

Abstract
Functional traits can covary to form “functional syndromes.” Describing and understanding functional syndromes is an important prerequisite for predicting the effects of organisms on ecosystem functioning. At the intraspecific level, functional syndromes have recently been described, but very little is known about their variability among populations and—if they vary—what the ecological and evolutionary drivers of this variation are. Here, we quantified and compared the variability in four functional traits (body mass, metabolic rate, excretion rate, and boldness), their covariations and the subsequent syndromes among thirteen populations of a common freshwater fish (the European minnow, Phoxinus phoxinus). We then tested whether functional traits and their covariations, as well as the subsequent syndromes, were underpinned by the phylogenetic relatedness among populations (historical effects) or the local environment (i.e., temperature and predation pressure), and whether adaptive (selection or plasticity) or nonadaptive (genetic drift) processes sustained among‐population variability. We found substantial among‐population variability in functional traits and trait covariations, and in the emerging syndromes. We further found that adaptive mechanisms (plasticity and/or selection) related to water temperature and predation pressure modulated the covariation between body mass and metabolic rate. Other trait covariations were more likely driven by genetic drift, suggesting that nonadaptive processes can also lead to substantial differences in trait covariations among populations. Overall, we concluded that functional syndromes are population‐specific, and that both adaptive and nonadaptive processes are shaping functional traits. Given the pivotal role of functional traits, differences in functional syndromes within species provide interesting perspectives regarding the role of intraspecific diversity for ecosystem functioning.
Keywords: adaptation, freshwater fish, genetic drift, intraspecific variation, rivers, syndrome, trait covariation
1. INTRODUCTION
Phenotypic variability measured within species has historically been the core of evolutionary studies, as it constitutes the visible outcome of evolutionary processes (Darwin, 1859; Roff, 1992; Stearns, 1992). It is now increasingly acknowledged that intraspecific phenotypic variability can strongly affect community structure and ecosystem functioning (Des Roches et al., 2018; Raffard, Santoul, Cucherousset, & Blanchet, 2018). In particular, functional traits, such as excretion rate, are extremely important for understanding and predicting how organisms affect their own biotic and abiotic environment (Díaz et al., 2013; Violle et al., 2007). Functional traits display variability both within and among populations (Helsen et al., 2017; Villéger, Brosse, Mouchet, Mouillot, & Vanni, 2017). For instance, the nutrient excretion rate (a trait potentially affecting nutrient availability in ecosystems, Vanni, 2002) can vary substantially among and within populations (Evangelista, Lecerf, Britton, & Cucherousset, 2017; Villéger, Grenouillet, Suc, & Brosse, 2012). Since functional traits determine the way organisms modulate the environment, it is important to investigate the spatial distribution of these traits (Funk et al., 2016; Villéger et al., 2017).
Functional traits are highly variable across landscapes. For instance, the metabolic rate of ectotherms is, on average, higher in warm than in cold environments (Brown, Gillooly, Allen, Savage, & West, 2004; Hildrew, Raffaelli, & Edmonds‐Browns, 2007). Moreover, trait covariations are also expected to be heterogeneous across landscapes (Reale et al., 2010). The covariations among multiple traits have been referred to as syndromes(Dingemanse et al., 2007). Syndromes have primarily been investigated for life‐history, behavioral, and physiological traits (Roff, 1992; Sih, Bell, & Johnson, 2004), and have greatly contributed to our understanding of life‐history strategies in wild populations (Reale et al., 2010). In the meantime, community ecologists have investigated how covariations in functional traits, measured at the community level, can affect ecosystem functioning (Díaz et al., 2016; Lavorel & Garnier, 2002). More recently, it has been demonstrated that functional trait covariations also occur within species, forming a so‐called functional syndrome (Raffard et al., 2017). Functional syndromes have been shown to exist in several species (e.g., Defossez, Pellissier, & Rasmann, 2018; Raffard et al., 2017), but the variability of these syndromes across populations and environmental conditions remains unexplored.
Functional syndromes are also expected to vary among populations within a single species (Peiman & Robinson, 2017). Indeed, it has been suggested that environmental conditions can modulate trait covariations and the associated syndromes (Killen, Marras, Metcalfe, McKenzie, & Domenici, 2013). Notably, experimental studies have demonstrated that some environmental conditions can induce biological constraints (e.g., energetic requirement) that modulate trait covariations (Finstad, Forseth, Ugedal, & NæSje, 2007; Killen, Marras, & McKenzie, 2011). For instance, food availability has been demonstrated to produce a covariation between metabolic rate and risk‐taking behavior in European sea bass (Dicentrarchus labrax), probably because individuals with high metabolic rate have high energetic demands and should be more active to acquire resources to sustain this demand (Killen et al., 2011). Variation in syndromes has also been reported among wild populations living in heterogeneous environments (Dingemanse et al., 2007; Peiman & Robinson, 2012; Pruitt et al., 2010; Závorka et al., 2017). Beyond the direct effect of environmental characteristics (e.g., temperature, predation) on syndromes, the evolutionary history of populations—such as the past demographic and colonization history that often generates bottlenecks and founder effects—may also play an underestimated role in shaping syndromes (Armbruster & Schwaegerle, 1996; Peiman & Robinson, 2017). For instance, populations can exhibit different syndromes because they may have been colonized by two independent lineages having evolved divergent syndromes in their past respective refuge (“the ghost of colonization past”). This past evolutionary legacy is likely to be identified at the level of the genetic lineages; two closely related populations being more likely to display similar syndromes than two distantly related populations. This possible evolutionary legacy of syndromes has—up to our knowledge—rarely been considered.
In this study, we investigated the variability of functional traits and the syndromes they form in wild populations inhabiting heterogeneous environments. Using a common freshwater fish (the European minnow, Phoxinus phoxinus) as a model species, we aimed at testing (a) whether functional traits and their covariations vary between populations, and (b) whether this variability is explained by environmental factors and/or the evolutionary history of populations. Focusing on four functional traits (i.e., excretion rate, metabolism, body mass, and boldness), we first expected that both mean values and covariations of traits differ between populations because of their contrasting environments and evolutionary histories. Second, we focused on two environmental characteristics (temperature and predation intensity) that affect functional traits (e.g., metabolism, Gillooly, 2001), and that are hence likely to also modulate their covariations. Temperature is indeed a key abiotic factor for ectotherms as it can affect their metabolic rate, behavior, and body mass (Biro, Beckmann, & Stamps, 2010; Brown et al., 2004; Gillooly, 2001). Additionally, predation risk can affect the physiology and behavior of individuals by inducing strong stresses (Bell & Sih, 2007; Hawlena & Schmitz, 2010). We concomitantly tested the contribution of the past evolutionary history of populations to explain variation in covariations among functional traits using phylogenetic models. Specifically, we assessed the relationships between genetic similarity (inferred from microsatellite markers) and syndrome similarity among populations. An influence of the environment on traits would suggest potential adaptation (or plasticity of these syndromes), and we hence finally used a quantitative genetic approach (P ST/F ST, Leinonen, McCairns, O'Hara, & Merilä, 2013) to infer the evolutionary processes (genetic drift vs. selection/plasticity) underlying differences in trait variation and covariation among populations.
2. MATERIALS AND METHODS
2.1. Model species
The European minnow (P. phoxinus) is an abundant species in Western Europe in cold lakes (e.g., mountains lakes) and rivers (e.g., from small rivers at intermediate altitude to mountain streams) with summer water temperature generally lower than 22–24°C (Keith, Persat, Feunteun, & Allardi, 2011). It is a small‐bodied fish species (<12 cm long, 5–8 cm long as an adult in general) living approximately 3 to 5 years, and which displays a generalist diet composed of small invertebrates, algae, or zooplankton (Collin & Fumagalli, 2011; Frost, 1943). The European minnow is considered as a genotypically and phenotypically variable species (Collin & Fumagalli, 2011, 2015; Fourtune et al., 2018).
2.2. Sampling sites and animal rearing
We focused on riverine European minnow populations from the Dordogne–Garonne river basin in southwestern France (Figure 1). We selected thirteen sites (coded from A to M) in different rivers to reflect their potential colonization history (Fourtune, Paz‐Vinas, Loot, Prunier, & Blanchet, 2016; Paz‐Vinas et al., 2018). Sampled rivers were selected based on previous knowledge in terms of environmental and geographic characteristics of the area (Fourtune et al., 2016, 2018).
Figure 1.
