

Abstract
The use of botanical dietary supplements is becoming increasingly popular for the alleviation of hormonal-based conditions such as hot flashes, premenstrual syndrome, and fertility. Estrogen and progesterone receptors (ER and PR) play an essential role in these processes. However, despite the fact that many therapies used to alleviate gynecological conditions act through PR-mediated mechanisms, few studies have investigated or identified any herbal natural product components that act on this receptor. In the current study, we used a progesterone response element (PRE)-luciferase (Luc) reporter assay to identify four phytoprogestins present in a standardized red clover extract. While looking for compounds in red clover, a synergistic interaction was identified between progesterone and irilone (1) in both endometrial and ovarian cancer cell lines. In these cancers, progesterone action is generally associated with positive outcomes, thus the synergistic activity of 1 may provide entirely new strategies for enhancing progesterone signaling as a means of mitigating conditions such as fibroids and endometriosis. Formononetin (3) and biochanin A (4) exhibited mixed antagonist activity, while prunetin (2) acted only as an antagonist. Collectively these results suggest that the effects of red clover extract repeatedly observed in cultured cells, and the inverse correlation between risk of various cancers and flavonoid intake may be due, in part, to altered progesterone signaling.
Graphical Abstract
The use of botanical dietary supplements is becoming increasingly popular for the alleviation of hormonal-based conditions such as hot flashes, premenstrual syndrome, and fertility.1 An estimated 13.7 million dollars is spent annually by Americans on alternative products, including on health supplements marketed specifically toward women. Estrogen and progesterone receptors (ER, PR), and their corresponding steroid hormones (estradiol and progesterone), act through complex mechanisms to regulate biological processes critical for woman’s health. These receptors play an essential role in the treatment of women who develop endometriosis, the relief of symptoms associated with hot flashes, and the incidence of reproductive cancers such as ovarian and breast.1,14–22 However, despite the fact that many drugs used to alleviate these issues act through PR-mediated mechanisms, few studies have investigated or identified the natural product components found in herbal supplements that act on the progesterone receptor, particularly when compared to literature that focuses on its estrogenic counterpart. Given the accepted role of estrogen receptor modulators in botanicals, ligands for other hormone receptors, such as the progesterone receptor, are likely present and bioactive in supplements used for women’s health.2 Investigations into botanicals as a source of progestins is significant, since several ubiquitously distributed natural product structural classes may interact with PR receptor binding, and as a result may currently be affecting the health of millions of women worldwide.3
One botanical supplement with reported ER/PR activity is red clover (Trifolium pratense L. (Fabaceae)), which is commonly used to alleviate menopausal symptoms. Though T. pratense is known to contain phytoestrogens, it was shown that extracts did not increase uterine weight in rats or the proliferative index in the endometrium of women.4,5 These data suggested that other components, such as progestins, may be inhibiting the observed estrogenic activity, since progestins inhibit estrogen-induced increases in uterine proliferation.6 We previously reported that the red clover constituent, kaempferol, exhibited progestogenic effects in ovariectomized rats, though its abundance and potency profile did not account for all of the progesterone activity observed in the crude extract.7,8 This suggested that other progesterone-like compounds were present in red clover. In the current study, we employed a progesterone response element (PRE)-luciferase (Luc) reporter assay in Ishikawa cells to characterize the agonist and antagonist effects of red clover extracts, and to identify molecules responsible for the activity.
RESULTS AND DISCUSSION
In order to identify phytoprogestins from T. pratense, we obtained a standardized phase II red clover extract. This extract was previously characterized by the Botanical Center at the University of Illinois at Chicago, and they described the identification of 23 components of isoflavone, flavonoid, pterocarpin, and coumarin structural classes.9 In the present study, Ishikawa cells that stably express progesterone receptor B (PR-B) were used to screen for compounds that activate or inhibit the receptor. Inhibition using this assay is based on the compound’s ability to block activation of the progesterone receptor in the presence of progesterone. Initially, 60 μg/mL of the red clover extract decreased luciferase activity by 44% (p<0.001) compared to a DMSO control. Furthermore, this same concentration of the extract blocked activation of PR-B in the presence of 100 μM progesterone, as evidenced by a reduction of luciferase activity by 94%, compared to the 98% reduction by RU486, a PR antagonist (Figure 1A and 1B).
