

Abstract
Since past century, vector-borne diseases have been a major public health concern in several states of Mexico. However, Mexico City continues to be free of endemic mosquito-borne viral diseases. The city is the most important politic and economic state of Mexico and one of the most important city of Latin America. Its subtropical highland climate and high elevation (2240 masl) had historically made the occurrence of Aedes species unlikely. However, the presence of other potential disease vectors (Culex spp, Culiseta spp), and the current intermittent introductions of Aedes aegypti, have revealed that control programs must adopt routine vector surveillance in the city. In this study, we provide an updated species list from a five-years of vector surveillance performed in Mexico City. A total of 18,553 mosquito larvae were collected. Twenty-two species from genus Culex, Aedes, Culiseta, Anopheles, Lutzia and Uranotaenia were observed. Nine new mosquito records for the city were found. Ae. albopictus was recorded for the first time in Mexico City. Interestingly, a new record, Ae. epactius was the most frequent species reported. Cx. pipiens quinquefasciatus exhibited the highest number of individuals collected. We detected six areas which harbor the highest mosquito species records in the city. Cemeteries included 68.9% of our collection sites. Temporarily ponds showed the highest species diversity. We detected an increasing presence of Ae. aegypti, which was detected for three consecutive years (2015–2017), predominantly in the warmer microclimates of the city. We found a possible correlation between increasing temperature and Ae. aegypti and Ae. albopictus expanding range. This study provides a starting point for developing strategies related to environmental management for mosquito control. The promotion of mosquito control practices through community participation, mass media and education programmes in schools should be introduced in the city.
Introduction
Between 225–247 species of mosquitoes from 20 genera are known to occur in the 32 states of México [1,2]. However, throughout the country, Aedes aegypti is the main vector of mosquito-borne viral diseases. Although the presence of Aedes albopictus has also been reported in several states [3], its impact as a relevant vector in Mexico is yet unknown. Interestingly, only Mexico City and the state of Tlaxcala remain without the endemic presence of Aedes spp mosquitoes. Thus far, both states continue to be free of endemic mosquito-borne viral diseases.
México City (19°25′57.85″N, 99°07′59.71″W) is the capital of Mexico and is the most important center for all types of financial, cultural and politic activities of the country. Is a destination and point of departure of most international flights, including those which use the city as a stopover from Central and South America, the US and Europe. It is the most urbanized and populous city of the country (8,985,339 people) with a relative small area (1485.5 km2) [4]. The city is located at the Basin of Mexico (Fig 1), in an elevated valley at an altitude of about 2,240 m [5]. The city is surrounded by the Trans-Mexican volcanic belt. It has an average annual temperature range of -2o C to 28o C with humid/subhumid/dry and cold/template climates, associated with seasonal rains in the summer and drier and colder weather in the winter (average annual rainfall: 600 to 1200 mm) (sources: Comisión Nacional del Agua-México and Universidad Nacional Autónoma de México). At the south, the city is bordered by the Ajusco-Chichinautzin mountain range and suburban areas including pine-oak forests and an agricultural lowland. The northern and eastern (drier) regions of the city are also bordered by mountain systems covered by grassland. (http://www.cuentame.inegi.org.mx/monografias/informacion/df/default.aspx?tema=me&e=09). These altitude and climate characteristics of the region have historically made the occurrence of Aedes spp. unlikely. However, a recent survey detected the intermittent presence of Ae. aegpyti larvae in the city [4]. Furthermore, previous surveys [4,6–10] had reported the presence of other competent arbovirus vectors including Culex, Culiseta and Ochlerotatus species, indicating that vector surveillance and control programs should adopt a routine monitoring and surveillance scheme in the city.
Fig 1. Mexico city location.
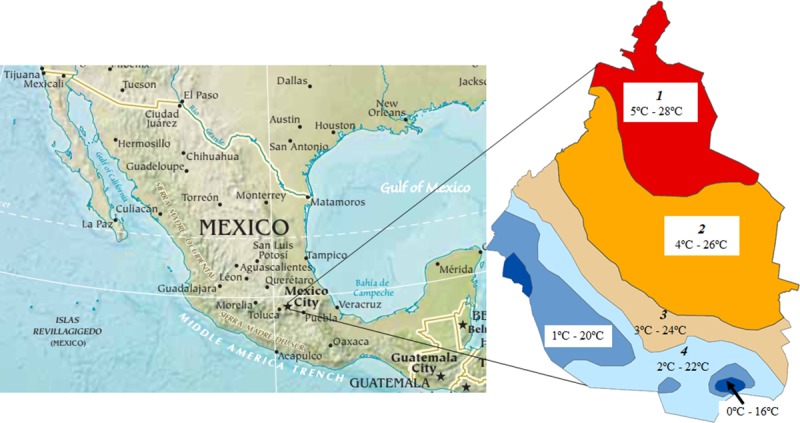
Lowest and highest average annual temperatures in the city are shown. Map modified from https://www.cia.gov/library/publications/the-world-factbook/attachments/docs/original/north_america.pdf?1528326233.
Ae. aegypti is a successful invasive species. Characteristics such as desiccation-resistant eggs [11] and exploitation of urban habitats [12] contribute to the likelihood of Aedes mosquitoes becoming established in non-native areas. The expansion of the range of Ae. aegypti into highland areas around the world might be facilitated by climate change due to rapid and extensive urbanization [13]. The expansion may negatively impact resident species because of interspecific competition (e.g. [14]). Likewise, competition could be altering species composition and may have effects on the vector potential of other medically important species (Aedes and non-Aedes) (e.g. [15]). Information on the diversity of mosquitoes is important to understand the effects of invasive species on native species.
In the present study, we provide an updated species list from five-years of mosquito surveillance performed in México City. Mosquito larvae were collected in urban areas (with high population density, public transportation and minimal green areas [8]) and semi-urban areas (a mix of buildings and green areas with sufficient public transportation but where people still partially maintain agricultural activities [8]). We discuss the potential establishment of Ae. aegypti and Ae. albopictus its impact on both the native mosquito species and on public health. Finally, we show a possible correlation between increasing temperatures in the city and the recent presence of Ae. aegypti and Ae. albopictus.
Methods
Larva collections came from 37 surveys performed across Mexico City over the period of 2012–2016. A total of 163 sites were inspected. Fourth instar larval samples were taken from a variety of habitats (natural and artificial) including: 1) cemeteries, 2) wetlands: non-tidal, semi-permanent wet areas, 3) municipal rain collectors: >25 m2 cement water containers, 4) natural lakes, 5) ponds: permanent bodies of water (natural or artificial) located in public parks or ecological preserves, 6) temporarily pools: seasonal small (< 5m2) bodies of water (natural or artificial) in parks and peri-domestic cemented tanks, 7) streams: natural or human-created shallow tributaries and low flow bodies of water 8) transportation canals: a 100-km network of 10–20 m width and 1–3 m deep canals, commonly used for tourism and food transportation, 8) Olympic canoe canals: a 27 ha and 2m deep artificial canoe sprint and rowing venue.
For larval collections, transfer pipettes and/or 500-ml plastic dippers were used. Collected larvae were placed into bottles containing 96% alcohol. Georeferenced coordinates for each collection site were obtained using a handheld GPS. Collected larvae were then transported to the laboratory. Larvae were counted and identified to species under 50x magnification of a stereo-microscope. Taxonomic identification was carried out using morphological identification keys [16], [17], [18].
The Jaccard similarity coefficient [19] was used to determine similarity in species composition among four temperature gradients where species occurred: microclimate one: 5°C—28°C, microclimate two: 4°C—26°C, microclimate three: 3°C—24°C, microclimate four: 2°C—22° (Figs 1 and 2). Values close to 0 indicate that the microclimates have no species in common and values close to 1 indicate that each species that occurs in one microclimate also occurs in the others.
Fig 2. Collecting sites and hotspots.
