

Abstract
Background
Edwardsiella ictaluri is a Gram-negative facultative intracellular anaerobe and the etiologic agent of enteric septicemia of channel catfish (ESC). To the catfish industry, ESC is a devastating disease due to production losses and treatment costs. Identification of virulence mechanisms of E. ictaluri is critical to developing novel therapeutic approaches for the disease. Here, we report construction of a transposon insertion library and identification of mutated genes in growth-delayed E. ictaluri colonies. We also provide safety and efficacy of transposon insertion mutants in catfish.
Results
An E. ictaluri transposon insertion library with 45,000 transposants and saturating 30.92% of the TA locations present in the E. ictaluri genome was constructed. Transposon end mapping of 250 growth-delayed E. ictaluri colonies and bioinformatic analysis of sequences revealed 56 unique E. ictaluri genes interrupted by the MAR2xT7 transposon, which are involved in metabolic and cellular processes and mostly localized in the cytoplasm or cytoplasmic membrane. Of the 56 genes, 30 were associated with bacterial virulence. Safety and vaccine efficacy testing of 19 mutants showed that mutants containing transposon insertions in hypothetical protein (Eis::004), and Fe-S cluster assembly protein (IscX, Eis::039), sulfurtransferase (TusA, Eis::158), and universal stress protein A (UspA, Eis::194) were safe and provided significant protection (p < 0.05) against wild-type E. ictaluri.
Conclusions
The results indicate that random transposon mutagenesis causing growth-delayed phenotype results in identification bacterial virulence genes, and attenuated strains with transposon interrupted virulence genes could be used as vaccine to activate fish immune system.
Keywords: pMAR2xT7, Enteric septicemia, Catfish, Virulence, Type III secretion system
Background
Enteric septicemia of catfish (ESC) is a devastating disease that causes significant production loss and treatment cost for the catfish aquaculture industry [1]. A few antimicrobials and a commercial live attenuated vaccine are available for treatment of ESC. However, treatment of sick catfish by medicated feed is not effective due to early onset of anorexia. The extensive use of antimicrobials can induce the appearance of resistant strains [2, 3]. The commercial ESC vaccine Aquavac-ESC has been available for the catfish industry for more than 15 years [4], but ESC is still one of the major diseases in the US catfish industry.
Edwardsiella ictaluri is well-adapted to channel catfish [5, 6] and some of the E. ictaluri virulence factors include lipopolysaccharide (LPS), flagella, outer membrane proteins (OMPs), and extracellular proteins [7–10]. There have been several reports on development of attenuated E. ictaluri strains by deleting genes involved in iron acquisition, tricarboxylic acid cycle, one-carbon metabolism, and amino acid biosynthesis [11–17]. However, virulence mechanisms of E. ictaluri are not understood well, and there is a need for identification of novel virulence-related genes to develop effective live attenuated vaccines.
Random transposon insertion is a high-throughput genetic manipulation tool that allows random mutation of genes at the genome level. Mariner family transposon Himar1 inserts itself randomly into “TA” nucleotide sequences [18, 19]. Mariner family transposons have been widely used to generate random mutagenesis in fish pathogen Mycobacterium marinum, and also human pathogens such as Pseudomonas aeruginosa, Campylobacter jejuni, Leptospira interrogans, and Rickettsia prowazekii [20–24].
In this research, MAR2xT7 transposon, a Himar1 derivative [20], was used to identify genes required for E. ictaluri growth on a solid complex medium. We expect that colonies exhibiting attenuated growth on solid media will have transposon insertions in important bacterial genes, and these mutants may also show attenuated virulence in the catfish host and potentiate catfish immune responses [14]. Therefore, attenuation and vaccine efficacy of 19 transposon mutants were evaluated in channel catfish.
Results
Transposon insertion library
By using MAR2xT7 transposon, an E. ictaluri transposon insertion library containing 45,000 transposants was constructed. Colonies with transposon insertion and delayed growth were observed on the BHI agar media after 48 h (Fig. 1). The initial overnight growth of these small colonies in BHI broth was also very slow compared to wild type, but this difference disappeared in later broth cultures (data not shown).
Fig. 1.
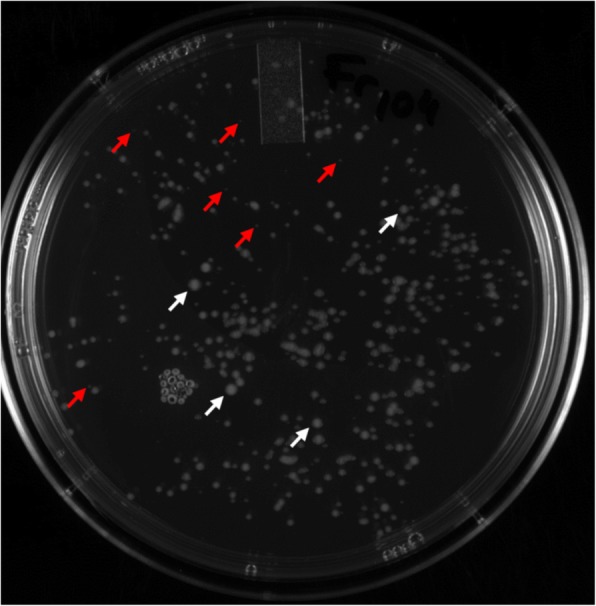
BHI agar plate showing transposon insertion colonies with delayed growth (red arrows) and normal size (white arrows) colonies after 48 h of incubation at 30 °C
The complete genome size of E. ictaluri strain 93–146 is 3,812,301 bp, which contains 3597 total genes. The number of TA locations in the entire E. ictaluri genome is 145,515. Thus, 45,000 transposants would saturate 30.92% of the potential MAR2xT7 transposon insertion sites available. The number of TA locations in the E. ictaluri open reading frames is 110,373, which represent 75.85% of all available MAR2xT7 transposon insertion sites. Thus, random insertion events would saturate 23,45% of the potential MAR2xT7 transposon insertion sites in the E. ictaluri open reading frames.
Gene identification
Transposon end amplification by single primer PCR yielded 151 samples with PCR products, of which 94 were sequenced successfully. After analysis, 56 unique genes containing transposon insertions were identified (Table 1). These unique genes contained a total number of 2235 MAR2xT7 transposon insertion sites, and the exact number of MAR2xT7 transposon insertion site in each gene was indicated in Table 1.
Table 1.
