 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kousa Y. A. and Schutte B. C. (2016). Toward an orofacial gene regulatory network. Dev. Dyn. 245, 220-232. 10.1002/dvdy.24341 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kousa Y. A., Roushangar R., Patel N., Walter A., Marangoni P., Krumlauf R., Klein O. D. and Schutte B. C. (2017). IRF6 and SPRY4 signaling interact in periderm development. J. Dent. Res. 96, 1306-1313. 10.1177/0022034517719870 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kouskoura T., Kozlova A., Alexiou M., Blumer S., Zouvelou V., Katsaros C., Chiquet M., Mitsiadis T. A. and Graf D. (2013). The etiology of cleft palate formation in BMP7-deficient mice. PLoS One 8, e59463 10.1371/journal.pone.0059463 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kratochwil K., Galceran J., Tontsch S., Roth W. and Grosschedl R. (2002). FGF4, a direct target of LEF1 and Wnt signaling, can rescue the arrest of tooth organogenesis in Lef1−/− mice. Genes Dev. 16, 3173-3185. 10.1101/gad.1035602 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kumar S. and Duester G. (2010). Retinoic acid signaling in perioptic mesenchyme represses Wnt signaling via induction of Pitx2 and Dkk2. Dev. Biol. 340, 67-74. 10.1016/j.ydbio.2010.01.027 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kurosaka H., Iulianella A., Williams T. and Trainor P. A. (2014). Disrupting hedgehog and WNT signaling interactions promotes cleft lip pathogenesis. J. Clin. Invest. 124, 1660-1671. 10.1172/JCI72688 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Lan Y. and Jiang R. (2009). Sonic hedgehog signaling regulates reciprocal epithelial-mesenchymal interactions controlling palatal outgrowth. Development 136, 1387-1396. 10.1242/dev.028167 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Lan Y., Ovitt C. E., Cho E. S., Maltby K. M., Wang Q. and Jiang R. (2004). Odd-skipped related 2 (Osr2) encodes a key intrinsic regulator of secondary palate growth and morphogenesis. Development 131, 3207-3216. 10.1242/dev.01175 [DOI] [PubMed] [Google Scholar]
- Lan Y., Ryan R. C., Zhang Z., Bullard S. A., Bush J. O., Maltby K. M., Lidral A. C. and Jiang R. (2006). Expression of Wnt9b and activation of canonical Wnt signaling during midfacial morphogenesis in mice. Dev. Dyn. 235, 1448-1454. 10.1002/dvdy.20723 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Lane J., Yumoto K., Pisano J., Azhar M., Thomas P. S. and Kaartinen V. (2014). Control elements targeting Tgfb3 expression to the palatal epithelium are located intergenically and in introns of the upstream Ift43 gene. Front Physiol. 5, 258 10.3389/fphys.2014.00258 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Lee J. M., Kim J. Y., Cho K. W., Lee M. J., Cho S. W., Kwak S., Cai J. and Jung H. S. (2008). Wnt11/Fgfr1b cross-talk modulates the fate of cells in palate development. Dev. Biol. 314, 341-350. 10.1016/j.ydbio.2007.11.033 [DOI] [PubMed] [Google Scholar]
- Leslie E. J., Mansilla M. A., Biggs L. C., Schuette K., Bullard S., Cooper M., Dunnwald M., Lidral A. C., Marazita M. L., Beaty T. H. et al. (2012). Expression and mutation analyses implicate ARHGAP29 as the etiologic gene for the cleft lip with or without cleft palate locus identified by genome-wide association on chromosome 1p22. Birth Defects Res. A Clin. Mol. Teratol 94, 934-942. 10.1002/bdra.23076 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Leslie E. J., Taub M. A., Liu H., Steinberg K. M., Koboldt D. C., Zhang Q., Carlson J. C., Hetmanski J. B., Wang H., Larson D. E. et al. (2015). Identification of functional variants for cleft lip with or without cleft palate in or near PAX7, FGFR2, and NOG by targeted sequencing of GWAS loci. Am. J. Hum. Genet. 96, 397-411. 10.1016/j.ajhg.2015.01.004 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Leslie E. J., Liu H., Carlson J. C., Shaffer J. R., Feingold E., Wehby G., Laurie C. A., Jain D., Laurie C. C., Doheny K. F. et al. (2016). A genome-wide association study of nonsyndromic cleft palate identifies an etiologic missense variant in GRHL3. Am. J. Hum. Genet. 98, 744-754. 10.1016/j.ajhg.2016.02.014 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Letra A., Menezes R., Granjeiro J. M. and Vieira A. R. (2009). AXIN2 and CDH1 polymorphisms, tooth agenesis and oral clefts. Birth Defects Res. A Clin. Mol. Teratol. 85, 169-173. 10.1002/bdra.20489 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Letra A., Bjork B., Cooper M. E., Szabo-Rogers H., Deleyiannis F. W. B., Field L. L., Czeizel A. E., Ma L., Garlet G. P., Poletta F. A. et al. (2012). Association of AXIN2 with non-syndromic oral clefts in multiple populations. J. Dent. Res. 91, 473-478. 10.1177/0022034512440578 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Leung A. W., Murdoch B., Salem A. F., Prasad M. S., Gomez G. A. and Garcia-Castro M. I. (2016). WNT/beta-catenin signaling mediates human neural crest induction via a pre-neural border intermediate. Development 143, 398-410. 10.1242/dev.130849 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Levi B., Brugman S., Wong V. W., Grova M., Longaker M. T. and Wan D. C. (2011). Palatogenesis: engineering, pathways and pathologies. Organogenesis 7, 242-254. 10.4161/org.7.4.17926 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Li C., Xiao J., Hormi K., Borok Z. and Minoo P. (2002). Wnt5a participates in distal lung morphogenesis. Dev. Biol. 248, 68-81. 10.1006/dbio.2002.0729 [DOI] [PubMed] [Google Scholar]
- Li B., Kuriyama S., Moreno M. and Mayor R. (2009). The posteriorizing gene Gbx2 is a direct target of Wnt signalling and the earliest factor in neural crest induction. Development 136, 3267-3278. 10.1242/dev.036954 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Li Q., Kim Y., Suktitipat B., Hetmanski J. B., Marazita M. L., Duggal P., Beaty T. H. and Bailey-Wilson J. E. (2015). Gene-gene interaction among WNT genes for oral cleft in trios. Genet. Epidemiol. 39, 385-394. 10.1002/gepi.21888 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Li J. Y., Zou J. J., Li Q., Chen L., Gao Y. L., Yan H., Zhou B. and Li J. (2016). Assessment of differentially expressed plasma microRNAs in nonsyndromic cleft palate and nonsyndromic cleft lip with cleft palate. Oncotarget 7, 86266-86279. 10.18632/oncotarget.13379 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Li C., Lan Y., Krumlauf R. and Jiang R. (2017). Modulating Wnt signaling rescues palate morphogenesis in Pax9 mutant mice. J. Dent. Res. 96, 1273-1281. 10.1177/0022034517719865 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Lin C., Fisher A. V., Yin Y., Maruyama T., Veith G. M., Dhandha M., Huang G. J., Hsu W. and Ma L. (2011). The inductive role of Wnt-β-Catenin signaling in the formation of oral apparatus. Dev. Biol. 356, 40-50. 10.1016/j.ydbio.2011.05.002 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Lipinski R. J., Song C., Sulik K. K., Everson J. L., Gipp J. J., Yan D., Bushman W. and Rowland I. J. (2010). Cleft lip and palate results from Hedgehog signaling antagonism in the mouse: phenotypic characterization and clinical implications. Birth Defects Res. A Clin. Mol. Teratol 88, 232-240. 10.1002/bdra.20656 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Liu P., Wakamiya M., Shea M. J., Albrecht U., Behringer R. R. and Bradley A. (1999). Requirement for Wnt3 in vertebrate axis formation. Nat. Genet. 22, 361-365. 10.1038/11932 [DOI] [PubMed] [Google Scholar]