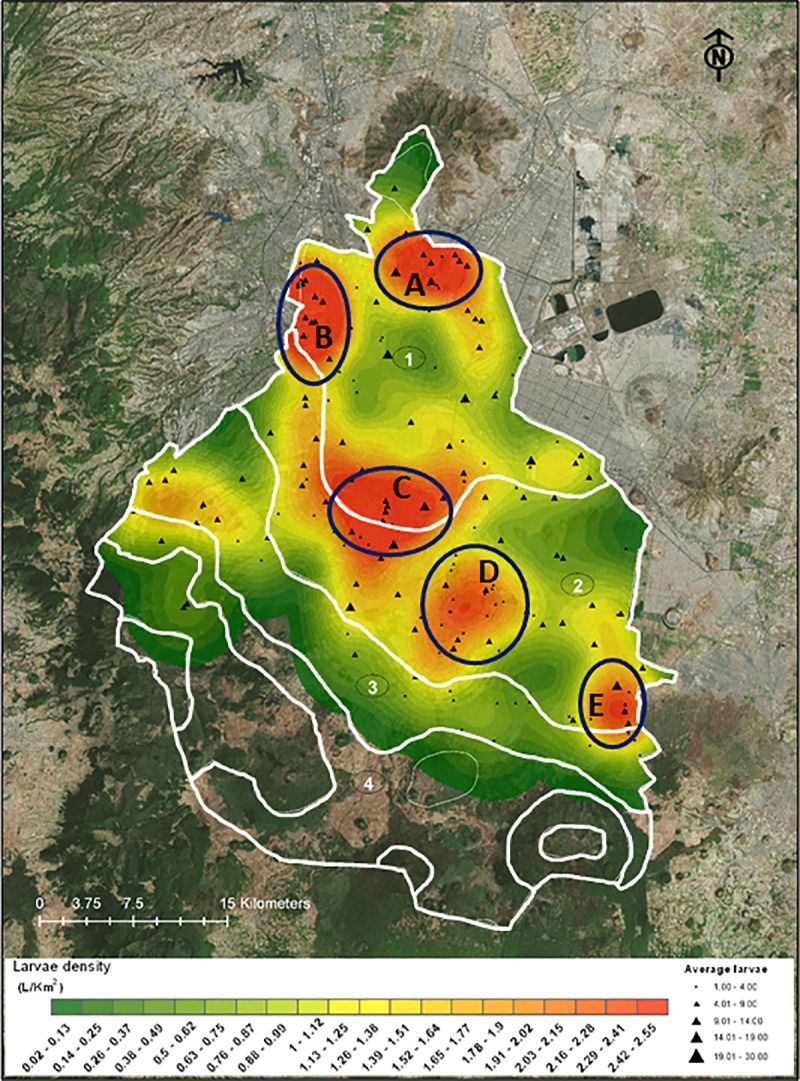
Six cluster of species (clusters A, B, C, D and E) are shown. Numbers indicate microclimates: 1) 5°C—28°C, 2) 4°C—26°C, 3) 3°C—24°C and 4) 2°C—22°C. Larvae density (larvae/km2; green- red areas) and average collected larvae (black triangles) are shown.
For Ae. aegypti and Ae. albopictus, collections (2015–2017) were performed using artificial ovitraps. The ovitraps used were 1-liter dark plastic cups, filled with tap water and lined with a strip of pellon paper along the water margin. In 2015–2016, eggs collected weekly from 2,253 ovitraps were transported and hatched. Fourth instar larvae were identified using a morphological identification key [10]. For the 2017 collection, eggs were hatched, and larvae were reared to adults and identified to species after emergence. Larvae were maintained at 28°C ± 2°C with 70–80% relative humidity and a photoperiod of 12:12 (L: D) h.
As a first step to detect the potential risk of Ae. aegypti introduction and establishment, the possible effects of climate change were assessed. Ae. aegypti distribution was mapped using Mexico City’s climate projection (modified from [20]). This projection reflects the past and current/future urban micro-climate changes arising from urban expansion and other physical characteristics, waste heat release, and regional climate factors. Maps visualizations (Figs 2 and 3, S2.1–S2.6 Fig) were performed using the ArcGIS version 10. The GPS information of every number of larvae of each specie was input. To make density maps (average larvae per collecting site; larvae/km2) the function “Spatial Analyst Tools—Density—Kernel Density” in ArcGIS 10.
Fig 3. Distribution of Aedes aegypti and Aedes albopictus of Mexico City eggs from ovitraps collected in the present study.
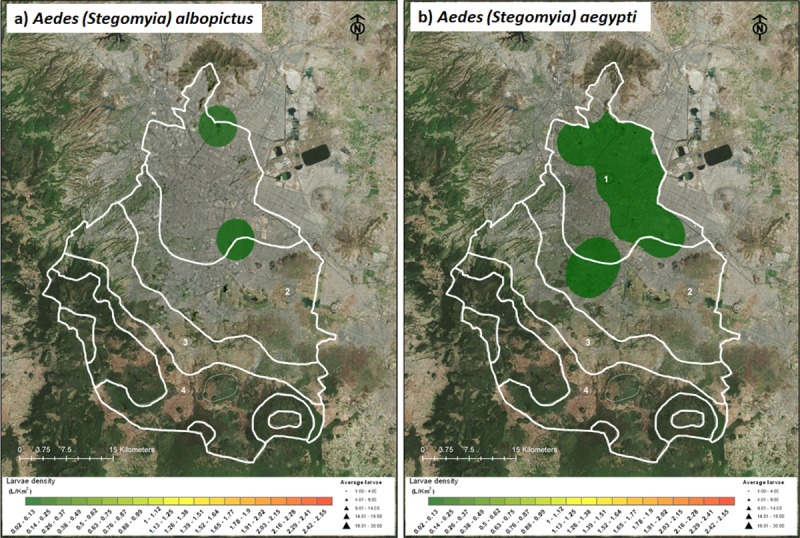
All collected specimens, were hatched, reared, identified and deposited in the Laboratory of Entomology at the Instituto de Diagnóstico y Referencia Epidemiológicos (Institute of Epidemiological Diagnosis and Reference, Mexico City, Mexico) and Unidad de Bioensayo-Centro Regional de Control de Vectores (Bioassay Unit-Regional Center of Vector Control Panchimalco, Morelos, Mexico).
Ethics statement
Note that: no specific permits were required for the described field studies; the location is not privately-owned; the field studies did not involve endangered or protected species.
Data availability statement
Data used in this study can be found at: dx.doi.org/10.6084/m9.figshare.7400018. Field larvae and ovitrap collection guides can be found at http://www.cenaprece.salud.gob.mx/programas/interior/vectores/descargas/pdf/guia_colecta_entomologica_InDRE.pdf and https://www.gob.mx/cms/uploads/attachment/file/37865/guia_vigilancia_entomologica_ovitrampas.pdf
Results
A total of 18,546 mosquito larvae belonging to twenty-two species were collected in urban and semi-urban areas of Mexico City (Fig 2, S1 Fig), including the following genus: Culex (thirteen species), Aedes (one specie;), Culiseta (two species), Anopheles (two species), Lutzia (one species) and Uranotaenia (two species) (Table 1, Fig 3 and S2–S7 Figs). Ae. aegypti and Ae. albopictus were collected as eggs only. Nine new mosquito records for the city were found: Anopheles punctipennis, Aedes albopictus, Aedes epactius, Aedes scapularis, Culex bidens, Culex pinarocampa, Lutzia bigoti, Culex erraticus, and Uranotaenia lowii (Tables 1 and 2).
Table 1. Percentage occurrence (from 163 collecting sites) and percentage of the total number of individuals (out of 18,546 collected larvae) of each mosquito species collected.
| Species | % occurrence | % individuals |
|---|---|---|
| Aedes (Ochlerotatus) epactius* | 90.8 | 27.40 |
| Culex (Culex) stigmatosoma | 78.53 | 16.10 |
| Culiseta (Culiseta) particeps | 67.48 | 14.34 |
| Culex (Culex) pipiens quinquefasciatus | 63.8 | 28.84 |
| Culex (Culex) pinarocampa* | 29.45 | 3.70 |
| Culiseta (Culiseta) inornata | 23.93 | 2.18 |
| Culex (Culex) thriambus | 18.4 | 0.65 |
| Culex (Culex) tarsalis | 16.65 | 1.02 |
| Culex (Culex) salinarius | 15.95 | 1.67 |
| Culex (Neoculex) arizonensis | 9.82 | 1.06 |
| Anopheles (Anopheles) aztecus | 6.75 | 1.27 |
| Culex (Culex) erythrothorax | 4.29 | 0.08 |
| Culex (Culex) bidens* | 3.68 | 0.06 |
| Culex (Neoculex) apicalis | 3.68 | 1.34 |
| Lutzia (Lutzia) bigoti* | 2.45 | 0.03 |
| Culex (Melanoconion) erraticus* | 2.45 | 0.09 |
| Uranotaenia (Uranotaenia) sapphirina | 2.45 | 0.04 |
| Anopheles (Anopheles) punctipennis* | 1.84 | 0.02 |
| Culex (Culex) coronator | 1.84 | 0.10 |
| Aedes (Ochlerotatus) scapularis* | 0.61 | 0.01 |
| Culex (Culex) restuans | 0.61 | 0.01 |
| Uranotaenia (Uranotaenia) lowii* | 0.61 | 0.01 |
*Species marked with asterisks are new records for Mexico City.