Figure 1.

Red clover contains progesterone antagonists. A–C) Progesterone-response element (PRE/Luc) activity in Ishikawa cells stably expressing progesterone receptor B. Cells were treated with red clover extract (A and B), and subsequent HPLC fractions (C) at 60 μg/mL. Significantly different from Control (DMSO), ***p<0.001, ****p<0.0001, n≥3.
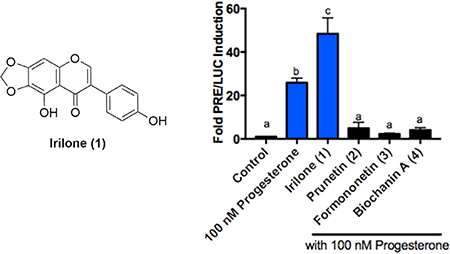
To determine which constituents of red clover were responsible for inhibiting PRE/Luc activity, the red clover extract was separated using reversed-phase HPLC to afford fractions RE-1 to RE-19. At 60 μg/mL, antagonist activity was greatest among the more non-polar fractions (RE-13 to RE-18, Figure 1C). Further exploration of these fractions led to the isolation of irilone (1), prunetin (2), formononetin (3) and biochanin A (4) (Figure 2A).
Figure 2.

A) Structures isolated from T. pratense. B) Structure of progesterone. C) PRE/Luc activity in Ishikawa PR-B cells treated with 50 μM of irilone (1), prunetin (2), formononetin (3), and biochanin A (4), and progesterone as indicated. Bars without the same letter (a-c) are significantly different from each other at p<0.05. n≥3.
Irilone (1) was identified upon comparison of 1H and 13C NMR shifts, and HRMS data to values reported in the literature.10 Formononetin (3) and biochanin A (4) were identified upon comparison of 1H and 13C NMR shifts and HRESIITTOF MS data to literature values,11,12 and comparison of retention times to authenticated standards. Prunetin (2) was previously characterized in the T. pratense extract by the UIC Botanical Center,9 and we confirmed its identity by HRESIITTOF MS analysis and comparison of its retention time to an authenticated standard. All relevant spectra (NMR and MS) and chromatograms can be found in the Supporting Information.
Biological Activity of Red Clover Components.
All biological assays were performed on the purified natural product (1) and the purchased standard compounds (2–4). Compounds 1–4 were tested in our PRE/Luc assay at 50 μM in the presence of 100 nM of progesterone (structure of progesterone, Figure 2B). Compounds 2–4 each inhibited the progesterone-induced increase in PRE/Luc activity by >90%. Surprisingly, 50 μM of irilone (1) resulted in an increase in PRE/Luc activity over that induced by 100 nM progesterone alone, increasing PRE/Luc induction from 25.7-fold to 48.4-fold (p<0.05; Figure 2C).
Compounds 2–4 Exhibit Antagonistic Activity with Progesterone.
To better characterize the effects of each compound, dose response curves were generated both in the presence of 100 nM of progesterone and in the absence of progesterone. Compounds 2–4 displayed strong antagonist activity for the progesterone receptor, with concentrations ≥ 25 μM dramatically reducing PRE/Luc activity in the presence of 100 nM of progesterone for all three compounds (p<0.01; Figure 3A–C). Even in the absence of progesterone, prunetin (2) decreased basal PRE/Luc activity at 25 and 50 μM (p<0.0001; Figure 3D). Formononetin (3) and biochanin A (4) decreased basal PRE/Luc activity at 50 μM (p<0.05; Figure 3E and 3F, respectively).
Figure 3.

