

Abstract
This study investigates the amphisbaenian species skull which includes cranium, lower jaw and hyoid apparatus. The medial dorsal bones comprise the premaxilla, nasal, frontal and parietal. The premaxilla carries a large medial tooth and two lateral ones. The nasals are paired bones and separated by longitudinal suture. Bones of circumorbital series are frontal, orbitosphenoid and maxilla. The occipital ring consists of basioccipital, supraoccipital and exooccipital. Supraoccipital and basioccipital are single bones while the exo-occipitals are paired. The bones of the palate comprise premaxilla, maxilla, septomaxilla, palatine, pterygoid, ectopterygoid, basisphenoid, parasphenoid, orbitosphenoid and laterosphenoid. Prevomer and pterygoid teeth are absent. Palatine represent by two separate bones. The temporal bones are clearly visible. The lower jaw consists of the dentary, articular, coronoid, supra-angular, angular and splenial. The hyoid apparatus is represented by a Y-shaped structure. The mandible is long and is suspended from the braincase via relatively short quadrate. There is an extensive contact between the long angular and the large triangular coronoid. Thus inter-mandibular joint is bridged completely by the angular and consequently, the lower jaws are relatively rigid and kinetic. The maxillae are suspended from the braincase largely by ligaments and muscles rather than through bony articulation. In conclusion, the skull shape affects feeding strategy in Diplometopon zarudnyi. The prey is ingested and transported via a rapid maxillary raking mechanism.
Keywords: Squamata, Amphisbaenia, Diplometopon zarudnyi, Skull, Morphology
1. Introduction
Amphisbaenians are a poorly known reptile with over 150 extant species occurring in suitable habitats over the world. They are fossorial reptiles, most of which are limbless. Many species of Amphisbaenians showed some adaptation in their cranium related to their mode of life and behaviors such as adaptive head for burrowing behavior in the friable or sandy soils where they live in (Kearney et al., 2005).
A morphological difference between extant and fossil amphisbaenians may be due to the progress in the evolution of the skull elements (Kearney, 2003). The scorbutic amphisbaenians are defined almost from a number of well-defined skulls which has been found in a good case and structures from ancient’s ages (Kearney et al., 2005, Maisano et al., 2006). Amphisbaena has a strong head with a wide snout, not discriminated from the neck (Gans et al., 1977). Most of the skull bones are solid and strong and have a characteristic single medial tooth in the upper jaw. As all lizards, amphisbaenians have not the outer ears and the eyes are deeply recessed and covered with skinned scales.
Snakes are reptiles that are specialized for eating large prey and some have number of features which are common with amphisbaenians and burrowing lizards. It has been suggested that snakes may have evolved from burrowing ancestors. The infra order Scolecophidia (blind snakes) which includes two families whose individuals are burrowing in the ground similar to amphisbaenians in mode of life. These two families are Typhlopidae (blind snakes) and Leptotyphlopidae (slender blind snakes) which considered as the most primitive snake (Allam and Abo-Eleneen, 2012) and more advanced than amphisbaenians which the most advanced lizard are. The amphisbaenian group takes a separate evolutionary path and is by no means intermediate between snakes and lizards (Gans, 1969).
Romer (1956) classified the amphisbaenians as an isolated infra-order among lizards. The morphology of both living and fossil members has been studied (Gans, 1968, Zangerel, 1945). Zangerl (1945) examined several species of Amphisbaena and reported a comparative study of their post-cranial osteology. Maisano et al. (2006) studied the cranial anatomy of spade-headed amphisbaenian. The nasal capsule of Amphisbaena cubana and Amphisbaena fuliginosa has been described by Zangerl (1944). Gans and Wever (1972) described the middle ear and measured the electrical potentials of the cochlea in certain species of Amphisbaena. The current study aimed to elaborate the morphology of amphisbaenian Diplometopon zarudnyi skull bones and the relation between mode of feeding and bones articulation and morphology.
2. Materials and methods
In the present study, 6 specimens of D. zarudnyi were collected in May from Qatif Oasis, Saudi Arabia. The animals were narcotized, labeled, injected with formalin and preserved in glass jars with 10% formalin solution. To show the whole mount morphology, the skull D. zarudnyi was investigated by X-ray.
2.1. Decalcification and serial histological studies
Five fresh specimens of D. zarudnyi were anesthetized in the lab using chloroform. The heads were taken and fixed in aqueous Bouin’s solution for 72 h. The fixed heads were washed in running water and decalcified with EDTA solution for about 20 days with changing the solution every 2 days for complete decalcification. At the end of this process, the heads were washed several times with 70% ethyl alcohol, embedd in molten paraffin wax and 5µm serial transverse sections (Tra) were prepared. The sections were stained with Ehrlich’s haematoxylin and eosin (Mallory, 1944) and Masson's trichrome stain (Drury et al., 1976). Periodic acid/Schiff’s (PAS) method was used for polysaccharide staining (McManus, 1946). To differentiate neutral mucopolysaccharides from the acid mucopolysaccharides, PAS/AB staining (Mowry, 1956) was employed. Bromophenol blue was used for staining proteins (Mazia et al., 1953). Erythrocytes and cytoplasm were identified by using Azur and Eosin (Lillie, 1965).
