

Abstract
Background
Synbranchidae or swamp eels are fishes belonging to the order Synbranchiformes that occur in both freshwater and occasionally in brackish. They are worldwide distributed in tropical and subtropical rivers of four different continents. A large degree of chromosomal variation has been found in this family, mainly through the use of conventional cytogenetic investigations. Inside this group, a still almost unexplored species under the cytogenetic point of view is the Asian swamp eel Monopterus albus, a widely distributed species throughout Asia. Here, we tested the hypothesis of chromosomal speciation, where a case of sympatric speciation may occur as the primary consequence of chromosomal rearrangements. We performed a comparative chromosomal analysis of M. albus from 22 different localities in Thailand, using distinct staining methods (C-banding, Ag-NO3, and Chromomycin A3), and FISH with repetitive DNA probes (5S rDNA, 18S rDNA, Rex1 element and microsatellite repeats).
Results
This approach evidenced two contrasting karyotypes (named karyomorphs A and B) that varied concerning their 2n and repetitive DNAs distribution, where chromosomal fusions and pericentric inversions were involved in such differentiation. While the karyomorph A has 2n = 24 chromosomes, the karyomorph B has only 2n = 18, both with NF = 24. In addition, karyomorph A contains only acrocentric chromosomes, while karyomorph B contains three unique metacentric pairs. These features highlight that M. albus has already gone through a significant genomic divergence, and may include at least two cryptic species.
Conclusions
This marked chromosomal differentiation, likely linked to the lifestyle of these fishes, point to the occurrence of a chromosomal speciation scenario, in which fusions and inversions had a prominent role. This highlights the biodiversity of M. albus and justifies its taxonomic revision, since this nominal species may constitute a species complex.
Keywords: Tropical freshwater fish, Reproductive isolation, Repetitive DNAs, Centric fusion, Species complex
Background
Freshwater habitats make up less than 0.01% of available aquatic habitat but contain almost half of all 34,000 valid fish species, making freshwater fishes an excellent model for studying speciation events [1, 2]. However, although freshwater environments are largely fragmented and isolated, which means allopatric speciation events are more frequently found, several known cases of sympatric speciation have already been identified [3, 4].
In recent years, cytogenetic studies have made important contributions toward a better understanding of recent speciation events, since chromosomal rearrangements can act as genetic barriers to gene flow, thus facilitating reproductive isolation [5–8]. The chromosomal rearrangements promote the reorganization of the genetic structure, and the evolutionary impact and the consequences at the speciation level can vary according to the rearrangement type, that is, inversion, fusion, fission or translocation [7–12]. Such chromosomal rearrangements can facilitate adaptation to heterogeneous environments by limiting genomic recombination [10].
Molecular cytogenetic studies using fluorescence in situ hybridization (FISH) to map repetitive DNA sequences have provided important contributions to the characterization of chromosomal rearrangements and the evolution of distinct fish groups (reviewed in [13]). Repetitive DNAs, widely distributed in the eukaryotic genomes, are generally divided into two classes, one comprising tandem sequences (satellite DNAs, minisatellites and microsatellites), and the other comprising interspersed sequences, such as transposons and retrotransposons [14].
Synbranchidae or swamp eels are fishes belonging to the order Synbranchiformes; they occur in freshwater and occasionally in brackish water. They are distributed worldwide in tropical and subtropical Asia, the Indo-Australian Archipelago, West Africa (Liberia), Mexico and Central and South America (Fig. 1) [1]. This family comprises four genera: Macrotrema, Monopterus, Ophisternon and Synbranchus [15]. While Macrotrema, Monopterus and Ophisternon are found in the Old World, Ophisternon and Synbranchus occur in the New World. Thus, the genus Ophisternon is currently found in both the Old and New Worlds. Macrotrema is restricted to Asia and Synbranchus to the Neotropical region. At present, 23 valid species are recognized: Macrotrema (1); Monopterus (13), Ophisternon (6) and Synbranchus (3) [16]. Macrotrema caligans (Cantor, 1849), Monopterus albus (Zuiew, 1793) and Ophisternon bengalense (McClelland, 1844) are the three species recorded in Thailand [1].
Fig. 1.
World map highlighting the actual distribution of Synbranchidae fish family. This map was created using the following softwares: QGis 3.4.3 and Inkscape 0.92
Monopterus albus, commonly known as the Asian swamp eel or rice field eel, is widely distributed throughout Asia, from northern India and Burma to China, Asiatic Russia, Japan and the Indo-Malayan Archipelago. However, cytogenetic studies conducted on this species are scarce. There are only three karyotype reports for M. albus from Thailand and China, showing 24 chromosomes and the fundamental number NF = 24 [17–19]. However, Donsakul and Magtoon [17] have also reported that M. albus from the central part of Thailand differs from the other populations, as it has 2n = 18 and NF = 24 (Table 1). Similarly, the Venezuelan Ophisternon aenigmaticum also highlights a karyotype diversification, including 2n = 45 and NF = 51 and 2n = 46 and NF = 52 [20], as well as the Synbranchus marmoratus species from Brazil and Argentina, which presents 2n ranging from 42 to 46 [21–25]. The Synbranchidae family has high karyotype diversity among its species, with 2n ranging from 18 to 46, mainly due to extensive chromosomal rearrangements (fusions/fissions and inversions) (Table 1).
