

Abstract
CD4 T cells differentiate into RORγt/IL-17A-expressing cells in the small intestine following colonization by segmented filamentous bacteria (SFB). However, it remains unclear whether SFB-specific CD4 T cells can differentiate directly from naïve precursors, and whether their effector differentiation is solely directed towards the Th17 lineage. In this study, we used adoptive T cell transfer experiments and showed that naïve CD4 T cells can migrate to the small intestinal lamina propria (sLP) and differentiate into effector T cells that synthesize IL-17A in response to SFB colonization. Using single cell RT-PCR analysis, we showed that the progenies of SFB responding T cells are not uniform but composed of transcriptionally divergent populations including Th1, Th17 and follicular helper T cells. We further confirmed this finding using in vitro culture of SFB specific intestinal CD4 T cells in the presence of cognate antigens, which also generated heterogeneous population with similar features. Collectively, these findings indicate that a single species of intestinal bacteria can generate a divergent population of antigen-specific effector CD4 T cells, rather than it provides a cytokine milieu for the development of a particular effector T cell subset.
Keywords: antigen-specific CD4 T cells, germ-free mice, segmented filamentous bacteria, single cell RT-PCR, small intestine
INTRODUCTION
After birth, vertebrates are colonized by trillions of commensal bacteria in the gut. They provide a number of beneficial effects such as increasing efficiency of nutrition absorption, producing vitamins, providing barrier against pathogenic microbes, and promoting maturation of mucosal immunity (LeBlanc et al., 2017; Lee and Mazmanian, 2010). The role of commensal bacteria in regulating the mucosal immune system is of particular interest and increasing evidence indicates that commensal bacteria play a pivotal role in controlling innate and adaptive immune cells for homeostatic regulation and maturation of gut immunity (Benson et al., 2009; Macdonald and Monteleone, 2005; Maynard et al., 2012). However, in dysbiotic conditions, an immune response towards commensal bacteria can triggers life-threatening chronic inflammatory diseases, such as Crohn’s disease and ulcerative colitis (Feng et al., 2010; Lodes et al., 2004; Powrie et al., 1994). Therefore, understanding the mechanisms of cellular and molecular interactions between the host immune system and the gut microbiome is essential to maintain healthy condition.
Gnotobiotic technology has been developed to study the role of commensal bacteria in germ-free (GF) mice system, by which, we can define specific bacterial strains with particular immune modulatory functions (Atarashi et al., 2011; Ivanov et al., 2009; Maynard et al., 2012; Round and Mazmanian, 2010). For example, Bacteroides fragilis and Clostridium species elicit the generation of peripherally induced regulatory T cells (pTregs) by promoting TGFβ production, and segmented filamentous bacteria (SFB) induces Th17 T cells by inducing serum amyloid A (SAA) production in intestinal epithelial cells. It has been proposed that there are two steps for Th17 generation by SFB (Sano et al., 2015): first, dendritic cells presenting SFB-derived antigens migrate to mesenteric lymph nodes and prime antigen-specific naïve T cells to become RORγt-expressing cells; second, conversion of RORγt-expressing cells to RORγt/IL-17A-expressing cells happens in the small intestinal ileum, where attachment of SFB induces serum amyloid A (SAA) production. Thus, the generation of RORγt-expressing T cells require only an interaction with SFB-antigen bearing dendritic cells, whereas the generation of functional Th17 cells require additional inflammatory signals from local tissues. The Th17-inducing property of SFB has been of special interest as these intestinal T cells are important for mucosal defense against extracellular pathogens (Aujla et al., 2007), but also trigger autoimmune diseases under particular conditions (Wu et al., 2010). These findings suggest that specific strains of gut bacteria can induce a certain type of effector T cell by providing them with a polarizing cytokine environment.
It is unclear whether the lineage differentiation of intestinal CD4 T cells is a result of stimulation to a particular lineage of antigen-specific naïve precursor, or competition amongst various lineages. As the intestine is exposed to many diverse luminal antigens of commensal microbiota and dietary foods (Kim et al., 2016), it is possible that SFB also acts on recently activated T cells with unrelated environmental antigens. However, it was previously shown that SFB induces only antigen specific Th17 cells (Goto et al., 2014; Yang et al., 2014), although there is some degree of flexibility on the fate of mature CD4 T cells in general (Murphy and Stockinger, 2010). Furthermore, T cells specific to SFB can differentiate into RORγt-expressing cells even when host mice were bi-colonized with SFB and Th1-inducing Listeria monocytogens. Collectively, these findings suggest that SFB preferentially induces Th17 CD4 T cells in the intestine. However, previous study focused on only a limited number of SFB-specific T cell clones (Yang et al., 2014) and it remains unclear whether SFB induces single type of intestinal CD4 T cell.
In the present study, we adoptively transferred naïve CD4 T cells to SFB monocolonized GF B6 mice and examined their fates at single cell level. Consistent with previous reports, (Ivanov et al., 2009), we found that SFB-monocolonization leads to the generation of mainly Th17 cells. We further analyzed these cells using single cell RT-PCR of 95 genes, and found that there is substantial heterogeneity within the SFB-induced intestinal CD4 T cells derived from naïve precursors. By co-culturing them with antigen-presenting cells (APC) loaded with SFB antigens, we further revealed that the in vitro re-stimulated SFB-specific T cells mimic the gene expression profiles of the in vivo SFB reactive T cells. Collectively, our data show that SFB colonization of the small intestine leads to the generation of transcriptionally diverse intestinal CD4 T cells derived from naïve precursors.