Formononetin (3) and biochanin A (4) have mixed antagonist activity, while prunetin (2) exhibited antagonist activity. A–C) PRE/Luc activity in Ishikawa PR-B cells treated with prunetin (A), formononetin (B), and biochanin A (C) in the presence of 100 nM of progesterone. D–F) PRE/Luc activity in Ishikawa PR-B cells treated with prunetin (D), formononetin (E), and biochanin A (F). Significantly different from 0 μM, *p<0.05, **p<0.01, ***p<0.001, ****p<0.0001, n≥3.
Formononetin (3) and biochanin A (4) also exhibited weak agonistic effects. In the presence of progesterone 1 μM of 3 increased PRE/Luc activity over that induced by progesterone by 57% (P<0.05; Figure 3A). In the absence of progesterone, 1 μM of 4 increased PRE/Luc activity by 55% (P<0.01; Figure 3F). In the presence of progesterone, 3 tended to increase PRE/Luc activity, but this did not reach statistical significance (P=0.097; Figure 3C). The agonistic effects of 3 and 4 are likely due to their weak phytoestrogen activity, which would increase endogenous PR expression.13
Irilone (1) Exhibits Synergistic Activity with Progesterone.
In the presence of 100 nM of progesterone, irilone (1) exhibited a dose-dependent increase in PRE/Luc activity, with 25 and 50 μM of 1 increasing PRE/Luc activity 138% and 151% higher than progesterone alone (p<0.01; Figure 4A). In the absence of progesterone, 1 (1–50 μM) had no observable effect on PRE/Luc activity (Figure 4B). This was a surprising finding, as previous studies found natural products that interacted with the PR had antagonist activity or no activity when tested in the presence of other PR ligands.7,8,14,15 Only one other natural product study reported synergistic activity with steroid signaling.16 In that study, extracts from Eucommia ulmoides synergistically increased androgen signaling, even in the presence of saturating concentrations of testosterone, but the compound responsible for this action was not isolated or further characterized.
Figure 4.

Irilone (1) exhibited synergistic activity with progesterone. A) PRE/Luc activity in Ishikawa PR-B cells treated with irilone (1) in the presence of 100 nM of progesterone. B) PRE/Luc activity after treating Ishikawa cells with 1. Significantly different from 0 μM, **p<0.01, n≥3.
It is currently not clear how these natural products could mechanistically exert the synergistic increase in steroid receptor signaling. The progesterone receptor interacts with numerous coactivators and corepressors that increase or decrease PR-mediated transcription, respectively.17 Therefore, one possibility is that irilone (1) induces synergistic activity by interacting with these coactivators and corepressors to enhance PR when bound to progesterone. Another potential explanation is that transcriptional activity of the PR is controlled by multiple phosphorylation sites, raising the possibility that synergy between 1 and progesterone could be due to 1 altering the phosphorylation status of the PR.18 In support of this possibility, a recent computer modeling study predicted irilone to have protein tyrosine kinase inhibitory activity.19 The PRE/Luc construct used in the current study has a simple thymidine kinase promoter with two PRE elements driving expression of luciferase. Future work should involve complex promoters that include distal regulatory units that may be important in controlling progesterone-stimulated gene expression.
Progesterone and synthetic compounds that interact with the PR frequently trigger degradation of the receptor.20 However, our previous report showed that kaempferol did not cause degradation of the PR.7 To determine if 1–4 trigger degradation of the PR, Ishikawa PR-B cells were treated with 25 and 50 μM of each compound for 24 hours. None of the compounds reduced PR-B levels (Figure 5). Absent or low PR expression is associated with more advanced endometrial carcinoma, due to losing the inhibitory actions of progesterone mediated by the receptor.21,22 Thus, natural products that activate the progesterone receptor without triggering proteolysis of the receptor may be beneficial in women at risk of developing endometrial cancer, as they are less likely to drive therapy resistance.
Figure 5.

