

Keywords: nerve regeneration, cerebral ischemia/reperfusion injury, estrogen, G protein-coupled estrogen receptor, G1, G15, endoplasmic reticulum stress, glucose-regulated protein 78, caspase-12, C/EBP homologous protein, neuronal apoptosis, neural regeneration
Abstract
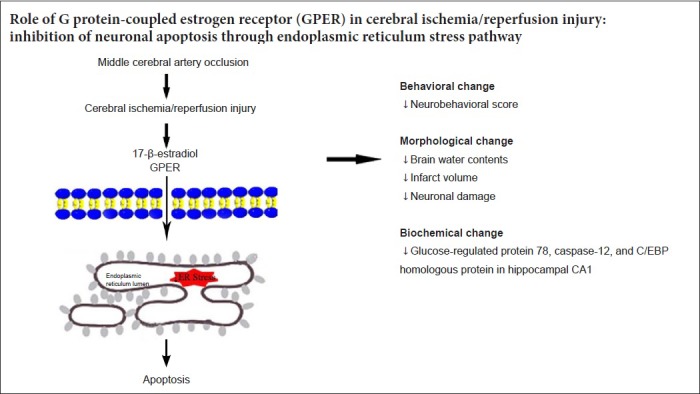
Studies have confirmed a strong association between activation of the endoplasmic reticulum stress pathway and cerebral ischemia/reperfusion (I/R) injury. In this study, three key proteins in the endoplasmic reticulum stress pathway (glucose-regulated protein 78, caspase-12, and C/EBP homologous protein) were selected to examine the potential mechanism of endoplasmic reticulum stress in the neuroprotective effect of G protein-coupled estrogen receptor. Female Sprague-Dawley rats received ovariectomy (OVX), and then cerebral I/R rat models (OVX + I/R) were established by middle cerebral artery occlusion. Immediately after I/R, rat models were injected with 100 μg/kg E2 (OVX + I/R + E2), or 100 μg/kg G protein-coupled estrogen receptor agonist G1 (OVX + I/R + G1) in the lateral ventricle. Longa scoring was used to detect neurobehavioral changes in each group. Infarct volumes were measured by 2,3,5-triphenyltetrazolium chloride staining. Morphological changes in neurons were observed by Nissl staining. Terminal dexynucleotidyl transferase-mediated nick end-labeling staining revealed that compared with the OVX + I/R group, neurological function was remarkably improved, infarct volume was reduced, number of normal Nissl bodies was dramatically increased, and number of apoptotic neurons in the hippocampus was decreased after E2 and G1 intervention. To detect the expression and distribution of endoplasmic reticulum stress-related proteins in the endoplasmic reticulum, caspase-12 distribution and expression were detected by immunofluorescence, and mRNA and protein levels of glucose-regulated protein 78, caspase-12, and C/EBP homologous protein were determined by polymerase chain reaction and western blot assay. The results showed that compared with the OVX + I/R group, E2 and G1 treatment obviously decreased mRNA and protein expression levels of glucose-regulated protein 78, C/EBP homologous protein, and caspase-12. However, the G protein-coupled estrogen receptor antagonist G15 (OVX + I/R + E2 + G15) could eliminate the effect of E2 on cerebral I/R injury. These results confirm that E2 and G protein-coupled estrogen receptor can inhibit the expression of endoplasmic reticulum stress-related proteins and neuronal apoptosis in the hippocampus, thereby improving dysfunction caused by cerebral I/R injury. Every experimental protocol was approved by the Institutional Ethics Review Board at the First Affiliated Hospital of Shihezi University School of Medicine, China (approval No. SHZ A2017-171) on February 27, 2017.
Chinese Library Classification No. R453; R364
Introduction
Stroke, a cerebrovascular condition, is increasing in prevalence (Shi et al., 2016; Zheng et al., 2018). Given that stroke is a condition in which blood flow to brain areas is disrupted, it is often associated with high rates of severe disabilities and multiple functional impairments. Moreover, reperfusion itself produces reactive oxygen species or overproduction of free radicals that can cause reperfusion injury (Chan, 1996). Despite advancements in the treatment of reperfusion injury in over the past few decades, clinical effects remain unsatisfactory. Notably, women have lower risk for ischemic stroke than men. However, this innate protection diminishes after menopause, which is believed to be associated with a loss of endogenous estrogen. Similarly, female animals demonstrate less tissue damage and improved functional outcome after experimental cerebral ischemia compared with their male or ovariectomized (OVX) female counterparts (Herson et al., 2009).
Apoptosis is a major pathophysiological change after ischemia/reperfusion (I/R) brain injury (Mattson et al., 2001). The endoplasmic reticulum stress pathway is a newly discovered apoptosis pathway that has been shown to induce apoptosis in Parkinson’s disease (Silva et al., 2005), atherosclerosis (Scull and Tabas, 2011), Alzheimer’s disease (Gwag et al., 2010), and other metabolic diseases (Sano and Reed, 2013).
There are three main types of estrogens in the human body: estradiol (17 beta-estradiol, E2), estriol, and estrone. However, E2 is the most biologically active natural estrogen. Animal experiments have confirmed that an appropriate dose of estrogen can reduce the area of cerebral infarction or hippocampal cell damage resulting from cerebral ischemia (Buttesmith et al., 2009; Lebesgue et al., 2009). However, owing to the special nature of estrogen, it cannot be used in male patients. Thus, clinical application of estrogen has many disadvantages, such as increased risk of cervical and breast cancers in female patients after perimenopause (Imoto et al., 1998).
Estrogen can regulate cell proliferation, apoptosis, and inflammatory response through rapid signal responses mediated by G protein-coupled estrogen receptor (GPER), a recently discovered estrogen-associated receptor (Prossnitz and Barton, 2014). The GPER agonist G1 and antagonist G15 play a significant role in the study of GPER (Morris et al., 1982; Bell et al., 2006). G1 can specifically excite GPER to play a protective role in many diseases and may replace estrogen in the future (Tropea et al., 2015). Several studies have shown that estrogen may protect the brain by inhibiting endoplasmic reticulum stress and apoptosis (Shughrue and Merchenthaler, 2003; Jia et al., 2009). However, the potential relationship between GPER and endoplasmic reticulum stress-induced apoptosis after I/R injury has not been elucidated. Thus, the present study investigated whether GPER protected the brain against I/R injury, and examined the role of the apoptotic mechanism mediated by I/R injury.
Materials and Methods
Animals
A total of 180 female Sprague-Dawley rats aged 8–12 weeks and weighing 230–300 g were purchased from the Experimental Animal Center of Xinjiang Medical University, China [certificate No. SCXK (Xin) 2003-0001]. Two or three rats were housed in each cage and under a reversed 12-hour light/dark cycle. Food and water were provided ad libitum, and rats were weighed twice a week.
Every experimental protocol was approved by the Institutional Ethics Review Board at the First Affiliated Hospital of Shihezi University School of Medicine, China (approval No. SHZ A2017-171) on February 27, 2017.