- Liu Z., Xu J., Colvin J. S. and Ornitz D. M. (2002). Coordination of chondrogenesis and osteogenesis by fibroblast growth factor 18. Genes Dev. 16, 859-869. 10.1101/gad.965602 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Liu W., Selever J., Lu M. F. and Martin J. F. (2003). Genetic dissection of Pitx2 in craniofacial development uncovers new functions in branchial arch morphogenesis, late aspects of tooth morphogenesis and cell migration. Development 130, 6375-6385. 10.1242/dev.00849 [DOI] [PubMed] [Google Scholar]
- Liu K. J., Arron J. R., Stankunas K., Crabtree G. R. and Longaker M. T. (2007). Chemical rescue of cleft palate and midline defects in conditional GSK-3beta mice. Nature 446, 79-82. 10.1038/nature05557 [DOI] [PubMed] [Google Scholar]
- Liu C., Lin C., Gao C., May-Simera H., Swaroop A. and Li T. (2014a). Null and hypomorph Prickle1 alleles in mice phenocopy human Robinow syndrome and disrupt signaling downstream of Wnt5a. Biology Open 3, 861-870. 10.1242/bio.20148375 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Liu B., Chen S., Johnson C. and Helms J. A. (2014b). A ciliopathy with hydrocephalus, isolated craniosynostosis, hypertelorism, and clefting caused by deletion of Kif3a. Reprod. Toxicol. 48, 88-97. 10.1016/j.reprotox.2014.05.009 [DOI] [PubMed] [Google Scholar]
- Liu Y., Wang M., Zhao W., Yuan X., Yang X., Li Y., Qiu M., Zhu X. J. and Zhang Z. (2015). Gpr177-mediated Wnt signaling is required for secondary palate development. J. Dent. Res. 94, 961-967. 10.1177/0022034515583532 [DOI] [PubMed] [Google Scholar]
- Lombardi M. P., Bulk S., Celli J., Lampe A., Gabbett M. T., Ousager L. B., van der Smagt J. J., Soller M., Stattin E. L., Mannens M. A. et al. (2011). Mutation update for the PORCN gene. Hum. Mutat. 32, 723-728. 10.1002/humu.21505 [DOI] [PubMed] [Google Scholar]
- Lough K. J., Byrd K. M., Spitzer D. C. and Williams S. E. (2017). Closing the gap: mouse models to study adhesion in secondary palatogenesis. J. Dent. Res. 96, 1210-1220. 10.1177/0022034517726284 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Lu Y. P., Han W. T., Liu Q., Li J. X., Li Z. J., Jiang M. and Xu W. (2015). Variations in WNT3 gene are associated with incidence of non-syndromic cleft lip with or without cleft palate in a northeast Chinese population. Genet. Mol. Res. 14, 12646-12653. 10.4238/2015.October.19.8 [DOI] [PubMed] [Google Scholar]
- Luis T. C., Weerkamp F., Naber B. A., Baert M. R., de Haas E. F., Nikolic T., Heuvelmans S., De Krijger R. R., van Dongen J. J. and Staal F. J. (2009). Wnt3a deficiency irreversibly impairs hematopoietic stem cell self-renewal and leads to defects in progenitor cell differentiation. Blood 113, 546-554. 10.1182/blood-2008-06-163774 [DOI] [PubMed] [Google Scholar]
- Machado R. A., Messetti A. C., de Aquino S. N., Martelli-Junior H., Swerts M. S., de Almeida Reis S. R., Moreira H. S., Persuhn D. C. and Coletta R. D. (2016). Association between genes involved in craniofacial development and nonsyndromic cleft lip and/or palate in the Brazilian population. Cleft Palate Craniofac. J. 53, 550-556. 10.1597/15-107 [DOI] [PubMed] [Google Scholar]
- Maher M. T., Flozak A. S., Stocker A. M., Chenn A. and Gottardi C. J. (2009). Activity of the β-catenin phosphodestruction complex at cell–cell contacts is enhanced by cadherin-based adhesion. J. Cell Biol. 186, 219-228. 10.1083/jcb.200811108 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Mangold E., Ludwig K. U., Birnbaum S., Baluardo C., Ferrian M., Herms S., Reutter H., de Assis N. A., Chawa T. A., Mattheisen M. et al. (2010). Genome-wide association study identifies two susceptibility loci for nonsyndromic cleft lip with or without cleft palate. Nat. Genet. 42, 24-26. 10.1038/ng.506 [DOI] [PubMed] [Google Scholar]
- Mangold E., Böhmer A. C., Ishorst N., Hoebel A.-K., Gültepe P., Schuenke H., Klamt J., Hofmann A., Gölz L., Raff R. et al. (2016). Sequencing the GRHL3 coding region reveals rare truncating mutations and a common susceptibility variant for nonsyndromic cleft palate. Am. J. Hum. Genet. 98, 755-762. 10.1016/j.ajhg.2016.02.013 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Mani P., Jarrell A., Myers J. and Atit R. (2010). Visualizing canonical Wnt signaling during mouse craniofacial development. Dev. Dyn. 239, 354-363. 10.1002/dvdy.22072 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Marçano A. C., Doudney K., Braybrook C., Squires R., Patton M. A., Lees M. M., Richieri-Costa A., Lidral A. C., Murray J. C. and Moore G. E. (2004). TBX22 mutations are a frequent cause of cleft palate. J. Med. Genet. 41, 68-74. 10.1136/jmg.2003.010868 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Marie P. J. and Haÿ E. (2013). Cadherins and Wnt signaling: a functional link controlling bone formation. Bonekey Rep. 2, 330 10.1038/bonekey.2013.64 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Martinelli M., Carinci F., Morselli P. G., Caramelli E., Palmieri A., Girardi A., Riberti C. and Scapoli L. (2011a). Evidence of LEF1 fetal-maternal interaction in cleft lip with or without cleft palate in a consistent Italian sample study. Int. J. Immunopathol. Pharmacol. 24, 15-19. 10.1177/03946320110240S204 [DOI] [PubMed] [Google Scholar]
- Martinelli M., Masiero E., Carinci F., Morselli P. G., Palmieri A., Girardi A., Baciliero U. and Scapoli L. (2011b). Evidence of an involvement of TFAP2A gene in non-syndromic cleft lip with or without cleft palate: an Italian study. Int. J. Immunopathol. Pharmacol. 24, 7-10. 10.1177/03946320110240S202 [DOI] [PubMed] [Google Scholar]
- Massink M. P., Créton M. A., Spanevello F., Fennis W. M., Cune M. S., Savelberg S. M., Nijman I. J., Maurice M. M., van den Boogaard M. J. and van Haaften G. (2015). Loss-of-function mutations in the WNT co-receptor LRP6 cause autosomal-dominant oligodontia. Am. J. Hum. Genet. 97, 621-626. 10.1016/j.ajhg.2015.08.014 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Matsui M. and Klingensmith J. (2014). Multiple tissue-specific requirements for the BMP antagonist Noggin in development of the mammalian craniofacial skeleton. Dev. Biol. 392, 168-181. 10.1016/j.ydbio.2014.06.006 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Matsumura K., Taketomi T., Yoshizaki K., Arai S., Sanui T., Yoshiga D., Yoshimura A. and Nakamura S. (2011). Sprouty2 controls proliferation of palate mesenchymal cells via fibroblast growth factor signaling. Biochem. Biophys. Res. Commun. 404, 1076-1082. 10.1016/j.bbrc.2010.12.116 [DOI] [PubMed] [Google Scholar]
- McGee-Lawrence M. E., Li X., Bledsoe K. L., Wu H., Hawse J. R., Subramaniam M., Razidlo D. F., Stensgard B. A., Stein G. S., van Wijnen A. J. et al. (2013). Runx2 protein represses Axin2 expression in osteoblasts and is required for craniosynostosis in Axin2-deficient mice. J. Biol. Chem. 288, 5291-5302. 10.1074/jbc.M112.414995 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Medio M., Yeh E., Popelut A., Babajko S., Berdal A. and Helms J. A. (2012). Wnt/β-catenin signaling and Msx1 promote outgrowth of the maxillary prominences. Front Physiol. 3, 375 10.3389/fphys.2012.00375 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Menezes R., Letra A., Kim A. H., Küchler E. C., Day A., Tannure P. N., Gomes da Motta L., Paiva K. B., Granjeiro J. M. and Vieira A. et al. (2010). Studies with Wnt genes and nonsyndromic cleft lip and palate. Birth Defects Res. A Clin. Mol. Teratol. 88, 995-1000. 10.1002/bdra.20720 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Mensah J. K., Ogawa T., Kapadia H., Cavender A. C. and D'Souza R. N. (2004). Functional analysis of a mutation in PAX9 associated with familial tooth agenesis in humans. J. Biol. Chem. 279, 5924-5933. 10.1074/jbc.M305648200 [DOI] [PubMed] [Google Scholar]
