

Abstract
Currently available antiepileptic drugs are effective; however, frequently associated with adverse effects that limit their therapeutic value. Compounds that target the molecular events underlying epilepsy, with minor or no adverse effects, would be of clinical value. Matrix metalloproteinase-9 (MMP-9) and the brain-derived neurotrophic factor (BDNF)-tropomyosin receptor kinase B (TrkB) signaling pathway may be involved in epileptogenesis. The current study investigated the effects of the plant-derived hydroxyflavone, myricetin, in a pentylenetetrazole (PTZ)-induced mouse model of epilepsy. Mice received an intraperitoneal injection of 35 mg/kg body weight PTZ on alternate days (13 injections) and were observed for 30 min following each PTZ injection. Myricetin (100 or 200 mg/kg body weight) was administered orally to the treatment groups (n=18/group) for 26 days, 30 min prior to each PTZ injection. Treatment with myricetin reduced seizure and mortality rates. Increased apoptotic cell count and elevated expression levels of apoptotic proteins caused by PTZ kindling were downregulated following treatment with myricetin. The BDNF-TrkB signaling pathway and MMP-9 expression levels were regulated by myricetin. Expression of γ-aminobutyric acid A (GABA) receptor and glutamic acid decarboxylase 65, as well as the glutamate/GABA balance, were restored following treatment with myricetin. The results of the present study indicated that myricetin may exert protective effects by regulating the molecular events associated with epileptogenesis.
Keywords: brain-derived neurotrophic factor-tropomyosin receptor kinase B pathway, epilepsy, γ-aminobutyric acid, matrix metalloproteinase, pentylenetetrazole
Introduction
Epilepsy is a neurological disorder with a complex etiology (1). Recurrent seizures in epilepsy are caused by an imbalance between cerebral excitability and inhibition (2,3). Epilepsy may be either symptomatic of an underlying disorder or idiopathic (4,5). Despite treatment with currently available antiepileptic drugs (AEDs), epilepsy remains refractory in 30% of patients (6). Additionally, previous studies have reported that recurring seizures, cognitive disturbance and psychiatric issues are associated with AEDs (7,8). Therefore, novel drugs that suppress seizures more efficiently and exert no or minor adverse effects are required.
Previous research has focused on the antiepileptic effects of plant-derived compounds (6,9). Myricetin, 3,3′,4′,4,5,5′,7-hexahydroxyflavone, is a flavonoid commonly extracted from the fruits, leaves and bark of the Chinese bayberry (Myrica rubra) and other edible plants, including broccoli, cabbage, French beans, garlic, peppers, tomato, cashew nuts, blueberries, and green and black tea (10–13). Myricetin exhibits several bioactive properties, including antioxidative (11) anticancer (14,15), nephroprotective (16) and anti-inflammatory effects (17). However, myricetin has not been previously evaluated for its potential antiepileptic effects. Therefore, the current study investigated whether myricetin exerts protective effects against experimentally induced epilepsy.
Brain-derived neurotrophic factor (BDNF) and its receptor tropomyosin receptor kinase B (TrkB) regulate the processes of synaptic plasticity and neurogenesis (18). The BDNF-TrkB signaling pathway serves a role in epileptogenesis (19,20). Increased expression levels of BDNF have been reported in epileptogenic conditions (21). Furthermore, a previous study used BDNF infusion into the hippocampus to induce seizures (22). The BDNF-TrkB signaling targets several downstream protein kinases to activate target proteins, leading to increased presynaptic release of mediators or modification of the activity of postsynaptic receptors, including the γ-aminobutyric acid A receptor (GABAAR) (23). GABA and GABAAR mediate inhibitory neurotransmission and attenuate overexcitation of neurons (24). Glutamic acid decarboxylase 65 (GAD65) is an enzyme catalyzing the conversion of glutamate to GABA via decarboxylation (25). Previous studies have demonstrated that the expression levels of GAD and GABA receptors in the brain are closely associated with epileptic conditions (26,27).
Matrix metalloproteinases (MMPs) serve roles in normal developmental processes, including tissue morphogenesis and cell migration, and are associated pathological conditions, including inflammation, metastasis, Alzheimer's disease, cerebral aneurysm and brain cancer (28,29). Furthermore, a previous study demonstrated an association between the expression of MMPs and epilepsy and epileptogenesis (30). Activation of MMP-2 and −9 was observed in the brains of rats with kainic acid (KA)-induced seizures (31). MMP-9 induces apoptosis by disrupting integrin-mediated cell survival signal in the pilocarpine-induced model of epilepsy (32). MMP-9 is also involved in tissue remodeling within the epileptic focus following seizures (33). Based on the association between the BDNF-TrkB signaling and MMP-9 in epilepsy, the current study further investigated whether myricetin reduced seizure rates and regulated MMP-9 expression and BDNF-TrkB pathway activity in a pentylenetetrazole (PTZ)-induced experimental model of epilepsy.
Materials and methods
Chemicals and reagents
PTZ, MMP-9 and myricetin were purchased from Sigma-Aldrich (Merck KGaA, Darmstadt, Germany). PTZ was dissolved in freshly prepared saline prior to treatment (100 mg/ml). Antibodies against cleaved caspase-3 (cat. no. 9661; 1:1,000; Cell Signaling Technology, Inc., Danvers, MA, USA), BDNF (cat. no. PA5-15198: 1:2,000; Invitrogen; Thermo Fishes Scientific, Inc., Waltham, MA, USA), TrkB (cat. no. 4606; 1:1,000; Cell Signaling Technology, Inc.), GABAAR (cat. no. PA5-78386; 1:1,000; Invitrogen; Thermo Fisher Scientific, Inc.), MMP-9 (cat. no. 3852; 1:1,000; Cell Signaling Technology, Inc.) and GAD65 (cat. no. PA5-22260; 1:2,000; Invitrogen; Thermo Fisher Scientific, Inc.) were obtained and Santa Cruz Biotechnology, Inc. (Dallas, TX, USA) provided antibodies against apoptosis regulator B-cell lymphoma-2 (Bcl-2; cat. no. sc509; 1:1,000), apoptosis regulator Bcl-2-like protein 4 (Bax; cat. no. sc-7480; 1:1,000), Bcl2-associated agonist of cell death (Bad; cat. no. sc-8004; 1:1,000), anti-apoptotic regulator Bcl extra large (Bcl-xL; cat. no. sc-136132; 1:1,000) and β-actin (cat. no. sc-47778; 1:1,000). All other standard chemicals used in the current study were purchased from Sigma-Aldrich; Merck KGaA.
