

Abstract
Background/Aim:
To determine whether Gramicidin S (GS)-nitroxide, JP4–039, esophageal radiation protection protected lung tumors in a transgenic model, LoxP-Stoop-LoxP Kristen Rat Sarcoma Viral oncogene (LSL-K-RAS) mice were administered intra-tracheal-Carbapenem-resistant Enterobacteriaceae (CRE) recombinase, bilateral lung tumors were confirmed at 11 weeks, then thoracic irradiation was delivered.
Materials and Methods:
Mice received single-fraction 15 Gy or 24 Gy to both lungs, in subgroups receiving intraesophageal administration 10 min before irradiation of JP4–039 (in F15 emulsion) tumor size reduction and survival were investigated. Mice were followed for survival, and reduction in tumor size.
Results:
There was no evidence of tumor radioprotection in mice receiving JP4–039/F15.
Conclusion:
Intraesophageal radioprotective small-molecule antioxidant therapy protects normal tissue but not tumor tissue in mice with transgenic lung tumors.
Keywords: GS-Nitroxide, esophageal radioprotection, transgenic lung cancer
A major complication of clinical radiotherapy of lung cancer is damage to normal tissues in the surrounding target volume. In particular, radiotherapy of non-small cell lung carcinoma is associated with significant esophagitis, which may be doselimiting, and can result in esophageal stricture in survivors. Animal models of lung cancer have facilitated the search for new radioprotector drugs (1–5). Local administration of both manganese superoxide dismutase-plasmid liposome (MnSOD-PL) gene therapy (6–28) and the small molecule GS-nitroxide conjugate JP4–039 (29–33) has been demonstrated to selectively protect the esophagus with no significant tumor radioprotection in orthotopic tumor models (16, 18, 20, 34). Intraesophageal administration of MnSOD-PL in mice with orthotopic Lewis lung carcinomas at the carina has been shown to reduce irradiation-induced esophagitis with no significant reduction in survival due to increase in tumor size (20, 34). In other experiments, single or multiple administrations of MnSOD-PL to the oral cavity during head and neck irradiation of mice with orthotopic squamous cell carcinoma in the floor of the mouth demonstrated no significant tumor radioprotection (16, 35). Recently, a small molecule GS-nitroxide has been reported to provide significant local tissue radioprotection when administered in a novel F15 lipid emulsion (32). Fluorochrome-labeled JP4–039-BODIPY has been demonstrated to target mitochondria of cells in vitro (4), supporting its proposed mechanism of action (33).
In the present studies, we tested the effect of intraesophageal administration of JP4–039/F15 in a model of transgenic lung carcinoma. LSL-K-RAS mice were administered intraesophageal CRE-recombinase, which has been shown to induce multi-focal bilateral lung tumors in the subsequent 11–15 weeks (36). Mice with established bilateral tumors received single-fraction whole-lung irradiation with the goal of providing tumor size reduction and long-term local control. In subgroups of mice that received intraesophageal administration of JP4–039/F15, we compared tumor size reduction by irradiation and survival. We sought to determine whether the known esophageal radioprotection mediated by JP4–039 (32) would also provide tumor radioprotection, and lead to more rapid death from faster tumor growth.
Materials and Methods
LSL-K-RAS transgenic tumor model.
LSL-K-RAS mice and control LSL mice (3, 37–40) were obtained from McGarry Houghton (36) and housed four per cage according to the University of Pittsburgh Institutional Animal Care and Use Committee (IACUC) regulations. Mice were fed standard laboratory chow. At ages of six to eight weeks, male and female mice were administered intratracheal CRE-recombinase according to published methods (36). Mice were followed for 11–14 weeks and at serial time points, representative animals were examined for the appearance of bilateral lung tumors. At the time when lung tumors of measurable size in all mice were detected, subgroups of 8 mice were randomized to receive radioprotective esophageal therapy prior to whole-lung irradiation.
Administration of JP4–039 in F15 emulsion.
The GS-nitroxide conjugate, JP4–039 (33), in F15 liposome emulsion (100 μl containing 400 μg drug/mouse) was administered orally prior to irradiation as published elsewhere (32).
Mouse lung irradiation.
Adult mice with bilateral CRE-recombinase-induced tumors, received single fraction 15 Gy or 24 Gy total-lung irradiation according to published methods (7). The head, neck, and lower body were shielded so that only the lung received irradiation (32). Mice were treated on a 6 MV linear accelerator (Varian Corporation, Palo Alto, CA, USA) according to published methods (32). Mice were followed-up for survival. When animals became moribund or lethargic, they were sacrificed.