Western blots for Ishikawa PR-B cells treated with 0, 25, or 50 μM of compounds 1–4 for 24 hours. Compounds 1–4 did not trigger degradation of the progesterone receptor B (PR-B) in Ishikawa PR-B cells.
Irilone (1) was previously reported to reduce proliferation of Ishikawa cells,23 consistent with a progestogenic, not estrogenic, effect in that cell line.24 Therefore, the ability of 1 to act synergistically at the functional level with progesterone was tested in the ovarian cancer cell line, PEO1. PEO1 cells express endogenous steroid receptors and a recent meta-analysis demonstrated that presence of the PR was associated with longer survival in ovarian cancer patients.25,26 Individually, neither progesterone (0.001–10 μM) nor 1 (1–50 μM) affected the proliferation of PEO1 cells (Figure 6A-B). However, in the presence of 100 nM of progesterone, both 25 and 50 μM 1 significantly reduced proliferation of PEO1 cells (p<0.05; Figure 6C). These data agree with a recent analysis showing an inverse relationship between flavonoid intake and epithelial ovarian cancer risk. However, the effects of individual flavonoids on ovarian cancer risk need to be further characterized, given the unique actions of 1 compared to the other flavonoids analyzed in the current study.27
Figure 6.

Irilone (1) and progesterone synergistically reduce proliferation of PEO1 cells. A-B) Proliferation of PEO1 cells treated with dose responses of progesterone (A) or irilone (B). C) Proliferation of PEO1 cells in response to increasing concentrations of 1 in the presence of 100 nM of progesterone. Significantly different from 0 μM, *p<0.05, **p<0.01, n≥3.
Irilone (1) is only 3.2% of the red clover extract by weight, making it much less abundant than formononetin (3) or biochanin A (4) (each approximately 14% by weight).9 Despite this, after ingestion of a hydrolyzed red clover supplement, irilone was measured to be the second most abundant isoflavone in serum, with only daidzein reaching higher concentrations (209±37 and 297±122 nM, respectively).28 This is likely due to irilone being resistant to metabolism by the microbiota in the gastrointestinal tract (GI) tract,29 and suggests that the biological effects of 1 may predominate over those of other isoflavones after oral ingestion of red clover extracts.
Progesterone signaling is thought to be critically important in ovarian, endometrial, and breast cancers.26,30 The current work showed a synergistic interaction between progesterone and 1 in both endometrial and ovarian cancer cell lines. In these cancers, progesterone action is generally associated with positive outcomes and reduced risk of disease occurance.21,26 However, in breast cancer progesterone signaling is generally associated with negative outcomes. This justifies future efforts to study the actions of irilone in breast cancer.31
We have identified four phytoprogestins present in a standardized red clover extract. Biochanin A (4) and formononetin (3) exhibited mixed antagonist activity in Ishikawa PR-B cells, increasing PRE/Luc activity at low concentrations and suppressing PRE/Luc activity at high concentrations. Prunetin (2) suppressed PRE/Luc activity in the same cells at high concentration but did not significantly increase PRE/Luc activity under any conditions. In contrast, irilone (1) synergistically increased PRE/Luc activity in the presence of progesterone in Ishikawa cells, and suppressed PEO1 cell growth. Collectively these results suggest that the effects of red clover extract repeatedly seen in cultured cells, and the inverse correlation between risk of various cancers and flavonoid intake may be due, in part, to altered progesterone signaling.27,32,33 Future work is needed to further elucidate the effects of each of these flavonoids in progesterone-responsive tissues so that women can make more informed decisions in regard to the safe use of supplements containing red clover.
EXPERIMENTAL SECTION
General Experimental Procedures.