Animal experiments
C57BL/6 male mice (n=9; weight, 20±2 g; age, 4–6 weeks) were obtained from the animal house of Hedong People's Hospital (Linyi, China) and housed under standard laboratory conditions (temperature, 23±1°C; humidity 50–60%; 12-h light/dark cycle). Mice had access to standard pellet diet and water ad libitum, and were allowed to acclimate to the laboratory conditions for 4–5 days prior to the experiments. The current study was approved by the Animal Studies Ethics Committee of Hedong People's Hospital and the protocols and handling of animals were in compliance with National Institutes of Health guidelines for the care and use of laboratory animals and the National Animal Welfare Law of China (34–36).
Animal kindling
Kindling is an experimental technique used to study epileptogenesis and epilepsy-associated cognitive and behavioral alterations, and to assess the efficacy of novel antiepileptic drugs (37,38). In the current study, the animals were given a total of 13 intraperitoneal (i.p.) injections of sub-effective dose of PTZ (35 mg/kg) on alternate days (9). The animals were observed for 30 min following each PTZ injection. Separate treatment groups (n=18/group) received myricetin at 100 or 200 mg/kg body weight, orally for 26 days, 30 min prior to each PTZ injection; the concentrations used in the study were based on the results of previous experiments using various concentrations of myricetin (data not shown). Control animals were treated with an equal volume of saline instead of administration of PTZ or myricetin. Epileptic control mice were treated with PTZ, but not treated with myricetin. Standard drug control mice were treated with PTZ and received valproate (100 mg/kg; i.p.) (6). The mice were anesthetized with isoflurane and perfused transcardially with 0.9% saline (100 ml), followed by 4% paraformaldehyde (200 ml) and sacrificed 24 h after the last PTZ challenge. The brain tissues were immediately collected and analyzed.
Seizure scoring
The mice were monitored for 30 min after each PTZ injection. Seizures were classified and scored according to the modified Racine scale (39): 0, no response; phase 1, hyperactivity and vibrissae twitching; phase 2, head nodding, head clonus and myoclonic jerk; phase 3, unilateral forelimb clonus; phase 4, rearing with bilateral forelimb clonus; phase 5, generalized tonic-clonic seizure with loss of postural control; and phase 6, absence of movement and loss of function of vital organs/mortality. Mortality rate was determined using the following formula: Mortality count/total number of animals in the particular experimental group ×100.
Histological Nissl staining
Nissl staining was performed to assess the effect of myricetin on the alterations in brain architecture following PTZ challenge. Coronal sections (7-µm-thick) were subjected to Nissl staining with toluidine blue (1%), as previously described (40). Viable cells in the CA1 and CA3 regions of the hippocampus per 1-mm length of bilateral hemisphere were counted under high magnification (×400) using a light microscope (Nikon Corporation, Tokyo, Japan).
Evaluation of neuroapoptosis by the terminal deoxynucleotidyl-transferase-mediated dUTP nick end labeling (TUNEL) assay
Neuroapoptosis was assessed using the TUNEL assay, as previously described by Li et al (41). Brain tissue sections (5-µm-thick) were sliced in the same plane of the hippocampal region of each mouse and subjected to analysis using the DeadEnd™ fluorometric TUNEL system (Promega Corporation, Madison, WI, USA). Tissue sections were protected from direct light during the assay. TUNEL-positive cells in the CA1 and CA3 regions of the hippocampus were observed and analyzed using NIS-Elements Basic Research microscope imaging software (version, 4.0; Nikon Corporation).
Analysis of brain GABA and glutamate levels
The mice were sacrificed as described above 50 min after the last PTZ injection. Brain tissues were isolated immediately, weighed and homogenized in 0.1 M PBS, and GABA and glutamate contents in the tissues were analyzed by high performance liquid chromatography with reference standards as described previously (42).
Reverse transcription-polymerase chain reaction (RT-PCR)
An RT-PCR analysis was performed to determine the effects of myricetin on MMP-9, BDNF and TrkB mRNA expression. Total RNA was extracted from the hippocampal tissues using TRIzol reagent (Invitrogen; Thermo Fisher Scientific, Inc., Waltham, MA, USA). First-strand cDNA was synthesized using the Revert Aid First Strand cDNA Synthesis kit (Fermentas; Thermo Fisher Scientific, Inc., Waltham, MA, USA). PCR reactions were performed using the 7300 Real-Time PCR system (Applied Biosystems; Thermo Fisher Scientific, Inc., Waltham, MA, USA). The following primers were used: Forward, 5′-CGAAGAGCTGCTGGATGAG-3′, and reverse, 5′-ATGGGATTACACTTGGTCTCG-3′, for BDNF; forward, 5′-CCTCCACGGATGTTGCTGA-3′, and reverse, 5′-GGCTGTTGGTGATACCGAAGTA-3′ for TrkB; forward, 5′-GTCTTCCCCTTCGTCTTCCT-3′, and reverse, 5′-GCTGGATGCCTTTTATGTCG-3′ for MMP-9; forward, 5′-CCGTATCGGACGCCTGGTTA-3′, and reverse, 5′-GGCTGTTGGTGATACCGAAGTA-3′, for GAPDH. GAPDH was used as the internal control. PCR products were separated by 1% agarose gel electrophoresis and visualized using ethidium bromide (0.05%) staining. The band intensities of the products were analyzed with the Bio-Gel imagery apparatus with Quantity One® 1-D analysis software (4.3.0; Bio-Rad Laboratories, Inc., Hercules, CA, USA).
Gel zymography
Gel zymography was performed to evaluate MMP-9 activity. The samples were prepared as previously described by Mizoguchi et al (43). The samples were subjected to SDS-PAGE using 10% gels containing 0.1% gelatin under non-reducing conditions. Triton X-100 gels (2.5%) were washed to remove the SDS, and further washed in incubation buffer (50 mM Tris-HCl, pH 7.4; 5 mM CaCl2; 200 mM NaCl and 2 µM ZnCl2) at 25±2°C for 30 min and subsequently incubated for 24 h at 37°C. Following the incubation, the gels were stained with Coomassie Brilliant Blue solution (1% Coomassie Brilliant Blue G-250; 30% methanol and 10% acetic acid) for 3 h at room temperature and destined using 7% acetic acid and 40% methanol until clear bands representing gelatinolysis were observed against a dark background. Total MMP-9 activity was determined using the ATTO Densitograph Software Library Lane Analyser (ATTO Corporation, Tokyo, Japan). Purified MMP-9 (Sigma-Aldrich; Merck KGaA) was used as the internal standard.