Establishment of middle cerebral artery occlusion rat models
The middle cerebral artery occlusion model was induced via an intraluminal suture technique under sterile conditions, as previously described (Offner et al., 2006; Hurn et al., 2007). Briefly, each rat was intraperitoneally anesthetized with 40 mg/kg pentobarbital and allowed to breathe spontaneously. A rectal probe was used to monitor body temperature, and a heat lamp was used to maintain normal body temperature. Surgery was performed on a stereotaxic apparatus. After anesthetization, rats were placed in a supine position, and a midline skin incision was made to expose the right common carotid artery, internal carotid artery, and external carotid artery. A 3-0 monofilament nylon thread with a rounded tip was inserted into the external carotid artery and gently advanced to a distance of 18.5 ± 0.5 mm, which blocked blood flow to the right middle cerebral artery. After 90 minutes, the 3-0 filament was pulled out, resulting in reperfusion. After the animal awoke, reperfusion was considered successful when the rat incurred a neurological behavior score of more than 1. Rats whose nylon filament did not completely block or penetrate blood vessels were excluded. Neurobehavioral scores were evaluated as previously described, with a neurobehavioral score ≥ 1 considered as successful modeling (Liu et al., 2012; Zhang et al., 2018).
Experimental groups and paraffin sectioning
One hundred eighty rats were randomly divided into 6 groups (n = 30). All drug treatment groups were administered once immediately upon reperfusion. (1) OVX group: Female rats with ovaries removed one week before transient focal cerebral ischemia. (2) OVX + I/R group: OVX rats with I/R model established through middle cerebral artery occlusion (Vavers et al., 2016). (3) OVX + I/R + vehicle group: OVX rats with I/R who received a lateral ventricle injection of dimethyl sulfoxide (DMSO; Sigma-Aldrich, St. Louis, MO, USA; high polarity, high boiling point, good thermal stability, miscibility with water, and good solvent for most organic substances). (4) OVX + I/R + E2 group: OVX rats with I/R who received a lateral ventricle injection of E2 (100 μg/kg, Sigma-Aldrich) diluted in DMSO. (5) OVX + I/R + G1 group: OVX rats with I/R who received a lateral ventricle injection of the GPER-specific agonist G1 (100 μg/kg, Sigma-Aldrich). (6) OVX + I/R + E2 + G15 group: OVX rats with I/R who received a lateral ventricle injection of E2 with the GPER-specific inhibitor G15 (100 μg/kg, Sigma-Aldrich).
Six rats were randomly selected from each group for euthanization and staining of paraffin-embedded sections for terminal dexynucleotidyl transferase (dUTP)-mediated nick end labeling (TUNEL), Nissl, and immunofluorescence.
Neurological scoring
Twenty-four hours after I/R, neurological deficits were evaluated blindly through using the Longa scoring system (Longa et al., 1989), which comprises five tests covering spontaneous activity, walking (but failing to fully extend the left forepaw), and mild focal neurological deficits. A score of 0 was assigned to the best test performance, whereas 4 points were assigned to the worst performance. Mean neurological scores were evaluated by two blinded observers for grading.
Brain water content
Six rats were randomly selected from each group at 24 hours after I/R injury. After neurobehavioral scoring, these rats were deeply anesthetized and sacrificed, and their brains were weighed immediately to determine the wet weight. Subsequently, samples were dried at 120°C for 24 hours and weighed again to determine the dry weight. Percentages of brain water content were calculated as [(wet weight − dry weight)/wet weight] × 100%.
2,3,5-Triphenyltetrazolium chloride staining
Twenty-four hours after I/R injury, six rats from each group were administered anesthesia, sacrificed, and their complete brain tissue was quickly removed and frozen in a −50°C refrigerator for 5 minutes. Frozen brain tissues were sliced to 2 mm-thick sections, quickly placed in 2% triphenyltetrazolium chloride (TTC) solution (Sigma-Aldrich), incubated at 37°C for 30 minutes, and fixed in 10% formaldehyde. Slices were taken out, and both sides of each brain slice was photographed with a digital camera. The infarction area of the two sides of each brain section was measured using a CM-2000B medical image analysis system. Red areas indicated normal brain tissue, whereas pale areas indicated the infarct area. Infarct volume percentages were calculated as (infarct volume / total volume) × 100%.
TUNEL staining
TUNEL analysis was performed using an in situ cell death assay kit (Roche Molecular Biochemicals, Mannheim, Germany) according to the manufacturer’s instructions in brain tissue at 24 hours after I/R. Stained brain sections were visualized with a confocal microscope (FV-1000; Olympus, Tokyo, Japan) and digital images were captured. For each group, TUNEL-positive cells were manually counted in the CA1 region (50 µm × 50 µm) of three sections (n = 6). Cells were blindly analyzed and TUNEL-positive cells were counted using a 20× objective.
Nissl staining
Twenty-four hours after I/R, randomly selected slices of the hippocampal CA1 region were flushed with 0.01 M phosphate-buffered saline (PBS) and dipped in dimethylbenzene I, II, and III at concentrations of 90%, 80%, and 70%, respectively, for 5 minutes each. Slices were then immersed in 5 g of Nissl solution (Solarbio Science & Technology Co., Ltd., Beijing, China) at 37°C for 20 minutes. After incubation, slices were washed with distilled water, followed by 95% alcohol for color separation. Microscopic examination showed distinct Nissl bodies. Slices were dehydrated with anhydrous alcohol, rendered transparent with xylene, and mounted with neutral balata.
Immunofluorescence
Paraffin sections of the hippocampal CA1 region were randomly selected at 24 hours after I/R, washed with PBS, dewaxed separately in xylene and alcohol, then repaired in citrate repair solution for 8 minutes. Sections were then washed three times with PBS for 5 minutes each, and then incubated in 1% bovine serum albumin for 1 hour at room temperature. Each section was soaked in rabbit anti-caspase-12 polyclonal antibody (1:500; Abcam, Cambridge, UK) diluted in PBS (1:125) at 4°C overnight. After washing with PBS, sections were incubated with a FITC-conjugated anti-rabbit secondary antibody (1:100; Santa Cruz Biotechnology, Dallas, TX, USA) for 1 hour at 37°C. After each slice was rinsed three times with PBS, sections were restained with 4′,6-diamidine-2-phenylindole (Sigma-Aldrich). Immunofluorescence staining of caspase-12 was observed under a confocal laser-scanning microscope (Zeiss, Oberkochen, Germany). Average optical densities were analyzed and calculated by ImageJ software (Media Cybernetics, Rockville, MD, USA). Five visual fields of each sample were randomly selected and analyzed at a high magnification. The fluorescence intensity of each group was analyzed statistically.
Quantitative real-time polymerase chain reaction assay
Twenty-four hours after I/R, the hippocampal CA1 region was taken from six rats of each group for quantitative real-time polymerase chain reaction (qRT-PCR) to determine mRNA expression levels of glucose-regulated protein (GRP78), caspase-12, and C/EBP homologous protein (CHOP). Total RNA was extracted from brain samples using a brain tissue RNA kit (Sangon Biotech, Shanghai, China) according to the manufacturer’s instructions. Isolated RNA was used for cDNA synthesis using a reverse transcription system. PCR analysis used the real-time SYBR Green PCR technology and primers shown in Table 1. A thresholding method was performed to analyze relative gene expression. Each sample was measured thrice. The 2−ΔΔCt method was used to express relative mRNA levels (Yuan et al., 2018).
