

Abstract
Background
From 2012 to 2013, an outbreak of cutaneous leishmaniasis by Leishmania braziliensis was detected in indigenous villages located in a remote rural area of Pernambuco state, north-eastern Brazil. Considering that the principal activities of this indigenous community are farming and crop plantation, and also that the outbreak involved many children, we investigated the presence of sand fly vectors inside human houses and also the exposure of dogs to leishmanial parasites. Our general objective was to gather epidemiological data that could indicate the occurrence of a peri-domestic/domestic transmission cycle of L. braziliensis in these indigenous villages.
Methods
From March 2015 to March 2016, sand flies were collected using light traps in the indoor and immediate outdoor environments in the three indigenous villages that reported the most cutaneous leishmaniasis cases during the 2012–2013 outbreak. Moreover, samples obtained from 300 dogs living in the outbreak villages and two nearby villages were tested by a rapid immunochromatographic test and by a real-time PCR for detecting anti-Leishmania antibodies and Leishmania DNA, respectively.
Results
In total, 5640 sand flies belonging to 11 species were identified. Males (n = 3540) predominated over females (n = 2100). Migonemyia migonei (84.3%) was the most abundant species, followed by Evandromyia lenti (5.5%), Lutzomyia longipalpis (4.1%), Nyssomyia intermedia (1.6%) and Micropygomyia capixaba (1.4%), representing together ~97% of the sand flies collected. Nine out of the 11 species identified in this study were found indoors, including M. migonei, L. longipalpis and N. intermedia, which are proven vectors of Leishmania spp. Out of 300 dogs tested, 26 (8.7%) presented anti-Leishmania antibodies and six (2%) were Leishmania DNA-positive. The level of exposure in dogs living in the indigenous villages where the 2012–2013 outbreak of human CL was detected was almost 2-fold higher than in the two nearby villages (11.0 vs 6.2% for serology and 2.6 vs 1.4% for real-time PCR).
Conclusions
The results suggest that different sand fly vectors may be adapted to human dwellings, thus increasing the risk of transmission in the indoor and immediate outdoor environments. The adaptation of sand flies to the indoor environment in the studied indigenous villages may be partly explained by the poor housing conditions and the proximity of the houses to crop plantations and forest fragments.
Electronic supplementary material
The online version of this article (10.1186/s13071-019-3383-1) contains supplementary material, which is available to authorized users.
Keywords: Cutaneous leishmaniasis, Dogs, Phlebotomine sand fly, Rural area
Background
Leishmaniases are neglected tropical diseases, which cause significant morbidity and mortality in endemic areas, particularly in tropical and subtropical regions of the world [1]. Brazil, India, Bangladesh, Sudan, South Sudan and Ethiopia account for more than 90% of the global cases of visceral leishmaniasis (VL), with an estimated 200,000 to 400,000 new cases per year [2]. About 75% of the global incidence of cutaneous leishmaniasis (CL) occurs in Afghanistan, Algeria, Colombia, Brazil, Iran, Syria, Ethiopia, Sudan, Costa Rica and Peru, with an estimated 0.7 to 1.2 million new cases per year [2]. From a global perspective, Brazil is one of the main foci of leishmaniases, with an annual average incidence of 1.7 and 8.0 new VL and CL cases per 100,000 population, respectively, during the period 2013–2017 [3].
CL and VL are primarily zoonoses, with wild animals (e.g. forest rodents) and domestic dogs, respectively, being involved as reservoirs in their zoonotic transmission cycles [4, 5]. Outbreaks of zoonotic CL in Brazil are commonly detected among males at working age who enter the forest for various reasons, such as for military training [6]. Deforestation (e.g. for road construction and crop plantation) and population movements from non-endemic to endemic areas (and vice versa) are also risk factors for both CL and VL [7]. In recent years, CL and VL have spread across different Brazilian regions, indicating the ineffectiveness of control measures to reduce the burden of the disease in both rural and urban areas [8, 9].
Degradation of natural habitats may force sand fly vectors to adapt to the modified environment [10]. Indeed, blood-feeding insects such as sand flies are commonly attracted to human dwellings, where they may find food sources (e.g. domestic animals and humans) [11], resting places and breeding sites [12]. Certainly, the adaptation of sand fly vectors to human dwellings may increase the risk of Leishmania spp. transmission in the peri-domestic and domestic environments.
The increasing interaction between humans, domestic and wild animals, in wild, peri-domestic and domestic environments has caused profound changes in the epidemiology of leishmaniases in the past decades [7]. For instance, CL caused by Leishmania braziliensis is a zoonosis maintained by multitude of small mammals (e.g. forest rodents and marsupials) and sand fly vectors, which have adapted to the peri-domestic and domestic environments. In endemic areas, dogs are frequently exposed to sand fly vectors and are often infected by L. braziliensis [13]. While dogs play no role as reservoirs of this parasite [14], they can play a useful role as a sentinel host [15].
From 2012 to 2013, an outbreak of CL was detected in indigenous villages located in a remote rural area of Pernambuco state, north-eastern Brazil. These villages are settled in legally-expropriated lands and are presently home to the tribe Xukuru de Ororubá. While many ancient indigenous traditions are maintained by the Xukuru people, their principal working activities are farming and crop plantation for their own subsistence. Bearing this in mind and also considering that the outbreak detected in 2012–2013 involved many children, we investigated the presence of sand fly vectors inside human houses and also the exposure of dogs to leishmanial parasites. Our general objective was to gather epidemiological data that could indicate the occurrence of a peri-domestic/domestic transmission cycle of L. braziliensis in these indigenous villages.
Methods
Study area
This study was carried out in the municipality of Pesqueira, which is located in the agreste region of Pernambuco, a narrow zone between the Atlantic forest zone (zona da mata) and the semiarid region (sertão). In particular, three indigenous villages were surveyed in this study: Guarda (V1; 8°21′49.6″S, 36°48′47.1″W, altitude: 844 m above sea level), Santana (V2; 8°20′12.3″S, 36°43′38.1″W, altitude: 850 m) and Afetos (V3; 8°19′06.1″S, 36°42′37.3″W, altitude: 965 m) (Fig. 1). Geographical coordinates and altitude of each village were recorded using a Garmin eTrex Venture HC GPS (Garmin International, Olathe, USA).
Fig. 1.

Location of the three indigenous villages (V1, Guarda; V2, Santana; and V3, Afetos) studied herein. Pesqueira, Pernambuco, Brazil. Copyright: Creative Commons Attribution-ShareAlike 3.0 Licence (https://creativecommons.org/licenses/by-sa/3.0/). The map was created using QGIS and publicly available shapefiles from QGIS web site [55]
These villages are part of the 24 indigenous villages of the tribe Xukuru de Ororubá, which occupy approximately 27,000 hectares of a chain of mountains named Serra do Ororubá. The villages are located in a rural area and the native vegetation is represented by semi-decidual and deciduous forests, although most of the original forest coverage has been substituted by crop plantations [16]. The landscape is represented by a xeric shrubland and thorn forest, which consists primarily of small, thorny trees that shed their leaves seasonally. The ground layer is made up of cacti, thick-stemmed plants, thorny brush and arid-adapted grasses. The climate of this area is semiarid, characterized by low humidity and little rainfall. The raining period ranges from February to July, with an annual average temperature of 26 °C (range, 24–27 °C), average relative humidity of 76% (range, 69–86%) and average precipitation of 700 mm3.
The population of the villages is currently around 2720 [17]. The local economy is mostly based on agriculture, with plantations of bananas, beans, cassava, corn and vegetables, as well as dairy cattle and goat farming [18]. Many of the houses are precarious and lack basic sanitation. Children and teenagers typically go to school in the morning and afternoon, respectively. In their spare time they play and are at home early in the evening, particularly young children. Domestic animals (e.g. dogs, cats and chickens) are common both indoors and outdoors of human houses.