Western blot analysis
Western blotting was performed to detect the protein expression levels. The hippocampal tissues were homogenized in lysis buffer (10 mM Tris-HCl pH 7.4, 2 mM EDTA, 150 mM NaCl and 0.5% Nonidet P-40) containing protease inhibitors (1 mg/ml leupeptin, 1 mg/ml aprotinin and 1 mg/ml pepstatin A) and incubated for 30 min on ice. Total protein contents of the tissues were determined using the bicinchoninic acid assay kit (Bio-Rad Laboratories, Inc.). Equal amounts of protein (60 µg/lane) from each group were separated by SDS-PAGE (10% gel), and the bands were transferred onto polyvinylidene fluoride membranes (Invitrogen; Thermo Fisher Scientific, Inc., Waltham, MA, USA). The membranes were blocked in 3% bovine serum albumin in TBS containing Tween-20 (0.1%; TBST) at room temperature for 40 min and incubated with specific primary antibodies, as described above, at 4°C overnight. The membranes were incubated for 2 h at room temperature with goat anti-mouse horseradish peroxidase-labelled secondary antibody (IgG; cat. no. ab97240; 1:2,000; Abcam, Cambridge, MA, USA) prior to washing with TBST. The enhanced chemiluminescence method (GE Healthcare, Chicago, IL, USA) was used to detect and analyze the immunoreactive bands.
Statistical analysis
Data are presented as the mean ± standard deviation, representative of six replicates. SPSS software (version 22.0; IBM Corp., Armonk, NY, USA) was used for the statistical analysis. One-way analysis of variance followed by Duncan's Multiple Range Test were used for multiple group comparisons. P<0.05 was considered to indicate a statistically significant difference.
Results
Myricetin reduces PTZ-induced seizure scores
Repeated administration of sub-convulsive dose of PTZ (35 mg/kg) on alternating days (13 injections) resulted in an increase in the seizure score compared with the control group, eventually leading to widespread clonic-tonic seizures (Racine scale 5). Administration of myricetin (100 or 200 mg) prior to each PTZ injection decreased seizure severity, as evidenced by a decline in the seizure score compared with the PTZ control group (Fig. 1). Furthermore, the mortality rate following the PTZ injections decreased significantly (P<0.05) in response to pre-treatment with myricetin and the group treated with 200 mg myricetin exhibited 5% mortality rate (Fig. 2).
Figure 1.

Myricetin reduces seizure scores following PTZ induction. Data are presented as the mean ± standard deviation, n=6. PTZ, pentylenetetrazole.
Figure 2.

Myricetin reduces mortality in a dose-dependent manner following PTZ induction. Data are presented as the mean ± standard deviation, n=6. aP<0.05 vs. control; bP<0.05 vs. PTZ. PTZ, pentylenetetrazole.
Myricetin reduces PTZ-induced neuronal loss
Mice were sacrificed 24 h after the last PTZ injection, and Nissl staining was used to determine seizure-induced neuronal loss in the CA1 and CA3 regions of the hippocampus. The viable cell count decreased significantly in the PTZ group compared with the normal control animals (P<0.05). Treatment with myricetin at both tested doses significantly attenuated seizure-induced neuronal loss (P<0.05) and the 200 mg dose exerted a higher protective effect compared with the 100 mg dose of myricetin (Fig. 3). Furthermore, negligible neuronal loss was observed among mice treated with valproate.
Figure 3.

Myricetin increases neuronal cell viability following PTZ induction in CA1 and CA3 regions of the hippocampus. Data are presented as the mean ± standard deviation, n=6. aP<0.05 vs. control; bP<0.05 vs. PTZ. PTZ, pentylenetetrazole.
Myricetin efficiently reduces neural degeneration and cell death following treatment with PTZ
TUNEL-positive cell counts in the CA1 and CA3 hippocampal regions were determined following the PTZ injections. Pre-treatment with myricetin reduced the TUNEL-positive cell counts (P<0.05) compared with the PTZ group, demonstrating inhibition of neuronal apoptosis (Fig. 4). Apoptotic protein expression was measured to determine the molecular mechanism underlying the antiapoptotic effects of myricetin. Increased Bad and Bax expression levels were observed in the PTZ control mice, compared with the untreated control groups (Fig. 5). PTZ group mice pre-treated with myricetin (100 or 200 mg) exhibited decreased Bad, Bax and cleaved caspase-3 expression levels. The expression levels of anti-apoptotic proteins, Bcl-2 and Bcl-xL, increased in mice with epilepsy following pre-treatment with myricetin, indicating inhibited apoptosis. These observations suggest that myricetin regulates the expression of proteins associated with the apoptotic pathway and reduces neuronal cell apoptosis to induce neuroprotective effects.
Figure 4.

Myricetin reduces PTZ-induced neuronal apoptosis. Data are presented as the mean ± standard deviation, n=6. aP<0.05 vs. control; bP<0.05 vs. PTZ. PTZ, pentylenetetrazole.
Figure 5.

Western blot analysis of the expression of apoptosis-associated proteins. PTZ, pentylenetetrazole; Bcl-2, B-cell lymphoma-2; Bcl-xL, Bcl extra large; Bad, Bcl2-associated agonist of cell death; Bax, Bcl-2-like protein 4; L1, control group; L2, PTZ group; L3, PTZ+100 mg myricetin group; L4, PTZ+200 mg myricetin group.
Myricetin inhibits the BDNF-TrkB signaling pathway following treatment with PTZ
The BDNF-TrkB signaling is pathway associated with epileptogenesis (19,44). BDNF potentiates glutamatergic neurotransmission and inhibits GABAergic transmission (45). In the current study, elevated BDNF and TrkB mRNA and protein expression levels were observed following the PTZ injections compared with the untreated control group, indicating sensitivity of the neurons and induction of epilepsy (Figs. 6 and 7). Pre-treatment with myricetin prior to the PTZ injections downregulated TrkB and BDNF protein expression in a dose-dependent manner and the 200 mg dose inhibited the BDNF-TrkB signaling pathway. This myricetin-mediated inhibition of the BDNF-TrkB signaling may contribute to the decreased seizure scores.
Figure 6.
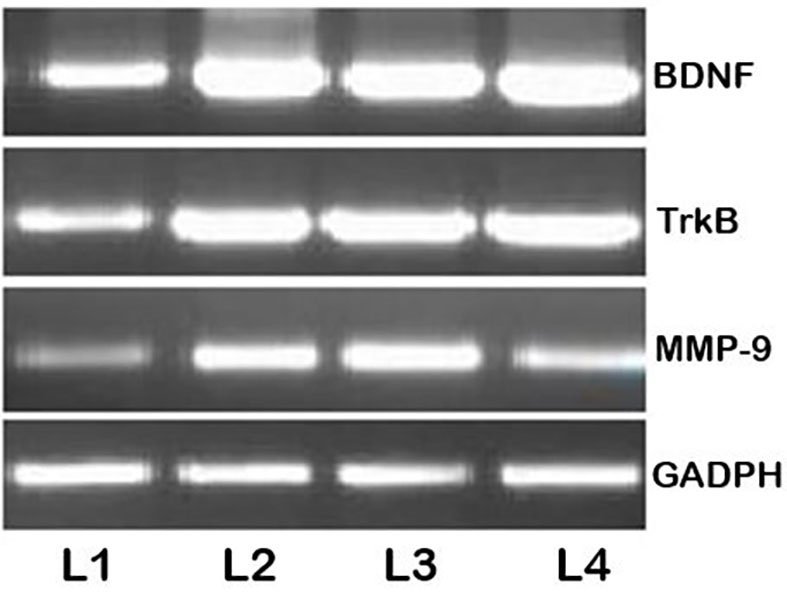
Reverse transcription-semi-quantitative polymerase chain reaction analysis following the PTZ injections. PTZ, pentylenetetrazole; MMP-9, matrix metalloproteinase-9; BDNF, brain-derived neurotrophic factor; TrkB, tropomyosin receptor kinase B; L1, control group; L2, PTZ group; L3, PTZ+100 mg myricetin group; L4, PTZ+200 mg myricetin group.
