

Abstract
Adenosine monophosphate-activated protein kinase (AMPK) is a highly conserved heterotrimeric complex that acts as an intracellular energy sensor. Based on recent observations of AMPK expression in all structures of the female reproductive system, we hypothesized that AMPK is functionally required for maintaining fertility in the female. This hypothesis was tested by conditionally ablating the two catalytic alpha subunits of AMPK, Prkaa1 and Prkaa2, using Pgr-cre mice. After confirming the presence of PRKAA1, PRKAA2 and the active phospho-PRKAA1/2 in the gravid uterus by immunohistochemistry, control (Prkaa1/2fl/fl) and double conditional knockout mice (Prkaa1/2d/d) were placed into a six-month breeding trial. While the first litter size was comparable between Prkaa1/2fl/fl and Prkaa1/2d/d female mice (p=0.8619), the size of all subsequent litters was dramatically reduced in Prkaa1/2d/d female mice (p=0.0015). All Prkaa1/2d/d female mice experienced premature reproductive senescence or dystocia by the fourth parity. This phenotype manifested despite no difference in estrous cycle length, ovarian histology in young and old nulliparous or multiparous animals, mid-gestation serum progesterone levels, or uterine expression of Esr1 or Pgr between Prkaa1/2fl/fl and Prkaa1/2d/d female mice suggesting that the hypothalamic-pituitary-ovary axis remained unaffected by PRKAA1/2 deficiency. However, an evaluation of uterine histology from multiparous animals identified extensive endometrial fibrosis and disorganized stromal-glandular architecture indicative of endometritis, a condition that causes subfertility or infertility in most mammals. Interestingly, Prkaa1/2d/d female mice failed to undergo artificial decidualization. Collectively, these findings suggest that AMPK plays an essential role in endometrial regeneration following parturition and tissue remodeling that accompanies decidualization.
Keywords: AMPK, development, fertility, fibrosis, mesenchymal stem cell, pregnancy, Prkaa1, Prkaa2, regeneration, uterus
INTRODUCTION
Pregnancy requires considerable energy expenditure by the mother. While some of the energy is transferred from the uterus to the embryo prior to placentation, energy is primarily used within the uterus for the purpose of tissue remodeling as pregnancy is established. Mechanisms regulating energy utilization and distribution in the uterus have not been well described. In most tissues, the AMP-activated protein kinase (AMPK) serves as a cellular sensor of energy status that allows cells to directly adapt to support survival and expansion, and this is particularly true in tissues with high cellular turnover (Viollet et al. 2009a). The AMPK is a highly conserved heterotrimeric protein complex consisting of a catalytic α subunit and structural β and γ subunits. There are twelve possible AMPK complexes as two α, two β, and three γ isoforms exist, each encoded by a distinct gene. The individual components of the AMPK heterotrimer are differentially expressed based on the tissue type and biological context. In response to a decrease in cellular adenosine triphosphate (ATP), as detected by the ratio of ATP with its dephosphorylated precursors adenosine diphosphate (ADP) or adenosine monophosphate (AMP), AMPK acts to initiate catabolic processes while concomitantly inhibiting heavily energy dependent anabolic pathways (Hardie et al. 2006, Horman et al. 2012, Hardie & Lin 2017). This metabolic switch is essential for maintenance of ATP homeostasis. Activation and enzyme kinetics of AMPK have been well studied and several steps are involved. First, AMP/ADP becomes bound by cystathione beta synthase (CBS) domains in the γ regulatory subunit exposing the catalytic domain of the α subunit (Viollet et al. 2009b). Second, the α subunit is subsequently phosphorylated on threonine-172, which amplifies its kinase activity. Three upstream enhancer kinases called AMPK kinases phosphorylate the threonine-172 residue, and these include liver kinase beta 1 (LKB1) (Hawley et al. 2003) calcium/calmodulin-dependent protein kinase kinase 2 (CaMKK2) (Hawley et al. 2005), and transforming growth factor β-activated kinase (TAK1) (Xie et al. 2006). Finally, conformational changes in the shape of AMPK due to phosphorylation of threonine-172 of the α subunit and binding of AMP/ADP to the γ subunit prevents phosphatases from cleaving activation phosphate groups from AMPK, thus allowing for sustained activation of the alpha subunit (Shaw et al. 2004). Aside from its well-established role in the maintenance of energy homeostasis, AMPK also regulates production of and protection from reactive oxygen species (ROS) (Shirwany & Zou 2014), is regulated by ROS (Rabinovitch et al. 2017), regulates remodeling of the extracellular matrix (Jiang et al. 2017), contributes to tissue regeneration (Mounier et al. 2015), plays an essential role in development (Viollet & Foretz 2016), and promotes mitochondrial biogenesis (O’Neill et al. 2013, Herzig & Shaw 2018).
It is known that AMPK is necessary for embryonic development in mice, as the null mutation of genes encoding the alpha 1 and alpha 2 subunits (i.e., Prkaa1 and Prkaa2), results in embryonic lethality around DOP 10.5 (Viollet et al. 2009a). Surprisingly very little is known about AMPK signaling in female reproductive tissues. Based primarily on pharmacological manipulation of AMPK activity, AMPK activation results in a decrease in steroidogenesis in the bovine and rat granulosa cells (Tosca et al. 2005, Tosca et al. 2006, Tosca et al. 2007a, Tosca et al. 2010). A role for AMPK in oocyte maturation is species-dependent with AMPK activation resulting in oocyte activation in mice, but inhibition in pigs and cattle (Mayes et al. 2007, Tosca et al. 2007b, Santiquet et al. 2014, Bertoldo et al. 2015a). Conditional mutagenesis of the gene encoding the α1 subunit of AMPK from murine oocytes resulted in a 27% decrease in litter size owing to altered gap junction formation and disruption of oocyte:granulosa cell communication (Bertoldo et al. 2015b). Within the uterus, polymorphisms in the PRKAA1 gene are hypothesized to influence uterine artery diameter, fetal growth, and birth weight outcomes, and to allow for adaptation to high altitudes during pregnancy in Andean women (Bigham et al. 2014). Within murine uterine epithelial tissue, steroid hormones were shown to regulate expression of facilitative glucose transporters in an AMPK-dependent manner (Kim & Moley 2009). More recently, p53 modulates uterine AMPK activity, which in turn controls the timing of parturition in a murine model of preterm birth (Deng et al. 2016). Beyond these limited findings, relatively little is known about the role of AMPK in the female reproductive system at the whole animal level.
The present studies were designed to test the hypothesis that AMPK is necessary for early postnatal development of the female reproductive tract, female fertility and normal reproductive lifespan. The specific objectives were to: 1) evaluate uterine expression of PRKAA1 and PRKAA2; and 2) determine the consequences of conditionally ablating Prkaa1 and Prkaa2 on female fertility and maintenance of uterine histoarchitecture.
MATERIALS AND METHODS
Animals
All animal protocols were reviewed and approved by the Washington State University Institutional Animal Care and Use Committee. Reproductive tissues were isolated from CD1 mice to first evaluate PRKAA1 and PRKAA2 expression in the gravid uterus. Female CD1 mice were mated and the presence of a vaginal plug marked day of pregnancy (DOP) 0.5. Implantation sites were collected from female mice on DOP 7.5 and prepared for PRKAA1, PRKAA2 and phospho-PRKAA1/2 immunohistochemistry as described below.
To evaluate the importance of AMPK in uterine development and function, Prkaa1 and Prkaa2 double conditional knockout (dcKO) mice were developed by crossing mice harboring floxed Prkaa1 and Prkaa2 genes with Pgr-cre mice (Viollet et al. 2003, Soyal et al. 2005, Miller et al. 2011, Boudaba et al. 2018). Use of Pgr-cre allowed for conditional ablation of any cell that normally expressed PGR, and this includes gonadotropes, mammary epithelium, and peri-ovulatory follicles, as well as stromal, epithelial and smooth muscle of the majority of the female reproductive tract. A PCR-based genotyping approach was used for identifying mice with floxed Prkaa1 and Prkaa2 alleles, as well as Pgr-cre using DNA isolated from tail snips. The presence of Prkaa1 and Prkaa2 was detected by 30 cycles of PCR (denaturation, 30 sec at 94°C; annealing, 60 sec at 51°C; extension, 60 sec at 72°C) with specific forward and reverse primers (Table 1). The presence of Pgr-cre was detected by 35 cycles of PCR (denaturation, 45 sec at 94°C; annealing, 60 sec at 61°C; extension 60 sec at 72°C) with specific forward and reverse primers. Four control (Pgr+/+;Prkaa1fl/fl;Prkaa2fl/fl or Prkaa1/2fl/fl) and six double conditional knockout (Pgrcre/+;Prkaa1fl/fl;Prkaa2fl/fl or Prkaa1/2d/d) female mice were entered into a six-month breeding trial with male mice of proven fertility. The date of birth, number of pups born per litter, parity interval, the total number of litters throughout the breeding trial, and the number and weight of male and female pups at weaning were recorded. Reproductive tracts were collected from Prkaa1/2fl/fl and Prkaa1/2d/d female mice at the end of the breeding trial, as well as from young (6–8 weeks) and aged (6 months) nulliparous female mice for histological analyses. Serum progesterone (P4) was measured at the University of Virginia Center for Research in Reproduction Ligand Assay and Analysis Core.
Table 1.