NMR spectra were obtained on a Bruker 800 MHz AVANCE NMR spectrometer equipped with a 5 mm TXI room temperature inverse probe with xyz-axis pfg and Topspin version 1.3 operating software, and a Bruker 900 (226.2) MHz AVANCE NMR spectrometer equipped with a 5 mm TCI cryogenic inverse probe with z-axis pfg and TopSpin version 2.1 operating software, at the University of Illinois at Chicago Center for Structural Biology. Chemical shifts (δ) are given in ppm and coupling constants (J) are reported in Hz. 1H and 13C NMR chemical shifts were referenced to the DMSO-d6 (δH 2.50 ppm and δC 39.5 ppm) and acetone-d6 solvent resonances (δH 2.05 ppm and δC 29.8 ppm). High resolution mass spectra were obtained on a Shimadzu HRESIITTOF mass spectrometer at the University of Illinois at Chicago Research Resources Center (UIC RRC). HPLC was performed on a Shimadzu UFLC LC-20AB equipped with a UV detector (Shimadzu SPD-20A). Detection at 260 nm occurred at room temperature.
Plant Material.
100 mg of the phase II red clover extract, which contained 30% isoflavones, was obtained from the UIC Botanical Center as previously described.9
Extraction and Isolation.
The extract was dissolved into methanol and was fractionated using RP-C18 semi-preparative HPLC (2 mL/min−1, gradient of MeOH-H2O with 0.1% formic acid (FA) from 6:4 to 9:1 over 20 min, followed by an isocratic flow of 100% MeOH with 0.1% FA for 10 min), to afford fractions RE-1 to RE-19. RE-14, −15, −17 and −18 exhibited biological activity (testing concentration 60 μg/mL) and RE-17 and −18 were identified as prunetin (2) and biochanin A (4), respectively. RE-14 and −15 were further purified using RP-phenyl-hexyl analytical HPLC (1 mL/min−1, gradient of MeOH-H2O with 0.1% FA from 6:4 to 9:1 over 30 min, followed by an isocratic flow of 100% MeOH with FA for 10 min) to afford irilone (1), and formononetin (3). These isolates afforded physical and spectroscopic data consistent with literature values.10–12
Retention Time Comparison of Compounds 2–4 with Authentic Samples by HPLC Analysis.
Compound 2 and an authentic sample of prunetin were each dissolved in methanol at 0.05 mg/mL. Compound 3, 4, and corresponding authentic samples, formononetin and biochanin A, were each prepared in similar solutions at 0.5 mg/mL. Each sample was analyzed by RP-C18 analytical HPLC (gradient of MeOH-H2O from 7:3 to 100% MeOH over 30 min, followed by an isocratic flow of 100% MeOH for 10 min). Peaks corresponding to compounds 2–4 were detected at 23.0, 21.1, and 23.5 min. Retention times for authentic samples were 23.0 (prunetin), 21.1 (formononetin), and 23.5 min (biochanin A), respectively. Co-injections of compound 2 with prunetin, compound 3 with formononetin, and compound 4 with biochanin A gave single peaks at 22.9, 21.1, and 23.5 min, respectively (Supplementary Figure S1–S3).
Authenticated Compounds.
Prunetin (2), formononetin (3) (F-103 and D-118, respectively; Indofine Chemical Company, Hillsborough, NJ), biochanin A (4) (D2016–100MG, Sigma Aldrich, St. Louis, MO), and RU486 (10006317, Cayman Chemical Ann Arbor, MI) were purchased from commercial sources. Irilone (1) was isolated from the standardized red clover extract using HPLC.
Cell Culture.
Ishikawa34 cells stably expressing PR-B under the control of CMV were graciously donated by Dr. Leen J. Blok, Department of Obstetrics and Gynecology, Erasmus Medical Center, Rotterdam, Netherlands. Ishikawa cells were maintained in steroid free media consisting of phenol red free DMEM/F12, 5% charcoal-stripped FBS, and selection antibiotic (500 μg/mL G418 and 250 μg/mL hygromycin) and were validated by STR analysis in October 2017. PEO1 cells, a gift from Kenneth Nephew at Indiana University, were maintained in RPMI (11875–085, ThermoFisher, Grand Island, NY) supplemented with 2 mM glutamine, 2 mM sodium pyruvate, 10% FBS, and 1% penicillin streptomycin. All cells were maintained in a humidified incubator at 37°C and 5% CO2 and passed at confluence.
