

Abstract
Burkholderia pseudomallei is a Gram-negative environmental bacterium and the aetiological agent of melioidosis, a life-threatening infection that is estimated to account for ~89,000 deaths per year worldwide. Diabetes mellitus is a major risk factor for melioidosis, and the global diabetes pandemic could increase the number of fatalities caused by melioidosis. Melioidosis is endemic across tropical areas, especially in southeast Asia and northern Australia. Disease manifestations can range from acute septicaemia to chronic infection, as the facultative intracellular lifestyle and virulence factors of B. pseudomallei promote survival and persistence of the pathogen within a broad range of cells, and the bacteria can manipulate the host’s immune responses and signalling pathways to escape surveillance. The majority of patients present with sepsis, but specific clinical presentations and their severity vary depending on the route of bacterial entry (skin penetration, inhalation or ingestion), host immune function and bacterial strain and load. Diagnosis is based on clinical and epidemiological features as well as bacterial culture. Treatment requires long-term intravenous and oral antibiotic courses. Delays in treatment due to difficulties in clinical recognition and laboratory diagnosis often lead to poor outcomes and mortality can exceed 40% in some regions. Research into B. pseudomallei is increasing, owing to the biothreat potential of this pathogen and increasing awareness of the disease and its burden; however, better diagnostic tests are needed to improve early confirmation of diagnosis, which would enable better therapeutic efficacy and survival.
Melioidosis is an infectious disease caused by the environmental Gram-negative bacterium Burkholderia pseudomallei. First recognised in 1911 (REF. 1) (FIG. 1), the organism is commonly found in the rhizosphere (the layer of soil directly influenced by root secretions and soil microorganisms)2 and surface groundwater of many tropical and subtropical regions3,4, and can infect humans and a wide range of animals.
Figure 1 |. Milestones in the history of melioidosis.
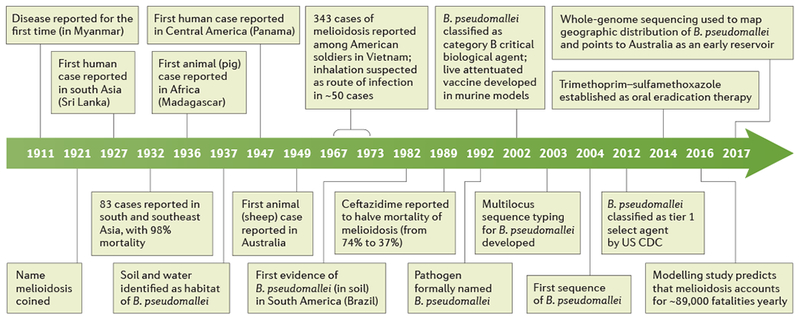
Melioidosis was first recognised in Rangoon in 1911 by the British doctor Alfred Whitmore and his assistant C. S. Krishnaswami, although the name of the disease was coined by Thomas Stanton and William Fletcher. From the time when the aetiological organism was first identified, it has been renamed many times: Bacterium (or Bacillus) whitmori, Malleomyces pseudomallei, Loefflerella pseudomallei, Pfeifferella whitmori, Pseudomonas pseudomallei and, finally, it was officially named Burkholderia pseudomallei in 1992. CDC, Centers for Disease Control and Prevention.
Naturally acquired infections in humans and animals results from exposure through broken skin, inhalation or ingestion of B. pseudomallei5; certain environmental conditions, such as tropical storms and specific occupations (for example, rice farming), are known to increase the risk of exposure6. B. pseudomallei infection can be acute, chronic or latent, although infection usually results in subclinical disease as the majority of immunocompetent individuals can clear the infection. Only those individuals with B. pseudomallei infection who develop clinical symptoms (either acute or chronic) are considered to have melioidosis.
Most cases of melioidosis (85%) result in acute infections from recent bacterial exposure7. The majority of patients with acute melioidosis present with sepsis (a life-threatening, dysregulated, systemic inflammatory and immune response that can cause organ dysfunction) with or without pneumonia, or localized abscesses, regardless of the route of infection. However, the presence of nonspecific signs and symptoms can often hinder the diagnosis and management of melioidosis, which has been nicknamed ‘the great mimicker’ (REF. 8). Chronic melioidosis is defined as a symptomatic infection that lasts >2 months, and it occurred in 11% of individuals infected with B. pseudomallei in a 20-year prospective Australian study7. The host’s immune response to acute infection is both humoral (involving cytokine release, especially interferon-γ (IFNγ)) and cell-mediated, and can completely eradicate or control the infection in most immunocompetent individuals. An unknown percentage of people exposed to B. pseudomallei can develop a latent infection (that is, the infection is asymptomatic and the pathogen is not cleared); activation from latency has been estimated to account for <5% of all melioidosis cases7, but may result in infection becoming apparent many years after exposure.
The case fatality rate of melioidosis is 10–50%6. Of the individuals who survive acute melioidosis, 5–28% experience recurrent infection, which could be due to recrudescence (that is, recurrence) of the original strain, which was not completely cleared and persisted in a dormant state, or reinfection with a different strain following re-exposure6,9–11. Approximately 80% of patients have known risk factors, mainly diabetes mellitus12 (BOX 1). The host-pathogen interplay is complicated by the tropism of the pathogen for a wide variety of cells and its ability to subvert and avoid the host innate immune response13.
Box 1 |. Diabetes mellitus and melioidosis.
There is a strong correlation between diabetes mellitus and Burkholderia pseudomallei infection, as 23–60% of patients with melioidosis also have diabetes12. Diabetes results in blunted B. pseudomallei-specific cellular responses during acute infection121, including decreased capacity for macrophages to phagocytose and kill the bacteria, reduced lipopolysaccharide-induced generation of CD4+ regulatory T (Treg) cells and impairment of Toll-like receptor-mediated myeloid differentiation primary response protein MyD88 inflammatory signalling. Dysregulated phosphorylation of nuclear factor-κB results in excessive tumour necrosis factor and IL-12 production by mononuclear cells, resulting in greater risk of septic shock222,223. Furthermore, disease progression and severity in diabetes is exacerbated by loss of effective proliferation of CD4+ T cells (which express higher levels of cytotoxic T lymphocyte protein 4) and loss of CD4+ T cell function, which is exacerbated by increased expression of programmed cell death 1 ligand 1 (a known regulator of T cell activation) on neutrophils; these neutrophils also inhibit interferon-γ production224.
In individuals with diabetes, several studies have demonstrated defects in neutrophil adhesion, chemotaxis and intracellular killing, but studies on the efficiency of neutrophil phagocytosis show mixed results225. Conflicting observations could be due to methodological differences; reduced phagocytosis could be explained by decreased opsonization of bacteria (a prerequisite for neutrophil uptake), possibly due to glucose affecting the thioester bond of complement C3 and thereby preventing binding to the bacterial surface225. Humoral responses are also poorer and could affect vaccination225.
However, diabetes was associated with a lower overall mortality in patients with melioidosis in Thailand, although only in those who were being treated with glibenclamide202. Glibenclamide is an anti-inflammatory agent that inhibits IL-1β secretion by monocytes and reduces neutrophil pro-inflammatory cytokine production by lowering free glutathione and enhancing IL-1 receptor-associated kinase 3 (IRAK3) pathways; this results in reduced IL-1β secretion in a dose-dependent fashion226,227. Patients taking glibenclamide prior to admission have attenuated inflammatory responses202.
Burkholderia mallei is a host-adapted (mainly causing infections in animals) species that originally derived from B. pseudomallei following substantial genome reduction (also known as genome degradation). B. mallei is extremely infectious, mainly to solipeds (mammals that have a single hoof on each foot; for example, horses) but can occasionally infect humans. B. mallei is the aetiological agent of glanders, a disease with similar manifestations to melioidosis. The US Centers for Disease Control and Prevention (CDC) have classified B. pseudomallei and B. mallei (which was used as a biological weapon in World War I)6 as tier 1 select agents because of their biothreat potential (tier 1 select agents present “the greatest risk of deliberate misuse with the most significant potential for mass casualties or devastating effect to the economy, critical infrastructure; or public confidence”)14. No vaccine for either is currently available15,16, which further exacerbates concerns of a possible emerging public health threat.
This Primer summarizes the state of the field in melioidosis research, focusing on epidemiology, pathophysiology (including host-pathogen interactions), diagnostics, screening, prevention and clinical management. In the Outlook, we explore future directions of research in the omics and cutting-edge immunology era, argue whether melioidosis should be recognized as a neglected tropical disease and discuss whether a viable vaccine is on the horizon.
Epidemiology
B. pseudomallei in the environment
B. pseudomallei is well-known to be present in soil and surface water in southeast Asia and northern Australia; however, case reports of melioidosis and predictive modelling studies suggest that it is probably widely present in many countries across the tropics (BOX 2).
Box 2 |. Genome and phylogeny of Burkholderia pseudomallei.
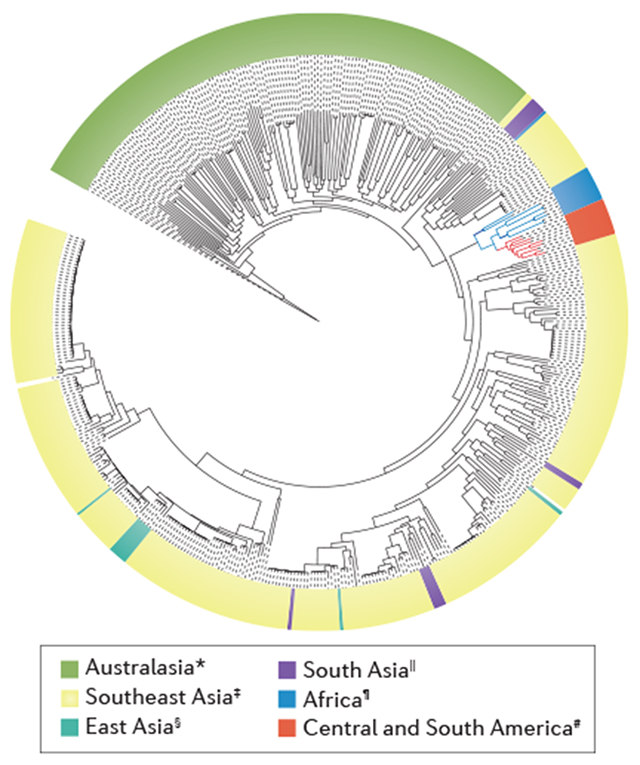
The genome of Burkholderia pseudomallei consists of two circular replicons — chromosome 1 (4.07Mb) and chromosome 2 (3.17 Mb). Chromosome 1 largely encodes proteins involved in core housekeeping functions, such as cell wall synthesis, metabolism and motility, whereas chromosome 2 mostly encodes proteins required for accessory functions involved in adaptation to environmental conditions228. Within this bipartite structure, horizontal gene transfer (transmission of genetic material other than by vertical transmission from parent to offspring) provides genetic plasticity, as represented by the large metabolic repertoire and intrinsic redundancy of virulence factors, such as type III secretion systems229. The pan-genome of B. pseudomallei shows substantial genetic heterogeneity between strains, which is largely influenced by horizontal gene transfer, recombination and mutations230,231.
This highly plastic and, as a consequence, highly variable genome across B. pseudomallei strains could also have a role in the various manifestations and disease courses of melioidosis98,229. Bacterial genetic mutations can also occur during the infection. For example, mutations in variable number tandem repeats were detected in isolates collected 2 weeks apart from a patient with acute melioidosis229,230. Geographical segregation could also contribute to different clinical manifestations, as region-specific genetic loci are associated with variability in survival and virulence232. Whole-genome tiling array expression data demonstrated that non-coding RNA could play an important part in virulence and host-pathogen interactions40.
Phylogenetic analysis demonstrates greater genetic diversity and a clear distinction between isolates from Australia and Asia, supporting the hypothesis that Australia was an early reservoir for the current global B. pseudomallei population232. Within the endemic zone of southeast Asia, the Mekong subregion has emerged as a hot spot for B. pseudomallei evolution232. Furthermore, isolates from Africa and Central and South America seem to have a common origin, as suggested by close ancestry that originated between the 17th and 19th centuries232.
*New Caledonia, Australia, Fiji and Papua New Guinea; ‡Brunei, Cambodia, Indonesia, Lao People’s Democratic Republic, Malaysia, Philippines, Singapore, Thailand and Vietnam; §China; ∥India and Bangladesh; ¶Burkina Faso, Chad, Gabon, Kenya, Madagascar, Mauritius and Nigeria; #Ecuador, Brazil, Martinique, Puerto Rico, Venezuela and the Virgin Islands. Figure adapted from (REF. 232), Macmillan Publishers Limited.
A consensus guideline for soil sampling for B. pseudomallei was proposed in 2013 with the goal of elucidating the global distribution of the bacterium17. B. pseudomallei is most abundant in soil at depths of ≥ 10 cm from the surface17; however, during the rainy season it can move from deeper soil layers to the surface, where it can then multiply17.
B. pseudomallei can survive in extreme conditions, such as in distilled (without nutrients) water (for ≥16 years)18, nutrient-depleted soil19 or desert environments20. Outbreaks of melioidosis from contaminated, unchlorinated water supplies have been reported in the Northern Territory, Australia21, and have been associated with chlorination failure (that is, insufficient addition of chlorine to the water) in Western Australia22. B. pseudomallei is also commonly found in unchlorinated water supplies and drinking water in rural areas in Thailand23. Nosocomial (originating in a hospital) infections have been attributed to B. pseudomallei-contaminated wound irrigation fluid, antiseptics and hand wash detergent24,25.
B. pseudomallei has rarely been detected in air. Aerosolized bacteria were first isolated in 1989 (REF. 26), and in 2015, B. pseudomallei DNA was detected in filtered air using quantitative PCR27. Whole-genome sequencing linked the bacteria found in an air isolate to the clinical isolate from a patient with mediastinal melioidosis (that is, with infection of the midline anatomical structures or connective tissue of the chest)28. Penetration through the skin, ingestion and inhalation are all important routes of infection with environmental B. pseudomallei5. Reported neonatal cases were probably caused by mother-to-child transmission (vertical transmission or breastfeeding)29, healthcare-associated infection29 or community-acquired infection29. Melioidosis is not contagious and human-to-human transmission has rarely been reported6.
Global burden of melioidosis
A 2016 modelling study estimated that there are ~165,000 cases of melioidosis in humans per year worldwide, of which 89,000 (54%) are estimated to be fatal3 (FIG. 2). This study highlights that underdiagnosis and under-reporting of melioidosis are a major issue, especially on the Indian subcontinent, where 44% of cases were predicted to occur (predicted incidence for India, Indonesia and Bangladesh are ~52,500, ~20,000 and ~16,900 cases per year, respectively). However, only ~1,300 cases were reported per year worldwide since 2010, which is <1% of the estimated annual incidence3. Melioidosis is prevalent in the Northern Territory, Australia, and northeast Thailand, where the annual incidence is up to 50 cases per 100,000 individuals4,7, and the emergence of melioidosis in areas where it was previously absent, for example, in northeastern Brazil, could be explained in part by the increasing recognition of this disease, owing to increased awareness and improved diagnostics30. Although reports of B. pseudomallei isolation from soil and animals in equatorial Africa are limited, they suggest that melioidosis is widely distributed across this region31–33. For example, Nigeria is predicted to have an incidence of ~13,400 cases per year, which is comparable to incidences observed in endemic regions such as India, Indonesia and Bangladesh3.
Figure 2 |. Estimated mortality and reported cases of melioidosis.
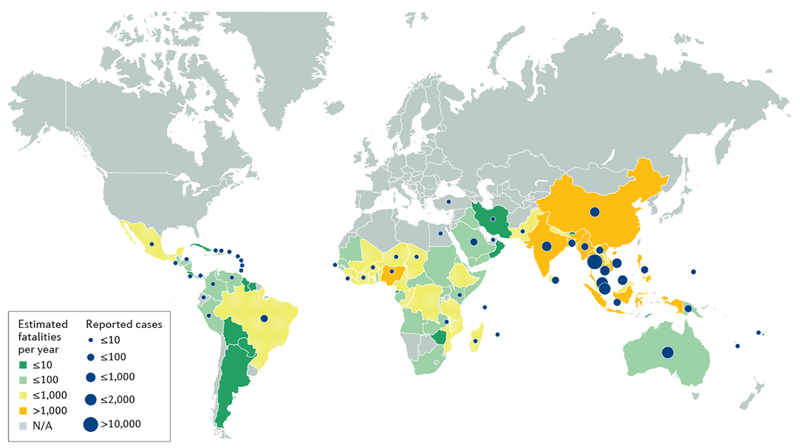
Only Australia, Brunei and Singapore have national surveillance data for melioidosis that are comparable to the estimates. Between 2010 and 2015, there were >100 culture-confirmed cases of melioidosis at a single hospital in Lao People’s Democratic Republic yearly247, a number that supports the estimated 420 cases per year countrywide3. However, ~20,000 cases of melioidosis per year are estimated in Indonesia, but only 64 have been reported in the country since 1921 (REF. 248). A large difference between the numbers of predicted and observed cases is also observed in Bangladesh, Brazil, China, India and Nigeria3. This discrepancy could be due to limitations of the model, underuse of clinical microbiology laboratories206, lack of awareness of melioidosis and poor disease reporting systems. Based on data from REF. 3. N/A, not applicable.