- Miles L. B., Dworkin S. and Darido C. (2017). Alternative splicing and start sites: Lessons from the Grainyhead-like family. Dev. Biol. 429, 12-19. 10.1016/j.ydbio.2017.06.018 [DOI] [PubMed] [Google Scholar]
- Morikawa Y., D'Autréaux F., Gershon M. D. and Cserjesi P. (2007). Hand2 determines the noradrenergic phenotype in the mouse sympathetic nervous system. Dev. Biol. 307, 114-126. 10.1016/j.ydbio.2007.04.027 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Mossey P. A., Little J., Munger R. G., Dixon M. J. and Shaw W. C. (2009). Cleft lip and palate. Lancet 374, 1773-1785. 10.1016/S0140-6736(09)60695-4 [DOI] [PubMed] [Google Scholar]
- Mostowska A., Hozyasz K. K., Biedziak B., Wojcicki P., Lianeri M. and Jagodzinski P. P. (2012). Genotype and haplotype analysis of WNT genes in non-syndromic cleft lip with or without cleft palate. Eur. J. Oral Sci. 120, 1-8. 10.1111/j.1600-0722.2011.00938.x [DOI] [PubMed] [Google Scholar]
- Nam J. S., Park E., Turcotte T. J., Palencia S., Zhan X., Lee J., Yun K., Funk W. D. and Yoon J. K. (2007). Mouse R-spondin2 is required for apical ectodermal ridge maintenance in the hindlimb. Dev. Biol. 311, 124-135. 10.1016/j.ydbio.2007.08.023 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Nawshad A. and Hay E. D. (2003). TGFβ3 signaling activates transcription of the LEF1 gene to induce epithelial mesenchymal transformation during mouse palate development. J. Cell Biol. 163, 1291-1301. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Nawshad A., Medici D., Liu C. C. and Hay E. D. (2007). TGFβ3 inhibits E-cadherin gene expression in palate medial-edge epithelial cells through a Smad2-Smad4-LEF1 transcription complex. J. Cell Sci. 120, 1646-1653. 10.1242/jcs.003129 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Neiswender H., Navarre S., Kozlowski D. J. and LeMosy E. K. (2017). Early craniofacial defects in zebrafish that have reduced function of a Wnt-interacting extracellular matrix protein, Tinagl1. Cleft Palate Craniofac. J. 54, 381-390. 10.1597/15-283 [DOI] [PubMed] [Google Scholar]
- Nelson E. R., Levi B., Sorkin M., James A. W., Liu K. J., Quarto N. and Longaker M. T. (2011). Role of GSK-3beta in the osteogenic differentiation of palatal mesenchyme. PLoS One 6, e25847 10.1371/journal.pone.0025847 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Neubüser A., Peters H., Balling R. and Martin G. R. (1997). Antagonistic interactions between FGF and BMP signaling pathways: a mechanism for positioning the sites of tooth formation. Cell 90, 247-255. 10.1016/S0092-8674(00)80333-5 [DOI] [PubMed] [Google Scholar]
- Niehrs C. (2012). The complex world of WNT receptor signalling. Nat. Rev. Mol. Cell Biol. 13, 767 10.1038/nrm3470 [DOI] [PubMed] [Google Scholar]
- Niemann S., Zhao C., Pascu F., Stahl U., Aulepp U., Niswander L., Weber J. L. and Muller U. (2004). Homozygous WNT3 mutation causes tetra-amelia in a large consanguineous family. Am. J. Hum. Genet. 74, 558-563. 10.1086/382196 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Nik A. M., Reyahi A., Ponten F. and Carlsson P. (2013). Foxf2 in intestinal fibroblasts reduces numbers of Lgr5(+) stem cells and adenoma formation by inhibiting Wnt signaling. Gastroenterology 144, 1001-1011. 10.1053/j.gastro.2013.01.045 [DOI] [PubMed] [Google Scholar]
- Nik A. M., Johansson J. A., Ghiami M., Reyahi A. and Carlsson P. (2016). Foxf2 is required for secondary palate development and Tgfbeta signaling in palatal shelf mesenchyme. Dev. Biol. 415, 14-23. 10.1016/j.ydbio.2016.05.013 [DOI] [PubMed] [Google Scholar]
- Nikopensius T., Jagomägi T., Krjutskov K., Tammekivi V., Saag M., Prane I., Piekuse L., Akota I., Barkane B., Krumina A. et al. (2010). Genetic variants in COL2A1, COL11A2, and IRF6 contribute risk to nonsyndromic cleft palate. Birth Defects Res. A Clin. Mol. Teratol 88, 748-756. 10.1002/bdra.20700 [DOI] [PubMed] [Google Scholar]
- Nikopensius T., Kempa I., Ambrozaityte L., Jagomägi T., Saag M., Matulevičiene˙ A., Utkus A., Krjutskov K., Tammekivi V., Piekuse L. et al. (2011). Variation in FGF1, FOXE1, and TIMP2 genes is associated with nonsyndromic cleft lip with or without cleft palate. Birth Defects Res. A Clin. Mol. Teratol 91, 218-225. 10.1002/bdra.20791 [DOI] [PubMed] [Google Scholar]
- Nishihara H., Kobayashi N., Kimura-Yoshida C., Yan K., Bormuth O., Ding Q., Nakanishi A., Sasaki T., Hirakawa M., Sumiyama K. et al. (2016). Coordinately co-opted multiple transposable elements constitute an enhancer for wnt5a expression in the mammalian secondary palate. PLoS Genet. 12, e1006380 10.1371/journal.pgen.1006380 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Noda K., Mishina Y. and Komatsu Y. (2016). Constitutively active mutation of ACVR1 in oral epithelium causes submucous cleft palate in mice. Dev. Biol. 415, 306-313. 10.1016/j.ydbio.2015.06.014 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Nunnally A. P. and Parr B. A. (2004). Analysis of Fz10 expression in mouse embryos. Dev. Genes Evol. 214, 144-148. 10.1007/s00427-004-0386-4 [DOI] [PubMed] [Google Scholar]
- Nusse R. and Clevers H. (2017). Wnt/beta-catenin signaling, disease, and emerging therapeutic modalities. Cell 169, 985-999. 10.1016/j.cell.2017.05.016 [DOI] [PubMed] [Google Scholar]
- Ockeloen C. W., Khandelwal K. D., Dreesen K., Ludwig K. U., Sullivan R., van Rooji I. A. L. M., Thonissen M., Swinnen S., Phan M., Conte F. et al. (2016). Novel mutations in LRP6 highlight the role of WNT signaling in tooth agenesis. Genet. Med. 18, 1158-1162. 10.1038/gim.2016.10 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Oishi I., Suzuki H., Onishi N., Takada R., Kani S., Ohkawara B., Koshida I., Suzuki K., Yamada G., Schwabe G. C. et al. (2003). The receptor tyrosine kinase Ror2 is involved in non-canonical Wnt5a/JNK signalling pathway. Genes Cells 8, 645-654. 10.1046/j.1365-2443.2003.00662.x [DOI] [PubMed] [Google Scholar]
- Okamoto M., Udagawa N., Uehara S., Maeda K., Yamashita T., Nakamichi Y., Kato H., Saito N., Minami Y., Takahashi N. et al. (2014). Non-canonical Wnt5a enhances Wnt/beta-catenin signaling during osteoblastogenesis. Sci. Rep. 4, 4493 10.1038/srep04493 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Okano J., Sakai Y. and Shiota K. (2008). Retinoic acid down-regulates Tbx1 expression and induces abnormal differentiation of tongue muscles in fetal mice. Dev. Dyn. 237, 3059-3070. 10.1002/dvdy.21715 [DOI] [PubMed] [Google Scholar]
- Okano J., Kimura W., Papaionnou V. E., Miura N., Yamada G., Shiota K. and Sakai Y. (2012). The regulation of endogenous retinoic acid level through CYP26B1 is required for elevation of palatal shelves. Dev. Dyn. 241, 1744-1756, 10.1002/dvdy.23862 [DOI] [PubMed] [Google Scholar]
- Okano J., Udagawa J. and Shiota K. (2014). Roles of retinoic acid signaling in normal and abnormal development of the palate and tongue. Congenit Anom (Kyoto) 54, 69-76. 10.1111/cga.12049 [DOI] [PubMed] [Google Scholar]