Table 1.
Primer sequence
| Gene | Primer sequence (5’–3’) | Product size (bp) |
|---|---|---|
| GRP78 | Forward: GAA CCA ACT CAC GTC CAA CC | 118 |
| Reverse: AAC CAC CTT GAA TGG CAA GA | ||
| Caspase-12 | Forward: TGG CCC ATG AAT CAC ATC TA | 143 |
| Reverse: TGT TGC AGA TGA TGA GAG CC | ||
| CHOP | Forward: CTG GAA GCC TGG TAT GAG GA | 127 |
| Reverse: GGG ATG CAG GGT CAA GAG TA | ||
| β-Actin | Forward: CAA CCT TCT TGC AGC TCC TC | 137 |
| Reverse: CGG TGT CCC TTC TGA GTG TT |
Caspase-12: Cysteinyl aspartate specific proteinase 12; CHOP: C/EBP homologous protein; GRP78: glucose-regulated protein 78.
Western blot assay
Twenty-four hours after I/R, six rats were euthanized for western blot assay in each group. Homogenates of hippocampal CA1 region were incubated at 4°C for 30 minutes, and then centrifuged at 10,450 × g for 15 minutes after extracting hippocampal proteins. Protein concentration in the supernatant was measured using a protein assay kit. Proteins were transferred to a polyvinylidene fluoride membrane (EMD Millipore, Billerica, MA, USA) after separation by Tris-glycine denaturing gradient gel electrophoresis. After blocking for 1 hour at 37°C in Tris-buffered saline with Tween (pH 8.0,10 mM Tris-HCl, 150 mM NaCl, and 0.2% Tween 20), membranes were incubated with the following antibodies overnight at 4°C: rabbit monoclonal anti-GRP78 (1:500; Abcam), monoclonal anti-mouse CHOP (1:500; Abcam), and rabbit polyclonal anti-caspase-12 (1:1000; Abcam). The blot was washed with Tris-buffered saline with Tween. After incubating with a horseradish peroxidase-conjugated secondary antibody (anti-rabbit or anti-mouse; 1:20,000; Santa Cruz Biotechnology) for 2 hours at room temperature, staining was visualized using enhanced chemiluminescence (GE Healthcare, Chicago, IL, USA). Target protein bands were visualized using an enhanced chemiluminescence kit in conjunction with X-ray film. Grayscale values were measured and normalized to the corresponding β-actin in the same sample.
Statistical analysis
Experimental data were processed using SPSS 17.0 statistical software (SPSS, Chicago, IL, USA). Statistical differences between two groups were analyzed by Student’s t test. One-way analysis of variance followed by Student-Newman-Keuls post hoc test was used to compare differences between more than two groups. P < 0.05 was considered statistically significant.
Results
Physiological evaluation
Mean arterial pressure (85–125 mmHg), partial pressure of oxygen (PO2, 80–95 mmHg), partial pressure of carbon dioxide (PCO2, 35–45 mmHg), arterial pH (7.35–7.45), and blood glucose levels (95–125 mg/dL) were monitored during the operation. Insignificant differences were observed between groups during the experiment (data not shown).
Neurological scoring
Compared with the OVX group, Longa scores in the OVX + I/R group were significantly increased (P < 0.05; Figure 1A). However, Longa scores were not significantly different between OVX + I/R + vehicle and OVX + I/R groups. Compared with the OVX + I/R group, scores were significantly increased in the OVX + I/R + E2 group, but significantly reduced in the OVX + I/R + G1 group (P < 0.05; Figure 1A). Compared with the OVX + I/R + E2 group, Longa scores were significantly increased in the OVX + I/R + E2 + G15 group (P < 0.05; Figure 1A).
Figure 1.

E2 and G1 reduce neurobehavioral scores and brain water content.
(A) Neurobehavioral score determination (Longa scoring system). (B) Brain water content determination. Data are expressed as the mean ± SEM (n = 6; one-way analysis of variance followed by Student-Newman-Keuls post hoc test). *P < 0.05, vs. OVX group; #P < 0.05, vs. OVX + I/R group; †P < 0.05, vs. OVX + I/R + E2 group. E2: 17-β-estradiol; G1: selective GPER agonist; G15: selective GPER inhibitor; I/R: ischemia/reperfusion; OVX: ovariectomized.
Effects of E2 and G1 on reduced brain water content after cerebral I/R injury in rats
Approximately 24 hours after I/R, brain water contents were significantly increased in OVX + I/R and OVX + I/R + vehicle groups compared with the OVX group (P < 0.05; Figure 1B). Brain water content was also reduced in OVX + I/R + E2 and OVX + I/R + G1 groups compared with the OVX + I/R group (P < 0.05; Figure 1B). However, brain water contents were not significantly different between OVX + I/R and OVX + I/R + vehicle groups (P < 0.05; Figure 1B). Compared with the OVX + I/R + E2 group, brain water content was significantly increased in the OVX + I/R + E2 + G15 group (P < 0.01; Figure 1B).
Effects of E2 and G1 on infarct volume after cerebral I/R injury in rats
TTC staining reflected the degree of cerebral ischemia (Figure 2A). Compared with the OVX group, infarct volumes were significantly increased in the OVX + I/R group (P < 0.05; Figure 2B). However, infarct volumes were significantly reduced in OVX + I/R + E2 and OVX + I/R + G1 groups compared with the OVX + I/R group (P < 0.01; Figure 2B). Compared with the OVX + I/R + E2 group, infarct volumes were significantly increased in the OVX + I/R + E2 + G15 group (P < 0.01; Figure 2B).
Figure 2.

E2 and G1 reduce infarct volume after I/R.
(A) Brain infarct region in TTC staining. Red: Non-ischemic area; white: ischemic area. (B) Percentage of infarct volume after 2,3,5-triphenyltetrazolium chloride staining. Data are expressed as mean ± SEM (n = 6; one-way analysis of variance followed by Student-Newman-Keuls post hoc test). *P < 0.05, vs. OVX group; #P < 0.05, vs. OVX + I/R group; †P < 0.05, vs. OVX + I/R + E2 group. E2: 17-β-estradiol; G1: selective GPER agonist; G15: selective GPER inhibitor; I/R: ischemia/reperfusion; OVX: ovariectomized.
Effects of E2 and G1 on neuronal morphology in the hippocampal CA1 region after cerebral I/R injury in rats
Rats from each group were euthanized 24 hours after cerebral I/R injury. Nissl staining was used to examine morphologic changes in CA1 neurons. In the OVX group, pyramidal neurons in the hippocampal CA1 subfield had clear cell outlines and abundant cytoplasm. Compared with the OVX group, neuronal damage was more severe in the OVX + I/R group, and injury in the hippocampal CA1 subfield was reduced in the OVX + I/R + E2 group. I/R-induced neuronal damage was clearly ameliorated in the OVX + I/R + G1 group compared with the OVX + I/R group. However, when G15 was injected, the neuroprotective effect of E2 was abolished compared with the OVX + I/R + E2 group (Figure 3).
Figure 3.

E2 and G1 reduce morphological alterations in the hippocampal CA1 region after I/R, as detected by Nissl staining.