The predicted mortality from melioidosis is comparable to that of measles (95,600 individuals per year) and higher than that for leptospirosis (50,000 individuals per year) and dengue infection (12,500 individuals per year), which are diseases that are considered of high priority by many international health organizations3. Melioidosis can affect all age groups. In prospective studies in Australia and Thailand, the median age of patients with melioidosis was 50 years, with 5–10% ofpatients of <15 years of age7,34,35.
Risk factors
The most common risk factor predisposing individuals to melioidosis is diabetes mellitus, which is present in >50% of all patients with melioidosis worldwide35,36 (BOX 1). Individuals with diabetes mellitus have a 12-fold higher risk of melioidosis after adjustment for age, sex and other risk factors35,36. Other known risk factors include exposure to soil or water (especially during the rainy season), male sex (probably because of a greater risk of environmental exposure), age of >45 years, excess alcohol consumption and liver disease, chronic lung disease, chronic kidney disease and thalassaemia (which probably causes neutrophil dysfunction due to iron overload)6,37. Prolonged steroid use and immunosuppression can also predispose individuals to infection. Nonetheless, >80% of paediatric patients34,38 and ~20% of adult patients have no recognized risk factors35,36. Melioidosis in adults who have no risk factors generally occurs in those who have been exposed to a high bacterial load, for example, by aspiration of surface water39. Zoonotic transmission to humans resulting from contact with livestock is extremely rare; only three possible cases have been reported in Australia6.
Mechanisms/pathophysiology
B. pseudomallei is an opportunistic, facultative intracellular, motile saprophyte (an organism that obtains its energy from decaying organic matter) that possesses a remarkable intrinsic array of virulence factors (TABLE 1) and broad antimicrobial drug resistance (BOX 3). B. pseudomallei is highly adaptable, a property that enables it to generate a variety of clinical manifestations, depending on the infected tissue, and to maintain a survival advantage in infected hosts and the environment40. Numerous studies have increased our insights into the pathogenesis of B. pseudomallei infection13,41–44 (FIG. 3).
Table 1.
Selected virulence factors of Burkholderia pseudomallei
| Gene | Antigen | Function | Refs |
|---|---|---|---|
| Adherence | |||
| pilA | Type IV pilin subunit protein PilA | • Temperature-dependent adherence and formation of microcolonies in some Burkholderia pseudomallei strains • Intracellular motility |
42,50 |
| boaA | Adhesins | • T5SS autotransporters • Role in cell attachment and possibly intracellular replication |
48,54,250 |
| boaB | |||
| bpaC | Adhesin | • T5SS trimeric autotransporter ad hesin • Protects from complement killing • Involved in cell attachment, which could be cell-specific (for example, in ciliated mucosal epithelial cells) |
43,48 |
| fliC | Flagellin structural component | • Reguired for flagellar assembly • Polar tuft of 2–4 flagella allows temperature-independent motility • Involved in cell adherence |
251,252 |
| Invasion | |||
| bopE | Guanine nucleotide exchange factor BopE | • T3SS effector • Targets CDC42 and RAC1, inducing actin rearrangements and aiding cellinvasion |
55 |
| bipB | Translocator protein | • T3SS components • Involved in endocytic vesicle survival, escape and cell invasion |
46,62 |
| bipC | Effector protein | ||
| bipD | Translocator protein | ||
| irlR | Transcriptional activator protein IrlR | Mutants displayed reduced invasion | 58 |
| Endocytic escape | |||
| bopA | Effector protein | • T3SS component • Involved in endocytic vesicle membrane disruption and avoidance of autophagy |
77,78 |
| bsaQ | T3SS structural component | Involved in endocytic vesicle escape, cell invasion and plaque formation | 45,253 |
| bsaZ | T3SS structural component | Implicated in endocytic vesicle escape and intracellular replication | 46,254 |
| bsaU | T3SS structural component | Involved in endocytic vesicle escape and early onset activation of the caspase 1 pathway in macrophages | 115,255 |
| CHBP (cif homologue) | ATP and GTP binding protein | • T3SS structural component • Delays host cell maturation, arresting cycle in G2-M and impeding apoptosis |
13,43,62,256 |
| Intracellular survival | |||
| purM | Phosphoribosylformylglycinamidine cyclo-ligase | • Purine biosynthetic pathway • Knockout models show decreased intracellular replication |
42,255 |
| purN | Phosphoribosylglycinamide formyltransferase | ||
| sodC | Superoxide dismutase | Resistance to oxidative stress | 70–73 |
| katG | Catalase-peroxidase | ||
| ahpC | Alkyl hydroperoxide reductase | ||
| dpsA | DNA starvation and stationary phase protection protein | ||
| rpoE | RNA polymerase σ-factor RpoE | • Biofilm formation, heat stress response via RNA polymerase σ-factor RpoH-regulated heat shock proteins, oxidative and osmotic stress • Mutants show reduced intracellular survival in macrophages |
44,257 |
| virAG locus | Two-component regulatory system | • Regulates T6SS transcription • Sensor of the iron-limiting environment of endocytic vesicles and histidine kinase • Upregulated at acidic pH, affecting T6SS secretion process |
13,229 |
| rpoS | RNA polymerase σ-factor RpoS | • Suppresses iNOS activity by upregulating SOCS3 and CIS cytokines • Could play a part in regulating genes involved in macrophage fusion |
68,258 |
| Actin-based motility | |||
| bimA | T5SS autotransporter | • Escape from phagosome and actin tail formation • Encephalomyelitis strongly correlated with BimABm allele |
80,81,83 |
| Multinuclear giant cell formation | |||
| hcp1 | Hcp1 family T6SS effector | • Role in cell fusion and macrophage cytotoxicity • Induces production of IL-10 and TGFβ |
259–262 |
| vgrG5 | Rhs element Vgr protein | • T6SS effector (tail-spike tip) • Functionally conserved across Burkholderia spp. |
263 |
| Others | |||
| tssM | TssM | • T2SS effector (a deubiquitinase) • Targets TRAF3, TRAF6 and NF-κB inhibitor-α inhibit type 1 interferon and NF-κB pathways |
47,264 |
| wcb operon | Capsular polysaccharides | • Protection from C3b complement and normal human serum • Protection from environmental dangers • Biofilm production, not essential for survival but contributes to chronic infection and latency |
265–267 |
| Various genes (including waaF, lytB (also known as ispH) and others) | Lipopolysaccharides | • Resistance to normal human serum and cationic peptides • Reduced minimal pyrogenic lethal toxicity (measure of inflammatory response) and macrophage activation • Length, number and position of fatty acyl chains can affect LPS bioactivity and vary between virulent strains |
44,111,112,268 |
| luxl and luxR homologues | N-Acyl homoserine lactones | • Quorum sensing (also mediated by a second system using HMAQ) • Upregulate transcription of genes simultaneously within a bacterial population involved in colonization, longer survival and higher LD50 • Regulation of siderophore synthesis |
269,270 |
| BLF1 | Burkholderia lethal factor 1 | • Glutamine deamidase, similar to Escherichia coli cytotoxic necrotizing factor • Irreversibly interferes with initiation of translation by inactivating EIF4A and thereby inhibiting recruitment of 40S ribosomal subunit and protein synthesis • Cell cytoskeleton alteration and cell death. Concentrations as low as 2.5 × 10−7 M can cause cell death with molecular turnover (a measure of enzymatic efficiency) like that of ricin |
271,272 |
| Multiple genes upregulated in tandem | Morphotype switching | • Seven morphotypes, wrinkled type 1 predominates • Strain differences in colony morphology phenotypically lead to changes in biofilm production, secreted enzymes and motility, hence influencing intracellular survival, lethality and persistence |
273 |
This is not a comprehensive list: the B. pseudomallei virulence factors shown are well-studied representative examples involved throughout the life cycle of B. pseudomallei. CDC42, cell division control protein 42 homologue; CIS, cytokine-inducible SH2-containing protein; EIF4A, eukaryotic initiation factor 4A; HMAQ, 4-hydroxy-3-methyl-2-alkylquinolone signalling molecules; iNOS, inducible nitric oxide synthase; NF-κB, nuclear factor-κB; LD50, median lethal dose (dose lethal to 50% of animals tested); LPS, lipopolysaccharide; RAC1, Ras-related C3 botulinum toxin substrate 1; SOCS3, suppressor of cytokine signalling 3; T5SS, type V secretion system; TGFβ, transforming growth factor-β; TNF, tumour necrosis factor; TRAF3, TNF receptor-associated factor 3; TssM, type VI secretion system ATPase and inner membrane protein.
Box 3 |. Antimicrobial drug resistance.
The Burkholderia pseudomallei genome contains several genes encoding Ambler class A, B and D β-lactamases. The most important is penA on chromosome 2, which encodes a class A membrane-bound lipoprotein that is secreted by the twin-arginine transport system with the ability to hydrolyse most β-lactams6,233.
Acquired antimicrobial resistance during melioidosis treatment is rare. Studies of acquired β-lactam resistance (including to carbapenems) whilst on therapy identified three distinct phenotypic changes, mainly resulting from penA mutations: derepression of the chromosomal enzyme, insensitivity to β-lactamase inhibitors and specific ceftazidime resistance6,234. In isolates from patients in Thailand who did not respond to ceftazidime, large segments of chromosome 2 were deleted, including three genes encoding putative penicillin-binding proteins, which are known targets of β-lactam antibiotics235.
Additionally, omp38, which encodes an outer membrane porin, is thought to contribute to ceftazidime and carbapenem resistance233. Metallo β-lactamase type 2 (encoded by blaNDM-1) is a lipoprotein carbapenemase expressed on the outer membrane of Gram-negative bacilli and can be shed in outer membrane vesicles, thereby representing a new mechanism of resistance dissemination that can confer phenotypic resistance to beneficiary bacteria236. B. pseudomallei penA could have a similar purpose236. Of note, 2017 reports from isogenic B. pseudomallei strains (isolated from the same patients at different time points and traced to a single ancestor) from patients on meropenem have shown increasing minimum inhibitory concentrations237.
B. pseudomallei lipopolysaccharide structure also plays an intrinsic part in resistance to cationic peptides, such as polymyxin B. B. pseudomallei encodes at least ten resistance nodulation division efflux pump systems, spanning both chromosomes, that confer at least partial resistance to six antibiotic classes, including aminoglycosides, fluoroquinolones and tetracyclines233. Mutations targeting folA, which encodes dihydrofolate reductase, and BpeEF-OprC efflux pump expression confer resistance to trimethoprim238. In vitro, B. pseudomallei cells growing as a biofilm were viable after 24 h of antibiotic (trimethoprim or ceftazidime) exposure, with a minimum inhibitory concentration of up to 200-times that of planktonic bacteria6. Inhibition of efflux pumps might lower resistance to ceftazidime and doxycycline in these biofilms239.
Figure 3 |. Schematic model of host-pathogen interactions and pathophysiology of melioidosis.
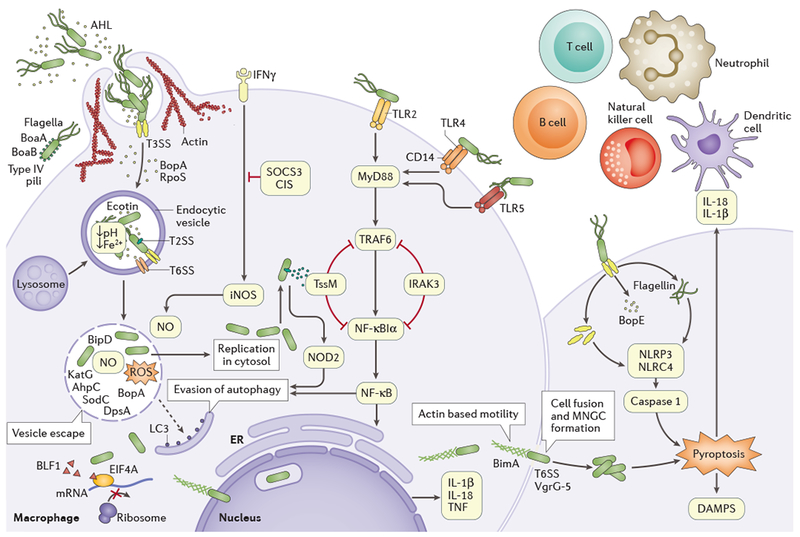
Burkholderia pseudomallei secretes N-acyl-homoserine lactones (AHL), which are signalling molecules involved in the quorum sensing machinery that is used to coordinate attacks against the host environment and biofilm formation. The type III secretion system (T3SS) effector proteins are necessary for invasion and escape from the endocytic vesicle; cell entry is aided by flagella, lipopolysaccharide (LPS), type IV pili and adhesins BoaA and BoaB. B. pseudomallei then guickly escapes the vesicle by lysing the membrane using T3SS, T6SS and T2SS. Metabolic flexibility (resistance to oxidative stress), resistance to antimicrobial cationic peptides and ecotin production enable bacteria to survive within an acidic endocytic environment. Effector protein BopA and translocator protein BipD further block sequestration in endocytic vesicles and prevent microtubule-associated proteins 1A/1B light chain 3B (LC3)-associated autophagy. Once free in the cytoplasm, B. pseudomallei replicates, induces the formation of actin-based membrane protrusions and can move via continuous polymerization of host cell actin at polar ends (a process regulated by autotransporter BimA), thereby facilitating spread to neighbouring cells, cell fusion and multinuclear giant cell (MNGC) formation. T6SS and the type IV secretion system (VgrG-5) are essential to this process. Toll-like receptors (TLRs) located on cell surfaces recognize pathogen-associated molecular patterns (such as LPS and flagella) and mediating nuclear factor-κB (NF-κB)-induced activation of the immune response, releasing pro-inflammatory cytokines IL-1β and IL-18. Intracellular inflammasome receptors such as NLR family CARD domain-containing protein 4 (NLRC4) and NACHT, LRR and PYD domains-containing protein 3 (NLRP3) recognize bacterial virulence factors and damage-associated molecular patterns (DAMPS), triggering caspase-1-mediated pyroptosis and further release of IL-1β and IL-18. IL-18 further ensures protective interferon-γ (IFNγ) production (mainly from natural killer cells). Neutrophils, dendritic cells, B cells and T cells are recruited towards the site of infection, and the complement and coagulation cascades are activated. AhpC, alkyl hydroperoxide reductase; BLF1, Burkholderia lethal factor 1; CIS, cytokine-inducible SH2-containing protein; DpsA, DNA starvation/stationary phase protection protein; EIF4A, eukaryotic initiation factor 4A; ER, endoplasmic reticulum; iNOS, inducible nitric oxide synthase; IRAK3, interleukin 1 receptor-associated kinase 3; KatG, catalase-peroxidase; MyD88, myeloid differentiation primary response protein; NF-κBIα, NF-κB inhibitor-α; NO, nitric oxide; NOD2, nucleotide-binding oligomerization domain-containing protein 2; ROS, reactive oxygen species; RpoS, RNA polymerase σ-factor RpoS; SOCS3, suppressor of cytokine signalling 3; SodC, copper/zinc superoxide dismutase; TNF, tumour necrosis factor; TRAF6, TNF receptor-associated factor 6; TssM, type VI secretion system.
B. pseudomallei infection
B. pseudomallei first enters and replicates in epithelial cells of the mucosal surface or broken skin, depending on the route of entry, and then spreads to various cell types. However, the mechanisms of cell invasion and replication are largely similar and are, therefore, discussed collectively (unless otherwise specified).
Epithelial attachment and cell invasion.
B. pseudomallei possesses multiple secretion systems, which are evolutionary apparatuses that enable the transport of proteins across cellular membranes in response to the environment and, therefore, host cell invasion. The secretion systems are classified depending on their structure, function and specificity. The type III secretion system (T3SS) comprises a molecular syringe (a structure made of a filamentous needle to translocate effector proteins into the surrounding milieu or cells) that is deployed on close contact with host cells45,46, T2SS is widely distributed in Gram-negative bacteria47 and T5SS secretes autotransporter proteins, which are usually bound to the outer membrane through some adhesin-like proteins48.
Attachment to human pharyngeal epithelial cells was initially thought to be mediated by capsular polysaccharides49 and type IV pili (hair-like structures on the bacterial surface)50). However, internalization into a cell line of human alveolar basal epithelial cells was increased in acapsular mutants compared with wild-type B. pseudomallei51. Furthermore, the type IV pilin protein PilA (encoded by pilA), a subunit type IV pili needed for adhesion to epithelial cells, could also have a role52.
Flagellar motility favours close contact with protective mucosal linings, but flagella are probably not a major adhesin for mammalian cells42,53. Two T5SS adhesin proteins, BoaA and BoaB (TABLE 1), can enhance adherence. However, double boaA and boaB knockouts show residual binding, indicating that multiple adhesins are required for cell adhesion54. Guanine nucleotide exchange factor BopE, a T3SS effector, causes rearrangement of the host actin cytoskeleton (membrane ruffling) and facilitates ingress55, and BsaQ (a conserved inner membrane T3SS protein) mutants displayed a 30% reduction in invasion42, suggesting that multiple T3SS effectors mediate cell invasion.