MATERIALS AND METHODS
Mice
Germ-free C57BL/6 (B6) mice were kindly provided by Drs. Andrew Macpherson (Bern Univ., Switzerland) and David Artis (Univ. Pennsylvania, USA) and maintained in sterile flexible film isolators (Class Biological Clean Ltd., USA). Specific pathogen-free (SPF) B6 mice and CD90.1 B6 mice were purchased from the Jackson Laboratory, and maintained in the animal facility of POSTECH Biotech Center. SPF Foxp3-GFP mice were a gift from Talal Chatila (Boston Children’s hospital) and bred onto CD90.1 B6 background. Mouse care and experimental procedures were performed in accordance with all institutional guidelines for the ethical use of non-human animals in research protocols approved by the Institutional Animal Care and Use Committees (IACUC) of the Pohang University of Science and Technology.
SFB colonization
SFB (or Arthromitus immunis) was kindly provided by Dr. Yoshinori Umesaki (Yakult Cent Inst, Japan) (Umesaki et al., 1999) and was maintained in monocolonized GF mice at POSTECH. For SFB colonization, fecal pellets from SFB-monocolonized mice were resuspended in sterile PBS and orally gavaged into GF mice. For a control group fecal pellets of GF mice were used.
Scanning electron microscopy
Scanning electron microscopy (SEM) was performed on ~1 cm pieces from terminal ileum. Tissues were extensively washed with PBS and fixed with 2% glutaraldehype (sigma) for 2 h at room temperature. Thereafter, tissues were post-fixed with 1% osmium tetroxide (sigma) for 1 h followed by 3 times wash with PBS. A series of ethanol (sigma) concentrations in distilled water was used for dehydration. Ethanol was completed removed by Critical-point dryer.
Cell isolation from tissues
Single cell suspensions were prepared from LN and spleen. To isolate T cells in small intestine, tissues were cut into small pieces after Peyer’s patches were removed. Chopped tissues were incubated for 30 minutes at 37°C in PBS buffer containing fetal bovine serum (FBS, 3% vol/vol), EDTA (10 mM), HEPES (20 mM), polymyxin B (10 mg/ml), penicillin (100 U/ml), streptomycin (100 mg/ml) and sodium pyruvate (1 mM) to remove epithelial cells. Tissues were extensively washed with PBS and digested with collagenase D and Dnase I in RPMI medium containing FBS (3% vol/vol), HEPES (20 mM), penicillin (100 U/ml), streptomycin (100 mg/ml), sodium pyruvate (1 mM) and non-essential amino acids (1 mM). Cell suspensions were enriched by 40:75% Percoll density gradient centrifugation.
Adoptive transfer
For naïve CD4 T cells, CD44loCD62LhiGFP− cells from CD90.1+ Foxp3-GFP reporter mice were purified by flow cytometry sorting. After separation, donor T cells were labelled with CellTrace Violet (CTV) as previously described (Martin et al., 2013). Varying doses of T cells were adoptively transferred by retro-orbital injection. For GF, donor T cells were manipulated aseptically, inserted into autoclaved amber glass vials capped with a silicone septum and transferred into GF isolaters after spraying with 2% peracetic acids.
Antibodies and staining reagents for flow cytometry and sorting
Isolated cells were stained with propidium iodide (Sigma) or Ghost viability dye (Tonbo) to eliminate dead cells. For surface staining, cells were stained with the following fluorochrome-conjugated antibodies (eBioscience, Biolegend): anti-CD4 (RM4-5), anti-Thy1.1 (HIS51), anti-CD44 (IM7), anti-CD62L (MEL-14). For intracellular staining, surface stained cells were fixed and permeabilized with a Foxp3 staining kit (eBioscience) according to manufacturer’s instruction and stained with the following antibodies: anti-Foxp3 (FJK-16s), anti-IFNγ (XMG1.2), anti-IL17A (eBio17/37). For intracellular staining of cytokines, isolated cells were cultured for 4 h in the presence of PMA/Ionomycin with protein transport inhibitors (eBioscience). Data from the stained cells were collected with LSR Forteassa with DIVA software (BD Bioscience) and were analyzed by FlowJo (TreeStar), or were sorted with Moflo-XDP (Beckman Coulter) for other experiments.
In vitro intestinal CD4 T cell proliferation with fecal antigens
Splenic APCs (5 × 105) was co-cultured for 4 days with 5 × 104 CTV-labeled purified sLP CD4 T cells from GF or GF mice mono-colonized with SFB. For fecal antigen preparation, we slightly modified protocol of a previous report (Goto et al., 2014). Briefly, 3 grams of fecal pellets from GF or SFB-monocolonized mice were homogenized in 10 ml PBS. Fecal suspensions were autoclaved and debris were removed by centrifugation at 3200g. Supernatant was used at 1:400 dilution for T cell culture.
Single cell RT-PCR
As previously described (Sanchez-Freire et al., 2012), single cell RT-PCR was performed by using Fluidigm C1 and Biomark. Single cells were captured in individual well and single cell content was manually confirmed under the light microscope. Lysis, reverse transcription and pre-amplification were performed thereafter. RT-PCR data were collected in Biomark. Heat map comparison followed with Euclidean distance clustering analysis was conducted using Morpheus (https://software.broadinstitute.org/morpheus/). tSNE analysis was conducted using FlowJo v10.
Statistical analyses
Mean and S.D. values were calculated by using Prism 5 (GraphPad Sotfware). Statistical significance between two variables was determined by unpaired two-tails t tests. P-values less than 0.05 were considered as significant.