Lung histopathology and evaluation of tumor size.
Mice dying of extension of lung cancer, radiation-associated morbidity, or at the end of the experiment at day 110 were sacrificed. The lungs were expanded with Optimal Cutting Temperature (OCT), excised, frozen, sectioned and hematoxylin and eosin (H&E)-stained lung sections were examined for the percentage of lung containing tumor. Histopathological evaluation of both lungs was carried out in serial sections from each animal. Ten sections of each lung were analyzed and the percentage of lung replaced by tumor was calculated. Histopathological findings of acute or late radiation toxicity in the lungs were documented as published (34).
Statistics.
Kaplan−Meier survival curves were plotted for each of the four treatment groups (namely, unirradiated control group; 19 Gy upper body irradiation only; F15 intraesophageal administration prior to irradiation; and intraesophageal JP4–039/F15 prior to irradiation). Comparison between any two groups was carried with the two-sided log-rank test. In the tumor growth study, sections of each lung were analyzed descriptively, where the percentage of lung replaced by tumor was calculated.
Results
CRE-recombinase induced bilateral lung cancers in LSL-K-RAS mice.
Mice treated with CRE-recombinase intra-nasal injection as described in the methods and as published elsewhere (36) were followed for 11–17 weeks and at serial times examined for the presence of bilateral lung cancer. Figure 1A shows the histological appearance of representative bilateral lung tumors in mice receiving intra-nasal CRE-recombinase. Lung tumors were evaluated histopathologically and diagnosed as either adenomas or squamous cell carcinomas. These results confirm and extend those of previous publications using the CRE-recombinase model for induction of bilateral transgenic lung cancer (3, 36–40).
Figure 1.
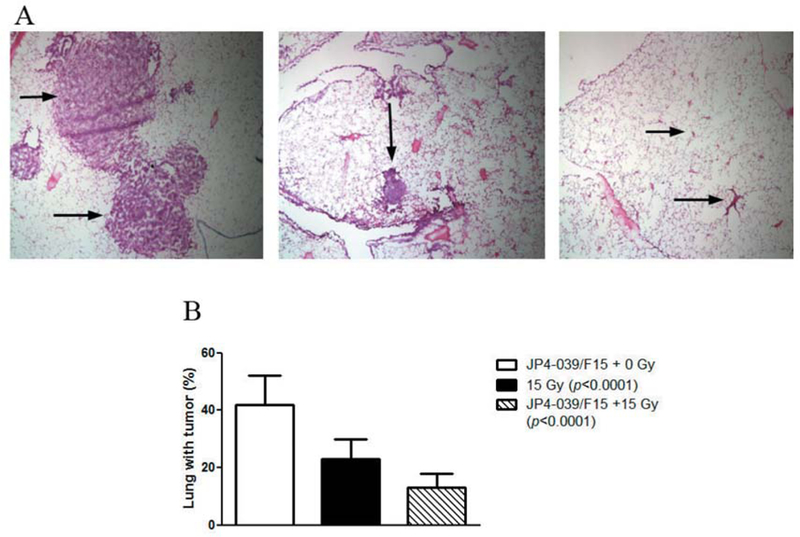
Intraesophageal swallow of JP4–039/F15 (100 μl containing 4 mg/ml drug) protects esophagus but not transgenic LSL-K-RAS_induced lung cancers from therapeutic irradiation. LSL-K-RAS male mice were injected intranasally with adeno CRE-recombinase as published (84). Fourteen weeks later the mice were treated with: intraesophageal F15 alone, JP4–039/F15, or no drug, then 15 Gy to the thoracic cavity. Seven days later, the mice were sacrificed, lungs expanded with OCT, excised, frozen in OCT, sectioned, and H&E-stained. The lungs were examined for the percentage of lung containing tumor. Each lung lobe was scored in 10 sagittal sections: A: Representative lung from tumor-bearing mouse (left), after irradiation at day 7 (middle), and at day 14 (right) each group (×4). B: Quantitation: JP4–039 + 15 Gy or 15 Gy-alone significantly reduced the percentage of lung tumor. Swallowed JP4–039 did not reduce the therapeutic effect of 15 Gy.
Intraesophageal absorption of JP4–039/F15 provides esophageal radiation protection with no detectable tumor protection.