Host factors also play a part in epithelial attachment. Protease-activated receptor 1 (PAR1, which belongs to the subfamily of G protein-coupled receptors) is expressed on several cell types (for example, endothelial cells, platelets and monocytes) and promotes B. pseudomallei cell invasion, growth and dissemination56. However, interestingly, PAR1 inactivation had no effect on B. pseudomallei-associated mortality in mouse models, whereas it delayed time to death when the mice were infected with pneumococci56.
Intracellular survival and replication.
B. pseudomallei can invade and propagate in both phagocytic and non-phagocytic cells57,58; the bacteria replicate intra-cellularly, cause lysis or spread to and infect adjacent cells. This process causes acute symptoms, which can vary depending on the tissue or organ infected.
Following endocytosis, B. pseudomallei can be seen in endocytic vesicles and later within the cytoplasm where it replicates59,60. The vesicles then fuse with lysosomes and acidify rapidly61. The T3SS is crucial for vesicle escape before the bacteria can be degraded, as multiple strains mutant for T3SS proteins showed downstream effects, including reduced formation of the actin tail (a comet-like filamentous tail made using actin molecules from the host that the bacteria use for intracellular motility), intracellular survival, cytotoxicity and intercellular spread46,62–64. Survival within the endocytic vesicle is aided by an ecotin (a periplasmic serine protease inhibitor) homologue, which is involved in resisting degradation by lysosomal enzymes65.
B. pseudomallei can multiply within phagocytes (including neutrophils, monocytes and macrophages) without activating a bactericidal response57,58. Despite detection of lysosome fusion within B. pseudomallei-infected macrophages (suggesting that degradation of the pathogens can occur to some extent), proliferation of the surviving bacteria ultimately overwhelms the macrophage66. However, macrophages activated by IFNγ (which mediates the immune response to intracellular pathogens) display improved killing of B. pseudomallei, probably via increased activation of inducible nitric oxide synthase (iNOS)67.
In fact, bacterial killing is predominantly mediated by reactive nitrogen intermediates and reactive oxygen species (ROS)67. Consequently, an important mechanism of B. pseudomallei pathogenesis is to suppress iNOS production by upregulating two negative regulatory cytokines: suppressor of cytokine signalling 3 (SOCS3) and cytokine-inducible SH2-containing protein (CIS)68,69. Superoxide (O2−) and H2O2 degrading enzymes have been associated with mediating B. pseudomallei resistance to oxidative stress70–73 (TABLE 1).
Evasion of autophagy and cell lysis.
B. pseudomallei may trigger autophagy by a T3SS-dependent process that involves the activation of nucleotide-binding oligomerization domain-containing protein 2 (NOD2, an intracellular pathogen recognition receptor)74, resulting in bacterial killing75. However, the exact role of NOD2 might not be clearcut, as another study shows that NOD2 promotes the upregulation of SOCS3 (REF. 76). Hence, the mechanisms by which NOD2 leads to containment of B. pseudomallei are probably not mediated by cytokine suppression in murine models. Interestingly, polymorphisms in the NOD2 region are associated with susceptibility to melioidosis74. However, the effectiveness of autophagy evasion is influenced by the expression of the T3SS effector protein Bop A. Loss of Bop A (as suggested by its increased colocalization with microtubule-associated proteins 1A/1B light chain 3B (LC3; also known as MAP1LC3B), an autophagy marker protein) leads to a substantial delay in efficient endosome escape, contributing towards reduced virulence77. T3SS probably plays a crucial part in the evasion of autophagy, as bacteria with mutant BopA are taken up by autophagic vesicles more efficiently and have decreased intracellular survival78. However, the complete mechanisms of autophagy escape remain to be defined.
B. pseudomallei cytotoxicity for certain cell types is also strain-dependent: some strains cause macrophage apoptosis60, some strains cause pyroptosis (an inflammatory form of caspase-1-dependent cell lysis)63 and others cause neither57. Macrophage lysis could represent an escape mechanism for B. pseudomallei once a threshold bacterial load has been reached44. By contrast, apoptosis and degradation of infected neutrophils by macrophages is delayed in melioidosis, favouring bacterial survival79.
Intercellular and secondary spread.
Intercellular spread of B. pseudomallei is facilitated by membranous protrusions (generated by bacteria-induced rearrangements of the cytoskeleton) formed by the host cell that extend into neighbouring cells, through which bacteria travel by actin-mediated motility60,80. The autotransporter BimA interacts with monomeric actin at the tail-end of the bacteria, where polymerization occurs81. Nerve root translocation and migration along infected neurons until B. pseudomallei reaches the central nervous system has been supported by animal studies82 and linked especially to the minority bacterial genotype carrying BimABm (B. mallei-like BimA) that is found mainly in Australia83. Such cell-to-cell spread occurring along nerve roots could explain the melioidosis encephalomyelitis syndrome (inflammation of the brain and spinal cord), with brainstem disease following nasal or throat infection, and myelitis (inflammation of the spinal cord) following infection through the skin on the limbs34.
Intercellular spread results in cell fusion and the formation of multinuclear giant cells (MNGCs)84, a hallmark of melioidosis44. Eventual death of MNGCs results in the formation of plaques (one or more MNGCs lyse, leaving a clear zone surrounded by a ring of fused cells) and subsequent damage to host cells, which may serve as a nidus for further B. pseudomallei replication or latent or persistent infection85.
As well as direct cell-to-cell spread, B. pseudomallei can also spread to the bloodstream, causing sepsis, and infect antigen-presenting cells, which then can transport the bacteria to the lymphatic system and contribute to dissemination of infection to secondary sites. However, the exact mechanism of secondary spreading remains elusive. Bacteria also remain viable in dendritic cells, inducing maturation and trafficking to secondary lymphoid organs86.
Latent or persistent infection.
B. pseudomallei can remain latent for extended periods before immunosuppression or other host stress responses reactivate bacterial proliferation and melioidosis develops. Reported latency periods have ranged from 19 years to 29 years87–89, indicating that B. pseudomallei can enter a dormant state and evade immune surveillance44. Neither the site (tissue or subcellular level) of latency nor the mechanisms by which B. pseudomallei remains undetected are clear90. By contrast, high antibody titres detected in patients years after an episode of acute melioidosis suggest continuous exposure or covert sequestration (bacteria hiding in cryptic sites with downregulation of products)91. B. pseudomallei has been found within the nucleus, which could potentially act as a persistence site for later recrudescence92. Strain variability or small colony variants could also play a part in determining whether latent or persistent infection is established93.
Some B. pseudomallei persistence (and possibly latency) factors have been characterized, including toxin-antitoxin systems (composed of a toxin (protein) and its cognate antitoxin (a protein or non-coding RNA)), metabolic enzymes and adaptive mutations94. By entering a slower growth rate, toxin-antitoxin systems enable bacteria to survive under stressful environments, whereas small colony variants can shift to an acid-tolerant state to survive in abscesses95,96. Furthermore, the host’s immune response and selection pressure of antibiotics can contribute to selecting resistance patterns that can also facilitate the establishment of persistent infection (BOX 3). Multiple genotypes have been identified within a single infection episode, which at least partly results from genetic adaptation to the human host, including inactivation of virulence and immunogenic factors and deletion of pathways involved in environmental survival97. Thus, bacterial isolates from patients with persistent or recurrent infection show extensive adaptive regulatory changes that favour bacterial persistence, including genome reduction and increased antibiotic resistance. Data do not yet support a correlation between phages and acquired pathogenicity in B. pseudomallei97,98.
Host immune response
Most patients with melioidosis have at least one predisposing risk factor, suggesting that initiation, progression and outcome of the disease are largely determined by the host’s immune status12,90. For example, genetic polymorphisms in TNF (encoding tumour necrosis factor), NOD2, TLR4 (encoding Toll-like receptor (TLR) 4) and TLR5 have all been linked to disease severity in patients with melioidosis74,99–101. Hypofunctional TLR5 was associated with decreased organ failure, improved survival and a functional cytokine response, possibly mediated by IL-10 ( REF. 102). Interestingly, individuals with the hyporesponsive TLR5 polymorphism display heightened susceptibility to invasive aspergillosis (diseases caused by infection of fungi of the genus Aspergillus) and Legionnaires’ disease (atypical pneumonia caused by Legionella bacteria)103.
Innate immune response.
B. pseudomallei activates the complement pathway, but the bactericidal activity of the complement membrane attack complex is hampered by the external capsular polysaccharides of the bacteria104. Owing to its capsule and lipopolysaccharides (LPSs), B. pseudomallei is also resistant to lysosomal defensins and cationic peptides (which contribute to bacterial killing by disrupting the structure of the cell membrane of the pathogen), enabling survival in human serum and within phagocytes44.
Neutrophil, macrophage and lymphocyte recruitment at the point of infection is triggered by activation of pattern recognition receptors, such as TLRs. Despite the possible detrimental effects of excessive neutrophil recruitment105, activated neutrophils play a pivotal part in early bacterial containment106.
TLRs recognize conserved pathogen-associated molecular patterns (PAMPs) and mediate an inflammatory immune response via various signalling adaptor proteins, including myeloid differentiation primary response protein MyD88 (FIG. 3). MyD88 downregulation in experimental melioidosis increases susceptibility to infection as a result of diminished neutrophil recruitment and activation107. B. pseudomallei triggers the upregulation of multiple TLRs including TLR2, TLR4 (and its co-receptor monocyte differentiation antigen CD14) and TLR5 in host cells, leading to the production of pro-inflammatory and anti-inflammatory cytokines via nuclear factor-κB (NF-κB)108–110. TLR signalling can be dampened or dysregulated by structural variants of LPSs111,112. In addition, LPS recognition seems to be model-dependent: it occurs solely through TLR4 in murine models, whereas in humans, TLR2 has an additional role113.
Phagosomal escape exposes B. pseudomallei to intra-cellular TLR-independent pattern recognition receptors, namely, NOD-like receptors, and activates the formation of the inflammasome, a multimeric protein complex that includes a sensor molecule and caspase 1 (REF. 114). Once the sensor molecule detects B. pseudomallei PAMPs, caspase 1 is activated and rapid pyroptosis ensues115 (FIG. 3). Additionally, the activation of caspase 1 releases active IL-1β and IL-18, which are both increased in patients with septic melioidosis105,116–118. IL-18 contributes to IFNγ induction and, therefore, has a protective effect against B. pseudomallei infection105,116, whereas IL-1β has a potential deleterious role owing to excessive recruitment of neutrophils. This interplay supports intracellular bacterial growth, tissue damage and inhibition of IFNγ production105.
Adaptive immune response.
Although B. pseudomallei antibodies (due to either past or asymptomatic infection) are common in individuals from melioidosis-endemic regions, their role in developing functional immunity to melioidosis is ambiguous, as reinfection from different strains is possible and, therefore, can occur even in the presence of high antibody levels91.
A strong, comprehensive, cell-mediated immune response is essential for protection against progression of infection and for bacterial clearance119. CD4+ T cells are paramount for B cell isotype switching and for activation of cytotoxic CD8+ T cells and macrophages120. Consistent with this, human survivors of melioidosis display increased levels of CD4+ and CD8+ T cells, whereas a decrease in the levels of these cells is specifically correlated with greater mortality. Moreover, vaccines that evoke an immune response skewed towards the activation of T helper 1 (TH1) cells (which promote cellular responses) provide protection against melioidosis, with potential to generate sterilizing immunity (which protects from the onset of both disease and infection)121,122.
Gradually, granulomas (containing neutrophils, macrophages, lymphocytes and MNGCs) form at the site of infection. Intracellular ‘globes’ of bacteria are seen within MNGCs in a background of acute necrotizing inflammation123, which could lead to the development of granulomas. Unfortunately, the dynamic of granuloma formation has not been studied as extensively in melioidosis as in tuberculosis, but it has been recognized as a source of bacterial reactivation in persistent or latent infections. Most clinicians would advocate active investigation of the nature of granulomas incidentally found in patients from melioidosis-endemic regions.
Unlike in infections caused by other organisms with pathogenicity mechanisms similar to those of B. pseudomallei (such as, for example, non-typhoidal Salmonella124), HIV infection does not seem to be a risk factor for melioidosis or for a more-severe or fatal outcome125. In individuals with HIV infection, macrophages show a dysregulated cytokine response (for example, of TNF, IL-10 and IL-12) owing to a low CD4+ T cell count but retain the capacity for bacterial internalization and intracellular killing126. In murine models of melioidosis, depletion of T cells and natural killer cells (hence a 95% reduction in IFNγ production) did not hamper bacterial control, suggesting substantial redundancy of defence mechanisms and the involvement of macrophages expressing major histocompatibility complex (MHC) class II127. For primary melioidosis, it has been suggested that bystander T cell activation is not required for host survival and could play a more substantial part during the antigen-induced activation phase than during the cytokine-mediated activation127. Paradoxically, a strong CD4+ and CD8+ T cell response was observed during acute infection in patients with melioidosis, and lower responses were associated with increased mortality121. Although the current weight of evidence favours a role for T cells in late stages of infection, little is known about the role of specific T cell subsets in regulating the speed of progression or course of B. pseudomallei infection15.
The inflammatory response.
The initial pro-inflammatory response to B. pseudomallei infection is a protective, bacterial killing mechanism. However, a dysregulated cytokine-mediated immune response could result in excessive inflammation with a potentially fatal outcome90. Elevated levels of pro-inflammatory cytokines (such as IL-6, IL-12, IL-15, IL-18, TNF and IFNγ) have been observed in patients with melioidosis, some of which have been correlated with a fatal outcome (for example, IL-6 and IL-18 are considered mortality predictors)116,128. IFNγ production activates T cells and natural killer cells; in murine models, natural killer cells are detected at the site of infection and produce 60–80% of the secreted IFNγ106,129.
Abrogation of TNF or its receptors (in knockout models) results in susceptibility to melioidosis, with increased neutrophil-based inflammatory influx and associated necrosis106,130,131. It is postulated that, in the absence of TNF, a hyperproduction of cytokines and chemokines occurs, leading to septic shock and mortality130.
Anti-inflammatory cytokines (such as IL-10 and IL-4), TNF receptor type I (TNFR1; also known as TNFRSF1A) and IL-1 receptor antagonist (IL1RA) are also upregulated during septic melioidosis. Significant increases in IL-1RA and TNFR1 expression were observed in non-survivors117,132. Fate-onset inflammatory mediators are also correlated with clinical outcome and mortality133,134: in experimental melioidosis, macrophage migration inhibitory factor (MIF) impairs bacterial defence, and neutralizing antibodies against high mobility group protein B1 (HMGB1) could be used as an adjunctive therapy to improve outcome. Coagulation also has a role in melioidosis severity (BOX 4).
Box 4 |. Role of coagulation in melioidosis.
New insights have enhanced our knowledge of the roles of coagulation and fibrinolysis and their interplay with inflammation in the pathogenesis of melioidosis (reviewed in REF. 240). There seems to be a bidirectional role of inflammation and coagulation during melioidosis: activation of coagulation and subsequent fibrin deposition plays an essential part in the host’s defence against infection; however, inflammation-induced coagulation could be detrimental if it is not adequately controlled and could lead to the clinical syndrome of disseminated intravascular coagulation. Plasma levels of anticoagulant vitamin-K-dependent protein C, vitamin-K-dependent protein S and antithrombin 3 are decreased in patients with acute severe melioidosis241,242. High ratios of thrombin–antithrombin complexes over plasmin-α2-antiplasmin complexes (which reflect the consumption of clotting factors and activation (high ratios) or inhibition (low ratios) of fibrinolysis pathways) indicate a predominance of procoagulant mechanisms in melioidosis, and elevated levels of soluble endothelial protein C receptor (whose function is less clear compared with the antithrombotic and anti-inflammatory effects of membrane-bound endothelial protein C receptor) on hospital admission are associated with increased mortality243. Furthermore, mice deficient in plasminogen activator inhibitor 1 (which have increased fibrinolysis and, therefore, decreased fibrin deposition) show heightened susceptibility to Burkholderia pseudomallei244. Activated protein C and the protein C system seem to have a bidirectional role, with a minimal amount of activated protein C required to support an appropriate antibacterial host response, whereas overexpression leads to a harmful phenotype243. Interestingly, the cytoprotective effects of activated protein C are independent of its anticoagulant function. The α2-antiplasmin, a major inhibitor of fibrinolysis, protects from experimental melioidosis by limiting bacterial growth, inflammation, tissue injury and coagulation245. The urokinase-type plasminogen activator receptor, which also plays a crucial part in fibrinolysis, protects against melioidosis by facilitating the migration of neutrophils to the site of infection and subsequently enabling the phagocytosis of B. pseudomallei, further underlying the bidirectional role between coagulation and inflammation in melioidosis240.
Diagnosis, screening and prevention
Melioidosis is grossly underdiagnosed worldwide3, mainly owing to a lack of diagnostic microbiological laboratories serving the low-income rural populations who are at greatest risk of infection and a lack of awareness of the disease amongst physicians and laboratory staff. Even in good microbiological laboratories, B. pseudomallei could initially be discarded as a contaminant (a commensal or non-pathogenic environmental species), especially in non-endemic areas135.