Unique E. ictaluri genes with transposon insertion
| Mutant | Locus | Product | E-valuea | Frequencyb | TA Frequency |
|---|---|---|---|---|---|
| Eis001 | NT01EI_1281 | NAD-dependent malic enzyme (NAD-ME) | 2.00E-72 | 64 | |
| Eis002 | NT01EI_1721 | PTS system, mannose/fructose/sorbose family, IIB component | 2.00E-106 | 32 | |
| Eis004 | NT01EI_0182 | Hypothetical protein | 7.00E-57 | 26 | |
| Eis006 | NT01EI_0085 | ATP-dependent DNA helicase Rep | 2.00E-114 | 46 | |
| Eis009 | NT01EI_1236 | Coproporphyrinogen III oxidase, aerobic | 6.00E-32 | 24 | |
| Eis011 | NT01EI_3690 | ABC transporter, periplasmic amino acid binding protein | 1.00E-74 | 50 | |
| Eis013 | NT01EI_2795 | Translocator protein, LysE family | 2.00E-115 | 36 | |
| Eis018 | NT01EI_0377 | Aspartate ammonia-lyase | 7.00E-145 | 59 | |
| Eis024 | NT01EI_3505 | Dihydrouridine synthase Dus | 1.00E-129 | 59 | |
| Eis027 | NT01EI_0408 | tRNA delta(2)-isopentenylpyrophosphate transferase | 5.00E-94 | 22 | |
| Eis028 | NT01EI_0277 | Transposase, IS4 family protein | 1.00E-73 | 4 | 17 |
| Eis029 | NT01EI_2683 | Membrane protein | 2.00E-88 | 27 | |
| Eis033 | pEI2_p2 | Putative Rep protein | 2.00E-123 | 5 | 31 |
| Eis035 | pEI1_p4 | Putative RNA one modulator protein | 9.00E-19 | 12 | |
| Eis037 | NT01EI_2355 | eseJ | 3.00E-129 | 2 | 88 |
| Eis038 | NT01EI_1334 | eseM | 5.00E-67 | 2 | 91 |
| Eis039 | NT01EI_3177 | FeS assembly protein IscX | 3.00E-38 | 7 | |
| Eis041 | NT01EI_0943 | eseC | 8.00E-116 | 53 | |
| Eis048 | NT01EI_3148 | Hypothetical protein | 4.00E-80 | 20 | |
| Eis055 | NT01EI_2314 | Prophage lambda integrase | 8.00E-128 | 56 | |
| Eis059 | NT01EI_1941 | Hypothetical protein | 3.00E-70 | 8 | |
| Eis065 | NT01EI_2281 | Excinuclease ABC subunit C | 3.00E-48 | 64 | |
| Eis068 | NT01EI_0448 | Polyprenyl synthetase | 1.00E-115 | 37 | |
| Eis080 | NT01EI_0981 | Hypothetical protein | 8.00E-129 | 9 | 58 |
| Eis086 | NT01EI_1237 | N-acetylmuramoyl-L-alanine amidase AmiA | 3.00E-67 | 25 | |
| Eis107 | NT01EI_0475 | DEAD box containing helicase | 1.00E-112 | 51 | |
| Eis110 | NT01EI_1332 | eseL | 4.00E-26 | 2 | 73 |
| Eis131 | NT01EI_2157 | Hypothetical protein | 4.00E-66 | 37 | |
| Eis152 | NT01EI_0224 | Transporter, major facilitator family | 1.00E-16 | 4 | 47 |
| Eis154 | NT01EI_3522 | Selenate reductase, FAD-binding subunit | 2.00E-73 | 23 | |
| Eis155 | NT01EI_0725 | Transcriptional regulator FruR | 3.00E-134 | 43 | |
| Eis156 | NT01EI_2381 | Ribonuclease, RNaseE/RNaseG family | 2.00E-04 | 72 | |
| Eis157 | NT01EI_0144 | Twin-arginine translocation protein subunit TatB | 8.00E-07 | 13 | |
| Eis158 | NT01EI_0022 | Sulfurtransferase, TusA | 8.00E-46 | 10 | |
| Eis171 | NT01EI_3723 | Magnesium-translocating P-type ATPase | 0 | 71 | |
| Eis172 | NT01EI_3786 | Hypothetical protein | 1.00E-26 | 24 | |
| Eis173 | NT01EI_3265 | Acyltransferase/AMP-dependent synthetase and ligase family | 0 | 2 | 54 |
| Eis174 | NT01EI_3721 | Hypothetical protein | 3.00E-25 | 7 | |
| Eis175 | NT01EI_3774 | IS1 transposase | 7.00E-93 | 19 | |
| Eis176 | NT01EI_0962 | esaT | 5.00E-129 | 27 | |
| Eis180 | NT01EI_3103 | UPF0126 domain protein | 7.00E-12 | 16 | |
| Eis183 | NT01EI_3105 | Chloride transporter, chloride channel (ClC) family | 6.00E-166 | 53 | |
| Eis184 | NT01EI_0419 | RNA methyltransferase, TrmH family, group 3 | 9.00E-27 | 21 | |
| Eis185 | NT01EI_3386 | TRAP transporter, DctM subunit | 7.00E-85 | 47 | |
| Eis192 | NT01EI_3147 | Hypothetical protein | 0 | 2 | 103 |
| Eis194 | NT01EI_1981 | Universal stress protein A uspA | 9.00E-86 | 3 | 20 |
| Eis195 | NT01EI_0376 | Anaerobic C4-dicarboxylate transporter DcuA | 2.00E-159 | 11 | 47 |
| Eis207 | NT01EI_1817 | Spermidine/putrescine transport system permease protein PotB | 4.00E-132 | 22 | |
| Eis210 | NT01EI_0800 | Prolipoprotein diacylglyceryl transferase | 5.00E-79 | 36 | |
| Eis220 | NT01EI_2076 | Hypothetical protein | 6.00E-20 | 5 | |
| Eis222 | NT01EI_0768 | Hypoxanthine phosphoribosyltransferase | 5.00E-127 | 24 | |
| Eis223 | NT01EI_1086 | Extracellular solute-binding protein, family 5 | 2.00E-142 | 48 | |
| Eis230 | NT01EI_3769 | Phosphoglycerate transporter family protein | 0 | 49 | |
| Eis232 | NT01EI_2010 | Hypothetical protein | 2.00E-136 | 3 | 58 |
| Eis233 | NT01EI_2530 | Putative permease, membrane region | 4.00E-81 | 65 | |
| Eis235 | NT01EI_3289 | Diaminopimelate decarboxylase | 9.00E-36 | 38 |
aBlastx E-value
bpMAR2xT7 insertion frequency
Functional annotation
Protein sequences of all 56 genes were annotated functionally and assigned to biological process (localization, cellular process, metabolic process, response to stimulus, biological regulation, signaling, multi-organism process, single-organism process, and biogenesis), cellular component (cell, macromolecular complex, and extracellular region), and molecular function (binding, transporter activity, catalytic activity, and nucleic acid binding transcription factor) (Fig. 2).
Fig. 2.

Gene ontology (GO) analysis of E. ictaluri transposon inserted genes. GO terms at level 2 according to biological process (a), GO terms at level 2 according to cellular component (b), GO terms at level 2 according to molecular function (c)
Subcellular localization
The locations of 15 proteins were unknown. Of the 41 proteins with known subcellular location, most were localized to the cytoplasm (20 proteins) and cytoplasmic membrane (16 proteins). Extracellular space, outer membrane, and periplasm contained very few proteins (3, 1, 1 proteins, respectively).
Proteins involved in host-pathogen interactions
Out of 56 identified unique proteins, 30 proteins had significant homology to Host-Pathogen Interaction Database (HPIDB) (Table 2). The proteins mostly matched to the Enterobacteriaceae (Yersinia pestis, Escherichia coli K12, Shigella flexneri), Francisellaceae (Francisella tularensis SCHU S4), and Bacillaceae families (Bacillus anthracis).
Table 2.