3. Results
The results of this study show the structure of skull compartments of D. zarudnyi. The three major compartments are: the cranium, the lower jaw and the hyoid apparatus (Fig. 1).
Fig. 1.
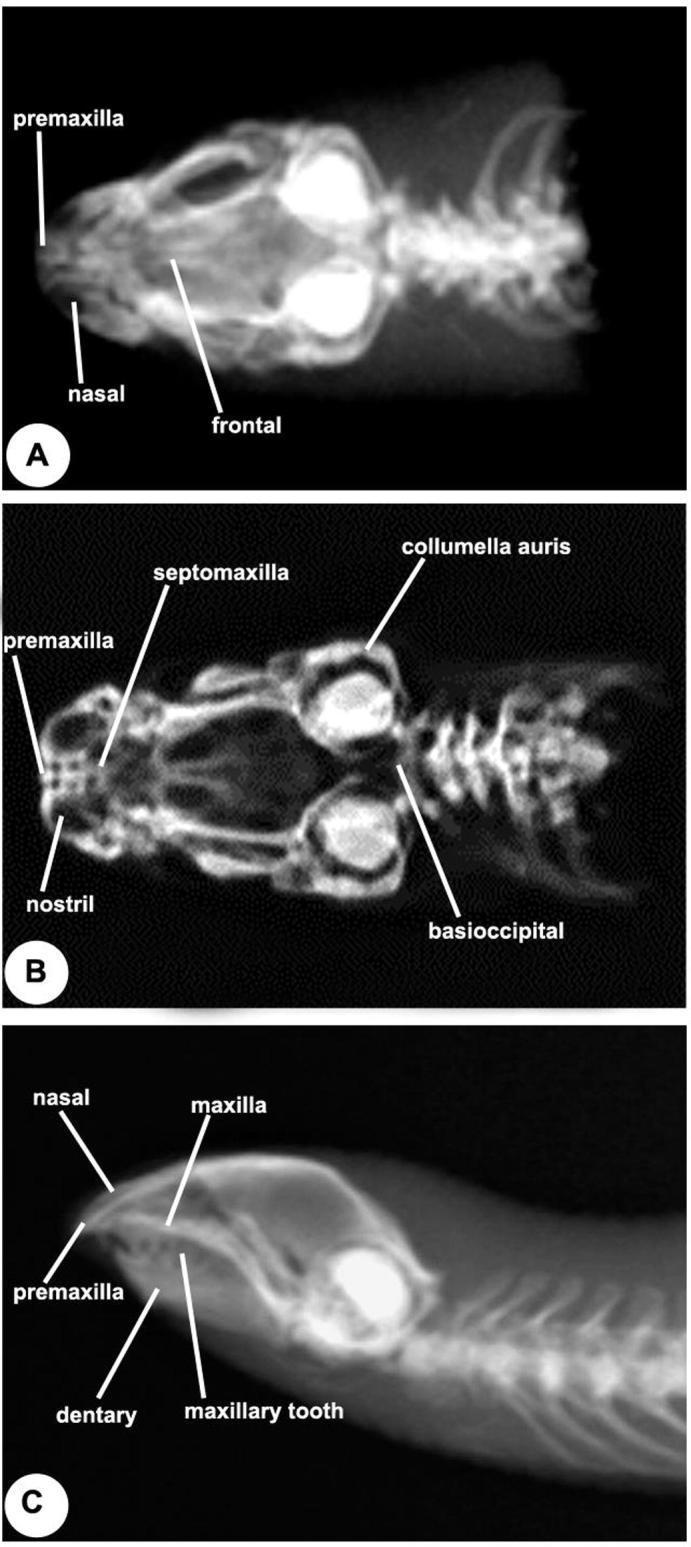
X-ray photos of skull of Diplometopon zarudnyi show that the nasofrontal joint located in-between the frontal bones, nasal and the snout unit consists of the nasals, premaxillae, septomaxillae and vomer. (A) Dorsal view, (B) Ventral view, (C) Lateral view.
I)- The cranium consists of the medial dorsal bones, the upper jaw bones, circumorbital elements series, basio occipital circle, otic capsule elements, palate and temporal bones.
-
(a)
The medial dorsal bones: These are further sub-divided into four categories.
1- Premaxillae (Fig. 1, Fig. 2, Fig. 3, Fig. 4; Tra 003- 029) combining into single short rounded broad bone forming the most anterior part of the cranium and bordering the external nostrils. The premaxilla bone sends a medial processus nasalis separating the two nasals and the anterior portion of the frontals. Ventrally, the premaxilla bone sends a backward palatal process to receive the anterior tip of the vomer. The palatal process of premaxilla carries large medial and two small lateral teeth.
Fig. 2.