Progesterone Response Element/Luciferase (PRE/Luc) Activity Assay.
Transcriptional activity was measured using a PRE/Luc assay as previously described.8 Cells (40,000 cells per well) were plated in 24 well plates in steroid-free media made without phenol red and using charcoal stripped FBS, as previously described.35 The next day, cells were transfected with 200 ng/well of a PRE/Luc construct and 100 ng/well of a construct containing β-galactosidase under the control of CMV. Transfections were carried out with LT1 transfection reagent per the manufacturer’s instructions (MIR 2304, Mirus Bio, Madison, WI). Cells were treated with crude fractions at 60 μg/mL or purified compounds at indicated concentrations. Twenty-four hours later, lysis buffer was added to each well and cells were frozen at −80 °C. Luciferase activity was measured on a synergy BioTek plate reader. The first six determinations of relative luminescence units (RLU) were averaged for each well. Luciferase activity was normalized to β-galactosidase and then normalized to control for each experiment.
Western Blots.
Ishikawa PR-B cells, in steroid free media, were treated with indicated concentrations of purified compounds for 24 hours. Cells were lysed in RIPA buffer containing protease and phosphatase inhibitors and the protein concentration was determined via BCA. Twenty-five micrograms of protein was separated on SDS-PAGE cells by electrophoresis and transferred to nitrocellulose membranes.36 Membranes were blocked for one hour in 5% milk and probed overnight for progesterone receptor (diluted 1:500, H-190, Santa Cruz Biotechnology) or β actin (diluted 1:1000, A2066, Sigma Aldrich, St. Louis MO). The next day, membranes were washed with TBS-T, incubated with anti-rabbit secondary antibody and developed with SuperSignal West Femto Substrate (34095, Thermo Scientific, Rockford IL). Images were captured with a FluorChem C (Protein Simple, San Jose, CA).
Proliferation Assay.
Proliferation was measured using the sulforhodamine B (SRB) assay.36 Briefly, 1,000 cells per well were seeded onto two 96-well plates in steroid free media. Two hours later, one plate was collected by discarding the cell media, adding 100 μL 20% trichloroacetic acid, and at storing the plate at 4°C. The other plate was treated with pure compounds at indicated concentrations. Five days later, the second plate was collected by discarding the cell culture media, adding 100 μL 20% trichloroacetic acid, and placing the plate at 4°C. The next day both plates were washed with tap water and the cells were stained with 0.4% SRB in 1% acetic acid. Plates were then washed four times with 1% acetic acid, dried, and SRB was dissolved in 10 mM TrisOH. Absorbance was measured at 505 nm. Absorbance on day 5 was normalized to day 0.
Statistical Analysis.
All data is presented as mean±SEM with n≥3. Due to the large number of samples, the screening of fractions produced from red clover was not statistically analyzed, and fractions with the largest effects were chosen for further evaluation. All other data were analyzed by ANOVA followed by a Tukey’s or Dunnett’s posthoc test. p<0.05 was considered significant. Analysis was performed using Prism version 7.0a.
Supplementary Material
ACKNOWLEDGEMENTS
This works was supported by grant R01 AT008824 to J.E.B. and B.T.M. and a fellowship supporting M.D. (T32 AT007533) both from the National Center for Complementary and Integrative Health (NCCIH). Finally, we would like to thank the UIC Botanical Center for supplying a standardized T. pratense extract.
Footnotes
The authors declare no competing financial interest.
ASSOCIATED CONTENT
Supporting Information. Additional mass spectra, HPLC chromatograms, and other materials are found in the Supporting Information. This material is available free of charge via the Internet at http://pubs.acs.org.