Table 1.
Comparative cytogenetic data of Symbranchidae species
| Species | Site sampling | 2n | NF | Karyotype | Reference |
|---|---|---|---|---|---|
| Monopterus albus | Central of Thailand | 18 | 24 | 6 m + 12a | [6] |
| Northeast of Thailand | 24 | 24 | 24a | [6] | |
| Central of Thailand | 24 | 24 | 24a | [8] | |
| China | 24 | 24 | 24a | [7] | |
| M. cuchia | India | 42 | 46 | 4sm + 38a | [33] |
| Ophisternon aenigmaticum | La Vega, Garcia, Venezuela | 45 | 51 | 6 m + 39a | [9] |
| EL Valle, Garcia, Venezuela | 46 | 52 | 6 m + 40a | ||
| O. bangalense | Southeast coast of India | 46 | 42 m + 4sm | [34] | |
| Synbranchus lampreia | Lago Catalão | 44 | 50 | 6 m + 2st + 36a | [19] |
| S. madeirae | Lago Catalão | 46 | 52 | 6 m + 2st + 38a | |
| S. marmoratus | Bataguassu – MS, Igaraçu do Tietê – SP, Pirassununga – SP, Icém – SP, Cáceres – MT, BR | 42 | 46 | 4 m + 12st + 26a | [14] |
| Synbranchus marmoratus | Coxim, MS, BR | 42 | 46 | 4 m,sm + 38st,a | [10] |
| São Simão, GO, BR | 42 | 46 | 4 m,sm + 38st,a | ||
| Nova Granada, SP, BR | 42 | 46 | 4 m,sm + 38st,a | ||
| Botucatu, SP, BR | 42 | 46 | 4 m,sm + 38st,a | [11] | |
| Birigui, SP, BR | 42 | 46 | 4 m,sm + 38st,a | ||
| ParaguaçuPaulista, SP, BR | 42 | 46 | 4 m,sm + 38st,a | ||
| Pirassununga, SP, BR | 42 | 48 | 6 m,sm + 36st,a | ||
| RibeirãoPreto, SP, BR | 42 | 48 | 6 m,sm + 36st,a | ||
| Bataguaçu, MS, BR | 42 | 48 | 6 m,sm + 36st,a | ||
| Guaíra – PR, BR | 42 | 48 | 6 m + 10st + 26a | [14] | |
| Londrina, PR, BR | 42 | 52 | 4 m + 2sm + 8st + 28a | [13] | |
| Guairá, PR, BR | 42 | 52 | 4 m + 2sm + 8st + 28a | ||
| Miranda, MS, BR | 42 | 52 | 4 m + 2sm + 8st + 28a | ||
| Pereiras, SP, BR | 42 | 52 | 4 m + 6sm + 8st + 24a | ||
| PresidenteEpitácio, SP, BR | 42 | 52 | 4 m + 6sm + 8st + 24a | ||
| Rio Claro, SP, BR | 44 | 48 | 4 m,sm + 40st,a | [10] | |
| Pentecostes, CE, BR | 44 | 48 | 4 m,sm + 40st,a | ||
| Botucatu, SP, BR | 44 | 48 | 4 m,sm + 40st,a | [11] | |
| Birigui, SP, BR | 44 | 48 | 4 m,sm + 40st,a | ||
| Bataguaçu, MS, BR | 44 | 48 | 4 m,sm + 40st,a | ||
| Ituzaingó, Corrientes, AR | 44 | 48 | 4 m,sm + 40st,a | [12] | |
| Reconquista, Santa Fé, AR | 44 | 48 | 4 m,sm + 40st,a | ||
| Garabato, Santa Fé, AR | 44 | 48 | 4 m,sm + 40st,a | ||
| Cerrito – RS, Rio Branco – AC, BR | 44 | 48 | 4 m + 10st + 30a | [14] | |
| Pirassununga, SP, BR | 44 | 50 | 4 m + 2sm + 8st + 30a | [13] | |
| Pirassununga, SP, BR | 46 | 50 | 4 m,sm + 42st,a | [11] | |
| RibeirãoPreto, SP, BR | 46 | 50 | 4 m,sm + 42st,a | ||
| Bandeirantes, PR, BR | 46 | 52 | 4 m + 2sm + 8st + 32a | [13] | |
| Igaraçu do Tietê – SP, Pirassununga – SP, BR | 46 | 52 | 4 m + 10st + 32a | [14] | |
| Pirassununga – SP, Icém – SP, BR | 46 | 52 | 6 m + 10st + 30a | ||
| Miranda, MS, BR | 46 | 54 | 6 m + 2sm + 6st + 32a | [13] |
Notes: BR Brazil, AR Argentina, 2n diploid number, NF fundamental number, m metacentric, sm submetacentric, st subtelocentric and a acrocentric chromosomes
This study presents a comparative chromosomal analysis of Monopterus albus from 22 different localities in Thailand (Fig. 2), using distinct staining methods (C-banding, Ag-NO3, and chromomycin A3) as well as FISH with repetitive DNA probes (5S rDNA, 18S rDNA, Rex1 element and microsatellite repeats). We tested the hypothesis of chromosomal speciation, where a case of sympatric speciation may occur as the primary consequence of chromosomal rearrangements. This approach provided an in-depth karyotype characterization of this taxon, evidencing the presence of two contrasting karyotypes occurring in sympatry and the occurrence of likely distinct species in this nominal species. This marked chromosomal differentiation, likely linked to the lifestyle of these fishes and their population fragmentation, protects from gene flow and therefore promotes speciation.