Collection and morphological identification of sand flies
Sand fly collections were carried out monthly, from March 2015 to March 2016 (except in October 2015, for logistic reasons) for two to three consecutive nights. Collection sites (houses) were chosen based on the occurrence of human cases of CL. Some of the houses were made of mud walls and thatched roofs, with obvious openings that may facilitate the entrance of insects during the night. Moreover, some of the houses were surrounded by native vegetation.
Each night, one to four CDC light traps were installed in each village, operating from 18:00 h to 6:00 h, for a total of 253 traps installed and 3036 cumulative hours of trapping. Each trap was positioned 1.5 m above the ground in two types of environments: indoor (living rooms and bedrooms) and outdoor (backyards with chicken coop, goats and/or dogs). All specimens collected were transferred to labelled vials containing 70% ethanol and subsequently identified using morphological keys for American sand flies based on characters of male genitalia, female spermathecae and pharyngeal armature [19]. The nomenclature of sand fly species followed Galati’s proposal [20]. Females collected indoors and outdoors during the last seven months of collection (from August 2015 to March 2016) were classified as engorged (blood in the abdomen, total or partial) or unfed (no visible blood in the abdomen).
Meteorological data
Monthly average temperature (°C), relative humidity (%) and rainfall (mm) data were obtained from station 82900 of the Technology Institute of Pernambuco (ITEP). The saturation deficit (SD) was calculated as follows: SD = (1 – RH/100) × 4.9463 × e0.0621 × T, where RH is relative humidity and T is temperature [6].
Canine blood collection and diagnostic procedures
Dogs living in the investigated indigenous villages (Guarda, Santana and Afetos) and two nearby villages (Cimbres and São José) were chosen as sentinel hosts. From March to June 2015, blood samples were collected (~5 ml) from 300 privately-owned dogs. Aliquots of ~2 ml and ~3 ml were added to EDTA tubes (Greiner Bio-One GmbH, Kremsmünster, Austria) and gel serum separator tubes (Greiner Bio-One GmbH), respectively. Gel serum separator tubes were centrifuged at 2000×g for 10 min for serum separation. The obtained sera and blood samples were stored at -20 °C.
Genomic DNA extraction from EDTA-blood samples was performed using the PureLink® Genomic DNA Mini Kit (Invitrogen, Carlsbad, CA, USA), according to the manufacturer’s instructions. The quantity and purity of the extracted DNA were assessed using a NanoDrop 2000c Spectrophotometer (Thermo Fisher Scientific, Waltham, MA, USA). Extracted DNA samples were stored at -20 °C until testing.
Real-time PCR reactions were performed using the primers LEISH-1 (5′-AAC TTT TCT GGT CCT CCG GGT AG-3′) and LEISH-2 (5′-ACC CCC AGT TTC CCG CC-3′) and the TaqMan® probe FAM-5′-AAA AAT GGG TGC AGA AAT-3′-non-fluorescent quencher-MGB, as described elsewhere [21]. The final reaction volume was adjusted to 25 μl containing 5.5 μl of type 1 (ultrapure) water, 2.25 μl of each primer (900 nM), 0.5 μl of probe (200 nM), 12.5 μl of TaqMan® Universal PCR Master Mix (Applied Biosystems, Carlsbad, CA, USA) and 2 μl of DNA template. PCR cycling conditions were 50 °C for 2 min, 95 °C for 10 min, then 40 cycles at 95 °C for 15 s and at 60 °C for 1 min. All samples were tested in duplicate and no template control (NTC) and positive controls (DNA extracted from cultured L. infantum promastigotes) were included in each PCR run. Reactions were run on a 7500 Real Time PCR System (Applied Biosystems, Foster City, CA, USA) and the results analysed using the 7500 software v.2.0.5.
Dog sera were tested using DPP LVC (Bio-Manguinhos, Rio de Janeiro, RJ, Brazil), according to the manufacturer’s instructions. This test uses recombinant antigens and it is the official screening test used by public health authorities in Brazil [22]. Results were read after 10 min and interpreted as follows: negative (only control red line present), positive (control and sample red lines present) and invalid (control red line absent).
Human cases
Secondary data regarding human cases of CL detected during the 2007–2017 in the indigenous villages were obtained from the Brazilian Information System on Diseases of Compulsory Declaration [3]. In particular, we were interested in all CL cases detected in the outbreak of 2012–2013. Variables of interest were year and month of notification, gender, place of residence, and age. Data were obtained and processed anonymously.
Diversity indices and statistical analyses
Sand fly species richness and diversity were assessed using the following parameters: species richness (S), number of individuals (n), Shannon’s diversity index (H’) and Pielou’s equitability index (J’). We also computed the species accumulation curve (sample-based rarefaction) as a function of number of samples using Maoʼs tau, with standard deviation; in the graphical plot, the standard errors were converted to 95% confidence intervals. Diversity indices and species accumulation curve were calculated using PAST, v.3.23 for Mac OS [23].
Before statistical analysis, normality of data was assessed using Lilliefors. Pearson’s (r) or Spearman’s (rs) correlation coefficients were used to determine the correlation between the meteorological variables and the relative frequencies of sand flies (i.e. number of individuals per hour of trapping). Student’s t-test was used for comparing the relative frequencies of sand flies collected monthly indoors vs outdoors. To compare the abundance of each species indoors vs outdoors, we calculated the index of species abundance (ISA), which was then converted to a scale of zero to one, through the standardized index of species abundance (SISA), where the value 1.00 represents the most abundant species [24]. The Kruskal-Wallis H-test (with Dunn’s post-hoc test) was used to compare the relative frequencies of sand flies collected monthly in the three surveyed indigenous villages. Chi-square test was used to assess whether positivity to Leishmania spp. varied according to dog data including sex (male, female), age (≤ 1 year, > 1 year), clinical status (healthy, sick) and housing condition (domiciled, semi-domiciled). We also used the Chi-square test to determine whether there was a significant difference between the frequencies of engorged and unfed females collected indoors vs outdoors. Statistical analyses were performed using BioEstat v.5.3 (Mamirauá Institute of Sustainable Development, Tefé, AM, Brazil) and P ≤ 0.05 was considered statistically significant.
Results
Sand fly species and numbers
A total of 5640 sand flies were collected and morphologically identified (Table 1). The species accumulation curve reached saturation at the 10th sampling event (Fig. 2), with 11 species identified.
Table 1.