Genes involved in host-pathogen interactions
| Mutant | Accession Nu. | Protein | Organism | E-value |
|---|---|---|---|---|
| Eis152 | YP_019321.1 | Oxalate:formate antiporter, putative | Bacillus anthracis | 2.00E-20 |
| Eis155 | YP_017710.1 | Sugar-binding transcriptional regulator, LacI family | Bacillus anthracis | 2.00E-21 |
| Eis207 | YP_002347936.1 | Inner membrane permease T of sulfate/thiosulfate ABC transporter | Yersinia pestis | 1.00E-12 |
| Eis011 | YP_017492.1 | Amino acid ABC transporter, amino acid-binding protein | Bacillus anthracis | 1.00E-15 |
| Eis013 | NP_670988.1 | Putative threonine efflux protein | Yersinia pestis | 5.00E-11 |
| Eis171 | YP_002345523.1 | Putative cation transport protein | Yersinia pestis | 4.00E-88 |
| Eis223 | YP_002345598.1 | HTH-type transcriptional regulator SgrR | Yersinia pestis | 2.00E-46 |
| Eis176 | NP_857736.1 | Yop proteins translocation protein T | Yersinia pestis | 9.00E-31 |
| Eis107 | YP_022388.1 | ATP-dependent RNA helicase, DEAD/DEAH box family | Bacillus anthracis | 1.00E-84 |
| Eis024 | NP_842644.2 | tRNA-dihydrouridine synthase | Bacillus anthracis | 6.00E-62 |
| Eis110 | NP_858359.2 | E3 ubiquitin-protein ligase ipaH9.8 | Shigella flexneri | 6.00E-90 |
| Eis086 | NP_667964.1 | N-acetylmuramoyl-l-alanine amidase II | Yersinia pestis | 1.00E-39 |
| Eis184 | YP_016695.1 | RNA methyltransferase, TrmH family, group 3 | Bacillus anthracis | 6.00E-43 |
| Eis180 | YP_002345138.1 | Putative membrane protein | Yersinia pestis | 4.00E-20 |
| Eis006 | YP_170066.1 | ATP-dependent DNA helicase | Francisella tularensis | 1.00E-171 |
| Eis009 | YP_170044.1 | Coproporphyrinogen-III oxidase, aerobic | Francisella tularensis | 9.00E-52 |
| Eis027 | YP_169650.1 | tRNA dimethylallyltransferase | Francisella tularensis | 8.00E-76 |
| Eis041 | YP_002345337.1 | Possible type III secretion protein | Yersinia pestis | 4.00E-154 |
| Eis173 | NP_994169.1 | Bifunctional protein aas | Yersinia pestis | 0 |
| Eis235 | YP_002345851.1 | Diaminopimelate decarboxylase | Yersinia pestis | 7.00E-177 |
| Eis156 | NP_669066.1 | RNase E | Yersinia pestis | 0 |
| Eis183 | NP_668136.1 | H(+)/Cl(−) exchange transporter ClcA | Yersinia pestis | 0 |
| Eis065 | NP_669748.1 | UvrABC system protein C | Yersinia pestis | 0 |
| Eis002 | YP_002346757.1 | PTS enzyme IIAB, mannose-specific | Yersinia pestis | 3.00E-149 |
| Eis222 | YP_646612.1 | Hypoxanthine phosphoribosyltransferase | Yersinia pestis | 6.00E-84 |
| Eis068 | YP_491372.1 | Octaprenyl-diphosphate synthase | Escherichia coli | 2.00E-157 |
| Eis001 | YP_002346527.1 | NAD-dependent malic enzyme | Yersinia pestis | 0 |
| Eis230 | YP_001608410.1 | Putative regulatory protein | Yersinia pestis | 0 |
| Eis018 | NP_667943.1 | Aspartate ammonia-lyase (Aspartase) | Yersinia pestis | 0 |
| Eis233 | YP_002346351.1 | Putative transport protein YPO1326/y2857/YP_1266 | Yersinia pestis | 0 |
Proteins involved in bacterial virulence
Out of 56 unique proteins, 30 matched significantly to known virulence-associated proteins from other Gram-negative and Gram-positive pathogenic bacteria in MVirDB (Table 3).
Table 3.
Genes involved in bacterial virulence
| Mutant | Locus | Number of hits | Lowest E-value | Protein | Location |
|---|---|---|---|---|---|
| Eis041 | NT01EI_0943 | 250 | 0 | esaC | Outer Membrane |
| Eis110 | NT01EI_1332 | 91 | 0 | eseL | Extracellular |
| Eis171 | NT01EI_3723 | 22 | 0 | Magnesium-translocating P-type ATPase | Cytoplasmic Membrane |
| Eis192 | NT01EI_3147 | 10 | 0 | Hypothetical protein | Unknown |
| Eis037 | NT01EI_2355 | 192 | 9.13E-144 | eseJ | Extracellular |
| Eis038 | NT01EI_1334 | 267 | 3.95E-121 | eseM | Extracellular |
| Eis001 | NT01EI_1281 | 6 | 1.97E-95 | NAD-dependent malic enzyme (NAD-ME) | Cytoplasmic |
| Eis195 | NT01EI_0376 | 5 | 8.13E-87 | Anaerobic C4-dicarboxylate transporter DcuA | Cytoplasmic Membrane |
| Eis173 | NT01EI_3265 | 221 | 8.89E-61 | Acyltransferase/AMP-dependent synthetase and ligase protein family | Cytoplasmic Membrane |
| Eis185 | NT01EI_3386 | 5 | 1.74E-56 | TRAP transporter DctM subunit | Cytoplasmic Membrane |
| Eis107 | NT01EI_0475 | 11 | 2.34E-56 | DEAD box containing helicase | Cytoplasmic |
| Eis230 | NT01EI_3769 | 250 | 1.16E-46 | Phosphoglycerate transporter family protein | Cytoplasmic Membrane |
| Eis028 | NT01EI_0277 | 8 | 4.26E-43 | Transposase, IS4 family protein | Unknown |
| Eis175 | NT01EI_3774 | 34 | 6.93E-41 | IS1 transposase | Unknown |
| Eis184 | NT01EI_0419 | 8 | 2.71E-40 | RNA methyltransferase TrmH family, group 3 | Cytoplasmic |
| Eis176 | NT01EI_0962 | 144 | 9.04E-38 | esaT | Cytoplasmic Membrane |
| Eis157 | NT01EI_0144 | 250 | 4.66E-29 | Twin-arginine translocation protein subunit TatB | Cytoplasmic Membrane |
| Eis068 | NT01EI_0448 | 2 | 4.13E-26 | Polyprenyl synthetase | Cytoplasmic |
| Eis155 | NT01EI_0725 | 9 | 4.05E-12 | Transcriptional regulator FruR | Cytoplasmic |
| Eis086 | NT01EI_1237 | 4 | 9.61E-12 | N-acetylmuramoyl-L-alanine amidase AmiA | Unknown |
| Eis006 | NT01EI_0085 | 40 | 9.81E-12 | ATP-dependent DNA helicase Rep | Cytoplasmic |
| Eis235 | NT01EI_3289 | 20 | 2.44E-10 | Diaminopimelate decarboxylase | Cytoplasmic |
| Eis055 | NT01EI_2314 | 22 | 7.18E-08 | Prophage lambda integrase | Cytoplasmic |
| Eis207 | NT01EI_1817 | 59 | 1.13E-07 | Spermidine/putrescine transport system permease protein PotB | Cytoplasmic Membrane |
| Eis080 | NT01EI_0981 | 10 | 1.52E-06 | Hypothetical protein | Cytoplasmic |
| Eis222 | NT01EI_0768 | 44 | 1.67E-06 | Hypoxanthine phosphoribosyltransferase | Cytoplasmic |
| Eis018 | NT01EI_0377 | 4 | 1.96E-06 | Aspartate ammonia-lyase | Cytoplasmic |
| Eis011 | NT01EI_3690 | 9 | 2.98E-04 | ABC transporter, periplasmic amino acid binding protein | Periplasmic |
| Eis131 | NT01EI_2157 | 21 | 9.55E-03 | Hypothetical protein | Unknown |
| Eis223 | NT01EI_1086 | 16 | 9.91E-03 | Extracellular solute-binding protein, family 5 | Unknown |
Safety and vaccine efficacy of mutants in catfish
Safety testing of transposon-derived mutants showed that all mutant strains were attenuated significantly compared to wild-type control (p < 0.05). Eis::004, Eis::039, Eis::041, Eis::176, and Eis::194 caused less than 5% mortality, while mutants, Eis::110, Eis::158, and Eis::195 caused less than 10% mortality (Fig. 3a). We noticed that mutants with gene products assigned to cytoplasm caused lower mortalities compared to those found in outer membrane or extracellular regions. Interestingly, proteins with unknown location (Eis::194 and Eis::039), did not show any mortalities in channel catfish. Mutants Eis::002, Eis::011, Eis::029, Eis::037, Eis::038, Eis::065, Eis::080, Eis::086, Eis::157, Eis::173, and Eis::232 were attenuated but caused over 20% mortality.
Fig. 3.

Virulence and efficacy of the transposon insertion mutants in channel catfish fingerlings. Percent mortalities and mutant names are indicated on the Y and X axis, respectively. Channel catfish fingerlings were infected with mutant strains to determine virulence and vaccinate the fish (a). After 21-days of post vaccination, fish were infected with wild-type E. ictaluri 93–146 to determine the efficacy of vaccination (b)
Edwardsiella ictaluri wild-type challenge of catfish 21 days post vaccination indicated that all mutants except Eis::041 and Eis::176 protected catfish significantly compared to sham vaccinated group (p < 0.05). Eis::004 and Eis::194 were the safest and most protective mutants, while Eis::157 protective but not safe (Fig. 3b). Although Eis::039, Eis::041, and Eis::176 were safe, they did not provide good immunization in catfish against E. ictaluri WT (Fig. 3).