- Okello D. O., Iyyanar P. P. R., Kulyk W. M., Smith T. M., Lozanoff S., Ji S. and Nazarali A. J. (2017). Six2 plays an intrinsic role in regulating proliferation of mesenchymal cells in the developing palate. Front Physiol. 8, 955 10.3389/fphys.2017.00955 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Ordόñez-Moran P., Irmisch A., Barbachano A., Chicote I., Tenbaum S., Landolfi S., Tabernero J., Huelsken J., Muñoz A. and Palmer H. G. (2014). SPROUTY2 is a beta-catenin and FOXO3a target gene indicative of poor prognosis in colon cancer. Oncogene 33, 1975-1985. 10.1038/onc.2013.140 [DOI] [PubMed] [Google Scholar]
- Ormestad M., Astorga J., Landgren H., Wang T., Johansson B. R., Miura N. and Carlsson P. (2006). Foxf1 and Foxf2 control murine gut development by limiting mesenchymal Wnt signaling and promoting extracellular matrix production. Development 133, 833-843. 10.1242/dev.02252 [DOI] [PubMed] [Google Scholar]
- Osei-Sarfo K. and Gudas L. J. (2014). Retinoic acid suppresses the canonical Wnt signaling pathway in embryonic stem cells and activates the non-canonical Wnt signaling pathway. Stem Cells 32, 2061-2071. 10.1002/stem.1706 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Otto F., Thornell A. P., Crompton T., Denzel A., Gilmour K. C., Rosewell I. R., Stamp G. W., Beddington R. S., Mundlos S., Olsen B. R. et al. (1997). Cbfa1, a candidate gene for cleidocranial dysplasia syndrome, is essential for osteoblast differentiation and bone development. Cell 89, 765-771. 10.1016/S0092-8674(00)80259-7 [DOI] [PubMed] [Google Scholar]
- Panamonta V., Pradubwong S., Panamonta M. and Chowchuen B. (2015). Global birth prevalence of orofacial clefts: a systematic review. J. Med. Assoc. Thai. 98 Suppl. 7, S11-S21. [PubMed] [Google Scholar]
- Panetta N. J., Gupta D. M., Slater B. J., Kwan M. D., Liu K. J. and Longaker M. T. (2008). Tissue engineering in cleft palate and other congenital malformations. Pediatr. Res. 63, 545 10.1203/PDR.0b013e31816a743e [DOI] [PubMed] [Google Scholar]
- Parada C. and Chai Y. (2012). Roles of BMP signaling pathway in lip and palate development. Front. Oral Biol. 16, 60-70. 10.1159/000337617 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Paré A., Kim M., Juarez M. T., Brody S. and McGinnis W. (2012). The functions of grainy head-like proteins in animals and fungi and the evolution of apical extracellular barriers. PLoS One 7, e36254 10.1371/journal.pone.0036254 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Park J. S., Ma W., O'Brien L. L., Chung E., Guo J. J., Cheng J. G. and McMahon A. P. (2012). Six2 and Wnt regulate self-renewal and commitment of nephron progenitors through shared gene regulatory networks. Dev. Cell 23, 637-651. 10.1016/j.devcel.2012.07.008 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Pauws E., Hoshino A., Bentley L., Prajapati S., Keller C., Hammond P., Martinez-Barbera J. P., Moore G. E. and Stanier P. (2009). Tbx22null mice have a submucous cleft palate due to reduced palatal bone formation and also display ankyloglossia and choanal atresia phenotypes. Hum. Mol. Genet. 18, 4171-4179. 10.1093/hmg/ddp368 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Pengelly R. J., Arias L., Martínez J., Upstill-Goddard R., Seaby E. G., Gibson J., Ennis S., Collins A. and Briceño I. (2016). Deleterious coding variants in multi-case families with non-syndromic cleft lip and/or palate phenotypes. Sci. Rep. 6, 30457 10.1038/srep30457 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Perrimon N., Pitsouli C. and Shilo B. Z. (2012). Signaling mechanisms controlling cell fate and embryonic patterning. Cold Spring Harb. Perspect. Biol. 4, a005975 10.1101/cshperspect.a005975 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Person A. D., Beiraghi S., Sieben C. M., Hermanson S., Neumann A. N., Robu M. E., Schleiffarth J. R., Billington C. J. Jr, van Bokhoven H., Hoogeboom J. M. et al. (2010). WNT5A mutations in patients with autosomal dominant Robinow syndrome. Dev. Dyn. 239, 327-337. 10.1002/dvdy.22156 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Peyrard-Janvid M., Leslie E. J., Kousa Y. A., Smith T. L., Dunnwald M., Magnusson M., Lentz B. A., Unneberg P., Fransson I., Koillinen H. K. et al. (2014). Dominant mutations in GRHL3 cause Van der Woude Syndrome and disrupt oral periderm development. Am. J. Hum. Genet. 94, 23-32. 10.1016/j.ajhg.2013.11.009 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Port F. and Basler K. (2010). Wnt trafficking: new insights into Wnt maturation, secretion and spreading. Traffic 11, 1265-1271. 10.1111/j.1600-0854.2010.01076.x [DOI] [PubMed] [Google Scholar]
- Proetzel G., Pawlowski S. A., Wiles M. V., Yin M., Boivin G. P., Howles P. N., Ding J., Ferguson M. W. and Doetschman T. (1995). Transforming growth factor-beta 3 is required for secondary palate fusion. Nat. Genet. 11, 409-414. 10.1038/ng1295-409 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Rafighdoost H., Hashemi M., Narouei A., Eskanadri-Nasab E., Dashti-Khadivaki G. and Taheri M. (2013). Association between CDH1 and MSX1 gene polymorphisms and the risk of nonsyndromic cleft lip and/or cleft palate in a Southeast Iranian population. Cleft Palate Craniofac. J. 50, e98-e104. 10.1597/12-144 [DOI] [PubMed] [Google Scholar]
- Rahimov F., Marazita M. L., Visel A., Cooper M. E., Hitchler M. J., Rubini M., Domann F. E., Govil M., Christensen K., Bille C. et al. (2008). Disruption of an AP-2α binding site in an IRF6 enhancer is strongly associated with cleft lip. Nat. Genet. 40, 1341-1347. 10.1038/ng.242 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Reinhold M. I. and Naski M. C. (2007). Direct interactions of Runx2 and canonical Wnt signaling induce FGF18. J. Biol. Chem. 282, 3653-3663. 10.1074/jbc.M608995200 [DOI] [PubMed] [Google Scholar]
- Reynolds P. R., Schaalje G. B. and Seegmiller R. E. (2003). Combination therapy with folic acid and methionine in the prevention of retinoic acid-induced cleft palate in mice. Birth Defects Res. A Clin. Mol. Teratol. 67, 168-173. [DOI] [PubMed] [Google Scholar]
- Rice R., Spencer-Dene B., Connor E. C., Gritli-Linde A., McMahon A. P., Dickson C., Thesleff I. and Rice D. P. (2004). Disruption of Fgf10/Fgfr2b-coordinated epithelial-mesenchymal interactions causes cleft palate. J. Clin. Invest. 113, 1692-1700. 10.1172/JCI20384 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Rifat Y., Parekh V., Wilanowski T., Hislop N. R., Auden A., Ting S. B., Cunningham J. M. and Jane S. M. (2010). Regional neural tube closure defined by the Grainy head-like transcription factors. Dev. Biol. 345, 237-245. 10.1016/j.ydbio.2010.07.017 [DOI] [PubMed] [Google Scholar]
- Riley B. M., Mansilla M. A., Ma J., Daack-Hirsch S., Maher B. S., Raffensperger L. M., Russo E. T., Vieira A. R., Dodé C., Mohammadi M. et al. (2007). Impaired FGF signaling contributes to cleft lip and palate. Proc. Natl. Acad. Sci. USA 104, 4512-4517. 10.1073/pnas.0607956104 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Roosenboom J., Claes P., Devriendt K., Dormaar T., Peeters H., Saey I., Schoenaers J., Vander Poorten V., Verdonck A. and Hens G. (2015). Review: Facial endophenotypes in non-syndromic orofacial clefting. B-ENT 11, 173-182. [PubMed] [Google Scholar]
