

Abstract
Information on genetic diversity and population structure are very important in any breeding programme for the improvement of traits of interest and the development of outstanding products for commercialization. In the present study, we assessed the genetic diversity of 94 early-maturing white and yellow tropical maize inbred lines using single nucleotide polymorphism (SNP) markers. The larger number of SNP markers used in this study allowed a clearer inference of the population structure of the 94 inbred lines. Cluster analysis resolved the inbred lines into different clusters based on their pedigree, selection history and endosperm colour. However, three heterotic groups were revealed by population structure analysis, but additional field evaluation could be more informative to confirm the heterotic groups identified. Nevertheless, wide genetic variability existed among the inbred lines making them unique with the potential to contribute new beneficial alleles to maize breeding programmes in the tropics, especially in the West and Central Africa (WCA) sub-region.
Introduction
For good progress from selection in any crop improvement programme, information on the genetic diversity and population structure of the base germplasm is crucial. Therefore, plant breeders routinely resort to newly available tools to make informed decisions on selection. In the past, a number of researchers have highlighted the importance and the need for accurate assessment of genetic diversity in applied breeding programmes. This allows the examination of issues relating to the assignment of inbred lines to heterotic groups, selection of efficient testers for testing of inbred lines in hybrid combinations, drawing inferences on the introgression of desirable genes from diverse germplasm sources into available genetic base, identification of diverse parental combinations to create segregating progenies with maximum genetic variability for further selection and the estimation of genetic diversity loss during conservation or selection [1–6]. Studies of the early and extra-early maize germplasm in sub-Saharan Africa (SSA) have shown conclusively that genetic diversity is of primary importance in the display of heterosis. Inbred lines from different heterotic groups produced higher-yielding hybrids than those of lines from within the same heterotic group [7]. Therefore, the assessment of genetic diversity within and between plant populations is routinely carried out via different marker techniques such as morphological, biochemical and molecular markers [8,9].
DNA based markers are preferred over morphological and biochemical markers because they are not affected by environmental factors and/or by the developmental stage of the plant [8,10]. Consequently, DNA markers have been an indispensable tool for characterizing genetic resources and providing breeders with more detailed information to assist in selecting diverse parents [11]. In the past, various molecular markers including simple sequence repeats (SSRs) have been extensively used in maize to study the correlation between genetic distance and hybrid performance, heterosis, combining ability and assign inbred lines into heterotic groups [12–17]. For example, forty-two early maturing inbred lines were genotyped by Oyekunle, Badu-Apraku, Hearne, and Franco [17] using 23 SSR markers and grouped them into five heterotic groups. The information generated also provided a better understanding of the genetic relationships among the early-maturing inbred lines and facilitated more effective utilization of the inbred lines in the breeding programme for the development of synthetic varieties and hybrids, and formation of heterotic populations used to derive promising inbred lines. However, the choice of either one or the other marker type, the number of loci, the cost of marker as well as the purpose of the study and the evolutionary history of the populations under investigation play a major role [18].
With the advances in marker technology, single nucleotide polymorphism (SNP) markers have become the choice due to their low cost per data point, high genomic abundance, locus-specificity, co-dominance, the potential for high throughput analysis and lower genotyping error rates [6,19–24]. Studies on the comparative utilization of either SSRs or SNPs have revealed that SSRs with moderate density seems to be more effective for diversity and population structure analysis in maize but as the number of SNP markers increases the results obtained are comparable [19,25].
In the 1990s, the maize improvement programme of IITA initiated a breeding programme for the extra-early- and early-maturing groups which resulted in the development of some multiple stress tolerant (drought, Striga, and streak resistant/tolerant) broad-based populations such as TZE-W Pop DT STR and TZE-Y Pop DT STR in the early maturing (90–95 days to physiological maturity) group, and TZEE-W Pop DT STR and TZEE-Y Pop DT STR in the extra-early (80–85 days to physiological maturity) maturing group during the first decade of the inception of the programme [26,27]. Subsequently, numerous open-pollinated varieties (OPVs) and inbred lines have been extracted from these multiple stress tolerant broad-based populations and their derivatives for all agro-ecologies of West and Central Africa (WCA) sub-regions by IITA and its National Agricultural Research (NARS) partners, which has ultimately played an important role in hybrid maize development in WCA. Most of these inbred lines have been characterized using both morphological and/or molecular markers especially SSR markers [28]. Given the cost-effectiveness of SNP markers and their growing use for a wide range of applications in different crops, it is of interest to shift to next-generation markers in the assessment of genetic differences and relationships among the new extra-early- and early-maturing white and yellow maize inbred lines that have been recently developed by IITA for their effective classification into heterotic groups. This will serve as a guide to parent selection for further hybrid development. Therefore, the objective of the present study was to examine the genetic diversity and population structure of Striga resistant and/or drought-tolerant maize inbred lines derived from different source populations selected from the panel of inbred lines of the IITA maize improvement programme using SNP markers.