References
- (1).Hajirahimkhan A; Dietz BM; Bolton JL Planta Med. 2013, 79, 538–553. [DOI] [PMC free article] [PubMed] [Google Scholar]
- (2).Patisaul HB; Jefferson W Front. Neuroendocrinol 2010, 31, 400–419. [DOI] [PMC free article] [PubMed] [Google Scholar]
- (3).Dean M; Murphy BT; Burdette JE Mol. Cell. Endocrinol 2017, 442, 98–105. [DOI] [PMC free article] [PubMed] [Google Scholar]
- (4).Hale GE; Hughes CL; Robboy SJ; Agarwal SK; Bievre M Menopause 2001, 8, 338–346. [DOI] [PubMed] [Google Scholar]
- (5).Overk CR; Guo J; Chadwick LR; Lantvit DD; Minassi A; Appendino G; Chen S-N; Lankin DC; Farnsworth NR; Pauli GF; van Breemen RB; Bolton JL Chem. Biol. Interact 2008, 176, 30–39. [DOI] [PMC free article] [PubMed] [Google Scholar]
- (6).Gielen SCJP; Hanekamp EE; Hanifi-Moghaddam P; Sijbers AM; van Gool AJ; Burger CW; Blok LJ; Huikeshoven FJ Int. J. Gynecol. Cancer 2006, 16, 110–120. [DOI] [PubMed] [Google Scholar]
- (7).Toh MF; Mendonca E; Eddie SL; Endsley MP; Lantvit DD; Petukhov PA; Burdette JE J. Steroids Horm. Sci 2014, 5, 136. [DOI] [PMC free article] [PubMed] [Google Scholar]
- (8).Toh MF; Sohn J; Chen SN; Yao P; Bolton JL; Burdette JE Steroids 2012, 77, 765–773. [DOI] [PMC free article] [PubMed] [Google Scholar]
- (9).Booth NL; Overk CR; Yao P; Burdette JE; Nikolic D; Chen S-N; Bolton JL; van Breemen RB; Pauli GF; Farnsworth NR J. Altern. Complement. Med. 2006, 12, 133–139. [DOI] [PMC free article] [PubMed] [Google Scholar]
- (10).Ibrahim SRM; Mohamed GA; Al-Musayeib NM Molecules 2012, 17, 2587–2598. [DOI] [PMC free article] [PubMed] [Google Scholar]
- (11).dos Santos SA; de Carvalho MG J. Braz. Chem. Soc 1995, 6, 349–352. [Google Scholar]
- (12).Kinjo J-E; Furusawa J-I; Baba J; Takeshita T; Yamasaki M; Nohara T Chem. Pharm. Bull 1987, 35, 4846–4850. [Google Scholar]
- (13).Kuiper GG; Lemmen JG; Carlsson B; Corton JC; Safe SH; van der Saag PT; van der Burg B; Gustafsson J-Å Endocrinology 1998, 139, 4252–4263. [DOI] [PubMed] [Google Scholar]
- (14).Roll DM; Barbieri LR; Bigelis R; McDonald LA; Arias DA; Chang L-P; Singh MP; Luckman SW; Berrodin TJ; Yudt MR J. Nat. Prod 2009, 72, 1944–1948. [DOI] [PubMed] [Google Scholar]
- (15).Burris TP; Montrose C; Houck KA; Osborne HE; Bocchinfuso WP; Yaden BC; Cheng CC; Zink RW; Barr RJ; Hepler CD; Krishnan V; Bullock HA; Burris LL; Galvin RJ; Bramlett K; Stayrook KR Mol. Pharmacol 2005, 67, 948–954. [DOI] [PubMed] [Google Scholar]
- (16).Ong VY; Tan BK BMC Complement. Altern. Med 2007, 7, 3. [DOI] [PMC free article] [PubMed] [Google Scholar]
- (17).Li X; O’Malley BW J. Biol. Chem 2003, 278, 39261–39264. [DOI] [PubMed] [Google Scholar]