Mice receiving intraesophageal JP4–039/F15 were irradiated to 15 Gy to the thoracic cavity. One group of control mice received 15 Gy irradiation with no prior JP4–039/F15. Another control group received F15-alone intraesophageal administration with no irradiation. As shown in Figure 1B at seven days after 15 Gy thoracic irradiation, tumor size was reduced by irradiation with no tumor protective effects (size increase) by JP4–039/F15.
In a second experiment, we observed mice for 110 days. As shown in Figure 2, a higher lung irradiation dose of 24 Gy resulted in a longer survival of mice to 50 days. There were no early deaths. Subgroups of animals receiving intraesophageal F15 emulsion alone prior to 24 Gy showed no deaths until 60 days. Those receiving JP4–039/F15 prior to irradiation showed no deaths until 50 days. The results demonstrated no significant decrease in survival in mice receiving intraesophageal radioprotective JP4–039/F15.
Figure 2.
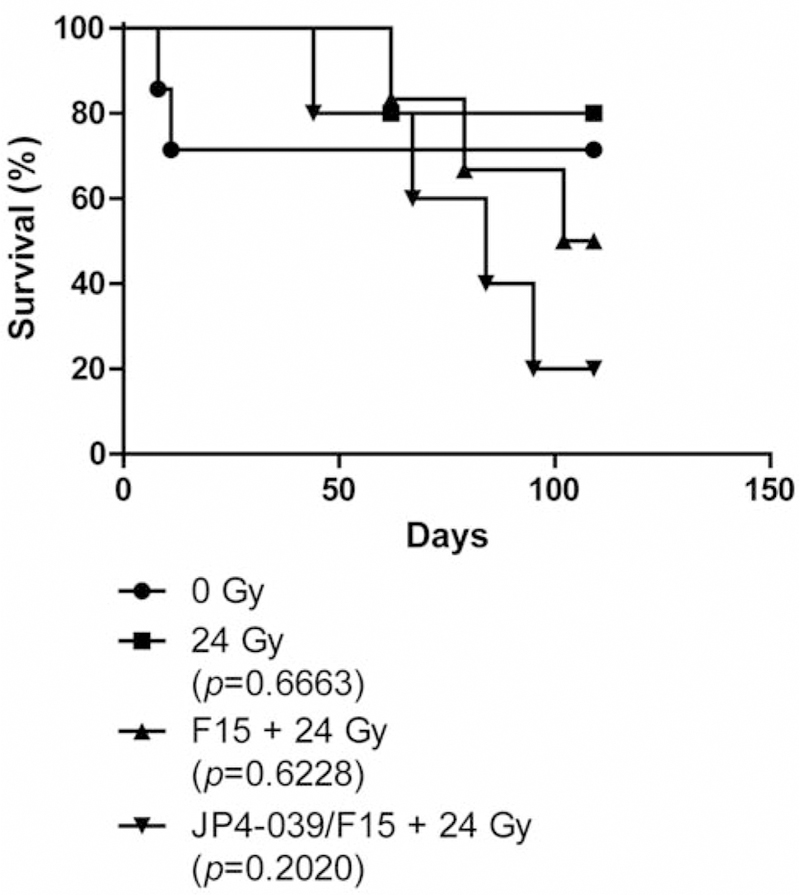
Effect of intra-oral JP4–039/F15 on thoracic irradiation of LSL-K-RAS tumor-bearing mice. Four groups of LSL-K-RAS mice (n=7 per group) were intra-nasally administered adeno-CRE recombinase (5×106 Plaque-forming unit (PFU)). Fourteen weeks later, the mice were randomly assigned to one of four treatments: 0 Gy, 24 Gy, F15 (100 μl) + 24 Gy, or JP4–039/F15 + 24 Gy (400 μg in 100 μl). Drug was administered 15 minutes before irradiation. Mice were irradiated to 24 Gy to the pulmonary cavity. The remainder of the head and neck and abdomen was shielded to preserve bone marrow and intestine. Mice were followed for detection of difficulty in breathing, which was an indication of tumor growth or development of irradiation pulmonary toxicity. All surviving mice were sacrificed at 110 days after irradiation, the lungs removed, and examined for residual tumor areas. No residual tumors were detected in any mice, including unirradiated controls. There was no significant difference in survival between irradiated groups determined using a log-rank statistical test with p-values comparing different treatment groups to the 0 Gy group. There was also no significant difference between irradiation groups.
The survival curve of mice with lung tumors not receiving irradiation demonstrated no late deaths from lung cancer after day 10, suggesting tumor regression in a sub-population of tumor-bearing mice (Figure 2).