Clinical diagnosis
The incubation period of acute infections is on average 9 days136, ranging from 1–21 days, although a more severe form of the disease with shorter incubation can occur after inhalation or aspiration of contaminated fresh water137. In patients with melioidosis, the clinical presentation, severity and outcome are affected by the presence or absence of risk factors, the route of infection and the bacterial load and strain, as well as the presence or absence of specific non-ubiquitous B. pseudomallei virulence genes12,83. The clinical spectrum of disease varies from localized cutaneous manifestations at the bacterial entry site with no systemic manifestations to sepsis and death (FIG. 4). Bacteraemia on admission occurs in 40–60% of all patients diagnosed with melioidosis, and septic shock occurs in ~20% of all cases. Pneumonia is the presenting illness in about half of all cases. Dissemination of the bacteria to internal organs is common, especially the spleen, prostate, liver and kidneys.
Figure 4 |. Clinical manifestations of melioidosis.
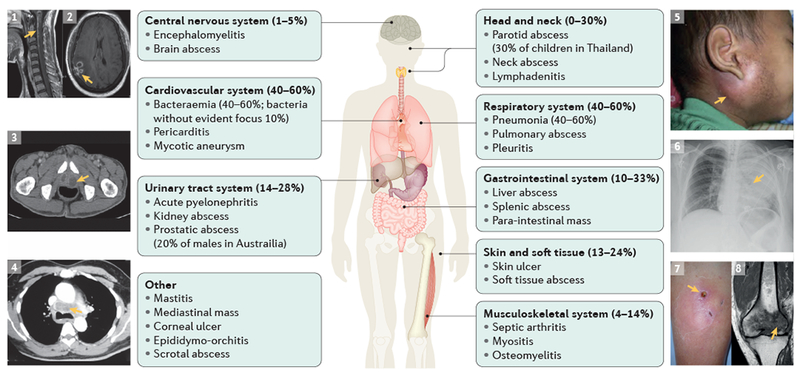
Examples of possible clinical presentations of melioidosis: an MRI of the brainstem and cervical spinal cord with inflammatory changes consistent with encephalomyelitis (arrow, part 1); a ring-enhancing lesion with surrounding oedema in the MRI image indicating cerebral abscesses (arrow, part 2); a CT image of prostatic abscesses (arrow, part 3); a CT image of a mediastinal mass (arrow, part 4); a child with tense parotitis (arrow, part 5); X-ray image of severe pneumonia (arrow, part 6); photo of a subcutaneous abscess (arrow, part 7); and an MRI image of osteomyelitis of the distal femur with surrounding inflammation (arrow, part 8). Clinical images 1–4, 6–8 courtesy of Bart J. Currie, Menzies School of Health, Australia. Clinical image 5 is reproduced with permission from (REF. 249), Elsevier.
Limitations of clinical diagnosis.
Making a diagnosis on clinical grounds alone is very difficult, although in known endemic areas, a patient with suggestive clinical and epidemiological (the presence of risk factors, such as diabetes mellitus and occupational or seasonal exposure) features should be treated empirically with antibiotics that target B. pseudomallei. As the clinical presentation of melioidosis can be nonspecific, the disease should be considered in anyone with a fever in endemic and potentially endemic countries (listed in REF. 3), in particular in individuals with abscesses (especially in the liver, spleen, prostate or parotid) or pneumonia. However, not all patients have risk factors. Laboratory tests are required to confirm the diagnosis of melioidosis138.
Microbiological diagnosis
Culture.
Culture remains the mainstay of melioidosis diagnosis. B. pseudomallei can grow on most routine laboratory media but might not be recognized and could be dismissed as a contaminant, or could be mis-identified as other bacteria (such as Pseudomonas spp. and Bacillus spp.) unless laboratory staff are familiar with its appearance138 (FIG. 5). B. pseudomallei is never part of the normal human flora, and its isolation from any clinical sample should be regarded as diagnostic of melioidosis. As B. pseudomallei is classified as a hazard group 3 pathogen and tier 1 select agent139, doctors should alert the hospital laboratory if melioidosis is suspected in an admitted patient to ensure that appropriate precautions can be followed. All microbiology laboratory staff in melioidosis-endemic areas should receive appropriate training regularly and follow local safety standards.
Figure 5 |. Identification of Burkholderia pseudomallei colonies on three common types of agar.
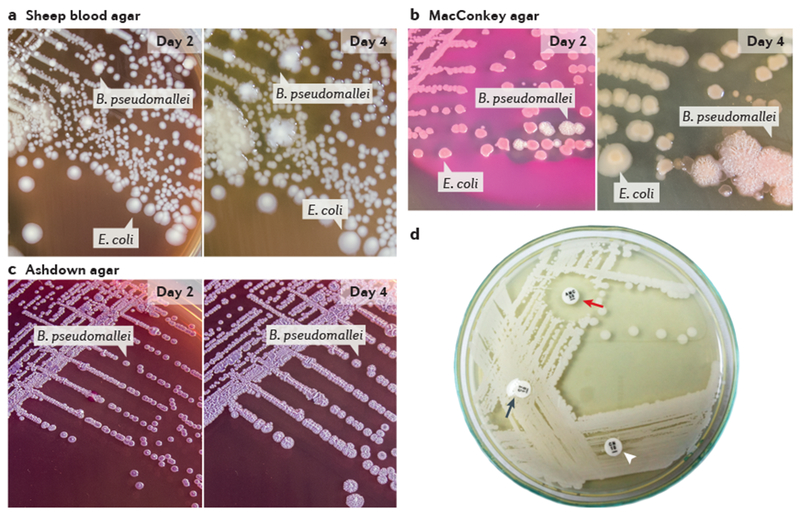
Typical appearance of Burkholderia pseudomallei and Escherichia coli isolated from non-sterile clinical samples. Suspected clinical specimens and suspected bacterial colonies should be processed in a biological safety cabinet. a | B. pseudomallei forms creamy, non-haemolytic colonies that resemble a coliform after 2 days of incubation; by day 4, the colonies are covered by a slight metallic sheen and become dry and wrinkled. b | B. pseudomallei colonies resemble a colourless, non-lactose fermenting coliform after 2 days of incubation; by day 4, the colonies appear dry and wrinkled. c | After 2 days of incubation, the first visible B. pseudomallei colonies are pinpoint with a clear to pale pink colour; by day 4, they become darker pink to purple, flat, slightly dry and wrinkled with a definite metallic sheen. E. coli fails to grow because it is inhibited by gentamicin in the agar. d | Three-disc diffusion antibiotic sensitivity testing: B. pseudomallei is resistant to colistin (or polymyxin) (black arrow) and gentamicin (arrowhead) (although sensitive isolates exist in some areas) and sensitive to co-amoxiclav (red arrow). Parts a–c courtesy of Direk Limmathurotsakul, Premjit Amornchai and Vanaporn Wuthiekanun, Mahidol-Oxford Tropical Medicine Research Unit, Thailand. Part d courtesy of Vanaporn Wuthiekanun, Mahidol-Oxford Tropical Medicine Research Unit, Thailand.
It is crucial that the appropriate clinical samples are collected and sent for culture to laboratories that are familiar with the disease and its causative organism. Blood cultures are the most important sample, as bacteraemia is common. Culture of throat or rectal swabs140,141 and the centrifuged deposit from urine142 are also recommended in all cases of suspected melioidosis, as these could be the only positive samples in some patients. Other samples that should be cultured include pus from abscesses and sputum in patients with pneumonia. Although blood cultures are positive in ~50% of patients overall7 and in up to 75% ofpatients in some reports143,144, this proportion is lower in children38, potentially reflecting that they usually do not have classic melioidosis risk factors. Because cultures have low sensitivity (60%)145, repeating cultures (especially of blood, sputum, urine and pus samples) should be considered in patients with strong indications of B. pseudomallei infection, as it is not uncommon to find subsequent samples positive despite initial negative results. If patients do not improve after 3–7 days of treatment for melioidosis and all culture results are negative, further investigations and a re-evaluation of the diagnosis and treatment should be considered.
B. pseudomallei grows on most routine laboratory media, but more slowly than many other organisms and, therefore, it might be outgrown in samples from sites that normally have a microbial flora; thus, selective media are preferable. Agar plates should be incubated and inspected daily for up to 4 days in suspected cases.
Standard biochemical tests and kit-based identification methods can be used to confirm the identity of colonies. As misidentification of B. pseudomallei with such methods are not uncommon138, the use of multiple methods is recommended. An antibody–antigen binding approach using a monoclonal antibody-based latex agglutination assay is very useful for screening suspect colonies146. Increasing numbers of clinical laboratories use matrix-assisted laser desorption/ionization time of flight (MALDI-TOF) mass spectrometry for bacterial identification, as this method (combined with appropriate databases) can provide an accurate identification of B. pseudomallei147. Numerous molecular approaches for species identification have been described, including 16S rDNA sequencing148 and specific PCRs149, although these tests are often only available in research and reference laboratories. The antimicrobial susceptibility pattern of B. pseudomallei is very characteristic, and in resource-limited areas, a simple disc diffusion antibiotic sensitivity test (to determine the resistance to gentamicin and colistin (or polymyxin) and the susceptibility to amoxicillin-clavulanic acid (also known as co-amoxiclav)) has been recommended to screen Gram-negative rod-shaped bacteria that produce cytochrome oxidase138,143, although it should be noted that gentamicin-susceptible isolates of B. pseudomallei predominate in some regions144.
Direct detection in clinical samples.
Because melioidosis can have severe, if not fatal, consequences, treatment should not be delayed by waiting days for culture results; thus, direct detection of the organism in clinical samples could provide a quick confirmation of the diagnosis. B. pseudomallei can be observed with light microscopy (the organism is often described in textbooks as a Gram-negative rod-shaped bacterium with bipolar staining that resembles a safety pin), but light microscopy lacks sensitivity and specificity150. Immunofluorescent microscopy has a specificity approaching 100%, although the sensitivity is <50% compared with culture151.
Alternative approaches using antigen detection or nucleic acid amplification have also been used. A lateral flow immunoassay that detects the extracellular capsular polysaccharides has been developed152, but it has not yet been extensively evaluated and, although it shows good specificity, it seems to have poor sensitivity, especially for blood specimens153. Numerous PCR assays with high specificity for B. pseudomallei have been developed since the 1990s and have undergone small-scale clinical evaluations. The most promising assay targets the T3SS gene cluster154, although the sensitivity in blood samples depends at least in part on an adequate bacterial concentration155. However, these PCR assays are not routinely used for clinical diagnosis in endemic areas, even in high-income countries such as Singapore and Australia: in addition to sensitivity issues, these tests are not cost-effective in providing the timely confirmation of diagnosis, which clinicians need to make therapeutic decisions.
Serology.
The serological diagnosis of melioidosis is difficult. Many different assays have been developed for detecting antibodies against B. pseudomallei, but most of them are based on poorly characterized antigens and have never been internationally standardized or subjected to extensive critical evaluation. The most widely used is an indirect haemagglutination test (a simple serological test used to detect antibodies raised against B. pseudomallei). The background seropositivity rates in the healthy population in some endemic areas are very high, presumably because of repeated exposures to B. pseudomallei or closely related organisms156,157. As a result, many patients presenting with fever are misdiagnosed with melioidosis in endemic countries in southeast Asia on the basis of a positive indirect haemagglutination test. By contrast, some patients with melioidosis never mount a good antibody response, perhaps because their immune system is compromised. The indirect haemagglutination test on admission has a reported sensitivity of only 56% in Australia158 and 73% in Thailand157, although in the Australian study, 68% of the patients whose tests were negative on admission subsequently showed seroconversion158. Thus, the diagnosis of melioidosis should not rely on the indirect haemagglutination test.
New assays based on purified antigens are being developed and have undergone small-scale evaluations, with some evidence of improved sensitivity and specificity159–161. A protein microarray that contains 20 recombinant and purified B. pseudomallei proteins provides a standardized, easy-to-perform test for the detection of B. pseudomallei-specific antibody patterns162 and could have the potential to improve the serodiagnosis of melioidosis in clinical settings.
Prevention
In northern Australia, basic public health advice is given every year to the general population, and especially to high-risk groups, such as avoiding direct contact with soil and water at the start of each rainy season163. In Thailand, evidence-based guidelines for the prevention of melioidosis recommend that residents, rice farmers and visitors should wear protective gear (such as boots and gloves) if direct contact with soil or water is necessary, only drink bottled or boiled water and avoid outdoor exposure to heavy rain or dust clouds5. The guidelines also encourage cessation of smoking (particularly in those with underlying conditions such as diabetes mellitus) and discourage the application of herbal remedies or organic substances to wounds5. However, the effectiveness of this advice in reducing the incidence of infection has not been proven. Large-scale water chlorination has been very successful in Australia despite theoretical concerns about B. pseudomallei survival in some conditions164. In low-income and middle-income countries, water should be boiled before consumption. Ultraviolet light treatment is effective for remediation of water contaminated with B. pseudomallei and could be recommended in high-income countries in households where individuals are at increased risk of contracting melioidosis165.
However, public awareness of melioidosis in developing tropical countries is limited, and preventive approaches are not always adopted166. A multifaceted intervention at community and government levels is required for successful prevention and is currently being prospectively evaluated in northeast Thailand167.
If high-risk laboratory exposure to B. pseudomallei occurs, post-exposure prophylaxis (PEP) is recommended; high-risk incidents include the exposure of penetrating injuries, mouth or eyes to B. pseudomallei-contaminated materials and the generation of aerosols outside of a biological safety cabinet168. PEP consists of oral antimicrobial treatment with trimethoprim–sulfamethoxazole or, if the organism is resistant or the patient is intolerant, doxycycline or co-amoxiclav for 21 days168. The potential benefit of PEP must be weighed against the fact that trimethoprim–sulfamethoxazole can have severe adverse effects: for individuals involved in a low-risk incident, the decision to begin PEP should be based on the presence of known risk factors for naturally acquired melioidosis. Individuals with known risk factors should be advised to receive PEP, whereas in the absence of known risk factors monitoring is sufficient143,168.
Management
Early diagnosis and the start of antimicrobial therapy specific to B. pseudomallei are crucial for melioidosis treatment. In locations with resources for rapid diagnosis, early implementation of optimal antibiotic therapy and state-of-the-art intensive care facilities for managing severe sepsis, mortality is ~10%12. Nevertheless, such resources are not available or are limited in many endemic regions, and in those circumstances, mortality is ≥40%12. The majority of B. pseudomallei isolates from primary infections have the same characteristic antimicrobial susceptibility profiles. B. pseudomallei is susceptible to β-lactam antibiotics (such as ceftazidime, meropenem, imipenem and co-amoxiclav), although the bactericidal activity of these drugs varies, and is almost always susceptible to doxycycline, chloramphenicol and trimethoprim–sulfamethoxazole169–171, although these agents only have bacteriostatic activity. Most isolates are susceptible in vitro to piperacillin, ceftriaxone and cefotaxime, but these agents are less effective clinically172. However, B. pseudomallei is resistant to penicillin, ampicillin, first-generation and second-generation cephalosporins, gentamicin, tobramycin, streptomycin, macrolides and polymyxins (BOX 3). Of note, clonal groups of isolates susceptible to gentamicin are common in Sarawak, Malaysia144.
On the back of the global concerns of antimicrobial resistance and the already limited options for treating melioidosis, new antimicrobials have been tested in vitro and in animal models, but none can yet replace ceftazidime and meropenem173. Doripenem has minimum inhibitory concentrations (the lowest concentrations that can prevent visible bacterial growth after over-night incubation) similar to meropenem, but ertapenem, tigecycline and moxifloxacin seem to have limited in vitro activity174.
As melioidosis is not a contagious disease, isolation of patients or special precautions are usually not required within endemic areas. However, as few nosocomial infections have been reported24,175,176, healthcare providers are recommended to follow universal precautions177 and standard infection control practices, including hand hygiene178. Potential contamination of the ward and local environment from patients with superficial lesions or pneumonia has been raised as a concern, but such contamination has never been documented.
Formal guidelines for melioidosis therapy, including recommended dosage and duration of each therapeutic phase, have been published by the CDC after a 2010 expert workshop that updated prior consensus-based guidelines168. Antimicrobial therapy consists of the initial intensive phase and the subsequent eradication phase (BOX 5).
Box 5 |. Antibiotic therapy for treatment of melioidosis.
Initial intensive therapy
Initial intensive therapy should last for a minimum of 10–14 days and consists of ceftazidime (in wards) 2 g (50 mg kg−1 up to 2 g in children (<15 years of age)) intravenous, 6-hourly; or meropenem (in intensive care units) 1 g (25 mg kg−1 up to 1 g in children) intravenous, 8-hourly.
Meropenem is the preferred initial intravenous therapy for neurological melioidosis (that is, in the presence of infection of the central nervous system), and the dose should be doubled.
Consider adding trimethoprim–sulfamethoxazole (which has excellent tissue penetration) in the doses recommended for eradication therapy from the start of the initial intensive therapy in cases of neurological melioidosis, osteomyelitis and septic arthritis, skin and soft tissue infections and genitourinary infection including prostatic abscesses12,168.
Long-term intravenous therapy (≥4–8 weeks) is recommended* where possible for complicated pneumonia, deep-seated infection (including prostatic abscesses), neurological melioidosis, osteomyelitis and septic arthritis168,246.