RESULTS
Diverse subsets of SFB-induced intestinal CD4 T cells develop from naïve precursors
SFB colonization leads to Th17 cell generation in the small intestine. To test whether naïve CD4 T cells can directly differentiate into Th17 cells upon SFB infection, we adoptively transferred GF B6 mice with CellTrace violet (CTV)-labeled purified naïve (CD44lo CD62Lhi) CD4 T cells from Thy1.1 Foxp3-GFP mice, followed by oral gavage of fecal suspensions from GF or SFB-monocolonized mice. At 14 d post-transfer, approximately 10% of donor T cells diluted out the CTV in the mesenteric lymph nodes (MLN) from both GF and SFB-monocolonized mice (Fig. 1A). Most of the donor T cells in the sLP were CTVneg, suggesting they encounter the antigens in the MLN and migrate into the sLP after extensive proliferation (Fig. 1B). Interestingly, there was no significant difference between frequencies of CTVneg donor CD4 T cells transferred into GF and SFB monocolonized hosts. In the GF condition, it might reflect homeostatic proliferation of naïve T cells upon response to innocuous self-antigens or dietary components. When we analyzed the cytokine production, only donor T cells transferred into SFB monocolonized mice produced IL-17A in the MLN and sLP (Fig. 1C). IFNγ producing cells, however, were not different probably because homeostatic expansion also induce it (Kawabe et al., 2017). Together, our data indicate that a small frequency of naïve CD4 T cells can undergo extensive proliferation and migrate into the local mucosal compartment, where they differentiate into Th17 cells in response to SFB antigens. We also confirmed this monocolonization using electron microscopy, as described previously (Ivanov et al., 2009; Klaasen et al., 1991; Umesaki et al., 1999)(Fig. 1D).
Fig. 1. SFB monocolonization induces effector T cells from a fraction of naïve CD4 T cells.
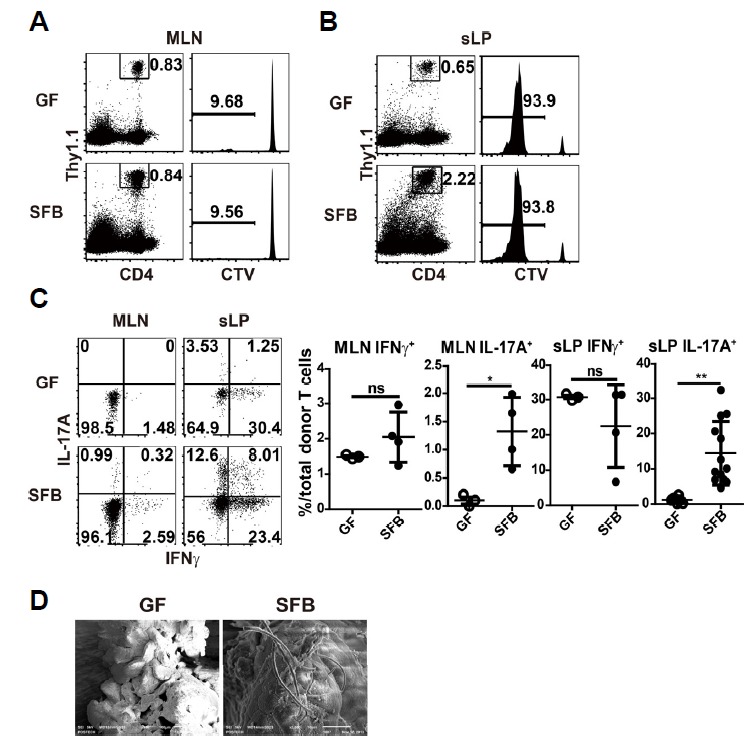
(A to C) Germ free (GF) B6 hosts were adoptively transferred with 1 × 107 CellTrace Violet (CTV)-labeled CD44lo CD62Lhi Foxp3-GFPneg CD4 T cells from Thy1.1 Foxp3-GFP mice. One day later, mice were SFB-monocolonized and analyzed 13 d later. Representative dot plots show frequencies of donor T cells; histograms show dilution of CTV in mesenteric lymph nodes (MLN) (A) and small intestinal lamina propria (sLP) (B). (C) Dot plots show IFNγ and IL-17A producing donor CD4 T cells after 4 h ex vivo re-stimulation with PMA/ionomycin in the presence of protein transport inhibitor (left). Graphs show the frequencies of indicated cytokine secreting cells (right). (D) Scanning electron microscopy (SEM) images showing SFB colonization in small intestinal ileum. Numbers indicate frequency of cells in adjacent gates or in each quadrant. Each dot represents an individual mouse and horizontal bars indicate mean values. P values were determined by Student’s t-test. Error bars show the mean ± SD. *P < 0.05; **P < 0.01. SFB, segmented filamentous bacteria.
According to previous reports, SFB colonization induces mainly Th17 cells, but not Th1 and pTregs (Geuking et al., 2011). However, we thought that analyzing SFB-induced T cells at the single cell level rather than at the population level might reveal some differences. To this end, we isolated a total of 95 donor derived CTVneg single cells from sLP of GF (47 cells) and SFB monocolonized GF (48 cells) host mice, and performed single cell RT-PCR using 98 primer sets that encompass various transcription factors and cytokine genes, as described previously (Sanchez-Freire et al., 2012). After normalization with three housekeeping genes, we generated a heat map for all 95 genes, and Euclidean-distance clustering analysis was performed (Fig. 2). We found that donor T cells isolated from SFB-monocolonized hosts consisted of three discrete clusters. For further in-depth analysis, the data were analyzed using t-distributed stochastic neighbor embedding (tSNE) algorithm. This analysis confirmed that SFB-reactive T cells are distributed into three distinct populations whereas T cells from the GF host were separately clustered (Fig. 3A). SFB-reactive T cell subset A (SFB A, ~35%) comprised of major Tbx21+Rora+Il22+ cells and minor populations of cells expressing Bcl6 or Il17b; SFB-reactive T cell subset B (SFB B, approximately 10%) comprised Tbx21+ Bcl6+ Rora+ Il22+ cells; and SFB-reactive T cell subset C (SFB C, approximately 52%) comprised Il22+Il17b+ cells (Figs. 3A and 3B). Of 95 genes tested, Notch1 most reliably separated SFB reactive T cells from those isolated form GF host (Fig. 3B). Collectively, these results show that we defined a diverse population of SFB-induced T cells, which could not be detected in previous analysis using flow cytometry.