Histological evidence of spontaneous tumor regression in mice surviving beyond 10 days.
Surviving LSL-K-RAS mice that had been injected with CRE-recombinase were analyzed at day 110 for histopathological evaluation of lung tissue. Mice dying at earlier time points were also analyzed. The gross appearance of the lungs from five representative mice at 110 days showed no evidence of tumor (Figure 3). Two sections from each of the representative mice are shown in Figure 3. Two mice (Figure 3 A and B) showed normal lung architecture, while others (Figure 3 C and E) showed some peri-alveolar bronchial infiltrates of inflammatory cells. H&E-stained frozen sections were examined for evidence of tumor (adenoma or adenocarcinomas). No tumors were observed in any of 690 lung sections examined in all dying mice or those sacrificed after day 10. All observed lesions of inflammatory infiltrates were within normal limits for LSL mice at 110 days of age. Inflammatory foci (infiltrates) were observed around some airways (Figure 3 C-E) and vessels were considered normal and unrelated to any tumor or disease.
Figure 3.
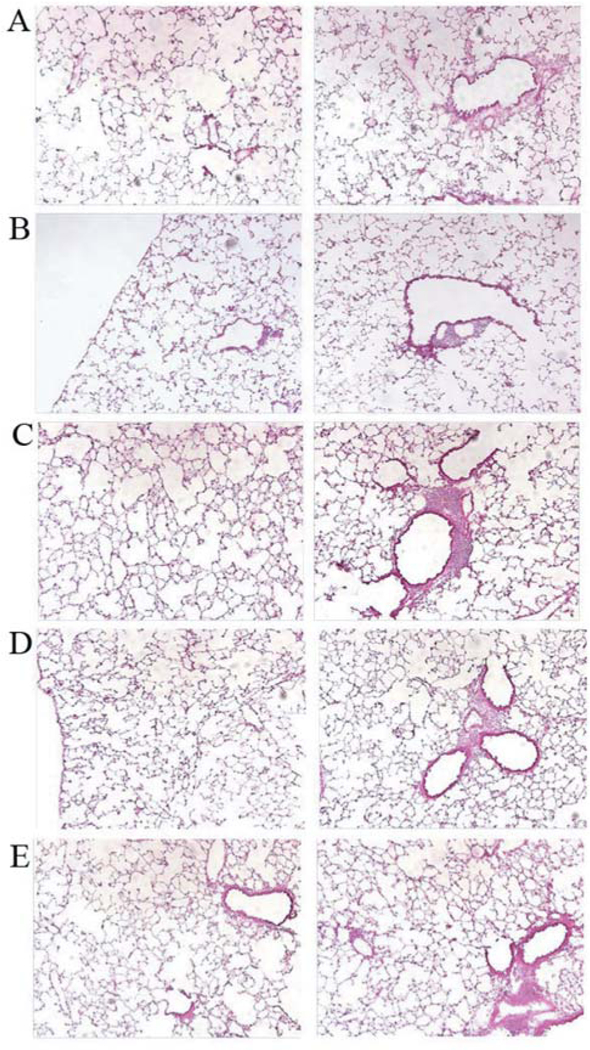
Lack of detectable residual lung tumors 110 days post-24 Gy thoracic irradiation. Lungs of five representative transgenic tumor-bearing mice show lack of detectable residual tumor 110 days post-24 Gy thoracic irradiation. Representative H&E-stained sections from two mice (A, B) with no residual tumor, or with peribronchial inflammatory cell infiltration (C-E) (×10).
The survival of non-irradiated control tumor-bearing mice beyond 10 days after detection of bilateral lung tumors suggested spontaneous resolution of these CRE-recombinase-induced lung cancers. Histopathological evaluation was carried-out in mice in all groups. There were no differences in lung morphology detected in the JP4–039/F15-or F15-treated groups when compared to the irradiated control group.
Discussion
The present results confirm and extend prior reports demonstrating the effect of esophageal radioprotection by swallowed JP4–039/F15 (32), now in another (LSL) mouse strain. In previous studies, C57BL/6NHsd mice received total-lung irradiation preceded by intraesophageal JP4–039/F15. There was a significant decrease in irradiation-induced esophagitis with higher doses of irradiation using JP4–039/F15 and decrease in early death with no significant morbidity of other tissues. Furthermore, there was no evidence of drug uptake in organs outside the oral cavity/esophagus, confirming and extending other publications demonstrating the safety of a local administration of radioprotective GS-nitroxide conjugates (32).