Eradication therapy
The eradication therapy should last for ≥3 months after the end of the initial intensive therapy and consists of trimethoprim–sulfamethoxazole‡ 6 + 30 mg kg−1 up to 240 + 1,200 mg in children, 240 + 1,200 mg in adults 40–60 kg and 320 + 1,600 mg in adults >60 kg orally, 12-hourly, and folic acid 5 mg (0.1 mg kg−1 up to 5 mg in children) orally, daily.
Longer eradication therapy (≥6 months) is recommended for neurological melioidosis and osteomyelitis.
*This treatment guidance is consistent with the most up-to-date recommendations by the International Melioidosis Society (http://www.melioidosis.info). Recommendations derived from Australian studies121,88 and apply to resource-rich countries. In melioidosis-endemic regions, such prolonged intravenous therapy is often either not available or not affordable. Nevertheless, a minimum of 10 days of intravenous therapy is recommended for all individuals with melioidosis, except for those with localized skin disease without sepsis168,171.
‡The trimethoprim–sulfamethoxazole dose is usually expressed as separate doses of each individual drug.
Initial intensive therapy
Intravenous ceftazidime or meropenem is the preferred choice179; the duration of initial intensive therapy should last a minimum of 10–14 days, with longer intensive therapy for critically ill patients, including those with extensive pulmonary disease, deep-seated collections or organ abscesses, osteomyelitis (infection of bone), septic arthritis or neurological melioidosis (BOX 5). The therapeutic response can be slow; the median time to resolution of fever is up to 9 days180, with longer times in patients with deep-seated abscesses. The addition of trimethoprim–sulfamethoxazole to ceftazidime for the intensive phase is used by some physicians for certain types of infection (BOX 5) but conferred no survival benefit in studies in Thailand181,182.
Imipenem and meropenem have the lowest minimum inhibitory concentrations against B. pseudomallei and have faster bacterial killing rates than ceftazidime in vitro183,184. The recommendation of meropenem as the drug of choice for severe melioidosis with septic shock is also supported by observational data from Australia185. However, ceftazidime remains the drug of choice for initial therapy for most patients with melioidosis, and there is no conclusive evidence that meropenem is superior to ceftazidime in patients who are not critically ill. In Australia, meropenem is switched to ceftazidime when those patients with severe disease recover and are well enough to be discharged from the intensive care unit to general wards.
Eradication therapy
After the initial intensive therapy, subsequent eradication therapy with oral antibiotics is recommended to prevent recrudescence of the disease or relapse of the patient. Trimethoprim–sulfamethoxazole is the preferred agent for eradication therapy (BOX 5), and co-amoxiclav or doxycycline is the second choice. Reports of primary resistance to trimethoprim–sulfamethoxazole in >10% of B. pseudomallei isolates from Thailand and other southeast Asian countries were demonstrated to be inaccurate171.
Based on empirical experience and therapeutic modelling, dosing recommendations for trimethoprim–sulfamethoxazole186 and co-amoxiclav187 in melioidosis are higher than the standard doses used with these antibiotics. Consequently, owing to the long-term course of trimethoprim–sulfamethoxazole required, adverse effects are reported in up to 40% of patients10. In Thailand, this drug is usually avoided in patients who have glucose-6-phosphate dehydrogenase (G6PD) deficiency, owing to the risk of haemolytic anaemia, although patients are not routinely screened for G6PD activity prior to treatment. Rash, gastrointestinal symptoms, hyperkalaemia (high serum potassium levels that can lead to muscle weakness and arrhythmias) and rising levels of serum creatinine (which could indicate renal dysfunction) could require dose modification or cessation of trimethoprim–sulfamethoxazole therapy, which can be replaced by doxycycline or co-amoxiclav. Desensitization (a strategy to safely induce drug tolerance and limit the possibility of a type I hypersensitivity reaction) should be considered for non-severe skin reactions attributed to trimethoprim–sulfamethoxazole.
In Australia, trimethoprim–sulfamethoxazole is the preferred eradication therapy for children and potentially in pregnant women after the first trimester (owing to the risk of neural tube or other congenital defects), whereas in some locations in Thailand co-amoxiclav has been used for eradication therapy in children and in pregnancy. However, dosing with co-amoxiclav is problematic, and acquired resistance is well documented when co-amoxiclav or doxycycline is used4.
Treatment duration
Lengthening the duration of the initial intensive therapy for patients with more-severe melioidosis has contributed to the decrease in mortality in regions with the necessary healthcare resources. A retrospective analysis of patients who were treated according to the Royal Darwin Hospital melioidosis treatment guidelines, which define the minimum recommended duration of initial intensive therapy based on the clinical presentation188, supports a longer intravenous therapy for critically ill patients. The median duration of the initial intensive therapy for such patients in that analysis was ~4 weeks and only 5 patients (1.2%) relapsed. Although non-compliance of patients during eradication therapy is common as patients stop taking the antibiotics early, skip doses or do not take any drug at all after discharge, the relapse rates in the Darwin study are low and are attributed to prolongation of the initial intensive therapy188. Studies are required to assess whether future guidelines can include options based solely on intravenous therapy without a long-term eradication phase.
Studies from Thailand showed that failure of the eradication therapy is associated with poor adherence to therapy, more-severe infections (for example, multifocal disease and bacteraemia) and a duration of eradication therapy of <8 weeks189–191. These findings support the recommendations for an eradication therapy of 3–6 months (BOX 5). Case series reported that selected patients with localized cutaneous disease could be successfully treated with oral trimethoprim–sulfamethoxazole for 3 months without preceding initial intensive therapy34,192,193. However, such a regimen must be restricted to patients who have no signs of sepsis or organ dysfunction, no underlying risk factors and no dissemination of the infection to distant sites, including regional lymph nodes.
Surgical aspects of therapy
Surgical drainage is usually required for single, large abscesses in the liver and muscles and prostatic abscesses194, but it is not necessary or possible for multiple small abscesses in the spleen, liver and kidneys. Septic arthritis (inflammation of the joints due to the infection) usually requires drainage and washout and might require repeated procedures. Other internal abscesses rarely need to be drained as they frequently resolve with medical therapy. Osteomyelitis can be very extensive when diagnosis and appropriate antibiotic therapy are delayed, and in such cases, aggressive and often repeated surgical debridement of the necrotic bone is usually necessary195. However, early long-bone osteomyelitis without abscess formation and vertebral osteomyelitis without epidural abscess might not require debridement. Mycotic aneurysms (caused by bacterial infiltration of the arterial wall) require urgent surgery, often with insertion of prosthetic vascular grafts. Lifelong suppressive therapy with trimethoprim–sulfamethoxazole might be indicated for patients who have received prosthetic grafts for mycotic aneurysms.
Adjuvant therapy
State-of-the-art intensive care management can sub-stantially decrease mortality in patients with melioidosis and sepsis or septic shock196,197, and appropriate guidelines are available198.
Granulocyte colony-stimulating factor (G-CSF, which stimulates the production of neutrophils in the bone marrow) has been used empirically in patients with septic shock. The rationale is to counteract the functional neutrophil defects that are thought to be crucial in the pathogenesis of severe melioidosis. Early observational data showed a significant improvement in survival with G-CSF, but this result was confounded by concomitant improvements in other clinical parameters199. A subsequent randomized controlled trial in Thailand showed no overall survival benefit with addition of G-CSF200; nevertheless, survival was significantly longer in the G-CSF group in settings with limited intensive care resources, and adjuvant G-CSF is still used for melioidosis-associated septic shock in some hospitals with state-of-the-art intensive care facilities. Preclinical studies have shown that administration of clinically available IL-1β blocking agents can protect mice against overt B. pseudomallei infection and mortality105,201. This finding is in line with reports that patients with melioidosis who have diabetes mellitus and take glibenclamide (also known as glyburide) have a lower mortality and attenuated inflammatory responses compared with patients who do not take glibenclamide202 (BOX 1). Given the crucial role of immune function in melioidosis pathogenesis, patients with melioidosis-associated sepsis or septic shock could benefit from newly available immune-modulating therapies, such as IL-7, granulocyte-macrophage colony-stimulating factor and anti-PD1 (programmed cell death protein 1).
Quality of life
Recrudescence and recurrence of melioidosis
Recrudescent melioidosis with a return of clinical illness and culture positivity can occur during the eradication treatment period if the initial intensive therapy is ceased too early, internal abscesses have not been diagnosed or adequately drained or if the oral eradication therapy is not adhered to or is ceased too early. Patients with recrudescence will need hospital admission, intravenous parenteral antimicrobials and re-examination for sites of infection.
Once both initial intensive and eradication therapies have been successfully completed, recurrent melioidosis could be due to either recrudescence of the original infection, as confirmed by bacterial isolate genotyping, or a new infection with a different strain190,203. With improvements in therapy, recrudescence decreased from ~10% of cases to <5%; new infections in survivors of melioidosis are now more common than recrudescence188.
Long-term sequelae
For survivors of melioidosis, the main determinants of future health are their underlying risk factors that predisposed them to the initial infection. Many of the patients in the Darwin Prospective Melioidosis Study have subsequently died or had a substantial and ongoing disability as a consequence of comorbidities such as diabetes mellitus, chronic kidney disease or malignancy (B.J.C., unpublished observation). The most severe melioidosis-associated disability is residual neurological deficit subsequent to melioidosis-associated encephalomyelitis. Although rare, neurological complications are particularly problematic in children and can range from severe residual quadriparesis (muscle weakness in all limbs) or severe flaccid paraparesis (partial or complete paralysis of both legs) to persisting isolated foot drop (an abnormal gait in which the forefoot drops owing to weakness)34. Limited range of motion, sinus tract (an infected tract connecting a deep-seated infection to the surface or another organ and often discharging pus) formation and joint deformities are also common in patients with bone and joint infections195,204.
Outlook
Melioidosis as a neglected disease
Neglected tropical diseases are understudied diseases that remain endemic in many developing countries around the world205. Melioidosis is not included in most lists of neglected tropical diseases, including the WHO list205, even though it has high mortality and is potentially preventable and treatable3. Thus, efforts by the international research community are needed to raise awareness of melioidosis within the WHO and regional and local health agencies, as well as in the general public in endemic areas. In 2015, the International Melioidosis Society (IMS) was formed by melioidosis researchers to raise awareness and knowledge of the disease amongst all stakeholders, and in 2016, a Research Collaboration Network (RCN) was formed to bring the disease to the attention of public health officials and policy makers in melioidosis-endemic countries. An interactive map and disease information are available on the IMS-RCN website (http://www.melioidosis.info).
Melioidosis in developing countries
Diagnosis.
In developing countries, melioidosis is most common amongst members of the rural population, who often have limited access to even simple diagnostics. Furthermore, even if clinical microbiology laboratories are available, they might be underused owing to costs or lack of trained staff206. Non-culture-based methods to diagnose bacterial infectious diseases are increasingly being encouraged207, but they have not been extensively developed and evaluated for melioidosis. Thus, the priorities for improving melioidosis case ascertainment in tropical endemic areas are the establishment of basic clinical microbiology laboratories and education of both clinicians and laboratory technicians about melioidosis and prevention.
Management.
To reduce acute melioidosis mortality, the availability and affordability of ceftazidime or carbapenems need to improve. In Thailand, an upper-middle-income country, a 14-day course of ceftazidime or carbapenem costs ~US$60 or US$1,080, respectively, and is covered by Thailand’s Universal Coverage Scheme208. Nonetheless, in other low-income and middle-income countries, these drugs could be more expensive, have limited availability or be excluded from the country’s universal health coverage, and patients frequently cannot afford to pay for the drugs themselves200. Studies on how best to allocate and use resources for melioidosis in low-income and middle-income countries are urgently needed.
The One Health Initiative
In 2016, melioidosis was highlighted as a sapronosis, a disease of animals and humans caused by an environmental organism; however, the disease in animals and geographical distribution of distinct human clinical manifestations (for example, parotitis is mainly observed in children within southeast Asia and encephalomyelitis mainly in northern Australia) are not well understood210. Thus, adopting the One Health Initiative (a strategy involving interdisciplinary collaborations of health professionals at the local, national and global levels in all aspects of healthcare for humans, animals and the environment211) will promote cooperation and strategic planning between physicians, ecologists, environmental scientists and veterinarians. Furthermore, interdisciplinary efforts will help to address the sapronotic spread of the disease and to establish effective melioidosis interventions.
To implement One Health Initiative approaches, potential technology-based solutions, such as wireless and mobile technologies for the delivery of health interventions and education and the interactive tools currently displayed on the aforementioned melioidosis website (http://www.melioidosis.info), should be implemented in resource-limited settings. One Health Initiatives for melioidosis could also lead to a broader engagement of organizations and individuals with experience in prevention, surveillance and clinical case management of neglected tropical diseases, as well as individuals with training in economic development, genomics, veterinary sciences, wildlife management, agriculture, molecular biology and bacteriology, ecology, policy and law211. The implementation of this interdisciplinary initiative would combine field efforts addressing both endemic and emerging melioidosis but requires effective global health governance210.
Advances in vaccine development
A vaccine could be a cost-effective intervention in tropical developing countries, particularly if used in high-risk populations such as individuals with diabetes mellitus, even if it produces only partial immunity212. The Steering Group on Melioidosis Vaccine Development has highlighted the need for standardized animal models and for advancing melioidosis vaccine candidates to preclinical and clinical studies, and the challenges posed by bacterial strains and routes of immunization. Furthermore, it recognized that vaccines designed for the military or other populations involved in biodefense have different requirements from those designed to prevent naturally acquired melioidosis in endemic areas213.
No melioidosis vaccine is currently available for human use; some vaccine candidates have been shown to provide partial protection against melioidosis or glanders in murine models of infection212,214,215, but few have been tested in non-human primates or humans216. Live attenuated vaccines induce a more-comprehensive immune response in animal models and are currently considered the best approach for generating protection against B. pseudomallei infection16,217. However, subunit-based vaccines provide a feasible alternative because of their increased safety and potential for large-scale production. Experimental evidence indicates that the combination of bacterial polysaccharides (LPSs or other capsular polysaccharides) with well-defined protein antigens (glycoconjugates) can generate substantial protection against Burkholderia infections217. This rapid advancement in vaccine design and optimization is very promising, and several vaccine candidates are currently being tested. However, a multivalent vaccine containing numerous immunogenic bacterial components will probably be necessary to achieve complete protection, as, in addition to a strong antibody response, both CD4+ and CD8+ T cells are also important in protection against human melioidosis.
Final thoughts
New global environmental sampling studies and improvements in diagnostic microbiology could enhance the understanding of the geographical distribution and burden of B. pseudomallei, and wide-scale whole-genome sequencing together with clinical details could provide new insights into the phylogeny and virulence of these bacteria. The application of such new techniques to isolates with well-characterized clinical and epidemiological metadata could provide further insights into melioidosis. New omics-based technologies will enable a better understanding of how B. pseudomallei evades immune surveillance and can remain latent for many years. The role of the human and environmental microbiota is only just beginning to emerge218 and could offer creative insights into how to tackle the infection in vivo and reduce exposure in terra.
Affordable, effective alternative drugs and new drug targets are needed to reduce mortality, relapse and the duration of treatment courses. More than 80% of individuals with diabetes mellitus live in low-income and middle-income countries, and the numbers are projected to increase by >55% globally by 2050, with tropical countries facing the brunt of this epidemic219. Case clusters of melioidosis have also been associated with severe weather events220,221, which, based on the current estimates of global climate change, might become increasingly common. This combination could cause a surge of melioidosis, particularly in countries where it has previously been under-reported, such as India. Calculating a representative disability-adjusted life year metric could also provide better comparative burden information and prompt regulating bodies to recognize melioidosis as a neglected tropical disease.
Acknowledgements
W.J.W. received a Vidi grant (91716475) from the Netherlands Organisation for Scientific Research (NWO) and Marie Curie Skledowska Innovative Training Network (MC-ITN) European Sepsis Academy, funded by the European Union’s Horizon 2020 programme. H.S.V. received a Marie Curie Skledowska fellowship under the European Sepsis Academy, funded by the European Union’s Horizon 2020 programme. A.G.T. is supported by the NIH and the National Institute of Allergy and Infectious Diseases (NIAID) R01 grant AI12660101. B.J.C. is supported by Australian National Health and Medical Research Council grants, including the HOT NORTH initiative. S.J.P. is an NIH Research Senior Investigator. D.A.B.D. is supported by The Wellcome Trust of Great Britain (grant number 106698/Z/14/Z). D.L. is supported by The Wellcome Trust Public Health and Tropical Medicine Intermediate Fellowship (grant number 101103/Z/13/Z). The authors thank G. Wongsuvan, P. Amornchai, P. Wannapinij and V. Wuthiekanun for their assistance with the images in FIG. 5.
Footnotes
Competing interests statement
D.A.B.D. acted as a consultant to Soligenix, Inc. All other authors declare no competing interests.