Number (n) and percentage (%) of sand flies collected indoors and outdoors in the surveyed indigenous villages (V1–V3) in Pesqueira, Pernambuco, Brazil, from March 2015 to March 2016, according to species and sex. Sex ratio (female:male) is also provided
| Species | Indoor | Outdoor | ||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Female | Male | Total | Sex ratio | Female | Male | Total | Sex ratio | |||||||
| n | % | n | % | n | % | n | % | n | % | n | % | |||
| Micropygomyia capixaba | 46 | 59.0 | 3 | 3.8 | 49 | 62.8 | 15.3 | 24 | 30.8 | 5 | 6.4 | 29 | 37.2 | 4.8 |
| Evandromyia evandroi | 5 | 7.8 | 1 | 1.6 | 6 | 9.4 | 5.0 | 53 | 82.8 | 5 | 7.8 | 58 | 90.6 | 10.6 |
| Micropygomyia villelai | 4 | 11.1 | 10 | 27.8 | 14 | 38.9 | 0.4 | 16 | 44.4 | 6 | 16.7 | 22 | 61.1 | 2.7 |
| Nyssomyia intermedia a | 3 | 3.4 | 12 | 13.5 | 15 | 16.9 | 0.3 | 34 | 38.2 | 40 | 44.9 | 74 | 83.1 | 0.9 |
| Evandromyia lenti | 18 | 5.8 | 30 | 9.7 | 48 | 15.6 | 0.6 | 72 | 23.4 | 188 | 61.0 | 260 | 84.4 | 0.4 |
| Lutzomyia longipalpis a | 25 | 10.9 | 46 | 20.0 | 71 | 30.9 | 0.5 | 50 | 21.7 | 109 | 47.4 | 159 | 69.1 | 0.5 |
| Migonemyia migonei a | 438 | 9.2 | 620 | 13.0 | 1058 | 22.3 | 0.7 | 1272 | 26.8 | 2423 | 51.0 | 3695 | 77.7 | 0.5 |
| Evandromyia sallesi | 0 | 0.0 | 0 | 0.0 | 0 | 0.0 | nc | 2 | 66.7 | 1 | 33.3 | 3 | 100.0 | 2.0 |
| Micropygomyia schreiberi | 0 | 0.0 | 0 | 0.0 | 0 | 0.0 | nc | 3 | 100.0 | 0 | 0.0 | 3 | 100.0 | nc |
| Sciopemyia sordellii | 0 | 0.0 | 7 | 24.1 | 7 | 24.1 | 0.0 | 9 | 31.0 | 13 | 44.8 | 22 | 75.9 | 0.7 |
| Micropygomyia trinidadensis | 10 | 21.3 | 6 | 12.8 | 16 | 34.0 | 1.7 | 16 | 34.0 | 15 | 31.9 | 31 | 66.0 | 1.1 |
| Total | 549 | 42.8 | 735 | 57.2 | 1284 | 22.8 | 0.7 | 1551 | 35.6 | 2805 | 64.4 | 4356 | 77.2 | 0.6 |
aProven vector species [24]
Abbreviation: nc, not calculated
Fig. 2.
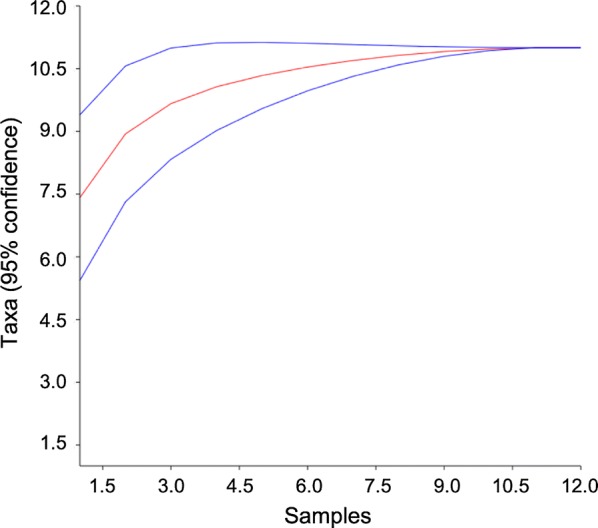
Species accumulation curve (Mao’s tau function) (red line) and 95% confidence interval (blue lines) for sand flies in Pesqueira, Pernambuco, Brazil
The percentage of females (37.2%, n = 2100) was lower than that of males (62.8%, n = 3540), with an overall female:male ratio of 0.6:1. A positive correlation between the monthly number of males and females collected during the study (r(10) = 0.86, P < 0.001) was found. However, the sex ratio varied according to sand fly species, type of environment (indoors vs outdoors) (Table 1) and village (Table 2).
Table 2.
Number (n) and percentage (%) of sand flies collected in three indigenous villages (V1–V3) in Pesqueira, Pernambuco, Brazil, from March 2015 to March 2016, according to species and sex. Sex ratio (female:male) is also provided
| Species | Guarda (V1) | Santana (V2) | Afetos (V3) | ||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Female | Male | Total | Sex ratio | Female | Male | Total | Sex ratio | Female | Male | Total | Sex ratio | ||||||||||
| n | % | n | % | n | % | n | % | n | % | n | % | n | % | n | % | n | % | ||||
| Micropygomyia capixaba | 31 | 4.5 | 7 | 0.7 | 38 | 2.2 | 4.4 | 3 | 1.6 | 0 | 0.0 | 3 | 0.8 | nc | 36 | 3.0 | 1 | 0.0 | 37 | 1.0 | 36.0 |
| Evandromyia evandroi | 32 | 4.6 | 2 | 0.2 | 34 | 2.0 | 16.0 | 9 | 4.8 | 1 | 0.5 | 10 | 2.5 | 9.0 | 17 | 1.4 | 3 | 0.1 | 20 | 0.6 | 5.7 |
| Micropygomyia villelai | 4 | 0.6 | 11 | 1.1 | 15 | 0.9 | 0.4 | 3 | 1.6 | 0 | 0.0 | 3 | 0.8 | nc | 13 | 1.1 | 5 | 0.2 | 18 | 0.5 | 2.6 |
| Nyssomyia intermedia a | 10 | 1.4 | 14 | 1.4 | 24 | 1.4 | 0.7 | 1 | 0.5 | 2 | 1.0 | 3 | 0.8 | 0.5 | 26 | 2.1 | 36 | 1.5 | 62 | 1.7 | 0.7 |
| Evandromyia lenti | 50 | 7.2 | 125 | 12.5 | 175 | 10.3 | 0.4 | 12 | 6.4 | 28 | 13.6 | 40 | 10.2 | 0.4 | 28 | 2.3 | 65 | 2.8 | 93 | 2.6 | 0.4 |
| Lutzomyia longipalpis a | 70 | 10.1 | 126 | 12.6 | 196 | 11.6 | 0.6 | 0 | 0.0 | 5 | 2.4 | 5 | 1.3 | 0.0 | 5 | 0.4 | 24 | 1.0 | 29 | 0.8 | 0.2 |
| Migonemyia migonei a | 483 | 69.8 | 704 | 70.5 | 1187 | 70.2 | 0.7 | 154 | 82.4 | 163 | 79.1 | 317 | 80.7 | 0.9 | 1073 | 88.0 | 2176 | 93.1 | 3249 | 91.4 | 0.5 |
| Evandromyia sallesi | 0 | 0.0 | 0 | 0.0 | 0 | 0.0 | nc | 2 | 1.1 | 1 | 0.5 | 3 | 0.8 | 2.0 | 0 | 0.0 | 0 | 0.0 | 0 | 0.0 | nc |
| Micropygomyia schreiberi | 0 | 0.0 | 0 | 0.0 | 0 | 0.0 | nc | 1 | 0.5 | 0 | 0.0 | 1 | 0.3 | nc | 2 | 0.2 | 0 | 0.0 | 2 | 0.1 | nc |
| Sciopemyia sordellii | 7 | 1.0 | 6 | 0.6 | 13 | 0.8 | 1.2 | 2 | 1.1 | 4 | 1.9 | 6 | 1.5 | 0.5 | 0 | 0.0 | 10 | 0.4 | 10 | 0.3 | 0.0 |
| Micropygomyia trinidadensis | 5 | 0.7 | 4 | 0.4 | 9 | 0.5 | 1.3 | 0 | 0.0 | 2 | 1.0 | 2 | 0.5 | 0.0 | 19 | 1.6 | 17 | 0.7 | 36 | 1.0 | 1.1 |
| Total | 692 | 40.9 | 999 | 59.1 | 1691 | 30.0 | 0.7 | 187 | 47.6 | 206 | 52.4 | 393 | 7.0 | 0.9 | 1219 | 34.3 | 2337 | 65.72 | 3556 | 63.0 | 0.5 |
aProven vector species [24]
Abbreviation: nc, not calculated
Most sand flies were collected outdoors (77.2%; n = 4356; mean = 2.36 sand flies per hour of trapping) as compared to indoors (22.8%; n = 1284; mean = 1.08 sand flies per hour of trapping) (Student’s t-test, t(22) = 3.14, P = 0.005). However, 9 out of 11 species found in this study were collected inside the houses. Some sand fly species (i.e. Evandromyia lenti, Migonemyia migonei and Lutzomyia longipalpis) were consistently collected indoors throughout the entire study period (Fig. 3, Additional file 1: Table S1). According to data recorded from August 2015 to March 2016, 28 out of 232 (12.1%) M. migonei females collected indoors and 98 out of 890 (11.0%) those collected outdoors were engorged (χ2 = 0.21, df = 1, P = 0.650). Additionally, 2 out of 32 (6.3%) Nyssomyia intermedia females collected outdoors were also engorged. All other females belonging to other species, which were collected in the aforementioned period, were unfed.