A new finding of the present experiments was the long-term survival of 70% of control non-irradiated mice with bilateral CRE-recombinase-induced tumors. Prior publications indicated that some of the tumors in CRE-recombinase-treated mice are not squamous cell carcinomas, but represent adenomas or benign lung tumors (3, 37–41). In the present studies, careful histological evaluation of lungs in long-term surviving mice demonstrated no evidence of tumors, not only in irradiated or JP4–039/F15 treated then irradiated groups, but also in control unirradiated groups. These results are consistent with the interpretation that some bilateral lung tumors in LSL-K-RAS, CRE-recombinase induced K-RAS transgenic mice are benign tumors and not associated with progression, invasion, or metastasis. The results suggest caution in using this CRE-recombinase-induced transgenic lung cancer model for studies of the therapy of aggressive rather than limitedstage lung cancers and suggest the need for use of another model, which includes a mutant p53 allele (41).
There has been controversy over the use of orthotopic tumors in mice for cancer therapy experiments rather than mice with transgenic cancer. Orthotopic tumor models have been criticized as unrealistic and not representative of true lung cancer in that they have been established by transplantation of documented tumor cell lines, including those removed from chemical carcinogen-or virally-induced established lung cancer (16, 18, 20, 34). In particular, the Lewis lung carcinoma model, which has been a mainstay of orthotopic tumor experiments for lung cancer, has been shown to produce locally invasive and metastatic lung cancer (34), but suffers as a model system as it is not representative of spontaneous lung cancer. Other models of orthotopic head and neck cancer have also been utilized for studies of normal tissue radioprotection (16), and similar criticism has been levied at the use of orthotopic squamous cell transplantation of established tumor cell lines in the floor of the mouth as a representative model system for head and neck cancer.
An alternative to the use of orthotopic tumor models is the transgenic tumor model. The LSL-K-RAS and C57BL/6-K-RAS models have been used as being representative of spontaneous true transgenic tumors and have been suggested as being more valuable (36), reproducible, and efficient for studies of tumor biology. Several transgenic models of lung carcinoma have included the hepatocyte growth factor (HGF) transgenic model (40) or spontaneous lung tumors arising in specific mouse strains.
In the present studies, we tested the LSL-K-RAS model as a system in which to document the effectiveness of radioprotective small-molecule therapy for the esophagus in a setting of true spontaneous tumors. The present results demonstrate the reproducible induction of bilateral transgenic lung tumors in LSL-K-RAS mice treated with intratracheal CRE-recombinase. However, 70% of these tumors spontaneously regressed in non-irradiated, non-drug-treated mice in the control group. Upon histopathological examination, no lung carcinomas were detected 110 days past the time-point when tumors were originally induced. In contrast, in orthotopic tumor-bearing mice, all would have died from progressive or metastatic cancer by this time point (16, 18, 20, 34). Further studies are required to identify an appropriate transgenic mouse model for high frequency detectable and reproducibly fatal lung carcinomas.
At present, orthotopic tumor models of localized yet progressive and fatal lung cancer may be more appropriate for evaluating the safety of intraesophageal radioprotective therapies in a setting in which all control non-irradiated mice would be expected to die of progressive lung cancer.
Acknowledgements
This manuscript is supported by the following grants: NIH R01-CA119927-11 and NIH/NIAID 1U19A168021-06. This project used the UPCI animal facility that is supported in part by award P30CA047904.