Fig. 2. Single cell RT-PCR reveals heterogeneity of CD4 T cells generated by SFB-colonization.
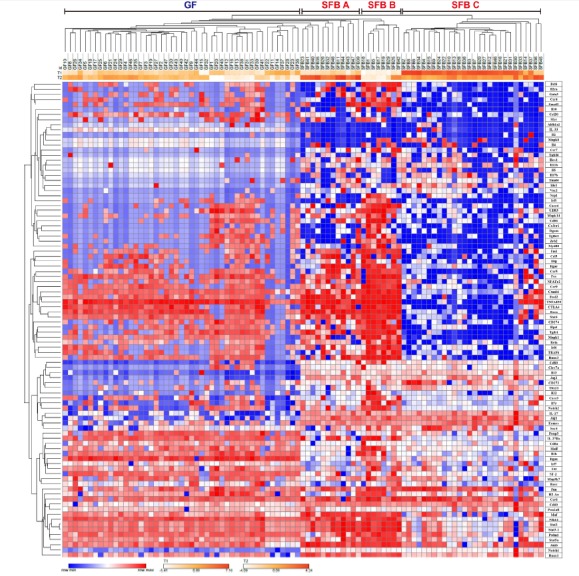
mRNA was extracted from 47 GF and 48 SFB-monocolonized single CTVneg donor T cells at 14 d post-injection in the small intestinal lamina propria as in Fig. 1. RT-PCR for 98 genes was performed, including the three housekeeping genes. Hierarchical clustering of isolated single T cells is shown for 95 genes with −ΔCT values.
Fig. 3. tSNE analysis of in vivo generated SFB reactive CD4 T cells.
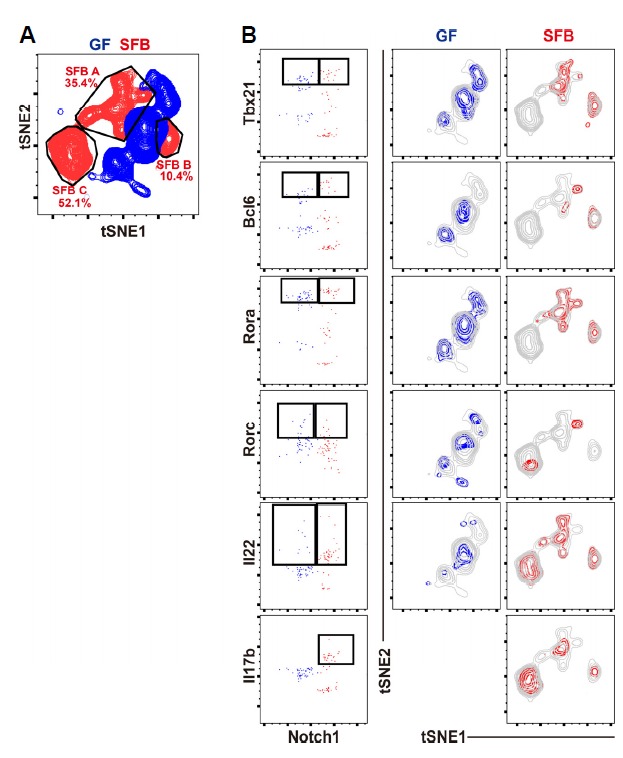
Data of Fig. 2 were further analyzed with FlowJo v10. t-distributed stochastic neighbor embedding (tSNE) analysis for 95 genes with −ΔCT values. (A) Distribution of donor T cells from GF (blue) or SFB-monocolonized (red) mice. (B) Distribution of single cells according to Tbx21, Bcl6, Rora, Rorc, Il22, or Il17b expression in the two-dimensional space.
SFB-specific intestinal CD4 T cells consist of heterogeneous subsets
The above-described single cell RT-PCR approach still had some limitations because we were not able to discriminate SFB-specific effector T cells from non-specifically proliferating T cells that were seen in the GF host (Fig. 1B). SFB-specific TCR transgenic mice and a SFB-specific MHCII tetramer (Yang et al., 2014) might be useful here, but these reagents also have limitations, as they do not represent all SFB reactive T cells. To resolve this issue, we generated SFB-specific CD4 T cells in vitro by co-culturing T-depleted splenic APCs loaded with SFB antigens and CTV-labeled intestinal CD4 T cells isolated from SFB-monocolonized mice (Fig. 4). This approach was previously described (Goto et al., 2014), but the only parameter they analyzed was IL-17A from proliferated SFB-specific CD4 T cells. As expected, CD4 T cells isolated from the sLP of SFB-monocolonized mice showed strong proliferation and increased granularity in response to cognate SFB fecal antigens, but not to GF fecal antigens (Fig. 4A). In contrast, intestinal CD4 T cells from GF mice showed a very low proliferative response against fecal antigens from both mice, although an anti-CD3 antibody was able to induce robust proliferation. Interestingly, intestinal CD4 T cells from SFB-monocolonized mice showed stronger proliferation than T cells from GF mice when stimulated with an anti-CD3 antibody. This could imply that intestinal CD4 T cells from SFB-monocolonized mice are in a higher activation state than those from GF mice due to an inflammatory response after SFB colonization.