- Rochard L., Monica S. D., Ling I. T. C., Kong Y., Roberson S., Harland R., Halpern M. and .Liao E. C. (2016). Roles of Wnt pathway genes wls, wnt9a, wnt5b, frzb and gpc4 in regulating convergent-extension during zebrafish palate morphogenesis. Development 143, 2541-2547. 10.1242/dev.137000 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Saal H. M., Prows C. A., Guerreiro I., Donlin M., Knudson L., Sund K. L., Chang C. F., Brugmann S. A. and Stottmann R. W. (2015). A mutation in FRIZZLED2 impairs Wnt signaling and causes autosomal dominant omodysplasia. Hum. Mol. Genet. 24, 3399-3409. 10.1093/hmg/ddv088 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Saito H., Yamamura K. and Suzuki N. (2012). Reduced bone morphogenetic protein receptor type1A signaling in neural-crest-derived cells causes facial dismorphism. Dis. Model Mech. 5, 948-955. 10.1242/dmm.009274 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Saket M., Saliminejad K., Kamali K., Moghadam F. A., Anvar N. E. and Khorram Khorshid H. R. (2016). BMP2 and BMP4 variations and risk of non-syndromic cleft lip and palate. Arch. Oral Biol. 72, 134-137. 10.1016/j.archoralbio.2016.08.019 [DOI] [PubMed] [Google Scholar]
- Salahshourifar I., Halim A. S., Wan Sulaiman W. A. and Zilfalil B. A. (2011). Contribution of MSX1 variants to the risk of non-syndromic cleft lip and palate in a Malay population. J. Hum. Genet. 56, 755-758. 10.1038/jhg.2011.95 [DOI] [PubMed] [Google Scholar]
- Sanford L. P., Ormsby I., Gittenberger-de Groot A. C., Sariola H., Friedman R., Boivin G. P., Cardell E. L. and Doetschman T. (1997). TGFbeta2 knockout mice have multiple developmental defects that are non-overlapping with other TGFbeta knockout phenotypes. Development 124, 2659-2670. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Santosh A. B. R. and Jones T. J. (2014). The epithelial-mesenchymal interactions: insights into physiological and pathological aspects of oral tissues. Oncol. Rev. 8, 239-239 10.4081/oncol.2014.239 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Sasaki Y., Taya Y., Saito K., Fujita K., Aoba T. and Fujiwara T. (2014). Molecular contribution to cleft palate production in cleft lip mice. Congenit Anom (Kyoto) 54, 94-99. 10.1111/cga.12038 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Satokata I. and Maas R. (1994). Msx1 deficient mice exhibit cleft palate and abnormalities of craniofacial and tooth development. Nat. Genet. 6, 348-356. 10.1038/ng0494-348 [DOI] [PubMed] [Google Scholar]
- Scharer C. D., McCabe C. D., Ali-Seyed M., Berger M. F., Bulyk M. L. and Moreno C. S. (2009). Genome-wide promoter analysis of the SOX4 transcriptional network in prostate cancer cells. Cancer Res. 69, 709-717. 10.1158/0008-5472.CAN-08-3415 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Schlessinger K., Hall A. and Tolwinski N. (2009). Wnt signaling pathways meet Rho GTPases. Genes Dev. 23, 265-277. 10.1101/gad.1760809 [DOI] [PubMed] [Google Scholar]
- Schock E. N., Struve J. N., Chang C. F., Williams T. J., Snedeker J., Attia A. C., Stottmann R. W. and Brugmann S. A. (2017). A tissue-specific role for intraflagellar transport genes during craniofacial development. PLoS One 12, e0174206 10.1371/journal.pone.0174206 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Schutte B. C. and Murray J. C. (1999). The many faces and factors of orofacial clefts. Hum. Mol. Genet. 8, 1853-1859. 10.1093/hmg/8.10.1853 [DOI] [PubMed] [Google Scholar]
- Schwabe G. C., Trepczik B., Suring K., Brieske N., Tucker A. S., Sharpe P. T., Minami Y. and Mundlos S. (2004). Ror2 knockout mouse as a model for the developmental pathology of autosomal recessive Robinow syndrome. Dev. Dyn. 229, 400-410. 10.1002/dvdy.10466 [DOI] [PubMed] [Google Scholar]
- Seelan R. S., Mukhopadhyay P., Pisano M. M. and Greene R. M. (2012). Developmental epigenetics of the murine secondary palate. ILAR J. 53, 240-252. 10.1093/ilar.53.3-4.240 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Self M., Lagutin O. V., Bowling B., Hendrix J., Cai Y., Dressler G. R. and Oliver G. (2006). Six2 is required for suppression of nephrogenesis and progenitor renewal in the developing kidney. Embo. J. 25, 5214-5228. 10.1093/ilar.53.3-4.240 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Shao R., Liu J., Yan G., Zhang J., Han Y., Guo J., Xu Z., Yuan Z., Liu J., Malumbres M. et al. (2016). Cdh1 regulates craniofacial development via APC-dependent ubiquitination and activation of Goosecoid. Cell Res. 26, 699-712. 10.1038/cr.2016.51 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Sharp G. C., Ho K., Davies A., Stergiakouli E., Humphries K., McArdle W., Sandy J., Davey Smith G., Lewis S. J. and Relton C. L. (2017). Distinct DNA methylation profiles in subtypes of orofacial cleft. Clin. Epigenetics 9, 63 10.1186/s13148-017-0362-2 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Shkoukani M. A., Lawrence L. A., Liebertz D. J. and Svider P. F. (2014). Cleft palate: a clinical review. Birth Defects Res. C Embryo Today 102, 333-342. 10.1002/bdrc.21083 [DOI] [PubMed] [Google Scholar]
- Slaney S. F., Oldridge M., Hurst J. A., Moriss-Kay G. M., Hall C. M., Poole M. D. and Wilkie A. O. (1996). Differential effects of FGFR2 mutations on syndactyly and cleft palate in Apert syndrome. Am. J. Hum. Genet. 58, 923-932. [PMC free article] [PubMed] [Google Scholar]
- Smith T. M., Wang X., Zhang W., Kulyk W. and Nazarali A. J. (2009). Hoxa2 plays a direct role in murine palate development. Dev. Dyn. 238, 2364-2373. 10.1002/dvdy.22040 [DOI] [PubMed] [Google Scholar]
- Smith T. M., Lozanoff S., Iyyanar P. P. and Nazarali A. J. (2012). Molecular signaling along the anterior–posterior axis of early palate development. Front Physiol. 3, 488 10.3389/fphys.2012.00122 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Song T., Li G., Jing G., Jiao X., Shi J., Zhang B., Wang L., Ye X. and Cao F. (2008). SUMO1 polymorphisms are associated with non-syndromic cleft lip with or without cleft palate. Biochem. Biophys. Res. Commun. 377, 1265-1268. 10.1016/j.bbrc.2008.10.138 [DOI] [PubMed] [Google Scholar]
- Song L., Li Y., Wang K., Wang Y. Z., Molotkov A., Gao L., Zhao T., Yamagami T., Wang Y., Gan Q. et al. (2009). Lrp6-mediated canonical Wnt signaling is required for lip formation and fusion. Development 136, 3161-3171. 10.1242/dev.037440 [DOI] [PubMed] [Google Scholar]
- Song H. Q., Wang X. T., Yan J. Q., Mi N., Jiao X. H., Hao Y. R., Zhang W. and Gao Y. W. (2017). Association of single-nucleotide polymorphisms of CDH1 with nonsyndromic cleft lip with or without cleft palate in a northern Chinese Han population. Medicine 96, e5574 10.1097/MD.0000000000005574 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Stamos J. L. and Weis W. I. (2013) The β-catenin destruction complex. Cold Spring Harbor Perspect. Biol. 5, a007898 10.1101/cshperspect.a007898 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Stanier P. and Pauws E. (2012). Development of the lip and palate: FGF signaling. Front. Oral Biol. 16, 71-80. 10.1159/000337618 [DOI] [PubMed] [Google Scholar]
- Suazo J., Santos J. L., Scapoli L., Jara L. and Blanco R. (2010). Association between TGFB3 and nonsyndromic cleft lip with or without cleft palate in a Chilean population. Cleft Palate Craniofac. J. 47, 513-517. 10.1597/09-015 [DOI] [PubMed] [Google Scholar]
- Sun D., McAlmon K. R., Davies J. A., Bernfield M. and Hay E. D. (1998). Simultaneous loss of expression of syndecan-1 and E-cadherin in the embryonic palate during epithelial-mesenchymal transformation. Int. J. Dev. Biol. 42, 733-736. [PubMed] [Google Scholar]