(A) OVX group; (B) OVX + I/R group; (C) OVX + I/R + vehicle group; (D) OVX + I/R + E2 group; (E) OVX + I/R + G1 group; (F) OVX + I/R + E2 + G15 group. In the OVX group, neurons were neatly arranged, cell morphology was intact, and nuclei were not condensed or dissolved. In the I/R group, the number of neurons was remarkably reduced and their arrangement was disordered, the cytoplasm was swollen, and cell bodies were shrunken. In OVX + I/R + E2 and OVX + I/R + G1 groups, neurons were relatively well-formed and neatly arranged. Neuronal morphology of the I/R + E2 + G15 group was similar to that of the I/R group. Arrows indicate Nissl bodies. Original magnification: 400×. Scale bars: 25 µm. E2: 17-β-estradiol; G1: selective GPER agonist; G15: selective GPER inhibitor; I/R: ischemia/reperfusion; OVX: ovariectomized.
E2 and G1 reduce apoptosis in the hippocampus of rats after cerebral I/R injury
Compared with the OVX group, percentages of apoptotic cells were significantly increased in OVX + I/R and OVX + I/R + vehicle groups (P < 0.05; Figure 4A). However, upon E2 and G1 injection, these groups exhibited significant decreases in numbers of TUNEL-positive cells (P < 0.05; Figure 4A) compared with the OVX+I/R group. A non-significant difference was observed between OVX + I/R and OVX + I/R + vehicle groups (P < 0.05; Figure 4A). Compared with the OVX + I/R + E2 group, the percentage of apoptotic cells was significantly increased in the OVX + I/R + E2 + G15 group (P < 0.05; Figure 4A).
Figure 4.
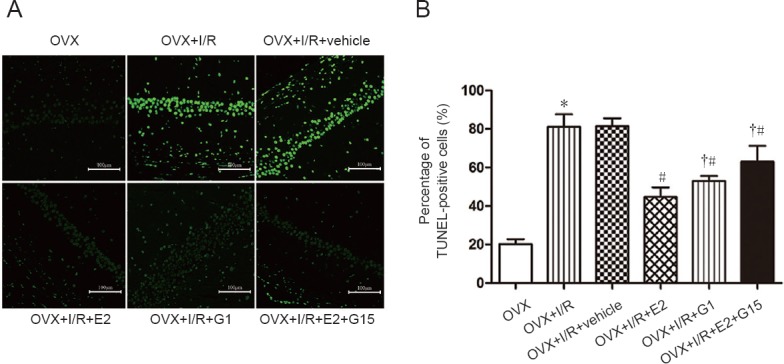
E2 and G1 reduce neuronal apoptosis in the hippocampal CA1 region after I/R.
(A) Neuronal apoptosis in the hippocampal CA1 region under confocal microscopy, as detected by TUNEL staining. Green: Apoptotic neurons. Fluorescent agent: FITC-conjugated anti-rabbit secondary antibody. Arrows indicate apoptotic neurons. Original magnification: 200×. Scale bars: 50 µm. (B) Number of TUNEL-positive cells. Data are expressed as the mean ± SEM (n = 6; one-way analysis of variance followed by Student-Newman-Keuls post hoc test). *P < 0.05, vs. OVX group; #P < 0.05, vs. OVX + I/R group; †P < 0.05, vs. OVX + I/R + E2 group. E2: 17-β-estradiol; G1: selective GPER agonist; G15: selective GPER inhibitor; I/R: ischemia/reperfusion; OVX: ovariectomized; TUNEL: terminal deoxynucleotidyl transferase (dUTP) nick end labeling.
Effects of E2 and G1 on the endoplasmic reticulum stress pathway after cerebral I/R injury in rats
Low expression levels of GRP78 (Figure 5B), caspase-12 (Figure 5C), and CHOP (Figure 5D) were observed in the OVX group. However, mRNA levels were significantly increased in OVX + I/R and OVX + I/R + vehicle groups compared with the OVX group 24 hours after brain I/R injury (P < 0.05; Figure 5). A non-significant difference was observed between OVX + I/R and OVX + I/R + vehicle groups (P > 0.05; Figure 5). After E2 or G1 treatment, levels of GRP78, caspase-12, and CHOP were significantly decreased compared with the OVX + I/R group (P < 0.05; Figure 5). Moreover, compared with the OVX + I/R + E2 group, these levels were significantly increased in the OVX + I/R + E2 + G15 group (P < 0.05; Figure 5).
Figure 5.
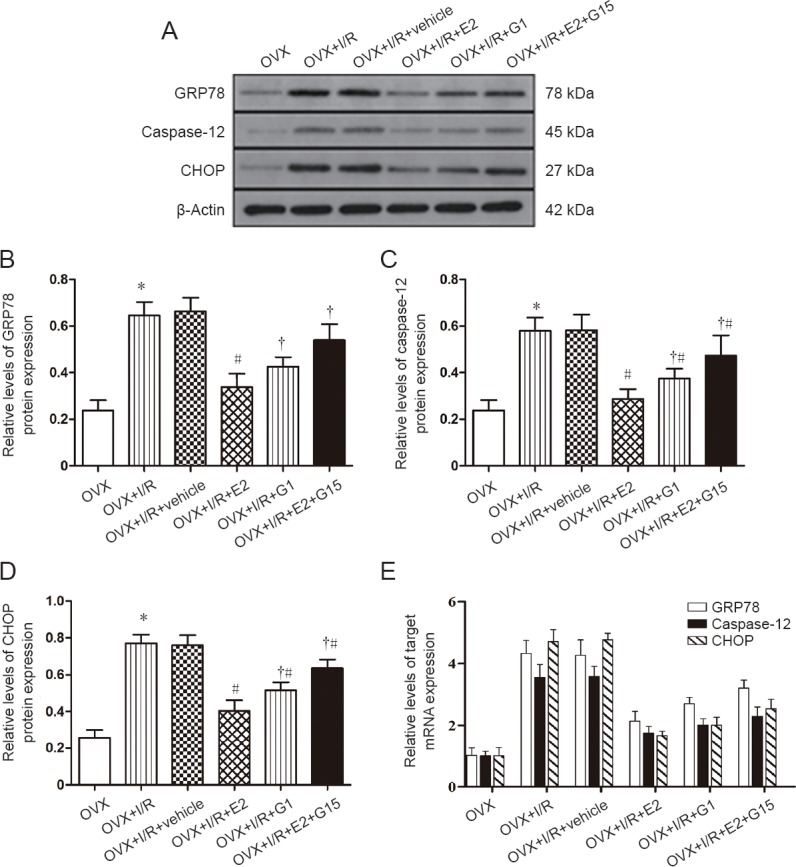
E2 and G1 inhibit expression of endoplasmic reticulum stress-related proteins in the hippocampal CA1 region after I/R.