Primers used for genotyping and qPCR
| Gene | Forward Primer | Reverse Primer (s) |
|---|---|---|
|
Prkaa1 (genotyping) |
5’-TATTGCTGCCATTAGGCTAC-3’ | 5’-GACCTGACAGAATAGGATATGC CCAACCTC-3’ |
|
Prkaa2 (genotyping) |
5’-GCTTAGCACGTTACCCTGGA TGG-3’ |
5’-GTTATCAGCCCAACTAATTACAC-3’ |
|
Pgr-cre (genotyping) |
5’-ATGTTTAGCTGGCCCAAATG-3’ | 5’-TATACCGATCTCCCTGGACG-3’ |
| 5’-CCCAAAGAGACACCAGGAAG-3’ | ||
|
Prkaa1 (qPCR) |
5’-CACGCTTGGTGTCGGC-3’ | 5’-GTTGACCGGACATAAAGTGGCT-3’ |
|
Prkaa2 (qPCR) |
5’-AACTAGCTGTGGATCGCCAA-3’ | 5’-CAGATATCGACCTCGGGACC-3’ |
|
Esr1 (qPCR) |
5’-CCAAAGCCTCGGGAATG-3’ | 5’-CTTTCTCGTTACTGCTGG-3’ |
|
Pgr (qPCR) |
5’-ATGGTCCTTGGAGGTCGTAA-3’ | 5’-CACCATCAGGCTCATCC-3’ |
|
Wnt4 (qPCR) |
5’-TGCAGATGTCAAACGGAAC-3’xg | 5’-TCCGGAACTGGTATTGGCAC-3’ |
|
Bmp2 (qPCR) |
5’-ACGAGAAAAGCGTCAAGCCA-3’ | 5’-CCAGTCATTCCACCCCACAT-3’ |
|
Rpl13a (qPCR) |
5’-TTGCTTACCTGGGGCGTCT-3’ | 5’-CCTTTTCCTTCCGTTTCTCCTCGC-3’ |
To evaluate estrous cyclicity at six weeks of age in Prkaa1/2fl/fl (n=3) and Prkaa1/2d/d (n=3) mice, vaginal smears were evaluated microscopically. Here, 50 μL saline was gently placed in the vaginal opening, collected, combined with 50 μL of 0.1% methylene blue dye, smeared on a microscope slide, and cover slipped. The stage of estrous cycle was then determined under 100X magnification. Female mice were evaluated on a daily basis for 21 days to determine the average cycle length for each mouse.
To determine if conditional ablation of Prkaa1 and Prkaa2 from the uterus impacted on the expression of the classical estrogen receptor (Esr1) and progesterone receptor (Pgr), uterine tissues were isolated from synchronized Prkaa1/2fl/fl and Prkaa1/2d/d mice (n=7 each). Synchronization was accomplished by ovariectomizing Prkaa1/2fl/fl and Prkaa1/2d/d mice and allowing them to clear endogenous steroids for one week. Mice were then given single daily injections of estradiol (E2, subcutaneous, 100 ng) for two days, followed by no treatment for four days and then a final single E2 injection. Uterine tissues were collected 24 h later and processed for quantitative PCR as described below.
Uterine decidualization was assessed artificially by first placing intact sexually mature Prkaa1/2fl/fl (n=4) and Prkaa1/2d/d (n=4) female mice with vasectomized male mice. Observation of a vaginal plug marked day of pseudopregnancy (DOPP) 0.5. On DOPP 4, 25 μl of sterile sesame oil was injected into the uterine horn through a dorsal incision using a burred 27-gauge needle. Four days following surgery, female mice were euthanized and uterine tracts collected, weighed and prepared for RNA isolation, as well as paraffin embedding and histological evaluation. To confirm this in greater detail, sexually mature Prkaa1/2fl/fl (n=4) and Prkaa1/2d/d (n=3) female mice were ovariectomized and allowed to rest for two weeks. Mice were then given a single daily injection of estradiol (100 ng in 50 μl sesame oil) for two days followed by two days of rest. Mice were then given single daily injections of estradiol (10 ng) + 1 mg progesterone for three days. In the late afternoon following the last injection, decidualization was stimulated in the left uterine horn by injecting 25 μl of sesame oil into the uterine lumen and scratching the antimesometrial surface with a burred needle. Mice were then given daily injections of progesterone (1 mg) for three days. Uteri were collected and the left uterine horn was weighed 24 h later.
To determine if conditional ablation of Prkaa1 and Prkaa2 altered expression of ESR1 and PGR protein expression, sexually mature nulliparous Prkaa1/2fl/fl and Prkaa1/2d/d female mice (n=7) were placed with vasectomized males to induce a state of pseudopregnancy so that mice could be synchronized. On DOPP 4, mice were artificially decidualized with sesame oil and uterine luminal scratching. Uterine tissues were collected 24 h later for fixation in 4% paraformaldehyde and paraffin embedding for thin sectioning and ESR1 and PGR immunohistochemistry (IHC).
RNA isolation and quantitative polymerase chain reaction (qPCR)
Uterine mRNA expression of Prkaa1, Prkaa2, Esr1, Pgr and Rpl13a was determined by qPCR using primer sets identified in Table 1. For these studies, RNA was isolated from liver and uteri obtained from Prkaa1/2fl/fl and Prkaa1/2d/d mice using Tri-Reagent (Sigma-Aldrich; St. Louis, MO) following the manufacture’s instruction. Liver RNA was isolated and used as a positive control for Prkaa1/2 expression to confirm conditional ablation of Prkaa1/2 from the uterus. Pgr-cre is not expressed in this liver and therefore Prkaa1/2 genes and transcribed mRNAs should remain intact. Isolated RNA was DNase treated (Roche Applied Science, Indianapolis, IN) and quantified using a NanoDrop 2000 Spectrophotometer (Thermo Scientific; Waltham, MA) prior to cDNA synthesis. Messenger RNA was then reverse transcribed using SuperScript II Reverse Transcriptase (Invitrogen; Carlsbad, CA). Quantitative PCR was completed using BioRad’s C1000 CFX96 Real-Time Thermal Cycler System with an annealing temperature of 56 ˚C (1 min) for 40 cycles. Data were analyzed with CFX Manager Software (BioRad; Hercules, CA). Rpl13a was used as an internal control. A no reverse transcriptase negative control was included to confirm the absence of genomic DNA.
Histology, immunohistochemistry and immunofluorescence
Collected tissues were fixed in 4% paraformaldehyde (PFA) overnight at 4°C. Tissues were then processed through a series of graded dehydrations in ethanol followed by xylenes and paraffin using Leica’s TP1020 automated system. Paraffin-embedded sections (5μm) were deparaffinized in xylenes followed by rehydration in graded ethanol (100, 95, 70, and 50%) and distilled water. Slides were then stained with hematoxylin and eosin (Scytek Laboratories Inc., Logan, UT) or trichrome stain (Abcam; Cambridge, MA) as per manufacturer’s instructions, or they were used for immunohistochemistry (IHC)/immunofluorescence (IF). For IHC, slides containing rehydrated tissue sections were peroxidase quenched (10 min in 10% hydrogen peroxide) and antigen retrieval was performed by boiling sections in a 0.1M sodium citrate buffer for 10 min. Upon cooling to room temperature, sections were blocked (0.1% bovine serum albumin, 0.1% normal goat serum, and 1% Triton-X in PBS) for one hour at room temperature. Sections were then incubated overnight at 4°C with primary antibody diluted in blocking solution as outlined in Table 2. Sections were washed in PBS 3X 10 min and incubated with secondary antibody for 45 min. Sections were washed as before and then incubated with horseradish peroxidase-conjugated streptavidin (Vector Laboratories, Burlingame, CA). Washes were then performed and the sections were exposed to 3,3’-diaminobenzidine (BD Biosciences, San Diego, CA) followed by a five-minute inactivation with PBS. Sections were then counterstained with methyl green, dehydrated and mounted. A similar approach was used for alpha-smooth muscle actin IF with the exception that the primary antibody was fluorescently tagged. All incubations were completed in a dark room. After the final wash (3X for 10 min) in PBS, mounting medium containing DAPI was applied and sections were cover slipped. Antibody specificity was established in negative control experiments where primary antibodies were omitted for all IHC and IF experiments.
Table 2.
Antibodies
| Target | Manufacturer, Catalog No. (Lot No.) |
Species, Clonal Status |
Dilution |
|---|---|---|---|
| Actin, alpha-Smooth Muscle |
Sigma-Aldrich, C6198 (042M4779) |
Mouse, monoclonal | 1:500 |
| ESR1 | Santa Cruz Biotech, SC-542 | Rabbit, polyclonal | 1:100 |
| PGR | Thermo Scientific, RM-9012 | Rabbit, monoclonal | 1:250 |
| PRKAA1 | Bioss, bs-1115R (120921) | Rabbit, polyclonal | 1:200 |
| PRKAA2 | Bioss, bs-2771R (130321) | Rabbit, polyclonal | 1:200 |
| pPRKAA1/2 | Cell Signaling, 2531 | Rabbit, polyclonal | 1:50 |
| Anti-rabbit IgG | Vector, BA-1000 (V0407) | Goat, polyclonal | 1:500 |
Data analyses
Animals were randomly assigned to the various treatment groups. All data are presented as the mean ± SEM for n=3–7 independent replicates. Individual animals represent a single experimental replicate within each experiment. Differences between treatment groups were established by Student’s t-test where the mean values of two groups were compared. A two-way analysis of variance was used to identify treatment effects in the breeding trials followed by a Bonferroni post-test. All data were analyzed using GraphPad 5.0 software (San Diego, CA) where a p-value of ≤0.05 was considered statistically significant.