Table 2. Updated list of urban and semi urban mosquitoes collected in Mexico city.
| Species | Temperature/climate range in Mexico City | Reported by | Pathogens detected elsewhere (but see †) |
|---|---|---|---|
| Aedes (Stegomyia) albopictus (Skuse 1894)* | 5°C—28°C; subhumid/template/dry | Present study (new record) | WNV, DV, CVV, ZIKV, CHIKV, JEV, EEEV, POTV, USUV, TENV, KEYV, LaCrosse, JCV [21, 22,23] |
| Aedes (Stegomyia) aegypti (Linnaeus 1762)* | 5°C—28°C, 4°C—26°C; subhumid/template/dry | Present study, Kuri- Morales et al. (2017) | DV, YF, ZIKV, CHIKV, RR, WNV, DNV, CPV [21, 24, 25] |
| Aedes (Ochlerotatus) epactius (Dyar & Knab 1908) | 5°C—28°C, 4°C—26°C, 3°C—24°C; subhumid/template/dry | Present study (new record) | WNV [21] |
| Aedes (Ochlerotatus) scapularis (Rondani 1848) | 4°C—26°C; subhumid/template | Present study (new record) | YF, REV, CVV, MV, IV, MyV, VEE [24,26,27] |
| Aedes (Ochlerotatus) trivittatus (Coquillett 1902) | No data | Diaz-Najera & Vargas (1973), Heinemann & Belkin (1977), Ibañez- Bernal & Martinez- Campos (1994) | WNV, TV [21,24] |
| Anopheles (Anopheles) aztecus (Hoffman 1935) | 4°C—26°C; subhumid/template | Present study, Heinemann & Belkin (1977), Ibañez- Bernal & Martinez- Campos (1994) | Plasmodium gonderi [28,29] |
| Anopheles (Anopheles) pseudopunctipennis (Theobald 1901) | No data | Heinemann & Belkin (1977) | Plasmodium spp [24] |
| Anopheles (Anopheles) punctipennis (Say 1823) | 4°C—26°C; subhumid/template | Present study (new record) | Plasmodium spp, WNV [21,24] |
| Culex (Culex) bidens (Dyar 1922) | 5°C—28°C, 4°C—26°C; subhumid/template/dry | Present study (new record) | VEE [30] |
| Culex (Culex) coronator (Dyar and Knab 1906) | 4°C—26°C; subhumid/template | Present study, Ibañez-Bernal & Martinez-Campos (1994) | WNV, ZIKV [21,25] |
| Culex (Culex) erythrothorax (Dyar 1907) | 4°C—26°C; subhumid/template | Present study, Martinez-Palacios (1952), Diaz-Najera & Vargas (1973) | WNV [21] |
| Culex (Culex) peus (Speiser 1904) ⁂ | No data | Diaz-Najera & Vargas (1973), Heinemann & Belkin (1977) | — |
| Culex (Culex) pinarocampa (Dyar & Knab 1908) | 5°C—28°C, 4°C—26°C, 3°C—24°C; subhumid/template/dry | Present study (new record) | — |
| Culex (Culex) pipiens quinquefasciatus (Say 1823) | 5°C—28°C, 4°C—26°C, 3°C—24°C; subhumid/template/dry | Present study, Diaz-Najera & Vargas (1973), Heinemann & Belkin (1977), Ibañez- Bernal & Martinez- Campos (1994), Diaz- Badillo et al. (2011) | WNV, SLE, NPV, CPV, †ZIKV in Mexico City [21,24,25,31] |
| Culex (Culex) pipiens pipiens (Linnaeus 1758) | 4°C—26°C, 3°C—24°C, 2°C—22°C; humid/subhumid/template | Diaz- Badillo et al. (2011) | WNV, SLE, NVP [21,24,31] |
| Culex (Culex) restuans (Theobald 1901) | 5°C—28°C; dry | Present study, Heinemann & Belkin (1977), Ibañez- Bernal & Martinez- Campos (1994) | WNV, NVP [21,24] |
| Culex (Culex) salinarius (Coquillett 1904) | 5°C—28°C, 4°C—26°C; subhumid/template/dry | Present study, Diaz-Najera & Vargas (1973), Ibañez-Bernal & Martinez-Campos (1994) | NVP, DNV [31] |
| Culex (Culex) stigmatosoma (Dyar 1907) | 5°C—28°C, 4°C—26°C, 3°C—24°C; subhumid/template/dry | Present study, Martinez-Palacios (1952), Ibañez-Bernal & Martinez-Campos (1994) | SLE [24] |
| Culex (Culex) tarsalis (Coquillett 1896) | 5°C—28°C, 4°C—26°C, 3°C—24°C; subhumid/template/dry | Present study, Martinez-Palacios (1952), Diaz-Najera & Vargas (1973), Heinemann & Belkin (1977), Ibañez- Bernal & Martinez- Campos (1994), Diaz- Badillo et al. (2011) | WNV, WEE, DNV, SLE, ZIKV [21,24,25,31,32] |
| Culex (Culex) thriambus (Dyar 1921) ⁂ | 5°C—28°C, 4°C—26°C, 3°C—24°C; subhumid/template/dry | Present study, Diaz-Najera & Vargas (1973), Ibañez- Bernal & Martinez- Campos (1994) | WNV, SLE [21,33] |
| Culex (Melanoconion) erraticus (Dyar & Knab 1906) | 4°C—26°C, 3°C—24°C; subhumid/template | Present study (new record) | WNV, VEE [21,34] |
| Culex (Neoculex) apicalis (Adams 1903) | 4°C—26°C; subhumid/template | Present study, Diaz-Najera & Vargas (1973) | WNV [21] |
| Culex (Neoculex) arizonensis (Bohart 1948) | 4°C—26°C, 3°C—24°C, 2°C—22°C; humid/subhumid/template/dry | Present study, Martinez-Palacios (1952), Diaz-Najera & Vargas (1973), Heinemann & Belkin (1977) | — |
| Culiseta (Culiseta) dugesi (Dyar and Knab, 1906)◆ | No data | Heinemann & Belkin (1977) | — |
| Culiseta (Culiseta) inornata (Williston 1893) | 5°C—28°C, 4°C—26°C, 3°C—24°C; subhumid/template/dry | Present study, Diaz-Najera & Vargas (1973), Heinemann & Belkin (1977), Ibañez- Bernal & Martinez- Campos (1994) | La Crosse, MV, WNV [21,24] |
| Culiseta (Culiseta) particeps (Adams 1903) ◆ | 5°C—28°C, 4°C—26°C, 3°C—24°C, 2°C—22°C; humid/subhumid/template/dry | Present study, Diaz-Najera & Vargas (1973), Ibañez-Bernal & Martinez-Campos (1994) | WNV [21] |
| Lutzia (Lutzia) bigoti (Bellardi 1862) | 4°C—26°C, 3°C—24°C; subhumid/template/ | Present study (new record) | — |
| Psorophora (Janthinosoma) cyanescens (Coquillett 1902) | No data | Diaz-Najera & Vargas (1973), Ibañez- Bernal & Martinez- Campos (1994) | Flavivirus, VEE [35,36] |
| Uranotaenia (Uranotaenia) lowii (Theobald 1901) | 4°C—26°C; subhumid/template | Present study (new record) | WNV [37] |
| Uranotaenia (Uranotaenia) sapphirina (Osten Sacken 1868) | 4°C—26°C; subhumid/template | Present study, Diaz-Najera & Vargas (1973), Ibañez-Bernal & Martinez-Campos (1994) | WNV, NPV, CPV [21,31] |
*Ae. aegypti and Ae. albopictus were collected using ovitraps.
⁂The name Cx. peus has been updated to Cx. thriambus.
◆Cs. particeps is the contemporary preferred name of Cs. dugesi.