Fig. 4. SFB colonization-induced intestinal CD4 T cells undergo robust proliferation in response to SFB fecal antigens.
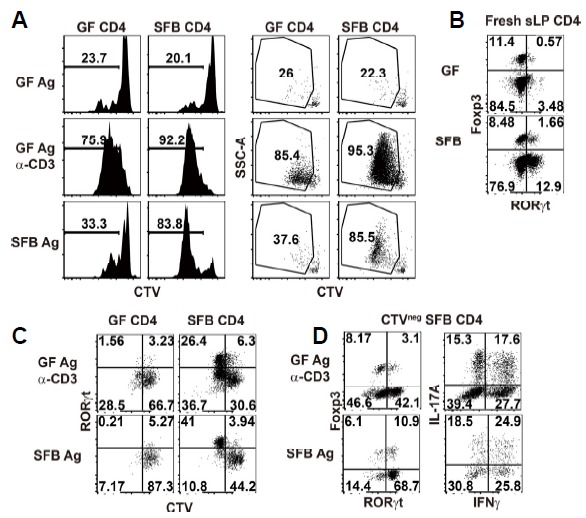
CellTrace Violet (CTV)-labeled small intestinal lamina propria (sLP) CD4 T cells from either GF- or SFB-monocolonized B6 mice were cultured. (A) Histograms show CTV-dilution profile of co-cultured CD4 T cells in the presence of GF fecal antigen, GF fecal Ag plus anti-CD3 antibody (2 μg/ml), or SFB fecal antigen. Dot plots show CTV-dilution and SSC pattern. (B) Dot plots showing RORγt and Foxp3 expression in sLP CD4 T cells from GF B6 mice (top) or GF B6 mice monocolonized with SFB 2 weeks before (bottom) (C and D). Dot plots showing phenotype of co-cultured T cells after proliferation in the presence of GF fecal Ag plus anti-CD3 antibody (2 μg/ml) or SFB fecal antigen alone. (C) CTV dilution profile and RORγt expression in co-cultured T cells. (D) Foxp3 and RORγt expression in CTVneg SFB-specific T cells (left). IFNγ and IL-17A expression in CTVneg SFB-specific T cells after 4 h re-stimulation with PMA/ionomycin in the presence of protein transport inhibitors (right). Numbers indicate frequency of cells in adjacent gates or in each quadrant. Representative data from at least two independent experiments are shown.
Next, we sought to determine additional phenotypes of the highly proliferative SFB-specific intestinal CD4 T cells under in vitro conditions. Consistent with prior findings (Atarashi et al., 2015; Sano et al., 2015), SFB-colonization clearly induced RORγt-expressing CD4 T cells in the small intestine (Fig. 4B). Upon in vitro stimulation with SFB fecal antigens, RORγt+ cells preferentially proliferated and these cells represented approximately 80% of the total CTVneg co-cultured T cells (Figs. 4C and 4D). This implies that most SFB-specific CD4 T cells are RORγt+ cells. Interestingly, however, not all of these cells were IL-17A-synthesizing cells, and instead were composed of various subsets that secreted IFNγ and/or IL-17A after 4 h stimulation with PMA/ionomycin in the presence of protein transport inhibitors (Fig. 4D). Thus, we were able to detect at least three distinct populations of SFB-specific CD4 T cells: IFNγ+, IFNγ+ IL-17A+, and IL-17A+ T cells.
We further analyze these SFB-specific CD4 T cells using a multiplexed single cell RT-PCR approach. CTVneg SSC-Ahi co-cultured T cells (45 cells) were sorted from the SFB CD4 plus SFB Ag condition shown in Fig. 4A (Fig. 5A). Including housekeeping genes, 48 genes were analyzed in SFB-specific CD4 T cells at the single cell level and ΔCT values were calculated by comparing the CT value for each gene with that of Gapdh, as shown in Fig. 2. Consistent with the flow cytometry data from in vitro co-culture assays (Fig. 4D), Euclidean distance clustering analysis with 46 genes revealed heterogeneity within the SFB-specific individual T cells. Although there were a number of homogeneously expressed genes, many showed distinct expression patterns within specific clusters (Fig. 5B and Fig. 6). Rora, Runx1, Runx3, and Prdm1 were induced in most cells and Rorc was induced in approximately 80% of cells, which correlates with our flow cytometry data (Fig. 4D). Other genes were heterogeneously expressed in each cluster. These in vitro re-stimulated SFB-specific T cells were clustered together with in vivo reactive SFB A and B subsets both in Euclidean-distance clustering (Fig. 5B) and tSNE analysis (Fig. 6A). These data indicate that in vitro re-stimulated SFB reactive T cells have a similar transcriptional profile to those of in vivo generated SFB reactive T cells, suggesting the SFB A and SFB B populations shown in Figs. 2 and 3 are more likely to represent actual SFB reactive T cells, rather than the homeostatic population observed in GF mice.
Fig. 5. In vitro re-stimulated SFB-specific CD4 T cells and in vivo SFB primed CD4 T cells have transcriptional similarity.