Figure 7.
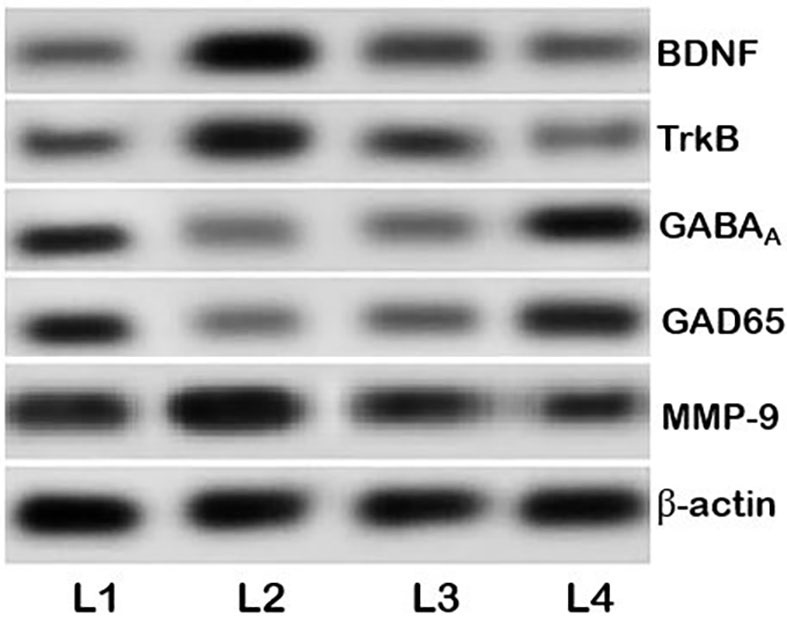
Myricetin regulates the BDNF-TrkB pathway and the expression of GABAA and GAD65. PTZ, pentylenetetrazole; MMP-9, matrix metalloproteinase-9; BDNF, brain-derived neurotrophic factor; TrkB, tropomyosin receptor kinase B; GABAA, γ-aminobutyric acid receptor; GAD65, glutamic acid decarboxylase 65; L1, control group; L2, PTZ group; L3, PTZ+100 mg myricetin group; L4, PTZ+200 mg myricetin group.
Myricetin induces the expression of GAD65 and GABAAR
GAD65 and GABAA expression levels decreased in the PTZ group compared with the untreated control group (Fig. 7). Pre-treatment with myricetin upregulated GAD65 and GABAAR protein expression levels, suggesting that myricetin may exhibit an anti-epileptic potential.
Myricetin regulates glutamate and GABA levels
A glutamate/GABA imbalance in the brain is associated with epileptogenesis (27). Myricetin (100 and 200 mg) significantly increased GABA levels while significantly decreasing glutamate levels compared with the PTZ group (both P<0.05), and normalized the glutamate/GABA ratio (Fig. 8).
Figure 8.

Myricetin regulates the expression levels of Glu and GABA. Data are presented as the mean ± standard deviation, n=6. aP<0.05 vs. control; bP<0.05 vs. PTZ. Glu, glutamate; GABA, γ-aminobutyric acid. PTZ, pentylenetetrazole.
Myricetin modulates the expression of MMP-9
MMP-2 and −9 serve roles in epilepsy (31,46). The effect of myricetin on the expression and activity of MMP-9 was assessed in the current study. The current study observed an increase in MMP-9 mRNA (Fig. 6) and protein (Fig. 7) levels at 24 h after the final PTZ injection, compared with the untreated control group. mRNA levels were unchanged following treatment with myricetin at both doses. The results of the gel zymography analysis revealed and increase in MMP-9 activity following PTZ kindling. MMP 9 activity was downregulated in a dose-dependent manner following treatment with myricetin (Fig. 9). These observations suggest that myricetin may regulate MMP-9 activity.
Figure 9.

Myricetin downregulates MMP-9. Representative gel zymography of MMP-9. PTZ, pentylenetetrazole; MMP-9, matrix metalloproteinase-9; L1, control group; L2, PTZ group; L3, PTZ+100 mg myricetin group; L4, PTZ+200 mg myricetin group.
Discussion
Epilepsy is a prevalent neurological disorder and kindling is a widely accepted experimental model of epileptogenesis (47,48). Kindling induces severe and recurrent seizures by altering neural circuits (49). PTZ is a widely used convulsant in epilepsy research, and subcutaneous injection of PTZ at a lower dose induces seizures (50–52).
In the current study, recurrent administration of a sub-effective dose of PTZ elicited severe seizures and high mortality. Myricetin efficiently decreased seizure severity and reduced mortality in PTZ-kindled mice. PTZ kindling causes hippocampal neuronal cell loss (53). The hippocampus is highly susceptible to seizures and is involved in epileptiform discharges (54).
Treatment with PTZ induced apoptotic neuronal cell death in the CA1 and CA3 regions of the hippocampus. Myricetin exerted protective effects by significantly reducing the apoptotic cell count, as demonstrated by the TUNEL assay and Nissl staining.
Several studies have suggested that seizures induce transient cerebral ischemia leading to neuronal death in the brain (55). To investigate the possible mechanism underlying the neuroprotective effects of myricetin, the expression levels of cleaved caspase-3 and other apoptotic pathway proteins were evaluated by western blot analysis. Increased protein expression levels of cleaved caspase-3, Bad and Bax in the brain of kindled mice indicated apoptosis. Treatment with myricetin significantly upregulated Bcl-xL and Bcl-2 expression, but downregulated the expression of caspase-3, Bad and Bax in a dose-dependent manner, suggesting suppressed apoptosis. This myricetin-mediated modulation of protein expression may facilitate reduced neuronal cell loss.