(A) Western blot assay of GRP78, caspase-12, and CHOP expression. Top panels show target bands for GRP78, caspase-12, and CHOP protein, while the bottom panel shows the loading control beta-actin. (B–E) Relative levels of GRP78 (B), caspase-12 (C), and CHOP (D) protein expression. (E) Relative levels of GRP78, caspase-12, and CHOP mRNA expression. Data are expressed as the mean ± SEM (n = 6; one-way analysis of variance followed by Student-Newman-Keuls post hoc test). *P < 0.05, vs. OVX group; #P < 0.05, vs. OVX + I/R group; †P < 0.05, vs. OVX + I/R + E2 group. Caspase-12: Cysteinyl aspartate-specific proteinase 12; CHOP: C/EBP homologous protein; E2: 17-β-estradiol; G1: selective GPER agonist; G15: selective GPER inhibitor; GRP78: Glucose-regulated protein 78.
Figure 5E shows the effects of E2 and G1 on mRNA expression of endoplasmic reticulum stress-related factors after cerebral I/R injury in rats. mRNA expression of GRP78, caspase-12, and CHOP increased in OVX + I/R and OVX + I/R + vehicle groups compared with the OVX group after cerebral I/R injury (P < 0.05; Figure 5E). There were no significant differences in mRNA expression of GRP78, caspase-12, or CHOP between OVX + I/R and OVX + I/R + vehicle groups (P > 0.05; Figure 5E). Administration of E2 or G1 alone decreased the expression of GRP78, caspase-12, and CHOP mRNA compared with the OVX + I/R group (P < 0.05; Figure 5E). However, administration of E2 and G15 increased mRNA levels of GRP78, caspase-12, and CHOP compared with the OVX + I/R + E2 group (P < 0.05; Figure 5E).
Detection of the endoplasmic reticulum stress-specific protein caspase-12 by immunofluorescence revealed significantly increased levels in OVX + I/R and OVX + I/R + vehicle groups compared with the OVX group 24 hours after brain I/R injury (P < 0.05; Figure 6). Injection of E2 and G1 after reperfusion resulted in significantly decreased expression levels of caspase-12 in rat brains compared with the OVX + I/R group (P < 0.05; Figure 6). Notably, caspase-12 expression was lower in the OVX + E2 group than in the OVX + G1 group (P < 0.05; Figure 6), although they were increased compared with OVX + E2 and OVX + G1 groups (P < 0.05; Figure 6). Immunofluorescence results were consistent with expression trends for these proteins identified by western blot assay (Figure 5A and C).
Figure 6.

E2 and G1 reduce expression and distribution of caspase-12 in the hippocampal CA1 region after I/R.
(A) Representative images of caspase-12 immunofluorescence. Blue: caspase-12; red: propidium iodide; fluorescent agent: FITC-conjugated anti-rabbit secondary antibody. Arrows indicate positive cells. Original magnification: 200×. Scale bar: 50 µm. (B) Fluorescence intensity of caspase-12. Data are expressed as the mean ± SEM (n = 6; one-way analysis of variance followed by Student-Newman-Keuls post hoc test). *P < 0.05, vs. OVX group; #P < 0.05, vs. OVX + I/R group; †P < 0.05, vs. OVX + I/R + E2 group. E2: 17-β-estradiol; G1: selective GPER agonist; G15: selective GPER inhibitor; I/R: ischemia/reperfusion; OVX: ovariectomized.
Discussion
In this study, we examined neuroprotective and anti-apoptotic effects of E2 and GPER activation on I/R injury in an animal model. We demonstrated that activation of GPER, similar to E2, noticeably reduces infarct volume and the percentage of neuronal apoptosis, and improves neurological function. Furthermore, E2 and GPER activation treatments remarkably increased brain water content and infarct volume after cerebral I/R injury in rats. Additionally, apoptosis was obviously attenuated, as expression levels of endoplasmic reticulum stress-related proteins GRP78, CHOP, and caspase-12 were decreased in OVX + I/R + E2 and OVX + I/R + G1 groups 24 hours after I/R injury. E2 and GPER activation efficiently suppressed apoptosis by inhibiting the endoplasmic reticulum stress-mediated apoptotic pathway. However, the above-mentioned effect of E2 on I/R injury was abolished by the GPER inhibitor G15.
G1 and G15 are highly selective agonists and inhibitors of GPER, respectively, and their discovery promotes the functional study of GPER in animal cells and tissues (Li et al., 2013; Barton and Prossnitz, 2015; Wang et al., 2018a, b). Many studies have shown that GPER activation has the same role as E2 in breast cancer cells, and inhibits the occurrence of breast cancer through the GPER/reactive oxygen species/p38MAPK/p21 signaling pathway; notably, G15 inhibits this effect (Santolla et al., 2015). The neural repair effect of E2 has been observed in ischemic stroke (Simpkins et al., 1997), experimental stroke (Toung et al., 1998), glioma, and neuroblastoma cells in vitro (Bishop and Simpkins, 1994). The results of this study showed that administration of the GPER receptor agonist G1 in the lateral ventricle can remarkably improve neurobehavioral function and reduce brain water content. Other experts have also shown that G1 ameliorates blood-brain barrier permeability after global cerebral ischemia in OVX rats (Lu et al., 2016). According to our results, G1 may also reduce the volume of cerebral infarction after I/R in OVX rats, consistent with the results of Zhang et al. (2010) showing that G1 reduced infarct size after experimental stroke. These results indicate that E2 and GPER activation may have considerable potential for treating neurodegenerative diseases and central nervous system injuries. Therefore, this study explored whether estrogen and activation of GPER have protective functions against I/R injury, and investigated potential mechanisms of these neuroprotective effects.
Since the discovery of GPER in the brain, some scholars have suggested that GPER acts as a classic estrogen receptor that can provide neuroprotection, such as against I/R injury (Owman et al., 1996a; Deschamps and Murphy, 2009; Li et al., 2015). The GPER receptor agonist used here (G1) does not interact with classical nuclear receptors (Bologa et al., 2006b). Thus, it can be used as an alternative drug for postmenopausal women, because it does not elicit adverse effects of hormone replacement therapy (Gillies and McArthur, 2010). Moreover, Nissl staining results showed distinct pyramidal neurons in the hippocampal CA1 subfield and cytoplasmic staining after the addition of G1 treatment. Our findings clearly indicate that G1 treatment can improve neurological deficits. The treatment selectively attenuated infarct damage in the OVX + I/R group, consistent with findings for the E2 treatment. However, these effects were abrogated by the addition of the GPER antagonist G15, thereby confirming that GPER plays an important role for the activity of many estrogen receptors. Further research on this topic is necessary.
To ensure that a high drug concentration was delivered to the damaged brain area, we employed lateral ventricle injection rather than intravenous or intramuscular injection. Given that 100 µg/kg estrogen is the best therapeutic dose for the employed I/R injury model (Yang et al., 2000), we used 100 µg/kg G1 and 100 µg/kg G15 in this preliminary experiment to compare their therapeutic effects with that of E2, considering factors such as the tolerance of rats and best experimental results. The frequency of cortical neuron apoptosis in OVX + I/R + E2 and OVX + I/R + G1 groups was reduced compared with the OVX + I/R group. However, when G15 was added, the therapeutic effect of E2 was noticeably reduced, indicating that GPER plays an important role in the therapeutic benefits elicited by E2.