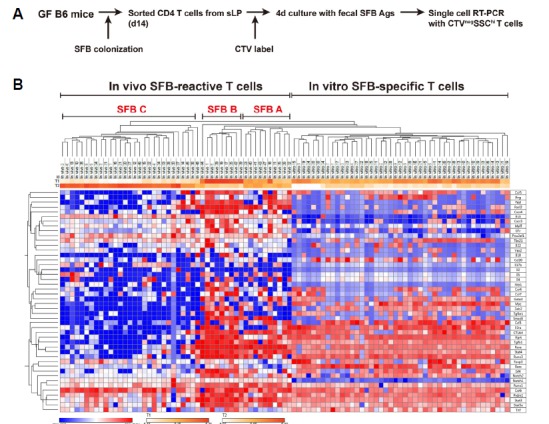
mRNA was extracted from CTVneg SSC-Ahi 45 single T cells after 4 d of in vitro stimulation with 5 × 105 T-depleted splenocytes loaded with SFB antigen as in Fig. 4A. RT-PCR was performed using primers for 46 target genes. (A) Schematic diagram of experimental strategy. (B) Hierarchical clustering shows in vitro activated SFB-specific CD4 T cells together with in vivo activated SFB reactive T cells from Fig. 2.
Fig. 6. Combined tSNE analysis of in vivo and in vitro generated SFB reactive CD4 T cells.
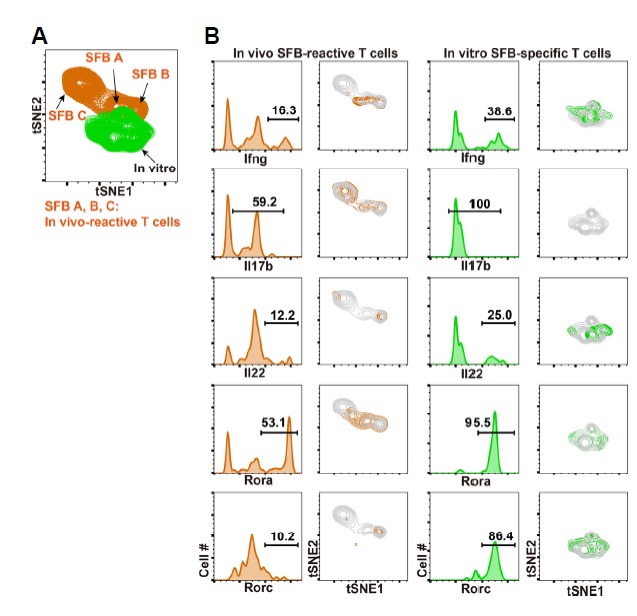
Single cell qPCR data from Fig. 5 were further analyzed with t-distributed stochastic neighbor embedding (tSNE) method for 46 genes with −ΔCT values. (A) Distributions of in vivo SFB-reactive T cells (In vivo A, B, and C in orange) and in vitro re-stimulated SFB-specific T cells (green) are shown. (B) Distributions of in vivo SFB-reactive T cells and in vitro SFB-specific T cells according to expression patterns of Ifng, Il17b, Il22, Rora, and Rorc in the two-dimensional space are shown.
DISCUSSION
In this paper, we used a microfluidic platform to evaluate gene expression profiles of SFB reactive T cells at the single cell level (Sanchez-Freire et al., 2012) and provided evidence that transcriptionally divergent populations of antigen-specific CD4 T cells are induced in the small intestine upon SFB colonization. Previous studies showed that SFB induces mainly a Th17 response (Goto et al., 2014; Ivanov et al., 2009) and we also showed that 80% of SFB-specific CD4 T cells express RORγt. Studies using SFB-specific TCR transgenic mice and a SFB specific pMHCII tetramer further confirmed that SFB induces only RORγt expressing CD4 T cells, but not IL-17A producing cells (Yang et al., 2014). However, previous studies did not address this issue on a single cell level. In the current study, we found that SFB induces at least two different types of effector T cells composed of Th17 and Th1 cells and possibly T follicular helpers (Tfh) expressing Bcl6. Although we could not analyze additional Tfh markers such as CXCR5 and PD1, the presence of Tfh cells is consistent with previous observation that SFB induces Tfh cells promoting autoantibody production (Teng et al., 2016).
With single cell RT-PCR analysis followed by tSNE analysis, we were able to evaluate the transcriptional profile of SFB reactive CD4 T cells at the single cell level. This method enabled us to find previously unrecognized heterogeneity of SFB reactive CD4 T cells in the small intestine. One may argue that our findings using in vivo SFB-reactive T cells could be a result from stochastic event during complex process of T cell differentiation (Kelso, 1999; Yoon et al., 2018). However, we additionally showed that in vitro SFB re-stimulated T cells have such a heterogeneity with substantial similarity (Fig. 6). Based on these observations, we concluded that SFB colonization induces transcriptionally divergent effector CD4 T cell subsets.
Previous reports showed that Th17 cells have substantial lineage plasticity and they can redirect their differentiation program to IFNγ producing Th1 cells by IL-23 (Basdeo et al., 2017; Hirota et al., 2011). Therefore, it is possible that SFB reactive non-Th17 CD4 T cells defined in our study is actually exTh17 cells, and some of our data supports this idea. First, Notch1, an essential component of Th17 differentiation (Keerthivasan et al., 2011), was constitutively upregulated in all SFB reactive T cells (Fig. 3). Second, Rora, which enhances Th17 differentiation program (Yang et al., 2008), and IL22, a cytokine secreted from Th17 cells, were co-expressed in Tbx21 expressing SFB A and SFB B populations. Third, 25% of IFNγ+ cells of SFB-specific CD4 T cells (Fig. 1C and data not shown), expressed Rorγt. These features suggest that substantial fraction of SFB-specific CD4 T cells share both Th1 and Th17 properties.