Fig. 2.
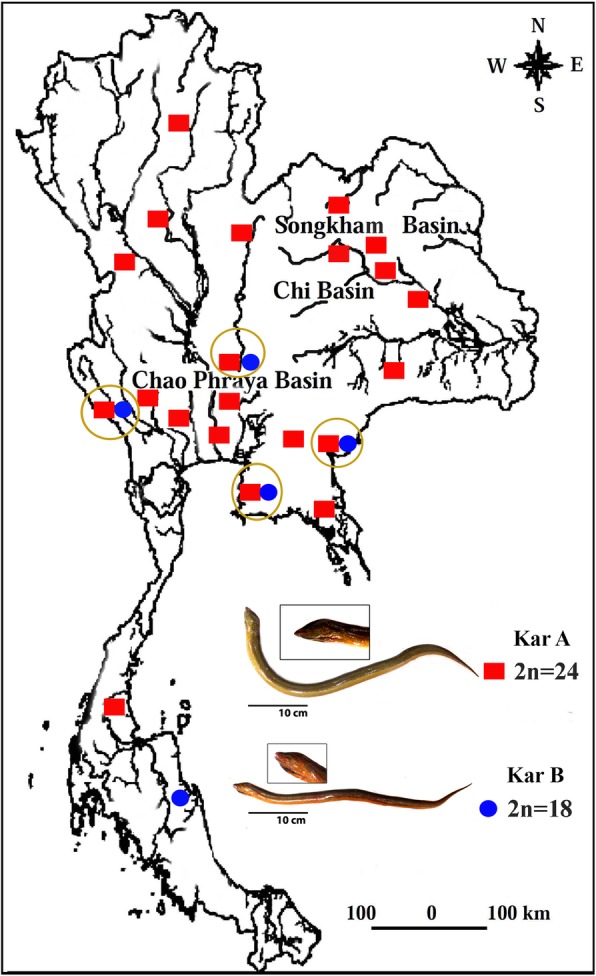
Map of Thailand highlighting the sample sites and specimens of two karyomorphs of the Asian swamp eel Monopterus albus. Red squares = karyomorph A (2n = 24) and blue circles = karyomorph B (2n = 18). Sympatry of both karyomorphs is indicated by circles. This map was created using the following software: Adobe Photoshop 7.0
Results
Karyotypes
M. albus from the populations analyzed presented two distinct karyotype forms, one with 2n = 24 (24a) and NF = 24 (karyomorph A) and the other with 2n = 18 (6 m + 12a) and NF = 24 (karyomorph B) (Fig. 3a). The first karyomorph was present in 21 localities (183 specimens), while the latter was found in five (17 specimens). Sympatry of both karyomorphs was observed in four localities, namely Nakhon Nayok, Kanchanaburi (Sri Yok), Chon Buri and Sa Kaeo Provinces (Table 2 and Fig. 2). No heterozygous karyotype forms were observed in the four localities of sympatry.
Fig. 3.

Karyotypes of the two Asian swamp eel Monopterus albus karyomorphs analyzed by (a) conventional staining and C-banding and (b) fluorescence in situ hybridization (FISH) with 5S (green) and 18S (red) rDNA probes. Inserted boxes indicate NOR-bearing chromosomes and CMA3 positive chromosomes. In (c) idiograms highlight the distribution of 18S rDNA (red) and 5S rDNA (green) in both karyomorphs. In the middle, the panel label highlights the main rearrangements related to karyotype differentiations, where centric fusions have played a major role. Scale bar = 5 μm
Table 2.