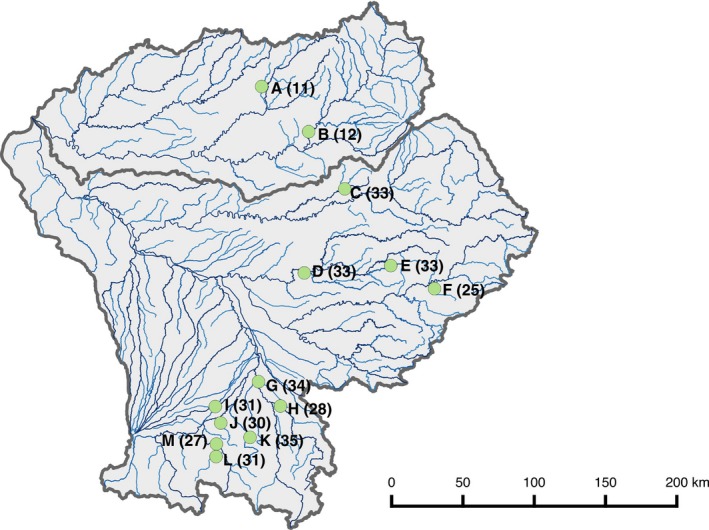
Distribution of the thirteen studied populations of European minnows (Phoxinus phoxinus). Names of populations were coded from A to M, and the number of individuals for each population is given as indication
For each site, we focused and measured two environmental variables that have been shown to modulate functional traits in ectotherms (Bestion, Teyssier, Aubret, Clobert, & Cote, 2014; Biro et al., 2010; Gillooly, 2001), and hence potentially their covariations. We first recorded water temperature, which was measured as the mean temperature between July and September 2017, using automatic sensors (HOBO®, one measure every hour). Mean summer water temperature varied from 15.5°C (site E) to 21.5°C (site D) (Figure 1). In addition, we measured the local predation pressure, a key biotic factor that can affect organisms’ phenotype (Langerhans, 2007). Predation pressure was calculated for each site as the density of piscivorous fishes (namely northern pike, Esox lucius; brown trout, Salmo trutta; rainbow trout, Oncorhynchus mykiss; European perch, Perca fluviatilis;pikeperch, Sander lucioperca; and European eel, Anguilla anguilla). This metric was similar to that described in Edeline, Lacroix, Delire, Poulet, and Legendre (2013). This index of predation was calculated by dividing the number of sampled predator individuals by the surface covered during sampling; these data—for each site—were sourced from Fourtune et al. (2016) and from the French Agency for Biodiversity (Poulet, Beaulaton, & Dembski, 2011).
In summer 2016, we collected adult fish on these thirteen sites using electrofishing (Figure 1). On each river, we collected approximately one hundred adults along a ~200‐m‐long river stretch to ensure representativeness of the fish habitat. Then, we randomly sampled 30–40 individuals among the sampled adults to have a representative subsample of each population. Electrofishing was performed under the authorization of “Arrêté Préfectoraux” delivered by the “Direction Départementale des Territoires” of each administrative department (Haute‐Garonne, Ariège, Aveyron, Lot, Tarn and Corrèze). Laboratory rearing of fish was performed under authorizations of the “Direction Départementale de la Cohésion Sociale et de la Protection des Populations (service Santé Protection des Animaux et Environnement) de l'Ariège,” Arrêté Préfectoraux SA‐013‐PB‐092 and Certificat de Capacité 09‐273. Fish were brought to the laboratory and maintained in a thermoregulated room for two to four weeks before experiments. Fish from the different populations were held in independent 150‐L tanks in which water temperature was set to 17°C and photoperiod to a light:dark cycle of 12:12 (Golovanov, 2013). They were fed with frozen bloodworms three times a week. Prior to experiments, fish were anesthetized (benzocaine, 25 mg/L), weighed (to the nearest 0.01 g), and tagged with a Passive Integrated Transponder (PIT) tags (8 × 1.4 mm, FDX‐B “skinny” PIT tag, Oregon RFID, USA) inserted in the general cavity using a sterile scalpel. Fish recovered and acclimatized to the rearing room for 10 days before the quantification of three functional traits in addition to body mass (boldness, excretion rate, and metabolic rate). Metabolic rate was measured on day 1 (morning), while excretion rate and boldness were measured on day 2 in the morning and in the afternoon, respectively. Before quantifying metabolic rate, individuals were starved for two days to ensure the same starvation level among individuals.
2.3. Boldness
Boldness was assessed for each individual independently in circular containers (30 cm in diameter) filled with 5L of dechlorinated tap water at 17°C and 500 ml of water from a tank containing conspecifics. The containers were surrounded by curtains to standardize light conditions and to hide the experimenter. A shelter (pipe, 7 cm length × 3 cm diameter) was added in each container to allow the fish to hide. After having introduced each individual into the shelter and after 10 min of acclimatization to reduce stress level induced by handling, the shelter was opened and each individual was filmed for fifteen minutes. Video footage was subsequently analyzed with the software “BORIS” (Friard & Gamba, 2016). Boldness was quantified as the time spent outside of the shelter. The order and the containers in which individuals were assayed were randomly attributed. All behavioral assays were performed in the afternoon (from 12:00 p.m. to 16:00 p.m.) to minimize the potential effects of circadian rhythms.
2.4. Excretion rate
Excretion rate was quantified using nitrogen excreted by organisms as the dissolved form of ammonium . Quantifying excretion rate on starved individuals was done to avoid an effect of differential consumption, which is a strong factor affecting the rate of nitrogen excretion (Glaholt & Vanni, 2005). Changes in concentration in water can affect ecosystem functioning through an increase in nutrient availability (Capps & Flecker, 2013) and primary production (Bassar et al., 2016; Schmitz, Hawlena, & Trussell, 2010). Following Villéger et al. (2012), individuals were placed in plastic bags containing 500 ml of spring bottled water for 1 hr at 17°C. Individuals were then removed and 100 ml of water was filtered through a glass microfiber filter (Whatman, GF/C, diameter = 25 mm), and samples were frozen at −20°C. Excretion rate ( in μg L−1 hr−1) was determined with a high‐performance ionic chromatograph (Dionex DX‐120).
2.5. Metabolic rate
We measured the oxygen consumption rate as a proxy of the metabolic rate of individuals. Fish were individually placed in a custom made metabolic chamber filled with 500 ml of dechlorinated tap water and hermetically sealed. Chambers were set in a thermoregulated room at 17°C in the dark to lower the stress level. We measured the metabolic rate just after handling so that the same stress was imposed to all individuals. Measurements of oxygen concentration were taken after 10 min, allowing individuals to acclimate, and continuously every five seconds for 50 min using oxygen probes (OXROB10, Pyroscience). The metabolic rate was calculated as the absolute slope between oxygen quantity in the chamber and time, reflecting the hourly consumption of oxygen (mg/hr).
2.6. Genetic analyses
Thirty additional adults from each of the thirteen sites were sampled for genetic material. For each of these thirty individuals, we collected and preserved in 70% ethanol a small piece of pelvic fin and individuals were then released in their respective sampling site. Genomic DNA was extracted using a salt‐extraction protocol (Aljanabi, 1997). Eighteen autosomal microsatellite markers were considered in this study: Polymerase chain reactions (PCR) and genotyping were performed as detailed in Supporting Information Appendix S1, resulting in a final data set of 357 genotypes. We checked for multilocus deviation from Hardy–Weinberg equilibrium (HWE) and for gametic disequilibrium using GENEPOP 4.2.1 (Rousset, 2008) after sequential Bonferroni correction to account for multiple related tests (Rice, 1989). The presence of null alleles was then assessed at each locus by analyzing homozygote excess in five populations that did not follow HWE, using MICROCHECKER 2.2.3 (Van Oosterhout, Hutchinson, Wills, & Shipley, 2004). We discarded from further analyses any locus showing significant gametic disequilibrium and/or evidence of null alleles, resulting in the withdrawal of one locus (CtoG‐075), for a total number of seventeen loci.
We computed Nei's standard genetic distance (Nei, 1973) between each pair of populations using the diveRsity R‐package (function diffCalc; Keenan, McGinnity, Cross, Crozier, & Prodöhl, 2013). A hierarchical cluster analysis was then performed to uncover genetic relatedness among the thirteen populations using the functions hclust (R‐package stats) and as.phylo (R‐package ape; Paradis, Claude, & Strimmer, 2004) to convert the genetic dissimilarity matrix into an unrooted phylogenetic tree based on complete linkage method.
Finally, we estimated the overall level of genetic differentiation F ST among the thirteen populations using the hierfstatR‐package (Goudet, 2005). The resulting global F ST corresponds to the inter‐population variance component in allelic frequencies (Yang, 1998), and to the level of differentiation among populations due to genetic drift only (Leinonen et al., 2013). This value is directly comparable to the interpopulation variance component in quantitative traits (P ST, see below). A 95% confidence interval (CI) was computed for the observed global F ST value using a classical cluster bootstrap procedure with 1,000 iterations (Field & Welsh, 2007): CI lower and upper bounds were computed as the 95% percentiles of a theoretical distribution of 1,000 F ST values obtained from the random sampling of the thirteen populations with replacement.