Compared to conventional Th1 or Th17 cells, ex-Th17 non-canonical Th1 cells were more proliferative and resistant to suppression by Tregs, and secreted more pro-inflammatory cytokines (Basdeo et al., 2017; Hirota et al., 2011). In humans, these cells were enriched in the joints of rheumatoid arthritis patients, suggesting ex-Th17 cells may play significant roles on its pathogenesis. Indeed, Th17-inducing Prevotella copri is abundantly found at early RA patients and microbiota from human RA patient induced IL17 dependent arthritis in GF SKG mice (Maeda and Takeda, 2017). However, in mice system, it was not clear whether SFB induces pathogen specific Th1 cells and how they can contribute to inflammatory disorders. For the future study, we plan to use IL17 fate-mapping reporter mice (Hirota et al., 2011) to see if IFNγ producing Th1 cells or Bcl6 positive CD4 T cells in SFB colonized mice were exTh17 cells. The results of this study will clarify the origin of these cells and fill the knowledge gaps between mouse and human studies.
ACKNOWLEDGMENTS
We thank A. Macpherson, K. McCoy, and D. Artis for generously providing various strains of germ-free mice; Tae-Kyu Kim, Jung-Wook Seo, Sang-Sik Kim, Haejin Jung, and Mi-Ok Lee for technical help. This work was supported by project IBS-R005-D1 of the Institute for Basic Science, Korean Ministry of Science, Information/Communication Technology and Future Planning.
REFERENCES
- Atarashi K., Tanoue T., Ando M., Kamada N., Nagano Y., Narushima S., Suda W., Imaoka A., Setoyama H., Nagamori T., et al. Th17 cell induction by adhesion of microbes to intestinal epithelial cells. Cell. 2015;163:367–380. doi: 10.1016/j.cell.2015.08.058. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Atarashi K., Tanoue T., Shima T., Imaoka A., Kuwahara T., Momose Y., Cheng G., Yamasaki S., Saito T., Ohba Y., et al. Induction of colonic regulatory T cells by indigenous Clostridium species. Science. 2011;331:337–341. doi: 10.1126/science.1198469. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Aujla S.J., Dubin P.J., Kolls J.K. Th17 cells and mucosal host defense. Semin Immunol. 2007;19:377–382. doi: 10.1016/j.smim.2007.10.009. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Basdeo S.A., Cluxton D., Sulaimani J., Moran B., Canavan M., Orr C., Veale D.J., Fearon U., Fletcher J.M. Ex-Th17 (nonclassical Th1) cells are functionally distinct from classical Th1 and Th17 cells and are not constrained by regulatory T cells. J Immunol. 2017;198:2249–2259. doi: 10.4049/jimmunol.1600737. [DOI] [PubMed] [Google Scholar]
- Benson A., Pifer R., Behrendt C.L., Hooper L.V., Yarovinsky F. Gut commensal bacteria direct a protective immune response against Toxoplasma gondii. Cell Host Microbe. 2009;6:187–196. doi: 10.1016/j.chom.2009.06.005. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Feng T., Wang L., Schoeb T.R., Elson C.O., Cong Y. Microbiota innate stimulation is a prerequisite for T cell spontaneous proliferation and induction of experimental colitis. J Exp Med. 2010;207:1321–1332. doi: 10.1084/jem.20092253. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Geuking M.B., Cahenzli J., Lawson M.A., Ng D.C., Slack E., Hapfelmeier S., McCoy K.D., Macpherson A.J. Intestinal bacterial colonization induces mutualistic regulatory T cell responses. Immunity. 2011;34:794–806. doi: 10.1016/j.immuni.2011.03.021. [DOI] [PubMed] [Google Scholar]
- Goto Y., Panea C., Nakato G., Cebula A., Lee C., Diez M.G., Laufer T.M., Ignatowicz L., Ivanov II. Segmented filamentous bacteria antigens presented by intestinal dendritic cells drive mucosal Th17 cell differentiation. Immunity. 2014;40:594–607. doi: 10.1016/j.immuni.2014.03.005. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Hirota K., Duarte J.H., Veldhoen M., Hornsby E., Li Y., Cua D.J., Ahlfors H., Wilhelm C., Tolaini M., Menzel U., et al. Fate mapping of IL-17-producing T cells in inflammatory responses. Nat Immunol. 2011;12:255–263. doi: 10.1038/ni.1993. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Ivanov II, Atarashi K., Manel N., Brodie E.L., Shima T., Karaoz U., Wei D., Goldfarb K.C., Santee C.A., Lynch S.V., et al. Induction of intestinal Th17 cells by segmented filamentous bacteria. Cell. 2009;139:485–498. doi: 10.1016/j.cell.2009.09.033. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kawabe T., Jankovic D., Kawabe S., Huang Y., Lee P.H., Yamane H., Zhu J., Sher A., Germain R.N., Paul W.E. Memory-phenotype CD4(+) T cells spontaneously generated under steady-state conditions exert innate TH1-like effector function. Sci Immunol. 2017;2:eaam9304. doi: 10.1126/sciimmunol.aam9304. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Keerthivasan S., Suleiman R., Lawlor R., Roderick J., Bates T., Minter L., Anguita J., Juncadella I., Nickoloff B.J., Le Poole I.C., et al. Notch signaling regulates mouse and human Th17 differentiation. J Immunol. 2011;187:692–701. doi: 10.4049/jimmunol.1003658. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kelso A. Educating T cells: early events in the differentiation and commitment of cytokine-producing CD4+ and CD8+ T cells. Springer Semin Immunopathol. 1999;21:231–248. doi: 10.1007/BF00812255. [DOI] [PubMed] [Google Scholar]