RESULTS
PRKAA1, PRKAA2 and pPRKAA1/2 expression in the gravid murine uterus
As shown in Fig. 1, PRKAA1 and PRKAA2 are expressed throughout much of the implantation site on DOP 7.5, and this includes both the mesometrial and antimesometrial poles. While PRKAA1 was restricted primarily to the stromal cells, PRKAA2 was also robustly expressed in endothelial cells of the mesometrial vascular sinuses. Phospho-AMPK (i.e., pPRKAA1/2), as detected using an antibody against phospho-threonine 172 of both the PRKAA1 and PRKAA2 catalytic subunits, was weakly present throughout much of the deciduum. However, rich pPRKAA1/2 immunostaining was observed in the terminally differentiated decidual cells at the antimesometrial pole.
Figure 1. PRKAA1, PRKAA2 and pPRKAA1/2 expression in the gravid uterus.
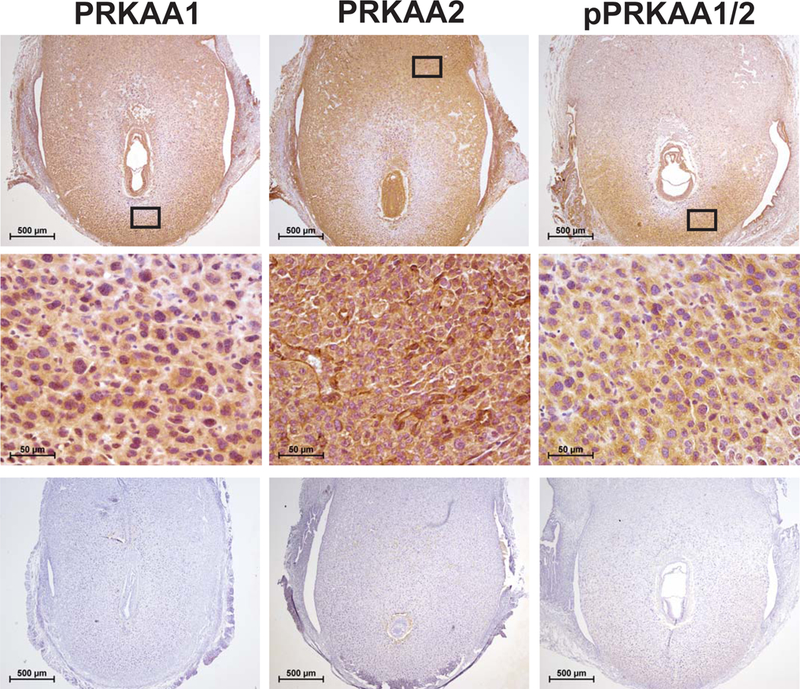
Pan-expression for the alpha catalytic subunits of AMPK (i.e., PRKAA1 and PRKAA2) on day of pregnancy (DOP) 7.5. Middle panels are higher magnification images of boxed areas in corresponding upper panels. Shown in the bottom panels are no primary antibody control sections, n=5
Conditional ablation of Prkaa1 and Prkaa2 causes progression to female infertility stemming from faulty endometrial regeneration
Prkaa1/2fl/fl and Prkaa1/2d/d mice were generated by crossing mice with floxed Prkaa1 and Prkaa2 alleles to mice expressing Pgr-cre. Genotyping was completed using tail snip DNA isolated from postnatal day (pnd) 19-21 mice (Fig. 2A and 2B). To confirm the conditional ablation of Prkaa1 and Prkaa2, mRNA was first isolated from female Prkaa1/2fl/fl and Prkaa1/2d/d livers and uteri. RT-qPCR revealed that while Prkaa1 and Prkaa2 expression did not differ between Prkaa1/2fl/fl and Prkaa1/2d/d livers, the expression of both genes was significantly reduced in uterine tissues obtained from Prkaa1/2fl/fl and Prkaa1/2d/d female mice confirming cre recombinase activity and genomic recombination (Fig. 2C and 2D). As shown in Fig. 3 the expression of Esr1 and Pgr mRNAs and ESR1 and PGR proteins were expressed at equitable levels in Prkaa1/2fl/fl and Prkaa1/2d/d female mice.
Figure 2. Validation of Prkaa1 and Prkaa2 ablation from the uterus.
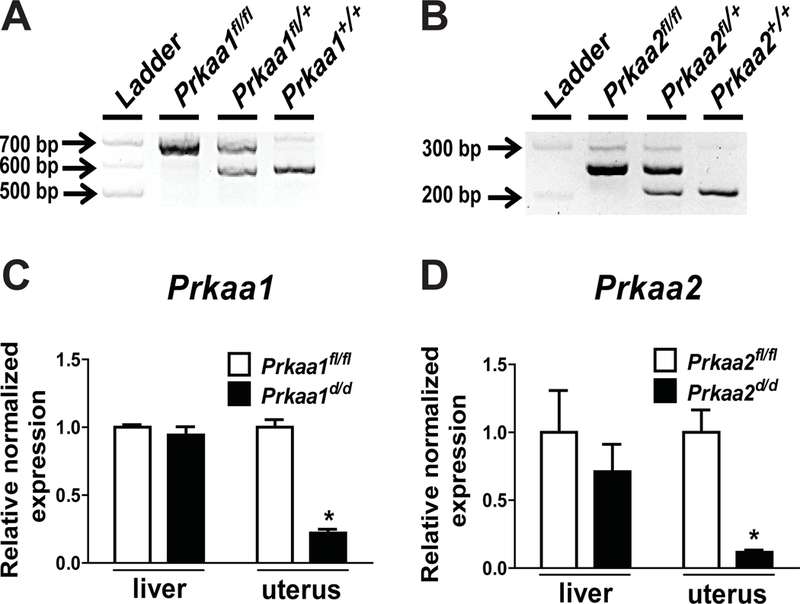
(A, B) Genotyping results identifying floxed (fl) and wild type (+) Prkaa1 and Prkaa2 alleles, respectively. (C, D) Relative normalized expression of Prkaa1 and Prkaa2 in livers and uteri obtained from Prkaa1/2fl/fl and Prkaa1/2d/d mice crossed with Pgr-cre driver mice as determined by RT-qPCR (n=4).
Figure 3. Equitable uterine expression of ESR1 and PGR in Prkaa1/2fl/fl and Prkaa1/2d/d female mice.
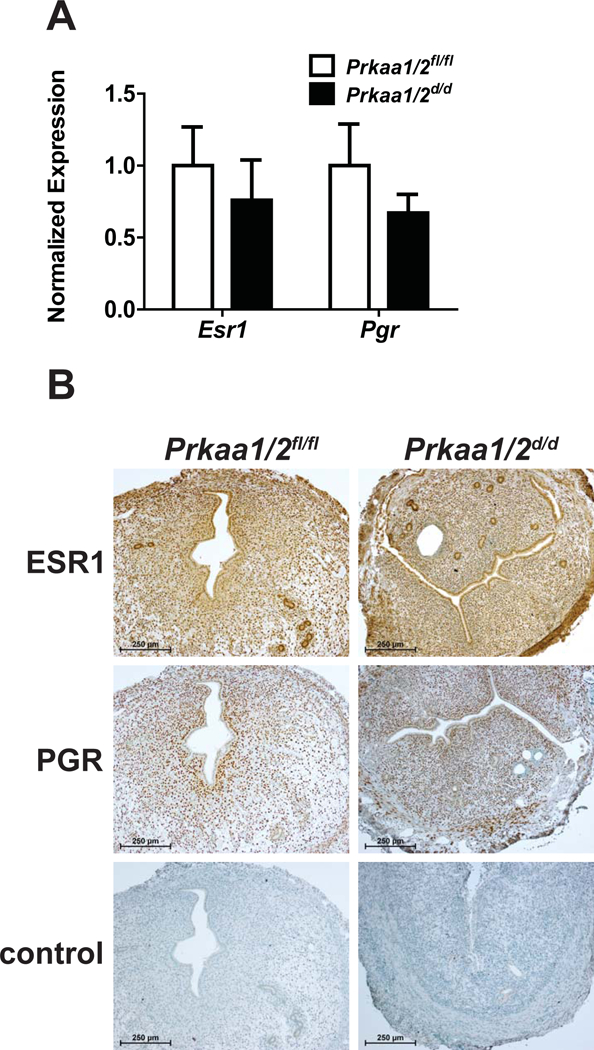
(A) RT-PCR was used to assess the expression of Esr1 and Pgr mRNAs in uteri obtained from synchronized (ovariectomy followed by estradiol treatment) Prkaa1/2fl/fl and Prkaa1/2d/d female mice (n=7). (B) ESR1 and PGR protein expression was also evaluated in uterine tissues obtained from Prkaa1/2fl/fl and Prkaa1/2d/d female mice. Here, uterine tissues were collected on DOPP 5, 24 h following oil induced decidualization as a means to synchronize mice (n=3–4).
We next completed a breeding trial to determine whether or not Prkaa1 and Prkaa2 are essential for female fertility. Cumulative breeding statistics are shown in Table 3. While there was no difference in the number of pups born in the first parity between Prkaa1/2fl/fl and Prkaa1/2d/d female mice (Fig. 4A; p=0.862), a significant and dramatic decline in pups born thereafter was noted in the Prkaa1/2d/d female mice (Fig. 4B; p=0.002). The parity interval increased greater than two-fold and mean litter size decreased significantly in Prkaa1/2d/d mice compared with Prkaa1/2fl/fl mice (Figs. 4C and4D). Complete reproductive senescence was observed in all Prkaa1/2d/d female mice after the fourth parity (Fig. 4E). Two Prkaa1/2d/d female mice were euthanized during parturition of the second and fourth pregnancy because of dystocia. No difference in estrous cycle length occurred between nulliparous Prkaa1/2fl/fl and Prkaa1/2d/d female mice (Fig. 4F), suggesting that the hypothalamic-pituitary-gonad axis was not affected by conditional ablation of Prkaa1 and Prkaa2 from these tissues. Furthermore, ovarian histology was consistent between 6–8 month old nulliparous Prkaa1/2fl/fl and Prkaa1/2d/d mice (Fig. 5). Similar findings were observed in ovaries obtained from multiparous Prkaa1/2fl/fl and Prkaa1/2d/d female mice at the end of the six-month breeding trial (Fig. 5).