References
- 1.Whitmore A An Account of a Glanders-like Disease occurring in Rangoon. J. Hyg 13, 1–34.1 (1913). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 2.Kaestli M et al. Out of the ground: aerial and exotic habitats of the melioidosis bacterium Burkholderia pseudomallei in grasses in Australia. Environ. Microbiol 14, 2058–2070 (2012). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 3.Limmathurotsakul D et al. Predicted global distribution of Burkholderia pseudomallei and burden of melioidosis. Nat. Microbiol 1, 15008 (2016). [DOI] [PubMed] [Google Scholar]; This is a key publication on the global burden of melioidosis that uses human, animal and environmental data to estimate the number of human melioidosis cases per year at 165,000 worldwide, of which 89,000 are fatal.
- 4.Wiersinga WJ, Currie BJ & Peacock SJ Melioidosis. N. Engl. J. Med 367, 1035–1044 (2012). [DOI] [PubMed] [Google Scholar]; This article reviews the clinical manifestations, epidemiology, pathogenesis, diagnosis and treatment of melioidosis, with an emphasis on clinical management.
- 5.Limmathurotsakul D et al. Activities of daily living associated with acquisition of melioidosis in northeast Thailand: a matched case-control study. PLoS Negl. Trop. Dis 7, e2072 (2013). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 6.Cheng AC & Currie BJ Melioidosis: epidemiology, pathophysiology, and management. Clin. Microbiol. Rev 18, 383–416 (2005). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 7.Currie BJ, Ward L & Cheng AC The epidemiology and clinical spectrum of melioidosis: 540 cases from the 20 year Darwin prospective study. PLoS Negl. Trop. Dis 4, e900 (2010). [DOI] [PMC free article] [PubMed] [Google Scholar]; This is the Darwin prospective study of melioidosis that has provided numerous new insights into the epidemiology and clinical spectrum of melioidosis. This article concludes that melioidosis should be regarded as an opportunistic infection that is unlikely to kill a healthy person in a resource-rich environment, provided the infection is diagnosed early.
- 8.Yee KC, Lee MK, Chua CT & Puthucheary SD Melioidosis, the great mimicker: a report of 10 cases from Malaysia. J. Trop. Med. Hyg 91, 249–254 (1988). [PubMed] [Google Scholar]
- 9.Maharjan B et al. Recurrent melioidosis in patients in northeast Thailand is frequently due to reinfection rather than relapse. J. Clin. Microbiol 43, 6032–6034 (2005). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 10.Chetchotisakd P et al. Trimethoprim-sulfamethoxazole versus trimethoprim-sulfamethoxazole plus doxycycline as oral eradicative treatment for melioidosis (MERTH): a multicentre, double-blind, non-inferiority, randomised controlled trial. Lancet 383, 807–814 (2014). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 11.Suputtamongkol Y et al. Amoxycillin-clavulanic acid treatment of melioidosis. Trans. R. Soc. Trop. Med. Hyg 85, 672–675 (1991). [DOI] [PubMed] [Google Scholar]
- 12.Currie BJ Melioidosis: evolving concepts in epidemiology, pathogenesis, and treatment. Semin. Respir. Crit. Care Med 36, 111–125 (2015). [DOI] [PubMed] [Google Scholar]
- 13.Willcocks SJ, Denman CC, Atkins HS & Wren BW Intracellular replication of the well-armed pathogen Burkholderia pseudomallei. Curr. Opin. Microbiol 29, 94–103 (2016). [DOI] [PubMed] [Google Scholar]
- 14.Centers for Disease Control and Prevention. Possession, use, and transfer of select agents and toxins; biennial review. Final rule. Fed. Regist 77, 61083–61115 (2012). [PubMed] [Google Scholar]
- 15.Silva EB & Dow SW Development of Burkholderia mallei and pseudomallei vaccines. Front. Cell. Infect. Microbiol 3, 10 (2013). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 16.Titball RW, Burtnick MN, Bancroft GJ & Brett P Burkholderia pseudomallei and Burkholderia mallei vaccines: are we close to clinical trials? Vaccine 35, 5981–5989 (2017). [DOI] [PubMed] [Google Scholar]; This is an up-to-date summary of vaccine research and front-line contenders with potential for success.
- 17.Limmathurotsakul D et al. Systematic review and consensus guidelines for environmental sampling of Burkholderia pseudomallei. PLoS Negl. Trop. Dis 7, e2105 (2013). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 18.Pumpuang A et al. Survival of Burkholderia pseudomallei in distilled water for 16 years. Trans. R. Soc. Trop. Med. Hyg 105, 598–600 (2011). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 19.Hantrakun V et al. Soil nutrient depletion is associated with the presence of Burkholderia pseudomallei. Appl. Environ. Microbiol 82, 7086–7092 (2016). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 20.Yip TW et al. Endemic melioidosis in residents of desert region after atypically intense rainfall in central australia, 2011. Emerg. Infect. Dis 21, 1038–1040 (2015). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 21.Currie BJ et al. A cluster of melioidosis cases from an endemic region is clonal and is linked to the water supply using molecular typing of Burkholderia pseudomallei isolates. Am. J. Trop. Med. Hyg 65, 177–179 (2001). [DOI] [PubMed] [Google Scholar]
- 22.Inglis TJ et al. Acute melioidosis outbreak in Western Australia. Epidemiol. Infect 123, 437–443 (1999). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 23.Limmathurotsakul D et al. Melioidosis caused by Burkholderia pseudomallei in drinking water, Thailand, 2012. Emerg. Infect. Dis 20, 265–268 (2014). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 24.Merritt AJ et al. Cutaneous melioidosis cluster caused by contaminated wound irrigation fluid. Emerg. Infect. Dis 22, 1420–1427 (2016). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 25.Gal D et al. Contamination of hand wash detergent linked to occupationally acquired melioidosis. Am. J. Trop. Med. Hyg 71, 360–362 (2004). [PubMed] [Google Scholar]
- 26.Kinoshita RE Epidemiology of melioidosis in an oceanarium: a clinical, environmental & molecular study. Thesis, Univ. of Hong Kong; (2003). [Google Scholar]
- 27.Chen PS. et al. Airborne transmission of melioidosis to humans from environmental aerosols contaminated with B. pseudomallei. PLoS Negl. Trop. Dis 9, e0003834 (2015). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 28.Currie BJ et al. Use of whole-genome sequencing to link Burkholderia pseudomallei from air sampling to mediastinal melioidosis, Australia. Emerg. Infect. Dis 21, 2052–2054 (2015). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 29.Thatrimontrichai A & Maneenil G Neonatal melioidosis: systematic review of the literature. Pediatr. Infect. Dis. J 31, 1195–1197 (2012). [DOI] [PubMed] [Google Scholar]
- 30.Rolim DB et al. Melioidosis, northeastern Brazil. Emerg. Infect. Dis 11, 1458–1460 (2005). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 31.Salam AP et al. Melioidosis acquired by traveler to Nigeria. Emerg. Infect. Dis 17, 1296–1298 (2011). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 32.Birnie E, Wiersinga WJ, Limmathurotsakul D & Grobusch MP Melioidosis in Africa: should we be looking more closely? Future Microbiol. 10, 273–281 (2015). [DOI] [PubMed] [Google Scholar]
- 33.Wiersinga WJ et al. Clinical, environmental, and serologic surveillance studies of melioidosis in Gabon, 2012–2013. Emerg. Infect. Dis 21, 40–47 (2015). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 34.McLeod C et al. Clinical presentation and medical management of melioidosis in children: a 24-year prospective study in the Northern Territory of Australia and review of the literature. Clin. Infect. Dis 60, 21–26 (2015). [DOI] [PubMed] [Google Scholar]
- 35.Limmathurotsakul D et al. Increasing incidence of human melioidosis in Northeast Thailand. Am. J. Trop. Med. Hyg 82, 1113–1117 (2010). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 36.Currie BJ et al. Melioidosis epidemiology and risk factors from a prospective whole-population study in northern Australia. Trop. Med. Int. Health 9, 1167–1174 (2004). [DOI] [PubMed] [Google Scholar]
- 37.Fong SM, Wong KJ, Fukushima M & Yeo TW Thalassemia major is a major risk factor for pediatric melioidosis in Kota Kinabalu, Sabah, Malaysia. Clin. Infect. Dis 60, 1802–1807 (2015). [DOI] [PubMed] [Google Scholar]
- 38.Turner P et al. A retrospective analysis of melioidosis in Cambodian children, 2009–2013. BMC Infect. Dis 16, 688 (2016). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 39.Lim MK Tan EH Soh CS. & Chang TL. Burkholderia pseudomallei infection in the Singapore Armed Forces from 1987 to 1994 — an epidemiological review. Ann. Acad. Med. Singapore 26, 13–17 (1997). [PubMed] [Google Scholar]
- 40.Ooi WF et al. The condition-dependent transcriptional landscape of Burkholderia pseudomallei. PLoS Genet. 9, e1003795 (2013). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 41.Wiersinga WJ, van der Poll T, White NJ, Day NP & Peacock SJ Melioidosis: insights into the pathogenicity of Burkholderia pseudomallei. Nat. Rev. Microbiol 4, 272–282 (2006). [DOI] [PubMed] [Google Scholar]
- 42.Allwood EM, Devenish RJ, Prescott M, Adler B & Boyce JD Strategies for intracellular survival of Burkholderia pseudomallei. Front. Microbiol 2, 170 (2011). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 43.Stone JK, DeShazer D, Brett PJ & Burtnick MN. Melioidosis: molecular aspects of pathogenesis. Expert Rev. Anti Infect. Ther 12, 1487–1499 (2014). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 44.Lazar Adler NR et al. The molecular and cellular basis of pathogenesis in melioidosis: how does Burkholderia pseudomallei cause disease? FEMS Microbiol. Rev 33, 1079–1099 (2009). [DOI] [PubMed] [Google Scholar]
- 45.Sun GW & Gan YH Unraveling type III secretion systems in the highly versatile Burkholderia pseudomallei. Trends Microbiol. 18, 561–568 (2010). [DOI] [PubMed] [Google Scholar]
- 46.Stevens MR et al. An Inv/Mxi-Spa-like type III protein secretion system in Burkholderia pseudomallei modulates intracellular behaviour of the pathogen. Mol. Microbiol 46, 649–659 (2002). [DOI] [PubMed] [Google Scholar]
- 47.Burtnick MN, Brett PJ & DeShazer D Proteomic analysis of the Burkholderia pseudomallei type II secretome reveals hydrolytic enzymes, novel proteins, and the deubiquitinase TssM. Infect. Immun 82, 3214–3226 (2014). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 48.Campos CG, Byrd MS & Cotter PA Functional characterization of Burkholderia pseudomallei trimeric autotransporters. Infect. Immun 81, 2788–2799 (2013). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 49.Ahmed K et al. Attachment of Burkholderia pseudomallei to pharyngeal epithelial cells: a highly pathogenic bacteria with low attachment ability. Am. J. Trop. Med. Hyg 60, 90–93 (1999). [DOI] [PubMed] [Google Scholar]
- 50.Essex-Lopresti AE et al. A type IV pilin, PilA, contributes to adherence of Burkholderia pseudomallei and virulence in vivo. Infect. Immun 73, 1260–1264 (2005). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 51.Phewkliang A, Wongratanacheewin S & Chareonsudjai S Role of Burkholderia pseudomallei in the invasion, replication and induction of apoptosis in human epithelial cell lines. Southeast Asian J. Trop. Med. Public Health 41, 1164–1176(2010). [PubMed] [Google Scholar]
- 52.David J, Bell RE & Clark GC Mechanisms of disease: host-pathogen interactions between Burkholderia species and lung epithelial cells. Front. Cell. Infect. Microbiol 5, 80 (2015). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 53.Chuaygud I, Tungpradabkul S, Sirisinha S, Chua KL & Utaisincharoen P A role of Burkholderia pseudomallei flagella as a virulent factor. Trans. R. Soc. Trop. Med. Hyg 102 (Suppl. 1), S140–S144 (2008). [DOI] [PubMed] [Google Scholar]
- 54.Balder R et al. Identification of Burkholderia mallei and Burkholderia pseudomallei adhesins for human respiratory epithelial cells. BMC Microbiol. 10, 250 (2010). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 55.Stevens MP et al. A Burkholderia pseudomallei type III secreted protein, BopE, facilitates bacterial invasion of epithelial cells and exhibits guanine nucleotide exchange factor activity. J. Bacteriol 185, 4992–4996 (2003). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 56.Kager LM, Wiersinga WJ, Roelofs JJ, van ‘t Veer C & van der Poll T Deficiency of protease-activated receptor-1 limits bacterial dissemination during severe Gram-negative sepsis (melioidosis). Microbes Infect. 16, 171–174 (2014). [DOI] [PubMed] [Google Scholar]
- 57.Pruksachartvuthi S, Aswapokee N & Thankerngpol K Survival of Pseudomonas pseudomallei in human phagocytes. J. Med. Microbiol 31, 109–114(1990). [DOI] [PubMed] [Google Scholar]
- 58.Jones AL, Beveridge TJ & Woods DE Intracellular survival of Burkholderia pseudomallei. Infect. Immun 64, 782–790 (1996). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 59.Harley VS, Dance DA, Tovey G, McCrossan MV & Drasar BS An ultrastructural study of the phagocytosis of Burkholderia pseudomallei. Microbios 94, 35–45(1998). [PubMed] [Google Scholar]
- 60.Kespichayawattana W, Rattanachetkul S, Wanun T, Utaisincharoen P & Sirisinha S Burkholderia pseudomallei induces cell fusion and actin-associated membrane protrusion: a possible mechanism for cell-to-cell spreading. Infect. Immun 68, 5377–5384 (2000). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 61.Ray K, Marteyn B, Sansonetti PJ & Tang CM Life on the inside: the intracellular lifestyle of cytosolic bacteria. Nat. Rev. Microbiol 7, 333–340 (2009). [DOI] [PubMed] [Google Scholar]
- 62.Stevens MP et al. Attenuated virulence and protective efficacy of a Burkholderia pseudomallei bsa type III secretion mutant in murine models of melioidosis. Microbiology 150, 2669–2676 (2004). [DOI] [PubMed] [Google Scholar]
- 63.Sun GW, Lu J, Pervaiz S, Cao WP & Gan YH Caspase-1 dependent macrophage death induced by Burkholderia pseudomallei. Cell. Microbiol 7, 1447–1458 (2005). [DOI] [PubMed] [Google Scholar]
- 64.Suparak S et al. Multinucleated giant cell formation and apoptosis in infected host cells is mediated by Burkholderia pseudomallei type III secretion protein BipB. J. Bacteriol 187, 6556–6560 (2005). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 65.Ireland PM, Marshall L, Norville I & Sarkar-Tyson M The serine protease inhibitor Ecotin is required for full virulence of Burkholderia pseudomallei. Microb. Pathog 67–68, 55–58 (2014). [DOI] [PubMed] [Google Scholar]
- 66.Nathan SA & Puthucheary SD An electronmicroscopic study of the interaction of Burkholderia pseudomallei and human macrophages. Malays. J. Pathol 27, 3–7 (2005). [PubMed] [Google Scholar]
- 67.Miyagi K, Kawakami K & Saito A Role of reactive nitrogen and oxygen intermediates in gamma interferon-stimulated murine macrophage bactericidal activity against Burkholderia pseudomallei. Infect. Immun 65, 4108–4113 (1997). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 68.Ekchariyawat P et al. Burkholderia pseudomallei-induced expression of suppressor of cytokine signaling 3 and cytokine-inducible src homology 2-containing protein in mouse macrophages: a possible mechanism for suppression of the response to gamma interferon stimulation. Infect. Immun 73, 7332–7339 (2005). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 69.Wiersinga WJ et al. Immunosuppression associated with interleukin-1R-associated-kinase-M upregulation predicts mortality in Gram-negative sepsis (melioidosis). Crit. Care Med 37, 569–576 (2009). [DOI] [PubMed] [Google Scholar]
- 70.Vanaporn M et al. Superoxide dismutase C is required for intracellular survival and virulence of Burkholderia pseudomallei. Microbiology 157, 2392–2400 (2011). [DOI] [PubMed] [Google Scholar]
- 71.Loprasert S, Whangsuk W, Sallabhan R & Mongkolsuk S Regulation of the katG-dpsA operon and the importance of KatG in survival of Burkholderia pseudomallei exposed to oxidative stress. FEBS Lett. 542, 17–21 (2003). [DOI] [PubMed] [Google Scholar]
- 72.Loprasert S, Sallabhan R, Whangsuk W & Mongkolsuk S Compensatory increase in ahpC gene expression and its role in protecting Burkholderia pseudomallei against reactive nitrogen intermediates. Arch. Microbiol 180, 498–502 (2003). [DOI] [PubMed] [Google Scholar]
- 73.Loprasert S, Whangsuk W, Sallabhan R & Mongkolsuk S DpsA protects the human pathogen Burkholderia pseudomallei against organic hydroperoxide. Arch. Microbiol 182, 96–101 (2004). [DOI] [PubMed] [Google Scholar]
- 74.Myers ND et al. The role of NOD2 in murine and human melioidosis. J. Immunol 192, 300–307 (2014). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 75.Rinchai D et al. Macroautophagy is essential for killing of intracellular Burkholderia pseudomallei in human neutrophils. Autophagy 11, 748–755 (2015). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 76.Pudla M, Kananurak A, Limposuwan K, Sirisinha S & Utaisincharoen R Nucleotide-binding oligomerization domain-containing protein 2 regulates suppressor of cytokine signaling 3 expression in Burkholderia pseudomallei-infected mouse macrophage cell line RAW 264.7. Innate Immun. 17, 532–540 (2011). [DOI] [PubMed] [Google Scholar]
- 77.Gong L et al. The Burkholderia pseudomallei type III secretion system and BopA are required for evasion of LC3-associated phagocytosis. PLoS ONE 6, el 7852 (2011). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 78.Cullinane M et al. Stimulation of autophagy suppresses the intracellular survival of Burkholderia pseudomallei in mammalian cell lines. Autophagy 4, 744–753 (2008). [DOI] [PubMed] [Google Scholar]
- 79.Chanchamroen S, Kewcharoenwong C, Susaengrat W, Ato M & Lertmemongkolchai G Human polymorphonuclear neutrophil responses to Burkholderia pseudomallei in healthy and diabetic subjects. Infect. Immun 77, 456–463 (2009). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 80.Breitbach K et al. Actin-based motility of Burkholderia pseudomallei involves the Arp 2/3 complex, but not N-WASP and Ena/VASP proteins. Cell. Microbiol 5, 385–393 (2003). [DOI] [PubMed] [Google Scholar]
- 81.Stevens MR et al. Identification of a bacterial factor required for actin-based motility of Burkholderia pseudomallei. Mol. Microbiol 56, 40–53 (2005). [DOI] [PubMed] [Google Scholar]
- 82.St John JA et al. Burkholderia pseudomallei penetrates the brain via destruction of the olfactory and trigeminal nerves: implications for the pathogenesis of neurological melioidosis. mBio 5, e00025 (2014). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 83.Sarovich DS et al. Variable virulence factors in Burkholderia pseudomallei (melioidosis) associated with human disease. PLoS ONE 9, e91682 (2014). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 84.Harley VS, Dance DA, Drasar BS & Tovey G Effects of Burkholderia pseudomallei and other Burkholderia species on eukaryotic cells in tissue culture. Microbios 96, 71–93 (1998). [PubMed] [Google Scholar]