The BDNF-TrkB signaling is a common pathway involved in a number of epilepsy models (19). BDNF promotes neuronal growth and survival, as well as the formation of synapses (18), and enhances hippocampal learning and memory processes (56). Epileptogenic brain insults increase BDNF synthesis and activate the TrkB receptor (19). In the current study, PTZ induction enhanced the expression of BDNF and TrkB, suggesting activation of the BDNF-TrkB pathway. Pre-treatment with myricetin inhibited BDNF and TrkB, and, therefore, downregulated the BDNF signaling. Both in vivo and in vitro studies demonstrated that the BDNF-TrkB signaling was activated in experimentally induced models of acute seizure, and promoted epileptiform activities in hippocampal neurons (20). Therefore, myricetin-mediated downregulation of the BDNF-TrkB pathway may promote seizure suppression.
BDNF potentiates glutamatergic neurotransmission, while inhibiting GABAergic neurotransmission (19,57). The balance between inhibitory and excitatory neurotransmitters is associated with epileptogenesis (58). A significant increase in glutamate levels, together with a decrease in GABA expression, was observed in the present study following PTZ-kindling, and resulted in a disturbed glutamate/GABA balance. Several previous studies demonstrated that glutamate/GABA imbalance is one of the factors contributing to epileptogenesis (58,59). Expression of GAD is associated with the severity of epilepsy (60,61). Upregulated expression of GAD65, an isoform of GAD, reduces sensitivity to epilepsy and improves the neuropathological alterations in mouse models of epilepsy (62). Increased levels of GABA, together with reduced glutamate levels in PTZ-kindled and myricetin-treated mice observed in the current study indicate restoration of the glutamate/GABA balance. Protein expression levels of GAD65, the key rate-limiting enzyme during GABA synthesis, were further assessed in the current study to determine whether GABA levels were regulated by myricetin (27,42). Previous studies have suggested that GAD65 and GABAA, the main GABA receptor, are potential targets for the development of novel antiepileptic drugs (27,58). Several studies have reported that the progression of kindling-induced seizures is associated with reduced levels of GABAA binding sites in the hippocampus (63) and increased glutamate release. The GABAA receptor-chloride channel is responsible for rapid hyperpolarization of the paroxysmal depolarizing shift implicated in kindling, leading to increased seizure severity (64).
In the present study, upregulated expression of GAD65 and GABAA was observed in mice treated with myricetin. Similar results were previously reported by Hao et al (5). Treatment with garcinol regulated the glutamate/GABA balance and restored GAD65 and GABAA expression in animals with PTZ-induced seizures (4). Restoration of the glutamate/GABA balance may occur due to increased expression of GAD65 and GABAA, indicating the anti-epileptic effects of myricetin.
A number of previous studies using animal models of epilepsy indicated that MMPs serve a role in epileptogenesis (30,32). MMPs are associated with apoptosis, necrotic cell death (32) and synaptic plasticity (30). Furthermore, MMP-9 serves a role in the pathogenesis of epilepsy and is associated with the formation of epileptic foci and tissue remodeling within these foci following seizures (65). Konopacki et al (66) demonstrated enhanced MMP-9 mRNA levels in dentate gyrus of KA-induced rats. Wilczynski et al (30) reported that the synaptic pool of MMP-9 is associated with the mechanisms underlying the development of seizures. Enhanced expression and enzymatic activity of MMP-9 in hippocampal regions have been reported in animal models with PTZ-induced epileptic seizures (67). Furthermore, MMP-9 converts pro-BDNF to mature BDNF, resulting in the activation of TrkB (68,69). PTZ kindling involves synaptic remodeling induced by prolonged MMP-9 activation and elevated BDNF levels (70). Similar to previous observations in experimental models of epilepsy, elevated MMP-9 expression was observed in the present study at both the mRNA and protein level following PTZ kindling (41,70). Gel zymography results revealed increased MMP-9 activity in the current study. Nevertheless, the treatment with myricetin reduced the expression and activity of MMP-9 in a dose-dependent manner. Mizoguchi et al (43) demonstrated that PTZ-induction increased expression of MMP-9 and that selective inhibition of MMP-9 confers neuroprotection in patients with epilepsy. Previously, a significant increase in MMP-9 mRNA was observed in rats 2 h following administration of 50 mg/kg PTZ (67). Therefore, downregulation of MMP-9 may indicate a neuroprotective effect of myricetin against PTZ kindling.
In conclusion, myricetin exhibited protective effects against PTZ-induced seizures by reducing seizure severity and regulating the GAD65/GABAA and BDNF-TrkB signaling. These observations suggest that myricetin may be used as an anti-epileptic drug in the future. However, clinical studies are required to further investigate its protective effects.
Acknowledgements
Not applicable.
Funding
No funding was received.
Availability of data and materials
The datasets used and/or analyzed during the current study are available from the corresponding author on reasonable request.
Authors' contributions
ZQS and LS contributed to the conception and design and were involved in drafting and revising the manuscript. FHM and LXT contributed to the acquisition of the data and the analysis and interpretation. All authors are accountable for all aspects of the work and read and approved the final version of the manuscript.
Ethics approval and consent to participate
The current study was approved by the Animal Studies Ethics Committee of Hedong People's Hospital.
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing interests.