†Determined at the Instituto de Diagnóstico y Referencia Epidemiológicos (Institute of Epidemiological Diagnosis and Reference, Mexico). Listed pathogens were intentionally limited to arbovirus and Plasmodium spp. CHIKV = Chikungunya virus, CPV = Cytoplasmic Polyhedrosis virus, CVV = Cache Valley virus, DNV = Densovirus, DV = Dengue virus, IV = Ilheus virus, EEEV = Eastern Equine Encephalitis virus, JCV = Jamestown Canyon virus, JEV = Japanese encephalitis virus KEYV = Keystone virus, MV = Melao virus, MyV = Mayaro virus, NPV = Nucleopolyhedrovirus, POTV = Potosi virus, REV = Rocio Encephalitis virus, RR = Ross River virus, SLE = Saint Louis Encephalitis, TENV = Tensaw virus, TV = Trivittatus virus, VEE = Venezuelan Equine Encephalitis virus, WEE = Western Equine Encephalomyelitis virus, USUV = Usuto virus, WNV = West Nile virus, YF = Yellow Fever, ZIKV = Zika virus.
Aedes epactius, Culex pipiens quinquefasciatus, Culex stigmatosoma, Culiseta particeps and Culex pinarocampa were the most frequent species collected in the city (occurring in 63–90% of collecting sites, Table 1). Ae. epactius was the most frequent species collected in the city (particularly in cemeteries) (S2 Fig) (Table 1). Aedes scapularis, Cx. restuans and Ur. lowii were the least frequent and numerous species (Table 1; S2, S4 and S6 Figs). Five species previously recorded in the city were not found during our surveys (see Table 2). Cx. peus can be excluded from previous records since it has been synonymized with Cx. thriambus [38]. Likewise, Cs. particeps is the contemporary preferred synonym of Cs. dugesi [1].
The taxonomical status of Cx pipiens and Cx. p. quinquefaciatus has not resulted in a consensus. Ortega-Morales et al. (2015), stated that true Cx. pipiens occurs in the northern United States and Canada and in Argentina and Uruguay. However, in Mexico City, the presence of Cx. p. quinquefasciatus, Cx. pipiens and hybrids has been previously detected by molecular methods [8]. In the present study specimens were classified as Cx. p. quinquefasciatus.
Fourteen species, including Ae. aegypti and Ae. albopictus, were reported in microclimate one, twenty-two (including Ae. aegypti and Ae. albopictus) in microclimate two, eleven in microclimate three and two in microclimate four (Table 2, Figs 2 and 3, S2–S7 Figs). Cx. restuans was only found in microclimate one (S4 Fig). Eight species were only found in microclimate two: Ae. scapularis, An. aztecus, An. Punctipennis (S2 Fig), Cx. coronator, Cx. Erythrothorax (S3 Fig), Cx. Apicalis (S5 Fig) and Ur. sapphirina (S7 Fig), while Cs. particeps was the only species distributed across the four microclimates (S6 Fig). The Jaccard similarity coefficients were: 0.31 between microclimate one and two; 0.33 between one and three; 0.31 between two and three; 0.15 between three and four; 0.08 between two and four; 0.06 between one and four.
We detected six clusters which harbored the greatest diversity of species in the city (Fig 2). The first cluster was located northwest of the city (spot A, Fig 2). This area corresponds to a set of cemeteries (Cementerio Español, San Isidro, Monte Sinai, Americano and Aleman) and a 77.4 ha park (Parque Ex Refineria 18 de Marzo Centenario). In 1991, this former refinery became a public park with artificial ponds and temporarily pools. The second cluster corresponds to the Basilica of Our Lady of Guadalupe and the foothills of “El Tepeyac” Hill National Park, on the north side of the city (spot B, Fig 2). The third cluster (spot C, Fig 2), corresponds to an urban developments called “Ciudad Jardin/Churubusco/Coyaacan” area (Spot D, Fig 2), which consists of buildings, parks and houses surrounded by small green areas. The fourth cluster and more extensive cluster (mid-south of the city, spot D, Fig 2), corresponds to the canals of the Ecological Park of “Xochimilco” (215 ha), along with several artificial agricultural plots called “chinampas”. This is also a tourist area with water-traffic corridors, wetlands, natural lakes, streams and a canoe canal. The area also contains a mix of urban and semi-urban settlements. Four species were only found in this area (An. aztecus, An. punctipennis, Ur. lowii and Ur. sapphirina) (S2, S6 and S7 Figs). Cluster six was located in the southeast (spot E, Fig 2), and corresponds to the “Milpa Alta” borough. It is a relatively new semi-urban area with surface water restricted to small springs, streams and several city rain collectors. It is part of the Chichinautzin biological corridor (which also includes the States of Mexico and Morelos).
Cemeteries included 68.9% of our collecting sites, and a great number of species were recorded in this habitat (Table 3). However, cemeteries did not present the highest species diversity; temporary ponds showed the highest diversity (15 species). Cemeteries and water corridors harbored 14 species each and 10 species each were recorded from city rain collectors and wetlands. Ponds, natural lakes and streams contained 8–9 species each. Canoe canals were the least preferred habitat (3 species recorded) (Table 3). Cs. inornata and Cx. stigmatosoma were the only species present in all habitats (Table 3, S4 and S6 Figs).
Table 3. Type of sampling habitat and number of records for each specie.
| Habitat | Specie | Records |
|---|---|---|
| Canoe canal | Cs. (Cs.) inornata | 1 |
| Cx. (Cx.) salinarius | 1 | |
| Cx. (Cx.) stigmatosoma | 1 | |
| Cemetery | Ae. (Och.) epactius | 170 |
| Cx. (Cx.) pipiens quinquefasciatus | 125 | |
| Cx. (Cx.) stigmatosoma | 121 | |
| Cs. (Cs.) particeps | 108 | |
| Cx. (Cx.) pinarocampa | 50 | |
| Cx. (Cx.) thriambus | 35 | |
| Cs. (Cs.) inornata | 16 | |
| Cx. (Cx.) tarsalis | 11 | |
| Cx. (Cx.) arizonensis | 9 | |
| Cx. (Cx.) bidens | 7 | |
| Cx. (Cx.) salinarius | 6 | |
| Lut. (Lut.) bigoti | 2 | |
| Cx. (Cx.) restuans | 1 | |
| Cx. (Cx.) erythrothorax | 1 | |
| City Rain Collector | Cs. (Cs.) particeps | 9 |
| Cx. (Cx.) stigmatosoma | 9 | |
| Cx. (Cx.) pipiens quinquefasciatus | 4 | |
| Cs. (Cs.) inornata | 3 | |
| Cx. (Cx.) tarsalis | 3 | |
| Cx. (Cx.) erraticus | 2 | |
| Cx. (Cx.) salinarius | 2 | |
| Cx. (Cx.) arizonensis | 1 | |
| Cx. (Cx.) pinarocampa | 1 | |
| Lut. (Lut.) bigoti | 1 | |
| Natural lake | Cs. (Cs.) inornata | 6 |
| Cx. (Cx.) stigmatosoma | 5 | |
| Cx. (Cx.) tarsalis | 5 | |
| Cx. (Cx.) salinarius | 4 | |
| Cx. (Cx.) pipiens quinquefasciatus | 2 | |
| Ur. (Ur.) sapphirina | 2 | |
| An. (An.) aztecus | 1 | |
| Cs. (Cs.) particeps | 1 | |
| Pond | Ae. (Och.) epactius | 3 |
| Cx. (Nx.) apicalis | 3 | |
| Cs. (Cs.) particeps | 2 | |
| Cx. (Cx.) arizonensis | 2 | |
| Cs. (Cs.) inornata | 1 | |
| Cx. (Cx.) coronstor | 1 | |
| Cx. (Cx.) erraticus | 1 | |
| Cx. (Cx.) pipiens quinquefasciatus | 1 | |
| Cx. (Cx.) stigmatosoma | 1 | |
| Stream | Cx. (Cx.) arizonensis | 6 |
| An. (An.) aztecus | 5 | |
| Cs. (Cs.) particeps | 4 | |
| An. (An.) punctipennis | 1 | |
| Cs. (Cs.) inornata | 1 | |
| Cx. (Cx.) erythrothorax | 1 | |
| Cx. (Cx.) salinarius | 1 | |
| Cx. (Cx.) stigmatosoma | 1 | |
| Temporarily pond | Ae. (Och.) epactius | 50 |
| Cs. (Cs.) particeps | 12 | |
| Cs. (Cs.) inornata | 11 | |
| Cx. (Cx.) stigmatosoma | 10 | |
| Cx. (Cx.) pipiens quinquefasciatus | 8 | |
| Cx. (Cx.) tarsalis | 7 | |
| Cx. (Cx.) salinarius | 4 | |
| Cx. (Cx.) thriambus | 4 | |
| An. (An.) aztecus | 2 | |
| Cx. (Cx.) coronstor | 2 | |
| Cx. (Cx.) pinarocampa | 2 | |
| Ae. (Och.) scapularis | 1 | |
| Cx. (Cx.) arizonensis | 1 | |
| Cx. (Cx.) erythrothorax | 1 | |
| Cx. (Nx.) apicalis | 1 | |
| Water traffic corridor | An. (An.) aztecus | 11 |
| Cs. (Cs.) inornata | 8 | |
| Cx. (Cx.) salinarius | 8 | |
| Cx. (Cx.) stigmatosoma | 8 | |
| Cx. (Cx.) tarsalis | 7 | |
| Cx. (Cx.) pipiens quinquefasciatus | 4 | |
| Cx. (Cx.) erythrothorax | 3 | |
| Ae. (Och.) epactius | 3 | |
| Cs. (Cs.) particeps | 2 | |
| Cx. (Nx.) apicalis | 2 | |
| Ur. (Ur.) sapphirina | 2 | |
| An. (An.) punctipennis | 1 | |
| Cx. (Cx.) erraticus | 1 | |
| Ur. (Ur.) lowii | 1 | |
| Lut. (Lut.) bigoti | 1 | |
| Wetland | An. (An.) aztecus | 5 |
| Cx. (Cx.) salinarius | 3 | |
| Cs. (Cs.) inornata | 2 | |
| Cs. (Cs.) particeps | 2 | |
| Cx. (Cx.) stigmatosoma | 2 | |
| Cx. (Cx.) tarsalis | 2 | |
| Cx. (Nx.) apicalis | 2 | |
| An. (An.) punctipennis | 1 | |
| Cx. (Cx.) erythrothorax | 1 | |
| Ur. (Ur.) sapphirina | 1 |
Cs. inornata and Cx. stigmatosoma (in bold) were recorded in all habitats.