Discussion
The bioinformatics analyses of 56 unique genes with transposon insertions showed that more than half (54%) were potential virulence factors in other pathogenic bacteria. Among the virulence factors, Type III secretion system (T3SS), twin-arginine translocation pathway (Tat), and ATP-binding cassette transporter (ABC) seem to be important for E. ictaluri virulence and invasion of the channel catfish [25–27].
The functional gene ontology analysis with Blast2GO indicated that most of the proteins participate in cellular and metabolic networks in the biological process while their molecular functions frequently matched to binding and catalytic activity. The proteins located in extracellular regions are part of the signaling process or response to any stimulus sensing bacteria in the biological process. Although several proteins showed transporter activity, these proteins account for localization and are in the cytoplasm.
The subcellular locations predicted by PSORTb revealed that most of the identified proteins are found in the cytoplasm and cytoplasmic membrane. Although many well-known virulence proteins are located in the outer membrane or periplasm in Gram-negative bacteria, only three proteins of T3SS are located in extracellular space and outer membrane, and one of the ABC transporter proteins was found in the periplasmic space.
The host-pathogen interaction examined by HPIDB proved that many proteins have a high similarity to other virulence-associated proteins in different pathogenic bacteria including Y. pestis, S. flexneri, and B. anthracis. The pathogenic Gram-negative bacteria Y. pestis and S. flexneri share the same evolutionary lineage with Edwardsiella sp. in Enterobacteriaceae [28]. Thus, most of the virulence-associated proteins may have a similar role in E. ictaluri. Two T3SS effector proteins EseJ and EseM have a predicted interaction with the channel catfish ubiquitin-conjugating enzyme E2 (XP_017323313). These two T3SS-related effector proteins known as E3 ubiquitin ligase play an important role in manipulation of host ubiquitination pathways.
The interrupted genes eseJ (Eis::037), eseM (Eis::038), esaC (Eis::041), eseL (Eis::110), and esaT (Eis::176) are part of T3SS, which are involved in export of proteins inside the host immune cells [29]. EsaC and EsaT are the structural membrane associated proteins of T3SS. Eis::41, YscC ring-shaped structure protein in the outer membrane, is required for a stable oligomeric complex to shape a T3SS in the outer membrane [30]. YscT inner membrane-embedded component is located in the cytoplasm, which has extended and helical regions that may form membrane-bound subunits. Insertions in the T3SS related genes, eseJ, eseM, and eseL, have been recently identified to be T3SS dependent effector proteins [31]. EseJ, EseL, and EseM proteins share high similarities with Salmonella T3SS effector proteins Ssph2 and SlrP. They are involved in ubiquitination of proteins, an important process regulating inflammatory response in eukaryotes. As a part of novel E3 ligases (NELs) protein family, EseJ, E3 ubiquitin-protein ligase (SspH2), and EseL, a new class of E3 ubiquitin ligase, have a role in T3SS that provides a strategy to exploit host cell ubiquitin pathway [32]. EseM, T3SS leucine rich repeat protein (SlrP), is also required to form a complex ubiquitin ligase enzyme [33]. T3SS effector protein mutants eseJ, eseM, and eseL, and T3SS structural mutants esaC and esaT showed significantly decreased virulence. However, in comparison of protection level of those two main groups, T3SS structural proteins EsaC and EsaT have been caused less protection in catfish. Mutation in T3SS effector proteins provides better protection against pathogenic bacteria [34–36]. EseL has provided significant protection among other T3SS related effector proteins. T3SS effector proteins could contribute the bacterial survival inside host immune cells [37, 38].
Transport processes in bacterial cells through outer membrane and periplasmic space are linked to E. ictaluri metabolism to survive in the host environment as well as switching between various biochemical processes during different stages of ESC. Eis::157, tatB, is located in the periplasmic space and is involved in the translocation of proteins including the components of respiratory complexes using a proton gradient as an energy source [39]. tatB mutant exhibited slow growth under low-iron conditions and observed a 10-fold decrease in Legionella pneumophila growth [40]. Eis::086, amiA, is a Tat pathway dependent substrate encoding a cell wall amidase. Tat pathway mutant causes mislocalization of AmiA protein, preventing translocation in the periplasm [41, 42]. Eis::011, ABC transporter periplasmic amino acid binding protein, is an important antigenic factor involved in adhesion and aspartate/glutamate transport in the microaerobic environment in Campylobacter jejuni [43]. Eis::207, potB, encodes a protein associated with spermidine/putrescine transport system. Polyamines are mostly involved in stabilization of DNA for stress resistance, intracellular signaling processes, and swarming motility [44, 45]. Polyamines are also associated with the virulence in the intracellular pathogen Salmonella enterica [46]. Eis::002, PTS system IIB component, is a cytoplasmic component of the major carbohydrate transport system highly conserved through bacteria [47]. PTS system participates in a variety of virulence mechanisms including biofilm formation, modulating the virulence gene expression, and regulating carbohydrate metabolism in pathogenic bacteria [48–50]. Eis::171, magnesium-translocating P-type ATPase, is an inducible magnesium transport system when bacteria grow at the low concentration of magnesium. Although Mg2+ is not essential for virulence, it participates in many cellular activities as a cofactor [51]. Magnesium is the part of the regulatory network that regulates the virulence-associated mechanisms in S. enterica [52]. Eis::195, dcuA, is encoded with aspartase in the same operon that is determined as an antiporter mechanism involved in the transport of aspartate under the anaerobic conditions [53]. DcuA function in the metabolic pathway under anaerobic conditions contributes the pathogenicity for the colonization in the lower oxygen level [54].
Pathogenic bacteria adapted different carbohydrate metabolism, which is activated by oxygen presence in the host environment. Eis::018, aspartate ammonia-lyase, is involved in the production of fumarate activated specifically under anaerobic conditions while there is no available electron acceptor. Bacteria encodes aspartate ammonia-lyase to utilize alternative carbon sources in the host environment if there are no available carbon sources [55, 56].
Bacterial stress related proteins induce the protective mechanisms under a variety of stress conditions to protect the bacterial cell inside or outside of the host [57]. Universal stress protein A (UspA) in Eis::194, is one of the stress proteins found in intracellular pathogenic bacteria. uspA expression reaches a high level when bacteria are exposed to heat, starvation, antimicrobial, and oxidative agents [58, 59]. UspA is a conserved protein that presents in Eubacteria, Archaea, plants, and fungi and the expression of UspA is triggered by exposure to oxidative agents in growth arrested cells [60–62]. UspA plays a significant role in the pathogenicity of bacteria, and uspA mutants are less virulent and sensitive to changes in the host environment. Mutation of S. typhimurium C5 uspA resulted in less virulence and more susceptibility to nutrient starvation oxidative agents [59]. In Listeria monocytogenes, uspA mutants were shown to have impaired activity in oxidative agent’s exposure to low pH conditions [58]. Deletion of uspA gene in Acinetobacter baumannii revealed that it has a significant role in protecting the bacteria from H2O2 and low pH [63].
IscX in Eis::S039, acts as a regulator for the Fe-S (iron-sulfur) cluster, which encodes proteins essential for cell activities [64]. FeS assembly protein IscX (YfhJ) is a part of the iron-sulfur cluster (ISC) mediated FeS cluster, which is a small acidic protein that binds IscC and Fe, and acts as a Fe donor in FeS cluster [65, 66]. ISC mediated FeS biogenesis is involved in survival of bacteria that face with iron starvation and oxidative stress. In S. flexneri, ISC mutants were less invasive and cannot form plaques on Henle cells monolayers [67]. ISC transcriptional regulator iscR mutant in Pseudomonas aeruginosa caused more susceptibility to oxidative agents and a significant decrease in virulence [68]. The importance of ISC system in bacterial virulence has been emphasized in different studies. However, limited information is known about the role of IscX in bacterial virulence.