Fig. 3.

Monthly relative frequency of sand flies (number of individuals per hour of trapping) collected indoors (a and b) and outdoors (c and d) according to species (a and c Migonemyia migonei; b and d remaining species), Pesqueira, Pernambuco, Brazil
Migonemyia migonei was the most abundant species both indoors (SISA = 1.00) and outdoors (SISA = 1.00). In the outdoor environment, L. longipalpis (SISA = 0.90) was the second most abundant species, followed by E. lenti (SISA = 0.67), Evandromyia evandroi (SISA = 0.67), N. intermedia (SISA = 0.58), Sciopemyia sordellii (SISA = 0.43), and other less abundant species (Additional file 2: Figure S1). In the indoor environment, Micropygomyia capixaba (SISA = 0.81) was the second most abundant species, followed by E. lenti (SISA = 0.78), Micropygomyia trinidadensis (SISA = 0.54), L. longipalpis (SISA = 0.48), Micropygomyia villelai (SISA = 0.46), and other less abundant species (Additional file 2: Figure S1).
Among the species collected, M. migonei (84.3%), E. lenti (5.5%), L. longipalpis (4.1%), N. intermedia (1.6%) and M. capixaba (1.4%) were the most frequent, representing together ~97% of the total sand flies captured. These three species were consistently trapped during the whole study period and in the three studied villages. However, M. migonei specimens were more frequently collected in V3, whereas most E. lenti and L. longipalpis in V1. Evandromyia sallesi (n = 3) and Micropygomyia schreiberi (n = 3) were the least representative species, being found only in V2 and V3 (Table 2).
The relative frequencies of sand flies collected monthly varied according to village (Kruskal-Wallis H-test, H = 18.24, df = 2, P < 0.001; Dunn’s post-hoc test, P < 0.05 for V1 vs V2 and V2 vs V3, and P > 0.05 for V1 vs V3). The highest number of sand flies was recorded in village V3 (63.0%; n = 3556; mean = 2.77 sand flies per hour of trapping), followed by V1 (30.0%; n = 1691; mean = 1.68 sand flies per hour of trapping) and V2 (7.0%; n = 393; mean = 0.53 sand flies per hour of trapping). The highest species richness was found in V2, where all 11 species found in this study were present (Table 3). The species diversity and equitability were higher in V1 (H’ = 1.06, J’ = 0.48) and V2 (H’ = 0.81, J’ = 0.34), corresponding to the villages with lower altitude (i.e. 844 and 850 m, respectively).
Table 3.
Diversity indices in three indigenous villages in Pesqueira, Pernambuco, Brazil, from March 2015 to March 2016
| Index | Guarda (V1) | Santana (V2) | Afetos (V3) |
|---|---|---|---|
| Species richness | 9 | 11 | 10 |
| Individuals (n) | 1691 | 393 | 3556 |
| Shannon (H’) | 1.06 | 0.81 | 0.46 |
| Equitability (J’) | 0.48 | 0.34 | 0.20 |
The months with the greatest number of sand flies collected were May 2015 and March 2016 (Fig. 4), when the monthly average precipitations were 179 and 69.2 mm, monthly average temperatures 26.4 and 27.4 °C and monthly average relative humidity 77.3 and 74%, respectively (Fig. 5).
Fig. 4.
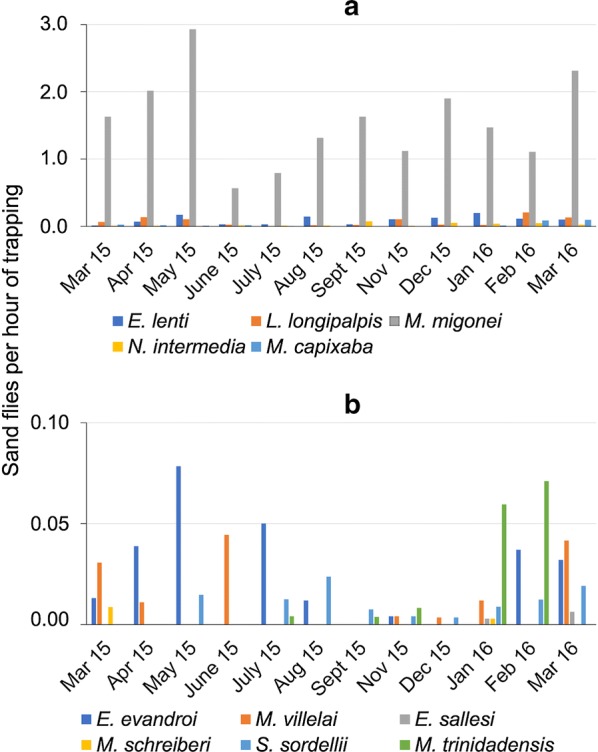
Monthly relative frequency of sand flies (number of individuals per hour of trapping) collected in Pesqueira, Pernambuco, Brazil (a the five most common species; b the remaining species)
Fig. 5.
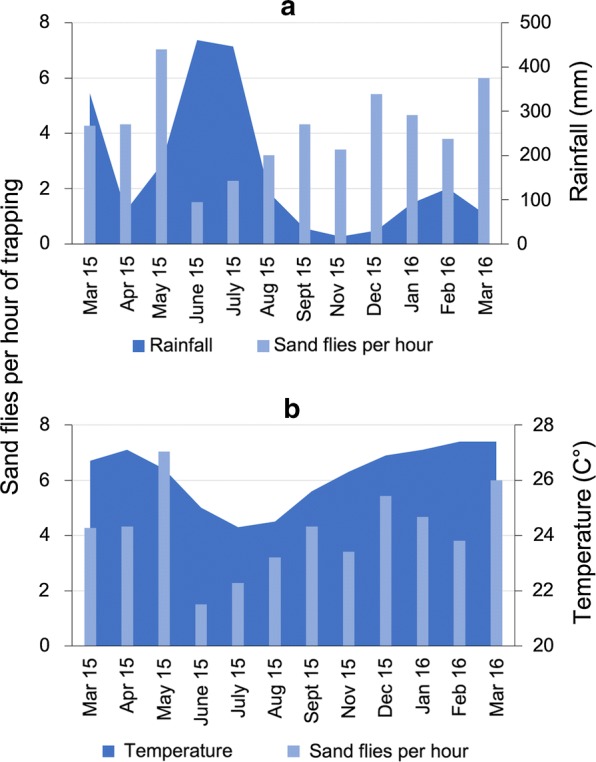
Monthly relative frequency of sand flies (number of individuals per hour of trapping) in relation to rainfall (a) and temperature (b) in Pesqueira, Pernambuco, Brazil
The months with higher temperatures and lower relative humidity coincided with peaks in sand fly population. Indeed, the relative frequency of sand flies (i.e. sand flies per hour of trapping) collected monthly was positively correlated with temperature (r(10) = 0.62, P = 0.033). On the other hand, no significant correlation was found between the relative frequency of sand flies with relative humidity (r(10) = -0.44, P = 0.152), precipitation (rs = -0.32, P = 0.308) or saturation deficit (r(10) = 0.49, P = 0.107). The most abundant species collected (i.e. M. migonei, E. lenti and L. longipalpis) were collected mainly in months with an average relative humidity < 75%.