- (18).Hagan CR; Daniel AR; Dressing GE; Lange CA Mol. Cell. Endocrinol 2012, 357, 43–49. [DOI] [PMC free article] [PubMed] [Google Scholar]
- (19).Hashemi M; Behrangi N; Borna H; Entezari MJ Med. Plants Res 2012, 6, 4375–4378. [Google Scholar]
- (20).Lange CA; Shen T; Horwitz KB PNAS 2000, 97, 1032–1037. [DOI] [PMC free article] [PubMed] [Google Scholar]
- (21).Zhang Y; Zhao D; Gong C; Zhang F; He J; Zhang W; Zhao Y; Sun J World J. Surg. Oncol 2015, 13, 208. [DOI] [PMC free article] [PubMed] [Google Scholar]
- (22).Hanekamp EE; Gielen SCJP; Smid-Koopman E; Kühne LCM; de Ruiter PE; Chadha-Ajwani S; Brinkmann AO; Grootegoed JA; Burger CW; Huikeshoven FJ; Blok LJ Clin. Cancer Res 2003, 9, 4190–4199. [PubMed] [Google Scholar]
- (23).Lutter S; Schmalbach K; Esch HL; Lehmann L Arch. Toxicol 2014, 88, 309–321. [DOI] [PubMed] [Google Scholar]
- (24).Croxtall JD; Elder MG; White JO J. Steroid Biochem 1990, 35, 665–669. [DOI] [PubMed] [Google Scholar]
- (25).O’Donnell AJM; Macleod KG; Burns DJ; Smyth JF; Langdon SP Endocr. Relat. Cancer 2005, 12, 851–866. [DOI] [PubMed] [Google Scholar]
- (26).Luo H; Li S; Zhao M; Sheng B; Zhu H; Zhu X Oncotarget 2017, 8, 36845–36856. [DOI] [PMC free article] [PubMed] [Google Scholar]
- (27).Cassidy A; Huang T; Rice MS; Rimm EB; Tworoger SS Am. J. Clin. Nutr 2014, 100, 1344–1351. [DOI] [PMC free article] [PubMed] [Google Scholar]
- (28).Maul R; Kulling SE Br. J. Nutr 2010, 103, 1569–1572. [DOI] [PubMed] [Google Scholar]
- (29).Braune A; Maul R; Schebb NH; Kulling SE; Blaut M Mol. Nutr. Food Res 2010, 54, 929–938. [DOI] [PubMed] [Google Scholar]
- (30).Kim JJ; Kurita T; Bulun SE Endocr. Rev 2013, 34, 130–162. [DOI] [PMC free article] [PubMed] [Google Scholar]
- (31).Brisken C Nat. Rev. Cancer 2013, 13, 385–396. [DOI] [PubMed] [Google Scholar]
- (32).Peterson J; Lagiou P; Samoli E; Lagiou A; Katsouyanni K; La Vecchia C; Dwyer J; Trichopoulos D Br. J. Cancer 2003, 89, 1255–1259. [DOI] [PMC free article] [PubMed] [Google Scholar]
- (33).Reiter E; Gerster P; Jungbauer A Gynecol. Endocrinol 2011, 27, 1037–1042. [DOI] [PubMed] [Google Scholar]
- (34).Smid-Koopman E; Kuhne LCM; Hanekamp EE; Gielen SCJP; De Ruiter PE; Grootegoed JA; Helmerhorst TJM; Burger CW; Brinkmann AO; Huikeshoven FJ; Blok LJ J. Soc. Gynecol. Investig 2005, 12, 285–292. [DOI] [PubMed] [Google Scholar]
- (35).Moyle-Heyrman G; Schipma MJ; Dean M; Davis DA; Burdette JE J. Ovarian Res. 2016, 9, 5. [DOI] [PMC free article] [PubMed] [Google Scholar]
- (36).Dean M; Davis DA; Burdette JE Cancer Lett. 2017, 391, 114–124. [DOI] [PMC free article] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.