Hypothetical protein in Eis::004 is located in the cytoplasm. There is no available information about the function of this hypothetical protein in any virulence related mechanisms. However, decreased virulence and significant protection against ESC revealed that Eis::004 mutant could be considered as a vaccine candidate for live attenuated vaccine development.
Conclusions
In summary, these results showed that random transposon mutagenesis in the E. ictaluri genome resulted in colonies with delayed growth on complex solid media, and many of the disrupted genes have important functions and potentially contribute to E. ictaluri virulence. Fish experiments showed that Eis::004, Eis::039, Eis::041, Eis::110, Eis::158, Eis::176, Eis::194, and Eis::195 mutants were significantly attenuated, and Eis::004 and Eis::194 provided good immunization in catfish.
Methods
Bacterial strains, plasmids, and growth conditions
Bacterial strains and plasmids used in this work are listed in Table 4. Edwardsiella ictaluri 93–146 carrying pAKgfplux1 [69] was grown at 30 °C using brain heart infusion (BHI) broth and agar plates (Difco, Sparks, MD). Escherichia coli SM10λpir donor strain carrying pMAR2xT7 [20] was grown at 37 °C using Luria-Bertani (LB) broth and agar plates (Difco). Antibiotics were added to the culture medium at the following concentrations: ampicillin (100 μg/ml), colistin (12.5 μg/ml), and gentamicin (12.5 μg/ml).
Table 4.
Bacterial strains and plasmids
Construction of transposon insertion library
Transposon insertion library was constructed by conjugation using the donor E. coli SM10λpir carrying pMAR2xT7 and the recipient E. ictaluri 93–146 wild type (WT) containing pAKgfplux1 [69]. Transposon insertion mutants were selected on selective BHI agar plates containing 100 μg/ml of ampicillin, 12.5 μg/ml of gentamicin, and 25 μg/ml colistin. Various sizes of gentamicin resistant transposon insertion colonies were observed on the selective BHI plates, and 250 smallest colonies compared to normal colony size were cultured in the BHI broth with colistin and gentamicin at 30 °C for 2 days. Finally, bacterial stocks were prepared in 20% glycerol and stored at − 80 °C freezer.
Transposon end mapping
Genomic DNA was isolated from the frozen E. ictaluri transposon insertion mutants using the heat denaturation method. Briefly, 100 μl frozen culture were added in 1 ml ddH2O and mixed well. Bacteria were collected by centrifugation and water was removed completely. After dissolving the bacterial pellet in 100 μl ddH2O, each sample was transferred to 200 μl PCR tubes and tubes were incubated at 100 °C for 10 min by using an Applied Biosystems 2720 Thermal Cycler (Life Technologies, Grand Island, NY). Samples were mixed well by vortexing, and bacterial cell debris was pelleted by centrifuging at 14,000 rpm for 5 min. The supernatant containing the genomic DNA was used as template in subsequent PCR reactions. Single primer PCR was performed by using a transposon-specific R1 primer (5`-CCGTATGCCCAACTTTGTATAGA-3`) to amplify the transposon end and flanking bacterial DNA [70]. Before sequencing, the PCR products were cleaned by using ExoSAP-IT for PCR Product Cleanup (Affymetrix, Santa Clara, CA). Sequencing was conducted at Eurofins MWG Operon LLC (Huntsville, AL) using a transposon-specific nested R3 primer (5`- TCTCGGCTTGAACGAATTGTT-3`).
Bioinformatics analyses
Transposon sequence removal and sequence trimming based on sequence quality scores were done by using the Sequencher DNA sequence analysis software v4.10.1 (Gene Codes Corp., Ann Arbor, MI). Trimmed sequences were searched against the available E. ictaluri 93–146 genome [29] by using basic local alignment search tool (Blast) at the National Center for Biotechnology Information (NCBI) for gene identification. Using the GI numbers, a FASTA file containing all protein sequences were downloaded from the Batch Entrez database of NCBI and used for downstream analysis. Gene Ontology (GO) annotation, visualization, and metabolic and cellular processes were determined by using Blast2GO [71] at the cut-off level 2. Subcellular localization of proteins was predicted by using PSORTb version 3.0.2 [72]. E. ictaluri proteins involving in host-pathogen interactions were identified by using the Host-Pathogen Interaction Database (HPIDB) at the cut-off level 0.0001. Bacterial proteins interacting with channel catfish proteins were determined at the cut-off level 0.00001, at identity filter 50% in bacterial proteins, and 70% in channel catfish proteins. [73]. The potential E. ictaluri virulence proteins were identified using the Microbial Virulence Database (MVirDB) at the cut-off level 0.5 [74]. TA sequence frequencies in the entire E. ictaluri genome, open reading frames, and genes with transposon insertion were calculated using CLC genomics workbench 11.0.1 (Qiagen, Redwood City, CA).
Safety and vaccine efficacy testing of mutants in catfish
Specific pathogen free (SPF) channel catfish was obtained from the fish hatchery of the College of Veterinary Medicine at Mississippi State University. All fish experiments were conducted under a protocol approved by the Institutional Animal Care and Use Committee at Mississippi State University (protocol number 12–042). In vivo experiments were conducted using catfish infection model to test 19 mutants. Briefly, four-month-old pathogen free channel catfish (11.58 ± 0.23 cm, 15.29 ± 0.95 g) were stocked at a rate of 20 fish/tank into 40 L tanks and maintained at 26 ± 2 °C throughout the experiment. Each transposon mutant, positive (E. ictaluri wild-type), and negative (BHI) controls were assigned to three or four tanks randomly. Catfish were challenged/vaccinated by immersion exposure using transposon mutants or wild type (3.09 × 107 CFU/ml of water) using published procedures [15]. Catfish mortalities were recorded for 21 days. After 21 days of the first vaccination, both vaccinated, and sham-vaccinated catfish were infected with E. ictaluri wild type by immersion exposure (3.27 × 107 CFU/ml of water). Catfish mortalities were recorded for two weeks.
Statistical analysis
We used SPSS V25 (IBM Corp., Armonk, NY) to conduct statistical analysis. For each strain, mean percent mortalities were calculated and arcsine-transformed. The one-way analysis of variance at significance level 0.05 was conducted using the “Univariate” function, in which strains were independent and arcsine-transformed mortalities were dependent variables. Because our data included different sample sizes, and variances were not equal, Games-Howell post hoc test was selected to identify significant differences between mutants and wild type or mutants and sham vaccinated group in virulence and efficacy experiments, respectively.
Acknowledgements
We thank the Laboratory Animal Resources and Care at the College of Veterinary Medicine for providing the specific pathogen free channel catfish.
Funding
This project was supported by Agriculture and Food Research Initiative competitive grant no. 2014–70007-22359 from the USDA National Institute of Food and Agriculture. The funding body did not have any role in the design of the study and collection, analysis, and interpretation of data and in writing the manuscript.
Availability of data and materials
The datasets used and/or analyzed during the current study are available from the corresponding author on reasonable request.
Abbreviations
- ESC
Enteric septicemia of catfish
- HPIDB
Host-Pathogen Interaction Database
- IACUC
Institutional Animal Care and Use Committee
- MVirDB
Microbial Virulence Database
- PSORTb
Subcellular Localization Prediction Tool
- T3SS
Type III Secretion System
- UspA
Universal Stress Protein A
Authors’ contributions
Conception and design of the study: MLL, AK. Performed experiments: SK, JL, HA, HCT. Analyzed data: SK. Wrote the manuscript: SK, MLL, AK. All authors read and approved the final manuscript.
Ethics approval and consent to participate
All fish experiments were conducted under a protocol approved by the Institutional Animal Care and Use Committee at Mississippi State University (protocol number 12–042).
Consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing interests.
Publisher’s Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Contributor Information
Safak Kalindamar, Email: sk1047@msstate.edu.
Jingjun Lu, Email: jl676@msstate.edu.