- Susman M. W., Karuna E. P., Kunz R. C., Gujral T. S., Cantú A. V., Choi S. S., Jong B. Y., Okada K., Scales M. K., Hum J. et al. (2017). Kinesin superfamily protein Kif26b links Wnt5a-Ror signaling to the control of cell and tissue behaviors in vertebrates. Elife 6, e26509 10.7554/eLife.26509 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Suzuki A., Sangani D. R., Ansari A. and Iwata J. (2016). Molecular mechanisms of midfacial developmental defects. Dev. Dyn. 245, 276-293. 10.1002/dvdy.24368 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Sweat Y. Y., Sweat M., Mansaray M., Saadi I., Lachke S., Butali A. and Amendt B. (2018). Six2 regulates palate development by inhibiting palatal bone formation during development. FASEB J. 32 Suppl. 1, 15.11-15.11. [Google Scholar]
- Szabo-Rogers H. L., Smithers L. E., Yakob W. and Liu K. J. (2010). New directions in craniofacial morphogenesis. Dev. Biol. 341, 84-94. 10.1016/j.ydbio.2009.11.021 [DOI] [PubMed] [Google Scholar]
- Szenker-Ravi E., Altunoglu U., Leushacke M., Bosso-Lefevre C., Khatoo M., Thi Tran H., Naert T., Noelanders R., Hajamohideen A., Beneteau C. et al. (2018). RSPO2 inhibition of RNF43 and ZNRF3 governs limb development independently of LGR4/5/6. Nature 557, 564-569. 10.1038/s41586-018-0118-y [DOI] [PubMed] [Google Scholar]
- Tai C. C., Sala F. G., Ford H. R., Wang K. S., Li C., Minoo P., Grikscheit T. C. and Bellusci S. (2009). Wnt5a knock-out mouse as a new model of anorectal malformation. J. Surg. Res. 156, 278-282. 10.1016/j.jss.2009.03.087 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Thomason H. A., Dixon M. J. and Dixon J. (2008). Facial clefting in Tp63 deficient mice results from altered Bmp4, Fgf8 and Shh signaling. Dev. Biol. 321, 273-282. 10.1016/j.ydbio.2008.06.030 [DOI] [PubMed] [Google Scholar]
- Thomason H. A., Zhou H., Kouwenhoven E. N., Dotto G. P., Restivo G., Nguyen B. C., Little H., Dixon M. J., van Bokhoven H. and Dixon J. (2010). Cooperation between the transcription factors p63 and IRF6 is essential to prevent cleft palate in mice. J. Clin. Invest. 120, 1561-1569. 10.1172/JCI40266 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Tian H., Feng J., Li J., Ho T. V., Yuan Y., Liu Y., Brindopke F., Figueiredo J. C., Magee W. III, Sanchez-Lara P. A. et al. (2017). Intraflagellar transport 88 (IFT88) is crucial for craniofacial development in mice and is a candidate gene for human cleft lip and palate. Hum. Mol. Genet. 26, 860-872. 10.1093/hmg/ddx002 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Ting S. B., Caddy J., Hislop N., Wilanowski T., Auden A., Zhao L.-l., Ellis S., Kaur P., Uchida Y., Holleran W. M. et al. (2005). A Homolog of Drosophila grainy head is essential for epidermal integrity in mice. Science 308, 411-413. 10.1126/science.1107511 [DOI] [PubMed] [Google Scholar]
- Tolarova M. M. and Cervenka J. (1998). Classification and birth prevalence of orofacial clefts. Am. J. Med. Genet. 75, 126-137.https://doi.org/<126::AID-AJMG2>3.0.CO;2-R [PubMed] [Google Scholar]
- Topczewski J., Dale R. M. and Sisson B. E. (2011). Planar cell polarity signaling in craniofacial development. Organogenesis 7, 255-259. 10.4161/org.7.4.18797 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Tribulo C., Aybar M. J., Nguyen V. H., Mullins M. C. and Mayor R. (2003). Regulation of Msx genes by a Bmp gradient is essential for neural crest specification. Development 130, 6441-6452. 10.1242/dev.00878 [DOI] [PubMed] [Google Scholar]
- Trokovic N., Trokovic R., Mai P. and Partanen J. (2003). Fgfr1 regulates patterning of the pharyngeal region. Genes Dev. 17, 141-153. 10.1101/gad.250703 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Truong A. B., Kretz M., Ridky T. W., Kimmel R. and Khavari P. A (2006) p63 regulates proliferation and differentiation of developmentally mature keratinocytes. Genes Dev. 20, 3185-3197. 10.1101/gad.1463206 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Tucci V., Kleefstra T., Hardy A., Heise I., Maggi S., Willemsen M. H., Hilton H., Esapa C., Simon M., Buenavista M. T., et al. (2014). Dominant β-catenin mutations cause intellectual disability with recognizable syndromic features. J. Clinic Invest. 124, 1468-1482. 10.1172/JCI70372 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Ulloa F., Itasaki N. and Briscoe J. (2007). Inhibitory Gli3 activity negatively regulates Wnt/β-catenin signaling. Curr. Biol. 17, 545-550. 10.1016/j.cub.2007.01.062 [DOI] [PubMed] [Google Scholar]
- Vacik T., Stubbs J. L. and Lemke G. (2011). A novel mechanism for the transcriptional regulation of Wnt signaling in development. Genes Dev. 25, 1783-1795. 10.1101/gad.17227011 [DOI] [PMC free article] [PubMed] [Google Scholar]
- van Bokhoven H. and Brunner H. G. (2002). Splitting p63. Am. J. Hum. Genet. 71, 1-13. 10.1086/341450 [DOI] [PMC free article] [PubMed] [Google Scholar]
- van Bokhoven H., Celli J., Kayserili H., van Beusekom E., Balci S., Brussel W. and Brunner H. G. (2000). Mutation of the gene encoding the ROR2 tyrosine kinase causes autosomal recessive Robinow syndrome. Nat. Genet. 25, 423-426. 10.1038/78113 [DOI] [PubMed] [Google Scholar]
- Vanderas A. P. (1987). Incidence of cleft lip, cleft palate, and cleft lip and palate among races: a review. Cleft Palate J. 24, 216-225. [PubMed] [Google Scholar]
- Velazquez-Aragón J. A., Alcántara-Ortigoza M. A., Estandia-Ortega B., Reyna-Fabian M. E., Mendez-Adame C. D. and Gonzalez-del Angel A. (2016). Gene interactions provide evidence for signaling pathways involved in cleft lip/palate in humans. J. Dent. Res. 95, 1257-1264. 10.1177/0022034516647034 [DOI] [PubMed] [Google Scholar]
- Vijayan V., Ummer R., Weber R., Silva R. and Letra A. (2018). Association of WNT pathway genes with nonsyndromic cleft lip with or without cleft palate. Cleft Palate Craniofac. J. 55, 335-341. 10.1177/1055665617732782 [DOI] [PubMed] [Google Scholar]
- Vogelaar I. P., Figueiredo J., van Rooij I. A., Simoes-Correia J., van der Post R. S., Melo S., Seruca R., Carels C. E., Ligtenberg M. J. and Hoogerbrugge N. (2013). Identification of germline mutations in the cancer predisposing gene CDH1 in patients with orofacial clefts. Hum. Mol. Genet. 22, 919-926. 10.1093/hmg/dds497 [DOI] [PubMed] [Google Scholar]
- von Gise A., Zhou B., Honor L. B., Ma Q., Petryk A. and Pu W. T. (2011). WT1 regulates epicardial epithelial to mesenchymal transition through beta-catenin and retinoic acid signaling pathways. Dev. Biol. 356, 421-431. 10.1016/j.ydbio.2011.05.668 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Wahl S. E., Kennedy A. E., Wyatt B. H., Moore A. D., Pridgen D. E., Cherry A. M., Mavila C. B. and Dickinson A. J. G. (2015). The role of folate metabolism in orofacial development and clefting. Dev. Biol. 405, 108-122. 10.1016/j.ydbio.2015.07.001 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Wallingford J. B. and Habas R. (2005). The developmental biology of Dishevelled: an enigmatic protein governing cell fate and cell polarity. Development 132, 4421 10.1242/dev.02068 [DOI] [PubMed] [Google Scholar]
- Wan W. D., Yang S. L., Liu J. Y., Cui Y. G., Zhou X. P., Guo F. F., Cheng H. Y., Cheng L., Xiao P. F. and Lu Z. H. (2009). Correlation of the SNPs of FGFR1, FGF10, FGF18 with nonsyndromic cleft lip with or without palate in Chinese population. Beijing Da Xue Xue Bao Yi Xue Ban 41, 409-413. [PubMed] [Google Scholar]
- Wang S. and Samakovlis C. (2012). Grainy head and its target genes in epithelial morphogenesis and wound healing. Curr. Top. Dev. Biol. 98, 35-63. 10.1016/B978-0-12-386499-4.00002-1 [DOI] [PubMed] [Google Scholar]