- 85.French CT et al. Dissection of the Burkholderia intracellular life cycle using a photothermal nanoblade. Proc. Natl Acad. Sci. USA 108, 12095–12100 (2011). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 86.Williams NL, Morris JL, Rush CM & Ketheesan N Migration of dendritic cells facilitates systemic dissemination of Burkholderia pseudomallei. Infect. Immun 82, 4233–4240 (2014). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 87.Newland RC Chronic melioidosis: a case in Sydney. Pathology 1, 149–152 (1969). [DOI] [PubMed] [Google Scholar]
- 88.Chodimella U, Hoppes WL, Whalen S, Ognibene AJ & Rutecki GW Septicemia and suppuration in a Vietnam veteran. Hosp. Pract 32, 219–221 (1997). [DOI] [PubMed] [Google Scholar]
- 89.Gee JE et al. Phylogeography of Burkholderia pseudomallei Isolates, Western Hemisphere. Emerg. Infect. Dis 23, 1133–1138 (2017). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 90.Gan YH Interaction between Burkholderia pseudomallei and the host immune response: sleeping with the enemy? J. Infect. Dis 192, 1845–1850 (2005). [DOI] [PubMed] [Google Scholar]
- 91.Vasu C, Vadivelu J & Puthucheary SD The humoral immune response in melioidosis patients during therapy. Infection 31, 24–30 (2003). [DOI] [PubMed] [Google Scholar]
- 92.Vadivelu J et al. Survival and intra-nuclear trafficking of Burkholderia pseudomallei: strategies of evasion from immune surveillance? PLoS Negl Trop. Dis 11, e0005241 (2017). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 93.Welkos SL. et al. Characterization of Burkholderia pseudomallei strains using a murine intraperitoneal infection model and in vitro macrophage assays. PLoS ONE 10, e0124667 (2015). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 94.Lewis ER & Torres AG The art of persistence-the secrets to Burkholderia chronic infections. Pathog. Dis 74, ftw070 (2016). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 95.Otsuka Y Prokaryotic toxin-antitoxin systems: novel regulations of the toxins. Curr. Genet 62, 379–382 (2016). [DOI] [PubMed] [Google Scholar]
- 96.Hamad MA et al. Adaptation and antibiotic tolerance of anaerobic Burkholderia pseudomallei. Antimicrob. Agents Chemother. 55, 3313–3323 (2011). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 97.Hayden HS et al. Evolution of Burkholderia pseudomallei in recurrent melioidosis. PLoS ONE 7, e36507 (2012). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 98.Price EP et al. Within-host evolution of Burkholderia pseudomallei over a twelve-year chronic carriage infection. mBio 4, e00388–13 (2013). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 99.Nuntayanuwat S, Dharakul T, Chaowagul W & Songsivilai S Polymorphism in the promoter region of tumor necrosis factor-alpha gene is associated with severe meliodosis. Hum. Immunol 60, 979–983 (1999). [DOI] [PubMed] [Google Scholar]
- 100.West TE et al. Toll-like receptor 4 region genetic variants are associated with susceptibility to melioidosis. Genes Immun. 13, 38–46(2012). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 101.Chantratita N et al. Screen of whole blood responses to fiagellin identifies TLR5 variation associated with outcome in melioidosis. Genes Immun. 15, 63–71 (2014). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 102.West TE et al. NLRC4 and TLR5 each contribute to host defense in respiratory melioidosis. PLoS Negl. Trop. Dis 8, e3178 (2014). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 103.Grube M et al. TLR5 stop codon polymorphism is associated with invasive aspergillosis after allogeneic stem cell transplantation. Med. Mycol 51, 818–825 (2013). [DOI] [PubMed] [Google Scholar]
- 104.Egan AM & Gordon DL Burkholderia pseudomallei activates complement and is ingested but not killed by polymorphonuclear leukocytes. Infect. Immun 64, 4952–4959 (1996). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 105.Ceballos-Olvera I, Sahoo M, Miller MA, Del Barrio L & Re F Inflammasome-dependent pyroptosis and IL-18 protect against Burkholderia pseudomallei lung infection while IL-1 beta is deleterious. PLoS Pathog. 7, e1002452 (2011). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 106.Easton A, Haque A, Chu K, Lukaszewski R & Bancroft GJ A critical role for neutrophils in resistance to experimental infection with Burkholderia pseudomallei. J. Infect. Dis 195, 99–107 (2007). [DOI] [PubMed] [Google Scholar]
- 107.Wiersinga WJ, Wieland CW, Roelofs JJ & van der Poll T MyD88 dependent signaling contributes to protective host defense against Burkholderia pseudomallei. PLoS ONE, 3 e3494 (2008). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 108.Wiersinga WJ et al. Toll-like receptor 2 impairs host defense in gram-negative sepsis caused by Burkholderia pseudomallei (Melioidosis). PLoS Med. 4, e248 (2007). [DOI] [PMC free article] [PubMed] [Google Scholar]; This is the first in-depth investigation of the expression and function of TLRs in human and murine melioidosis.
- 109.Hii CS et al. Interleukin-8 induction by Burkholderia pseudomallei can occur without Toll-like receptor signaling but requires a functional type III secretion system. J. Infect. Dis 197, 1537–1547 (2008). [DOI] [PubMed] [Google Scholar]
- 110.Wiersinga WJ et al. CD 14 impairs host defense against gram-negative sepsis caused by Burkholderia pseudomallei in mice. J. Infect. Dis 198, 1388–1397 (2008). [DOI] [PubMed] [Google Scholar]
- 111.Novem V et al. Structural and biological diversity of lipopolysaccharides from Burkholderia pseudomallei and Burkholderia thailandensis. Clin. Vaccine Immunol 16, 1420–1428 (2009). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 112.Korneev KV et al. Structural Relationship of the Lipid A Acyl Groups to Activation of Murine Toll-Like Receptor 4 by Lipopolysaccharides from Pathogenic Strains of Burkholderia mallei. Acinetobacter baumannii, and Pseudomonas aeruginosa. Front. Immunol 6, 595 (2015). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 113.Weehuizen TA et al. Differential Toll-like receptor-signalling of Burkholderia pseudomallei lipopolysaccharide in murine and human models. PLoS ONE 10, e0145397 (2015). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 114.Teh BE et al. Type three secretion system-mediated escape of Burkholderia pseudomallei into the host cytosol is critical for the activation of NFκB . BMC Microbiol. 14, 115 (2014). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 115.Bast A et al. Caspase-1 -dependent and -independent cell death pathways in Burkholders a pseudomallei infection of macrophages. PLoS Pathog. 10, e1003986 (2014). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 116.Wiersinga WJ et al. Endogenous interleukin-18 improves the early antimicrobial host response in severe melioidosis. Infect. Immun 75, 3739–3746 (2007). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 117.Wiersinga WJ et al. High-throughput mRNA profiling characterizes the expression of inflammatory molecules in sepsis caused by Burkholderia pseudomallei. Infect. Immun 75, 3074–3079 (2007). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 118.Lauw FN et al. Elevated plasma concentrations of interferon (IFN)-gamma and the IFN-gamma-inducing cytokines interleukin (IL)-18, IL-12, and IL-15 in severe melioidosis. J. Infect. Dis 180, 1878–1885 (1999). [DOI] [PubMed] [Google Scholar]
- 119.Barnes JL et al. Adaptive immunity in melioidosis: a possible role for T cells in determining outcome of infection with Burkholderia pseudomallei. Clin. Immunol 113, 22–28 (2004). [DOI] [PubMed] [Google Scholar]
- 120.Ulett GC, Ketheesan N & Hirst RG Macrophage-lymphocyte interactions mediate anti-Burkholderia pseudomallei activity. FEMS Immunol. Med. Microbiol 21, 283–286(1998). [DOI] [PubMed] [Google Scholar]
- 121.Jenjaroen K et al. T-Cell responses are associated with survival in acute melioidosis patients. PLoS Negl. Trop. Dis 9, e0004152 (2015). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 122.Aschenbroich SA, Lafontaine ER & Hogan RJ Melioidosis and glanders modulation of the innate immune system: barriers to current and future vaccine approaches. Expert Rev. Vaccines 15, 1163–1181 (2016). [DOI] [PubMed] [Google Scholar]
- 123.Wong KT, Puthucheary SD & Vadivelu J The histopathology of human melioidosis. Histopathology 26, 51–55 (1995). [DOI] [PubMed] [Google Scholar]
- 124.Taramasso L, Tatarelli P & Di Biagio A Bloodstream infections in HIV-infected patients. Virulence 7, 320–328 (2016). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 125.Chierakul W et al. Short report: disease severity and outcome of melioidosis in HIV coinfected individuals. Am. J. Trop. Med. Hyg 73, 1165–1166 (2005). [PubMed] [Google Scholar]
- 126.Gordon MA et al. Primary macrophages from HIV-infected adults show dysregulated cytokine responses to Salmonella, but normal internalization and killing. AIDS 21, 2399–2408 (2007). [DOI] [PubMed] [Google Scholar]
- 127.Haque A et al. Role of T cells in innate and adaptive immunity against murine Burkholderia pseudomallei infection. J. Infect. Dis 193, 370–379 (2006). [DOI] [PubMed] [Google Scholar]
- 128.Simpson AJ et al. Prognostic value of cytokine concentrations (tumor necrosis factor-alpha, interleukin-6, and interleukin-10) and clinical parameters in severe melioidosis. J. Infect. Dis 181, 621–625 (2000). [DOI] [PubMed] [Google Scholar]
- 129.Lauw FN et al. The CXC chemokines gamma interferon (IFN-gamma)-inducible protein 10 and monokine induced by IFN-gamma are released during severe melioidosis. Infect. Immun 68, 3888–3893 (2000). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 130.Barnes JL, Williams NL & Ketheesan N Susceptibility to Burkholderia pseudomallei is associated with host immune responses involving tumor necrosis factor receptor-1 (TNFR1) and TNF receptor-2 (TNFR2). FEMS Immunol. Med. Microbiol 52, 379–388 (2008). [DOI] [PubMed] [Google Scholar]
- 131.Ekchariyawat P et al. Expression of suppressor of cytokine signaling 3 (SOCS3) and cytokine-inducible Src homology 2-containing protein (CIS) induced in Burkholderia pseudomallei—infected mouse macrophages requires bacterial internalization. Microb. Pathog 42, 104–110 (2007). [DOI] [PubMed] [Google Scholar]
- 132.Massey S et al. Comparative Burkholderia pseudomallei natural history virulence studies using an aerosol murine model of infection. Sci. Rep 4, 4305 (2014). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 133.Wiersinga WJ et al. Expression and function of macrophage migration inhibitory factor (MIF) in melioidosis. PLoS Negl. Trop. Dis 4, e605 (2010). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 134.Charoensup J et al. High HMGB1 level is associated with poor outcome of septicemic melioidosis. Int. J. Infect. Dis 28, 111–116 (2014). [DOI] [PubMed] [Google Scholar]
- 135.Doker TJ et al. Fatal Burkholderia pseudomallei infection initially reported as a Bacillus species, Ohio, 2013. Am. J. Trop. Med. Hyg 91, 743–746 (2014). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 136.Currie BJ, Fisher DA, Anstey NM & Jacups SP Melioidosis: acute and chronic disease, relapse and re-activation. Trans. R. Soc. Trop. Med. Hyg 94, 301–304 (2000). [DOI] [PubMed] [Google Scholar]
- 137.Chierakul W et al. Melioidosis in 6 tsunami survivors in southern Thailand. Clin. Infect. Dis 41, 982–990 (2005). [DOI] [PubMed] [Google Scholar]
- 138.Hoffmaster AR et al. Melioidosis diagnostic workshop, 2013. Emerg. Infect. Dis 10.3201/eid2102.141045 (2015). [DOI] [PMC free article] [PubMed] [Google Scholar]; This is a CDC workshop paper involving the efforts of a large working group to update the diagnosis for melioidosis.
- 139.Centers for Disease Control and Prevention. Federal Select Agent Program https://www.selectagents.gov/ (2017).
- 140.Wuthiekanun V, Suputtamongkol Y, Simpson AJ, Kanaphun P & White NJ Value of throat swab in diagnosis of melioidosis. J. Clin. Microbiol 39, 3801–3802 (2001). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 141.Cheng AC et al. Role of selective and nonselective media for isolation of Burkholderia pseudomallei from throat swabs of patients with melioidosis. J. Clin. Microbiol 44, 2316 (2006). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 142.Limmathurotsakul D et al. Role and significance of quantitative urine cultures in diagnosis of melioidosis. J. Clin. Microbiol 43, 2274–2276 (2005). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 143.Dance DAB, Limmathurotsakul D & Currie BJ Burkholderia pseudomallei: challenges for the clinical microbiology laboratory — a response from the front line. J. Clin. Microbiol 55, 980–982 (2017). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 144.Podin Y et al. Burkholderia pseudomallei isolates from Sarawak, Malaysian Borneo, are predominantly susceptible to aminoglycosides and macrolides. Antimicrob. Agents Chemother 58, 162–166 (2014). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 145.Limmathurotsakul D et al. Defining the true sensitivity of culture for the diagnosis of melioidosis using Bayesian latent class models. PLoS ONE 5, e12485 (2010). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 146.Duval BD et al. Evaluation of a latex agglutination assay for the identification of Burkholderia pseudomallei and Burkholderia mallei. Am. J. Trop. Med. Hyg 90, 1043–1046 (2014). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 147.Suttisunhakul V et al. Matrix-assisted laser desorption/ionization time-of-flight mass spectrometry for the identification of Burkholderia pseudomallei from Asia and Australia and differentiation between Burkholderia species. PLoS ONE 12, e0175294 (2017). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 148.Gee JE et al. Use of 16S rRNA gene sequencing for rapid identification and differentiation of Burkholderia pseudomallei and B. mallei. J. Clin. Microbiol 41, 4647–4654 (2003). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 149.Koh SF et al. Development of a multiplex PCR assay for rapid identification of Burkholderia pseudomallei, Burkholderia thailandensis, Burkholderia mallei and Burkholderia cepacia complex. J. Microbiol. Methods 90, 305–308 (2012). [DOI] [PubMed] [Google Scholar]
- 150.Sheridan EA et al. Evaluation of the Wayson stain for the rapid diagnosis of melioidosis. J. Clin. Microbiol 45, 1669–1670 (2007). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 151.Tandhavanant S et al. Monoclonal antibody-based immunofluorescence microscopy for the rapid identification of Burkholderia pseudomallei in clinical specimens. Am. J. Trop. Med. Hyg 89, 165–168 (2013). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 152.Houghton RL et al. Development of a prototype lateral flow immunoassay (LFI) for the rapid diagnosis of melioidosis. PLoS Negl. Trop. Dis 8, e2727 (2014). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 153.Robertson G et al. Rapid diagnostics for melioidosis: a comparative study of a novel lateral flow antigen detection assay. J. Med. Microbiol 64, 845–848 (2015). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 154.Kaestli M et al. Comparison of TaqMan PCR assays for detection of the melioidosis agent Burkholderia pseudomallei in clinical specimens. J. Clin. Microbiol 50, 2059–2062 (2012). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 155.Richardson LJ et al. Towards a rapid molecular diagnostic for melioidosis: comparison of DNA extraction methods from clinical specimens. J. Microbiol. Methods 88, 179–181 (2012). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 156.Chaowagul W et al. Melioidosis: a major cause of community-acquired septicemia in northeastern Thailand. J. Infect. Dis 159, 890–899 (1989). [DOI] [PubMed] [Google Scholar]
- 157.Cheng AC et al. Prospective evaluation of a rapid immunochromogenic cassette test for the diagnosis of melioidosis in northeast Thailand. Trans. R. Soc. Trop. Med. Hyg 100, 64–67 (2006). [DOI] [PubMed] [Google Scholar]
- 158.Cheng AC, O’Brien M, Freeman K, Lum G & Currie BJ Indirect hemagglutination assay in patients with melioidosis in northern Australia. Am. J. Trop. Med. Hyg 74, 330–334 (2006). [PubMed] [Google Scholar]
- 159.Pumpuang A et al. Comparison of O-polysaccharide and hemolysin co-regulated protein as target antigens for serodiagnosis of melioidosis. PLoS Negl. Trop. Dis 11, e0005499 (2017). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 160.Suttisunhakul V et al. Development of rapid enzyme-linked immunosorbent assays for detection of antibodies to Burkholderia pseudomallei. J. Clin. Microbiol 54, 1259–1268 (2016). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 161.Suttisunhakul V et al. Evaluation of polysaccharide-based latex agglutination assays for the rapid detection of antibodies to Burkholderia pseudomallei. Am. J. Trop. Med. Hyg 93, 542–546 (2015). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 162.Kohler C et al. Rapid and sensitive multiplex detection of Burkholderia pseudomallei-specific antibodies in melioidosis patients based on a protein microarray approach. PLoS Negl. Trop. Dis 10, e0004847 (2016). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 163.Boyd R, McGuiness S, Draper AD, Neilson M & Krause V Melioidosis awareness campaign…. Don’t get melioidosis. Northern Territory Dis. Control Bull 23, 1–6 (2016). [Google Scholar]
- 164.Howard K & Inglis TJ The effect of free chlorine on Burkholderia pseudomallei in potable water. Water Res. 37, 4425–4432 (2003). [DOI] [PubMed] [Google Scholar]
- 165.McRobb E et al. Melioidosis from contaminated bore water and successful UV sterilization. Am. J. Trop. Med. Hyg 89, 367–368 (2013). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 166.Suntornsut P et al. Barriers and recommended interventions to prevent melioidosis in Northeast Thailand: a focus group study using the behaviour change wheel. PLoS Negl. Trop. Dis 10, e0004823 (2016). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 167.US National Library of Medicine. ClinicalTrials.gov, https://clinicaltrials.gov/ct2/show/NCT02089152 (2016). [DOI] [PubMed] [Google Scholar]
- 168.Lipsitz R et al. Workshop on treatment of and postexposure prophylaxis for Burkholderia pseudomallei and B. mallei Infection, 2010. Emerg. Infect. Dis 18, e2 (2012). [DOI] [PMC free article] [PubMed] [Google Scholar]; This is a CDC workshop paper involving the efforts of a large working group to update the treatment for melioidosis.