References
- 1.Meuwissen R and Berns A: Mouse models for human lung cancer. Genes Dev 19: 643–664, 2005. [DOI] [PubMed] [Google Scholar]
- 2.Stabile LP, Lyker JS, Land SR, Daeie S, Zamboni BA and Siegfried JM: Transgenic mice overexpressing hepatocyte growth factor in the airways show increased susceptibility to lung cancer. Carcinogenesis 27: 1547–1555, 2006. [DOI] [PubMed] [Google Scholar]
- 3.Jackson EL, Willis N, Mercer K, Bronson RT, Crowley D, Montoya R, Jacks T and Tuveson DA: Analysis of lung tumor initiation and progression using conditional expression of oncogenic K-ras. Genes Dev 15: 3243–3248, 2001. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 4.Bernard ME, Kim H, Berhane H, Epperly MW, Franicola D, Zhang X, Houghton F, Shields D, Wang H, Bakkenist CJ, Frantz M-C, Wipf P and Greenberger JS: GS-nitroxide (JP4–039) mediated radioprotection of human Fanconi anemia cell lines. Radiat Res 176: 603–612, 2011. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 5.Winslow MM, Dayton TL, Verhaak RGW, Kim-Kiselak C, Snyder EL, Feldser DM, Hubbard DD, DuPage MJ, Whittaker CA, Hoersch S, Yoon S, Crowley D, Bronson RT, Chiang DY, Meyerson M and Jacks T: Suppression of lung adenocarcinoma progression by Nkx2–1. Nature 473: 101–104, 2011 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 6.Epperly MW, Gretton JE, Sikora CA, Jefferson M, Bernarding M, Nie S and Greenberger JS: Mitochondrial localization of superoxide dismutase is required for decreasing radiation-induced cellular damage. Radiat Res 160: 568–578, 2003. [DOI] [PubMed] [Google Scholar]
- 7.Epperly MW, Gretton JA, DeFilippi SJ, Sikora CA, Liggitt D, Koe G and Greenberger JS: Modulation of radiation-induced cytokine elevation associated with esophagitis and esophageal stricture by manganese superoxide dismutase-plasmid/liposome (SOD-PL) gene therapy. Radiat Res 155: 2–14, 2001. [DOI] [PubMed] [Google Scholar]
- 8.Tarhini AA, Luketich JD, Ramalingam SS, Argiris A, Gooding W, Petro D, Kane K, Liggitt D, Championsmith T, Zhang X, Epperly MW and Greenberger JS: A phase I study of concurrent chemotherapy (paclitaxel and carboplatin) and thoracic radiotherapy with swallowed manganese superoxide dismutase (MnSOD) plasmid liposome (PL) protection in patients with locally advanced stage III non-small cell lung cancer. Hum Gene Ther 22: 336–342, 2011. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 9.Stickle RL, Epperly MW, Klein E, Bray JA and Greenberger JS: Prevention of irradiation-induced esophagitis by plasmid/liposome delivery of the human manganese superoxide dismutase (MnSOD) transgene. Radiat Oncol Invest Clin−Basic Res 7(6): 204–217, 1999. [DOI] [PubMed] [Google Scholar]
- 10.Epperly MW, Sikora C, Defilippi S, Bray J, Koe G, Liggitt D, Luketich JD and Greenberger JS: Plasmid/liposome transfer of the human manganese superoxide dismutase (MnSOD) transgene prevents ionizing irradiation-induced apoptosis in human esophagus organ explants culture. Int J Cancer (Radiat Oncol Invest) 90(3): 128–137, 2000. [DOI] [PubMed] [Google Scholar]
- 11.Epperly MW, Zhang X, Nie S, Cao S, Kagan V, Tyurin V and Greenberger JS: MnSOD-plasmid liposome gene therapy effects on ionizing irradiation induced lipid peroxidation of the esophagus. In Vivo 19: 997–1004, 2005. [PubMed] [Google Scholar]
- 12.Epperly MW, Goff JP, Sikora CA, Shields DS and Greenberger JS: Bone marrow origin of cells with capacity for homing and differentiation to esophageal squamous epithelium. Radiat Res 162: 233–240, 2004. [DOI] [PubMed] [Google Scholar]
- 13.Niu Y, Epperly MW, Shen H, Smith T, Lewis D, Gollin S and Greenberger JS: Intraesophageal MnSOD-plasmid liposome administration enhances engraftment and self-renewal capacity of bone marrow derived progenitors of esophageal squamous epithelium. Gene Therapy 15: 347–356, 2008. [DOI] [PubMed] [Google Scholar]
- 14.Epperly MW, Shen H, Zhang X, Nie S, Cao S and Greenberger JS: Protection of esophageal stem cells from ionizing irradiation by MnSOD-plasmid liposome gene therapy. In Vivo 19: 965–974, 2005. [PubMed] [Google Scholar]