The present results show that neuronal apoptosis in the hippocampus and expression levels of key apoptotic proteins, such as GRP78, CHOP, and caspase-12, were increased in rat brains after I/R injury. However, the number of apoptotic neurons and level of protein expression decreased after treatment with GPER agonists. Many experimental studies have indicated that estrogen signaling affects stroke-induced injury by modulating apoptotic signaling mechanisms (Dubal et al., 1999; Rau et al., 2003; Liu et al., 2009, 2011). However, this novel study of E2 and GPER indicates that they can protect neurons in the hippocampus through the endoplasmic reticulum stress pathway. GRP78, which is important in cellular responses to stress, is involved in endoplasmic reticulum (ER) glycoprotein trafficking and molecular chaperone activity (Yong et al., 1987; Wooden et al., 1991; Little et al., 1994; Brostrom and Brostrom, 1997). In normal conditions, GRP78 associates with ER-localized transmembrane proteins and maintains an inactive form. However, under pathological conditions, GRP78 dissociates from these receptors, thereby resulting in activation of these proteins. Moreover, when severe endoplasmic reticulum stress disrupts ER function, the ER induces apoptosis by activating downstream signaling molecules, such as CHOP and caspase-12. In the present experiment, E2 and G1 considerably reduced neuronal apoptosis and expression levels of GRP78, CHOP, and caspase-12 in the hippocampus of I/R injury rats. Notably, the therapeutic effect of E2 was slightly stronger than that of G1. However, protective effects were inhibited after the addition of G15. Our results indicate that G1 can reduce apoptosis induced by the endoplasmic reticulum stress pathway.
Rats deficient for the caspase-12 gene were unable to induce apoptosis in endoplasmic reticulum stress. In other words, caspase-12 is essential in the mechanism of endoplasmic reticulum stress-mediated apoptosis and specific to endemic stress in the ER (Nakagawa et al., 2000). Immunofluorescence experiments revealed noticeably increased caspase-12 levels after I/R injury, although G1 injection after reperfusion obviously decreased caspase-12 expression levels in rat brains. This further shows that G1 has a specific effect on endoplasmic reticulum stress.
This study has some limitations. The most effective dosages of E2 and G1, and time window of treatment require further investigation. Moreover, potential effects of non-endoplasmic reticulum stress apoptotic proteins or other signaling pathways on this protective mechanism need to be elucidated. Therefore, future studies must focus on the interactions between these pathways.
In summary, GPER activation protects the brain after I/R injury and reduces apoptosis via the endoplasmic reticulum stress-mediated apoptotic pathway. This result provides new insight into treatment of I/R injury. Nevertheless, the results require further validation.
Additional file: Open peer review report 1 (61.8KB, pdf) .
Acknowledgments
We thank the Key Laboratory of Xinjiang Endemic and Ethnic Diseases of Xinjiang Provincial Department of Physiology, School of Medicine, Shihezi University for excellent technical assistance.
Footnotes
Conflicts of interest: The authors declare that there are no conflicts of interest associated with this manuscript.
Financial support: This study was supported by the National Natural Science Foundation of China, No. 81560175, No. 81260159 (both to LL). The funding sources had no role in study design, conception, analysis or interpretation of data, writing and deciding to submit this paper for publication.
Institutional review board statement: The experimental protocol was approved by the Institutional Ethics Review Board at the First Affiliated Hospital of Shihezi University School of Medicine, China (approval No. SHZ A2017-171) on February 27, 2017. All experimental procedures described here were in accordance with the National Institutes of Health (NIH) guidelines for the Care and Use of Laboratory Animals.
Copyright license agreement: The Copyright License Agreement has been signed by all authors before publication.
Data sharing statement: Datasets analyzed during the current study are available from the corresponding author on reasonable request.
Plagiarism check: Checked twice by iThenticate.
Peer review: Externally peer reviewed.
Open peer reviewer: Fabricio Ferreira de Oliveira, Universidade Federal de Sao Paulo, Brazil.
Funding: This study was supported by the National Natural Science Foundation of China, No. 81560175, 81260159 (both to LL).
P-Reviewer: Ferreira de Oliveira F; C-Editor: Zhao M; S-Editors: Wang J, Li CH; L-Editors: Deusen AV, Qiu Y, Song LP; T-Editor: Liu XL
References
- 1.Barton M, Prossnitz ER. Emerging roles of GPER in diabetes and atherosclerosis. Trends Enocrinol Metab. 2015;26:185–192. doi: 10.1016/j.tem.2015.02.003. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 2.Bell KF, Ducatenzeiler A, Ribeiro-Da-Silva A, Duff K, Bennett DA, Cuello AC. The amyloid pathology progresses in a neurotransmitter-specific manner. Neurobiol Aging. 2006;27:1644–1657. doi: 10.1016/j.neurobiolaging.2005.09.034. [DOI] [PubMed] [Google Scholar]
- 3.Bishop J, Simpkins JW. Estradiol treatment increases viability of glioma and neuroblastoma cells in vitro. Mol Cell Neurosci. 1994;5:303–308. doi: 10.1006/mcne.1994.1036. [DOI] [PubMed] [Google Scholar]
- 4.Bologa CG, Revankar CM, Young SM, Edwards BS, Arterburn JB, Kiselyov AS, Parker MA, Tkachenko SE, Savchuck NP, Sklar LA. Virtual and biomolecular screening converge on a selective agonist for GPR30. Nat Chem Biol. 2006a;2:207–212. doi: 10.1038/nchembio775. [DOI] [PubMed] [Google Scholar]
- 5.Bologa CG, Revankar CM, Young SM, Edwards BS, Arterburn JB, Kiselyov AS, Parker MA, Tkachenko SE, Savchuck NP, Sklar LA, Oprea TI, Prossnitz ER. Virtual and biomolecular screening converge on a selective agonist for GPR30. Nat Chem Biol. 2006b;2:207–212. doi: 10.1038/nchembio775. [DOI] [PubMed] [Google Scholar]
- 6.Brostrom CO, Brostrom MA. Regulation of translational initiation during cellular responses to stress. Prog Nucleic Acid Res Mol Biol. 1997;58:79–125. doi: 10.1016/s0079-6603(08)60034-3. [DOI] [PubMed] [Google Scholar]