Hossam Abdelhamed, Email: abdelhamed@cvm.msstate.edu.
Hasan C. Tekedar, Email: hct37@msstate.edu
Mark L. Lawrence, Email: lawrence@cvm.msstate.edu
Attila Karsi, Email: karsi@cvm.msstate.edu.
References
- 1.Highlights of NAHMS catfish 2003: part II. USDA APHIS. 2003.
- 2.Hawke JP, Durborow RM, Thune RL, Camus AC. ESC-Enteric Septicemia of Catfish. Southern Regional Aquaculture Center Publication No. 477. 1998.
- 3.Tu TD, Haesebrouck F, Nguyen AT, Sorgeloos P, Baele M, Decostere A. Antimicrobial susceptibility pattern of Edwardsiella ictaluri isolates from natural outbreaks of bacillary necrosis of Pangasianodon hypophthalmus in Vietnam. Microb Drug Resist. 2008;14(4):311–316. doi: 10.1089/mdr.2008.0848. [DOI] [PubMed] [Google Scholar]
- 4.Klesius PH, Shoemaker CA. Development and use of modified live Edwardsiella ictaluri vaccine against enteric septicemia of catfish. Adv. Vet. Med. 1999;41:523–537. doi: 10.1016/S0065-3519(99)80039-1. [DOI] [PubMed] [Google Scholar]
- 5.Booth NJ, Elkamel A, Thune RL. Intracellular replication of Edwardsiella ictaluri in channel catfish macrophages. J Aquat Anim Health. 2006;18(2):101–108. doi: 10.1577/H05-025.1. [DOI] [Google Scholar]
- 6.Ainsworth AJ, Chen DX. Differences in the phagocytosis of four bacteria by channel catfish neutrophils. Dev Comp Immunol. 1990;14(2):201–209. doi: 10.1016/0145-305X(90)90091-R. [DOI] [PubMed] [Google Scholar]
- 7.Lawrence ML, Banes MM, Williams ML. Phenotype and virulence of a transposon-derived lipopolysaccharide O side-chain mutant strain of Edwardsiella ictaluri. J Aquat Anim Health. 2001;13(4):291–299. doi: 10.1577/1548-8667(2001)013<0291:PAVOAT>2.0.CO;2. [DOI] [Google Scholar]
- 8.Newton JC, Triche PL. Isolation and characterization of flagella from Edwardsiella ictaluri. J Aquat Anim Health. 1993;5(1):16–22. doi: 10.1577/1548-8667(1993)005<0016:IACOFF>2.3.CO;2. [DOI] [Google Scholar]
- 9.Bader JA, Shoemaker CA, Klesius PH. Immune response induced by N-lauroylsarcosine extracted outer-membrane proteins of an isolate of Edwardsiella ictaluri in channel catfish. Fish & shellfish immunology. 2004;16(3):415–428. doi: 10.1016/j.fsi.2003.07.003. [DOI] [PubMed] [Google Scholar]
- 10.Klesius PH, Sealey WM. Chemotactic and Chemokinetic responses of channel catfish macrophages to exoantigen from Edwardsiella ictaluri. J Aquat Anim Health. 1996;8(4):314–318. doi: 10.1577/1548-8667(1996)008<0314:CACROC>2.3.CO;2. [DOI] [Google Scholar]
- 11.Lawrence ML, Cooper RK, Thune RL. Attenuation, persistence, and vaccine potential of an Edwardsiella ictaluri purA mutant. Infect Immun. 1997;65(11):4642–4651. doi: 10.1128/iai.65.11.4642-4651.1997. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 12.Thune RL, Fernandez DH, Battista JR. An aroA mutant of Edwardsiella ictaluri is safe and efficacious as a live, attenuated vaccine. J Aquat Anim Health. 1999;11(4):358–372. doi: 10.1577/1548-8667(1999)011<0358:AAMOEI>2.0.CO;2. [DOI] [Google Scholar]
- 13.Lawrence ML, Banes MM. Tissue persistence and vaccine efficacy of an O polysaccharide mutant strain of Edwardsiella ictaluri. J Aquat Anim Health. 2005;17(3):228–232. doi: 10.1577/H04-049.1. [DOI] [Google Scholar]
- 14.Karsi A, Gulsoy N, Corb E, Dumpala PR, Lawrence ML. High-throughput bioluminescence-based mutant screening strategy for identification of bacterial virulence genes. Appl Environ Microbiol. 2009;75(7):2166–2175. doi: 10.1128/AEM.02449-08. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 15.Abdelhamed H, Lu J, Shaheen A, Abbass A, Lawrence ML, Karsi A. Construction and evaluation of an Edwardsiella ictaluri fhuC mutant. Vet Microbiol. 2013;162(2–4):858–865. doi: 10.1016/j.vetmic.2012.11.006. [DOI] [PubMed] [Google Scholar]
- 16.Dahal N, Abdelhamed H, Lu J, Karsi A, Lawrence ML. Tricarboxylic acid cycle and one-carbon metabolism pathways are important in Edwardsiella ictaluri virulence. PLoS One. 2013;8(6):e65973. doi: 10.1371/journal.pone.0065973. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 17.Santander J, Golden G, Wanda SY, Curtiss R., 3rd Fur-regulated iron uptake system of Edwardsiella ictaluri and its influence on pathogenesis and immunogenicity in the catfish host. Infect Immun. 2012;80(8):2689–2703. doi: 10.1128/IAI.00013-12. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 18.Akerley BJ, Rubin EJ, Camilli A, Lampe DJ, Robertson HM, Mekalanos JJ. Systematic identification of essential genes by in vitro mariner mutagenesis. Proc Natl Acad Sci U S A. 1998;95(15):8927–8932. doi: 10.1073/pnas.95.15.8927. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 19.Rubin EJ, Akerley BJ, Novik VN, Lampe DJ, Husson RN, Mekalanos JJ. In vivo transposition of mariner-based elements in enteric bacteria and mycobacteria. Proc Natl Acad Sci U S A. 1999;96(4):1645–1650. doi: 10.1073/pnas.96.4.1645. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 20.Liberati NT, Urbach JM, Miyata S, Lee DG, Drenkard E, Wu G, Villanueva J, Wei T, Ausubel FM. An ordered, nonredundant library of Pseudomonas aeruginosa strain PA14 transposon insertion mutants. Proc Natl Acad Sci U S A. 2006;103(8):2833–2838. doi: 10.1073/pnas.0511100103. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 21.Hendrixson DR, Akerley BJ, DiRita VJ. Transposon mutagenesis of campylobacter jejuni identifies a bipartite energy taxis system required for motility. Mol Microbiol. 2001;40(1):214–224. doi: 10.1046/j.1365-2958.2001.02376.x. [DOI] [PubMed] [Google Scholar]
- 22.Murray GL, Morel V, Cerqueira GM, Croda J, Srikram A, Henry R, Ko AI, Dellagostin OA, Bulach DM, Sermswan RW, et al. Genome-wide transposon mutagenesis in pathogenic Leptospira species. Infect Immun. 2009;77(2):810–816. doi: 10.1128/IAI.01293-08. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 23.Liu ZM, Tucker AM, Driskell LO, Wood DO. Mariner-based transposon mutagenesis of rickettsia prowazekii. Appl Environ Microbiol. 2007;73(20):6644–6649. doi: 10.1128/AEM.01727-07. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 24.Gao LY, Groger R, Cox JS, Beverley SM, Lawson EH, Brown EJ. Transposon mutagenesis of Mycobacterium marinum identifies a locus linking pigmentation and intracellular survival. Infect Immun. 2003;71(2):922–929. doi: 10.1128/IAI.71.2.922-929.2003. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 25.Galán JE, Collmer A. Type III secretion machines: bacterial devices for protein delivery into host cells. Science. 1999;284(5418):1322–1328. doi: 10.1126/science.284.5418.1322. [DOI] [PubMed] [Google Scholar]
- 26.Ochsner UA, Snyder A, Vasil AI, Vasil ML. Effects of the twin-arginine translocase on secretion of virulence factors, stress response, and pathogenesis. Proc Natl Acad Sci U S A. 2002;99(12):8312–8317. doi: 10.1073/pnas.082238299. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 27.Davidson AL, Chen J. ATP-binding cassette transporters in bacteria. Annu Rev Biochem. 2004;73:241–268. doi: 10.1146/annurev.biochem.73.011303.073626. [DOI] [PubMed] [Google Scholar]