- 15.Epperly MW, Smith T, Wang H, Schlesselman J, Franicola D and Greenberger JS: Modulation of total body irradiation induced life shortening by systemic intravenous MnSOD-plasmid liposome gene therapy. Radiat Res 170(4): 437–444, 2008. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 16.Epperly MW, Wegner R, Kanai AJ, Kagan V, Greenberger EE, Nie S and Greenberger JS: Irradiated murine oral cavity orthotopic tumor antioxidant pool destabilization by MnSOD-plasmid liposome gene therapy mediates tumor radiosensitization. Radiat Res 267: 289–297, 2007. [DOI] [PubMed] [Google Scholar]
- 17.Niu Y, Wang H, Wiktor-Brown D, Rugo R, Shen H, Huq MS, Engelward B, Epperly M and Greenberger JS: Irradiated esophageal cells are protected from radiation-induced recombination by MnSOD gene therapy. Radiat Res 173: 453–461, 2010. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 18.Epperly MW, Lai SM, Mason N, Lopresi B, Dixon T, Franicola D, Niu Y, Wilson WR, Kanai AJ and Greenberger JS: Effectiveness of combined modality radiotherapy of orthotopic human squamous cell carcinomas in Nu/Nu mice using cetuximab, tirapzamine, and MnSOD-plasmid liposome gene therapy. In Vivo 24: 1–8, 2010. [PMC free article] [PubMed] [Google Scholar]
- 19.Rajagopalan MS, Stone B, Rwigema J-C, Salimi U, Epperly MW, Goff J, Franicola D, Dixon T, Cao S, Zhang X, Buchholz BM, Bauer AJ, Choi S, Bakkenist C, Wang H and Greenberger JS: Intraesophageal manganese superoxide dismutase-plasmid liposomes ameliorates novel total body and thoracic irradiation sensitivity of homologous deletion recombinant negative nitric oxide synthase-1 (NOS1−/−) mice. Radiat Res 174: 297–312, 2010. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 20.Epperly MW, Defilippi S, Sikora C, Gretton J, Kalend K and Greenberger JS: Intratracheal injection of manganese superoxide dismutase (MnSOD) plasmid/liposomes protects normal lung but not orthotopic tumors from irradiation. Gene Ther 7(12): 1011–1018, 2000. [DOI] [PubMed] [Google Scholar]
- 21.Epperly MW, Kagan VE, Sikora CA, Gretton JE, Defilippi SJ, Bar-Sagi D and Greenberger JS: Manganese superoxide dismutase-plasmid liposome (MnSOD-PL) administration protects mice from esophagitis associated with fractionated irradiation. Int J Cancer (Radiat Oncol Invest) 96(4): 221–233, 2001. [DOI] [PubMed] [Google Scholar]
- 22.Epperly MW, Carpenter M, Agarwal A, Mitra P, Nie S and Greenberger JS: Intra-oral manganese superoxide dismutase plasmid liposome radioprotective gene therapy decreases ionizing irradiation-induced murine mucosal cell cycling and apoptosis. In Vivo 18: 401–410, 2004. [PubMed] [Google Scholar]
- 23.Greenberger JS and Epperly MW: Pleiotrophic stem cell and tissue effects of ionizing irradiation protection by MnSOD-plasmid liposome gene therapy. In: Progress in Gene TherapyFrank Columbus (ed.). Nova Science Publications, pp. 110–118, 2005. [Google Scholar]
- 24.Greenberger JS, Epperly MW, Gretton J, Jefferson M, Nie S, Bernarding M, Kagan V and Guo HL: Radioprotective gene therapy. Curr Gene Ther 3: 183–195, 2003. [DOI] [PubMed] [Google Scholar]
- 25.Greenberger JS and Epperly MW: Radioprotective antioxidant gene therapy: Potential mechanisms of action. Gene Ther Mol Biol 8: 31–44, 2004. [Google Scholar]
- 26.Epperly MW, Bray JA, Esocobar P, Bigbee WL, Watkin S and Greenberger JS: Overexpression of the human MnSOD transgene in subclones of murine hematopoietic progenitor cell line 32D cl 3 decreases irradiation-induced apoptosis but does not alter G2/M or G1/S phase cell cycle arrest. Radiat Oncol Invest Clin− Basic Res 7: 331–342, 1999. [DOI] [PubMed] [Google Scholar]
- 27.Epperly MW, Travis EL, Sikora C and Greenberger JS: Magnesium superoxide dismutase (MnSOD) plasmid/liposome pulmonary radioprotective gene therapy: Modulation of irradiation-induced mRNA for IL-1, TNF-α, and TGF-β correlates with delay of organizing alveolitis/fibrosis. BiolBlood Marrow Transpl 5: 204–214, 1999. [DOI] [PubMed] [Google Scholar]