Selected transverse sections (Tra) of Diplometopon zarudnyi skull. A, Tra001 Masson's trichrome, B, Tra002 PAS-Alcian blue, C, Tra003 bromophenol blue, D, Tra005 Masson's trichrome, E, Tra007 Masson's trichrome, F, Tra008 PAS-Alcian blue. X 40. d, dentary; dn, duct of naso-lacrymal; fr, frontal; mu, muscle; mx, maxilla; n, nasal; na, nasal anteroom; nc, nasal chamber; ng, nasal gland; nln, nervus lateralis nasi; nmn, nervus medialis nasi; pfr, prefrontal; pmx, premaxilla; smx, septomaxilla; snc, septum nasi cartilagineum; t, tooth; to, tongue.
Fig. 3.

Selected transverse sections (Tra) of Diplometopon zarudnyi skull. A, Tra009 bromophenol blue, B, Tra011 Masson's trichrome, C, Tra013 Masson's trichrome, D, Tra014 PAS-Alcian blue, E, Tra015 bromophenol blue, F, Tra017 Masson's trichrome. X 40. dng, duct of nasal gland; jo, jacobson's organ; lng, lateral nasal gland; mt, maxillary tooth; pc, pulp cavity.
Fig. 4.

Selected transverse sections (Tra) of Diplometopon zarudnyi skull. A, Tra019 Masson's trichrome, B, Tra023 Masson's trichrome, C, Tra024 Masson's trichrome, D, Tra025 Masson's trichrome, E, 027 H.E., F, Tra029 Masson's trichrome. X 40. aiaf, anterior inferior alveolar foramen; cf, conchal fold; dc, dentary canal; dnt, dentary tooth; p, parietal; slg, sublingual gland; tc, trabecula communis; tel, telencephalon; v, vomer.
2- Nasal (Fig. 1, Fig. 2, Fig. 3, Fig. 4; Tra 001- 023) is paired and forms the dorsal roof of the external nostrils and laterally articulated with the maxilla bones. Each nasal bone is articulated laterally with the processus nasalis through a longitudinal suture. The nasal bone is rectangular in shape.
3- Frontal (Fig. 1, Fig. 2, Fig. 3, Fig. 4; Tra 007 - 023) is paired and separated at the mid-line by the posterior tip of the processus nasalis of the premaxilla. The frontal extended anteriorly as three large processes to fit with the nasal bone by one medial process and the maxillary bones by two lateral processes. The posterior end of the frontals carried also two processes which are firmly articulated with the anterior border of the parietal.
4- Parietal (Fig. 4, Fig. 5, Fig. 6, Fig. 7, Fig. 8; Tra 027 - 100) is a single unpaired bone forming most of the dorsal roof of the cranium. The parietal bone is joined posterodorsally with the anterior margin of the otic capsule of each side. Laterally, the parietal bone sends two lateral down growth which are totally separated in the midventral line.
-
(b)
Bones of the upper jaw: These further divided into two categories:
Fig. 5.

Selected transverse sections (Tra) of Diplometopon zarudnyi skull. A, Tra030 Masson's trichrome., X 40, B, Tra031 Masson's trichrome., X 40, C, Tra034 Masson's trichrome., X 40, D, Tra041 Azur., X 200, E, Tra037 Masson's trichrome., X 40, F, Tra043 Masson's trichrome., X 40. ang, angular; cb, ciliary body; chv, choanal vault; co, coronoid; ec, ectopterygoid; hg, harderian gland; l, lens; opc, optic capsule; os, orbitosphenoid; pal, palatine.
Fig. 6.

Selected transverse sections (Tra) of Diplometopon zarudnyi skull. A, Tra049, B, 054, C, 056, D, 061, E, 062, F, 065. Masson's trichrome., X 40. cc, cranial cavity; mb, mixed bone; mc, meckelian canal; pbs, parabasisphenoid; sp, splenial.
Fig. 7.

Selected transverse sections (Tra) of Diplometopon zarudnyi skull. A, Tra066, B, 068, C, 070, D, 075, E, 078, F, 082. Masson's trichrome., X 40. appr, alar process of prootic; exc, extracolumella; gf, gasserian foramen; pt, pterygoid; q, quadrate; sc, semicircular canal; vc, vidian canal.
Fig. 8.

Selected transverse sections (Tra) of Diplometopon zarudnyi skull. A, Tra084, B, 085, C, 087, D, 094, E, 095, F, 100. Masson's trichrome., X 40. avsc, anterior vertical semi-circular canal; bo, basioccipital; c, columella; enc, endolymphatic canal; jr, jugular process; lsph, laterosphenoid; ooc, otic-occipital complex; otc, otic capsule; pas, processus ascendes of supraoccipital; pd, perilymphatic duct; pop, paraoccipital process; pro, prootic, rst, recessus scalae tympani; sq, squamosal; vb, vestibule.
1- Maxilla (Fig. 1, Fig. 2, Fig. 3, Fig. 4; Tra 001-019) are two large, strong and separated bones articulating anteriorly with the nasals. Laterally, each maxilla bone has a small protrusion interlocked with the corresponding frontal bones. Each maxilla bone carries a single row of maxilla teeth on its ventral surface.