Chromosomal data for Monopterus albus populations from different Thailand regions
| Region | Population/Province | Number of specimen | 2n | NF | Karyotype | NORs |
|---|---|---|---|---|---|---|
| Central | Nakhon Nayok | 5 | 18 | 24 | 6 m + 12a | 2 (near telomere) |
| 7 | 24 | 24 | 24a | 2 (near cetromere) | ||
| Nakhon Pathom | 15 | 24 | 24 | 24a | 2 (near cetromere) | |
| Bangkok | 8 | 24 | 24 | 24a | 2 (near cetromere) | |
| Sing Buri | 10 | 24 | 24 | 24a | 2 (near cetromere) | |
| South | Nakhon Si Thammarat | 8 | 18 | 24 | 6 m + 12a | 2 (near telomere) |
| Chumphon | 8 | 24 | 24 | 24a | 2 (near cetromere) | |
| West | Kanchanaburi | 6 | 18 | 24 | 6 m + 12a | 2 (near telomere) |
| (Sri Yok) | 8 | 24 | 24 | 24a | 2 (near cetromere) | |
| Kanchanaburi (Sri Sawat) | 11 | 24 | 24 | 24a | 2 (near cetromere) | |
| East | Chon Buri | 6 | 18 | 24 | 6 m + 12a | 2 (near telomere) |
| 10 | 24 | 24 | 24a | 2 (near cetromere) | ||
| Sa Kaeo (including | 9 | 18 | 24 | 6 m + 12a | 2 (near telomere) | |
| Campodia) | 6 | 24 | 24 | 24a | 2 (near cetromere) | |
| Chanthaburi | 15 | 24 | 24 | 24a | 2 (near cetromere) | |
| PrachinBuri | 12 | 24 | 24 | 24a | 2 (near cetromere) | |
| North | Tak | 8 | 24 | 24 | 24a | 2 (near cetromere) |
| Phayao | 10 | 24 | 24 | 24a | 2 (near cetromere) | |
| Sukhothai | 10 | 24 | 24 | 24a | 2 (near cetromere) | |
| Northeast | KhonKaen | 12 | 24 | 24 | 24a | 2 (near cetromere) |
| Loei | 8 | 24 | 24 | 24a | 2 (near cetromere) | |
| Roi Et | 8 | 24 | 24 | 24a | 2 (near cetromere) | |
| Kalasin | 5 | 24 | 24 | 24a | 2 (near cetromere) | |
| Buri Ram | 6 | 24 | 24 | 24a | 2 (near cetromere) | |
| UdonThani | 8 | 24 | 24 | 24a | 2 (near cetromere) | |
| Mahasarakham | 4 | 24 | 24 | 24a | 2 (near cetromere) |
Notes: 2n diploid number, NF fundamental number, m metacentric and a acrocentric chromosomes
C-banding, ag-NORs and Chromomycin A3 staining
C-positive heterochromatic bands were observed in the centromeric/pericentromeric region of all chromosomes as well as in the telomeric region of several pairs in both karyomorphs. Interstitial heterochromatic blocks were also found in chromosome pair no. 4 in both karyomorphs (Fig. 3a). Ag-NORs sites were present in the centromeric region of pair no. 7 and on the telomeric region of pair no. 3 in karyomorphs A and B, respectively. These Ag-NORs were the only observed GC-rich regions in the karyotype (Fig. 3a, boxed).
Chromosome mapping of 5S and 18S rDNAs
5S rDNA sequences were found in the pericentromeric region of the q arms of chromosome pair no. 7 in karyomorph A and in the pericentromeric region of the p arms of chromosome pair no. 3 in karyomorph B (Fig. 3b). Concerning the 18S rDNA sequences, chromosome pair no. 7 of karyomorph A also displayed sites in the pericentromeric region of the q arms, while in karyomorph B, in addition to the signals in the pericentromeric region, chromosomal pair no. 3 also showed telomeric markings (Fig. 3b). Therefore, the 5S and 18S rDNAs are located together in one chromosome pair in both karyomorphs.
Microsatellites and Rex1 distribution
The (GC)15, (CAA)10, (CAC)10, (CAG)10, (CAT)10, (CGG)10, (GAA)10 and (GAG)10 repeats displayed scattered hybridization signals throughout the genome of both karyomorphs, some showing more concentrated signals. However, while the (CA)15 and (TA)15 sequences also show a scattered distribution in karyomorph B, they clearly accumulate at the centromeric region of several chromosomes in karyomorph A. In turn, the (GA)15 and the retroelement Rex1 sequences present a strong, dispersed distribution without preferential accumulation in any chromosome pairs of either karyomorph (Figs. 4 and 5).
Fig. 4.

Metaphase plates of Monopterus albus karyomorph A mapped with di- and trinucleotide microsatellites and Rex1 as probes. Scale bar = 5 μm
Fig. 5.

Metaphase plates Monopterus albus karyomorph B mapped with di- and trinucleotide microsatellites and Rex1 as probes. Scale bar =5 μm
Discussion
The occurrence of distinct M. albus karyomorphs living in sympatry with the absence of natural hybrids, as found in the present study in Nakhon Nayok, Kanchanaburi (Sri Yok), Chon Buri and Sa Kaeo Provinces, reinforces the hypothesis that these karyomorphs represent two reproductively isolated biological units. Although easily distinguishable through cytogenetic analysis, specimens from both karyomorphs have the same morphology, making difficult the identification of such probable new species.
The integration of both conventional and molecular cytogenetic approaches allowed the proposal of some chromosomal rearrangements probably related to the differentiation of both M. albus karyomorphs, where centric fusions appear as the main evolutionary sources shaping such a process (Fig. 3c). The different karyotype composition among individuals allowed us to identify two distinct karyomorphs, named A and B, which presented 2n of 24 and 18 chromosomes, respectively, both with NF = 24. The chromosomal divergence as well as the relation between the karyomorphs is clearly evidenced by their same NF and different karyotype formulas. This indicates that karyomorph B originated from karyomorph A (with only acrocentric chromosomes), where centric fusions were the most probable mechanism behind the presence of six metacentric chromosomes in karyomorph B (Fig. 3c). However, another scenario can also not be ruled out, in which fission-type rearrangements would have originated the additional 12 acrocentric chromosomes in karyomorph A. Once the majority of chromosomal rearrangements involve heterochromatic regions, especially in fish species [26, 27], this explains the centromeric rearrangements in M. albus. The co-localization of CMA3 positive heterochromatin with the NOR loci also occurs in other synbranchids [25, 28], probably because of local changes in base composition (increase in GC content) due to the so-called GC-biased gene conversion that involves rDNA in many vertebrates, including ray-finned fishes [29].