The presence of Ae. aegypti was detected over three consecutive years (2015–2017) using ovitraps (Table 4, Fig 3). Occurrence was detected predominantly in the warmer microclimate (5°C—28°C: in the north of the city) (Tables 2 and 4). Although each year has seen increases in the number of positive ovitraps, at the sites have not been consistent, with the exception of “Tapo” Bus Terminal (19o 25’ 44” N, 99o 06’46” W), where the occurrence of Ae. aegypti was recorded for two consecutive years (2016–2017) (Table 4). The average number of eggs laid in each ovitrap was 21.33, where the percentage of Ae. aegypti eggs hatched was 8 (37%).
Table 4. Location of Aedes aegypti and Aedes albopictus eggs collected by ovitraps in Mexico city.
| Year | Positive ovitrap site | Description | Min-max climate range in Mexico City | Latitude (N) | Longitude (W) |
|---|---|---|---|---|---|
| Aedes aegypti | |||||
| 2015 | Casa del Peregrino | Pilgrim house | 5°C—28°C | 19o 28’ 57” | 99o 06’ 37” |
| Estación Pantaco | Train station | 5°C—28°C | 19o 28’ 59” | 99o 10’ 09” | |
| 2016 | CONALEP Aeropuerto | Voc-Tech high school | 5°C—28°C | 19 o 25’ 26” | 99o 03’ 26” |
| TAPO | Bus terminal | 5°C—28°C | 19o 25’ 44” | 99o 06’ 46” | |
| Alameda Oriente | Public park | 5°C—28°C | 19o 26’ 08” | 99o 03’ 08” | |
| 2017 | Parque Santa Úrsula | Public park | 4°C—26°C | 19o 18’ 12” | 99o 09’ 26” |
| Museo Diego Rivera Anahuacalli | Museum | 4°C—26°C | 19o 19’ 22” | 99o 08’ 38” | |
| Alberca Salvador Allende | Community swimming pool | 5°C—28°C | 19o 21’ 29” | 99o 03’ 02” | |
| Parque del Pueblo Cuitláhuac | Farm park & zoo | 5°C—28°C | 19o 21’ 41” | 99o 02’ 38” | |
| Central de Abastos | Food supply center | 5°C—28°C | 19o 22’ 45” | 99o 05’ 35” | |
| Deportivo Venustiano Carranza | Community sport center | 5°C—28°C | 19o 25’ 35” | 99o 07’ 06” | |
| TAPO | Bus terminal | 5°C—28°C | 19o 25’ 44” | 99o 06’ 46” | |
| Gimnasio “Smart Fit” Molina | Gym parking lot | 5°C—28°C | 19o 29’ 51” | 99o 05’ 23” | |
| Parque Nacional El Tepeyac | National Park | 5°C—28°C | 19o 30’ 14” | 99o 06’ 28” | |
| Panteón Ticomán | Cemetery | 5°C—28°C | 19o 30’ 18” | 99o 07’ 16” | |
| Deportivo Atlético Mexicano | Community sport center | 5°C—28°C | 19o 22’ 29” | 99o 03’ 46” | |
| Zoológico de San Juan de Aragón | Zoo park | 5°C—28°C | 19o 27’ 43” | 99o 05’ 02” | |
| Jardín Francisco J. Múgica | Public park | 5°C—28°C | 19o 20’ 57” | 99o 03’ 41” | |
| Aedes albopictus | |||||
| 2017 | Cerro de la Estrella | National Archeological Park | 5°C—28°C | 19o 21’ 06” | 99o 05’ 30” |
| Parque Nacional El Tepeyac | National Park | 5°C—28°C | 19o 29’ 58” | 99o 06’ 23” | |
For the first time, Ae. albopictus was detected in the city. In 2017, Ae. albopictus were detected in two sites in the warm northern region of Mexico City in national parks (“El Tepeyac” and “Cerro de la Estrella”) (Table 4, Fig 3). At the “El Tepeyac” park, 125 eggs were collected, but only two hatched larvae were identified as Ae. albopictus. At “Cerro de la Estrella” park, 27 eggs were collected, but only one larva was recognized as Ae. albopictus.
When overlaying Ae. aegypti and Ae. albopictus distribution on the temperature projection, collection sites and predicted warmer temperatures areas were highly correlated (Fig 4). These zones might provide climatic and habitat suitability which could promote the long-term establishment of the species.
Fig 4. Possible correlation between the projected increase in temperatures (modified from [20]) and the current presence of Ae. aegypti (yellow dots) and Ae. albopictus (green dots) in the city.

Discussion
The updated list now includes a total of 26 mosquito species (excluding Cx. peus and Cs. Dugesi, because synonymy) in Mexico City. Nine new records, and the intermittent but increased presence of Ae. aegypti, were documented. The present surveillance effort was the first to sample mosquitoes over a large extension of the city. Since previous collections may not have surveyed all the water bodies as in our present study, the newly recorded species may have been historically present.
New records include: Ae. scapularis, a common species in Mexico, mainly with neotropical distribution; An. punctipennis and Cx. erraticus, nearctic common species with some neotropical distribution; Cx. bidens, a species with nearctic and neotropical distribution; Lut. bigoti, mainly a neotropical species, distributed in the Balsas Basin and South Pacific Zone; Ur. lowii, a neotropical common species with some nearctic distribution [7,10].
A particularly interesting new record was Ae. epactius. Its presence, but mostly its great abundance and distribution, was surprising given that there were no previous records for this species in the city. Although Diaz-Badillo et al. [8] reported the presence of Ochlerotatus spp, identification was not made at the species level. Ae. epactius is a common species in the southern United States and Mexico [10,17] at low, mid and high elevations [39]. Lozano-Fuentes et al. [39] had speculated that a possible reason for increased distribution at higher elevations could be climate change. The lack of previous records of this species in the city may be explained by a possible event of recent colonization. The presence of Cx. pinarocampa was another interesting finding. Previous records only reported this species from five states of Mexico (Oaxaca, Chiapas, Veracruz, Guerrero and Estado de Mexico) [7]. This species occurred in 29% of our collecting sites, however, the number of collected individuals was low.
Microclimates and clusters
Microclimates one-two, 1–3 and 2–3 shared several species (Jaccard index = 0.31, 0.33 and 0.31 respectively). It could be argued that the continued increase in temperature can explain this pattern. High urban temperatures may have been promoting the expansion of species ranges (see Jauregui 1997), with, some species now adapted to a wide range of temperatures. For the case of Cs. particeps (distributed in the four microclimates), the increasing temperatures could be changing their distribution in the city.