2- Quadrate (Fig. 1, Fig. 7, Fig. 8; Tra 078-087) is a short, stout and nearly triangular bone directed anteroventrally to take an oblique position. The proximal portion of the quadrate is articulated with the lower jaw, while the distal part is attached to the otic capsule. Laterally, the pterygoid is fixed to the inner side of the distal portion of the quadrate.
-
(c)
Bones of the circumorbiatal series:
In D. zarudnyi, the orbit bones are the frontal, orbitosphenoid and maxilla. The maxilla forms the ventral and anterior borders of the orbit; the orbitosphenoid is a triangular bone extending along the ventral rim of the cranium and share in forming the inner walls of the orbit while the parietal forms its roof.
-
(d)
The occipital Ring: These are further sub-divided into three categories.
1- The Basioccipital (Fig. 1, Fig. 8; Tra 084-100) constructs the most posterior portion of the cranial floor. The basioccipital bones possess two lateral elevations on both sides of its anterior part. There are two sutures surrounding the basioccipital, one is located anteriorly between the basioccipital and basisphenoid, while the other is found between the basioccipital and exoccipital.
2- The Supraoccipital (Fig. 1, Fig. 8; Tra 084-100) is a single medial bone roofing the foramen magnum and laterally is attached to the otic capsule. A distinct suture is present between the supraoccipital and parietal bone.
3- The Exoccipitals (Fig. 1, Fig. 8; Tra 084-100) are two separate bones. Each one is found to be fused with the supraoccipital and the otic capsule without any distinct sutures. Ventrally, there is a clearly visible suture between the exoccipitals and basioccipitals.
-
(e)
The otic capsule:
The otic capsule is composed of three bones which formed the lateral side of the posterior part of the cranium. These bones are the prootic, laterosphenoid and opisthotic. It is almost fused with the occipital bones forming a single structure. Laterally, it is overlapped by the posterior edge of the parietal plate from which it can be detached. It is perforated by a large rounded fenestra ovalis in which the plate of the collumella auris is fitted. The Lateral shenoid is a small bone present on each side of the parasphenoid and shares in the formation of the side wall of the cranial cavity (Tra 087-100).
-
(f)
Bones of the palate: These are further sub-divided into seven categories.
1- The Vomer (Fig. 1, Fig. 3, Fig. 4, Fig. 5; Tra 023-037) is elongated; roughly triangular paired element situated in the anterior part of the palate and formed the floor of the nasal caspsule. The anterior part of it extends to articulate with the palatal process of premaxilla. Small part of the septomaxilla is clearly visible from the palatal view of the cranium and between the palatal process of maxilla and the vomer bone.
2- Palatine (Fig. 1, Fig. 5, Fig. 6; Tra 034-054) is two separate elongated bones lying on both sides of the vomer. Each palatine has a deep concavity on its ventral surface. Anterolaterally, the palatine articulates with the maxilla and ectopterygoid. Posteriorly, the vomer extends to a small distance from the palatine bone.
3- Pterygoid (Fig. 7, Fig. 8; Tra 078-082) is a triangular bone situated on both sides of the palate and extended from the palatine towards the head of the quadrate. It is terminated into a narrow curved end which embraces the inner side of the head of the quadrate.
4- Ectopterygoid (Fig. 5, Fig. 6; Tra 043-049) is a small bone found in front of the pterygoid forming a considerable part of the cranial floor.
5- Parasphenoid (Fig. 6, Fig. 7, Fig. 8; Tra 054-082) is totally fused with the basisphenoid forming most of the cranial floor.
6- Orbitosphenoid (Fig. 5, Fig. 6, Fig. 7; Tra 043-075) bone extends along the ventral surface of the cranium between the down growths of the frontal and the parietal bones.
7- Temporal bones (Fig. 8; Tra082-100) are represented by small supratemporal found just above the fenestra ovalis. The proximal end of the quadrate is attached to the otic capsule, while the distal end is attached to the lower jaw. The small supratemporal bone serves to attach the narrow posterior end of the proximal part of the quadrate to the cranium just above the fenestra ovalis.
II) The lower jaw (Fig. 1, Fig. 2, Fig. 3, Fig. 4, Fig. 5, Fig. 6; Tra 007-062) ramus is nearly triangular in shape. The ramus composed of dentary, articular, coronoid, supra-angular, angular and splenial. The dentary, is the largest bone forming most of the ramus and overlapping the articular bone posteriorly. The articular forms a concave condyle to receive the quadrate bone. The coronoid is clearly visible from the inner side overlapping the supra-angular. Meanwhile, on the inner side of each ramus, the angular bone extended posteriorly to reach the splenial.