Although the 18S and 5S sites are located in an acrocentric pair in karyomorph A and in a metacentric pair in karyomorph B, it is possible to infer that both pairs are related in both karyomorphs. As karyomorph B contains additional 18S rDNA signals in the telomeric region in pair no. 3, it is likely that a pericentric inversion has divided the pericentromeric 18S rDNA loci into two parts and has transposed one of them near the telomere. Centric fusions would be the most suitable rearrangements to illustrate this scenario.
The differential distribution of the microsatellite motifs (CA)15 and (TA)15 between the two karyomorphs reinforces the interpretation that they represent, in fact, different species. A genome-wide analysis found that microsatellites have a repeat- and chromosome-biased distribution in M. albus from China, mainly located in noncoding regions (98,602, 99%) [30]. Differences both in the abundance and in the chromosomal location of several microsatellite motifs have been reported among closely related fish species also involved in recent speciation events [31, 32]. Similar to microsatellites, the Rex1 sequences are also resolutive markers for comparative genomic studies, as was already shown for several Asian fish species [19, 30, 31]. In the present study, the interspersed distribution of the retroelement Rex1 contrasts with that reported by [19].
Inter- and/or intrapopulation diversity has also been found in the karyotypes of other Synbranchidae species. For example, Ophisternon aenigmaticum and Synbranchus marmoratus possess high karyotype variability, with several karyomorphs described among their populations; they are also considered species complexes, where independent and bidirectional rearrangements, such as fusion and fission events, were responsible for the distinct 2n and karyotypic characteristics observed (Table 1). Biological, physiological and/or reproductive characteristics of Synbranchidae fishes may facilitate the intra- and interspecific karyotype variability observed among species and populations, especially in Monopterus and Synbranchus. These species tolerate a wide range of water oxygen levels, being able to obtain up to 25% of air oxygen by the cutaneous surface, allowing them to survive up to nine months in a drying burrow [33]. They also form small populations, and although they prefer freshwater habitats, these fishes tolerate brackish and saline conditions [1]. On the other hand, as many individuals can be isolated in small lakes during dry years, the capacity of sex reversal observed in synbranchids could contribute to the viability of such populations, increasing the probability of the fixation of genetic differences and speciation processes [25, 28].
Conclusions
In summary, the present scenario points to the occurrence of a chromosomal speciation scenario, in which fusions and inversions had a prominent role in the diversification of two distinct karyomorphs that differ with respect to diploid number, chromosome features and repetitive DNA distributions. However, this does not necessarily mean that a sympatric speciation is the only viable alternative, since such karyomorphs could have been established in allopatry, where a secondary join between them originated the present distribution. The karyotypic features highlight the biodiversity of M. albus and justify a taxonomic revision, since this nominal species may actually constitute a species complex.
Methods
Individuals examined
Two hundred specimens of M. albus were collected in 22 localities from distinct Thai regions (Fig. 2 and Table 2). The specimens were caught using traps, and after capture, the animals were placed in sealed plastic bags containing oxygen and were transported to the research station. The specimens were deposited in the fish collection of the Cytogenetic Laboratory, Department of Biology, Faculty of Science, Khon Kaen University. All the experiments followed ethical protocols, and anesthesia with clove oil was used prior to sacrificing the animals to minimize suffering. The fishes were then immersed in an ice-slurry to achieve death by hypothermia. The process was approved by the Animal Ethics Committee of Khon Kaen University based on the Ethics of Animal Experimentation of the National Research Council of Thailand AEKKU23/2558.
Chromosome preparation and C-banding, ag- and CMA3 staining
Mitotic chromosomes were obtained from the cell suspensions of the anterior kidney, using the conventional air-drying method [34]. Conventional staining was done using 5% Giemsa solution in phosphate buffer, pH 6.8, for 10 min. Chromosomes were analyzed after silver nitrate staining [35] in order to visualize the nucleolar organizing regions (Ag-NORs), and C-banding was also employed to detect the C-positive heterochromatin [36]. GC-specific fluorochrome chromomycin A3 (CMA3) was carried out following the method of Amemiya and Gold [37] to detect CG-rich regions on the chromosomes.
Preparation of FISH probes derived from repetitive sequences
Two tandemly arrayed rDNA sequences isolated from the genome of an Erythrinidae fish species, Hoplias malabaricus, were used as probes, as described in details in [38, 39]. The 5S and 18S rDNA probes were labeled with Spectrum Green dUTP and Spectrum Orange dUTP, respectively, using nick translation according to the manufacturer’s recommendations (Roche, Mannheim, Germany).
The microsatellites (CA)15, (GA)15,(GC)15, (TA)15, (CAA)10, (CAC)10, (CAG)10, (CAT)10, (CGG)10, (GAA)10 and (GAG)10 were synthesized according to Kubat et al. [40]. During synthesis by Sigma (St. Louis, MO, USA), these sequences were directly labeled with Cy3 at the 5’terminus. The retrotransposable element Rex1 sequence was prepared by PCR, using primers described in Volff et al. [41]. The Rex1 probe was directly labeled with Spectrum Orange dUTP by nick translation, according to the manufacturer’s recommendations (Roche, Mannheim, Germany).
Detection of repetitive DNA sequences by FISH
All FISH experiments with repetitive DNA probes were essentially carried out according to the protocol described in Yano et al. [42]. The first post-hybridization wash was performed with 2x SSC for 5 min at 42 °C, and a final wash was performed at room temperature in 1x SSC for 5 min. Finally, the slides were counterstained with DAPI and mounted in an antifade solution (Vectashield from Vector Laboratories).