Microclimate two (4°C—26°C), showed 22 species, eight were exclusively found here and four of these were confined to cluster five (spot D “Xochimilco”). The “Xochimilco” area is set of canals and wetlands, both remnants of the extinct large saline Lake of Xochimilco [40]. In 1987, the area was included on UNESCO’s World Heritage List [41]. However, previous high deforestation rates, rapid ongoing illegal urbanization and overexploitation of groundwater have caused a significant reduction of the wetland area [42]. For the four species that were only found here, a process of intense local adaptation could had caused their restricted distribution.
Habitat loss may have also promoted the isolation of populations. For example, Cx. erraticus Ae. scapularis, Cx. apicalis, Cx coronator and Cx erythrothorax were only present in ecological preserves (Pedregal: 19o19’06” N, 99o11’04” W and Xochimilco) and/or well-preserved fragmented areas (Milpa Alta, cluster E). The same could be occurring for some species found in clusters four and six. Current trends in urbanization may threaten the persistence of native mosquitos in the city.
Clusters “A, B and C” were completely immersed in urban areas and showed high larval productivity. In this area there is an assemblage of untreated habitats, useful for both possible vectors and/or nuisance mosquitoes. Clusters D and E included ecological or national parks, surrounded by residential or industrial areas. The bordering urban sites may provide natural and artificial containers that could be used as larval habitats. They could also be transition areas between sylvatic and truly urban zones.
Habitats
The abundance of larvae appeared to be most influenced by habitat type. For Aedes and Culex spp. [43,44], cemeteries have long been recognized as major breeding sites with suitable conditions for larval development (abundant containers, vegetation cover and landscape connectivity). For these reasons, they have been under constant monitoring. As expected, cemeteries showed the greatest productivity and diversity of mosquitoes (14 species, 10 belonging to the Culex genus). Ae. epactius, Cx. p. quinquefasciatus and Cx. stigmatosoma, were the most frequent observed species. Cemeteries should remain as priority sites for the monitoring of mosquitoes.
Mexico City is a heterogeneous mosaic of residential and commercial areas, parks, and other land-use types. This also provides an array of temporary pools that are being frequently used by mosquitoes. Although temporary pools could also be in the same areas as cemeteries, the edaphic conditions and assemblage of predators and prey may be completely different. This could help explain the differences related to species assemblage between cemeteries and temporary pools.
Rain collectors were located in the Milpa Alta borough, in microclimate three. Meanwhile, two streams were situated in microclimate four and one stream in the Xochimilco area. Milpa Alta and microclimate four were the less urbanized and colder zones. These areas could provide harborage and dispersal routes for sylvatic/native species.
Wetlands, water corridors, natural lakes and a pond are part of the remnants of the Xochimilco and Chalco extinct saline lakes (located in the southwestern part of the urban area of the city). These water bodies have alkaline pH and a high content of organic matter [45,46]. Interestingly, regardless of its size and freshwater capacity (470, 625 m3; [47]), canoe canals harbored only three species: Cx. salinarius, Cs. inornata and Cx. stigmatosoma. Pollution could be an important consideration in the canal. Algae and cyanobacteria toxicity blooms are constant [47,48], limiting habitat suitability for some species. The salinity, pollution and pH of these habitats could explain the presence of few species and the low abundance of species like Ae. epactius and Cx. p. quinquefasciatus. In our study, Cs. inornata and Cx. stigmatosoma were the only species capable of exploiting all available habits. It is unknown if larvae of both species are adapting to polluted sites or to man-made environments.
Aedes aegypti and Aedes albopictus presence: Risks and challenges
Our study created a starting point for future efforts aimed to addressing Ae. aegypti and Ae. albopictus distribution across the city. The presence of Ae. aegypti has significantly increased. Unsurprisingly, positive ovitrap collections were recorded in the warmer areas, in the north and northeast of the city. The first records of Ae. aegypti in 2015 occurred in a train station and a pilgrim house. Both sites are places that experience massive local and non-local human transit. We presume that an accidental introduction of eggs, larvae, pupae, and/or adults into the region occurred by human transit [4]. However, the 2016–2017 collections were recorded at public parks, museums, residential areas and cemeteries, indicating the possible presence of transitory colonies. To date, it remains unclear if true colonization has occurred since no larvae or adults have been collected.
Ae. albopictus was only collected at the warmer area of the city. The National park “El Tepeyac” is next to the Basilica of Guadalupe, one of the most important pilgrimage sites of Catholicism in the Americas which is annually visited by millions of people from different states and countries. The National Park “Cerro de la Estrella”, and its surroundings, are the venues for the Iztapalapa Catholic Passion Play (a representation of the crucifixion of Jesus). Annually, almost 2 million persons (local and non-local) gather to observe the play [49]. Given the high level of transit into the parks from areas with endemic Ae. albopictus, independent introductions of adults or immature stages are likely to occur. However, it also remains unclear if colonization has occurred.
Temperature and habitat availability are two important factors affecting the presence of mosquitoes. Climate change might be a causative factor for introduction of Ae. aegypti [13], especially in urban areas, given that water and air temperature in urban areas are higher than in suburban ones [50]. Reports of Ae. aegypti being in areas either with elevated altitudes and/or cold temperatures show that migration and colonization in colder areas seems to be becoming more common [51–53]. Deforestation, poor housing and insufficient sewer and waste management systems are consequences of uncontrolled urbanization. All these factors could be leading to the increase of suitable habitats for Aedes spp. However, not all water bodies in the city are necessarily at risk; for example, the Xochimilco area includes turbid, saline or polluted breeding sites where Aedes larvae are not likely to occur.
Historically, the altitude and geographical location of Mexico City could have limited Ae. albopictus and Ae. aegypti presence. Increasing levels of urbanization put Mexico City and neighboring areas at risk of becoming areas where vector mosquitoes (and the diseases they carry) could become established. Because of their close association with humans, surveillance for larval and adult Ae. aegypti and Ae. albopictus should occur near human dwellings, schools and residential/commercial areas (F. Castelo, pers.comm). Cemeteries should also be priority sites for monitoring and mosquito control [54].
Cemeteries seem to be the most suitable habitats for Ae. aegypti and Ae. albopictus to colonize. However, we have detected the presence of multiple other species in these habitats. Competition among larvae is an important factor regulating mosquito populations [55]. Aedes spp are known to alter competitive interactions, declining the presence of other species [14]. Nevertheless, it is also possible that native species could influence the population growth of invasive larvae, imposing barriers to a successful invasion. The potential impact of species competition among native species and Ae. aegypti should be evaluated.
Competition and climate change may also impact adult mosquito susceptibility to arboviral infections [56]. Since vector borne diseases can also be influenced by climate, extreme weather may impact the presence of several infectious diseases. It is possible that warmer temperatures could trigger the introduction of pathogens. It has been proposed that climate change will contribute to an extensive increase in the number of people at risk of dengue fever [57], although recent findings suggest a low potential for ZIKV transmission at high elevations [58]. Twenty-three species reported here have medical importance. Consequently, a constant surveillance of arboviral diseases and vectors must be a priority in the city
Conclusions
Our findings provide a starting point to create a suitable plan for mosquito control in Mexico City. A deeper understanding of the spatio-temporal dynamics of breeding sites and microecological habitat characteristics in Mexico City is required. Identifying associations between biological diversity and habitats may us enable to predict how populations will respond to habitat reduction, species competition and climate change.
Currently, Ae. aegypti and Ae. albopictus do not seem to be established in Mexico City, however, they were included in the updated list because of their rapid and successful colonizing abilities. The presence of the main vector of dengue, chikungunya and Zika shows that Mexico City should no longer be considered exempt from the occurrence of vector-borne disease outbreaks. Thus, the practice of arbovirus control through community participation, mass media and education programmes in schools should be introduced.
Supporting information
All sites were peri-domestic areas, including: cemeteries, public (free access) parks, museums or community centers.
(TIF)
(TIF)
(TIF)
(TIF)
(TIF)
(TIF)
(TIF)
Acknowledgments
We would like to thank the control personnel from the Ministry of Health of Mexico City for their assistance with fieldwork. To the Entomology Laboratory staff at the InDRE and the staff of the Unidad de Bioensayos-Centro Regional de Control de Vectores Panchimalco, Jojutla, Morelos. To Fabio Castello for useful comments and discussions. To Priscila Bascuñan-Garcia for invaluable comments. We are extremely grateful to Audrey Lenhart for her comments and proofreading this paper for the use of English.
Data Availability
All data are available from the figshare database at: dx.doi.org/10.6084/m9.figshare.7400018.
Funding Statement
The authors received no specific funding for this work.