Canine exposure to Leishmania spp
Out of 300 dogs tested, 26 (8.7%) were positive by serology and six (2%) by real-time PCR. Six dogs were simultaneously positive for both the tests. The highest positivity rates for both serology and real-time PCR were recorded in the outbreak villages (serology = 11.0%, 17/155; real-time PCR = 2.6%, 4/155), as compared with nearby villages (serology = 6.2%, 9/145; real-time PCR = 1.4%, 2/145). Most of the dogs presented apparent clinical signs (n = 179) and were semi-domiciled (n = 265). No significant association was found between positivity (serology and/or real-time PCR) and variables such as the sex (χ2 = 1.11, df = 1, P = 0.293), age (χ2 = 2.78, df = 1, P = 0.096), clinical status (χ2 = 3.52, df = 1; P = 0.061) and housing condition (χ2 = 1.69, df = 1, P = 0.194).
Human cases
From 2007 to 2017, 49 human cases of CL were notified in 12 indigenous villages of Pesqueira. Of these, 40 cases were diagnosed from February 2012 to December 2013, with 77.5% (n = 31) of the cases being diagnosed in 2012. During this outbreak, 67.5% of the cases were detected in V1 (50%, n = 20), V2 (12.5%, n = 5) and V3 (5%, n = 2). Cases were notified during almost all months of the year, except in May and June. The months with more cases recorded during outbreak were: March 2012 (n = 7) and October 2012 (n = 14). Men (n = 29) were more frequently affected than women (n = 11), and most cases (62.5%) were from 3 to 10 years-old (25%) and 11 to 17 years-old (37.5%), with age ranging from 3 to 66 years.
Discussion
Throughout the study period, we identified 11 sand fly species in the investigated indigenous villages. Migonemyia migonei, E. lenti, L. longipalpis, N. intermedia and M. capixaba were the most abundant species, present in all villages, both indoors and outdoors. From an epidemiological point of view, this finding is very important because it may suggest a year-long risk for the transmission of Leishmania infantum and L. braziliensis in this region. Lutzomyia longipalpis and M. migonei are incriminated as vectors of L. infantum and L. braziliensis, respectively, in Brazil and other Latin American countries [25]. In addition, M. migonei has been strongly suggested as a vector of L. infantum in some foci [26, 27] and a recent laboratory study reinforced this hypothesis [28]. Furthermore, N. intermedia is also a proven vector of L. braziliensis in Brazil [25]. Considering the presence of potential vectors during the whole year, further research focused on detecting DNA (by PCR) or promastigotes (by dissection and microscopical examination) of Leishmania spp. in sand flies could provide valuable data on the transmission pattern in this area, which may include the participation of multiple vectors.
Our results support our initial hypothesis that sand flies may be adapted to human dwellings in the studied indigenous villages. For instance, M. capixaba is a sylvatic species, generally found in forests and marginal areas [29]. However, in the present study, this species was the second most abundant species indoors (SISA = 0.81), after M. migonei (SISA = 1.00). Notably, 46 out of 49 M. capixaba specimens caught indoors were female, which could suggest an endophilic behaviour, although none of them were engorged. Future studies, with a larger number of specimens, are needed to assess the blood meals of M. capixaba females collected indoors and outdoors in these villages. The houses where M. capixaba were found indoors have openings, as did most of the houses in the studied indigenous villages. Moreover, some of the houses were surrounded by native vegetation, which may have favoured the encounter of M. capixaba in their interior. A study carried out in São Vicente Férrer (agreste region of Pernambuco) [30] reported a single female in the peridomicile and 24 males and 31 females in the forest environment. In Caruaru (agreste region of Pernambuco) [31] reported only three females of M. capixaba in the intradomicile. These findings suggest that sylvatic sand fly species (e.g. M. capixaba) may find a home inside human houses in the studied indigenous villages. Indeed, also sand flies incriminated as vectors of Leishmania spp. (i.e. M. migonei, N. intermedia and L. longipalpis) were consistently collected inside the investigated houses during this study. Moreover, 12.1% of M. migonei females caught indoors contained fresh blood in their abdomen, suggesting an endophilic behaviour. Overall, these findings may indicate a constant, close contact between sand fly vectors, domestic animals and humans, potentially increasing the risk of Leishmania spp. transmission. Several factors may drive the adaptation of sand flies to human dwellings, including deforestation, construction of houses close to forest fragments, poor housing conditions and presence of animal sheds in the backyards. All these factors were observed in the indigenous villages surveyed in this study.
Similar studies conducted in Pernambuco reported a species richness ranging between 4–25 species [6, 27, 30–32]. Until this study, 41 sand fly species were considered to be present in Pernambuco [29]. With the record of M. trinidadensis, this study increases the number of sand fly species of Pernambuco to 42, corresponding to approximately 4.3 species per 10,000 km2. Pernambuco has a rich sand fly fauna as compared with other Brazilian states [29], such as Alagoas (3.2 per 10,000 km2) and São Paulo (3.1 per 10,000 km2). Incidentally, some authors have mentioned the presence of M. trinidadensis in Pernambuco [33], but provided no evidence or reference supporting this statement. Indeed, this species was not considered in subsequent sand fly species checklists of this state [13, 29].
The distribution of CL appears to be influenced by altitude. We observed a high species diversity and lower species dominance in the villages with altitude ~850 m (V1 and V2), where 62.5% of the CL cases reported from 2007 to 2017 were concentrated. A study conducted in south-eastern Brazil showed that the number of CL cases decreased progressively with altitude [34]; most cases occurred at 650–750 m and no case occurred at 850–950 m. This is in partial agreement with our results, since CL cases were detected in V3, which is located at an altitude of 965 m. It is worth noting that the highest number of the potential vectors M. migonei and N. intermedia were found exactly in V3, where only two CL cases were reported in ten years. This suggests that the risk of CL in this area may not be directly correlated with sand fly abundance.
The overall number of males was higher than females, as reported in different studies conducted in other regions of Brazil [35–38]. It is acknowledged that the sex ratio may be influenced by trapping methods, with light traps usually attracting more males than females [39]. However, the sex ratio varied widely according to species and environment (indoors vs outdoors, e.g. M. villelai), being close to unity in some (e.g. M. trinidadensis) and female-biased in others (e.g. M. capixaba and E. evandroi). This indicates that females of some species may be more phototropic than others, as emphasized elsewhere [39].
The highest sand fly population peaks occurred in May 2015 and March 2016, corresponding to the pre-rainy season in 2015 and following the first rains in February 2016, respectively. The number of sand flies collected was positively correlated with temperature. It means that sand flies were more frequently trapped during hot months. It is worth noting that in the agreste region of Pernambuco the rains are unevenly distributed throughout the year, occurring mainly from February to July. This factor may be an important driver of the seasonality of sand flies in this region, similarly to what occurs in the semiarid region of Ceará state [40]. The decline in the sand fly collections during torrential rains (June and July) may also be attributed to inherent difficulties in collecting sand flies using light traps under the rain [6]. In other words, the lower trapping success during this period probably related to a reduced flying activity of sand flies during the raining nights rather than due to their absence. In a study conducted in Passira, another municipality located in the agreste region of Pernambuco, the authors reported that 82.4% of the L. longipalpis specimens were collected in months with relative humidity surpassing 75% [31]. In the present study, most L. longipalpis specimens (79.1%) were collected in months with a relative humidity less than 75%. These divergent results reinforce the hypothesis that L. longipalpis in north-eastern Brazil is less dependent on climate [31] or that the relationship between climate and L. longipalpis population may vary locally.
It is worth noting that 50% of the CL cases reported during the outbreak from 2012 to 2013 in Pesqueira were concentrated at V1. Interestingly, most M. migonei females (50.2%) collected indoors in this study were collected in V1. While this species displays a sylvatic behaviour in some Brazilian regions, it is recognized that in north-eastern Brazil this species is adapted to different environments, including forest fragments, animal sheds and human houses [41, 42].