2.7. Statistical analyses
2.7.1. Trait variability among populations
For each of the four traits separately, we tested whether there was significant variability among the thirteen populations using an analysis of variance (ANOVA) with the population of origin as the explicative categorical variable. To meet the assumptions of Gaussian models (normality of the residuals and homoscedasticity), data were transformed: Body mass, metabolic rate, and excretion rate were log‐transformed and boldness was square‐root‐transformed.
2.7.2. Heterogeneity in trait covariations among populations
We tested whether covariations among the four traits (i.e., syndromes) were different among the thirteen populations. We first synthetized and described, for each population, patterns of trait covariation using path analysis. Traits were scaled to the mean within each population (i.e., each population displays a mean of zero with a variance of one for each trait), and a general path analysis linking each trait to the others (saturated path analysis) was computed for each population independently using the lavaan R‐package (Rosseel, 2012). These resulted in thirteen path models (each path model corresponding to a population's syndrome) and thirteen associated covariance matrices. Then, we then tested whether these path models (and hence trait covariations) varied among populations using a test of heterogeneity on covariance matrices among groups (metaSEM R‐package, Cheung, 2015). Briefly, this analysis allows assessing the heterogeneity of covariance matrices with a combination of indices (Hooper, Coughlan, & Mullen, 2008): (a) root mean square error of approximation (RMSEA, expected to be higher than 0.06 if the matrices are heterogeneous), (b) standardized root mean square residual (SRMR, expected to be higher than 0.09 if the matrices are heterogeneous), and (c) comparative fit index (CFI, expected to be lower than 0.96 if the matrices are heterogeneous).
2.7.3. Heterogeneity of pairwise covariations
We tested whether the six covariations considered separately differed among populations using a test of heterogeneity (Rosenberg, Adams, & Gurevitch, 1997). We estimated and extracted the covariations between each pair of traits (six pairs in total: mass–metabolism; mass–excretion; mass–boldness; metabolism–excretion; metabolism–boldness; and excretion–boldness) from the path models described above so as to control for all relationships among traits simultaneously. We applied meta‐analytic tools to analyze the heterogeneity in covariances. We applied the Z‐Fisher transformation to each covariance value (Cov) to obtain a standardized Zr using the following formula: , and we calculated the corresponding standard error as: (Nakagawa & Cuthill, 2007) where n is the sample size of the considered population. We estimated the degree of variability of Zr for each pair of traits among populations with a test of heterogeneity (Higgins & Thompson, 2002; Viechtbauer, 2010). This index (H) indicates the percentage of heterogeneity and tests whether heterogeneity in a data set is higher than that expected by chance. The standard error of Zr was added as a pondering parameter to the heterogeneity test to give more weight to populations with more individuals.
2.7.4. Effect of phylogeny
We tested whether phylogenetically related populations displayed similar traits and trait covariations using phylogenetic models (PGLS, Garland & Ives, 2000). These models allow incorporating the genetic relatedness among populations through a phylogenetic tree used to estimate a λ value corresponding to the degree of phylogenetic conservatism in the response variable. λ is expected to vary between 0 and 1, where 0 means no phylogenetic dependence in a trait among populations, and 1 means that the focal trait is phylogenetically conserved (Comte, Murienne, & Grenouillet, 2014; Harvey & Purvis, 1991). We calculated λ independently for each trait and each covariation (calculated from path analyses; see above) using only the intercept as fixed effect.
2.7.5. Effect of environmental characteristics
We used phylogenetic models to assess the effects of temperature and predation on traits and covariations. We ran PGLS for each trait and covariation (Zr) independently, with temperature, predation pressure (measured at the site level), and the resulting two‐term interaction as explanatory variables. The phylogenetic tree based on microsatellite markers was incorporated into each model to account for genetic relatedness among populations. When λ = 0, the model is equivalent to a classical linear model, whereas when λ = 1 it accounts for phylogenetic conservatism in trait. We then used an information‐theoretic approach, based on Akaike Information Criteria (AIC) comparisons, to select the model(s) that best fit the data. We considered model(s) that fell within a ΔAIC <4 as “best” model(s) as they would maximize the likelihood of the model while taking into account the number of parameters, and we rejected those with a ΔAIC>4 (Burnham & Anderson, 2002). We ran PGLS models using the pgls function from the caper R‐package (Orme et al., 2013).
2.7.6. F ST /P ST comparison
Finally, we tested whether variability in traits and covariations among populations were higher or not than expected under the hypothesis that differentiation is due to genetic drift only. To do so, we compared F ST calculated on neutral genetic markers (corresponding to the level of differentiation among populations expected if genetic drift only is affecting traits) to P ST values calculated for each trait and covariation independently. P ST is the phenotypic equivalent of the Q ST index, although calculated for wild populations when no information on the parental relatedness among individuals is available (Leinonen, Cano, MäKinen, & Merilä, 2006). A P ST value higher than the global F ST value indicates that phenotypic differentiation among populations is higher than expected by genetic drift only, and that mechanisms such as plasticity and/or selection might explain these differences (Leinonen et al., 2013). The use of F0 individuals allows comparison of natural trait variability and covariations existing among wild populations. However, this approach does not enable to tease apart genetic and plastic contributions to trait variability and covariations. Therefore, P ST here represents the level of phenotypic differentiation that is due to both genetic and developmental components. We estimated a P ST for each trait as: where and were, respectively, the among‐ and within‐population variance in the considered trait (Leinonen et al., 2013). Among‐ and within‐population variance components were estimated from generalized linear mixed models with the trait as response variable, the intercept as a fixed effect, and the population as a random effect (Leinonen et al., 2013).
In the case of covariations, among‐ and within‐population variance components were calculated in a similar way but with the addition of a random slope, corresponding to the covariable trait (Supporting Information Appendix S2). This allows estimating among‐ and within‐population variance in the covariation between each pair of traits (Mazé‐Guilmo, Blanchet, Rey, Canto, & Loot, 2016). The generalized linear mixed models were run using the lme4 R‐package (Bates, Maechler, Bolker, & Walker, 2014). We applied a classical bootstrap clustering procedure with 1,000 iterations (Field & Welsh, 2007) to assess the 95% confidence interval for P ST. We then compared the CI of P ST for each trait and each covariation (i.e., 10 P ST quantified in total: 4 single traits and 6 covariations among them) to the CI of F ST. All analyses were performed using R (R Core Team, 2013).
3. RESULTS
3.1. Trait variability among populations
Body mass (F = 29.859, df = 12, 349, p < 0.001), metabolic rate (F = 14.538, df = 12, 350, p < 0.001), excretion rate (F = 14.842, df = 12, 322, p < 0.001), and boldness (F = 5.179, df = 12, 329, p < 0.001) were all significantly different among populations (Figure 2). There was no strong evidence for phylogenetic conservatism for any of the traits (see Supporting Information Figure S1): λ was highest for body mass (λ = 0.87) and metabolic rate (λ = 0.74), although none of these values were significantly different from zero (Table 1). Regarding determinants of trait means, the best models explaining body mass included temperature, predation pressure, and their interaction (Table 1). Body mass increases as temperature decreases (negative relationship), and this increase was exacerbated as predation pressure increased (Figure 3a). The model selection for the three other traits led to equivalent models, and the null models were, in all‐three cases, the best models (Table 1). This suggested that metabolic rate, excretion rate, and boldness were neither—or weakly—related to temperature, nor to predation pressure. Finally, the estimates of phenotypic differentiation among populations (P ST) were high for body mass, metabolic rate, and excretion, and were significantly higher than the level of neutral genetic differentiation (F ST) (Figure 4). Phenotypic differentiation measured for boldness was not different from what was expected under the drift hypothesis.
Figure 2.