III) The hyoid apparatus (Fig. 1C) is consisted of an inverted Y-shaped structure to which two pairs of rods on both sides are attached. The Y-shaped medial structure represents the hyoid corpus. It is extended anterior forming the processus lingualis and posteriorly forming two second ceratobranchials. The anterolateral rods represent the hyoid arch. These rods are not fused with the hyoid corpus. The postero-lateral rods represent the first ceratobranchials which may represent the posterior cornua. The first ceratobranchials are attached posteriorly with a small cartilaginous epibranchial.
4. Discussion
D. zarudnyi is a worm-like burrowing reptile classified at one time with the lizards but now generally given a subordinal rank within the Squamata in parallel with lizards and snakes since it has some characters typical of snakes and lizards. It is limbless, long and cylindrical with blunt head, short pointed tail and very small eyes. The skull of D. zarudnyi, exhibits the general pattern of the amphisbaenid skull although certain minor differences exist. The bones roofing the cranium have interlocking sutures as mentioned by Bellairs (1950) for burrowing lizards. The snout is adapted for digging as the premaxillary bone is large, rounded and possesses an elongated processus nasalis wedged between the nasals and reaching the frontals posteriorly. The premaxillae are fused together forming the anterior most part of the skull. In Amphisbaenidae, the fusion of the premaxillae into a single bone was reported by El-Assy and Al-Nassar (1976). This condition is commonly found in other ophidian families (Abdeen, 1991). The premaxillary bone sends dorsally a strong medial processus nasalis which separate the nasals and extends to wedge between the anterior ends of the frontals. This condition also faced Zangerl (1944) in Bipes biporus, Amphisbaena cubana, Geocalamus acutus, Monopeltis capensis and Leposternon microcephalum. The two premaxillae are fused to form the anterior part of the rostrum. It seems that this character reflects the burrowing mode of life.
The palatal process of premaxillae carries large medial strong teeth and two smaller lateral ones behind. The teeth are strong, sharply pointed directed backwards. Zangerl (1944) found only a single medial premaxillary tooth in Rhineura floridana. In D. zarudnyi, the fused premaxillae are connected with the maxillae through distinct sutures. However, in most advanced snakes, the premaxillae are separated from the maxillae (Zaher, 1990, Abdeen, 1991).
The mode of feeding in amphisbaenians which was described by Gans (1969) is by biting pieces of their prey where the grasping effectiveness is enhanced by the jaw mechanism of many species in which the mandibular condyle lies depressed far below the toothline. Gans (1969) further added that the closing of the jaws induces a backward directed force which pushes the prey between the teeth and locks it in position. Moreover, Gans (1968) considered the short and heavy jaws of amphisbaenids and the overlapped dentition between them to squash the small arthropods and pang chunks out of other large animals.
The paired appearance of the nasal bone which is clearly recognized in the species study is also found in most Typhlops (El-Sadany, 2014). Moreover, a paired nasal was recorded in most advanced snakes (Abdeen, 1991). On the other hand, the nasals are absolutely absent in Chelonia (Kingsley, 1925). The two naslas are entirely separated from each other by the extended processus nasalis of the premaxillae. Gans et al. (1977) described such a process in D. zarudnyi, as a finger of the nasal investing into the frontal almost one-third of the frontal length. In some amphisbaenids, the nasals articulate suturally with each other in the middle dorsal line as found in Amphisbaena fuliginosa and Rhineura floridana (Zangerl, 1944).
In D. zarudnyi, the frontals are paired elements. This character is generally found in blind snakes (El-Sadany, 2014). Moreover, paired frontals are found in some geckos, skinks, lacertids, amphisbaenids and varanids (Bahl, 1937, El–Toubi, 1938, Kamal, 1965).
The fused configuration of the parietals in D. zarudnyi represents a general rule among advanced snakes (El–Toubi and El–Assy, 1972). The processes of the frontals which interlock with those of the parietals and extending radially outward was reported to act as a device which function to strengthen the edge and the roof of the orbit against the stresses tending to twist or bend the skull around its long axis as documented by Gans (1969).
Bones of the upper jaw usually comprise the premaxillae, maxilla and quadrate, however, jugal and quadrate-jugal are entirely absent. The maxillae are strong bones found on both sides of the premaxillae and resting on their palatal processes. However, the maxilla does not reach the premaxilla in the colubrid snakes studied by Zaher (1990). Romer (1956) mentioned that the maxilla lacks the primitive contact with the premaxilla in ophidia except in typhlopids, leptotyphlopids and uropeltid groups.
The quadrate is a large triangular bone; its proximal part is attached to the cranium while its distal part articulates with the lower jaw. In lacertilia, the quadrate articulates not only with the squamosal, but also with the paraoccipital process of the exoccipital, while in ophidia it never articulates with this process (Kingsley, 1925). The inner side of the distal end of the quadrate is fixed with the pterygoid bone. However, in Acontias meleagris (Brock, 1941) the quadrate is entirely separated from the pterygoid.