Table 3.
Summary of the 6-month breeding trial
| Genotype | Females | Litters | Pups | Pups/litter | Litters/female |
|---|---|---|---|---|---|
|
| |||||
| Prkaa1/2 fl/fl | 4 | 24 | 159 | 6.300 ± 0.0752 | 6.000 ± 0.408 |
| Prkaa1/2 d/d | 4 | 12 | 49 | 4.563 ± 0.8683 | 3.000 ± 0.577 |
Figure 4. Conditional ablation of Prkaa1 and Prkaa2 results in premature reproductive senescence.
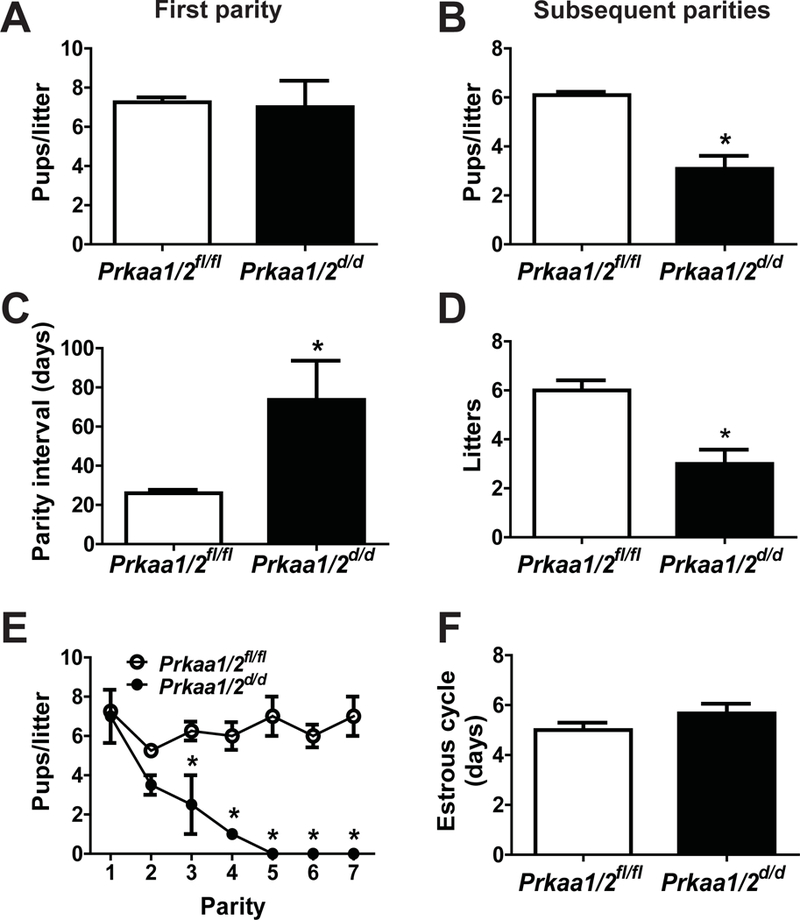
A six-month breeding trial was completed to assess the functional requirement of Prkaa1 and Prkaa2 for female fertility. The mean number of pups per first litter (A), the mean number of pups in all subsequent litters (B), the parity interval (C), the mean number of litters through the six-month trial (D), the mean number of pups/litter versus parity (E) and mean estrous cycle length (F) were determined for Prkaa1/2fl/fl and Prkaa1/2d/d mice. Values are expressed as mean ± SEM. *P≤0.05 compared with Prkaa1/2fl/fl control, n=3–5.
Figure 5. Prkaa1/2d/d mice have normal ovarian histology.

Shown are medial histological thin sections of ovaries obtained from nulliparous (nulli, 6–8 months of age) and multiparous (multi, 8 months of age at end of breeding trial) Prkaa1/2fl/fl and Prkaa1/2d/d mice (n=4–6).
The dramatic decline in fecundity after the first parity could be an age-dependent phenotype, or it could be explained by faulty regeneration of the uterus following parturition. To address this, nulliparous Prkaa1/2fl/fl and Prkaa1/2d/d female mice were aged to 4 months and then placed with males of proven breeding capacity. As shown in Fig. 6, there was no difference in the number of implantation sites (p=0.2782), the number of resorption sites (p=0.5945), or the mass of implantation sites (p=0.5897) between Prkaa1/2fl/fl and Prkaa1/2d/d female mice on DOP 10.5. Furthermore, serum P4 did not differ (p=0.5281) between these two genotypically distinct groups of mice providing further support that luteal function is not disrupted in Prkaa1/2d/d female mice. This was repeated with females at 6 months of age and again there is no difference in the number of implantation sites on DOP 7.5 (Fig. 6E).
Figure 6. Premature reproductive senescence is parity-dependent suggesting disrupted endometrial regeneration following parturition.
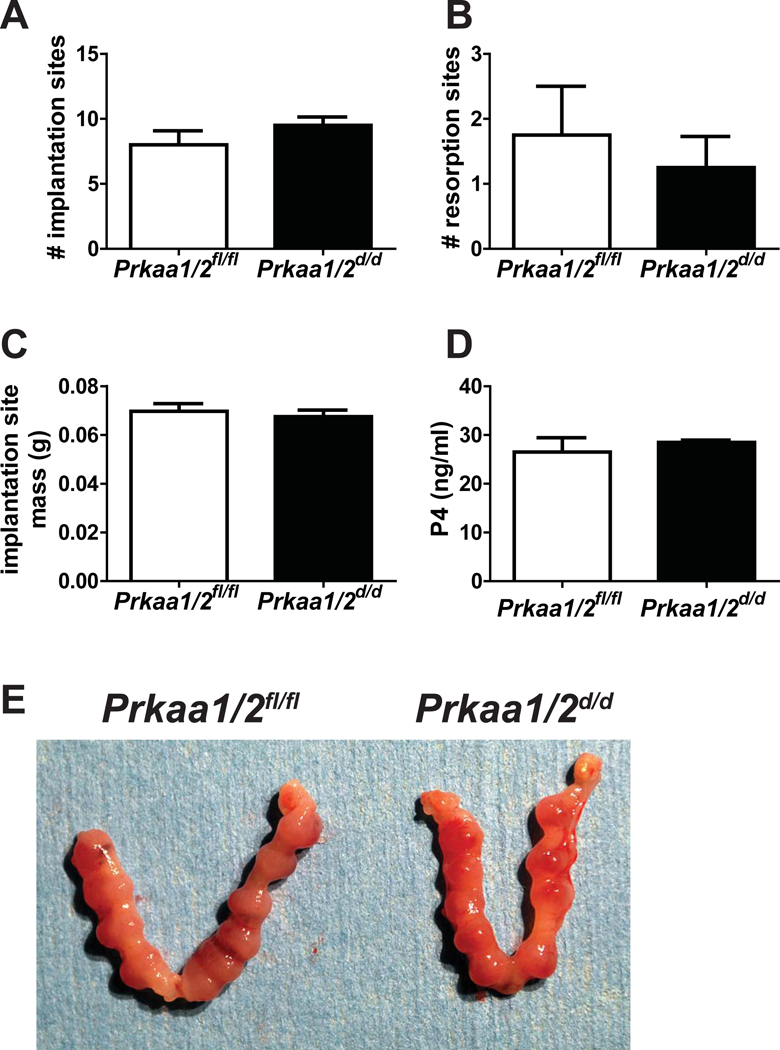
To determine if the observed premature reproductive senescence in Prkaa1/2d/d mice was due to parity or age, Prkaa1/2fl/fl and Prkaa1/2d/d female mice were aged to 4 months and then placed with males of proven breeding capacity. There was no difference in the number of implantation sites (A), the number of resorption sites (B), implantation site mass (C), or serum progesterone (P4, D) on DOP 10.5 (n=4). (E) Shown are uteri obtained from 6-month old Prkaa1/2fl/fl and Prkaa1/2d/d mice on DOP 7.5 (n=4). Values are expressed as mean ± SEM.
Results from the aging experiment prompted us to next evaluate the capacity for decidualization in Prkaa1/2d/d female mice. Here, 6–10 week old nulliparous Prkaa1/2fl/fl and Prkaa1/2d/d female mice placed with vasectomized males were induced to undergo decidualization on DOPP 4. As shown in Figure 7, artificial decidualization was completely absent in Prkaa1/2d/d female mice on DOPP 8 as shown grossly, by assessing uterine wet weight, histologically and by evaluating expression of decidualization markers Wingless-related murine mammary tumor virus integration site 4 (Wnt4) and bone morphogenetic protein 2 (Bmp2). Importantly, ovariectomized female mice treated with a hormone regimen consistent with early pregnancy showed similar results in that Prkaa1/2d/d female mice failed to decidualize (Fig. 7C), suggesting that the observed decidualization phenotype derives from faulty uterine function rather than ovarian dysregulation.
Figure 7. Uterine decidualization is compromised by Prkaa1 and Prkaa2 deficiency.