Materials and methods
Germplasm
The 94 inbred lines used in this study were developed by the West and Central Africa Collaborative Maize Research Network/International Institute of Tropical Agriculture (WECAMAN/IITA), with tolerance or resistance to Striga and maize streak virus (MSV), and/or tolerance to drought (S1 Table). The inbred lines were extracted from twelve broad-based and two narrow-based source populations developed from both local and exotic germplasm identified based on several years of extensive testing for adaptation to the Guinea and Sudan savanna agro-ecologies of WCA (Table 1). The first generation of inbred lines were obtained following six generations of self-pollination in four early (TZE-W Pop DT STR C0, TZE-Y Pop DT STR C0, TZE Comp5-Y C6 and WEC STR) and two extra-early (TZEE-W Pop DT STR C0 and TZEE-Y Pop DT STR C0) broad-based populations with varied levels of drought tolerance and Striga resistance. A few inbred lines extracted from the two extra-early source populations, were categorized as early maturing because the flowering dates were typical of lines of the early maturing group. A second generation of inbred lines was developed from two F2 populations derived from the bi-parental crosses, TZEI 1 x TZEI 2 and TZEI 11 x TZEI 8 which involved elite parental inbred lines from two of the four broad-based early maturing populations. Another set of inbred lines was extracted from two other bi-parental crosses, TZE-W Pop x 1368 STR and TZE-W Pop x LD. The second generation of inbred lines was in all cases obtained following 6–7 cycles of self-pollination and selection for drought tolerance and/or resistance to Striga.
Table 1. Source populations of 94 maize inbred lines used in the present study.
| S/N | Source population | Number of extracted inbred line | Grain colour |
|---|---|---|---|
| 1 | TZE-W Pop x 1368 STR | 4 | White |
| 2 | TZE-W Pop x LD | 1 | White |
| 3 | TZE-W Pop STR 108 | 10 | White |
| 4 | TZE-W Pop STR Co | 4 | White |
| 5 | TZE-W POP STR 104 | 10 | White |
| 6 | TZEE-W POP STR 104 | 4 | White |
| 7 | TZEE-W POP STR 108 | 1 | White |
| 8 | WEC STR | 2 | White |
| 9 | (TZEI 1 x TZEI 2) | 10 | White |
| 10 | TZE-Y Pop STR Co | 12 | Yellow |
| 11 | TZE-Y Pop STR 106 | 5 | Yellow |
| 12 | TZEE-Y Pop STR 106 | 9 | Yellow |
| 13 | TZE Comp5-Y C6 | 3 | Yellow |
| 14 | (TZEI 11 x TZEI 8) | 19 | Yellow |
| Total | 94 |
DNA extraction and genotyping
Fresh leaves samples were collected from three weeks old seedlings within each genotype and stored in a deep freezer at -80°C. Prior to genomic DNA extraction, each sample was dried in a Labconco Freezone 2.5L System lyophilizer (Marshall Scientific, USA) followed by grinding using SpexTM Sample Prep 2010 Geno/Grinder (Thomas Scientific, USA). Total genomic DNA extraction was performed using the DArT protocol (www.diversityarrays.com/files/DArT_DNA_isolation.pdf). The quality and quantity of DNA in each sample was determined on 2% agarose gel followed by quantification using an ND-1000 Spectrophotometer (Nanodrop Technologies). For genotyping, DNA samples were sent to Diversity Arrays Platform [29]. Library construction, sequencing and SNP calling was performed at the Diversity Arrays Facility (Canberra, Australia).
Statistics and population structure analysis
SNP markers with more than 20% of missing data, 20% of heterozygosity and the minor allele frequency lower than 0.05 were eliminated resulting in 15,047 SNP markers which were used for further analysis [30]. The polymorphic information content (PIC), major allele frequency, the number of alleles, heterozygosity and gene diversity were estimated with the aid of PowerMarker V3.2.5 [31].
The data from the 15,047 SNP markers were subjected to population structure analysis based on the admixture model-based clustering method using the software package STRUCTURE 2.3.4 [32]. The best k was identified by inputting the data into the STRUCTURE HARVESTER software utilizing the Evanno method [33,34]. The initial model was run by varying the number of clusters (k) from 1 to 20 with 10 to 20 alterations for each K. Finally, the number of clusters was set at 10 with 10 alterations for each K. The results were obtained by running the data against 10,000 Markov Chain Monte Carlo and 10,000 burn-in as previously described [30]. Each genotype was assigned to a cluster at a 90% threshold, while genotypes with less than this value were assigned to an additional cluster designated as mixed cluster.
Cluster analysis
Following the determination of the number of clusters using STRUCTURE, the 15,047 SNP marker data were analyzed using DARwin software [35]. The neighbor-joining method (NJ) under 30,000 bootstraps was used. Genetic distance matrix was generated using the Jaccard similarity test in the DARwin software [36], using the formula dij = (b + c)/ (a+ (b +c)) where dij is the dissimilarity between units i and j, a is the number of variables where Xi is present and Xj is present, b is the number of variables where Xi is present and Xj is absent, c is the number of variables where Xi is absent and Xj is present. To generate the final phylogenetic tree, the results obtained from DARwin were loaded into FigTree version 1.4.3 software [37].