References
- 1.Duncan JS, Sander JW, Sisodiya SM, Walker MC. Adult epilepsy. Lancet. 2006;367:1087–1100. doi: 10.1016/S0140-6736(06)68477-8. [DOI] [PubMed] [Google Scholar]
- 2.Yang T, Kong B, Gu JW, Kuang YQ, Cheng L, Yang WT, Cheng JM, Ma Y, Yang XK. Anticonvulsant and sedative effects of paederosidic acid isolated from Paederia scandens (Lour.) Merrill. in mice and rats. Pharmacol Biochem Behav. 2013;111:97–101. doi: 10.1016/j.pbb.2013.08.015. [DOI] [PubMed] [Google Scholar]
- 3.Prilipko L, deBoer MH, Dua T, Bertolote J. Epilepsy care-The WHO/ILAE/IBE global campaign against epilepsy. US Neurological Disease. 2006:39–40. [Google Scholar]
- 4.Motamedi G, Meador K. Epilepsy and cognition. Epilepsy Behav. 2003;4:25–38. doi: 10.1016/j.yebeh.2003.07.004. [DOI] [PubMed] [Google Scholar]
- 5.Hao F, Jia LH, Li XW, Zhang YR, Liu XW. Garcinol upregulates GABAA and GAD65 expression, modulates BDNF-TrkB pathway to reduce seizures in pentylenetetrazole (PTZ)-induced epilepsy. Med Sci Monit. 2016;22:4415–4425. doi: 10.12659/MSM.897579. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 6.Rahmati B, Khalili M, Roghani M, Ahghari P. Anti-epileptogenic and antioxidant effect of Lavandula officinalis aerial part extract against pentylenetetrazol-induced kindling in male mice. J Ethnopharmacol. 2013;148:152–157. doi: 10.1016/j.jep.2013.04.004. [DOI] [PubMed] [Google Scholar]
- 7.Elger CE, Helmstaedter C, Kurthen M. Chronic epilepsy and cognition. Lancet Neurol. 2004;3:663–672. doi: 10.1016/S1474-4422(04)00906-8. [DOI] [PubMed] [Google Scholar]
- 8.Schmitz B. Effects of antiepileptic drugs on mood and behavior. Epilepsia. 2006;47:28–33. doi: 10.1111/j.1528-1167.2006.00684.x. [DOI] [PubMed] [Google Scholar]
- 9.Xie T, Wang WP, Jia LJ, Mao ZF, Qu ZZ, Luan SQ, Kan MC. Effects of epigallocatechin-3-gallate on pentylenetetrazole-induced kindling, cognitive impairment and oxidative stress in rats. Neurosci Lett. 2012;516:237–241. doi: 10.1016/j.neulet.2012.04.001. [DOI] [PubMed] [Google Scholar]
- 10.Ross JA, Kasum CM. Dietary flavonoids: Bioavailability, metabolic effects, and safety. Ann Rev Nutr. 2002;22:19–34. doi: 10.1146/annurev.nutr.22.111401.144957. [DOI] [PubMed] [Google Scholar]
- 11.Shimosaki S, Tsurunaga Y, Itamura H, Nakamura M. Anti-allergic effect of the flavonoid myricitrin from Myrica rubra leaf extracts in vitro and in vivo. Nat Prod Res. 2011;25:374–380. doi: 10.1080/14786411003774320. [DOI] [PubMed] [Google Scholar]
- 12.Kim HH, Kim DH, Kim MH, Oh MH, Kim SR, Park KJ, Lee MW. Flavonoid constituents in the leaves of Myrica rubra sieb. et zucc. with anti-inflammatory activity. Arch Pharm Res. 2013;36:1533–1540. doi: 10.1007/s12272-013-0147-x. [DOI] [PubMed] [Google Scholar]
- 13.Guitard R, Paul JF, Nardello-Rataj V, Aubry JM. Myricetin, rosmarinic and carnosic acids as superior natural antioxidant alternatives to α-tocopherol for the preservation of omega-3 oils. Food Chem. 2016;213:284–295. doi: 10.1016/j.foodchem.2016.06.038. [DOI] [PubMed] [Google Scholar]
- 14.Pan H, Hu Q, Wang J, Liu Z, Wu D, Lu W, Huang J. Myricetin is a novel inhibitor of human inosine 5′-monophosphate dehydrogenase with anti-leukemia activity. Biochem Biophys Res Commun. 2016;477:915–922. doi: 10.1016/j.bbrc.2016.06.158. [DOI] [PubMed] [Google Scholar]
- 15.Wang G, Wang JJ, Tang XJ, Du L, Li F. In vitro and in vivo evaluation of functionalized chitosan-Pluronic micelles loaded with myricetin on glioblastoma cancer. Nanomedicine. 2016;12:1263–1278. doi: 10.1016/j.nano.2016.02.004. [DOI] [PubMed] [Google Scholar]
- 16.Hassan SM, Khalaf MM, Sadek SA, Abo-Youssef AM. Protective effects of apigenin and myricetin against cisplatin-induced nephrotoxicity in mice. Pharm Biol. 2017;55:766–774. doi: 10.1080/13880209.2016.1275704. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 17.Cho BO, Yin HH, Park SH, Byun EB, Ha HY, Jang SI. Anti-inflammatory activity of myricetin from Diospyros lotus through suppression of NF-κB and STAT1 activation and Nrf2-mediated HO-1 induction in lipopolysaccharide-stimulated RAW264.7 macrophages. Biosci Biotechnol Biochem. 2016;80:1520–1530. doi: 10.1080/09168451.2016.1171697. [DOI] [PubMed] [Google Scholar]
- 18.Huang EJ, Reichardt LF. Neurotrophins: Roles in neuronal development and function. Annu Rev Neurosci. 2001;24:677–736. doi: 10.1146/annurev.neuro.24.1.677. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 19.Binder DK, Croll SD, Gall CM, Scharfman HE. BDNF and epilepsy: Too much of a good thing? Trends Neurosci. 2001;24:47–53. doi: 10.1016/S0166-2236(00)01682-9. [DOI] [PubMed] [Google Scholar]
- 20.Papaleo F, Silverman JL, Aney J, Tian Q, Barkan CL, Chadman KK, Crawley JN. Working memory deficits, increased anxiety-like traits, and seizure susceptibility in BDNF overexpressing mice. Learn Mem. 2011;18:534–544. doi: 10.1101/lm.2213711. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 21.Elmér E, Kokaia Z, Kokaia M, Carnahan J, Nawa H, Lindvall O. Dynamic changes of brain-derived neurotrophic factor protein levels in the rat forebrain after single and recurring kindling-induced seizures. Neurosci. 1998;83:351–362. doi: 10.1016/S0306-4522(97)00387-4. [DOI] [PubMed] [Google Scholar]
- 22.Scharfman HE, Goodman JH, Sollas AL, Croll SD. Spontaneous limbic seizures after intrahippocampal infusion of brain-derived neurotrophic factor. Exp Neurol. 2002;174:201–214. doi: 10.1006/exnr.2002.7869. [DOI] [PubMed] [Google Scholar]
- 23.Kafitz KW, Rose CR, Thoenen H, Konnerth A. Neurotrophin evoked rapid excitation through TrkB receptors. Nature. 1999;401:918–921. doi: 10.1038/44847. [DOI] [PubMed] [Google Scholar]
- 24.Suzdak PD, Jansen JA. A review of the preclinical pharmacology of tiagabine: A potent and selective anticonvulsant GABA uptake inhibitor. Epilepsia. 1995;36:612–626. doi: 10.1111/j.1528-1157.1995.tb02576.x. [DOI] [PubMed] [Google Scholar]