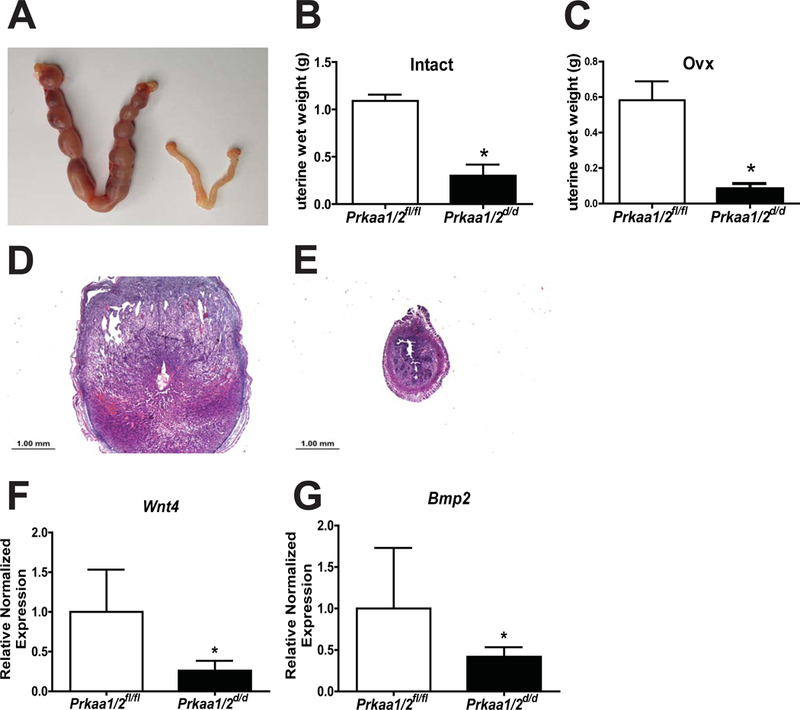
Artificial decidualization is blocked in intact Prkaa1/2d/d mice as shown by wet weight (A, B). Similar findings were observed when Prkaa1/2fl/fl and Prkaa1/2d/d mice were ovariectomized, given a steroid hormone consistent with early pregnancy, and artificially decidualized (C). This is supported in intact mice histologically (D, E) and in the expression of decidualization markers Wnt4 (F) and Bmp2 (G) on DOPP 8.5. Values are expressed as mean ± SEM, n=3–6. *P≤0.05 compared with Prkaa1/2fl/fl control.
Decidualization and regeneration following parturition both require extensive remodeling of the cellular composition and extracellular matrix of the endometrium. Given that each of these events becomes disrupted in Prkaa1/2d/d mice, we next completed a histological evaluation of uteri obtained from nulliparous and multiparous Prkaa1/2fl/fl and Prkaa1/2d/d female mice. Uteri from nulliparous Prkaa1/2fl/fl mice aged to 6–8 months showed expected histology that included increased glandular content with some enlarged glands (Fig 8). Uteri from nulliparous Prkaa1/2fl/fl and Prkaa1/2d/d mice have a well-defined lumen and distinct stromal and myometrial layers. In contrast, uteri collected from the multiparous animals displayed distinct differences (Fig. 8). Whereas uteri from multiparous Prkaa1/2fl/fl mice were overall very similar to uteri from aged nulliparous Prkaa1/2fl/fl mice, uteri from multiparous Prkaa1/2d/d mice had diminished luminal epithelial content and disorganized glandular tissue that was uncharacteristically present at both the antimesometrial and mesometrial aspects of the luminal compartment (Fig. 8). The stromal-myometrial interface was often indistinct and glandular tissue was routinely observed within intermixed stromal and myometrial tissues. Most noticeable was the reduced cellular content within the stromal compartment that seemed to be replaced by extracellular material. Two to nine large fibrotic lesions were identified in uterine sections obtained from multiparous, but not nulliparous, Prkaa1/2d/d mice (Fig. 9B). Many of these lesions were surrounded by hemosiderin positive cells (Fig. 9C). Unlike glandular epithelium of Prkaa1/2fl/fl mice, glands of Prkaa1/2d/d mice were commonly found to be surrounded by excessive fibrotic tissue indicative of endometritis, a condition that causes infertility in most mammals (Figs. 9D and9E). Polymorphonucleocytes could be found throughout the endometrium of multiparous Prkaa1/2d/d mice, particularly within the luminal compartment and underlying stromal tissue (Fig. 9F). An absence of α-smooth muscle actin staining confirmed that the fibrotic lesions were likely not leiomyomas or sarcomas (Fig. 9G). The acellular regions within the endometrium stained heavily with trichrome stain, suggesting high collagen content within the fibrotic lesions (Fig. 9H and9I).
Figure 8. Disorganization of the Prka1/2d/d endometrium and inner circular smooth muscle following parturition.
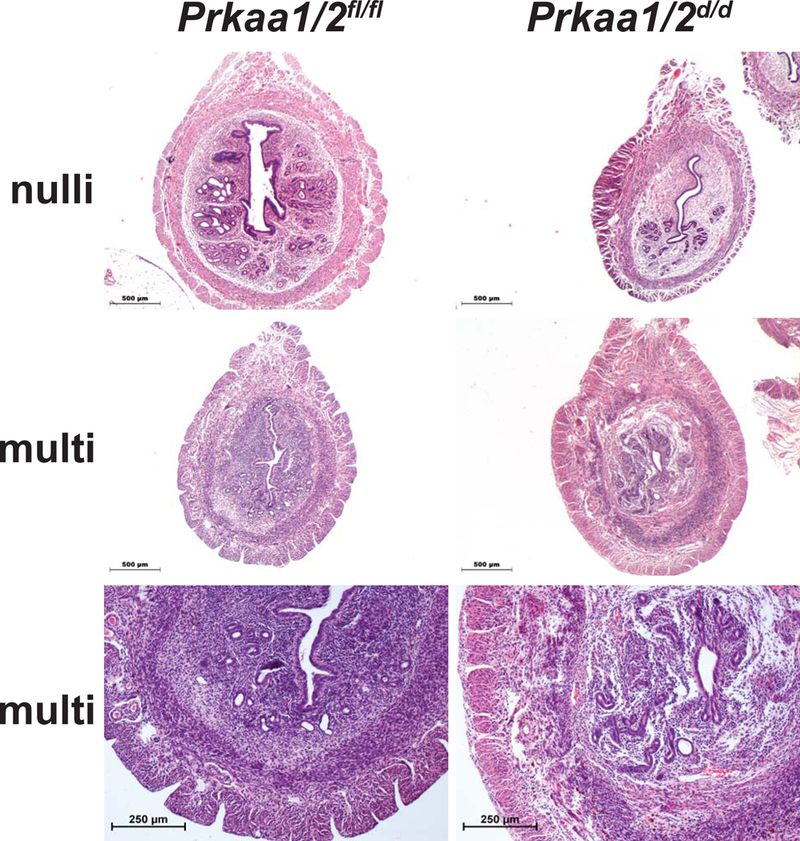
Representative images of uterine cross-sections from nulliparous (nulli; 6–8 months of age) and multiparous (multi; taken from breeding trial) Prkaa1/2fl/fl and Prkaa1/2d/d mice. At higher magnification, note the disorganized uterine histoarchitecture in multiparous Prkaa1/2d/d mice compared with Prkaa1/2fl/fl mice (bottom panels, n=4–8).
Figure 9. Uteri from Prkaa1/2d/d mice develop endometrial fibrotic lesions.
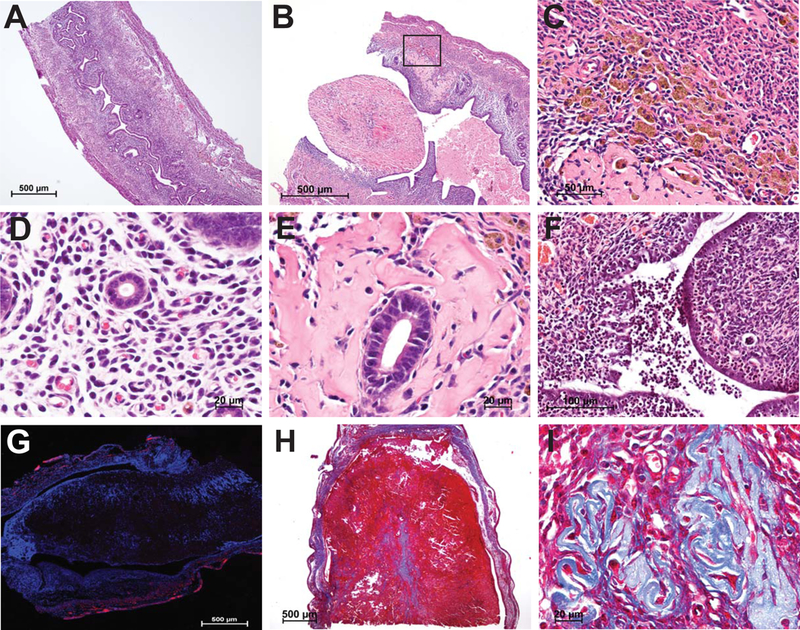
Compared with uteri from multiparous Prkaa1/2fl/fl female mice (A), endometrial fibrotic lesions are observed in Prkaa1/2d/d female mice (B), which are abundant in hemosiderin positive cells (C). Likewise, while uteri from multiparous Prkaa1/2fl/fl female mice show a close association of glandular epithelium and surrounding stroma (D), uteri from Prkaa1/2d/d female mice display with heavy circumferential glandular fibrosis indicative of endometritis (E). Furthermore, uteri from multiparous Prkaa1/2d/d female mice become infiltrated with polymorphonucleocytes (F) and develop large fibrotic α-smooth muscle actin negative (G; note immunofluorescent stain restricted to myometrial tissue) lesions that are rich in collagen fibers as evidenced in trichrome stained sections (H, I). n=3–8.