- 7.Buttesmith MD, Gulinello M, Zukin RS, Etgen AM. Chronic estradiol treatment increases CA1 cell survival but does not improve visual or spatial recognition memory after global ischemia in middle-aged female rats. Horm Behav. 2009;55:442–453. doi: 10.1016/j.yhbeh.2008.11.011. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 8.Chan PH. Role of oxidants in ischemic brain damage. Stroke. 1996;27:1124–1129. doi: 10.1161/01.str.27.6.1124. [DOI] [PubMed] [Google Scholar]
- 9.Deschamps AM, Murphy E. Activation of a novel estrogen receptor, GPER, is cardioprotective in male and female rats. Am J Physiol Heart Circ Physiol. 2009;297:H1806–H1813. doi: 10.1152/ajpheart.00283.2009. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 10.Dubal DB, Shughrue PJ, Wilson ME, Merchenthaler I, Wise PM. Estradiol modulates bcl-2 in cerebral ischemia: a potential role for estrogen receptors. J Neurosci. 1999;19:6385. doi: 10.1523/JNEUROSCI.19-15-06385.1999. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 11.Feng YH, Zhu ZH, Wu CX, Zhou GP. Effects of electroacupuncture at points selected by orthogonal experiment on the extracellular signal regulated kinase signal pathway in a rat model of cerebral ischenia-reperfusion injury. Zhongguo Zuzhi Gongcheng Yanjiu. 2016;20:5953–5958. [Google Scholar]
- 12.Gillies GE, McArthur S. Estrogen actions in the brain and the basis for differential action in men and women: a case for sex-specific medicines. Pharmacol Rev. 2010;62:155–198. doi: 10.1124/pr.109.002071. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 13.Gupta S, Gupta YK. Combination of Zizyphus jujuba and silymarin showed better neuroprotective effect as compared to single agent in MCAo-induced focal cerebral ischemia in rats. J Ethnopharmacol. 2017;197:118–127. doi: 10.1016/j.jep.2016.07.060. [DOI] [PubMed] [Google Scholar]
- 14.Gwag BJ, Moon GJ, Suh J, Jin HL, Sun JS, Sun MW, Park UJ. Induction of the unfolded protein response and cell death pathway in Alzheimer’s disease, but not in aged Tg2576 mice. Exp Mol Med. 2010;42:386. doi: 10.3858/emm.2010.42.5.040. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 15.Herson PS, Koerner IP, Hurn PD. Sex, sex steroids, and brain injury. In: Seminars in reproductive medicine. Semin Reprod Med. 2009 doi: 10.1055/s-0029-1216276. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 16.Hurn PD, Subramanian S, Parker SM, Afentoulis ME, Kaler LJ, Vandenbark AA, Offner H. T- and B-cell-deficient mice with experimental stroke have reduced lesion size and inflammation. J Cereb Blood Flow Metab. 2007;27:1798–1805. doi: 10.1038/sj.jcbfm.9600482. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 17.Imoto S, Ohkura H, Sugano K, Sasaki Y, Ito K, Igarashi T, Ohtsu T, Fujii H, Minami H, Hasebe T. Determination of cytosol c-erbB-2 protein in breast cancer by sandwich enzyme immunoassay. Jpn J Clin Oncol. 1998;28:92. doi: 10.1093/jjco/28.2.92. [DOI] [PubMed] [Google Scholar]
- 18.Jia J, Guan D, W, Alkayed N, Wang M, Hua Z, Xu Y. Estrogen inhibits Fas-mediated apoptosis in experimental stroke. Exp Neurol. 2009;215:48–52. doi: 10.1016/j.expneurol.2008.09.015. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 19.Lebesgue D, Chevaleyre V, Zukin RS, Etgen AM. Estradiol rescues neurons from global ischemia-induced cell death: multiple cellular pathways of neuroprotection. Steroids. 2009;74:555–561. doi: 10.1016/j.steroids.2009.01.003. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 20.Li WL, Xiang W, Ping Y. Activation of novel estrogen receptor GPER results in inhibition of cardiocyte apoptosis and cardioprotection. Mol Med Rep. 2015;12:2425–2430. doi: 10.3892/mmr.2015.3674. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 21.Li YR, Ren C, Zhang Q, Li JC, Chian RC. Expression of G protein estrogen receptor (GPER) on membrane of mouse oocytes during maturation. J Assist Reprod Genet. 2013;30:227–232. doi: 10.1007/s10815-013-9942-z. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 22.Little E, Ramakrishnan M, Roy B, Gazit G, Lee AS. The glucose-regulated proteins (GRP78 and GRP94): functions, gene regulation, and applications. Crit Rev Eukaryot Gene Expr. 1994;4:1–18. doi: 10.1615/critreveukargeneexpr.v4.i1.10. [DOI] [PubMed] [Google Scholar]
- 23.Liu F, Li Z, Li J, Siegel C, Yuan R, Mccullough LD. Sex differences in caspase activation after experimental stroke. Stroke. 2009;40:1842. doi: 10.1161/STROKEAHA.108.538686. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 24.Liu F, Lang J, Li J, Benashski SE, Siegel M, Xu Y, Mccullough LD. Sex differences in the response to poly (ADP-ribose) polymerase-1 deletion and caspase inhibition after stroke. Stroke. 2011;42:1090. doi: 10.1161/STROKEAHA.110.594861. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 25.Liu SB, Zhang N, Guo YY, Zhao R, Shi TY, Feng SF, Wang SQ, Yang Q, Li XQ, Wu YM, Ma L, Hou Y, Xiong LZ, Zhang W, Zhao MG. G-protein-coupled receptor 30 mediates rapid neuroprotective effects of estrogen via depression of NR2B-containing NMDA receptors. J Neurosci. 2012;32:4887–4900. doi: 10.1523/JNEUROSCI.5828-11.2012. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 26.Longa EZ, Weinstein PR, Carlson S, Cummins R. Reversible middle cerebral artery occlusion without craniectomy in rats. Stroke. 1989;20:84. doi: 10.1161/01.str.20.1.84. [DOI] [PubMed] [Google Scholar]
- 27.Lu D, Qu Y, Shi F, Feng D, Tao K, Gao G, He S, Zhao T. Activation of G protein-coupled estrogen receptor 1 (GPER-1) ameliorates blood-brain barrier permeability after global cerebral ischemia in ovariectomized rats. Biochem Biophys Res Commun. 2016;477:209–214. doi: 10.1016/j.bbrc.2016.06.044. [DOI] [PubMed] [Google Scholar]
- 28.Ma J, Bao L, Xia X, Feng Q, Zhou Y, Wang Y, Cao Z. miRNA-128b promotes cerebral infarction by regulating the expressions of Bcl-2 and Caspase 3. World Neurosurg. 2018;15:198. doi: 10.1016/j.wneu.2018.11.144. [DOI] [PubMed] [Google Scholar]
- 29.Mattson MP, Duan W, Pedersen WA, Culmsee C. Neurodegenerative disorders and ischemic brain diseases. Apoptosis. 2001;6:69–81. doi: 10.1023/a:1009676112184. [DOI] [PubMed] [Google Scholar]
- 30.Morris RGM, Garrud P, Rawlins JNP, O’Keefe J. Place navigation impaired in rats with hippocampal lesions. Nature. 1982;297:681–683. doi: 10.1038/297681a0. [DOI] [PubMed] [Google Scholar]
- 31.Nakagawa T, Zhu H, Morishima N, Li E, Xu J, Yankner BA, Yuan J. Caspase-12 mediates endoplasmic-reticulum-specific apoptosis and cytotoxicity by amyloid-beta. Nature. 2000;403:98–103. doi: 10.1038/47513. [DOI] [PubMed] [Google Scholar]
- 32.Offner H, Subramanian S, Parker SM, Afentoulis ME, Vandenbark AA, Hurn PD. Experimental stroke induces massive, rapid activation of the peripheral immune system. J Cereb Blood Flow Metab. 2006;26:654–665. doi: 10.1038/sj.jcbfm.9600217. [DOI] [PubMed] [Google Scholar]