Results
Panel summary statistics
The results of the summary statistics of the SNP markers are presented in Table 2. Heterozygosity averaged 0.07 and varied from 0.00 to 0.20. Gene diversity ranged from 0.01 to 0.50 with an average of 0.22. The major allele frequencies of the 15,047 primers averaged 0.84 with a range from 0.50 to 0.99. The PIC ranged from 0.01 to 0.38 with an average of 0.19 (S2 Table). The highest and lowest minor allele frequency recorded were 0.50 and 0.01, respectively.
Table 2. Diversity indices statistics of the 94 maize inbred lines based on 15,047 SNP markers.
| MaF | GD | He | PIC | MAF | |
|---|---|---|---|---|---|
| Minimum | 0.50 | 0.01 | 0.00 | 0.01 | 0.01 |
| Maximum | 0.99 | 0.50 | 0.20 | 0.38 | 0.50 |
| Mean | 0.84 | 0.22 | 0.07 | 0.19 | 0.16 |
MaF = Major allele frequency, GD = gene diversity; He = Heterozygosity, PIC = polymorphic information content, MAF = Minor allele frequency
Genetic distance and population structure using SNP markers
The genetic distance between pairwise comparisons of the inbred lines varied from 0.06 to 0.65, and the overall average distance was 0.38. The majority (67.70%) of the genetic distances fell between 0.32 and 0.42 (Fig 1). The lowest genetic distance (0.06) was observed between inbred TZdEI 24 and TZdEI 68 (S3 Table). Both inbred lines have the same descent and selection history (S1 Table). The highest genetic distance (0.65) was observed between inbred TZEI 135 and TZEI 26. These inbred lines were derived from different source populations; TZEI 26 was extracted from WEC STR, which has white endosperm colour, while TZEI 135 was derived from TZE-Y Pop STR C0, which has yellow endosperm colour (S1 Table).
Fig 1. Frequency distribution of pairwise genetic distances calculated based on Euclidean method for 94 tropical maize inbred lines genotyped with 15,047 SNPs.

Based on 15,047 SNPs, population structure analysis revealed three distinct sub-populations in the 94 inbred lines (Fig 2A and 2B). The sub-population 1 consisted of 8.5% (8 lines) of the inbred lines, 20.2% (19 lines) were grouped into sub-population 2 and 36.1% (34 lines) in sub-population 3. A 35.1% (33 lines) of the inbred lines had a probability of association less than 90%, and were grouped into mixed populations (S1 Table). The three sub-populations were separated mainly on the basis of the endosperm color, sub-populations 1 and 2 comprising only yellow endosperm maize inbred lines while majority of inbred lines (94%) in sub-population 3 consisted of white endosperm maize (S1 Table). Results showed that sub-population 1 contained only inbred lines derived from the source population TZEE-Y Pop STR 106 while the sub-population 2 comprised about 97% of inbred lines extracted from (TZEI 11 x TZEI 8). Sub-population 3 had the greatest diversity comprising six diverse source populations, namely TZE-W Pop STR 108, TZE-W Pop STR 104, (TZEI 1 x TZEI 2), TZE-W Pop x 1368 STR, WEC STR and TZE-W Pop STR Co. The expected heterozygosity among inbred lines within the three sub-populations ranged between 0.06 for sub-population 1 and 0.34 for sub-population 3 with an average of 0.19. Sub-populations 1, 2 and 3 had FST values of 0.83, 0.49 and 0.01, respectively. The highest allele frequency divergence of 0.15 was recorded between sub-populations 2 and 1, followed by sub-population 3 and 1 with 0.11, while the least allele frequency divergence of 0.07 was recorded between sub-populations 2 and 3.
Fig 2. The three sub-populations of the 94 tropical maize inbred lines using SNP markers.
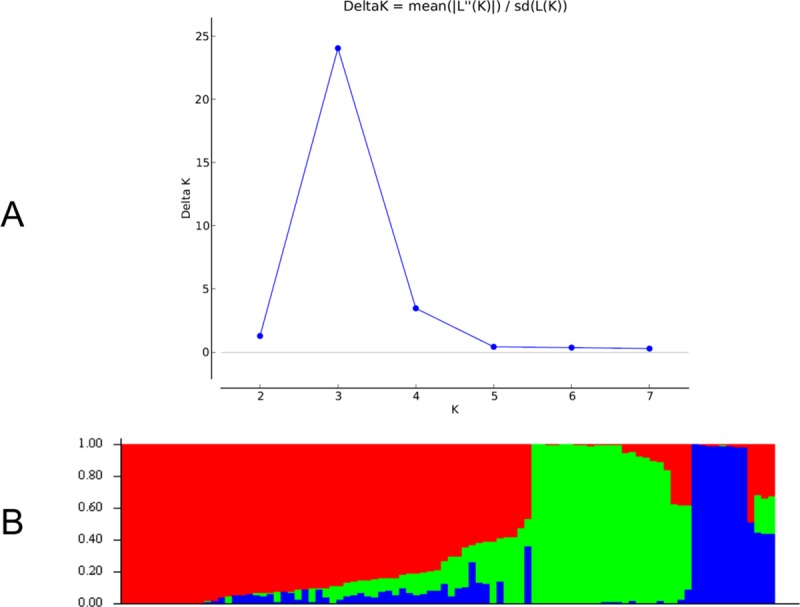
A. Best delta K estimation by Evanno method. B. Estimated population structure of 94 tropical maize inbred lines as revealed by 15,047 SNP markers for K = 3. Blue, green and red colour represents sub-population 1, 2, and 3, respectively.