DISCUSSION
It was previously shown that phospho-PRKAA1/2 is regulated by sex steroid hormones in the uterus and is more abundant in the artificially decidualized uterus than in non-decidualized uterus (Kim & Moley 2009). Our studies extend these findings to the gravid uterus where PRKAA1 and PRKAA2 were robustly expressed at the maternal:embryo interface on DOP 7.5. While PRKAA1 is the more abundant isoform throughout much of the gravid uterus, PRKAA2 was heavily localized to the uterine vasculature with weaker staining throughout the stroma and myometrium. Interestingly, phospho-PRKAA1/2 immunostaining was most intense in the terminally differentiated cells of the antimesometrial pole, indicating an important role for AMPK in the decidualization process. More importantly, we established that uterine ablation of Prkaa1 and Prkaa2 resulted in reduced lifetime fecundity in the female due to premature reproductive senescence caused by faulty endometrial regeneration following parturition. In comparison to Prkaa1/2fl/fl female mice, which produced 6.3 pups/litter for each of six litters over a six-month period of time, Prkaa1/2d/d female mice produced only 4.6 pups/litter in each of three litters over the same period of time. All Prkaa1/2d/d female mice became infertile or were euthanized because of dystocia by the fourth parity. While a role for AMPK deficiency in dystocia has not yet been reported, previous findings demonstrate that AMPK activation attenuates heightened mTORC1 signaling present in mouse models of preterm birth and Trp53 conditional mutant mice that display a preterm birth phenotype (Hirota et al. 2011, Deng et al. 2016, Sun et al. 2018). We also noted an increase in parity interval. Given the fibrotic endometrial phenotype, we interpret this to mean that embryos are unable to properly implant into the scarred endometrium following the first parturition and only do so with delayed frequency after the endometrium has had additional time to repair. Additional studies are now needed to evaluate the role of individual Prkaa genes in female fertility.
Because the classical progesterone receptor (PGR) is expressed in gonadotrophs, granulosa cells of peri-ovulatory follicles and most of the cells of the female reproductive tract, it is likely that Prkaa1 and Prkaa2 are deleted from these tissues when crossed with Pgr-cre mice. Although additional studies are needed to evaluate the HPG axis in greater detail, four lines of indirect evidence suggest that the observed loss of fertility most likely stems from dysfunctional regeneration of the uterus. First, estrous cyclicity was not different between sexually mature Prkaa1/2fl/fl and Prkaa1/2d/d female mice, suggesting that the hypothalamic-pituitary-ovary (HPO) axis remained intact in Prkaa1/2d/d female mice and that Prkaa1 and Prkaa2 are dispensable for gonadotroph responses to GnRH such as the production and secretion of gonadotropins. Second, there were no histological differences in ovaries obtained from aged (6–8 months) nulliparous or multiparous Prkaa1/2fl/fl and Prkaa1/2d/d female mice. Here, all of the expected follicles at different stages of development, as well as corpora lutea were identified as histologically normal in both Prkaa1/2fl/fl and Prkaa1/2d/d female mice regardless of parity status or age. Third, serum progesterone levels were not different between Prkaa1/2fl/fl and Prkaa1/2d/d female mice on DOP 10.5. The nulliparous female mice used in this study were first aged to 4 months and then placed with males to establish pregnancy. This corresponds to the third parity of female mice placed in the breeding trial and a time marked by a significant decline in fecundity in Prkaa1/2d/d female mice. When combining the normal ovarian histology and progesterone production in Prkaa1/2d/d female mice, it is unlikely that the progressive subfertility phenotype is due to luteal insufficiency. Finally, to eliminate the possibility of an age-dependent decline in the functional activity of the HPO axis, two sets of nulliparous Prkaa1/2fl/fl and Prkaa1/2d/d female mice were aged to either 4 or 6 months before being placed with males of proven breeding capacity. We observed no difference in the number of implantation sites, resorption sites, implantation site mass, or serum P4 on DOP 7.5 or 10.5.
While the involvement of PRKAA1/2 deficiency in other reproductive cells that express Pgr-cre as contributing factors cannot be completely ruled out, the most likely explanation for the progressive fertility defect is faulty endometrial regeneration following parturition and development of severe endometrial fibrosis indicative of endometritis. Endometritis is a spectral condition that commonly results in infertility. Asherman’s Syndrome is an example of this in the human. Beyond serving as an immune privileged environment that gestates a histocompatibly unique embryo/fetus, the uterus has unique regenerative capacity distinct from most other mammalian tissues. Unlike most tissues where fibrosis is commonly associated with wound repair and regeneration, the endometrium undergoes natural cycles of degeneration and scar-free regeneration in menstrual cycling animals, as well as following parturition in all mammals. In fact, the regenerative process that follows menstruation is thought to precondition the uterus for subsequent successful pregnancy (Brosens et al. 2009). Interestingly, damaging the uterus of women with recurrent pregnancy loss during biopsy actually increases fertility rates, suggesting that regeneration is a natural process needed to prepare the endometrium for subsequent embryo implantation (Barash et al. 2003). This is thought to occur in response to inflammation that facilitates natural repair of the endometrium (Gnainsky et al. 2015). Through conditional mutagenesis studies or pharmacological activation, AMPK has been shown to reduce fibrosis and/or failure of scars to mature following injury in the liver (Ramezani-Moghadam et al. 2015, Yang et al. 2015, Chen et al. 2017, Liang et al. 2017), heart (Hermida et al. 2013, Noppe et al. 2014), lung (King et al. 2012, Sato et al. 2016, Tao et al. 2017), kidney (Kim et al. 2015, Thakur et al. 2015), intestine (Cieslik et al. 2017), peritoneum (Shin et al. 2017, Zheng et al. 2017), and adipose (Luo et al. 2016). Our findings place AMPK at the center of the unique regenerative process within the endometrium that is essential for maintenance of fertility in the female.
In addition to the well-established role of AMPK in regulating extracellular remodeling during tissue regeneration, AMPK also plays a prominent role in regulating glucose uptake in various tissues and the embryo (Kemp et al. 1999, Eng et al. 2007). By example, translocation of glucose transporters (i.e., GLUT1 and GLUT4) to the plasma membrane is mediated by AMPK in skeletal muscle and the heart (Russell et al. 1999, Li et al. 2004, Wu et al. 2013, Kjobsted et al. 2018). Given the presence of phospho-PRKAA1/2 in terminally differentiated stromal cells at the antimesometrial pole, AMPK may be involved in regulating glucose uptake or in how glucose is metabolized to generate ATP energy. In the presence of sufficient glucose, glycolysis is actually more efficient than oxidative phosphorylation in providing ATP energy and metabolic intermediates necessary for the biosynthesis of other macromolecules such as nucleotides, amino acids and fatty acids. The decidualization process, which is the terminal differentiation of endometrial stromal cells into mature decidual cells, is preceded by several rounds of proliferation. To satisfy the bioenergetic demands and provide the essential organic building blocks for generating macromolecules during proliferation and terminal differentiation, endometrial stromal cells transition from oxidative phosphorylation to glycolysis. Indeed, a Warburg-like glycolysis and lactate shuttle has been well-established during decidualization of early pregnancy (Zuo et al. 2015). This transition to glycolysis is accompanied by an acceleration in the glycolytic flux, which is fundamentally required for decidualization (Kommagani et al. 2013). Interestingly, a precedence for AMPK in stimulating Warburg-like glycolysis has been established (Teperino et al. 2012, Gingras & Pause 2014, Hart et al. 2015). By example, PRKAA1 is essential for satellite cell activation and transition to Warburg-like glycolysis during skeletal muscle regeneration (Fu et al. 2015).
As with endometrial regeneration, substantial remodeling of the endometrial extracellular compartment also accompanies decidualization. Here, much of the extracellular material is removed between stromal cells as they terminally differentiate into epithelial-like decidual cells. It is interesting to note that uterine decidualization was unaffected by PRKAA1/2 deficiency in the presence of the embryo during gestation. In contrast, artificial decidualization using mechanical stimulation was completely blocked in uteri from Prkaa1/2d/d female mice. We propose that AMPK plays a fundamental role in either remodeling the extracellular matrix as a component of decidualization or is necessary for stromal cell transdifferentiation. The presence of an embryo during pregnancy compensates for uterine PRKAA1/2 deficiency through an as-yet unknown mechanism. Other examples demonstrating an important role for embryonic signaling as a driver of decidualization exist. For instance, uterine ablation of Fkbp52 or Nr3c1 (i.e., glucocorticoid receptor) results in a similar phenotype where artificial decidualization fails, while natural decidualization in the presence of an embryo occurs, although at a reduced level (Tranguch et al. 2007, Whirledge et al. 2015). Here is yet another clear example demonstrating that it is the collective contributions of both the embryonic and maternal factors that are necessary for optimizing uterine remodeling during gestation. As a final note, obesity and fertility issues are both on the rise in the human population. One of the features of obesity is systemic diminished AMPK signaling. By example, obesity negatively impacts on proper murine skeletal muscle regeneration following damage through its inhibition of AMPK signaling (Fu et al. 2016). A clearer understanding of AMPK signaling at the maternal:embryo interface, as well as during endometrial regeneration, will likely enhance efforts to mitigate the negative impact of obesity on human pregnancy outcomes.
ACKNOWLEGMENTS
We thank The University of Virginia Center for Research in Reproduction Ligand Assay and Analysis Core, supported by the Eunice Kennedy Shriver NICHD/NIH P50 grant (HD28934), for performing the serum P4 hormone assays. We are grateful for funding support from the National Institutes of Health (OD016564 and HD086402).