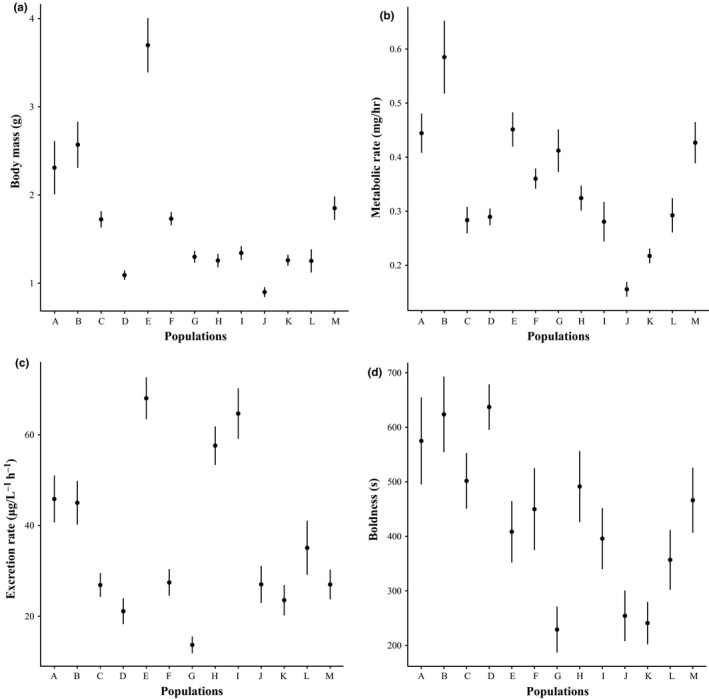
Mean trait values for body mass (a), metabolic rate (b), excretion rate (c), and boldness (d) in function of the population origin of fish. Error bars represent ±1SE
Table 1.
Results of the model selection to explain the variability of functional traits and their covariations among populations. All possible phylogenetic models (PGLS, see the main text) were run for each trait and then compared based on AIC. Bold values represent models that fell in a ΔAIC <4
| λ (p‐value) | Models | |||||
|---|---|---|---|---|---|---|
| Null | Temperature | Predation | Temperature and predation | Temperature‐by‐predation | ||
| Mass | 0.87 (0.12) | 7.982 | 7.018 | 9.997 | 0 | 0.194 |
| Metabolism | 0.74 (0.19) | 0 | 1.451 | 1.997 | 2.907 | 4.547 |
| Excretion | 0 (1) | 0 | 1.016 | 1.67 | 2.952 | 4.521 |
| boldness | 0.55 (1) | 0 | 1.982 | 1.932 | 3.925 | 5.924 |
| Mass–metabolism | 0 (1) | 4.123 | 3.528 | 1.966 | 3.929 | 0 |
| Mass–excretion | 0 (1) | 0 | 1.411 | 1.8 | 0.732 | 1.617 |
| Mass–boldness | 0 (1) | 0 | 1.93 | 1.057 | 2.559 | 2.27 |
| Metabolism–excretion | 0 (1) | 2.719 | 4.611 | 4.64 | 5.963 | 0 |
| Metabolism–boldness | 0 (1) | 0.35 | 1.698 | 0 | 1.757 | 3.332 |
| Excretion–boldness | 0 (1) | 0 | 1.102 | 1.853 | 2.862 | 3.639 |
Figure 3.
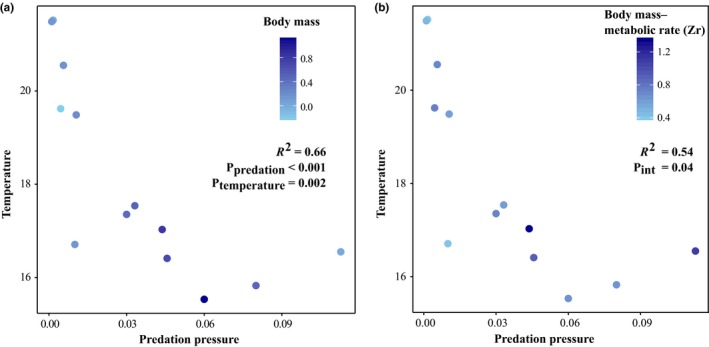
Interaction between temperature (°C) and predation pressure (ind.m2) explains the variation in body mass (a), and in the covariation between body mass and metabolic rate (b). The R 2 and the p‐values are extracted from the best models based on AIC selection (see Table 1), and “Pint” represents the p‐value for the interaction between temperature and predation
Figure 4.
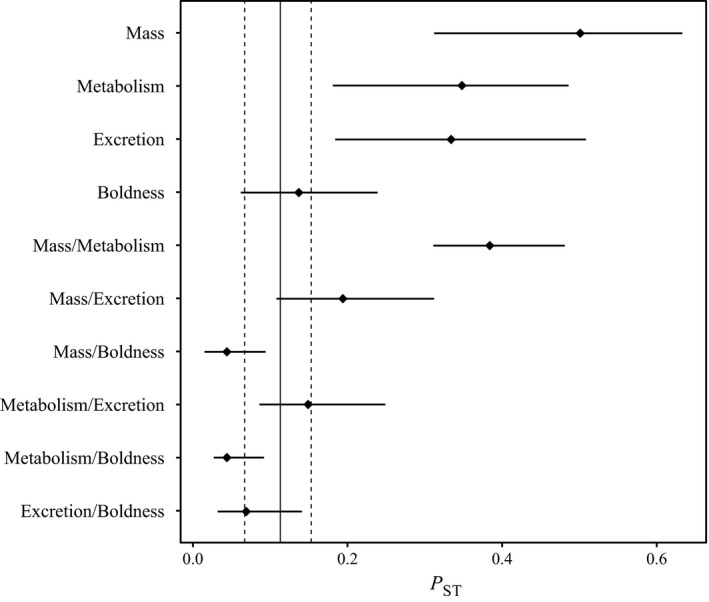
Estimates of P ST for each trait (body mass, metabolic rate, excretion rate, and boldness) and for each covariation (body mass–metabolic rate, body mass–excretion rate, body mass–boldness, metabolic rate–excretion rate, metabolic rate–boldness, and excretion rate–boldness), and F ST (vertical straight line) on neutral microsatellite markers. Horizontal bars represent 95% confident interval of P ST , and vertical dotted line represents 95% confident interval of F ST that were calculated using cluster bootstrap procedure
3.2. Among population heterogeneity in functional trait syndromes and covariations
We found that populations varied in their syndromes of functional traits since the matrices of covariations were heterogeneous (RMSEA = 0.266, CFI = 0.602, SRMR = 0.263, Supporting Information Figure S2). For instance, the syndrome in the population F was characterized by positive covariations among body mass, metabolic rate, and excretion rate, and a negative covariation between boldness and excretion rate (Figure 5a); whereas population L displayed negative covariations between body mass and boldness, boldness and metabolic rate, and metabolic and excretion rates, while the body mass–metabolic rate covariation was positive (Figure 5b).
Figure 5.
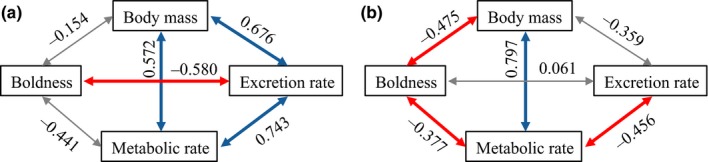
Syndromes of functional traits among populations of European minnow. Populations F and L were represented as examples in panel (a) and (b), respectively. Blue and red arrows denote significant positive and negative covariances, respectively, while the gray arrow represents nonsignificant covariance. Syndromes in all populations are displayed in Supporting Information Figure S3
This was confirmed since we also found strong significant heterogeneity among populations for several trait covariations. In particular, the covariations measured between body mass and excretion rate (H = 72.03%, Q = 45.837, df = 12, p < 0.001), between excretion rate and metabolic rate (H = 69.20%, Q = 41.229, df = 12, p < 0.001), and between excretion rate and boldness (H = 58.26%, Q = 31.296, df = 12, p = 0.002) strongly (and significantly) varied among populations (Figure 6b,e, and f). For instance, the covariation between metabolic and excretion rates was significantly positive for six populations, significantly negative for one population, and nonsignificant for the remaining populations (Figure 6e). The covariations between body mass and metabolic rate, between metabolic rate and boldness, and between body mass and boldness were homogeneous (p > 0.052, Figure 6a,c, and d).
Figure 6.