- Kim K.S., Hong S.W., Han D., Yi J., Jung J., Yang B.G., Lee J.Y., Lee M., Surh C.D. Dietary antigens limit mucosal immunity by inducing regulatory T cells in the small intestine. Science. 2016;351:858–863. doi: 10.1126/science.aac5560. [DOI] [PubMed] [Google Scholar]
- Klaasen H.L., Koopman J.P., Van den Brink M.E., Van Wezel H.P., Beynen A.C. Mono-association of mice with non-cultivable, intestinal, segmented, filamentous bacteria. Arch Microbiol. 1991;156:148–151. doi: 10.1007/BF00290989. [DOI] [PubMed] [Google Scholar]
- LeBlanc J.G., Chain F., Martin R., Bermudez-Humaran L.G., Courau S., Langella P. Beneficial effects on host energy metabolism of short-chain fatty acids and vitamins produced by commensal and probiotic bacteria. Microb Cell Fact. 2017;16:79. doi: 10.1186/s12934-017-0691-z. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Lee Y.K., Mazmanian S.K. Has the microbiota played a critical role in the evolution of the adaptive immune system? Science. 2010;330:1768–1773. doi: 10.1126/science.1195568. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Lodes M.J., Cong Y., Elson C.O., Mohamath R., Landers C.J., Targan S.R., Fort M., Hershberg R.M. Bacterial flagellin is a dominant antigen in Crohn disease. J Clin Invest. 2004;113:1296–1306. doi: 10.1172/JCI20295. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Macdonald T.T., Monteleone G. Immunity, inflammation, and allergy in the gut. Science. 2005;307:1920–1925. doi: 10.1126/science.1106442. [DOI] [PubMed] [Google Scholar]
- Maeda Y., Takeda K. Role of gut microbiota in rheumatoid arthritis. J Clin Med. 2017;6:60. doi: 10.3390/jcm6060060. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Martin C.E., Frimpong-Boateng K., Spasova D.S., Stone J.C., Surh C.D. Homeostatic proliferation of mature T cells. Methods Mol Biol. 2013;979:81–106. doi: 10.1007/978-1-62703-290-2_9. [DOI] [PubMed] [Google Scholar]
- Maynard C.L., Elson C.O., Hatton R.D., Weaver C.T. Reciprocal interactions of the intestinal microbiota and immune system. Nature. 2012;489:231–241. doi: 10.1038/nature11551. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Murphy K.M., Stockinger B. Effector T cell plasticity: flexibility in the face of changing circumstances. Nat Immunol. 2010;11:674–680. doi: 10.1038/ni.1899. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Powrie F., Leach M.W., Mauze S., Menon S., Caddle L.B., Coffman R.L. Inhibition of Th1 responses prevents inflammatory bowel disease in scid mice reconstituted with CD45RBhi CD4+ T cells. Immunity. 1994;1:553–562. doi: 10.1016/1074-7613(94)90045-0. [DOI] [PubMed] [Google Scholar]
- Round J.L., Mazmanian S.K. Inducible Foxp3+ regulatory T-cell development by a commensal bacterium of the intestinal microbiota. Proc Natl Acad Sci USA. 2010;107:12204–12209. doi: 10.1073/pnas.0909122107. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Sanchez-Freire V., Ebert A.D., Kalisky T., Quake S.R., Wu J.C. Microfluidic single-cell real-time PCR for comparative analysis of gene expression patterns. Nat Protoc. 2012;7:829–838. doi: 10.1038/nprot.2012.021. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Sano T., Huang W., Hall J.A., Yang Y., Chen A., Gavzy S.J., Lee J.Y., Ziel J.W., Miraldi E.R., Domingos A.I., et al. An IL-23R/IL-22 Circuit Regulates Epithelial Serum Amyloid A to Promote Local Effector Th17 Responses. Cell. 2015;163:381–393. doi: 10.1016/j.cell.2015.08.061. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Teng F., Klinger C.N., Felix K.M., Bradley C.P., Wu E., Tran N.L., Umesaki Y., Wu H.J. Gut microbiota drive autoimmune arthritis by promoting differentiation and migration of peyer’s patch T follicular helper cells. Immunity. 2016;44:875–888. doi: 10.1016/j.immuni.2016.03.013. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Umesaki Y., Setoyama H., Matsumoto S., Imaoka A., Itoh K. Differential roles of segmented filamentous bacteria and clostridia in development of the intestinal immune system. Infect Immun. 1999;67:3504–3511. doi: 10.1128/iai.67.7.3504-3511.1999. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Wu H.J., Ivanov II, Darce J., Hattori K., Shima T., Umesaki Y., Littman D.R., Benoist C., Mathis D. Gut-residing segmented filamentous bacteria drive autoimmune arthritis via T helper 17 cells. Immunity. 2010;32:815–827. doi: 10.1016/j.immuni.2010.06.001. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Yang X.O., Pappu B.P., Nurieva R., Akimzhanov A., Kang H.S., Chung Y., Ma L., Shah B., Panopoulos A.D., Schluns K.S., et al. T helper 17 lineage differentiation is programmed by orphan nuclear receptors ROR alpha and ROR gamma. Immunity. 2008;28:29–39. doi: 10.1016/j.immuni.2007.11.016. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Yang Y., Torchinsky M.B., Gobert M., Xiong H., Xu M., Linehan J.L., Alonzo F., Ng C., Chen A., Lin X., et al. Focused specificity of intestinal TH17 cells towards commensal bacterial antigens. Nature. 2014;510:152–156. doi: 10.1038/nature13279. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Yoon B.H., Kim M., Kim M.H., Kim H.J., Kim J.H., Kim J.H., Kim J., Kim Y.S., Lee D., Kang S.J., et al. Dynamic transcriptome, DNA methylome, and DNA hydroxymethylome networks during T-cell lineage commitment. Mol Cells. 2018;41:953–963. doi: 10.14348/molcells.2018.0213. [DOI] [PMC free article] [PubMed] [Google Scholar]