Funding: NIH OD016564 and HD086402
Footnotes
Competing Financial Interests: The authors declare no competing or financial interests.
REFERENCES
- Barash A, Dekel N, Fieldust S, Segal I, Schechtman E & Granot I 2003. Local injury to the endometrium doubles the incidence of successful pregnancies in patients undergoing in vitro fertilization. Fertil Steril 79 1317–1322. [DOI] [PubMed] [Google Scholar]
- Bertoldo MJ, Faure M, Dupont J & Froment P 2015a. AMPK: a master energy regulator for gonadal function. Front Neurosci 9 235. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Bertoldo MJ, Guibert E, Faure M, Rame C, Foretz M, Viollet B, Dupont J & Froment P 2015b. Specific deletion of AMP-activated protein kinase (alpha1AMPK) in murine oocytes alters junctional protein expression and mitochondrial physiology. PLoS One 10 e0119680. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Bigham AW, Julian CG, Wilson MJ, Vargas E, Browne VA, Shriver MD & Moore LG 2014. Maternal PRKAA1 and EDNRA genotypes are associated with birth weight, and PRKAA1 with uterine artery diameter and metabolic homeostasis at high altitude. Physiol Genomics 46 687–697. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Boudaba N, Marion A, Huet C, Pierre R, Viollet B & Foretz M 2018. AMPK Re-Activation Suppresses Hepatic Steatosis but its Downregulation Does Not Promote Fatty Liver Development. EBioMedicine 28 194–209. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Brosens JJ, Parker MG, McIndoe A, Pijnenborg R & Brosens IA 2009. A role for menstruation in preconditioning the uterus for successful pregnancy. Am J Obstet Gynecol 200 615 e611–616. [DOI] [PubMed] [Google Scholar]
- Chen M, Liu J, Yang L & Ling W 2017. AMP-activated protein kinase regulates lipid metabolism and the fibrotic phenotype of hepatic stellate cells through inhibition of autophagy. FEBS Open Bio 7 811–820. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Cieslik KA, Trial J & Entman ML 2017. Aicar treatment reduces interstitial fibrosis in aging mice: Suppression of the inflammatory fibroblast. J Mol Cell Cardiol 111 81–85. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Deng W, Cha J, Yuan J, Haraguchi H, Bartos A, Leishman E, Viollet B, Bradshaw HB, Hirota Y & Dey SK 2016. p53 coordinates decidual sestrin 2/AMPK/mTORC1 signaling to govern parturition timing. J Clin Invest 126 2941–2954. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Eng GS, Sheridan RA, Wyman A, Chi MM, Bibee KP, Jungheim ES & Moley KH 2007. AMP kinase activation increases glucose uptake, decreases apoptosis, and improves pregnancy outcome in embryos exposed to high IGF-I concentrations. Diabetes 56 2228–2234. [DOI] [PubMed] [Google Scholar]
- Fu X, Zhu M, Zhang S, Foretz M, Viollet B & Du M 2016. Obesity Impairs Skeletal Muscle Regeneration Through Inhibition of AMPK. Diabetes 65 188–200. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Fu X, Zhu MJ, Dodson MV & Du M 2015. AMP-activated protein kinase stimulates Warburg-like glycolysis and activation of satellite cells during muscle regeneration. J Biol Chem 290 26445–26456. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Gingras MC & Pause A 2014. FLCN: A new regulator of AMPK-dependent Warburg metabolic reprogramming. Mol Cell Oncol 1 e961819. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Gnainsky Y, Granot I, Aldo P, Barash A, Or Y, Mor G & Dekel N 2015. Biopsy-induced inflammatory conditions improve endometrial receptivity: the mechanism of action. Reproduction 149 75–85. [DOI] [PubMed] [Google Scholar]
- Hardie DG, Hawley SA & Scott JW 2006. AMP-activated protein kinase--development of the energy sensor concept. J Physiol 574 7–15. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Hardie DG & Lin SC 2017. AMP-activated protein kinase - not just an energy sensor. F1000Res 6 1724. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Hart PC, Mao M, de Abreu AL, Ansenberger-Fricano K, Ekoue DN, Ganini D, Kajdacsy-Balla A, Diamond AM, Minshall RD, Consolaro ME, et al. 2015. MnSOD upregulation sustains the Warburg effect via mitochondrial ROS and AMPK-dependent signalling in cancer. Nat Commun 6 6053. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Hawley SA, Boudeau J, Reid JL, Mustard KJ, Udd L, Makela TP, Alessi DR & Hardie DG 2003. Complexes between the LKB1 tumor suppressor, STRAD alpha/beta and MO25 alpha/beta are upstream kinases in the AMP-activated protein kinase cascade. J Biol 2 28. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Hawley SA, Pan DA, Mustard KJ, Ross L, Bain J, Edelman AM, Frenguelli BG & Hardie DG 2005. Calmodulin-dependent protein kinase kinase-beta is an alternative upstream kinase for AMP-activated protein kinase. Cell Metab 2 9–19. [DOI] [PubMed] [Google Scholar]
- Hermida N, Markl A, Hamelet J, Van Assche T, Vanderper A, Herijgers P, van Bilsen M, Hilfiker-Kleiner D, Noppe G, Beauloye C, et al. 2013. HMGCoA reductase inhibition reverses myocardial fibrosis and diastolic dysfunction through AMP-activated protein kinase activation in a mouse model of metabolic syndrome. Cardiovasc Res 99 44–54. [DOI] [PubMed] [Google Scholar]
- Herzig S & Shaw RJ 2018. AMPK: guardian of metabolism and mitochondrial homeostasis. Nat Rev Mol Cell Biol 19 121–135. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Hirota Y, Cha J, Yoshie M, Daikoku T & Dey SK 2011. Heightened uterine mammalian target of rapamycin complex 1 (mTORC1) signaling provokes preterm birth in mice. Proc Natl Acad Sci U S A 108 18073–18078. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Horman S, Beauloye C, Vanoverschelde JL & Bertrand L 2012. AMP-activated protein kinase in the control of cardiac metabolism and remodeling. Curr Heart Fail Rep 9 164–173. [DOI] [PubMed] [Google Scholar]
- Jiang S, Li T, Yang Z, Yi W, Di S, Sun Y, Wang D & Yang Y 2017. AMPK orchestrates an elaborate cascade protecting tissue from fibrosis and aging. Ageing Res Rev 38 18–27. [DOI] [PubMed] [Google Scholar]
- Kemp BE, Mitchelhill KI, Stapleton D, Michell BJ, Chen ZP & Witters LA 1999. Dealing with energy demand: the AMP-activated protein kinase. Trends Biochem Sci 24 22–25. [DOI] [PubMed] [Google Scholar]
- Kim H, Moon SY, Kim JS, Baek CH, Kim M, Min JY & Lee SK 2015. Activation of AMP-activated protein kinase inhibits ER stress and renal fibrosis. Am J Physiol Renal Physiol 308 F226–236. [DOI] [PubMed] [Google Scholar]
- Kim ST & Moley KH 2009. Regulation of facilitative glucose transporters and AKT/MAPK/PRKAA signaling via estradiol and progesterone in the mouse uterine epithelium. Biol Reprod 81 188–198. [DOI] [PMC free article] [PubMed] [Google Scholar]
- King JD Jr., Lee J, Riemen CE, Neumann D, Xiong S, Foskett JK, Mehta A, Muimo R & Hallows KR 2012. Role of binding and nucleoside diphosphate kinase A in the regulation of the cystic fibrosis transmembrane conductance regulator by AMP-activated protein kinase. J Biol Chem 287 33389–33400. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kjobsted R, Hingst JR, Fentz J, Foretz M, Sanz MN, Pehmoller C, Shum M, Marette A, Mounier R, Treebak JT, et al. 2018. AMPK in skeletal muscle function and metabolism. FASEB J 32 1741–1777. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kommagani R, Szwarc MM, Kovanci E, Gibbons WE, Putluri N, Maity S, Creighton CJ, Sreekumar A, DeMayo FJ, Lydon JP, et al. 2013. Acceleration of the glycolytic flux by steroid receptor coactivator-2 is essential for endometrial decidualization. PLoS Genet 9 e1003900. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Li J, Hu X, Selvakumar P, Russell RR 3rd, Cushman SW, Holman GD & Young LH 2004. Role of the nitric oxide pathway in AMPK-mediated glucose uptake and GLUT4 translocation in heart muscle. Am J Physiol Endocrinol Metab 287 E834–841. [DOI] [PubMed] [Google Scholar]
- Liang Z, Li T, Jiang S, Xu J, Di W, Yang Z, Hu W & Yang Y 2017. AMPK: a novel target for treating hepatic fibrosis. Oncotarget 8 62780–62792. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Luo T, Nocon A, Fry J, Sherban A, Rui X, Jiang B, Xu XJ, Han J, Yan Y, Yang Q, et al. 2016. AMPK Activation by Metformin Suppresses Abnormal Extracellular Matrix Remodeling in Adipose Tissue and Ameliorates Insulin Resistance in Obesity. Diabetes 65 2295–2310. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Mayes MA, Laforest MF, Guillemette C, Gilchrist RB & Richard FJ 2007. Adenosine 5’-monophosphate kinase-activated protein kinase (PRKA) activators delay meiotic resumption in porcine oocytes. Biol Reprod 76 589–597. [DOI] [PubMed] [Google Scholar]