References
- 1.Darsie RF. A Survey and bibliography of the mosquito fauna of Mexico (Diptera:Culicidae). J Am Mosq Control Assoc. 1996;12: 298–306. [PubMed] [Google Scholar]
- 2.Ibáñez-Bernal S., Strickman D, Martínez-Campos C. Culicidae (Diptera) In: Llorente-Bousquets J, García AN, González-Soriano E, editors. Biodiversidad, taxonomía y biogeografía de artrópodos de México: hacia una síntesis de su conocimiento Vol 1 Mexico City: Universidad Nacional Autónoma de México; 1996. pp. 591–602. [Google Scholar]
- 3.Pech-May A, Moo-Llanes DA, Puerto-Avila MB, Casas M, Danis-Lozano R, Ponce G, et al. Population genetics and ecological niche of invasive Aedes albopictus in Mexico. Acta Trop. 2016;157: 30–41. 10.1016/j.actatropica.2016.01.021 [DOI] [PubMed] [Google Scholar]
- 4.Kuri-Morales P, Correa-Morales F, González-Acosta C, Sánchez-Tejeda G, Dávalos-Becerril E, Fernanda Juárez-Franco M, et al. First report of Stegomyia aegypti (= Aedes aegypti) in Mexico City, Mexico. J Med Vet Entomol. 2017;31: 240–242. [DOI] [PubMed] [Google Scholar]
- 5.United Nations. The World’s Cities in 2016. Data Booklet; Nairobi; 2016. [Google Scholar]
- 6.Díaz-Nájera A, Vargas L. Mosquitos mexicanos distribución geográfica actualizada. Rev Invest Salud Publica. 1973; 33: 111–125. [PubMed] [Google Scholar]
- 7.Martinez-Palacios A. Notas sobre la distribucion de los mosquitos Culex en Mexico (Dipetra: Culicidae). Rev Soc Mex Hist Nat. 1952;13: 75–87. [Google Scholar]
- 8.Diaz-Badillo A, Bolling BG, Perez-Ramirez G, Moore CG, Martinez-Muñoz JP, Padilla-Viveros AA, et al. The distribution of potential West Nile virus vectors, Culex pipiens pipiens and Culex pipiens quinquefasciatus (Diptera: Culicidae), in Mexico City. Parasit Vectors. 2011; 4: 70 10.1186/1756-3305-4-70 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 9.Heinemann S, Belkin J. Collection records of the project “Mosquitoes of Middle America” 9. Mexico (MEX, MF, MT, MX). Mosq Syst. 1977; 9: 483–535. [Google Scholar]
- 10.Ibanez-Bernal S, Martinez-Campos C. Clave para la identificación de larvas de mosquitos comunes en las áreas urbanas y suburbanas de la República Mexicana (Diptera: Culicidae). Folia Entomológica Mex. 1994; 92: 43–73. [Google Scholar]
- 11.Clements AN. The Biology of Mosquitoes. Vol 1: development, nutrition and reproduction. London, UK: CABI Publishing; 1992. [Google Scholar]
- 12.Cheong WH. Preferred Aedes aegypti larval habitats in urban areas. Bull World Health Organ. 1967;36: 586–589. [PMC free article] [PubMed] [Google Scholar]
- 13.Jansen CC, Beebe NW. The dengue vector Aedes aegypti: what comes next. Microbes Infect. 2010;12: 272–279. 10.1016/j.micinf.2009.12.011 [DOI] [PubMed] [Google Scholar]
- 14.Barrera R. Competition and resistance to starvation in larvae of container-inhabiting Aedes mosquitoes. Ecol Entomol. 1996; 21: 117–127. [Google Scholar]
- 15.Bevins SN. Invasive mosquitoes, larval competition, and indirect effects on the vector competence of native mosquito species (Diptera: Culicidae). Biol Invasions. 2008;10: 1109–1117. [Google Scholar]
- 16.Carpenter SJ, LaCasse WJ. Mosquitoes of North America (North of Mexico) Berkeley, CA: University of California Press; 1955. [Google Scholar]
- 17.Darsie RF, Ward RA. Identifaction and Geographical Distribution of the Mosquitoes of North America, North of Mexico. 2nd ed. Gainesville, FL: University Press of Florida; 2005. [Google Scholar]
- 18.Ibáñez-Bernal S, Martínez-Campos C. Aedes albopictus in Mexico. J Am Mosq Control Assoc. 1994;10: 231–232. [PubMed] [Google Scholar]
- 19.Brower JE, Zar JH. Field and Laboratory Methods for General Ecology. Dubuque, IA: Wm. C. Brown Company Publishers; 1984. [Google Scholar]
- 20.Magaña-Rueda V, Méndez-Pérez J, Galván-Ortiz L, López-Bravo L, Méndez-Antonio B. Estudio de vulnerabilidad diferenciada ante eventos extremos del clima en la Zona Metropolitana de la Ciudad de México: aspectos climáticos In: Leon C, Neri C, editors. Informe final Estudio: pobreza urbana y cambio climático para la Ciudad de México. Gobierno del Distrito Federal; 2010. pp. 72–114. Available: http://www.museodehistorianatural.df.gob.mx/sedema/images/archivos/temas-ambientales/cambio-climatico/informe-pobreza-cambio-climatico-ciudad-de-mexico-informe-ejecutivo.pdf [Google Scholar]
- 21.CDC. Mosquito species in which West Nile virus has been detected, United States, 1999–2012 [Internet]. Atlanta, GA; 2012. Available: https://www.cdc.gov/westnile/resources/pdfs/mosquito species 1999-2012.pdf
- 22.Paupy C, Delatte H, Bagny L, Corbel V, Fontenille D. Aedes albopictus, an arbovirus vector: From the darkness to the light. Microbes Infect. 2009; 11: 1177–1185. 10.1016/j.micinf.2009.05.005 [DOI] [PubMed] [Google Scholar]
- 23.Puggioli A, Bonilauri P, Calzolari M, Lelli D, Carrieri M, Urbanelli S, et al. Does Aedes albopictus (Diptera: Culicidae) play any role in Usutu virus transmission in Northern Italy? Experimental oral infection and field evidences. Acta Trop. 2017; 172: 192–196. 10.1016/j.actatropica.2017.05.006 [DOI] [PubMed] [Google Scholar]
- 24.Foster WA, Walker ED. Mosquitoes (Culicidae) In: Mullen G, Durden L, editors. Medical and Veterinary Entomology. San Diego, CA: Academic Press; 2002. pp. 203–262. [Google Scholar]
- 25.Elizondo-Quiroga D, Medina-Sánchez A, Sánchez-González JM, Eckert KA, Villalobos-Sánchez E, Navarro-Zúñiga AR, et al. Zika virus in salivary glands of five different species of wild-caught mosquitoes from Mexico. Sci Rep. 2018;8: 809 10.1038/s41598-017-18682-3 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 26.Downs WG, Spence L, Aitken TH, Whitman LE. Cache Valley virus, isolated from a Trinidadian mosquito, Aedes scapularis. West Indian Med J. 1961;10: 13–15. [PubMed] [Google Scholar]
- 27.Forattini O. Culicidologia Médica Identificação, Biologia, Epidemiologia. Vol. 2 São Paulo: Universidade de São Paulo; 2002. [Google Scholar]
- 28.Perez-Reyes R. Anopheles aztecus (Hoffman, 1938) a new definitive host for the cyclical transmission of Plasmodium berghei Vinoke and Lips, 1948. J Parasitol. 1953;39: 603–604. [PubMed] [Google Scholar]
- 29.Downs W, Bordas E. Hallazgo de Anopheles aztecus Hoffmann naturalmente infectado con Plasmodium en la zona de Xochimilco D. F. Rev del Inst Salubr y Enfermedades Trop Mex. 1949;10: 321–325. [Google Scholar]
- 30.Sabattini MS, Avilés G, Monath T. Historical, epidemiological and ecological aspects of arboviruses in Argentina: Flaviviridae, Bunyaviridae and Rhabdoviridae In: Travassos da Rosa APA, Vasconcelos PFC, Travassos da Rosa JFS, editors. An Overview of Arbovirology in Brazil and Neighboring Countries. Belem, Brasil: Instituto Evandro Chagas; 1998. pp. 113–134. [Google Scholar]
- 31.Shapiro AM, Becnel JJ, White SE. A nucleopolyhedrovirus from Uranotaenia sapphirina (Diptera: Culicidae). J Invertebr Pathol. 2004; 86: 96–103. 10.1016/j.jip.2004.04.005 [DOI] [PubMed] [Google Scholar]