- Wang B., Fallon J. F. and Beachy P. A. (2000). Hedgehog-regulated processing of Gli3 produces an anterior/posterior repressor gradient in the developing vertebrate limb. Cell 100, 423-434. 10.1016/S0092-8674(00)80678-9 [DOI] [PubMed] [Google Scholar]
- Wang T., Tamakoshi T., Uezato T., Shu F., Kanzaki-Kato N., Fu Y., Koseki H., Yoshida N., Sugiyama T. and Miura N. (2003). Forkhead transcription factor Foxf2 (LUN)-deficient mice exhibit abnormal development of secondary palate. Dev. Biol. 259, 83-94. 10.1016/S0012-1606(03)00176-3 [DOI] [PubMed] [Google Scholar]
- Wang Y., Song L. and Zhou C. J. (2011). The canonical Wnt/beta-catenin signaling pathway regulates Fgf signaling for early facial development. Dev. Biol. 349, 250-260. 10.1016/j.ydbio.2010.11.004 [DOI] [PubMed] [Google Scholar]
- Wang X., Shaffer J. R., Zeng Z., Begum F., Vieira A. R., Noel J., Anjomshoaa I., Cuenco K. T., Lee M. K., Beck J. et al. (2012a). Genome-wide association scan of dental caries in the permanent dentition. BMC Oral Health 12, 57 10.1186/1472-6831-12-57 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Wang H., Hetmanski J. B., Ruczinski I., Liang K. Y., Fallin M. D., Redett R. J., Raymond G. V., Chou Y. H., Chen P. K., Yeow V. et al. (2012b). ROR2 gene is associated with risk of non-syndromic cleft palate in an Asian population. Chin. Med. J. (Engl.) 125, 476-480. [PMC free article] [PubMed] [Google Scholar]
- Wang J., Bai Y., Li H., Greene S. B., Klysik E., Yu W., Scwartz R. J., Williams T. J. and Martin J. F. (2013a). MicroRNA-17-92, a direct Ap-2α transcriptional target, modulatesT-box factor activity in orofacial clefting. PLoS Genet. 9, e1003785 10.1371/journal.pgen.1003785 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Wang C., Chang J. Y., Yang C., Huang Y., Liu J., You P., McKeehan W. L., Wang F. and Li X. (2013b). Type 1 fibroblast growth factor receptor in cranial neural crest cell-derived mesenchyme is required for palatogenesis. J. Biol. Chem. 288, 22174-22183. 10.1074/jbc.M113.463620 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Wang H., Zhang T., Wu T., Hetmanski J. B., Ruczinski I., Schwender H., Liang K. Y., Murray T., Fallin M. D., Redett R. J. et al. (2013)., The FGF and FGFR gene family and risk of cleft lip with or without cleft palate. Cleft Palate Craniofac. J. 50, 96-103. 10.1597/11-132 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Wang Y., Sun Y., Huang Y., Pan Y., Jia Z., Ma L., Ma L., Lan F., Zhou Y., Shi J. et al. (2016). Association study between Van der Woude Syndrome causative gene GRHL3 and nonsyndromic cleft lip with or without cleft palate in a Chinese cohort. Gene 588, 69-73. 10.1016/j.gene.2016.04.045 [DOI] [PubMed] [Google Scholar]
- Wang S., Sun C., Meng Y., Zhang B., Wang X., Su Y., Shi L. and Zhao E. (2017a). A pilot study: screening target miRNAs in tissue of nonsyndromic cleft lip with or without cleft palate. Exp. Ther. Med. 13, 2570-2576. 10.3892/etm.2017.4248 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Wang Y., Sun Y., Huang Y., Pan Y., Shi B., Ma J., Ma L., Lan F., Zhou Y., Shi J. et al. (2017b). The association study of nonsyndromic cleft lip with or without cleft palate identified risk variants of the GLI3 gene in a Chinese population. J. Genet. 96, 687-693. 10.1007/s12041-017-0808-5 [DOI] [PubMed] [Google Scholar]
- Warner D. R., Smith H. S., Webb C. L., Greene R. M. and Pisano M. M. (2009). Expression of Wnts in the developing murine secondary palate. Int. J. Dev. Biol. 53, 1105-1112. 10.1387/ijdb.082578dw [DOI] [PMC free article] [PubMed] [Google Scholar]
- Warner D. R., Smith S. C., Smolenkova I. A., Pisano M. M. and Greene R. M. (2016). Inhibition of p300 histone acetyltransferase activity in palate mesenchyme cells attenuates Wnt signaling via aberrant E-cadherin expression. Exp. Cell Res. 342, 32-38. 10.1016/j.yexcr.2016.02.015 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Watanabe A., Akita S., Tin N. T., Natsume N., Nakano Y., Niikawa N., Uchiyama T. and Yoshiura K. (2006). A mutation in RYK is a genetic factor for nonsyndromic cleft lip and palate. Cleft Palate Craniofac J. 43, 310-316. 10.1597/04-145.1 [DOI] [PubMed] [Google Scholar]
- Welsh I. C., Hagge-Greenberg A. and O'Brien TP (2007). A dosage-dependent role for Spry2 in growth and patterning during palate development. Mech. Dev. 124, 746-761. 10.1016/j.mod.2007.06.007 [DOI] [PMC free article] [PubMed] [Google Scholar]
- White J., Mazzeu J. F., Hoischen A., Jhangiani S. N., Gambin T., Alcino M. C., Penney S., Saraiva J. M., Hove H., Skovby F. et al. (2015). DVL1 frameshift mutations clustering in the penultimate exon cause autosomal-dominant Robinow syndrome. Am. J. Hum. Genet. 96, 612-622. 10.1016/j.ajhg.2015.02.015 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Willems B., Tao S., Yu T., Huysseune A., Witten P. E. and Winkler C. (2015). The Wnt co-receptor Lrp5 is required for cranial neural crest cell migration in zebrafish. PLoS One 10, e0131768 10.1371/journal.pone.0131768 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Wils L. J. and Bijlsma M. F. (2018). Epigenetic regulation of the Hedgehog and Wnt pathways in cancer. Crit. Rev. Oncol. Hematol. 121, 23-44. 10.1016/j.critrevonc.2017.11.013 [DOI] [PubMed] [Google Scholar]
- Wu N., Rollin J., Masse I., Lamartine J. and Gidrol X. (2012). p63 regulates human keratinocyte proliferation via MYC-regulated gene network and differentiation commitment through cell adhesion-related gene network. J. Biol. Chem. 287, 5627-5638. 10.1074/jbc.M111.328120 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Wu W., Gu S., Sun C., He W., Xie X., Li X., Ye W., Qin C., Chen Y., Xiao J. et al. (2015). Altered FGF signaling pathways impair cell proliferation and elevation of palate shelves. PLoS One 10, e0136951.s 10.1371/journal.pone.0136951 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Xiao J., Zhu E.-X., Nagatsuka H., Gunduz M., Li C., Minoo P. and Nagai N. (2005). Wnt5a gene plays a role in mouse embryonic orofacial development. J. Hard Tissue Biol. 14, 355-356. 10.2485/jhtb.14.355 [DOI] [Google Scholar]
- Xiong W., He F., Morikawa Y., Yu X., Zhang Z., Lan Y., Jiang R., Cserjesi P. and Chen Y. (2009). Hand2 is required in the epithelium for palatogenesis in mice. Dev. Biol. 330, 131-141. 10.1016/j.ydbio.2009.03.021 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Xu J., Liu H., Lan Y., Aronow B. J., Kalinichenko V. V. and Jiang R. (2016). A Shh-Foxf-Fgf18-Shh molecular circuit regulating palate development. PLoS Genet. 12, e1005769 10.1371/journal.pgen.1005769 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Xu M., Horrell J., Snitow M., Cui J., Gochnauer H., Syrett C. M., Kallish S., Seykora J. T., Liu F., Gaillard D. et al. (2017). WNT10A mutations causes ectodermal dysplasia by impairing progenitor cell proliferation and KLF4-mediated differentiation. Nat. Commun. 8, 15397 10.1038/ncomms15397 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Yamaguchi T. P., Bradley A., McMahon A. P. and Jones S. (1999). A Wnt5a pathway underlies outgrowth of multiple structures in the vertebrate embryo. Development 126, 1211-1223. [DOI] [PubMed] [Google Scholar]
- Yamada W., Nagao K., Horikoshi K., Fujikura A., Ikeda E., Inagaki Y., Kakitani M., Tomizuka K., Miyazaki H., Suda T. et al. (2009). Craniofacial malformation in R-spondin2 knockout mice. Biochem. Biophys. Res. Commun. 381, 453-458. 10.1016/j.bbrc.2009.02.066 [DOI] [PubMed] [Google Scholar]