Microscopic analysis and image processing
At least 30 metaphase spreads per individual were analyzed to confirm the 2n, karyotype structure and FISH results. Images were captured using an Olympus BX50 microscope (Olympus Corporation, Ishikawa, Japan) with Cool SNAP, and the images were processed using Image Pro Plus 4.1 software (Media Cybernetics, Silver Spring, MD, USA). Chromosomes were classified as acrocentric (a) or metacentric (m), according to their arm ratios [43].
Acknowledgements
Not applicable.
Funding
This study had financial support by the Young Researcher Development Project of Khon Kaen University (Year 2015); Toxic Substances in Livestock; Aquatic Animals Research Group; Post-Doctoral Training Program from Research Affairs and Graduate School (Grant no 59255), Khon Kaen University, Thailand; by the Conselho Nacional de Desenvolvimento Científico e Tecnológico- CNPq (Proc. nos 304992/2015–1 and 401962/2016–4) and CAPES/Alexander von Humboldt (Proc. No. 88881.136128/2017–01). This study was financed in part by the Coordenação de Aperfeiçoamento de Pessoal de Nível Superior - Brasil (CAPES) - Finance Code 001. These funding sources had no role in the design of the study and collection, analysis, and interpretation of data and in writing the manuscript.
Availability of data and materials
The datasets used and/or analyzed during the current study available from the corresponding author on reasonable request.
Abbreviations
- 2n
Diploid chromosome number
- a
Acrocentric chromosome
- CMA3
Chromomycin A3
- DAPI
4′,6-diamidino-2-phenylindole
- dUTP
2′-Deoxyuridine-5′-Triphosphate
- FISH
Fluorescence in situ hybridization
- FN
Fundamental number
- m
Metacentric chromosome
- NOR
Nucleolar organizer region
- NTS
Non-transcribed spacer
- PCR
Polymerase chain reaction
- rDNA
ribosomal DNA
- sm
Submetacentric chromosome
- st
Subtelocentric chromosome
Authors’ contributions
WS, CFY, EAO and MBC carried out the molecular cytogenetic analysis and drafted the manuscript. KP, KS, PS,TL and SP helped in analysis and drafted the manuscript. LACB, TL, MBC and AT coordinated the study, drafted and revised the manuscript. All authors read and approved the final version of the manuscript.
Ethics approval
All the experiments followed ethical protocols and anesthesia with clove oil was used prior to sacrificing the animals to minimize suffering. The process was approved by the Animal Ethics Committee of Khon Kaen University based on the Ethic of Animal Experimentation of National Research Council of Thailand AEKKU23/2558.
Consent for publication
Not applicable
Competing interests
The authors declare that they have no competing interests.
Publisher’s Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Contributor Information
Weerayuth Supiwong, Email: supiwong@hotmail.com.
Krit Pinthong, Email: k_pinthong@yahoo.com.
Kriengkrai Seetapan, Email: kook82@hotmail.com.
Pasakorn Saenjundaeng, Email: pasafish@yahoo.com.
Luiz A. C. Bertollo, Email: bertollo@ufscar.br
Ezequiel A. de Oliveira, Email: ezekbio@gmail.com
Cassia F. Yano, Email: yanocassia@gmail.com
Thomas Liehr, Email: Thomas.Liehr@med.uni-jena.de.
Sumalee Phimphan, Email: joodoof@gmail.com.
Alongklod Tanomtong, Email: tanomtong@hotmail.com.
Marcelo B Cioffi, Phone: +55 16 3351 8431, Email: mbcioffi@ufscar.br.
References
- 1.Nelson JS, Grande TC, MVH W. Fishes of the World. Hoboken: Wiley; 2016.
- 2.Bloom DD, Weir JT, Piller KR, Lovejoy NR. Do freshwater fishes diversify faster than marine fishes? A test using state-depedent diversification analyses and molecular phylogenetics of new world silversides (Atherinopsidae) Evolution. 2013;67:2040–2057. doi: 10.1111/evo.12074. [DOI] [PubMed] [Google Scholar]
- 3.Bolnick DI, Fitzpatrick BM. Sympatric Speciation: Models and Empirical Evidence. Annu Rev Ecol Evol Syst. 2007.