- 28.Epperly MW, Guo HL, Jefferson M, Wong S, Gretton J, Bernarding M, Bar-Sagi D and Greenberger JS: Cell phenotype specific duration of expression of epitope-tagged HA-MnSOD in cells of the murine lung following intratracheal plasmid liposome gene therapy. Gene Therapy 10: 163–171, 2003. [DOI] [PubMed] [Google Scholar]
- 29.Goff JP, Epperly MW, Shields D, Wipf P, Dixon T and Greenberger JS: Radiobiologic effects of GS-nitroxide (JP4–039) in the hematopoietic syndrome. In Vivo 25: 315–324, 2011. [PMC free article] [PubMed] [Google Scholar]
- 30.Jiang J, Belikova NA, Xiao J, Zhao Q, Greenberger JS, Wipf P and Kagan VE: A mitochondria-targeted nitroxide/ hemigramicidin S conjugate protects mouse embryonic cells against y-irradiation. Int J Radiat Oncol Biol Phys 70(3): 816–825, 2008. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 31.Rajagopalan MS, Gupta K, Epperly MW, Franicola D, Zhang X, Wang H, Zhao H, Tyurin V, Kagan VE, Wipf P, Kanai A and Greenberger JS: The mitochondria-targeted nitroxide JP4–039 augments potentially lethal irradiation damage repair. In Vivo 23: 717–725, 2009. [PMC free article] [PubMed] [Google Scholar]
- 32.Epperly MW, Goff J, Li S, Gao X, Wipf P, Wang H, Franicola D, Shen H, Rwigema J-CM, Kagan V, Bernard M and Greenberger JS: Intraesophageal administration of GS-nitroxide (JP4–039) protects against ionizing irradiation-induced esophagitis. In Vivo 24: 811–819, 2010. [PMC free article] [PubMed] [Google Scholar]
- 33.Rwigema J-CM, Beck B, Wang W, Doemling A, Epperly MW, Shields D, Franicola D, Dixon T, Frantz M-C, Wipf P, Tyurina Y, Kagan VE, Wang H and Greenberger JS: Two strategies for the development of mitochondrial-targeted small molecule radiation damage mitigators. Int J Radiat Oncol Biol Phys 80(3): 860–868, 2011. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 34.Guo H, Epperly MW, Bernarding M, Nie S, Gretton J, Jefferson M and Greenberger JS: Manganese superoxide dismutase-plasmid liposome (MnSOD-PL) intratracheal gene therapy reduction of irradiation-induced inflammatory cytokines does not protect orthotopic Lewis lung carcinomas. In Vivo 17: 13–22, 2003. [PubMed] [Google Scholar]
- 35.Guo HL, Seixas-Silva JA, Epperly MW, Gretton JE, Shin DM and Greenberger JS: Prevention of irradiation-induced oral cavity mucositis by plasmid/liposome delivery of the human manganese superoxide dismutase (MnSOD) transgene. Radiat Res 159: 361–370, 2003. [DOI] [PubMed] [Google Scholar]
- 36.Houghton AM, Rzymkiewicz DM, Ji H, Gregory AD, Egea EE, Metz HE, Stolz DB, Land SR, Marconcini LA, Kliment CR, Jenkins KM, Beaulieu KA, Mouded M, Frank SJ, Wong KK and Shapiro SD: Neutrophil elastase-mediated degradation of IRS-1 accelerates lung tumor growth. Nat Med 16(2): 219–223, 2010. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 37.Meuwissen R, Linn SC, van der Valk M, Mooi WJ and Berns A: Mouse model for lung tumorigenesis through Cre/Lox controlled sporadic activation of the K-Ras oncogene. Oncogene 20: 6551–6558, 2001. [DOI] [PubMed] [Google Scholar]
- 38.Fisher GH, Wellen SL, Klimstra D, Lenczowski JM, Tichelaar JW, Lizak MJ, Whitsett JA, Koretsky A and Varmus HE: Induction and apoptotic regression of lung adenocarcinomas by regulation of a K-Ras transgene in the presence and absence of tumor-suppressor genes. Genes Dev 15: 3249–3262, 2001. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 39.Johnson L, Mercer K, Greenbaum D, Bronson RT, Crowley D, Tuveson DA and Jacks T: Somatic activation of the K-ras oncogene causes early onset lung cancer in mice. Nature 410: 1111–1116, 2001. [DOI] [PubMed] [Google Scholar]
- 40.Tuveson DA and Jacks T: Modeling human lung cancer in mice: Similarities and shortcomings. Oncogene 18: 5318–5324, 1999. [DOI] [PubMed] [Google Scholar]
- 41.Jackson EL, Olive KP, Tuveson DA, Bronson R, Crowley D, Brown M and Jacks T: The differential effects of mutant p53 alleles on advanced murine lung cancer. Cancer Res 65(22): 10280–10288, 2005. [DOI] [PubMed] [Google Scholar]