- Yan C., Deng-Qi H., Li-Ya C., Mang Y. and Ke-Hu Y. (2018). Transforming growth factor alpha taq I polymorphisms and nonsyndromic cleft lip and/or palate risk: a meta-analysis. Cleft Palate Craniofac. J. 55, 814-820. 10.1597/16-008 [DOI] [PubMed] [Google Scholar]
- Yanagisawa H., Clouthier D. E., Richardson J. A., Charite J. and Olson E. N. (2003). Targeted deletion of a branchial arch-specific enhancer reveals a role of dHAND in craniofacial development. Development 130, 1069-1078. 10.1242/dev.00337 [DOI] [PubMed] [Google Scholar]
- Yanaka Y., Muramatsu T., Uetake H., Kozaki K. and Inazawa J. (2015). miR-544a induces epithelial-mesenchymal transition through the activation of WNT signaling pathway in gastric cancer. Carcinogenesis 36, 1363-1371. 10.1093/carcin/bgv106 [DOI] [PubMed] [Google Scholar]
- Yang L. T. and Kaartinen V. (2007). Tgfb1 expressed in the Tgfb3 locus partially rescues the cleft palate phenotype of Tgfb3 null mutants. Dev. Biol. 312, 384-395. 10.1016/j.ydbio.2007.09.034 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Yang Y., Topol L., Lee H. and Wu J. (2003). Wnt5a and Wnt5b exhibit distinct activities in coordinating chondrocyte proliferation and differentiation. Development 130, 1003-1015. 10.1242/dev.00324 [DOI] [PubMed] [Google Scholar]
- Yang T., Jia Z., Bryant-Pike W., Chandrasekhar A., Murray J. C., Fritzsch B. and Bassuk A. G. (2014). Analysis of PRICKLE1 in human cleft palate and mouse development demonstrates rare and common variants involved in human malformations. Mol. Genet. Genomic. Med. 2, 138-151. 10.1002/mgg3.53 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Yao T., Yang L., Li P. Q., Wu H., Xie H. B., Shen X. and Xie X. D. (2011). Association of Wnt3A gene variants with non-syndromic cleft lip with or without cleft palate in Chinese population. Arch. Oral Biol. 56, 73-78. 10.1016/j.archoralbio.2010.09.002 [DOI] [PubMed] [Google Scholar]
- Yasuhara R., Yuasa T., Williams J. A., Byers S. W., Shah S., Pacifici M., Iwamoto M. and Enomoto-Iwamoto M. (2010). Wnt/beta-catenin and retinoic acid receptor signaling pathways interact to regulate chondrocyte function and matrix turnover. J. Biol. Chem. 285, 317-327. 10.1074/jbc.M109.053926 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Yu L., Gu S., Alappat S., Song Y., Yan M., Zhang X., Zhang G., Jiang Y., Zhang Z., Zhang Y. et al. (2005a). Shox2-deficient mice exhibit a rare type of incomplete clefting of the secondary palate. Development 132, 4397-4406. 10.1242/dev.02013 [DOI] [PubMed] [Google Scholar]
- Yu H. M., Jerchow B., Sheu T. J., Liu B., Costantini F., Puzas J. E., Birchmeier W. and Hsu W. (2005b). The role of Axin2 in calvarial morphogenesis and craniosynostosis. Development 132, 1995-2005. 10.1242/dev.01786 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Yu H., Smallwood P. M., Wang Y., Vidaltamayo R., Reed R. and Nathans J. (2010). Frizzled 1 and frizzled 2 genes function in palate, ventricular septum and neural tube closure: general implications for tissue fusion processes. Development 137, 3707-3717. 10.1242/dev.052001 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Yu H., Ye X., Guo N. and Nathans J. (2012). Frizzled 2 and frizzled 7 function redundantly in convergent extension and closure of the ventricular septum and palate: evidence for a network of interacting genes. Development 139, 4383-4394. 10.1242/dev.083352 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Yu Q., He S., Zeng N., Ma J., Zhang B., Shi B. and Jia Z. (2015). BMP7 gene involved in nonsyndromic orofacial clefts in Western Han Chinese. Med. Oral Patol Oral Cir. Bucal 20, e298-e304. 10.4317/medoral.20335 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Yu K., Deng M., Naluai-Cecchini T., Glass I. A. and Cox T. C. (2017). Differences in oral structure and tissue interactions during mouse vs. human palatogenesis: Implications for the translation of findings from mice. Fronti. Physiol. 8, 154 10.3389/fphys.2017.00154 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Yuan G., Yang G., Zheng Y., Zhu X., Chen Z., Zhang Z. and Chen Y. (2015). The non-canonical BMP and Wnt/β-catenin signaling pathways orchestrate early tooth development. Development 142, 128-139. 10.1242/dev.117887 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Yuan X., Qiu L., Pu Y., Liu C., Zhang X., Wang C., Pu W. and Fu Y. (2016). Histone acetylation is involved in TCDD–induced cleft palate formation in fetal mice. Mol. Med. Rep. 14, 1139-1145. 10.3892/mmr.2016.5348 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Zehir A., Hua L., Maska E., Morikawa Y. and Cserjesi P. (2010). Dicer is required for survival of differentiating neural crest cells. Dev. Biol. 340, 459-467. 10.1016/j.ydbio.2010.01.039 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Zhan T., Rindtorff N. and Boutros M. (2017). Wnt signaling in cancer. Oncogene 36, 1461-1473. 10.1038/onc.2016.304 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Zhang Z., Song Y., Zhao X., Zhang X., Fermin C. and Chen Y. (2002). Rescue of cleft palate in Msx1-deficient mice by transgenic Bmp4 reveals a network of BMP and Shh signaling in the regulation of mammalian palatogenesis. Development 129, 4135-4146. [DOI] [PubMed] [Google Scholar]
- Zhang M., Yan Y., Lim Y., Tang D., Xie R., Chen A., Tai P., Harris S. E., Xing L., Qin Y. X. et al. (2009). BMP-2 modulates β-catenin signaling through stimulation of Lrp5 expression and inhibition of β-TrCP expression in osteoblasts. J. Cell. Biochem. 108, 896-905. 10.1002/jcb.22319 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Zhang R., Oyajobi B. O., Harris S. E., Chen D., Tsao C., Deng H. W. and Zhao M. (2013). Wnt/beta-catenin signaling activates bone morphogenetic protein 2 expression in osteoblasts. Bone 52, 145-156. 10.1016/j.bone.2012.09.029 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Zhao X. and Duester G. (2009). Effect of retinoic acid signaling on Wnt/beta-catenin and FGF signaling during body axis extension. Gene Expr. Patterns 9, 430-435. 10.1016/j.gep.2009.06.003 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Zhou C. J., Wang Y. Z., Yamagami T., Zhao T., Song L. and Wang K. (2010). Generation of Lrp6 conditional gene-targeting mouse line for modeling and dissecting multiple birth defects/congenital anomalies. Dev. Dyn. 239, 318-326. 10.1002/dvdy.22054 [DOI] [PubMed] [Google Scholar]
- Zhu X. J., Liu Y. D., Yuan X. Y., Wang M., Zhao W. X., Yang X. Q., Zhang X. Y., Hsu W., Qiu M. S., Zhang Z. et al. (2016). Ectodermal Wnt controls nasal pit morphogenesis through modulation of the BMP/FGF/JNK signaling axis. Dev. Dyn. 245, 414-426. 10.1002/dvdy.24376 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Zhu X. X., Li Y., Zhou R. M., Wang N. and Kang S. (2018). Knockdown of E-cadherin expression of endometrial epithelial cells may activate Wnt/beta-catenin pathway in vitro. Arch. Gynecol. Obstet. 297, 117-123. 10.1007/s00404-017-4560-0 [DOI] [PubMed] [Google Scholar]
- Zhuang L., Hulin J.-A., Gromova A., Tran Nguyen T. D. T., Yu R. T., Liddle C., Downes M., Evans R. M., Makarenkova H. P. and Meech R. (2014). Barx2 and Pax7 have antagonistic functions in regulation of Wnt signaling and satellite cell differentiation. Stem Cells. 32, 1661-1673. 10.1002/stem.1674 [DOI] [PMC free article] [PubMed] [Google Scholar]