- 28.Yang M, Lv Y, Xiao J, Wu H, Zheng H, Liu Q, Zhang Y, Wang Q. Edwardsiella comparative phylogenomics reveal the new intra/inter-species taxonomic relationships, virulence evolution and niche adaptation mechanisms. PLoS One. 2012;7(5):e36987. doi: 10.1371/journal.pone.0036987. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 29.Williams ML, Gillaspy AF, Dyer DW, Thune RL, Waldbieser GC, Schuster SC, Gipson J, Zaitshik J, Landry C, Banes MM, et al. Genome sequence of Edwardsiella ictaluri 93-146, a strain associated with a Natural Channel catfish outbreak of enteric septicemia of catfish. J Bacteriol. 2012;194(3):740–741. doi: 10.1128/JB.06522-11. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 30.Koster M, Bitter W, de Cock H, Allaoui A, Cornelis GR, Tommassen J. The outer membrane component, YscC, of the Yop secretion machinery of Yersinia enterocolitica forms a ring-shaped multimeric complex. Mol Microbiol. 1997;26(4):789–797. doi: 10.1046/j.1365-2958.1997.6141981.x. [DOI] [PubMed] [Google Scholar]
- 31.Dubytska LP, Rogge ML, Thune RL. Identification and Characterization of Putative Translocated Effector Proteins of the Edwardsiella ictaluri Type III Secretion System. mSphere. 2016;1(3):e00039–16. [DOI] [PMC free article] [PubMed]
- 32.Hicks SW, Galan JE. Hijacking the host ubiquitin pathway: structural strategies of bacterial E3 ubiquitin ligases. Curr Opin Microbiol. 2010;13(1):41–46. doi: 10.1016/j.mib.2009.11.008. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 33.Bernal-Bayard J, Cardenal-Munoz E, Ramos-Morales F. The Salmonella type III secretion effector, salmonella leucine-rich repeat protein (SlrP), targets the human chaperone ERdj3. J Biol Chem. 2010;285(21):16360–16368. doi: 10.1074/jbc.M110.100669. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 34.Haque A, Chu K, Easton A, Stevens MP, Galyov EE, Atkins T, Titball R, Bancroft GJ. A live experimental vaccine against Burkholderia pseudomallei elicits CD4+ T cell-mediated immunity, priming T cells specific for 2 type III secretion system proteins. J Infect Dis. 2006;194(9):1241–1248. doi: 10.1086/508217. [DOI] [PubMed] [Google Scholar]
- 35.Farfán MJ, Toro CS, Barry EM, Nataro JP. Shigella enterotoxin-2 is a type III effector that participates in Shigella-induced interleukin 8 secretion by epithelial cells. FEMS Immunol. Med. Microbiol. 2011;61(3):332–339. doi: 10.1111/j.1574-695X.2011.00778.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 36.Zhang Y, Bliska JB. YopJ-promoted cytotoxicity and systemic colonization are associated with high levels of murine interleukin-18, gamma interferon, and neutrophils in a live vaccine model of Yersinia pseudotuberculosis infection. Infect Immun. 2010;78(5):2329–2341. doi: 10.1128/IAI.00094-10. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 37.Figueira R, Watson KG, Holden DW, Helaine S. Identification of salmonella pathogenicity island-2 type III secretion system effectors involved in intramacrophage replication of S. enterica serovar typhimurium: implications for rational vaccine design. mBio. 2013;4(2):e00065. doi: 10.1128/mBio.00065-13. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 38.Whitlock GC, Estes DM, Young GM, Young B, Torres AG. Construction of a reporter system to study Burkholderia mallei type III secretion and identification of the BopA effector protein function in intracellular survival. Trans R Soc Trop Med Hyg. 2008;102(Suppl 1):S127–S133. doi: 10.1016/S0035-9203(08)70029-4. [DOI] [PubMed] [Google Scholar]
- 39.Muller M. Twin-arginine-specific protein export in Escherichia coli. Res Microbiol. 2005;156(2):131–136. doi: 10.1016/j.resmic.2004.09.016. [DOI] [PubMed] [Google Scholar]
- 40.Rossier O, Cianciotto NP. The Legionella pneumophila tatB gene facilitates secretion of phospholipase C, growth under iron-limiting conditions, and intracellular infection. Infect Immun. 2005;73(4):2020–2032. doi: 10.1128/IAI.73.4.2020-2032.2005. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 41.Ize B, Stanley NR, Buchanan G, Palmer T. Role of the Escherichia coli tat pathway in outer membrane integrity. Mol Microbiol. 2003;48(5):1183–1193. doi: 10.1046/j.1365-2958.2003.03504.x. [DOI] [PubMed] [Google Scholar]
- 42.Craig M, Sadik AY, Golubeva YA, Tidhar A, Slauch JM. Twin-arginine translocation system (tat) mutants of Salmonella are attenuated due to envelope defects, not respiratory defects. Mol Microbiol. 2013;89(5):887–902. doi: 10.1111/mmi.12318. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 43.Leon-Kempis Mdel R, Guccione E, Mulholland F, Williamson MP, Kelly DJ. The campylobacter jejuni PEB1a adhesin is an aspartate/glutamate-binding protein of an ABC transporter essential for microaerobic growth on dicarboxylic amino acids. Mol Microbiol. 2006;60(5):1262–1275. doi: 10.1111/j.1365-2958.2006.05168.x. [DOI] [PubMed] [Google Scholar]
- 44.Kurihara S, Suzuki H, Tsuboi Y, Benno Y. Dependence of swarming in Escherichia coli K-12 on spermidine and the spermidine importer. FEMS Microbiol Lett. 2009;294(1):97–101. doi: 10.1111/j.1574-6968.2009.01552.x. [DOI] [PubMed] [Google Scholar]
- 45.Chattopadhyay MK, Tabor CW, Tabor H. Polyamines protect Escherichia coli cells from the toxic effect of oxygen. Proc Natl Acad Sci U S A. 2003;100(5):2261–2265. doi: 10.1073/pnas.2627990100. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 46.Jelsbak L, Thomsen LE, Wallrodt I, Jensen PR, Olsen JE. Polyamines are required for virulence in Salmonella enterica serovar typhimurium. PLoS One. 2012;7(4):e36149. doi: 10.1371/journal.pone.0036149. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 47.Postma PW, Lengeler JW, Jacobson GR. Phosphoenolpyruvate:carbohydrate phosphotransferase systems of bacteria. Microbiol rev. 1993;57(3):543–594. doi: 10.1128/mr.57.3.543-594.1993. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 48.Houot L, Chang S, Pickering BS, Absalon C, Watnick PI. The phosphoenolpyruvate phosphotransferase system regulates Vibrio cholerae biofilm formation through multiple independent pathways. J Bacteriol. 2010;192(12):3055–3067. doi: 10.1128/JB.00213-10. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 49.Wang Q, Millet YA, Chao MC, Sasabe J, Davis BM, Waldor MK. A genome-wide screen reveals that the Vibrio cholerae phosphoenolpyruvate phosphotransferase system modulates virulence gene expression. Infect Immun. 2015;83(9):3381–3395. doi: 10.1128/IAI.00411-15. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 50.Moye ZD, Zeng L, Burne RA. Modification of gene expression and virulence traits in Streptococcus mutans in response to carbohydrate availability. Appl Environ Microbiol. 2014;80(3):972–985. doi: 10.1128/AEM.03579-13. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 51.Tao T, Snavely MD, Farr SG, Maguire ME. Magnesium transport in Salmonella typhimurium: mgtA encodes a P-type ATPase and is regulated by Mg2+ in a manner similar to that of the mgtB P-type ATPase. J Bacteriol. 1995;177(10):2654–2662. doi: 10.1128/jb.177.10.2654-2662.1995. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 52.Choi E, Lee KY, Shin D. The MgtR regulatory peptide negatively controls expression of the MgtA Mg2+ transporter in Salmonella enterica serovar typhimurium. Biochem Biophys Res Commun. 2012;417(1):318–323. doi: 10.1016/j.bbrc.2011.11.107. [DOI] [PubMed] [Google Scholar]