- 32.Hammon WMCD Reeves WC, Brookman B, Izumi EM, Gjullin CM. Isolation of the viruses of Western Equine and St. Louis Encephalitis from Culex tarsalis mosquitoes. Science. 1941;94: 328–330. 10.1126/science.94.2440.328 [DOI] [PubMed] [Google Scholar]
- 33.Reeves WC, Hardy JL, Scrivani RP, Presser SB, Rosen L. Experimental transovarial transmission of St. Louis Encephalitis Virus by Culex and Aedes mosquitoes. Am J Trop Med Hyg. 1984;33: 166–175. [DOI] [PubMed] [Google Scholar]
- 34.Chamberlain RW, Sudia WD, Work TH, Coleman PH, Newhouse VF, Johnston J. G. JR. Arbovirus studies in South Florida, with emphasis on Venezuelan Equine Encephalomyelitis Virus1. Am J Epidemiol. 1969;89: 197–210. [DOI] [PubMed] [Google Scholar]
- 35.Farfan-Ale JA, Loroño-Pino MA, Garcia-Rejon JE, Soto V, Lin M, Staley M, et al. Detection of Flaviviruses and Orthobunyaviruses in mosquitoes in the Yucatan Peninsula of Mexico in 2008. Vector Borne Zoonotic Dis. 2010;10: 777–783. 10.1089/vbz.2009.0196 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 36.Sudia WD, Newhouse VF, Beadle ID, Miller DL, Johnston JGJ, Young R, et al. Epidemic Venezuelan equine encephalitis in North America in 1971: vector studies. Am J Epidemiol. 1975; 101: 17–35. [DOI] [PubMed] [Google Scholar]
- 37.Cruz Cruz M, Gonzalez Escalona A. Culícidos transmisores del virus del Nilo Occidental en el municipio de Jesús Menéndez. Rev Electrónica Dr Zoilo E Mar Vidaurreta. 2015; 40: 1–6. [Google Scholar]
- 38.Strickman D. Redescription of the holotype of Culex (Culex) peus Speiser and taxonomy of Culex (Culex) stigmatosoma Dyar and thriambus Dyar (Diptera: Culicidae). Proc Entomol Soc Washingt. 1988; 90: 484–494. [Google Scholar]
- 39.Lozano-Fuentes S, Welsh-Rodriguez C, Hayden MH, Tapia-Santos B, Ochoa-Martinez C, Kobylinski KC, et al. Aedes (Ochlerotatus) epactius along an elevation and climate gradient in Veracruz and Puebla States, México. J Med Entomol. Entomological Society of America; 2012;49: 1244–1253. [DOI] [PubMed] [Google Scholar]
- 40.Alcocer-Durand J, Escobar-Briones EG. The aquatic biota of the now extinct lacustrine complex of the Mexico basin. Freshw Forum. 1992; 2: 171–183. [Google Scholar]
- 41.ICOMOS (International Council on Monuments and Sites). World Heritage List Advisory Board Evaluation No. 412. Paris: UNESCO; 1987. [Google Scholar]
- 42.Narchi NE, Cristiani BC. Subtle tyranny: divergent constructions of nature and the erosion of traditional ecological knowledge in Xochimilco. Lat Am Perspect. 2015; 42: 90–108. [Google Scholar]
- 43.Vezzani D. Review: Artificial container-breeding mosquitoes and cemeteries: a perfect match. Trop Med Int Heal. 2007;12: 299–313. [DOI] [PubMed] [Google Scholar]
- 44.Vezzani D, Albicócco AP. The effect of shade on the container index and pupal productivity of the mosquitoes Aedes aegypti and Culex pipiens breeding in artificial containers. Med Vet Entomol. 2009;23: 78–84. 10.1111/j.1365-2915.2008.00783.x [DOI] [PubMed] [Google Scholar]
- 45.Gayosso-Morales MA, Nandini S, Martinez-Jernonimo FF, Sarma SSS. Effect of organic and inorganic turbidity on the zooplankton community structure of a shallow waterbody in Central Mexico (Lake Xochimilco, Mexico). J Environ Biol. 2017; 38: 1183–1196. [Google Scholar]
- 46.Ramos Bello R, Cajuste LJ, Flores Román D, García Calderón NE. Heavy metals, salts and sodium in chinampa soils in México. Agrociencia. 2001; 35: 385–395. [Google Scholar]
- 47.Pereyra B. Cuemanco: cuando entrenar da asco. Proceso; 2017: 2110, April 9. Available: http://www.proceso.com.mx/482034/cuemanco-cuando-entrenar-da-asco [Google Scholar]
- 48.Pineda-Mendoza RM, Olvera-Ramírez R, Martínez-Jerónimo F. Microcystins produced by filamentous cyanobacteria in urban lakes. A case study in Mexico City. Hidrobiológica. 2012; 22: 290–298. [Google Scholar]
- 49.Zepeda-Lopez HM, Perea-Araujo L, Miliar-García A, Dominguez-López A, Xoconostle-Cázarez B, et al. Inside the outbreak of the 2009 influenza A (H1N1)v virus in Mexico. PLoS One. 2010; 8: e13256. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 50.Li Y, Kamara F, Zhou G, Puthiyakunnon S, Li C, Liu Y, et al. Urbanization increases Aedes albopictus larval habitats and accelerates mosquito development and survivorship. PLoS Negl Trop Dis. 2014; 8: e3301 10.1371/journal.pntd.0003301 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 51.Ruiz-López F, González-Mazo A, Vélez-Mira A, Gómez GF, Zuleta L, Vélez-Bernal ID. Presencia de Aedes (Stegomyia) aegypti (Linnaeus, 1762) y su infección natural con el virus del dengue en alturas no registradas para Colombia. Biomédica. 2016;36: 303–308. 10.7705/biomedica.v36i2.3301 [DOI] [PubMed] [Google Scholar]
- 52.Zanotti G, De Majo MS, Alem I, Schweigmann N, Campos RE, Fischer S. New records of Aedes aegypti at the southern limit of its distribution in Buenos Aires province, Argentina. J Vector Ecol. 2015;40: 408–411. 10.1111/jvec.12181 [DOI] [PubMed] [Google Scholar]
- 53.De Majo MS, Montini P, Fischer S. Egg hatching and survival of immature stages of Aedes aegypti (Diptera: Culicidae) under natural temperature conditions during the cold season in Buenos Aires, Argentina. J Med Entomol. 2016; 54: 106–113. 10.1093/jme/tjw131 [DOI] [PubMed] [Google Scholar]
- 54.WHO. Prevention and Control of Dengue and Dengue Haemorrhagic Fever—Comprehensive Guidelines. World Health Organization, South-East Asia Regional Office, New Delhi; 1999. [Google Scholar]
- 55.Noden BH, O’Neal PA, Fader JE, Juliano SA. Impact of inter- and intra-specific competition among larvae on larval, adult, and life-table traits of Aedes aegypti and Aedes albopictus females. Ecol Entomol. 2016;41: 192–200. 10.1111/een.12290 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 56.Alto BW, Lounibos LP, Higgs S, Juliano S a. Larval competition differentially affects arbovirus infection in Aedes mosquitoes. Ecology. 2005;86: 3279–3288. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 57.Hales S, de Wet N, Maindonald J, Woodward A. Potential effect of population and climate changes on global distribution of dengue fever: an empirical model. Lancet. 2002;360: 830–834. 10.1016/S0140-6736(02)09964-6 [DOI] [PubMed] [Google Scholar]
- 58.Watts AG, Miniota J, Joseph HA, Brady OJ, Kraemer MUG, Grills AW, et al. Elevation as a proxy for mosquito-borne Zika virus transmission in the Americas. PLoS One. 2017;12: e0178211 10.1371/journal.pone.0178211 [DOI] [PMC free article] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.
Supplementary Materials
All sites were peri-domestic areas, including: cemeteries, public (free access) parks, museums or community centers.
(TIF)
(TIF)
(TIF)
(TIF)
(TIF)
(TIF)
(TIF)
Data Availability Statement
All data are available from the figshare database at: dx.doi.org/10.6084/m9.figshare.7400018.
Data used in this study can be found at: dx.doi.org/10.6084/m9.figshare.7400018. Field larvae and ovitrap collection guides can be found at http://www.cenaprece.salud.gob.mx/programas/interior/vectores/descargas/pdf/guia_colecta_entomologica_InDRE.pdf and https://www.gob.mx/cms/uploads/attachment/file/37865/guia_vigilancia_entomologica_ovitrampas.pdf