- Miller RA, Chu Q, Le Lay J, Scherer PE, Ahima RS, Kaestner KH, Foretz M, Viollet B & Birnbaum MJ 2011. Adiponectin suppresses gluconeogenic gene expression in mouse hepatocytes independent of LKB1-AMPK signaling. J Clin Invest 121 2518–2528. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Mounier R, Theret M, Lantier L, Foretz M & Viollet B 2015. Expanding roles for AMPK in skeletal muscle plasticity. Trends Endocrinol Metab 26 275–286. [DOI] [PubMed] [Google Scholar]
- Noppe G, Dufeys C, Buchlin P, Marquet N, Castanares-Zapatero D, Balteau M, Hermida N, Bouzin C, Esfahani H, Viollet B, et al. 2014. Reduced scar maturation and contractility lead to exaggerated left ventricular dilation after myocardial infarction in mice lacking AMPKalpha1. J Mol Cell Cardiol 74 32–43. [DOI] [PubMed] [Google Scholar]
- O’Neill HM, Holloway GP & Steinberg GR 2013. AMPK regulation of fatty acid metabolism and mitochondrial biogenesis: implications for obesity. Mol Cell Endocrinol 366 135–151. [DOI] [PubMed] [Google Scholar]
- Rabinovitch RC, Samborska B, Faubert B, Ma EH, Gravel SP, Andrzejewski S, Raissi TC, Pause A, St-Pierre J & Jones RG 2017. AMPK Maintains Cellular Metabolic Homeostasis through Regulation of Mitochondrial Reactive Oxygen Species. Cell Rep 21 1–9. [DOI] [PubMed] [Google Scholar]
- Ramezani-Moghadam M, Wang J, Ho V, Iseli TJ, Alzahrani B, Xu A, Van der Poorten D, Qiao L, George J & Hebbard L 2015. Adiponectin reduces hepatic stellate cell migration by promoting tissue inhibitor of metalloproteinase-1 (TIMP-1) secretion. J Biol Chem 290 5533–5542. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Russell RR 3rd, Bergeron R, Shulman GI & Young LH 1999. Translocation of myocardial GLUT-4 and increased glucose uptake through activation of AMPK by AICAR. Am J Physiol 277 H643–649. [DOI] [PubMed] [Google Scholar]
- Santiquet N, Sasseville M, Laforest M, Guillemette C, Gilchrist RB & Richard FJ 2014. Activation of 5’ adenosine monophosphate-activated protein kinase blocks cumulus cell expansion through inhibition of protein synthesis during in vitro maturation in Swine. Biol Reprod 91 51. [DOI] [PubMed] [Google Scholar]
- Sato N, Takasaka N, Yoshida M, Tsubouchi K, Minagawa S, Araya J, Saito N, Fujita Y, Kurita Y, Kobayashi K, et al. 2016. Metformin attenuates lung fibrosis development via NOX4 suppression. Respir Res 17 107. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Shaw RJ, Kosmatka M, Bardeesy N, Hurley RL, Witters LA, DePinho RA & Cantley LC 2004. The tumor suppressor LKB1 kinase directly activates AMP-activated kinase and regulates apoptosis in response to energy stress. Proc Natl Acad Sci U S A 101 3329–3335. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Shin HS, Ko J, Kim DA, Ryu ES, Ryu HM, Park SH, Kim YL, Oh ES & Kang DH 2017. Metformin ameliorates the Phenotype Transition of Peritoneal Mesothelial Cells and Peritoneal Fibrosis via a modulation of Oxidative Stress. Sci Rep 7 5690. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Shirwany NA & Zou MH 2014. AMPK: a cellular metabolic and redox sensor. A minireview. Front Biosci (Landmark Ed) 19 447–474. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Soyal SM, Mukherjee A, Lee KY, Li J, Li H, DeMayo FJ & Lydon JP 2005. Cre-mediated recombination in cell lineages that express the progesterone receptor. Genesis 41 58–66. [DOI] [PubMed] [Google Scholar]
- Sun X, Tavenier A, Deng W, Leishman E, Bradshaw HB & Dey SK 2018. Metformin attenuates susceptibility to inflammation-induced preterm birth in mice with higher endocannabinoid levels. Biol Reprod 98 208–217. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Tao L, Cao J, Wei W, Xie H, Zhang M & Zhang C 2017. Protective role of rhapontin in experimental pulmonary fibrosis in vitro and in vivo. Int Immunopharmacol 47 38–46. [DOI] [PubMed] [Google Scholar]
- Teperino R, Amann S, Bayer M, McGee SL, Loipetzberger A, Connor T, Jaeger C, Kammerer B, Winter L, Wiche G, et al. 2012. Hedgehog partial agonism drives Warburg-like metabolism in muscle and brown fat. Cell 151 414–426. [DOI] [PubMed] [Google Scholar]
- Thakur S, Viswanadhapalli S, Kopp JB, Shi Q, Barnes JL, Block K, Gorin Y & Abboud HE 2015. Activation of AMP-activated protein kinase prevents TGF-beta1-induced epithelial-mesenchymal transition and myofibroblast activation. Am J Pathol 185 2168–2180. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Tosca L, Chabrolle C, Uzbekova S & Dupont J 2007a. Effects of metformin on bovine granulosa cells steroidogenesis: possible involvement of adenosine 5’ monophosphate-activated protein kinase (AMPK). Biol Reprod 76 368–378. [DOI] [PubMed] [Google Scholar]
- Tosca L, Froment P, Solnais P, Ferre P, Foufelle F & Dupont J 2005. Adenosine 5’-monophosphate-activated protein kinase regulates progesterone secretion in rat granulosa cells. Endocrinology 146 4500–4513. [DOI] [PubMed] [Google Scholar]
- Tosca L, Rame C, Chabrolle C, Tesseraud S & Dupont J 2010. Metformin decreases IGF1-induced cell proliferation and protein synthesis through AMP-activated protein kinase in cultured bovine granulosa cells. Reproduction 139 409–418. [DOI] [PubMed] [Google Scholar]
- Tosca L, Solnais P, Ferre P, Foufelle F & Dupont J 2006. Metformin-induced stimulation of adenosine 5’ monophosphate-activated protein kinase (PRKA) impairs progesterone secretion in rat granulosa cells. Biol Reprod 75 342–351. [DOI] [PubMed] [Google Scholar]
- Tosca L, Uzbekova S, Chabrolle C & Dupont J 2007b. Possible role of 5’AMP-activated protein kinase in the metformin-mediated arrest of bovine oocytes at the germinal vesicle stage during in vitro maturation. Biol Reprod 77 452–465. [DOI] [PubMed] [Google Scholar]
- Tranguch S, Wang H, Daikoku T, Xie H, Smith DF & Dey SK 2007 FKBP52 deficiency-conferred uterine progesterone resistance is genetic background and pregnancy stage specific. J Clin Invest 117 1824–1834. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Viollet B, Andreelli F, Jorgensen SB, Perrin C, Geloen A, Flamez D, Mu J, Lenzner C, Baud O, Bennoun M, et al. 2003. The AMP-activated protein kinase alpha2 catalytic subunit controls whole-body insulin sensitivity. J Clin Invest 111 91–98. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Viollet B, Athea Y, Mounier R, Guigas B, Zarrinpashneh E, Horman S, Lantier L, Hebrard S, Devin-Leclerc J, Beauloye C, et al. 2009a. AMPK: Lessons from transgenic and knockout animals. Front Biosci (Landmark Ed) 14 19–44. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Viollet B & Foretz M 2016. Animal Models to Study AMPK. EXS 107 441–469. [DOI] [PubMed] [Google Scholar]
- Viollet B, Guigas B, Leclerc J, Hebrard S, Lantier L, Mounier R, Andreelli F & Foretz M 2009b. AMP-activated protein kinase in the regulation of hepatic energy metabolism: from physiology to therapeutic perspectives. Acta Physiol (Oxf) 196 81–98. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Whirledge SD, Oakley RH, Myers PH, Lydon JP, DeMayo F & Cidlowski JA 2015. Uterine glucocorticoid receptors are critical for fertility in mice through control of embryo implantation and decidualization. Proc Natl Acad Sci U S A 112 15166–15171. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Wu N, Zheng B, Shaywitz A, Dagon Y, Tower C, Bellinger G, Shen CH, Wen J, Asara J, McGraw TE, et al. 2013. AMPK-dependent degradation of TXNIP upon energy stress leads to enhanced glucose uptake via GLUT1. Mol Cell 49 1167–1175. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Xie M, Zhang D, Dyck JR, Li Y, Zhang H, Morishima M, Mann DL, Taffet GE, Baldini A, Khoury DS, et al. 2006. A pivotal role for endogenous TGF-beta-activated kinase-1 in the LKB1/AMP-activated protein kinase energy-sensor pathway. Proc Natl Acad Sci U S A 103 17378–17383. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Yang Y, Zhao Z, Liu Y, Kang X, Zhang H & Meng M 2015. Suppression of oxidative stress and improvement of liver functions in mice by ursolic acid via LKB1-AMP-activated protein kinase signaling. J Gastroenterol Hepatol 30 609–618. [DOI] [PubMed] [Google Scholar]
- Zheng W, Song J, Zhang Y, Chen S, Ruan H & Fan C 2017. Metformin prevents peritendinous fibrosis by inhibiting transforming growth factor-beta signaling. Oncotarget 8 101784–101794. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Zuo RJ, Gu XW, Qi QR, Wang TS, Zhao XY, Liu JL & Yang ZM 2015. Warburg-like Glycolysis and Lactate Shuttle in Mouse Decidua during Early Pregnancy. J Biol Chem 290 21280–21291. [DOI] [PMC free article] [PubMed] [Google Scholar]