In general, most human cases of CL diagnosed in rural and/or forested areas in north-eastern Brazil are males involved in occupational activities that increase their exposure to sand flies [6, 43–45]. In rural settings where children are frequently affected by CL, the transmission cycle of L. braziliensis is probably taking place in the indoor or immediate outdoor environments [46], where domestic dogs are also frequently exposed. In the present study, a relatively low (8.7%) overall level of exposure to Leishmania spp. infection was detected among dogs living in the studied indigenous villages and two nearby villages, as compared to other studies conducted in Pernambuco [47–50] or elsewhere in Brazil [51, 52]. While the overall level of exposure to Leishmania spp. infection in dogs was relatively low, data from notified CL cases and from our serological survey suggest that transmission is taking place in the peri-domestic and/or domestic environments. Indeed, these dogs are typically free roaming during the day, but stay around or inside the houses during the night. Furthermore, considering that remarkably anthropophilic vector species (e.g. N. intermedia) [10] were found indoors, the risk of exposure to L. braziliensis may be ever higher in humans as compared to dogs. This could partially explain the 2012–2013 outbreak in humans and the relatively low exposure to leishmanial parasites in dogs. Nevertheless, the level of exposure in dogs living in the indigenous villages where the 2012–2013 outbreak of human CL was detected was almost 2-fold higher than in the two nearby villages (11.0 vs 6.2% for serology and 2.6 vs 1.4% for real-time PCR).
While this was not our primary objective for screening dogs, the low exposure to leishmanial parasites in dogs suggests that these animals are not playing any role as reservoirs of L. braziliensis in the study area, which is in line with the current notion that dogs are mere accidental hosts of this parasite [14]. Indeed, previous studies conducted in other endemic foci in Pernambuco have indicated sylvatic and synanthropic rodents as the reservoirs of L. braziliensis [4, 53], a hypothesis also supported by an experimental study [54].
Conclusions
In conclusion, we confirm that proven sand fly vectors are present in the indoor and immediate outdoor environments in indigenous villages where CL is endemic. The adaptation of sand flies to the indoor environment may be related to the poor housing conditions observed in these villages and the proximity of houses to green areas (e.g. crop plantations and forest fragments).
Additional files
Additional file 1: Table S1. Number of sand flies collected indoors and outdoors in each village (V1–V3) in Pesqueira, Pernambuco, Brazil, from March 2015 to March 2016.
Additional file 2: Figure S1. Standardized index of species abundance (SISA) of sand fly species collected indoors and outdoors in Pesqueira, Pernambuco, Brazil, from March 2015 to March 2016.
Acknowledgements
Thanks to Pólo Indígena dos Xukurus and especially to Cacique Marquinhos for their full support during the fieldwork. Thanks also to the Secretaria de Saúde de Pesqueira and IV Gerência Regional de Saúde (IV Geres) for their support with sand fly collection. Thanks to Lucas C. de Sousa-Paula (Aggeu Magalhães Institute, Fiocruz, Brazil) for elaboration of Fig. 1. Publication of this paper has been sponsored by Bayer Animal Health in the framework of the 14th CVBD World Forum Symposium.
Funding
This study was financially supported by Fundação de Amparo à Ciência e Tecnologia de Pernambuco (FACEPE - APQ-0281-2.13/13). FDT is the recipient of a research fellowship from Conselho Nacional de Desenvolvimento Científico e Tecnológico (CNPq; 313118/2018-3).
Availability of data and materials
The data supporting the conclusions of this article are included within the article and its additional files. Raw data can be shared with other researchers upon specific request.
Authors’ contributions
FDT conceived the study. KGSS, LAF and PLC conducted the fieldwork. KGSS and LAF conducted the laboratory testing. KGSS, DEOM and FJS processed and identified the sand flies. FDT and KGSS conducted the literature review and wrote the manuscript. SPBF participated in data interpretation and reviewed the manuscript. All authors read and approved the final manuscript.
Ethics approval and consent to participate
The Committee on Ethics in the Use of Animals CEUA (number: 56/2013) of the Aggeu Magalhães Institute (Fiocruz) approved the procedures used in this study. Dog owners signed a consent form before the inclusion of their animals in this study. Residents of the houses where sand fly collections were conducted authorized the installation of the traps in their houses.
Consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing interests.
Publisher’s Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Contributor Information
Kamila Gaudêncio da Silva Sales, Email: kamilasalesg@gmail.com.
Débora Elienai de Oliveira Miranda, Email: deboraomyranda@gmail.com.
Pietra Lemos Costa, Email: lemos.pietra@gmail.com.
Fernando José da Silva, Email: fs@cpqam.fiocruz.br.
Luciana Aguiar Figueredo, Email: vet.luciana@gmail.com.
Sinval Pinto Brandão-Filho, Email: sinval@cpqam.fiocruz.br.
Filipe Dantas-Torres, Email: filipe.dantas@cpqam.fiocruz.br.
References
- 1.Cantacessi C, Dantas-Torres F, Nolan MJ, Otranto D. The past, present, and future of Leishmania genomics and transcriptomics. Trends Parasitol. 2015;31:100–108. doi: 10.1016/j.pt.2014.12.012. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 2.Alvar J, Vélez ID, Bern C, Herrero M, Desjeux P, Cano J, et al. Leishmaniasis worldwide and global estimates of its the incidence. PLoS ONE. 2012;7:e35671. doi: 10.1371/journal.pone.0035671. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 3.SINAN: Sistema de Informação de Agravos de Notificação. Ministério da Saúde. 2018. http://www.portalsinan.saude.gov.br/. Accessed 27 Feb 2018.
- 4.Lima BS, Dantas-Torres F, de Carvalho MR, Marinho-Junior JF, de Almeida EL, Brito ME, et al. Small mammals as hosts of Leishmania spp in a highly endemic area for zoonotic leishmaniasis in north-eastern Brazil. Trans R Soc Trop Med Hyg. 2013;107:592–597. doi: 10.1093/trstmh/trt062. [DOI] [PubMed] [Google Scholar]
- 5.Maia C, Dantas-Torres F, Campino L. Parasite biology: the reservoir hosts. In: Bruschi F, Gradoni L, editors. The leishmaniases: Old neglected tropical diseases. Cham: Springer; 2018. pp. 79–106. [Google Scholar]
- 6.Dantas-Torres F, Sales KGS, Miranda DEO, da Silva FJ, Figueredo LA, de Melo FL, et al. Sand fly population dynamics and cutaneous leishmaniasis among soldiers in an Atlantic forest remnant in northeastern Brazil. PLoS Negl Trop Dis. 2017;11:e0005406. doi: 10.1371/journal.pntd.0005406. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 7.Desjeux P. The increase in risk factors for leishmaniasis worldwide. Trans R Soc Trop Med Hyg. 2001;95:239–243. doi: 10.1016/S0035-9203(01)90223-8. [DOI] [PubMed] [Google Scholar]
- 8.Antonialli SAC, Torres TG, Paranhos Filho AC, Tolezano FE. Spatial analysis of American visceral leishmaniasis in Mato Grosso do Sul State, Central Brazil. J Infect. 2007;54:509–514. doi: 10.1016/j.jinf.2006.08.004. [DOI] [PubMed] [Google Scholar]
- 9.Harhay MO, Olliaro PL, Costa DL, Costa CH. Urban parasitology: visceral leishmaniasis in Brazil. Trends Parasitol. 2011;27:403–409. doi: 10.1016/j.pt.2011.04.001. [DOI] [PubMed] [Google Scholar]
- 10.Rangel EF, Lainson R. Proven and putative vectors of American cutaneous leishmaniasis in Brazil: aspects of their biology and vectorial competence. Mem Inst Oswaldo Cruz. 2009;104:937–954. doi: 10.1590/S0074-02762009000700001. [DOI] [PubMed] [Google Scholar]
- 11.Sales KG, Costa PL, de Morais RC, Otranto D, Brandão-Filho SP, Cavalcanti MP, Dantas-Torres F. Identification of phlebotomine sand fly blood meals by real-time PCR. Parasit Vectors. 2015;8:230. doi: 10.1186/s13071-015-0840-3. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 12.Sangiorgi B, Miranda DN, Oliveira DF, Santos EP, Gomes FR, Santos EO, et al. Natural breeding places for phlebotomine sand flies (Diptera: Psychodidae) in a semiarid region of Bahia state, Brazil. J Trop Med. 2012;2012:124068. doi: 10.1155/2012/124068. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 13.Dantas-Torres F, de Andrade AJ, Tenório KE, Andrade Filho JD, Balbino VQ, Brandão-Filho SP. Phlebotomine sand flies (Diptera: Psychodidae: Phlebotominae) in the State of Pernambuco. Rev Soc Bras Med Trop. 2010;43:733–736. doi: 10.1590/S0037-86822010000600028. [DOI] [PubMed] [Google Scholar]
- 14.Dantas-Torres F. The role of dogs as reservoirs of Leishmania parasites, with emphasis on Leishmania (Leishmania) infantum and Leishmania (Viannia) braziliensis. Vet Parasitol. 2007;149:139–146. doi: 10.1016/j.vetpar.2007.07.007. [DOI] [PubMed] [Google Scholar]
- 15.Cleaveland S, Meslin FX, Breiman R. Dogs can play useful role as sentinel hosts for disease. Nature. 2006;440:605. doi: 10.1038/440605b. [DOI] [PubMed] [Google Scholar]
- 16.Camara Municipal de Pesqueira. http://www.camarapesqueira.pe.gov.br/oMunicipio.php. Accessed 14 Dec 2018.