Cluster analysis
The results of the population structure analysis were confirmed by the phylogenetic tree which showed that the 94 inbred lines genotyped formed three clusters and each cluster was further partitioned into sub-clusters (Fig 3). In each cluster, inbred lines were coloured based on the results of the population structure analysis. Each colour represents specific sub-population such as lines coloured with blue, green, red and black corresponding to sub-population (SP) 1, 2, 3 and 4, respectively. The sub-population 4 represents the mixture of individuals at a 90% threshold. The inbred lines were clustered based on their pedigree, selection history and endosperm colour (S1 Table). Phylogenetic cluster 1 had a total of 14 inbred lines and represented a very clear sub-cluster of 8 lines (blue colour). This sub-cluster corresponded to sub-population 1 in population structure analysis (Fig 2B), with the remaining lines considered as mixed genotypes. All 14 lines were derived from the TZE-Y POP STR 106 and were characterized by yellow endosperm maize lines. The second cluster generated by the phylogenetic analysis included 32 inbred lines classified into several sub-clusters. However, it is clear that the grouping of 20 maize inbred lines based on the phylogenetic analysis was similar to that based on the population structure analysis (SP2; green color) (Figs 2B and 3). Ninety-five percent of the lines were extracted from the bi-parental cross TZEI 11 x TZEI 8 while one of the inbred lines was sourced from TZE Comp5-Y C6. The remaining 17 lines were derived from two source populations TZE-Y Pop STR Co and TZE Comp5-Y C6 and were identified as mixed genotypes by structure analysis. The third and last cluster was the largest and most diverse among the three main clusters with 48 maize inbred lines. This cluster included inbred lines from seven source populations, namely TZE-W POP STR 108, TZE-W POP STR 104, TZE-W Pop x 1368 STR, TZE-W Pop x LD, (TZEI 1 x TZEI 2), WEC STR and TZE-Y Pop STR Co (S2). All the inbred lines in this phylogenetic cluster belong to sub-population 3 (red colour) except 12 lines which were classified by the population analysis into the admixture group (Fig 3).
Fig 3. Clustering of 94 tropical maize inbred lines based on 15,047 SNP markers.

Clusters resulting from structure analysis are shown in blue, green and red colours. Mixed individuals that did not belong to any particular group at a 90% threshold are represented by the black colour.
Discussion
Genetic diversity studies are very essential in the selection of individual genotypes among closely related groups for initiation of new breeding activities. It is particularly important that the genetic similarity or dissimilarity among inbred lines are established to facilitate the development of productive hybrids which are often a product of crosses involving inbred lines of opposing heterotic groups. Heterotic populations are also developed from inbred lines sharing common ancestry. New set of inbred lines are derived from the heterotic populations and the inbred lines are expected to combine well with inbred lines developed from other opposing heterotic populations. Genotyping has proved to be one of the reliable approaches of establishing such phylogenetic relationships among a set of inbred lines [8]. In the present study, the PIC average was of 0.19, a value found lower than ones described by some researchers in wheat (0.44; [38]), maize (0.25–0.39, [30,39–43]) and sugar beet (0.28; [44]), but similar to the results of Cruz et al. (0.21; [45]) in an oilseed crop Lesquerella, Bisen et al. [46] in soybean, Ramakrishnan et al. [47] in finger millet, Singh et al. [48] in rice and Oyekunle et al. [17], Dao et al. [40] and Senior et al. [49] in maize. Approximately 49% of the SNPs presented a MAF<0.1 and 4.5% of the SNPs showed almost equal allele frequencies (MAF ~0.5) for the two alternative alleles. Previously, it was shown in maize by using a set of 1,057 SNPs that IITA maize materials presented an average value of 0.218 and 0.202 for PIC and MAF values, respectively. Similar values were found for CIMMYT maize inbred lines [40]. The variation between our results and the findings of earlier researchers could be attributed to the differences in the composition of experimental material, population size and number of markers involved in the studies [50,51].