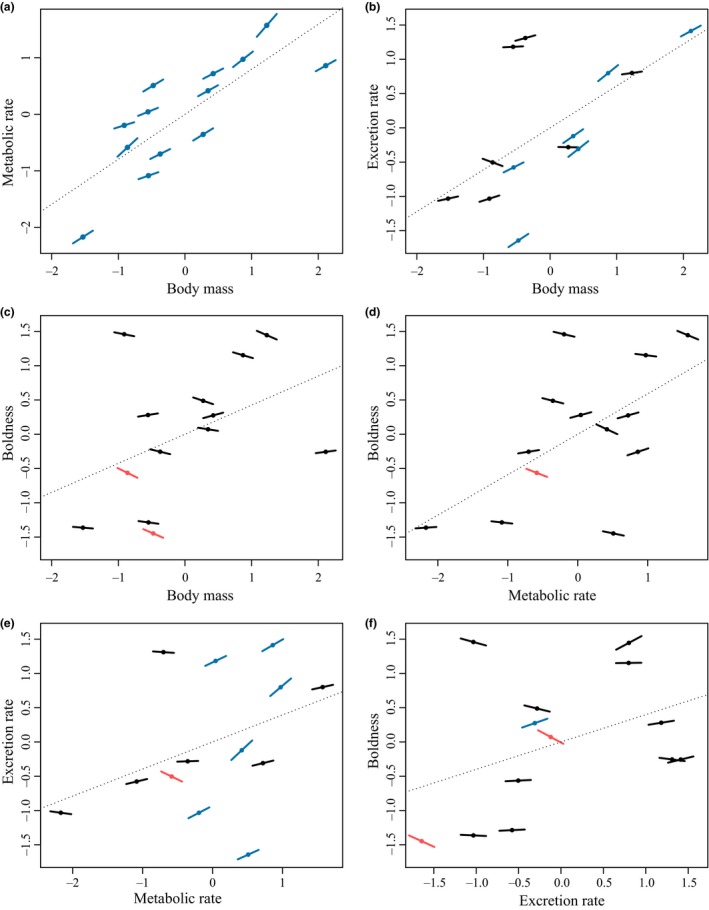
Covariations between each pair of functional traits: (a) body mass–metabolic rate, (b) body mass–excretion rate, (c) body mass–boldness, (d) metabolic rate–excretion rate, (e) metabolic rate–boldness, and (f) excretion rate–boldness. Points represent the average trait value for each population, and lines on points represent the covariations (i.e., the slope) between traits within each population. Blue and red lines indicate significant (α = 0.05) positive and negative covariations, respectively. The dotted lines represent the relationship between traits across the thirteen populations
We did not find evidence for significant phylogenetic conservatism for any of the covariations (Table 1 and Supporting Information Figure S3). The best models explaining the covariation between body mass and metabolic rate included temperature, predation, and the temperature‐by‐predation interaction term (Table 1). For this covariation, the null model was strongly rejected from the set of best‐supported models (ΔAIC >4), and the results suggested that the strength of the covariation tended to increase as the temperature decreased, and when the predation pressure increased (Figure 3b). Regarding other covariations, models including temperature and predation pressure were not strongly supported by the data as the null models were always selected within the set of models displaying a ΔAIC <4 (Table 1).
Finally, covariation measured between body mass and metabolic rate displayed a P ST value that was significantly higher than the global F ST value (Figure 4). P ST measured for the covariation between body mass and excretion rate was higher than the global F ST, but the CIs of the two estimates overlapped. For other trait covariations, P ST values were not significantly different from the global F ST value (Figure 4).
4. DISCUSSION
We demonstrated that functional traits, trait covariations, and syndromes they form strongly varied across populations of European minnow sampled in a large riverscape. We further found that multiple processes explained variability in functional traits, their covariations, and hence in syndromes of functional traits. For instance, we found evidence for adaptive mechanisms (plasticity and/or selection) related to water temperature and/or predation for explaining the covariation between body mass and metabolic rate. In parallel, we found that other traits and covariations were consistent with the hypothesis that genetic drift is sufficient to explain variability, which would suggest that even nonadaptive processes could sustain intraspecific variation in functional traits. Finally, we do not detect any evidence of evolutionary conservatism in any of the functional traits nor in their covariations.
We showed that body mass, metabolic rate, and excretion rate differed among populations more than expected by genetic drift only, suggesting trait divergences arose from selection and/or developmental plasticity. Although our design does not allow selection to be teased apart from developmental plasticity, our findings are theoretically sound and may suggest adaptation to environmental conditions since the decrease in body mass with temperature is expected for ectotherms (Daufresne, Lengfellner, & Sommer, 2009). Here, we found that both temperature and predation intensity affected body mass. We can speculate that higher body mass could allow minnows to reach a size refuge from predators, and/or to increase their locomotor performances to escape predators (Domenici, 2001; Villéger et al., 2017). Nonetheless, this result should be interpreted with care since our statistical power is weak and because of collinearity between water temperature and predation. Indeed, we could alternatively argue (based on the visual inspection of biplot, Supporting Information Figure S4) that a quadratic relationship (Supporting Information Figure S4) exists between body mass and predation pressure that we may fail to properly identify because of the small sample size and the collinearity with water temperature (Prunier & Blanchet, 2018). Collinearity can, in some cases, lead to inappropriate conclusions since it is difficult to discriminate the causal links among explicative variables, or because model estimates may be biased (Prunier & Blanchet, 2018; Prunier, Colyn, Legendre, Nimon, & Flamand, 2015). However, since the results are biologically sound, we are confident that body mass is adaptively related to environmental variables. We also found high variability in metabolic and excretion rates, which were also likely driven by adaptive mechanisms (Figure 4). Nonetheless, we failed to detect the environmental pressures driving divergences in these two traits. The variability in excretion rate probably stands in trophic and stoichiometric factors, such as trophic niche, elemental composition of resources, or allochthonous nutrient inputs (El‐Sabaawi, Warbanski, Rudman, Hovel, & Matthews, 2016; Evangelista et al., 2017), which could be characteristic of each geographical site. Hence, measuring stoichiometric variability of individuals and populations would benefit to infer hypotheses regarding variability in excretion rate.
We found that traits can not only vary among populations, but also that functional traits formed different syndromes among populations of European minnow. Indeed, the sets of covariations were different among populations, and multiple patterns were identified, with some trait covariations being more robust than others. For instance, the allometric relationships between body mass and metabolic rate, and between body mass and excretion rate were both positive across all populations, but the former was homogeneous among populations (i.e., stable), whereas the latter was heterogeneous and hence more flexible among populations (Figure 6). Similarly, the covariation between excretion rate and boldness was flexible, confirming that relationships between behavioral and physiological traits can be complex (Killen et al., 2013). These various functional trait covariations among populations subsequently generated variability in syndromes. Such variability has been documented in behavioral traits (Dingemanse et al., 2007) and morphological traits (Berner, Stutz, & Bolnick, 2010), but rarely among multiple types of traits. The various biological mechanisms—such as pleiotropy or allometry—underlying the links among traits might therefore be modulated differently among populations, resulting in difference of syndromes (Peiman & Robinson, 2017). Hence, it would be worth further investigating the biological mechanisms driving trait covariations to better appraise the variability of functional syndromes (Killen, Atkinson, & Glazier, 2010; Raffard et al., 2017).
Although we detected variability in syndromes of functional traits, the lack of determinants (i.e., temperature or predation) and the low P ST values for most covariations suggest that a non‐negligible part of the heterogeneity in syndromes variability may—in our case—arise from the effect of genetic drift. Actually, the relationship between body mass and metabolic rate was the only covariation whose variability was likely driven by adaptive mechanisms. Indeed, as revealed by the P ST/F ST analysis and the trait–environment analysis, we found evidence that selection and/or plasticity associated with predation pressure and water temperature may drive variation observed among populations. Previous works have reported variability in the allometric relationship between body mass and metabolic rate at both inter‐ and intraspecific levels in many organisms (Bokma, 2004; Glazier, 2005; Seibel, 2007). Here, covariations increase as temperature decreases and predation increases (Figure 2b). Although this should be interpreted with care (see statistical caution above), the metabolic allometry might vary allowing individuals to optimize energetic efficiency under different environmental constraints (Glazier, 2005; Killen et al., 2010). Fish can notably adapt their lifestyle to increase or decrease their energetic assimilation in order to cope with biotic and abiotic constraints, such as predation (Killen et al., 2010). This confirms that trait architecture within populations can be complex, and—in some cases—allow individuals to adapt/acclimatize to their environment (Peiman & Robinson, 2017).
To conclude, we found that syndromes in functional traits can strongly vary among populations, and that both adaptive (natural selection and/or plasticity) and nonadaptive processes (genetic drift) are driving intraspecific heterogeneity in these syndromes. Since functional traits can affect ecological processes (Lavorel & Garnier, 2002; Raffard et al., 2017; Violle et al., 2007), the variability in functional syndromes may exert puzzling effects on ecological processes. For instance, the variability in covariations involving excretion rate may have implications for the dynamics of nutrient recycling and ecological stoichiometry (Atkinson, Capps, Rugenski, & Vanni, 2017; Vanni, 2002); while in some populations, large individuals should excrete a high quantity of nitrogen, they should excrete a low quantity of nitrogen in other populations, with potential consequences for primary production (Evangelista et al., 2017; McIntyre et al., 2008). Variability of syndromes may have further ecological effects through trophic mechanisms since individuals with different functional traits may have different trophic niches (Villéger et al., 2017). Trophic variability can subsequently affect community structure and ecosystem functioning (Des Roches, Shurin, Schluter, & Harmon, 2013). Further studies should aim to experimentally test how heterogeneity in functional syndromes is acting on ecological dynamics.