- 17.SIASI: Sistema de informação da Atenção da Saúde Indígena. Ministério da Saúde. 2018. http://sis.funasa.gov.br/siasi/. Accessed 14 Dec 2018.
- 18.Instituto Socioambiental. Povos indígenas no Brasil. 2018. https://pib.socioambiental.org/pt/Povo:Xukuru. Accessed 30 Dec 2018.
- 19.Young DG, Duncan MA. Guide to the identification and geographic distribution of Lutzomyia sand flies in Mexico, the West Indies, Central and South America (Diptera: Psychodidae) Mem Am Entomol Inst. 1994;54:1–881. [Google Scholar]
- 20.Galati EAB. Phlebotominae (Diptera, Psychodidae): classification, morphology and terminology of adults and identification of American taxa. In: Rangel E, Shaw J, editors. Brazilian sand flies. Switzerland: Springer; 2018. pp. 9–212. [Google Scholar]
- 21.Francino O, Altet L, Sánchez-Robert E, Rodriguez A, Solano-Gallego L, Alberola J, et al. Advantages of real-time PCR assay for diagnosis and monitoring of canine leishmaniosis. Vet Parasitol. 2006;137:214–221. doi: 10.1016/j.vetpar.2006.01.011. [DOI] [PubMed] [Google Scholar]
- 22.Dantas-Torres F, Sales KGS, da Silva LG, Otranto D, Figueredo LA. Level of agreement between two commercially available rapid serological tests and the official screening test used to detect Leishmania seropositive dogs in Brazil. Vet J. 2018;234:102–104. doi: 10.1016/j.tvjl.2018.02.007. [DOI] [PubMed] [Google Scholar]
- 23.Hammer O, Harper DAT, Ryan PD. PAST: Paleontological Statistics Software Package for education and data analysis. Palaeontol Electron. 2001;4:9. [Google Scholar]
- 24.Roberts DR, Hsi BP. An index of species abundance for use with mosquito surveillance data. Environ Entomol. 1979;8:1007–1013. doi: 10.1093/ee/8.6.1007. [DOI] [Google Scholar]
- 25.Brazil RP, Rodrigues AAF, Filho JDA. Sand fly vectors of Leishmania in the Americas - a mini review. Entomol Ornithol Herpetol. 2015;4:144. [Google Scholar]
- 26.Carvalho GM, Gontijo CM, Falcão AL, Andrade Filho JD. Study of phlebotomine sand flies (Diptera: Psychodidae) collected in a Leishmania-endemic area of the metropolitan region of Belo Horizonte, Brazil. J Med Entomol. 2010;47:972–976. doi: 10.1603/ME09127. [DOI] [PubMed] [Google Scholar]
- 27.Guimarães VCFV, Costa PL, Silva FJ, Silva KT, Silva KG, Araújo AIF, et al. Phlebotomine sandflies (Diptera: Psychodidae) in São Vicente Férrer, a sympatric area to cutaneous and visceral leishmaniasis in the state of Pernambuco, Brazil. Rev Soc Bras Med Trop. 2012;45:66–70. doi: 10.1590/S0037-86822012000100013. [DOI] [PubMed] [Google Scholar]
- 28.Guimarães VCFV, Pruzinova K, Sadlova J, Volfova V, Myskova J, Brandão-Filho SP, Volf P. Lutzomyia migonei is a permissive vector competent for Leishmania infantum. Parasit Vectors. 2016;9:159. doi: 10.1186/s13071-016-1444-2. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 29.Aguiar GM, Vieira VR. Regional distribution and habitats of Brazilian phlebotomine species. In: Rangel E, Shaw J, editors. Brazilian sand flies. Switzerland: Springer; 2018. pp. 251–298. [Google Scholar]
- 30.Agra MCR, Costa PL, Duque AES, Soares ENL, Alves LC, Ramos RAN, et al. Sandflies (Diptera: Psychodidae) in an urban area of northeastern Brazil. Rev Soc Bras Med Trop. 2016;49:698–702. doi: 10.1590/0037-8682-0147-2016. [DOI] [PubMed] [Google Scholar]
- 31.Costa PL, Dantas-Torres F, Silva FJ, Guimarães VCFV, Gaudêncio K, Brandão-Filho SP. Ecology of Lutzomyia longipalpis in an area of visceral leishmaniasis transmission in north-eastern Brazil. Acta Trop. 2013;126:99–102. doi: 10.1016/j.actatropica.2013.01.011. [DOI] [PubMed] [Google Scholar]
- 32.Miranda DE, Sales KG, Faustino MA, Alves LC, Brandão-Filho SP, Dantas-Torres F, et al. Ecology of sand flies in a low-density residential rural area, with mixed forest/agricultural exploitation, in north-eastern Brazil. Acta Trop. 2015;146:89–94. doi: 10.1016/j.actatropica.2015.03.011. [DOI] [PubMed] [Google Scholar]
- 33.Castellon EG, Arias JR, Freitas RA, Naiff RD. Sand flies of the Amazonian region Manaus-Humaitá Highway, State of Amazonas, Brazil (Diptera: Psychodidae: Phlebotominae) Acta Amazon. 1994;24:91–102. doi: 10.1590/1809-43921994242102. [DOI] [Google Scholar]
- 34.Ferreira AL, Sessa PA, Varejão JB, Falqueto A. Distribution of sand flies (Diptera: Psychodidae) at different altitudes in an endemic region of American cutaneous leishmaniasis in the State of Espírito Santo, Brazil. Mem Inst Oswaldo Cruz. 2001;96:1061–1067. doi: 10.1590/S0074-02762001000800006. [DOI] [PubMed] [Google Scholar]
- 35.Michalsky EM, Franca-Silva JC, Barata RA, Lara e Silva FO, Loureiro AMF, Fortes-Dias CL, et al. Phlebotominae distribution in Janaúba, an area of transmission for visceral leishmaniasis in Brazil. Mem Inst Oswaldo Cruz. 2009;104:56–61. doi: 10.1590/S0074-02762009000100009. [DOI] [PubMed] [Google Scholar]
- 36.Almeida OS, Minzão ER, Minzão LD, Silva SR, Ferreira AD, Faccenda O, et al. Aspectos ecológicos de flebotomíneos (Diptera: Psychodidae) em área urbana do município de Ponta Porã, Estado de Mato Grosso do Sul. Rev Soc Bras Med Trop. 2010;43:723–727. doi: 10.1590/S0037-86822010000600025. [DOI] [PubMed] [Google Scholar]
- 37.Thies SF, Bronzoni RVMB, Espinosa MM, Souza CO, Ribeiro ALM, Santos ES, et al. Frequency and diversity of phlebotomine sand flies (Diptera: Psychodidae) in Sinop, State of Mato Grosso, Brazil. Rev Soc Bras Med Trop. 2016;49:544–552. doi: 10.1590/0037-8682-0251-2016. [DOI] [PubMed] [Google Scholar]