- 33.Owman C, Blay P, Nilsson C, Lolait SJ. Cloning of human cDNA encoding a novel heptahelix receptor expressed in Burkitt’s lymphoma and widely distributed in brain and peripheral tissues. Biochem Biophys Res Commun. 1996a;228:285–292. doi: 10.1006/bbrc.1996.1654. [DOI] [PubMed] [Google Scholar]
- 34.Prossnitz ER, Barton M. Estrogen biology: New insights into GPER function and clinical opportunities. Mol Cell Endocrinol. 2014;389:71–83. doi: 10.1016/j.mce.2014.02.002. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 35.Rau SW, Dubal DB, Böttner M, Gerhold LM, Wise PM. Estradiol attenuates programmed cell death after stroke-like injury. J Neurosci. 2003;23:11420–11426. doi: 10.1523/JNEUROSCI.23-36-11420.2003. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 36.Sano R, Reed JC. ER stress-induced cell death mechanisms. Biochim Biophys Acta. 2013;1833:3460–3470. doi: 10.1016/j.bbamcr.2013.06.028. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 37.Santolla MF, Avino S, Pellegrino M, Francesco EMD, Marco PD, Lappano R, Vivacqua A, Cirillo F, Rigiracciolo DC, Scarpelli A. SIRT1 is involved in oncogenic signaling mediated by GPER in breast cancer. Cell Death Dis. 2015;6:e1834. doi: 10.1038/cddis.2015.201. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 38.Scull CM, Tabas I. Mechanisms of ER stress-induced apoptosis in atherosclerosis. Arterioscler Thromb Vasc Biol. 2011;31:2792. doi: 10.1161/ATVBAHA.111.224881. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 39.Shughrue PJ, Merchenthaler I. Estrogen prevents the loss of CA1 hippocampal neurons in gerbils after ischemic injury. Neuroscience. 2003;116:851–861. doi: 10.1016/s0306-4522(02)00790-x. [DOI] [PubMed] [Google Scholar]
- 40.Shi D, Li Z, Yang J, Liu BZ, Xia H. Symptom experience and symptom burden of patients following first-ever stroke within 1 year: a cross-sectional study. Neural Regen Res. 2018;13:1907–1912. doi: 10.4103/1673-5374.239440. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 41.Silva RM, Ries V, Oo TF, Yarygina O, Jackson‐Lewis V, Ryu EJ, Lu PD, Marciniak SM, Ron D, Przedborski S. CHOP/GADD153 is a mediator of apoptotic death in substantia nigra dopamine neurons in an in vivo neurotoxin model of parkinsonism. J Neurochem. 2005;95:974–986. doi: 10.1111/j.1471-4159.2005.03428.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 42.Simpkins JW, Rajakumar G, Zhang YQ, Simpkins CE, Greenwald D, Yu CJ, Bodor N, Day AL. Estrogens may reduce mortality and ischemic damage caused by middle cerebral artery occlusion in the female rat. J Neurosurg. 1997;87:724–730. doi: 10.3171/jns.1997.87.5.0724. [DOI] [PubMed] [Google Scholar]
- 43.Toung TJK, Traystman RJ, Hurn PD. Estrogen-mediated neuroprotection after experimental stroke in male rats. Stroke. 1998;29:1666. doi: 10.1161/01.str.29.8.1666. [DOI] [PubMed] [Google Scholar]
- 44.Tropea T, Francesco EMD, Rigiracciolo D, Maggiolini M, Wareing M, Osol G, Mandalà M. Pregnancy augments g protein estrogen receptor (GPER) induced vasodilation in rat uterine arteries via the nitric oxide - cGMP signaling pathway. PLoS One. 2015;10:e0141997. doi: 10.1371/journal.pone.0141997. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 45.Vavers E, Zvejniece L, Svalbe B, Volska K, Makarova E, Liepinsh E, Rizhanova K, Liepins V, Dambrova M. The neuroprotective effects of R-phenibut after focal cerebral ischemia. Pharmacol Res. 2016;113:796–801. doi: 10.1016/j.phrs.2015.11.013. [DOI] [PubMed] [Google Scholar]
- 46.Wang C, Li J, Ye S, Zhang Y, Li P, Wang L, Wang TH. Oestrogen inhibits VEGF expression and angiogenesis in triple-negative breast cancer by activating GPER-1. J Cancer. 2018a;9:3802–3811. doi: 10.7150/jca.29233. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 47.Wang Z, Chen X, Zhao Y, Jin Y, Zheng J. G-protein-coupled estrogen receptor suppresses the migration of osteosarcoma cells via post-translational regulation of Snail. J Cancer Res Clin Oncol. 2018b doi: 10.1007/s00432-018-2768-4. doi: 10.1007/s00432-018-2768-4. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 48.Wooden SK, Li LJ, Navarro D, Qadri I, Pereira L, Lee AS. Transactivation of the grp78 promoter by malfolded proteins, glycosylation block, and calcium ionophore is mediated through a proximal region containing a CCAAT motif which interacts with CTF/NF-I. Mol Cell Biol. 1991;11:5612. doi: 10.1128/mcb.11.11.5612. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 49.Yang SH, Shi J, Day AL, Simpkins JW. Estradiol exerts neuroprotective effects when administered after ischemic insult. Stroke. 2000;31:745–749. doi: 10.1161/01.str.31.3.745. [DOI] [PubMed] [Google Scholar]
- 50.Yong KK, Kim KS, Lee AS. Regulation of the glucose-regulated protein genes by β-mercaptoethanol requires de novo protein synthesis and correlates with inhibition of protein glycosylation. J Cell Physiol. 1987;133:553–559. doi: 10.1002/jcp.1041330317. [DOI] [PubMed] [Google Scholar]
- 51.Yuan M, Ge M, Yin J, Dai Z, Xie L, Li Y, Liu X, Peng L, Zhang G, Si J, Ma K, Wang S. Isoflurane post-conditioning down-regulates expression of aquaporin 4 in rats with cerebral ischemia/reperfusion injury and is possibly related to bone morphogenetic protein 4/Smad1/5/8 signaling pathway. Biomed Pharmacother. 2018;97:429–438. doi: 10.1016/j.biopha.2017.10.082. [DOI] [PubMed] [Google Scholar]
- 52.Zhang B, Subramanian S, Dziennis S, Jia J, Uchida M, Akiyoshi K, Migliati E, Lewis AD, Vandenbark AA, Offner H, Hurn PD. Estradiol and G1 reduce infarct size and improve immunosuppression after experimental stroke. J Immunol. 2010;184:4087–4094. doi: 10.4049/jimmunol.0902339. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 53.Zhang Z, Pei Q, Deng Y, Zhi M, Hang G, Guo H, Hou Y, Wang S, Zou W, Sun Y. The novel estrogenic receptor GPR30 alleviates ischemic injury by inhibiting TLR4-mediated microglial inflammation. J Neuroinflammation. 2018;15:206. doi: 10.1186/s12974-018-1246-x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 54.Zheng HZ, Jiang W, Zhao XF, Du J, Liu PG, Chang LD, Li WB, Hu HT, Shi XM. Electroacupuncture induces acute changes in cerebral cortical miRNA profile, improves cerebral blood flow and alleviates neurological deficits in a rat model of stroke. Neural Regen Res. 2016;11:1940–1950. doi: 10.4103/1673-5374.197135. [DOI] [PMC free article] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.