Genetic distance provides a measure of the degree of relatedness between individuals in a population [52]. The results obtained revealed a wide genetic variability among the inbred lines with lower number of pairwise individuals with low genetic distances, suggesting that most of the inbred line used are unique and each of them has the potential to contribute new alleles to the breeding programme.
The SNP markers clustered the inbred lines based on their ancestry, selection history and endosperm colour (Figs 2 and 3; S1 Table). However, the clustering of some inbred lines was not based on shared ancestry, indicating that inbred lines extracted from the same source population do not necessarily have the same selection history [53]. The lack of association between clustering patterns and phenotypes, environmental adaptation, maturity and the heterotic groups has also been previously observed in CIMMYT maize germplasm [51,54]. Warburton et al. [54] suggested that markers may be better indicators of relatedness of the inbred lines in cases where inbreds derived from the same source population are more different than those extracted from different source populations.
Population structure analysis is a process of inferring individual ancestry of inbred lines from genotypic information [55]. The studied SNPs revealed the presence of three sub-populations (K = 3) within the 94 inbred lines. Inbred lines with similar pedigree tended to cluster into the same group. The SNP markers clearly assigned inbred lines into heterotic groups based on the source populations, with individuals of the same endosperm colour and similar genetic background placed in the same sub-population. The grouping of inbred lines using SNP markers on the basis of similarity of pedigree and phenotype indicated that the SNP markers were very effective in assigning the inbred lines into homogenous groups. Our findings might have been influenced by the relatively larger number of SNPs used in the present study which would facilitate better interpretation of the results and inference about population structure [51].
The lower levels of heterozygosity observed among inbred lines within the three sub-populations suggested that the SNPs were effective in forming homogeneous sub-populations. The very large FST values obtained for sub-populations and the moderate allele frequency divergence observed between sub-populations indicates that these inbred lines are fixed and can be classified into genetically distinct groups (heterotic groups). These characteristics make them a valuable resource for genetic studies in maize and association mapping where uniformity of inbred lines and genetic divergence are required. For future development of productive hybrids, crosses should be made between parent lines from different sub-populations, particularly between inbred lines in sub-populations 1 and 2.
Conclusion
High genetic distances obtained among paired inbred lines revealed the uniqueness of the studied lines and existence of substantial genetic variability that could be exploited for the development of productive hybrids. The inbred lines were assigned into heterotic groups based on similarity of ancestry, selection history and endosperm colour. This study showed that SNP markers were more reliable in categorizing maize inbred lines into groups on the basis of shared phylogeny. The low heterozygosity observed among inbred lines within sub-populations and the moderate divergence among sub-populations suggested that the inbred lines could be used in the development of productive hybrids breeding or heterotic populations for the West African Sub-region.
Supporting information
(XLSX)
(XLSX)
(XLSX)
Acknowledgments
We are most grateful to CIMMYT for the SNP genotyping. We also acknowledge the contributions of Dr Talabi Abidemi and Mr Obisesan Oluwafemi for their helpful comments in the manuscript. Authors are very thankful to anonymous reviewers for their comments and suggestions.
Data Availability
The dataset on Genetic Diversity and Population Structure of Early-maturing Tropical Inbred Lines Using SSR and SNP Markers has been uploaded to CKAN. DOI has also been minted for the dataset. You can click any of the link below to view the uploaded dataset. Link on CKAN: http://data.iita.org/dataset/genetic-diversity-and-population-structure-of-early-maturiing-tropical-inbred-lines; DOI: https://dx.doi.org/10.25502/20181017/0932/AG.
Funding Statement
This research was funded by the Bill and Melinda Gates Foundation under Drought tolerant Maize for Africa and Stress Tolerant Maize for Africa projects of CIMMYT and IITA (OPP1134248 to BBA and MG). The funders had no role in study design, data collection and analysis, decision to publish, or preparation of the manuscript.