The prefrontal, postfrontal, postorbital, lachrymal and jugal are completely disappeared. However, the presence of lachrymal or prefrontal bone in amphisbaenids was reported by some authors. Kritzinger (1946) emphasized its presence in Monopeltis. Also, Zangerl (1944) referred to the presence of prefrontal in some amphisbaenid skull. In D. zarudnyi, the prefrontal is absent. On the other hand, in Liotyphlops albirostris, the prefrontal has an anterior enlarged portion which articulates with the maxilla and a posterior enlarged portion which articulates with the lateral process of the frontal. The postfrontal bone is completely absent, as indicated in Amphisbaena alba (Williston, 1925).
The basioccipital is a single unpaired bone and shares with the exoccipital in the formation of the single tripartite occipital condyle. Similar observation was recorded by Abdeen (1991). In D. zarudnyi, the supraoccipital is a single bone. Similar observation was recorded by Maisano et al. (2006).The two exoccipitals border the foramen magnum laterally, wheras they are totally fused with the supraoccipital above and with the otic capsules in the regions of the opisthotic bone. On the other hand, in Anniella pulehra (Coe and Kunkel, 1906), the exoccipitals are fused with the basi and supraoccipital bones. Moreover, the exoccipitals are fused with the supraoccipitals in Typhlops braminus and Typhlops boettgeri, while in Typhlops. lineatus a complete fusion between the exoccipitals, supraoccipitals and the prootics was recorded (List, 1966).
In D. zarudnyi, the otic capsules are fused with the occipital bones forming a single structure overlapped by the posterior edges of the parietal plate. Thus the prootic bones are not clearly distinct from the otic capsule. In the amphisbanids, the prootics and parietals are found in broad contact which seems to be a characteristic feature in burrowing reptiles (Romer, 1956). In the present species of D. zarudnyi, the laterosphenoid is a small bone found on each side of the parabasisphenoid and share in the formation of the side wall of the cranial cavity. It articulates anteriorly with the orbitosphenoid and excluding it from the foramen of the Gasserian ganglion. The laterosphenoid articulates antero-dorsally with the parietal downgrowths and postero-laterally with the otic capsule. The morphology of the columella auris in the D. zarudyni, is described a large fenestra ovalis, a very short columellar part of the stapes and a cartilaginous extracolumella which lies in a shallow groove in the outer surface of the mandible. Similar observations were recorded by Maisano et al. (2006).
The vomer in the present study appeared as two conjugated bones which form the floor of the vomero-nasal compartment. The current vomer configuration may be resembles to Rhineura vomer anatomy as stated by Maisano et al. (2006). Also, the vomer contributes to the the sides of the choana, internarial girder and the roof of the palate and as reported by Maisano et al. (2006). The septomaxilla is an elongated bone wedged between the premaxilla, the nasal, the prevomer. The septomaxilla really forms considerable portion in the anterior region of the palate, which is considered as a distinctive character among snakes. However, in lizards the septomaxilla takes no part in the formation of the palate (Kamal et al., 1970). The septomaxilla bone is absent in Chelonia and Crocodilia (Williston, 1925). In D. zarudnyi, the palatines are in close contact with the vomers on both sides and they extend posteriorly beyond and diverge on both sides of the rostrum of the parabasisphenoid. Zangerl (1944) mentioned that the palatines in Amphisbaena cubana are joined medially in front of the parabasisphenoid bone, thus separating the frontal end of the parabasisphenoid from the backward ends of the vomers. The pterygoid is toothless, as in other Leptotyphlopids and Typhlopid snakes (Abdeen et al., 1991a, Abdeen et al., 1991b, El–Sadany, 2014). In D. zarudnyi, the epipterygoid is also lacking. This was also confirmed by Maisano et al. (2006). In D. zarudnyi, the ectopterygoid is present. In contrast, this bone is absent in Leptotyphlops (Abdeen, 1991). The parabasisphenoid complex is totally separate in the early embryonic stages and fused in the adult. These observations were recorded by (Abdeen and Zaher, 1992). El-assy and Al-Nassar (1976) referred to the presence of orbitosphenoid bone in the amphisbaenid skull D. zarudnyi. They gave a detailed description and recorded that the two orbitosphenoids of both sides are fused together into a medain thick bony attached to the ventrolateral sides of the frontal and parietal downgrowths. The orbitosphenoids fused in many amphisbaenians, highly increased and possibly augmented by a dermal component (Maisano et al., 2006).
In D. zarudnyi, the proximal part of the quadrate is found to be attached to the otic capsule of the cranium and the distal part articulates with the lower jaw. The narrow posterior end of the proximal part of the quadrate is found to be attached to a small- scale like supratemporal bone just above the fenestra ovalis. The supratemporal bone was previously applied by Kritzinger (1946) in Monopeltis capensis and by Bellairs (1950) in Anniella pulchera.