- 25.Vianello M, Tavolato B, Giometto B. Glutamic acid decarboxylase autoantibodies and neurological disorders. Neurol Sci. 2002;23:145–151. doi: 10.1007/s100720200055. [DOI] [PubMed] [Google Scholar]
- 26.Ha JH, Lee DU, Lee JT, Kim JS, Yong CS, Kim JA, Ha JS, Huh K. 4-Hydroxybenzaldehyde from Gastrodia elata B1 is active in the antioxidation and GABAergic neuromodulation of the rat brain. J Ethnopharmacol. 2000;73:329–333. doi: 10.1016/S0378-8741(00)00313-5. [DOI] [PubMed] [Google Scholar]
- 27.Xiang J, Jiang Y. Antiepileptic potential of matrine via regulation the levels of gamma-aminobutyric acid and glutamic acid in the brain. Int J Mol Sci. 2013;14:23751–23761. doi: 10.3390/ijms141223751. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 28.Yong VW. Metalloproteinases: Mediators of pathology and regeneration in CNS. Nat Neurosci. 2005;6:931–944. doi: 10.1038/nrn1807. [DOI] [PubMed] [Google Scholar]
- 29.Rivera S, Khrestchatisky M, Kaczmarek L, Rosenberg GA, Jaworski DM. Metzincin proteases and their inhibitors, foes or friends in nervous system physiology? J Neurosci. 2010;30:15337–15357. doi: 10.1523/JNEUROSCI.3467-10.2010. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 30.Wilczynski GM, Konopacki FA, Wilczek E, Lasiecka Z, Gorlewicz A, Michaluk P, Wawrzyniak M, Malinowska M, Okulski P, Kolodziej LR, et al. Important role of matrix metalloproteinase-9 in epileptogenesis. J Cell Biol. 2008;180:1021–1035. doi: 10.1083/jcb.200708213. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 31.Zhang JW, Deb S, Gottschall PE. Regional and age-related expression of gelatinases in the brains of young and old rats after treatment with kainic acid. Neurosci Lett. 2000;295:9–12. doi: 10.1016/S0304-3940(00)01582-2. [DOI] [PubMed] [Google Scholar]
- 32.Kim GW, Kim HJ, Cho KJ, Kim HW, Cho YJ, Lee BI. The role of MMP-9 in integrin-mediated hippocampal cell death after pilocarpine-induced status epilepticus. Neurobiol Dis. 2009;36:169–180. doi: 10.1016/j.nbd.2009.07.008. [DOI] [PubMed] [Google Scholar]
- 33.Szklarczyk A, Lapinska J, Rylski M, McKay RD, Kaczmarek L. Matrix metalloproteinase-9 undergoes expression and activation during dendritic remodeling in adult hippocampus. J Neurosci. 2002;22:920–930. doi: 10.1523/JNEUROSCI.22-03-00920.2002. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 34.Garber JC. Washington, DC, USA: National Academy of Sciences; 2011. (Chair): Committee for the Update of the Guide for the Care and Use of Laboratory Animals. Guide for the Care and Use of Laboratory Animals, eighth ed. [Google Scholar]
- 35.Wang J. The evolving regulatory environment. In: Pankevich DE, Wizemann TM, Mazza A, Altevogt BM, editors. International Animal Research Regulations: Impact on neuroscience research-workshop summary. The National Academies Press; Washington, DC: 2012. pp. 13–16. [PubMed] [Google Scholar]
- 36.The National People's Congress of China, corp-author. Animal Husbandry Law of the People's Republic of China 2005. Adopted at the 19th Meeting of the Standing Committee of the Tenth National People's Congress on December 29, 2005. http://english.agri.gov.cn/governmentaffairs/lr/ap/201305/t20130508_19602.htm. [May 8;2013 ];
- 37.Löscher W. Animal models of epilepsy for the development of antiepileptogenic and disease-modifying drugs. A comparison of the pharmacology of kindling and post-status epilepticus models of temporal lobe epilepsy. Epilepsy Res. 2002;50:105–123. doi: 10.1016/S0920-1211(02)00062-1. [DOI] [PubMed] [Google Scholar]
- 38.Bertram E. The relevance of kindling for human epilepsy. Epilepsia. 2007;48(Suppl 2):S65–S74. doi: 10.1111/j.1528-1167.2007.01068.x. [DOI] [PubMed] [Google Scholar]
- 39.Racine RJ. Modification of seizure activity by electrical stimulation. II. Motor seizure. Electroencephalogr Clin Neurophysiol. 1972;32:281–294. doi: 10.1016/0013-4694(72)90177-0. [DOI] [PubMed] [Google Scholar]
- 40.Hui L, Pei DS, Zhang QG, Guan QH, Zhang GY. The neuroprotection of insulin on ischemic brain injury in rat hippocampus through negative regulation of JNK signaling pathway by PI3 K/Akt activation. Brain Res. 2005;1052:1–9. doi: 10.1016/j.brainres.2005.05.043. [DOI] [PubMed] [Google Scholar]
- 41.Li Y, Liang G, Wang S, Meng Q, Wang Q, Wei H. Effects of fetal exposure to isoflurane on postnatal memory and learning in rats. Neuropharmacology. 2007;53:942–950. doi: 10.1016/j.neuropharm.2007.09.005. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 42.Monge-Acuña AA, Fornaguera-Trías J. A high performance liquid chromatography method with electrochemical detection of gamma-aminobutyric acid, glutamate and glutamine in rat brain homogenates. J Neurosci Methods. 2009;183:176–181. doi: 10.1016/j.jneumeth.2009.06.042. [DOI] [PubMed] [Google Scholar]
- 43.Mizoguchi H, Nakade J, Tachibana M, Ibi D, Someya E, Koike H, Kamei H, Nabeshima T, Itohara S, Takuma K, et al. Matrix metalloproteinase-9 contributes to kindled seizure development in pentylenetetrazole-treated mice by converting pro-BDNF to mature BDNF in the hippocampus. J Neurosci. 2011;31:12963–12971. doi: 10.1523/JNEUROSCI.3118-11.2011. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 44.Lähteinen S, Pitkänen A, Saarelainen T, Nissinen J, Koponen E, Castrén E. Decreased BDNF signaling in transgenic mice reduces epileptogenesis. Eur J Neurosci. 2002;15:721–734. doi: 10.1046/j.1460-9568.2002.01897.x. [DOI] [PubMed] [Google Scholar]
- 45.Zhu WJ, Roper SN. Brain-derived neurotrophic factor enhances fast excitatory synaptic transmission in human epileptic dentate gyrus. Ann Neurol. 2001;50:188–194. doi: 10.1002/ana.1074. [DOI] [PubMed] [Google Scholar]
- 46.Hoehna Y, Uckermann O, Luksch H, Stefovska V, Marzahn J, Theil M, Gorkiewicz T, Gawlak M, Wilczynski GM, Kaczmarek L, Ikonomidou C. Matrix metalloproteinase 9 regulates cell death following pilocarpine-induced seizures in the developing brain. Neurobiol Dis. 2012;48:339–347. doi: 10.1016/j.nbd.2012.06.023. [DOI] [PubMed] [Google Scholar]