CONFLICT OF INTEREST
None declared.
AUTHOR CONTRIBUTIONS
AR, SB, FS, and JC designed the study. AR and SB performed fieldwork, and AR performed trait measurement. GL carried out the processing of genetic samples. JGP analyzed genetic data and AR performed statistical analyses. All authors interpreted and discussed the results. AR and SB wrote the first draft of the paper. All authors corrected and improved the paper, and approved this version of the manuscript.
Supporting information
ACKNOWLEDGMENTS
We thank Kéoni Saint‐Pe, Geoffrey Marselli, and other students for their helps during fieldworks. We also thank Antoine Fargette for his help on trait measurement and animal husbandry. AR is financially supported by a Doctoral scholarship from the Université Fédérale de Toulouse. This work was undertaken at SETE, which is part of the “Laboratoire d'Excellence” (LABEX) entitled TULIP (ANR‐10‐LABX‐41).
Raffard A, Cucherousset J, Prunier JG, Loot G, Santoul F, Blanchet S. Variability of functional traits and their syndromes in a freshwater fish species (Phoxinus phoxinus): The role of adaptive and nonadaptive processes. Ecol Evol. 2019;9:2833–2846. 10.1002/ece3.4961
DATA ACCESSIBILITY
Data are available on Figshare. https://doi.org/10.6084/m9.figshare.7623854.
REFERENCES
- Aljanabi, S. (1997). Universal and rapid salt‐extraction of high quality genomic DNA for PCR‐ based techniques. Nucleic Acids Research, 25, 4692–4693. 10.1093/nar/25.22.4692 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Armbruster, W. S. , & Schwaegerle, K. E. (1996). Causes of covariation of phenotypic traits among populations. Journal of Evolutionary Biology, 9, 261–276. 10.1046/j.1420-9101.1996.9030261.x [DOI] [Google Scholar]
- Atkinson, C. L. , Capps, K. A. , Rugenski, A. T. , & Vanni, M. J. (2017). Consumer‐driven nutrient dynamics in freshwater ecosystems: From individuals to ecosystems. Biological Reviews, 92, 2003–2023. 10.1111/brv.12318 [DOI] [PubMed] [Google Scholar]
- Bassar, R. D. , Bryan, B. L. , Marshall, M. C. , Pringle, C. M. , Reznick, D. N. , & Travis, J. (2016). Local adaptation of fish consumers alters primary production through changes in algal community composition and diversity. Oikos, 594–603. [Google Scholar]
- Bates, D. , Maechler, M. , Bolker, B. , & Walker, S. (2014). lme4: Linear mixed‐effects models using Eigen and S4 . R package version 1.1‐7.
- Bell, A. M. , & Sih, A. (2007). Exposure to predation generates personality in threespined sticklebacks (Gasterosteus aculeatus). Ecology Letters, 10, 828–834. 10.1111/j.1461-0248.2007.01081.x [DOI] [PubMed] [Google Scholar]
- Berner, D. , Stutz, W. E. , & Bolnick, D. I. (2010). Foraging trait (co)variances in stickleback evolve deterministically and do not predict trajectories of adaptive diversification. Evolution, 64, 2265–2277. 10.1111/j.1558-5646.2010.00982.x [DOI] [PubMed] [Google Scholar]
- Bestion, E. , Teyssier, A. , Aubret, F. , Clobert, J. , & Cote, J. (2014). Maternal exposure to predator scents: Offspring phenotypic adjustment and dispersal. Proceedings of the Royal Society B‐Biological Sciences, 281, 20140701 10.1098/rspb.2014.0701 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Biro, P. A. , Beckmann, C. , & Stamps, J. A. (2010). Small within‐day increases in temperature affects boldness and alters personality in coral reef fish. Proceedings of the Royal Society B‐Biological Sciences, 277, 71–77. 10.1098/rspb.2009.1346 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Bokma, F. (2004). Evidence against universal metabolic allometry. Functional Ecology, 18, 184–187. 10.1111/j.0269-8463.2004.00817.x [DOI] [Google Scholar]
- Brown, J. H. , Gillooly, J. F. , Allen, A. P. , Savage, V. M. , & West, G. B. (2004). Toward a metabolic theory of ecology. Ecology, 85, 1771–1789. 10.1890/03-9000 [DOI] [Google Scholar]
- Burnham, K. P. , & Anderson, D. R. (2002). Model selection and multimodel inference: A practical information‐theoretic approach. New York, NY: Springer. [Google Scholar]
- Capps, K. A. , & Flecker, A. S. (2013). Invasive aquarium fish transform ecosystem nutrient dynamics. Proceedings of the Royal Society B‐Biological Sciences, 280, 20131520. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Cheung, M.‐W.‐L. (2015). metaSEM: An R package for meta-analysis using structural equation modeling. Frontiers in Psychology, 5, 1521 10.3389/fpsyg.2014.01521 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Collin, H. , & Fumagalli, L. (2011). Evidence for morphological and adaptive genetic divergence between lake and stream habitats in European minnows (Phoxinus phoxinus, Cyprinidae). Molecular Ecology, 20, 4490–4502. 10.1111/j.1365-294X.2011.05284.x [DOI] [PubMed] [Google Scholar]
- Collin, H. , & Fumagalli, L. (2015). The role of geography and ecology in shaping repeated patterns of morphological and genetic differentiation between European minnows (Phoxinus phoxinus) from the Pyrenees and the Alps. Biological Journal of the Linnean Society, 116, 691–703. [Google Scholar]
- Comte, L. , Murienne, J. , & Grenouillet, G. (2014). Species traits and phylogenetic conservatism of climate‐induced range shifts in stream fishes. Nature Communications, 5, 5053 10.1038/ncomms16201 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Darwin, C. (1859). On the origin of species by natural selection, or the preservation of favoured races in the struggle for life. London, UK: John Murray. [PMC free article] [PubMed] [Google Scholar]
- Daufresne, M. , Lengfellner, K. , & Sommer, U. (2009). Global warming benefits the small in aquatic ecosystems. Proceedings of the National Academy of Sciences of the United States of America, 106, 12788–12793. 10.1073/pnas.0902080106 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Defossez, E. , Pellissier, L. , & Rasmann, S. (2018). The unfolding of plant growth form‐defence syndromes along elevation gradients. Ecology Letters, 21, 609–618. 10.1111/ele.12926 [DOI] [PubMed] [Google Scholar]
- Des Roches, S. , Shurin, J. B. , Schluter, D. , & Harmon, L. J. (2013). Ecological and evolutionary effects of stickleback on community structure. PLoS ONE, 8, e59644 10.1371/journal.pone.0059644 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Des Roches, S. , Post, D. M. , Turley, N. E. , Bailey, J. K. , Hendry, A. P. , Kinnison, M. T. , … Palkovacs, E. P. (2018). The ecological importance of intraspecific variation. Nature Ecology & Evolution, 2, 57–64. 10.1038/s41559-017-0402-5 [DOI] [PubMed] [Google Scholar]
- Díaz, S. , Kattge, J. , Cornelissen, J. H. C. , Wright, I. J. , Lavorel, S. , Dray, S. , … Gorné, L. D. (2016). The global spectrum of plant form and function. Nature, 529, 167–171. 10.1038/nature16489 [DOI] [PubMed] [Google Scholar]
- Díaz, S. , Purvis, A. , Cornelissen, J. H. C. , Mace, G. M. , Donoghue, M. J. , Ewers, R. M. , … Pearse, W. D. (2013). Functional traits, the phylogeny of function, and ecosystem service vulnerability. Ecology and Evolution, 3, 2958–2975. 10.1002/ece3.601 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Dingemanse, N. J. , Wright, J. , Kazem, A. J. N. , Thomas, D. K. , Hickling, R. , & Dawnay, N. (2007). Behavioural syndromes differ predictably between 12 populations of three‐spined stickleback. Journal of Animal Ecology, 76, 1128–1138. 10.1111/j.1365-2656.2007.01284.x [DOI] [PubMed] [Google Scholar]
- Domenici, P. (2001). The scaling of locomotor performance in predator‐prey encounters: From fish to killer whales. Comparative Biochemistry and Physiology, 131, 169–182. 10.1016/S1095-6433(01)00465-2 [DOI] [PubMed] [Google Scholar]