References
- 1.Carena MJ, Hallauer AR, Miranda Filho JB. Quantitative Genetics in Maize Breeding [Internet]. 2nd ed PROHENS J, NUEZ F, CARENA MJ, editors. Springer, Dordrecht; 2010. 680 p. Available from: http://link.springer.com/10.1007/978-1-4419-0766-0 [Google Scholar]
- 2.Thompson JA, Nelson RL, Vodkin LO. Identification of diverse soybean germplasm using RAPD markers. Crop Sci. 1998;38(5):1348–55. [Google Scholar]
- 3.Mohammadi SA, Prasanna BM. Analysis of genetic diversity in crop plants—Salient statistical tools and considerations. Crop Sci. 2003;43(4):1235–48. [Google Scholar]
- 4.Reif JC, Melchinger AE, Xia XC, Warburton ML, Hoisington DA, Vasal SK, et al. Genetic distance based on simple sequence repeats and heterosis in tropical maize populations. Crop Sci. 2003;43(4):1275–82. [Google Scholar]
- 5.Tian F, Bradbury PJ, Brown PJ, Hung H, Sun Q, Flint-Garcia S, et al. Genome-wide association study of leaf architecture in the maize nested association mapping population. Nat Genet. 2011;43(2):159–62. 10.1038/ng.746 [DOI] [PubMed] [Google Scholar]
- 6.Semagn K, Magorokosho C, Vivek BS, Makumbi D, Beyene Y, Mugo S, et al. Molecular characterization of diverse CIMMYT maize inbred lines from eastern and southern Africa using single nucleotide polymorphic markers. BMC Genomics. 2012;13(1). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 7.Talabi AO, Badu-Apraku B, Fakorede MAB. Genetic variances and relationship among traits of an early maturing maize population under drought-stress and low nitrogen environments. Crop Sci. 2017;57(2):681–92. [Google Scholar]
- 8.Govindaraj M, Vetriventhan M, Srinivasan M. Importance of genetic diversity assessment in crop plants and its recent advances: An overview of its analytical perspectives. Genet Res Int. 2015;2015(Fig 1). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 9.Badu-Apraku B, Fakorede MAB. Advances in genetic enhancement of early and extra-early maize for sub-Saharan Africa. 2017. 1–604 p. [Google Scholar]
- 10.Govindaraj M, Shanmugasundaram P, Sumathi P, Muthiah A. Simple, Rapid And Cost Effective Screening Method For Drought Resistant Breeding In Pearl Millet. Electron J Plant Breed. 2010;1(4):590–9. [Google Scholar]
- 11.Collard BCY, Mackill DJ. Marker-assisted selection: An approach for precision plant breeding in the twenty-first century. Philos Trans R Soc B Biol Sci. 2008;363(1491):557–72. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 12.Larnkey KR, Staub JE, Melchinger AE, Gumber RK. Overview of Heterosis and Heterotic Groups in Agronomic Crops. Concepts Breed heterosis Crop plants [Internet]. 1998;(25):29–44. Available from: https://dl.sciencesocieties.org/publications/books/abstracts/cssaspecialpubl/conceptsandbree/29 [Google Scholar]
- 13.Liu K, Goodman M, Muse S, Smith JS, Buckler E, Doebley J. Genetic Structure and Diversity among Maize Inbred Lines as Inferred from DNA Microsatellites. Genetics. 2003;165(4):2117–28. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 14.Menkir A, Kling JG, Badu-Apraku B, Ingelbrecht I. Molecular marker-based genetic diversity assessment of Striga-resistant maize inbred lines. Theor Appl Genet. 2005;110(6):1145–53. 10.1007/s00122-005-1946-3 [DOI] [PubMed] [Google Scholar]
- 15.Warburton ML, Reif JC, Frisch M, Bohn M, Bedoya C, Xia XC, et al. Genetic diversity in CIMMYT non-temperate maize germplasm: Landraces, open pollinated varieties, and inbred lines. Crop Sci. 2008;48(2):617–24. [Google Scholar]
- 16.Akaogu IC, Badu-Apraku B, Adetimirin VO, Vroh-Bi I, Oyekunle M, Akinwale RO. Genetic diversity assessment of extra-early maturing yellow maize inbreds and hybrid performance in Striga-infested and Striga-free environments. J Agric Sci. 2013;151(4):519–37. [Google Scholar]
- 17.Oyekunle M, Badu-Apraku B, Hearne S, Franco J. Genetic diversity of tropical early-maturing maize inbreds and their performance in hybrid combinations under drought and optimum growing conditions. F Crop Res [Internet]. 2015;170(2015):55–65. Available from: 10.1016/j.fcr.2014.10.005 [DOI] [Google Scholar]
- 18.Mohan Jain S, Brar DS. Molecular techniques in crop improvement: 2nd edition. Mol Tech Crop Improv 2nd Ed [Internet]. 2009;1:1–772. Available from: http://link.springer.com/10.1007/978-90-481-2967-6 [Google Scholar]
- 19.Jones ES, Sullivan H, Bhattramakki D, Smith JSC. A comparison of simple sequence repeat and single nucleotide polymorphism marker technologies for the genotypic analysis of maize (Zea mays L.). Theor Appl Genet [Internet]. 2007;115(3):361–71. Available from: 10.1007/s00122-007-0570-9 [DOI] [PubMed] [Google Scholar]
- 20.Wen W, Araus JL, Shah T, Cairns J, Mahuku G, Bänziger M, et al. Molecular characterization of a diverse maize inbred line collection and its potential utilization for stress tolerance improvement. Crop Sci. 2011;51(6):2569–81. [Google Scholar]
- 21.Prasanna BM. Diversity in global maize germplasm: Characterization and utilization. J Biosci. 2012;37(5):843–55. [DOI] [PubMed] [Google Scholar]
- 22.Raza S, Shoaib MW, Mubeen H. Genetic Markers: Importance, uses and applications. Int J Sci Res Publ [Internet]. 2016;6(3):221–2250. Available from: www.ijsrp.org [Google Scholar]
- 23.Zhangxiong L, Li H, Wen Z, Fan X, Li Y, Guan R, et al. Comparison of Genetic Diversity between Chinese and American Soybean (Glycine max (L.)) Accessions Revealed by High-Density SNPs. Front Plant Sci [Internet]. 2017;8(November):1–13. Available from: http://journal.frontiersin.org/article/10.3389/fpls.2017.02014/full [DOI] [PMC free article] [PubMed] [Google Scholar]
- 24.Mengesha WA, Menkir A, Unakchukwu N, Meseka S, Farinola A, Girma G, et al. Genetic diversity of tropical maize inbred lines combining resistance to Striga hermonthica with drought tolerance using SNP markers. Plant Breed. 2017;136(3):338–43. [Google Scholar]
- 25.Hamblin MT, Warburton ML, Buckler ES. Empirical comparison of simple sequence repeats and single nucleotide polymorphisms in assesment of maize diversity and relatedness. PLoS One [Internet]. 2007;2(12):1–9. Available from: http://dx.plos.org/10.1371/journal.pone.0001367 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 26.Badu-Apraku B, Fakorede MAB, Ouedraogo M, Carsky RJ. Impact, challenges, and prospects of maize research and development in West and Central Africa. In: Proc Regional Maize Workshop, IITA, Cotonou, Benin Republic. WECAMAN/IITA, Ibadan, Nigeria.; 2001. p. 4–7.