- 4.Seehausen O, Wagner CE. Speciation in freshwater fishes. Annu Rev Ecol Evol Syst. 2014;45:621–651. doi: 10.1146/annurev-ecolsys-120213-091818. [DOI] [Google Scholar]
- 5.Rieseberg LH. Chromosomal rearrangements and speciation. Trends Ecol Evol. 2001;16:351–358. doi: 10.1016/S0169-5347(01)02187-5. [DOI] [PubMed] [Google Scholar]
- 6.Navarro A, Barton NH. Chromosomal speciation and molecular divergence--accelerated evolution in rearranged chromosomes. Science. 2003;300:321–324. doi: 10.1126/science.1080600. [DOI] [PubMed] [Google Scholar]
- 7.Lowry DB, Willis JH. A widespread chromosomal inversion polymorphism contributes to a major life-history transition, local adaptation, and reproductive isolation. PLoS Biol. 2010;8:e1000500. doi: 10.1371/journal.pbio.1000500. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 8.Jay P, Whibley A, Frézal L, Rodríguez de Cara MÁ, Nowell RW, Mallet J, et al. Supergene Evolution Triggered by the Introgression of a Chromosomal Inversion. Curr Biol. 2018;28:1839–1845. doi: 10.1016/j.cub.2018.04.072. [DOI] [PubMed] [Google Scholar]
- 9.Berg PR, Star B, Pampoulie C, Bradbury IR, Bentzen P, Hutchings JA, et al. Trans-oceanic genomic divergence of Atlantic cod ecotypes is associated with large inversions. Heredity. 2017;119:418–428. doi: 10.1038/hdy.2017.54. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 10.Mérot C, Berdan EL, Babin C, Normandeau E, Wellenreuther M, Bernatchez L. Intercontinental karyotype-environment parallelism supports a role for a chromosomal inversion in local adaptation in a seaweed fly. Proc R Soc B Biol Sci. 2018;285:20180519. [DOI] [PMC free article] [PubMed]
- 11.Bakloushinskaya IY. Chromosomal rearrangements, genome reorganization, and speciation. Biol Bull. 2016;43:759–775. doi: 10.1134/S1062359016080057. [DOI] [Google Scholar]
- 12.Baker RJ, Bickham JW. Speciation by monobrachial centric fusions. Proc Natl Acad Sci. 1986;83:8245–8248. doi: 10.1073/pnas.83.21.8245. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 13.Cioffi MB, Bertollo LAC. Chromosomal distribution and evolution of repetitive DNAs in fish. In: Garrido-Ramos MA, editor. Repetitive DNA. Basel: Karger Publishers; 2012. 197–221. [DOI] [PubMed]
- 14.López-Flores I, Garrido-Ramos MA. The repetitive DNA content of eukaryotic genomes. In: Garrido-Ramos MA, editor. Repetitive DNA. Basel: Karger Publishers; 2012. pp. 1–28. [DOI] [PubMed] [Google Scholar]
- 15.Rosen DE, Rumney A. Evidence of a second species of Synbranchus (Pisces, Teleostei) in South America. American museum novitates; 2497. American Museum of Natural History: New York, NY; 1972. [Google Scholar]
- 16.Eschmeyer WN, Fong J. Species by family/subfamily in the catalog of fishes, electronic version). 2017. http://researcharchive.calacademy.org/research/ichthyology/catalog/SpeciesByFamily.asp. Accessed 31 Jul 2017.
- 17.Donsakul T, Magtoon W. A chromosome study of rice field eel, Fluta alba from Central and Northeastern parts of Thailand. Srinakharinwirot University; 1996.
- 18.Ji FY, Yu QX, Li K, Ren XH. Ag-staining pattern, FISH and ISH with rDNA probes in the rice field eel (Monopterus albus Zuiew) chromosomes. Hereditas. 2003;138:207–212. doi: 10.1034/j.1601-5223.2003.01643.x. [DOI] [PubMed] [Google Scholar]
- 19.Suntronpong A, Thapana W, Twilprawat P, Prakhongcheep O, Somyong S, Muangmai N, et al. Karyological characterization and identification of four repetitive element groups (the 18S–28S rRNA gene, telomeric sequences, microsatellite repeat motifs, rex retroelements) of the Asian swamp eel (Monopterus albus) Comp Cytogenet. 2017;11:435. doi: 10.3897/compcytogen.v11i3.11739. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 20.Nirchio M, Mariguela TC, Ferreira IA, Foresti F, Oliveira C. Karyotype and nucleolus organizer regions of Ophisternon aenigmaticum (Teleostei: Synbranchiformes: Synbranchidae) from Venezuela. Interciencia. 2011;3:229–233. [Google Scholar]
- 21.Foresti F, Oliveira C, Tien OS. Cytogenetic studies of the genus Synbranchus (Pisces, Synbranchiformes, Synbranchidae) Naturalia. 1992;17:129–138. [Google Scholar]
- 22.Melillo MIF, Foresti F, Oliveira C. Additional cytogenetic studies on local populations of Synbranchus marmoratus (Pisces, Synbranchiformes, Synbranchidae) Naturalia. 1996;21:201–208. [Google Scholar]
- 23.Sanchez S. Fenocchio a. S. Karyotypic analysis in three populations of the south- American eel like fish Synbranchus marmoratus. Caryologia. 1996;49:65–71. doi: 10.1080/00087114.1996.10797351. [DOI] [Google Scholar]
- 24.Torres RA, Roper JJ, Foresti F, Oliveira C. Surprising genomic diversity in the Neotropical fish Synbranchus marmoratus (Teleostei: Synbranchidae): how many species? Neotrop Ichthyol. 2005;3:277–284. doi: 10.1590/S1679-62252005000200005. [DOI] [Google Scholar]