- Edeline, E. , Lacroix, G. , Delire, C. , Poulet, N. , & Legendre, S. (2013). Ecological emergence of thermal clines in body size. Global Change Biology, 19, 3062–3068. 10.1111/gcb.12299 [DOI] [PubMed] [Google Scholar]
- El‐Sabaawi, R. W. , Warbanski, M. L. , Rudman, S. M. , Hovel, R. , & Matthews, B. (2016). Investment in boney defensive traits alters organismal stoichiometry and excretion in fish. Oecologia, 181, 1209–1220. 10.1007/s00442-016-3599-0 [DOI] [PubMed] [Google Scholar]
- Evangelista, C. , Lecerf, A. , Britton, J. R. , & Cucherousset, J. (2017). Resource composition mediates the effects of intraspecific variability in nutrient recycling on ecosystem processes. Oikos, 126, 1439–1450. 10.1111/oik.03787 [DOI] [Google Scholar]
- Field, C. A. , & Welsh, A. H. (2007). Bootstrapping clustered data. Journal of the Royal Statistical Society: Series B (Statistical Methodology), 69, 369–390. 10.1111/j.1467-9868.2007.00593.x [DOI] [Google Scholar]
- Finstad, A. G. , Forseth, T. , Ugedal, O. , & NæSje, T. F. (2007). Metabolic rate, behaviour and winter performance in juvenile Atlantic salmon. Functional Ecology, 21, 905–912. 10.1111/j.1365-2435.2007.01291.x [DOI] [Google Scholar]
- Fourtune, L. , Paz‐Vinas, I. , Loot, G. , Prunier, J. G. , & Blanchet, S. (2016). Lessons from the fish: A multi‐species analysis reveals common processes underlying similar species‐genetic diversity correlations. Freshwater Biology, 61, 1830–1845. 10.1111/fwb.12826 [DOI] [Google Scholar]
- Fourtune, L. , Prunier, J. G. , Mathieu‐Begne, E. , Canto, N. , Veyssiere, C. , Loot, G. , & Blanchet, S. (2018). Intraspecific genetic and phenotypic diversity: parallel processes and correlated patterns? bioRxiv, 10.1101/288357 [DOI] [Google Scholar]
- Friard, O. , & Gamba, M. (2016). BORIS: A free, versatile open‐source event‐logging software for video/audio coding and live observations. Methods in Ecology and Evolution, 7, 1325–1330. 10.1111/2041-210X.12584 [DOI] [Google Scholar]
- Frost, W. E. (1943). The natural history of the minnow, Phoxinus phoxinus . The Journal of Animal Ecology, 12, 139. [Google Scholar]
- Funk, J. L. , Larson, J. E. , Ames, G. M. , Butterfield, B. J. , Cavender‐Bares, J. , Firn, J. , … Wright, J. (2016). Revisiting the Holy Grail: Using plant functional traits to understand ecological processes. Biological Reviews, 92, 1156–1173. [DOI] [PubMed] [Google Scholar]
- Garland, T. , & Ives, A. R. (2000). Using the past to predict the present: Confidence intervals for regression equations in phylogenetic comparative methods. American Naturalist, 155, 346–364. 10.1086/303327 [DOI] [PubMed] [Google Scholar]
- Gillooly, J. F. (2001). Effects of size and temperature on metabolic rate. Science, 293, 2248–2251. 10.1126/science.1061967 [DOI] [PubMed] [Google Scholar]
- Glaholt, S. P. , & Vanni, M. J. (2005). Ecological responses to simulated benthic‐derived nutrient subsidies mediated by omnivorous fish. Freshwater Biology, 50, 1864–1881. 10.1111/j.1365-2427.2005.01456.x [DOI] [Google Scholar]
- Glazier, D. S. (2005). Beyond the: Variation in the intra‐ and interspecific scaling of metabolic rate in animals. Biological Reviews, 80, 611. [DOI] [PubMed] [Google Scholar]
- Golovanov, V. K. (2013). Ecophysiological patterns of distribution and behavior of freshwater fish in thermal gradients. Journal of Ichthyology, 53, 252–280. 10.1134/S0032945213030016 [DOI] [Google Scholar]
- Goudet, J. (2005). hierfstat, a package for r to compute and test hierarchical F‐statistics. Molecular Ecology Notes, 5, 184–186. 10.1111/j.1471-8286.2004.00828.x [DOI] [Google Scholar]
- Harvey, P. H. , & Purvis, A. (1991). Comparative methods for explaining adaptations. Nature, 351, 619–624. 10.1038/351619a0 [DOI] [PubMed] [Google Scholar]
- Hawlena, D. , & Schmitz, O. J. (2010). Physiological stress as a fundamental mechanism linking predation to ecosystem functioning. American Naturalist, 176, 537–556. 10.1086/656495 [DOI] [PubMed] [Google Scholar]
- Helsen, K. , Acharya, K. P. , Brunet, J. , Cousins, S. A. O. , Decocq, G. , Hermy, M. , … Graae, B. J. (2017). Biotic and abiotic drivers of intraspecific trait variation within plant populations of three herbaceous plant species along a latitudinal gradient. BMC Ecology, 17, 38 10.1186/s12898-017-0151-y [DOI] [PMC free article] [PubMed] [Google Scholar]
- Higgins, J. P. T. , & Thompson, S. G. (2002). Quantifying heterogeneity in a meta‐analysis. Statistics in Medicine, 21, 1539–1558. 10.1002/sim.1186 [DOI] [PubMed] [Google Scholar]
- Hildrew, A. G. , Raffaelli, D. , & Edmonds‐Browns, R. (2007). Body size: The structure and function of aquatic ecosystems. Cambridge, UK: Cambridge University Press. [Google Scholar]
- Hooper, D. , Coughlan, J. , & Mullen, M. R. (2008). Structural equation modelling: Guidelines for determining model fit. Electronic Journal on Business Research Methods, 6, 8. [Google Scholar]
- Keenan, K. , McGinnity, P. , Cross, T. F. , Crozier, W. W. , & Prodöhl, P. A. (2013). diveRsity: An R package for the estimation and exploration of population genetics parameters and their associated errors. Methods in Ecology and Evolution, 4, 782–788. [Google Scholar]
- Keith, P. , Persat, H. , Feunteun, E. , & Allardi, J. (2011). Les poissons d’eau douce de France. Paris and Mèze: Muséum National d’Histoire Naturelle and Publications Biotope. [Google Scholar]
- Killen, S. S. , Atkinson, D. , & Glazier, D. S. (2010). The intraspecific scaling of metabolic rate with body mass in fishes depends on lifestyle and temperature. Ecology Letters, 13, 184–193. 10.1111/j.1461-0248.2009.01415.x [DOI] [PubMed] [Google Scholar]
- Killen, S. S. , Marras, S. , & McKenzie, D. J. (2011). Fuel, fasting, fear: Routine metabolic rate and food deprivation exert synergistic effects on risk‐taking in individual juvenile European sea bass. Journal of Animal Ecology, 80, 1024–1033. 10.1111/j.1365-2656.2011.01844.x [DOI] [PubMed] [Google Scholar]
- Killen, S. S. , Marras, S. , Metcalfe, N. B. , McKenzie, D. J. , & Domenici, P. (2013). Environmental stressors alter relationships between physiology and behaviour. Trends in Ecology & Evolution, 28, 651–658. 10.1016/j.tree.2013.05.005 [DOI] [PubMed] [Google Scholar]
- Langerhans, R. B. (2007). Evolutionary consequences of predation: Avoidance, escape, reproduction, and diversification In Elewa A. M. T. (Ed.), Predation in organisms (pp. 177–220). Berlin Heidelberg: Springer. [Google Scholar]
- Lavorel, S. , & Garnier, E. (2002). Predicting changes in community composition and ecosystem functioning from plant traits: Revisiting the Holy Grail. Functional Ecology, 16, 545–556. 10.1046/j.1365-2435.2002.00664.x [DOI] [Google Scholar]
- Leinonen, T. , Cano, J. M. , MäKinen, H. , & Merilä, J. (2006). Contrasting patterns of body shape and neutral genetic divergence in marine and lake populations of threespine sticklebacks. Journal of Evolutionary Biology, 19, 1803–1812. 10.1111/j.1420-9101.2006.01182.x [DOI] [PubMed] [Google Scholar]
- Leinonen, T. , McCairns, R. J. S. , O’Hara, R. B. , & Merilä, J. (2013). QST–FST comparisons: Evolutionary and ecological insights from genomic heterogeneity. Nature Reviews Genetics, 14, 179–190. 10.1038/nrg3395 [DOI] [PubMed] [Google Scholar]
- Mazé‐Guilmo, E. , Blanchet, S. , Rey, O. , Canto, N. , & Loot, G. (2016). Local adaptation drives thermal tolerance among parasite populations: A common garden experiment. Proceedings of the Royal Society B‐Biological Sciences, 283, 20160587 10.1098/rspb.2016.0587 [DOI] [PMC free article] [PubMed] [Google Scholar]