- 27.Estep MC, Gowda BS, Huang K, Timko MP, Bennetzen JL. Genomic Characterization for Parasitic Weeds of the Genus by Simple Sequence Analysis In: The Plant Genome Journal [Internet]. WECAMAN/IITA, Ibadan, Nigeria; 2012. p. 30 Available from: https://www.crops.org/publications/tpg/abstracts/5/1/30 [Google Scholar]
- 28.Oyekunle M, Badu-Apraku B. Agronomic performance of drought-tolerant maize hybrids in diverse environments of lowland tropics. J Crop Improv. 2017;31(6):743–57. [Google Scholar]
- 29.Jaccoud D. Diversity Arrays: a solid state technology for sequence information independent genotyping. Nucleic Acids Res [Internet]. 2001;29(4):25e–25. Available from: https://academic.oup.com/nar/article-lookup/doi/10.1093/nar/29.4.e25 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 30.Zhang X, Zhang H, Li L, Lan H, Ren Z, Liu D, et al. Characterizing the population structure and genetic diversity of maize breeding germplasm in Southwest China using genome-wide SNP markers. BMC Genomics [Internet]. 2016;17(1):1–16. Available from: 10.1186/s12864-016-3041-3 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 31.Liu K, Muse S V. PowerMaker: An integrated analysis environment for genetic marker analysis. Bioinformatics. 2005;21(9):2128–9. 10.1093/bioinformatics/bti282 [DOI] [PubMed] [Google Scholar]
- 32.Earl DA, vonHoldt BM. STRUCTURE HARVESTER: A website and program for visualizing STRUCTURE output and implementing the Evanno method. Conserv Genet Resour. 2012;4(2):359–61. [Google Scholar]
- 33.Evanno G, Regnaut S, Goudet J. Detecting the number of clusters of individuals using the software STRUCTURE: A simulation study. Mol Ecol. 2005;14(8):2611–20. 10.1111/j.1365-294X.2005.02553.x [DOI] [PubMed] [Google Scholar]
- 34.Falush D, Stephens M, Pritchard JK. Inference of population structure using multilocus genotype data: Dominant markers and null alleles. Mol Ecol Notes. 2007;7(4):574–8. 10.1111/j.1471-8286.2007.01758.x [DOI] [PMC free article] [PubMed] [Google Scholar]
- 35.Perrier and Jacquemoud-Collet. DARwin software [Internet]. 2006. Available from: http://http//darwin.cirad.fr [Google Scholar]
- 36.Jaccard P. Nouvelles Recherches sur la distribuition Florale. Bull la Sociéetée Vandoise des Sci Nat. 1908;44(44):223–70. [Google Scholar]
- 37.Rambaut A. FigTree v1.4.3. Molecular evolution, phylogenetics and epidemiology. 2016. [Google Scholar]
- 38.A HR, Stodart BJA, B CC, Mackay MC, B MM, A AM, et al. Molecular diversity and genetic structure of modern and traditional landrace cultivars of wheat (Triticum aestivum L.). 2010;222–9. [Google Scholar]
- 39.van Inghelandt D, Melchinger AE, Lebreton C, Stich B. Population structure and genetic diversity in a commercial maize breeding program assessed with SSR and SNP markers. Theor Appl Genet. 2010;120(7):1289–99. 10.1007/s00122-009-1256-2 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 40.Dao A, Sanou J, Mitchell SE, Gracen V, Danquah EY. Genetic diversity among INERA maize inbred lines with single nucleotide polymorphism (SNP) markers and their relationship with CIMMYT, IITA, and temperate lines. BMC Genet. 2014;15(1):1–14. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 41.Yang X, Gao S, Xu S. Characterization of a global germplasm collection and its potential utilization for analysis of complex quantitative traits in maize. Mol Breed. 2011;(2011)(28):511–26. [Google Scholar]
- 42.Wu X, Li Y, Li X, Li C, Shi Y, Song Y, et al. Analysis of genetic differentiation and genomic variation to reveal potential regions of importance during maize improvement. 2015;1–13. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 43.Wu X, Li Y, Shi Y, Song Y. Fine genetic characterization of elite maize germplasm using high ‑ throughput SNP genotyping. Theor Appl Genet. 2014;(127):621–31. [DOI] [PubMed] [Google Scholar]