- 25.Utsunomia R, Pansonato-Alves JC, Costa-Silva GJ, Mendonça FF, Scacchetti PC, Oliveira C, et al. Molecular and cytogenetic analyses of cryptic species within the Synbranchus marmoratus Bloch, 1795 (Synbranchiformes: Synbranchidae) grouping: species delimitations, karyotypic evolution and intraspecific diversification. Neotrop Ichthyol. 2014;12:903–912. doi: 10.1590/1982-0224-20140039. [DOI] [Google Scholar]
- 26.Oliveira C, Almeida-Toledo LF, Mori L, Toledo-Filho SA. Cytogenetic and DNA content in six genera of the family Callichthyidae (Pisces, Siluriformes) Caryologia. 1993;46:171–188. doi: 10.1080/00087114.1993.10797258. [DOI] [Google Scholar]
- 27.Yano CF, Margarido VP. First cytogenetic studies of the genus Heptapterus (Actinopterygii, Siluriformes): karyotype differentiation and review of cytogenetic data on the Heptapteridae family. J Fish Biol. 2012;81:939–953. doi: 10.1111/j.1095-8649.2012.03314.x. [DOI] [PubMed] [Google Scholar]
- 28.Carvalho NDM, Gross MC, Schneider CH, Terencio ML, Zuanon J, Feldberg E. Cytogenetics of Synbranchiformes: a comparative analysis of two Synbranchus Bloch, 1795 species from the Amazon. Genetica. 2012;140:149–158. doi: 10.1007/s10709-012-9666-5. [DOI] [PubMed] [Google Scholar]
- 29.Escobar JS, Glémin S, Galtier N. GC-biased gene conversion impacts ribosomal DNA evolution in vertebrates, angiosperms, and other eukaryotes. Mol Biol Evol. 2011;28:2561–2575. doi: 10.1093/molbev/msr079. [DOI] [PubMed] [Google Scholar]
- 30.Li Z, Chen F, Huang C, Zheng W, Yu C, Cheng H, et al. Genome-wide mapping and characterization of microsatellites in the swamp eel genome. Sci Rep. 2017;7:31–57. doi: 10.1038/s41598-017-00044-8. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 31.Supiwong W, Liehr T, Cioffi MB, Chaveerach A, Kosyakova N, Pinthong K, et al. Chromosomal evolution in naked catfishes (Bagridae, Siluriformes): a comparative chromosome mapping study. Zool Anz. 2014;253:316–320. doi: 10.1016/j.jcz.2014.02.004. [DOI] [Google Scholar]
- 32.Cioffi MB, LAC B, Villa MA, de Oliveira EA, Tanomtong A, Yano CF, et al. Genomic organization of repetitive DNA elements and its implications for the chromosomal evolution of channid fishes (Actinopterygii, Perciformes) PLoS One. 2015;10:e0130199. doi: 10.1371/journal.pone.0130199. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 33.Graham JB. Air-breathing fishes: evolution, diversity, and adaptation. San Diego: Academic Press; 1997. [Google Scholar]
- 34.Supiwong W, Jearranaiprepame P, Tanomtong A. A new report of karyotype in the chevron snakehead 2009;74:317–322.
- 35.Howell WM, Black DA. Controlled silver-staining of nucleolus organizer regions with a protective colloidal developer: a 1-step method. Experientia. 1980;36:1014–1015. doi: 10.1007/BF01953855. [DOI] [PubMed] [Google Scholar]
- 36.Sumner AT. A simple technique for demonstrating centromeric heterochromatin. Exp Cell Res. 1972;75:304–306. doi: 10.1016/0014-4827(72)90558-7. [DOI] [PubMed] [Google Scholar]
- 37.Amemiya CT, Gold JR. Chromomycin a 3 stains nucleolus organizer regions of fish chromosomes. Copeia. 1986;1986:226–231. doi: 10.2307/1444915. [DOI] [Google Scholar]
- 38.Martins C, Ferreira IA, Oliveira C, Foresti F, Galetti PM. A tandemly repetitive centromeric DNA sequence of the fish Hoplias malabaricus (Characiformes: Erythrinidae) is derived from 5S rDNA. Genetica. 2006;127:133–141. doi: 10.1007/s10709-005-2674-y. [DOI] [PubMed] [Google Scholar]
- 39.Cioffi MB, Martins C, Centofante L, Jacobina U, Bertollo LAC. Chromosomal variability among allopatric populations of Erythrinidae fish Hoplias malabaricus: mapping of three classes of repetitive DNAs. Cytogenet Genome Res. 2009;125:132–141. doi: 10.1159/000227838. [DOI] [PubMed] [Google Scholar]
- 40.Kubat Z, Hobza R, Vyskot B, Kejnovsky E. Microsatellite accumulation in the Y chromosome of Silene latifolia. Genome. 2008;51:350–356. doi: 10.1139/G08-024. [DOI] [PubMed] [Google Scholar]
- 41.Volff JN, Körting C, Froschauer A, Sweeney K, Schartl M. Non-LTR retrotransposons encoding a restriction enzyme-like endonuclease in vertebrates. J Mol Evol. 2001;52:351–360. doi: 10.1007/s002390010165. [DOI] [PubMed] [Google Scholar]
- 42.Yano CF, Bertollo LAC, Cioffi MB. FISH-FISH: molecular cytogenetics in fish species. In: Liehr T, editor. Fluorescence In Situ Hybridization (FISH)- Application Guide. 2. Berlin: Springer; 2017. pp. 429–444. [Google Scholar]
- 43.Levan A, Fredga K, Sandberg AA. Nomenclature for centromeric position on chromosomes. Hereditas. 1964;52:201–220. doi: 10.1111/j.1601-5223.1964.tb01953.x. [DOI] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.
Data Availability Statement
The datasets used and/or analyzed during the current study available from the corresponding author on reasonable request.