- McIntyre, P. B. , Flecker, A. S. , Vanni, M. J. , Hood, J. M. , Taylor, B. W. , & Thomas, S. A. (2008). Fish distributions and nutrient cycling in streams: Can fish create biogeochemical hotspots. Ecology, 89, 2335–2346. 10.1890/07-1552.1 [DOI] [PubMed] [Google Scholar]
- Nakagawa, S. , & Cuthill, I. C. (2007). Effect size, confidence interval and statistical significance: A practical guide for biologists. Biological Reviews, 82, 591–605. 10.1111/j.1469-185X.2007.00027.x [DOI] [PubMed] [Google Scholar]
- Nei, M. (1973). Analysis of gene diversity in subdivided populations. Proceedings of the National Academy of Sciences of the United States of America, 70, 3321–3323. 10.1073/pnas.70.12.3321 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Orme, D. Freckleton, R. , Thomas, G. , Petzoldt, T. , Fritz, S. , Isaac, N. , & Pearse, W. (2013). caper: Comparative analyses of phylogenetics and evolution in R. Retrieved from https://cran.r-project.org/web/packages/caper/index.html
- Paradis, E. , Claude, J. , & Strimmer, K. (2004). APE: Analyses of phylogenetics and evolution in R language. Bioinformatics, 20, 289–290. 10.1093/bioinformatics/btg412 [DOI] [PubMed] [Google Scholar]
- Paz‐Vinas, I. , Loot, G. , Hermoso, V. , Veyssiere, C. , Poulet, N. , Grenouillet, G. , & Blanchet, S. (2018). Systematic conservation planning for intraspecific genetic diversity. Proceedings of the Royal Society B, 285, 20172746. 10.1098/rspb.2017.2746 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Peiman, K. S. , & Robinson, B. W. (2012). Diversifying and correlational selection on behavior toward conspecific and heterospecific competitors in brook stickleback (Culaea inconstans). Ecology and Evolution, 2, 2141–2154. 10.1002/ece3.339 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Peiman, K. S. , & Robinson, B. W. (2017). Comparative analyses of phenotypic trait covariation within and among populations. American Naturalist, 190, 451–468. 10.1086/693482 [DOI] [PubMed] [Google Scholar]
- Poulet, N. , Beaulaton, L. , & Dembski, S. (2011). Time trends in fish populations in metropolitan France: Insights from national monitoring data. Journal of Fish Biology, 79, 1436–1452. 10.1111/j.1095-8649.2011.03084.x [DOI] [PubMed] [Google Scholar]
- Pruitt, J. N. , Riechert, S. E. , Iturralde, G. , Vega, M. , Fitzpatrick, B. M. , & Avilés, L. (2010). Population differences in behaviour are explained by shared within‐population trait correlations. Journal of Evolutionary Biology, 23, 748–756. 10.1111/j.1420-9101.2010.01940.x [DOI] [PubMed] [Google Scholar]
- Prunier, J. G. , & Blanchet, S. (2018). Using connectivity to identify climatic drivers of local adaptation: A response to Macdonald et al . Ecology Letters, 21, 1121–1123. 10.1111/ele.12972 [DOI] [PubMed] [Google Scholar]
- Prunier, J. G. , Colyn, M. , Legendre, X. , Nimon, K. F. , & Flamand, M. C. (2015). Multicollinearity in spatial genetics: Separating the wheat from the chaff using commonality analyses. Molecular Ecology, 24, 263–283. 10.1111/mec.13029 [DOI] [PubMed] [Google Scholar]
- R Core Team (2013). R: A language and environment for statistical computing. Vienna, Austria: R Foundation for Statistical Computing. [Google Scholar]
- Raffard, A. , Lecerf, A. , Cote, J. , Buoro, M. , Lassus, R. , & Cucherousset, J. (2017). The functional syndrome: Linking individual trait variability to ecosystem functioning. Proceedings of the Royal Society B‐Biological Sciences, 284, 20171893 10.1098/rspb.2017.1893 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Raffard, A. , Santoul, F. , Cucherousset, J. , & Blanchet, S. (2018). The community and ecosystem consequences of intraspecific diversity: A meta‐analysis. Biological Reviews. 10.1111/brv.12472 [DOI] [PubMed] [Google Scholar]
- Reale, D. , Garant, D. , Humphries, M. M. , Bergeron, P. , Careau, V. , & Montiglio, P.‐O. (2010). Personality and the emergence of the pace‐of‐life syndrome concept at the population level. Philosophical Transactions of the Royal Society B: Biological Sciences, 365, 4051–4063. 10.1098/rstb.2010.0208 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Rice, W. R. (1989). Analyzing tables of statistical tests. Evolution, 43, 223. [DOI] [PubMed] [Google Scholar]
- Roff, D. A. (1992). The evolution of life histories. Theory and analysis. New York, NY: Chapman & Hall. [Google Scholar]
- Rosenberg, M. S. , Adams, D. C. , & Gurevitch, J. (1997). MetaWin. In Statistical software for conducting meta-analysis: Fixed effect models, mixed effect models, and resampling tests. Version 1.0. Sunderland, MA: Sinauer Associates. [Google Scholar]
- Rosseel, Y. (2012). lavaan: An R package for structural equation modeling. Journal of Statistical Software, 48, 1–36. [Google Scholar]
- Rousset, F. (2008). genepop’007: A complete re‐implementation of the genepop software for Windows and Linux. Molecular Ecology Resources, 8, 103–106. 10.1111/j.1471-8286.2007.01931.x [DOI] [PubMed] [Google Scholar]
- Schmitz, O. J. , Hawlena, D. , & Trussell, G. C. (2010). Predator control of ecosystem nutrient dynamics. Ecology Letters, 13, 1199–1209. 10.1111/j.1461-0248.2010.01511.x [DOI] [PubMed] [Google Scholar]
- Seibel, B. A. (2007). On the depth and scale of metabolic rate variation: Scaling of oxygen consumption rates and enzymatic activity in the Class Cephalopoda (Mollusca). Journal of Experimental Biology, 210, 1–11. 10.1242/jeb.02588 [DOI] [PubMed] [Google Scholar]
- Sih, A. , Bell, A. , & Johnson, J. C. (2004). Behavioral syndromes: An ecological and evolutionary overview. Trends in Ecology & Evolution, 19, 372–378. 10.1016/j.tree.2004.04.009 [DOI] [PubMed] [Google Scholar]
- Stearns, S. (1992). The evolution of life histories. Oxford, UK: Oxford University Press. [Google Scholar]
- Van Oosterhout, C. , Hutchinson, W. F. , Wills, D. P. M. , & Shipley, P. (2004). micro‐checker: Software for identifying and correcting genotyping errors in microsatellite data. Molecular Ecology Notes, 4, 535–538. 10.1111/j.1471-8286.2004.00684.x [DOI] [Google Scholar]
- Vanni, M. J. (2002). Nutrient cycling by animals in freshwater ecosystems. Annual Review of Ecology and Systematics, 33, 341–370. 10.1146/annurev.ecolsys.33.010802.150519 [DOI] [Google Scholar]
- Viechtbauer, W. (2010). Conducting meta‐analyses in R with the metafor package. Journal of Statistical Software, 36, 1–48. 10.18637/jss.v036.i03 [DOI] [Google Scholar]
- Villéger, S. , Brosse, S. , Mouchet, M. , Mouillot, D. , & Vanni, M. J. (2017). Functional ecology of fish: Current approaches and future challenges. Aquatic Sciences, 79, 783–801. 10.1007/s00027-017-0546-z [DOI] [Google Scholar]
- Villéger, S. , Grenouillet, G. , Suc, V. , & Brosse, S. (2012). Intra‐ and interspecific differences in nutrient recycling by European freshwater fish. Freshwater Biology, 57, 2330–2341. 10.1111/fwb.12009 [DOI] [Google Scholar]
- Violle, C. , Navas, M.‐L. , Vile, D. , Kazakou, E. , Fortunel, C. , Hummel, I. , & Garnier, E. (2007). Let the concept of trait be functional! Oikos, 116, 882–892. [Google Scholar]
- Yang, R.‐C. (1998). Estimating hierarchical F ‐statistics. Evolution, 52, 950–956. [DOI] [PubMed] [Google Scholar]
- Závorka, L. , Koeck, B. , Cucherousset, J. , Brijs, J. , Näslund, J. , Aldvén, D. , … Johnsson, J. I. (2017). Co‐existence with non‐native brook trout breaks down the integration of phenotypic traits in brown trout parr. Functional Ecology, 31, 1582–1591. 10.1111/1365-2435.12862 [DOI] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.
Supplementary Materials
Data Availability Statement
Data are available on Figshare. https://doi.org/10.6084/m9.figshare.7623854.