- 44.Simko I, Eujayl I, Hintum TJL Van. Plant Science Empirical evaluation of DArT, SNP, and SSR marker-systems for genotyping, clustering, and assigning sugar beet hybrid varieties into populations. Plant Sci [Internet]. 2012;184:54–62. Available from: 10.1016/j.plantsci.2011.12.009 [DOI] [PubMed] [Google Scholar]
- 45.Cruz VM V, Kilian A, Dierig DA. Development of DArT Marker Platforms and Genetic Diversity Assessment of the U. S. Collection of the New Oilseed Crop Lesquerella and Related Species. PLoS One. 2013;8(5):1–13. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 46.Bisen A, Khare D, Nair P, Tripathi N. SSR analysis of 38 genotypes of soybean (Glycine Max (L.) Merr.) genetic diversity in India. Physiol Mol Biol Plants. 2014;21(1):109–15. 10.1007/s12298-014-0269-8 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 47.Ramakrishnan M, Antony Ceasar S, Duraipandiyan V, Al-Dhabi NA, Ignacimuthu S. Assessment of genetic diversity, population structure and relationships in Indian and non-Indian genotypes of finger millet (Eleusine coracana (L.) Gaertn) using genomic SSR markers. Springerplus [Internet]. 2016;5(1):1–11. Available from: http://www.ncbi.nlm.nih.gov/pubmed/26900542%5Cnhttp://www.pubmedcentral.nih.gov/articlerender.fcgi?artid=PMC4749518 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 48.Singh N, Choudhury DR, Singh AK, Kumar S, Srinivasan K, Tyagi RK, et al. Comparison of SSR and SNP markers in estimation of genetic diversity and population structure of Indian rice varieties. PLoS One. 2013;8(12):1–14. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 49.Senior ML, Murphy JP, Goodman MM, Stuber CW. Utility of SSRs for determining genetic similarities and relationships in maize using an agarose gel system. Crop Sci. 1998;38(4):1088–98. [Google Scholar]
- 50.Yan J, Yang X, Shah T, Sánchez-Villeda H, Li J, Warburton M, et al. High-throughput SNP genotyping with the Goldengate assay in Maize. Mol Breed. 2010;25(3):441–51. [Google Scholar]
- 51.Xia XC, Reif JC, Hoisington DA, Melchinger AE, Frisch M, Warburton ML. Genetic diversity among CIMMYT maize inbred lines investigated with SSR markers: I. Lowland tropical maize. Crop Sci. 2004;44(6):2230–7. [Google Scholar]
- 52.Garcia AAF, Benchimol LL, Barbosa AMM, Geraldi IO, Souza CL, de Souza AP. Comparison of RAPD, RFLP, AFLP and SSR markers for diversity studies in tropical maize inbred lines. Genet Mol Biol [Internet]. 2004;27(4):579–88. Available from: http://www.scielo.br/scielo.php?script=sci_arttext&pid=S1415-47572004000400019&lng=en&tlng=en [Google Scholar]
- 53.Badu-Apraku B, Menkir A, Fakorede MAB, Fontem Lum A, Obeng-Antwi K. Multivariate analyses of the genetic diversity of forty-seven Striga resistant tropical early maturing maize inbred lines. Maydica. 2006;51(3–4):551–9. [Google Scholar]
- 54.Warburton ML, Xianchun X, Crossa J, Franco J, Melchinger AE, Frisch M, et al. Genetic characterization of CIMMYT inbred maize lines and open pollinated populations using large scale fingerprinting methods. Crop Sci. 2002;42(6):1832–40. [Google Scholar]
- 55.Lawson DJ, Falush D. Population Identification Using Genetic Data. Annu Rev Genomics Hum Genet [Internet]. 2012;13(1):337–61. Available from: http://www.annualreviews.org/doi/10.1146/annurev-genom-082410-101510 [DOI] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.
Supplementary Materials
(XLSX)
(XLSX)
(XLSX)
Data Availability Statement
The dataset on Genetic Diversity and Population Structure of Early-maturing Tropical Inbred Lines Using SSR and SNP Markers has been uploaded to CKAN. DOI has also been minted for the dataset. You can click any of the link below to view the uploaded dataset. Link on CKAN: http://data.iita.org/dataset/genetic-diversity-and-population-structure-of-early-maturiing-tropical-inbred-lines; DOI: https://dx.doi.org/10.25502/20181017/0932/AG.