- 38.Fernandes WS, Borges LM, Casaril AE, Oliveira EF, Infran JOM, Piranda EM, et al. Sandfly fauna (Diptera: Psychodidae) in an urban area, Central-West of Brazil. Rev Inst Med Trop Sao Paulo. 2017;59:e54. doi: 10.1590/S1678-9946201759054. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 39.Alexander B. Sampling methods for phlebotomine sandflies. Med Vet Entomol. 2000;14:109–122. doi: 10.1046/j.1365-2915.2000.00237.x. [DOI] [PubMed] [Google Scholar]
- 40.Deane LM, Deane MP. Encontro de Leishmania nas vísceras e na pele de uma raposa, em zona endêmica de calazar, nos arredores de Sobral no Ceará. Hosp. 1954;45:419–421. [PubMed] [Google Scholar]
- 41.Andrade Filho JD, Brazil RP. Phlebotomine sand flies (Diptera: Psychodidae: Phlebotominae) of Alagoas state, northeast of Brazil. Neotrop Entomol. 2009;38:688–690. doi: 10.1590/S1519-566X2009000500022. [DOI] [PubMed] [Google Scholar]
- 42.Souza FMT, Santos CFR, de Andrade EM, Marcondes CB, de Queiroz Balbino V, Pessoa FAC. New records of phlebotomine sand flies (Diptera: Psychodidae) from the state of Alagoas, northeast of Brazil. J Med Entomol. 2018;55:242–247. doi: 10.1093/jme/tjx175. [DOI] [PubMed] [Google Scholar]
- 43.Araújo AR, Portela NC, Feitosa AP, Silva OA, Ximenes RA, Alves LC, et al. Risk factors associated with American cutaneous leishmaniasis in an endemic area of Brazil. Rev Inst Med Trop Sao Paulo. 2016;58:86. doi: 10.1590/S1678-9946201658086. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 44.Brilhante AF, Melchior LAK, Nunes VLB, Cardoso CO, Galati EAB. Epidemiological aspects of American cutaneous leishmaniasis (ACL) in an endemic area of forest extractivist culture in western Brazilian Amazonia. Rev Inst Med Trop Sao Paulo. 2017;59:e12. doi: 10.1590/s1678-9946201759012. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 45.Turetz ML, Machado PR, Ko AI, Alves F, Bittencourt A, Almeida RP, et al. Disseminated leishmaniasis: a new and emerging form of leishmaniasis observed in northeastern Brazil. J Infect Dis. 2002;186:1829–1834. doi: 10.1086/345772. [DOI] [PubMed] [Google Scholar]
- 46.Guerra JA, Barbosa M, Loureiro AC, Coelho CP, Rosa GG, Coelho LI. American tegumentary leishmaniasis in children: epidemiological aspects of cases treated in Manaus, Amazonas, Brazil. Cad Saude Publica. 2007;23:2215–2223. doi: 10.1590/S0102-311X2007000900029. [DOI] [PubMed] [Google Scholar]
- 47.Dantas-Torres F, Brito MEF, Brandão-Filho SP. Seroepidemiological survey on canine leishmaniasis among dogs from an urban area of Brazil. Vet Parasitol. 2006;31:54–60. doi: 10.1016/j.vetpar.2006.03.008. [DOI] [PubMed] [Google Scholar]
- 48.Dantas-Torres F, de Paiva-Cavalcanti M, Figueredo LA, Melo MF, da Silva FJ, da Silva AL, et al. Cutaneous and visceral leishmaniasis in dogs from a rural community in northeastern Brazil. Vet Parasitol. 2010;170:313–317. doi: 10.1016/j.vetpar.2010.02.019. [DOI] [PubMed] [Google Scholar]
- 49.Santos JML, Dantas-Torres F, Mattos MRF, Lino FRL, Andrade LSS, Souza RCA, et al. Prevalence of anti-Leishmania spp antibodies in dogs from Garanhuns, in the middle scrub zone (Agreste) of Pernambuco. Rev Soc Bras Med Trop. 2010;43:41–45. doi: 10.1590/S0037-86822010000100010. [DOI] [PubMed] [Google Scholar]
- 50.Figueredo LA, Sales KGS, Deuster K, Pollmeier M, Otranto D, Dantas-Torres F. Exposure to vector-borne pathogens in privately owned dogs living in different socioeconomic settings in Brazil. Vet Parasitol. 2017;243:18–23. doi: 10.1016/j.vetpar.2017.05.020. [DOI] [PubMed] [Google Scholar]
- 51.França-Silva JC, Costa RT, Siqueira AM, Machado-Coelho GLL, Costa CA, Mayrink W, et al. Epidemiology of canine visceral leishmaniosis in the endemic area of Montes Claros Municipality, Minas Gerais State, Brazil. Vet Parasitol. 2003;111:161–173. doi: 10.1016/S0304-4017(02)00351-5. [DOI] [PubMed] [Google Scholar]
- 52.Malaquias LCC, Romualdo RC, Anjos JB, Giunchetti RC, Corrêa-Oliveira R, Reis AB. Serological screening confirms the re-emergence of canine leishmaniosis in urban and rural areas in Governador Valadares, Vale do Rio Doce, Minas Gerais, Brazil. Parasitol Res. 2007;100:233–239. doi: 10.1007/s00436-006-0259-z. [DOI] [PubMed] [Google Scholar]
- 53.Brandão-Filho SP, Brito ME, Carvalho FG, Ishikawa EA, Cupolillo E, Floeter-Winter L, et al. Wild and synanthropic hosts of Leishmania (Viannia) braziliensis in the endemic cutaneous leishmaniasis locality of Amaraji, Pernambuco State, Brazil. Trans R Soc Trop Med Hyg. 2003;97:291–296. doi: 10.1016/S0035-9203(03)90146-5. [DOI] [PubMed] [Google Scholar]
- 54.Andrade MS, Courtenay O, Brito MEF, Carvalho FG, Carvalho AWS, Soares F, et al. Infectiousness of sylvatic and synanthropic small rodents implicates a multi-host reservoir of Leishmania (Viannia) braziliensis. PLoS Negl Trop Dis. 2015;9:e0004137. doi: 10.1371/journal.pntd.0004137. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 55.QGIS Development Team. QGIS Geographic Information System. Open Source Geospatial Foundation Project. https://www.qgis.org/en/site/. Accessed 13 Dec 2018.
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.
Supplementary Materials
Additional file 1: Table S1. Number of sand flies collected indoors and outdoors in each village (V1–V3) in Pesqueira, Pernambuco, Brazil, from March 2015 to March 2016.
Additional file 2: Figure S1. Standardized index of species abundance (SISA) of sand fly species collected indoors and outdoors in Pesqueira, Pernambuco, Brazil, from March 2015 to March 2016.
Data Availability Statement
The data supporting the conclusions of this article are included within the article and its additional files. Raw data can be shared with other researchers upon specific request.