- 47.Schmidt D, Rogawski MA. New strategies for the identification of drugs to prevent the development or progression of epilepsy. Epilepsy Res. 2002;50:71–78. doi: 10.1016/S0920-1211(02)00070-0. [DOI] [PubMed] [Google Scholar]
- 48.Fatih S, Faruk B, Acar MD, Cafer M. Influence of carbenoxolone on the anticonvulsant efficacy of phenytoin in pentylenetetrazole kindled rats. Acta Neurobiol Exp (Wars) 2012;72:177–184. doi: 10.55782/ane-2012-1890. [DOI] [PubMed] [Google Scholar]
- 49.He XP, Wen R, McNamara JO. Impairment of kindling development in phospholipase Cγ1 heterozygous mice. Epilepsia. 2014;55:456–463. doi: 10.1111/epi.12536. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 50.Liu X, Liu J, Liu J, Liu XL, Jin LY, Fan W, Ding J, Peng LC, Wang Y, Wang X. BDNF-TrkB signaling pathway is involved in pentylenetetrazole-evoked progression of epileptiform activity in hippocampal neurons in anesthetized rats. Neurosci Bull. 2013;29:565–575. doi: 10.1007/s12264-013-1326-y. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 51.Mortazavi F, Ericson M, Story D, Hulce VD, Dunbar GL. Spatial learning deficits and emotional impairments in pentylenetetrazole-kindled rats. Epilepsy Behav. 2005;7:629–638. doi: 10.1016/j.yebeh.2005.08.019. [DOI] [PubMed] [Google Scholar]
- 52.Pitkänen A, Schwartzkroin PA, Moshe SL. Burlington, VT: Elsevier Academic Press, Elsevier Inc; 2006. Models of seizures and epilepsy; pp. 345–346. [Google Scholar]
- 53.Pavlova T, Stepanichev M, Gulyaeva N. Pentylenetetrazole kindling induces neuronal cyclin B1 expression in rat hippocampus. Neurosci Lett. 2006;392:154–158. doi: 10.1016/j.neulet.2005.09.021. [DOI] [PubMed] [Google Scholar]
- 54.Scharfman HE. Epilepsy as an example of neural plasticity. Neuroscientist. 2002;8:154–173. doi: 10.1177/107385840200800211. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 55.Diemer NH, Jørgensen MB, Johansen FF, Sheardown M, Honoré T. Protection against ischemic hippocampal CA1 damage in the rat with a new non-NMDA antagonist NBQX. Acta Neurol Scand. 1992;86:45–49. doi: 10.1111/j.1600-0404.1992.tb08052.x. [DOI] [PubMed] [Google Scholar]
- 56.Yamada K, Nabeshima T. Brain-derived neurotrophic factor/TrkB signaling in memory processes. J Pharmacol Sci. 2003;91:267–270. doi: 10.1254/jphs.91.267. [DOI] [PubMed] [Google Scholar]
- 57.Tanaka T, Saito H, Matsuki N. Inhibition of GABAA synaptic responses by brain-derived neurotrophic factor (BDNF) in rat hippocampus. J Neurosci. 1997;17:2959–2966. doi: 10.1523/JNEUROSCI.17-09-02959.1997. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 58.Svenningsen AB, Madsen KD, Liljefors T, Stafford GI, Stafford GI, van Staden J, Jäeger AK. Biflavones from Rhus species with affinity for the GABAA/benzodiazepine receptor. J Ethnopharmacol. 2006;103:276–80. doi: 10.1016/j.jep.2005.08.012. [DOI] [PubMed] [Google Scholar]
- 59.Ren LX, Wu YL, Luo YF, Li X, Zuo DY. Effects of amiloride on behavior and extracellular glutamate level of frontal cortex in acute epileptic mice induced by penicillin. Chin Pharmacol Bull. 2006;22:1274–1275. (In Chinese) [Google Scholar]
- 60.Tian FF, Xie GJ, Yang QD, Chen ZC, Lv BQ. Relationship between the epileptic seizure and both the GABA concentration, and GAD activity in temporal tissue. Clin J Neurol. 2000;33:224–226. (In Chinese) [Google Scholar]
- 61.Yang DB, Wang L, Huang M, Yu JM, Wang XM, Luo JM. Effects of pretreatment with repetitive transcranial magnetic stimulation on development of seizures induced by pilocarpine and expression of GAD65 in rat hippocampus. Chin J Clin Neurosci. 2009;17:337–340. [Google Scholar]
- 62.Liu X, Qi ZG, Xie P. Transplantation of GAD65-engineered neural stem cells for treatment of epilepsy: A study in rat model. Laser J. 2011;32:54–56. [Google Scholar]
- 63.Bazyan AS, Zhulin VV, Karpova MN, Klishina NY, Glebov RN. Long-term reduction of benzodiazepine receptor density in the rat cerebellum by acute seizures and kindling and its recovery 6 months later by a pentylenetetrazole challenge. Brain Res. 2001;888:212–220. doi: 10.1016/S0006-8993(00)03045-6. [DOI] [PubMed] [Google Scholar]
- 64.Armijo JA, de las Cuevas I, Adín J. Ion channels and epilepsy. Rev Neurol. 2000;30(Suppl 1):S25–S41. (In Spanish) [PubMed] [Google Scholar]
- 65.Sharma AK, Reams RY, Jordan WH, Miller MA, Thacker HL, Snyder PW. Mesial temporal lobe epilepsy: Pathogenesis, induced rodent models and lesions. Toxicol Pathol. 2007;35:984–999. doi: 10.1080/01926230701748305. [DOI] [PubMed] [Google Scholar]
- 66.Konopacki FA, Rylski M, Wilczek E, Amborska R, Detka D, Kaczmarek L, Wilczynski GM. Synaptic localization of seizure-induced matrix metalloproteinase-9 mRNA. Neuroscience. 2007;150:31–39. doi: 10.1016/j.neuroscience.2007.08.026. [DOI] [PubMed] [Google Scholar]
- 67.Rylski M, Amborska R, Zybura K, Michaluk P, Bielinska B, Konopacki FA, Wilczynski GM, Kaczmarek L. Jun-B is a repressor of MMP-9 transcription in depolarized rat brain neurons. Mol Cell Neurosci. 2009;40:98–110. doi: 10.1016/j.mcn.2008.09.005. [DOI] [PubMed] [Google Scholar]
- 68.Hwang JJ, Park MH, Choi SY, Koh JY. Activation of the Trk signaling pathway by extracellular zinc. Role of metalloproteinases. J Biol Chem. 2005;280:11995–12001. doi: 10.1074/jbc.M403172200. [DOI] [PubMed] [Google Scholar]
- 69.Ethell IM, Ethell DW. Matrix metalloproteinases in brain development and remodeling: Synaptic functions and targets. J Neurosci Res. 2007;85:2813–2823. doi: 10.1002/jnr.21273. [DOI] [PubMed] [Google Scholar]
- 70.Mizoguchi H, Yamada K. Roles of matrix metalloproteinases and their targets in epileptogenesis and seizures. Clin Psychopharmacol Neurosci. 2013;11:45–52. doi: 10.9758/cpn.2013.11.2.45. [DOI] [PMC free article] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.
Data Availability Statement
The datasets used and/or analyzed during the current study are available from the corresponding author on reasonable request.