In D. zarudnyi, the dentary is the largest bone forming most of the ramus laterally. It carries a row of six acrodont teeth, while Rhineura carried seven subpleurodont teeth and others Amphisbaena carried eight subpleurodont teeth (Maisano et al., 2006). The articular bone prolongates into a small retroarticular process ventral to the articular condyle which receives that of the quadrate. The current splenial is elongated bone extending in front of the angular and the suture between the two bones was not quite distinct. However, Zangerl (1944) mentioned that the splenial in the amphisbaenid skull is very small and not distinct. In D. zarudnyi, the angular was only found in the inner side of the ramus facing the splenial. However, Camp (1923) referred to the fusion of the angular with the articular in Amphisbaenidae. In D. zarudnyi, the current results showed that the coronoid is a simple and small bone that forms the dorsal rim of the coronoid process. The current study confirms the observations of other previous study concerned to amphisbaenians coronoid interfering with dentary to form dentary coronoid process (Kearney, 2003, Maisano et al., 2006). Moreover, the coronoid bone was found in snake families belonging to Annillidae (Hoge, 1962). Others studies reported that the coronoid is totally absent in Colubridae and Elapidae (Kamal and Hammouda, 1967, Mohammed, 1988).
In D. zarudnyi, the hyoid apparatus consisted of a cartilaginous inverted Y-shaped structure to which are attached two pairs of rods on side, an antero-lateral cartilaginous pair and a posterolateral pair which is bony. The Y-shaped medial structure represents the hyoid corpus. Cope, 1892, Maisano et al., 2006) described the hyoid of Amphisbaena sp but he neglected the presence of epibranchial and the anterior cornua was shown fused with the hyoid corpus. In conclusion, the skull shape affects feeding strategy in D. zarudnyi. The prey is ingested and transported via a rapid maxillary raking mechanism.
Acknowledgement
The authors would like to thank the Deanship of Scientific Research at Princess Nourah bint Abdulrahman University for funding this work through the Research Groups Program grant no. (RG-1438-002).
Footnotes
Peer review under responsibility of King Saud University.
References
- Abdeen A.M. Further studies on the ophidian cranial osteology: the skull of the Egyptian black cobra, Walterinnesia aegyptia, (Family Elapidae). A. The dorsal bones, bones of the upper jaw, circumorbital series and occipital ring. J. Egypt Ger. Soc. Zool. 1991;5:367–389. [Google Scholar]
- Abdeen A.M., Zaher M.M. Futher studies on the ophidian osteocrania: the osteocranium of the snake Eryx jaculus (Family: Boidae) J. Egypt Ger. Soc. Zool. 1992;7(B):295–333. [Google Scholar]
- Abdeen A.M., Abo-Taira A.M., Zaher M.M. Further studies on the Ophidian cranial osteology: the skull of the Egyptian blind snake Leptotyphlops cairi (Family, Leptotyphlopidae) I. The cranium. A-The medial dorsal bones, bones of the upper jaw, circumorbital series and occipital ring. J. Egypt Ger. Soc. Zool. 1991;5:417–437. [Google Scholar]
- Abdeen A.M., Abo-Taira A.M., Zaher M.M. Further studies on the Ophidian cranial osteology: the skull of the Egyptian blind snake Leptotyphlops cairi (Family, Leptotyphlopidae) I. The cranium. B-The otic capsule, palate and temporal bones. J. Egypt Ger. Soc. Zool. 1991;5:439–455. [Google Scholar]
- Allam A.A., Abo El–Eneen R.E. Scales microstructure of some snakes inhabited the Egyptian Area. Zool. Sci. 2012;29(11):770–775. doi: 10.2108/zsj.29.770. [DOI] [PubMed] [Google Scholar]
- Bahl K.N. The skull of Varanus monitor (linn.) Rec. Indian Mus. 1937;39:133–147. [Google Scholar]
- Bellaris A.D. Observations on the cranial anatomy of Anniella and a comparison with that of other burrowing lizards. Proc. Zool. Soc. London. 1950;119:887–904. [Google Scholar]
- Brock G.T. The skull of Acontias meleagris, with a study of the affinities between lizards and snakes. J. Linn. Soc. 1941;42:71–88. [Google Scholar]
- Camp C.L. Classification of the lizards. Bull. Am. Mus. Nat. Hist. 1923;48:289–481. [Google Scholar]
- Coe W.R., Kunkel B.W. Studies on the California limbless lizard, Anniella. Trans. Conn. Acad. 1906;12:366–403. [Google Scholar]
- Cope E.D. The osteology of the lacertilia. Proc. Am. Philos. Soc. 1892;30:185–222. [Google Scholar]
- Drury R.A., Wallington E.A., Cancerson R. fourth ed. Oxford University Press; Oxford, London, New York: 1976. Carlton’s Histopathological Techniques. [Google Scholar]