- 53.Six S, Andrews SC, Unden G, Guest JR. Escherichia coli possesses two homologous anaerobic C4-dicarboxylate membrane transporters (DcuA and DcuB) distinct from the aerobic dicarboxylate transport system (Dct) J Bacteriol. 1994;176(21):6470–6478. doi: 10.1128/jb.176.21.6470-6478.1994. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 54.Nogrady N, Imre A, Rychlik I, Barrow PA, Nagy B. Genes responsible for anaerobic fumarate and arginine metabolism are involved in growth suppression in Salmonella enterica serovar typhimurium in vitro, without influencing colonisation inhibition in the chicken in vivo. Vet Microbiol. 2003;97(3–4):191–199. doi: 10.1016/j.vetmic.2003.08.011. [DOI] [PubMed] [Google Scholar]
- 55.Novik V, Hofreuter D, Galan JE. Identification of campylobacter jejuni genes involved in its interaction with epithelial cells. Infect Immun. 2010;78(8):3540–3553. doi: 10.1128/IAI.00109-10. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 56.Jacobsen I, Hennig-Pauka I, Baltes N, Trost M, Gerlach GF. Enzymes involved in anaerobic respiration appear to play a role in Actinobacillus pleuropneumoniae virulence. Infect Immun. 2005;73(1):226–234. doi: 10.1128/IAI.73.1.226-234.2005. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 57.Kvint K, Nachin L, Diez A, Nystrom T. The bacterial universal stress protein: function and regulation. Curr Opin Microbiol. 2003;6(2):140–145. doi: 10.1016/S1369-5274(03)00025-0. [DOI] [PubMed] [Google Scholar]
- 58.Seifart Gomes C, Izar B, Pazan F, Mohamed W, Mraheil MA, Mukherjee K, Billion A, Aharonowitz Y, Chakraborty T, Hain T. Universal stress proteins are important for oxidative and acid stress resistance and growth of listeria monocytogenes EGD-e in vitro and in vivo. PLoS One. 2011;6(9):e24965. doi: 10.1371/journal.pone.0024965. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 59.Liu WT, Karavolos MH, Bulmer DM, Allaoui A, Hormaeche RD, Lee JJ, Khan CM. Role of the universal stress protein UspA of Salmonella in growth arrest, stress and virulence. Microb Pathog. 2007;42(1):2–10. doi: 10.1016/j.micpath.2006.09.002. [DOI] [PubMed] [Google Scholar]
- 60.Nystrom T, Neidhardt FC. Cloning, mapping and nucleotide sequencing of a gene encoding a universal stress protein in Escherichia coli. Mol Microbiol. 1992;6(21):3187–3198. doi: 10.1111/j.1365-2958.1992.tb01774.x. [DOI] [PubMed] [Google Scholar]
- 61.Nystrom T, Neidhardt FC. Expression and role of the universal stress protein, UspA, of Escherichia coli during growth arrest. Mol Microbiol. 1994;11(3):537–544. doi: 10.1111/j.1365-2958.1994.tb00334.x. [DOI] [PubMed] [Google Scholar]
- 62.Nachin L, Nannmark U, Nystrom T. Differential roles of the universal stress proteins of Escherichia coli in oxidative stress resistance, adhesion, and motility. J Bacteriol. 2005;187(18):6265–6272. doi: 10.1128/JB.187.18.6265-6272.2005. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 63.Elhosseiny NM, Amin MA, Yassin AS, Attia AS. Acinetobacter baumannii universal stress protein a plays a pivotal role in stress response and is essential for pneumonia and sepsis pathogenesis. Int J Med Microbiol. 2015;305(1):114–123. doi: 10.1016/j.ijmm.2014.11.008. [DOI] [PubMed] [Google Scholar]
- 64.Kim JH, Bothe JR, Frederick RO, Holder JC, Markley JL. Role of IscX in iron-sulfur cluster biogenesis in Escherichia coli. J Am Chem Soc. 2014;136(22):7933–7942. doi: 10.1021/ja501260h. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 65.Shimomura Y, Takahashi Y, Kakuta Y, Fukuyama K. Crystal structure of Escherichia coli YfhJ protein, a member of the ISC machinery involved in assembly of iron-sulfur clusters. Proteins. 2005;60(3):566–569. doi: 10.1002/prot.20481. [DOI] [PubMed] [Google Scholar]
- 66.Pastore C, Adinolfi S, Huynen MA, Rybin V, Martin S, Mayer M, Bukau B, Pastore A. YfhJ, a molecular adaptor in iron-sulfur cluster formation or a frataxin-like protein? Structure. 2006;14(5):857–867. doi: 10.1016/j.str.2006.02.010. [DOI] [PubMed] [Google Scholar]
- 67.Runyen-Janecky L, Daugherty A, Lloyd B, Wellington C, Eskandarian H, Sagransky M. Role and regulation of iron-sulfur cluster biosynthesis genes in Shigella flexneri virulence. Infect Immun. 2008;76(3):1083–1092. doi: 10.1128/IAI.01211-07. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 68.Kim SH, Lee BY, Lau GW, Cho YH. IscR modulates catalase a (KatA) activity, peroxide resistance and full virulence of Pseudomonas aeruginosa PA14. J Microbiol Biotechnol. 2009;19(12):1520–1526. doi: 10.4014/jmb.0906.06028. [DOI] [PubMed] [Google Scholar]
- 69.Karsi A, Lawrence ML. Broad host range fluorescence and bioluminescence expression vectors for gram-negative bacteria. Plasmid. 2007;57(3):286–295. doi: 10.1016/j.plasmid.2006.11.002. [DOI] [PubMed] [Google Scholar]
- 70.Karlyshev AV, Pallen MJ, Wren BW. Single-primer PCR procedure for rapid identification of transposon insertion sites. Biotechniques. 2000;28(6):1078. doi: 10.2144/00286bm05. [DOI] [PubMed] [Google Scholar]
- 71.Conesa A, Gotz S, Garcia-Gomez JM, Terol J, Talon M, Robles M. Blast2GO: a universal tool for annotation, visualization and analysis in functional genomics research. Bioinformatics. 2005;21(18):3674–3676. doi: 10.1093/bioinformatics/bti610. [DOI] [PubMed] [Google Scholar]
- 72.Yu NY, Wagner JR, Laird MR, Melli G, Rey S, Lo R, Dao P, Sahinalp SC, Ester M, Foster LJ, et al. PSORTb 3.0: improved protein subcellular localization prediction with refined localization subcategories and predictive capabilities for all prokaryotes. Bioinformatics. 2010;26(13):1608–1615. doi: 10.1093/bioinformatics/btq249. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 73.Kumar R, Nanduri B. HPIDB--a unified resource for host-pathogen interactions. BMC bioinformatics. 2010;11(Suppl 6):S16. doi: 10.1186/1471-2105-11-S6-S16. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 74.Zhou CE, Smith J, Lam M, Zemla A, Dyer MD, Slezak T. MvirDB--a microbial database of protein toxins, virulence factors and antibiotic resistance genes for bio-defence applications. Nucleic Acids Res. 2007;35(Database issue):D391–D394. doi: 10.1093/nar/gkl791. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 75.de Lorenzo V, Timmis KN. Analysis and construction of stable phenotypes in gram-negative bacteria with Tn5 and Tn10-derived minitransposons. Methods Enzymol. 1994;235:386–405. doi: 10.1016/0076-6879(94)35157-0. [DOI] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.
Data Availability Statement
The datasets used and/or analyzed during the current study are available from the corresponding author on reasonable request.