- El–Assy Y.S., Al–Nassar N.A. Morphological study of the cranial osteology of the amphisbaenian D. zarudnyi. J. Univ. Kuwait (Sci.) 1976;3:113–142. [Google Scholar]
- El–Sadany D.A. Beni–Suef Univ; Egypt: 2014. Anatomical Studies on the Skull and the Digestive System of the Blind Snake, Rhamphotyphlops braminus. M Sc Thesis. [Google Scholar]
- El-Toubi M.R. The osteology of the lizard Scincus scincus. Bull. Fac. Sci. Cairo Univ. 1938;14:5–38. [Google Scholar]
- El–Toubi M.R., El–Assy Y.S. A comparative study of the cranial osteology of the Egyptian snakes from the family Colubridae. Proc. Zool. Soc. AR Egypt. 1972;4:289–328. [Google Scholar]
- Gans C. Relative success of divergent pathways in Amphisbaenian specialization. Am. Nat. 1968;102:345–362. [Google Scholar]
- Gans C. Amphisbaenian–reptiles specialized for a burrowing existence. Endeavour. 1969;28(105):140–151. [Google Scholar]
- Gans C., Wever E.G. The ear and hearing in Amphisbaenia (Reptilia) J. Exp. Zool. 1972;179:17–34. [Google Scholar]
- Gans C., Carl A., Sandra S. Amphisbaena medemi, an interesting new species from Colombia (Amphisbaenia, Reptilia) Fieldiana Zool. (Field Museum of Natural History) 1977;72:2–40. [Google Scholar]
- Hoge A.R. A notes on Xenopeltis unicolor. Mem. Inst. Butantan. 1962;30:31–33. [Google Scholar]
- Kamal A.M. The cranial osteology of adult Chalcides ocellatus. Anat. Anz. 1965;117:338–370. [PubMed] [Google Scholar]
- Kamal A.M., Hammouda H.G. The cranial osteology of the adult Psammophis sibilans. Bull. Fac. Sci. Cairo Univ. 1967;41:119–149. [Google Scholar]
- Kamal A.M., Hammouda H.G., Mokhtar F.M. The development of the osteocranium of the Egyptian Cobra. III. The otic capsule, palate, temporal bones, lower jaw and hyoid apparatus of the adult osteocranium. Acta Zool. 1970;51:31–42. [Google Scholar]
- Kearney M. Systematics of the Amphisbaenia (Lepidosauria: Squamata) based on morphological evidence from Recent and fossil forms. Herpetol. Monogr. 2003;17:1–74. [Google Scholar]
- Kearney M., Maisano J.A., Rowe T. Cranial anatomy of the extinct amphisbaenian Rhineura hatcherii (Squamata, Amphisbaenia) based on high-resolution X-ray computed tomography. J. Morphol. 2005;264:1–33. doi: 10.1002/jmor.10210. [DOI] [PubMed] [Google Scholar]
- Kingsley J.S. The vertebrate Skeleton. Jhon Marry London. 1925:1220–1320. [Google Scholar]
- Kritzinger C.C. The cranial anatomy and kinesis of the South African amphisbaenid Monopeltis capensis, Smith. S Afr. J. Sci. 1946;42:175–204. [Google Scholar]
- Lillie R.D. third ed. McGraw–Hill; New York: 1965. Histopathologic Technic and Practical histochemistry. [Google Scholar]
- List G.C. Comparative osteology of the snake families Typhlopidae and Leptotyphlopidae. Illinois Biol. Monogr. 1966;36:1–112. [Google Scholar]
- Maisano J.A., Kearney M., Rowe T. Cranial Anatomy of the spade–headed amphisbaenian D. zarudnyi (Squamata, Amphisbaenia) based on high–resolution X-ray computed tomography. J. Mprphol. 2006;267:70–102. doi: 10.1002/jmor.10388. [DOI] [PubMed] [Google Scholar]
- Mallory F.B. WB Saunders Philadelphia; UK: 1944. Pathological Technique. [Google Scholar]
- Mazia D., Brewer A., Alfert M. The cytochemical staining and measurement of protein with mercuric bromophenol blue. Biol. Bull. 1953;104:57–67. [Google Scholar]
- McManus J.F. The histological demonstration of mucin after periodic acid. Nature. 1946;158:202–211. doi: 10.1038/158202a0. [DOI] [PubMed] [Google Scholar]
- Mohammed M.H. The skeletal anatomy of Cerastes vipera (Viperidae, Reptilia) Proc. Zool. Soc. ARE. 1988;15:223–236. [Google Scholar]
- Mowry R.W. Alcian blue technique for histochemical study of acidic carbohydrates. J. Histochem. Cytochem. 1956;4:407–412. [Google Scholar]
- Romer A.S. The Univ. of Chicago press; Chicago–London: 1956. Osteology of the Reptilia. [Google Scholar]
- Williston S.W. Harvard Univ.; 1925. The Osteology of the Reptiles. 340–620. [Google Scholar]
- Zaher M.M. Further studies on the ophidian cranial osteology, The skull of the colubrid snake Natrix tessellata. I. The cranium. A. The medial dorsal bones, bones of the upper jaw, circumorbital series and occipital ring. J. Egypt Ger. Soc. Zool. 1990;2:309–327. [Google Scholar]
- Zangerel R. Contributions to the osetology of the skull of the Amphisbaenidae. Am. Midl. Nat. 1944;31:417–454. [Google Scholar]
- Zangerel R. Contributions to the osetology of the post-cranial skeleton of the Amphisbaenidae. Am. Midl. Nat. 1945;33(3):464–780. [Google Scholar]