- 169.Crowe A, McMahon N, Currie BJ & Baird RW Current antimicrobial susceptibility of first-episode melioidosis Burkholderia pseudomallei isolates from the Northern Territory. Australia. Int. J. Antimicrob. Agents 44, 160–162 (2014). [DOI] [PubMed] [Google Scholar]
- 170.Dance DA et al. Trimethoprim/sulfamethoxazole resistance in Burkholderia pseudomallei. Int. J. Antimicrob. Agents 44, 368–369 (2014). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 171.Saiprom N et al. Trimethoprim/sulfamethoxazole resistance in clinical isolates of Burkholderia pseudomallei from Thailand. Int. J. Antimicrob. Agents 45, 557–559 (2015). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 172.Chaowagul W, Simpson AJ, Suputtamongkol Y & White NJ Empirical cephalosporin treatment of melioidosis. Clin. Infect. Dis 28, 1328 (1999). [DOI] [PubMed] [Google Scholar]
- 173.Dance D Treatment and prophylaxis of melioidosis. Int. J. Antimicrob. Agents 43, 310–318 (2014). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 174.Harris P, Engler C & Norton R Comparative in vitro susceptibility of Burkholderia pseudomallei to doripenem, ertapenem, tigecycline and moxifloxacin. Int. J. Antimicrob. Agents 37, 547–549 (2011). [DOI] [PubMed] [Google Scholar]
- 175.Ashdown LR Nosocomial infection due to Pseudomonas pseudomallei: two cases and an epidemiologic study. Rev. Infect. Dis 1, 891–894 (1979). [DOI] [PubMed] [Google Scholar]
- 176.Markovitz A Inoculation by bronchoscopy. West. J. Med 131, 550 (1979). [PMC free article] [PubMed] [Google Scholar]
- 177.Kelen GD, Hansen KN, Green GB, Tang N & Ganguli C Determinants of emergency department procedure- and condition-specific universal (barrier) precaution requirements for optimal provider protection. Ann. Emerg. Med 25, 743–750 (1995). [DOI] [PubMed] [Google Scholar]
- 178.Pittet D, Allegranzi B, Boyce J & World Health Organization World Alliance for Patient Safety First Global Patient Safety Challenge Core Group of Experts. The World Health Organization guidelines on hand hygiene in health care and their consensus recommendations. Infect. Control Hosp. Epidemiol 30, 611–622 (2009). [DOI] [PubMed] [Google Scholar]
- 179.White NJ et al. Halving of mortality of severe melioidosis by ceftazidime. Lancet 2, 697–701 (1989). [DOI] [PubMed] [Google Scholar]
- 180.Simpson AJ et al. Comparison of imipenem and ceftazidime as therapy for severe melioidosis. Clin. Infect. Dis 29, 381–387 (1999). [DOI] [PubMed] [Google Scholar]
- 181.Chierakul W et al. Two randomized controlled trials of ceftazidime alone versus ceftazidime in combination with trimethoprim-sulfamethoxazole for the treatment of severe melioidosis. Clin. Infect. Dis 41, 1105–1113 (2005). [DOI] [PubMed] [Google Scholar]
- 182.Chierakul W et al. Addition of trimethoprim-sulfamethoxazole to ceftazidime during parenteral treatment of melioidosis is not associated with a long-term outcome benefit. Clin. Infect. Dis 45, 521–523 (2007). [DOI] [PubMed] [Google Scholar]
- 183.Smith MD, Wuthiekanun V, Walsh AL & White NJ Susceptibility of Pseudomonas pseudomallei to some newer beta-lactam antibiotics and antibiotic combinations using time-kill studies. J. Antimicrob. Chemother 33, 145–149 (1994). [DOI] [PubMed] [Google Scholar]
- 184.Smith MD, Wuthiekanun V, Walsh AL & White NJ In vitro activity of carbapenem antibiotics against beta-lactam susceptible and resistant strains of Burkholderia pseudomallei. J. Antimicrob. Chemother 37, 611–615 (1996). [DOI] [PubMed] [Google Scholar]
- 185.Cheng AC et al. Outcomes of patients with melioidosis treated with meropenem. Antimicrob. Agents Chemother 48, 1763–1765 (2004). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 186.Cheng AC et al. Dosing regimens of cotrimoxazole (trimethoprim–sulfamethoxazole) for melioidosis. Antimicrob. Agents Chemother 53, 4193–4199 (2009). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 187.Cheng AC et al. Consensus guidelines for dosing of amoxicillin-clavulanate in melioidosis. Am. J. Trop. Med. Hyg 78, 208–209 (2008). [PMC free article] [PubMed] [Google Scholar]
- 188.Pitman MC et al. Intravenous therapy duration and outcomes in melioidosis: a new treatment paradigm. PLoS Negl. Trop. Dis 9, e0003586 (2015). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 189.Chaowagul W et al. Relapse in melioidosis: incidence and risk factors. J. Infect. Dis 168, 1181–1185 (1993). [PubMed] [Google Scholar]
- 190.Limmathurotsakul D et al. Risk factors for recurrent melioidosis in northeast Thailand. Clin. Infect. Dis 43, 979–986 (2006). [DOI] [PubMed] [Google Scholar]
- 191.Limmathurotsakul D et al. A simple scoring system to differentiate between relapse and re-infection in patients with recurrent melioidosis. PLoS Negl. Trop. Dis 2, e327 (2008). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 192.Lumbiganon P, Chotechuangnirun N, Kosalaraksa P & Teeratakulpisarn J Localized melioidosis in children in Thailand: treatment and long-term outcome. J. Trop. Pediatr 57, 185–191 (2011). [DOI] [PubMed] [Google Scholar]
- 193.Pagnarith Y et al. Emergence of pediatric melioidosis in Siem Reap, Cambodia. Am. J. Trop. Med. Hyg 82, 1106–1112 (2010). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 194.Morse LP et al. Osteomyelitis and septic arthritis from infection with Burkholderia pseudomallei: a 20-year prospective melioidosis study from northern Australia. J. Orthop 10, 86–91 (2013). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 195.Shetty RP et al. Management of melioidosis osteomyelitis and septic arthritis. Bone Joint J. 97-B, 277–282 (2015). [DOI] [PubMed] [Google Scholar]
- 196.Cheng AC, West TE, Limmathurotsakul D & Peacock SJ Strategies to reduce mortality from bacterial sepsis in adults in developing countries. PLoS Med. 5, e175 (2008). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 197.Stephens DP, Thomas JH, Ward LM & Currie BJ Melioidosis causing critical illness: a review of 24 yearn of experience from the Royal Darwin Hospital ICU. Crit. Care Med 44, 1500–1505 (2016). [DOI] [PubMed] [Google Scholar]
- 198.Rhodes A et al. Surviving Sepsis Campaign: International Guidelines for Management of Sepsis and Septic Shock: 2016. Intensive Care Med. 43, 304–377 (2017). [DOI] [PubMed] [Google Scholar]
- 199.Cheng AC, Stephens DP, Anstey NM & Currie BJ Adjunctive granulocyte colony-stimulating factor for treatment of septic shock due to melioidosis. Clin. Infect. Dis 38, 32–37 (2004). [DOI] [PubMed] [Google Scholar]
- 200.Cheng AC et al. A randomized controlled trial of granulocyte colony-stimulating factor for the treatment of severe sepsis due to melioidosis in Thailand. Clin. Infect. Dis 45, 308–314 (2007). [DOI] [PubMed] [Google Scholar]
- 201.Weehuizen TA et al. Therapeutic administration of a monoclonal anti-II-1 beta antibody protects against experimental melioidosis. Shock 46, 566–574 (2016). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 202.Koh GC et al. Glyburide is anti-inflammatory and associated with reduced mortality in melioidosis. Clin. Infect. Dis 52, 717–725 (2011). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 203.Sarovich DS et al. Recurrent melioidosis in the Darwin Prospective Melioidosis Study: improving therapies mean that relapse cases are now rare. J. Clin. Microbiol 52, 650–653 (2014). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 204.Teparrakkul P et al. Rheumatological manifestations in patients with melioidosis. Southeast Asian J. Trop. Med. Publ. Health 39, 649–655 (2008). [PubMed] [Google Scholar]
- 205.Molyneux DH, Savioli L & Engels D Neglected tropical diseases: progress towards addressing the chronic pandemic. Lancet 389, 312–325 (2017). [DOI] [PubMed] [Google Scholar]
- 206.Teerawattanasook N et al. Capacity and utilisation of blood culture in two referral hospitals in Indonesia and Thailand. Am. J. Trop. Med. Hyg 97, 1257–1261 (2017). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 207.Skvarc M, Stubljar D, Rogina P & Kaasch AJ Non-culture-based methods to diagnose bloodstream infection: Does it work? Eur. J. Microbiol. Immunol 3, 97–104 (2013). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 208.Paek SC, Meemon N &Wan TT Thailand’s universal coverage scheme and its impact on health-seeking behavior. Springerplus 5, 1952 (2016). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 209.van Dijk DP, Dinant G & Jacobs JA Inappropriate drug donations: what has happened since the 1999 WHO guidelines? Educ. Health 24, 462 (2011). [PubMed] [Google Scholar]
- 210.Schweizer HP, Tuanyok A & Bertherat E Eighth World Melioidosis Congress, 2016: presenting an emerging infectious disease in the context of “One Health”. Wkly Epidemiol. Rec 91, 543–547 (2016). [PubMed] [Google Scholar]
- 211.Gibbs P Origins of One Health and One Medicine. Vet. Rec 174, 152 (2014). [DOI] [PubMed] [Google Scholar]
- 212.Peacock SJ et al. Melioidosis vaccines: a systematic review and appraisal of the potential to exploit biodefense vaccines for public health purposes. PLoS Negl. Trop. Dis 6, e 1488 (2012). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 213.Limmathurotsakul D et al. Consensus on the development of vaccines against naturally acquired melioidosis. Ernerg. Infect. Dis 10.3201/eid2106.141480 (2015). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 214.Sarkar-Tyson M & Titball RW Progress toward development of vaccines against melioidosis: A review. Clin. Ther 32, 1437–1445 (2010). [DOI] [PubMed] [Google Scholar]
- 215.Patel N et al. Development of vaccines against Burkholderia pseudomallei. Front. Microbiol 2, 198 (2011). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 216.Torres AG et al. Protection of non-human primates against glanders with a gold nanoparticle glycoconjugate vaccine. Vaccine 33, 686–692 (2015). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 217.Muruato LA & Torres AG Melioidosis: where do we stand in the development of an effective vaccine? Future Microbiol. 11, 477–480 (2016). [DOI] [PubMed] [Google Scholar]
- 218.Lankelma JM et al. The gut microbiota as a modulator of innate immunity during melioidosis. PLoS Negl. Trop. Dis 11, e0005548 (2017). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 219.van Crevel R, van de Vijver S & Moore DAJ. The global diabetes epidemic: what does it mean for infectious diseases in tropical countries? Lancet Diabetes Endocrinol. 5, 457–468 (2017). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 220.Cheng AC, Jacups SP, Gal D, Mayo M & Currie BJ Extreme weather events and environmental contamination are associated with case-clusters of melioidosis in the Northern Territory of Australia. Int. J. Epidemiol 35, 323–329 (2006). [DOI] [PubMed] [Google Scholar]
- 221.Liu CL, Huang JJ, Lin HC, Huang ST. & Liu DP Investigation and analysis of melioidosis outbreak after Typhoon Nanmadol in Southern Taiwan, 2011. Int. J. Infect. Dis 16, e351 (2012). [Google Scholar]
- 222.Wang J et al. Multiple mechanisms involved in diabetes protection by lipopolysaccharide in non-obese diabetic mice. Toxicol. Appl. Pharmacol 285, 149–158 (2015). [DOI] [PubMed] [Google Scholar]
- 223.Maniam P, Nurul Aiezzah Z, Mohamed R, Embi N & Hasidah MS Regulatory role of GSK3beta in the activation of NF-kappaB and modulation of cytokine levels in Burkholderia pseudomallei-infected PBMC isolated from streptozotocin-induced diabetic animals. Trop. Biorned 32, 36–48 (2015). [PubMed] [Google Scholar]
- 224.Buddhisa S, Rinchai D, Ato M, Bancroft GJ & Lertmemongkolchai G Programmed death ligand 1 on Burkholderia pseudomallei-infected human polymorphonuclear neutrophils impairs T cell functions. J. Immunol 194, 4413–4421 (2015). [DOI] [PubMed] [Google Scholar]
- 225.Koh GC, Peacock SJ, van der Poll T & Wiersinga WJ The impact of diabetes on the pathogenesis of sepsis. Eur J. Clin. Microbiol. Infect. Dis 31, 379–388 (2012). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 226.Liu X et al. Sulphonylurea usage in melioidosis is associated with severe disease and suppressed immune response. PLoS Negl. Trop. Dis 8, e2795 (2014). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 227.Kewcharoenwong C et al. Glibenclamide impairs responses of neutrophils against Burkholderia pseudomallei by reduction of intracellular glutathione. Sci. Rep 6, 34794 (2016). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 228.Galyov EE, Brett PJ & DeShazer D Molecular insights into Burkholderia pseudomallei and Burkholderia mallei pathogenesis. Annu. Rev. Microbiol 64, 495–517 (2010). [DOI] [PubMed] [Google Scholar]; This is a review article that presents a comprehensive history of the mechanisms of pathogenesis associated with both B. pseudomallei and B. mallei (up to 2010).
- 229.Holden MT et al. Genomic plasticity of the causative agent of melioidosis, Burkholderia pseudomallei. Proc. Natl Acad. Sci. USA 101, 14240–14245 (2004). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 230.Price EP et al. Within-host evolution of Burkholderia pseudomallei in four cases of acute melioidosis. PLoS Pathog. 6, e1000725 (2010). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 231.Tumapa S et al. Burkholderia pseudomallei genome plasticity associated with genomic island variation. BMC Genom. 9, 190 (2008). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 232.Chewapreecha C et al. Global and regional dissemination and evolution of Burkholderia pseudomallei. Nat. Microbiol 2, 16263 (2017). [DOI] [PMC free article] [PubMed] [Google Scholar]; This is a study in which whole-genome sequencing of 469 B. pseudomallei isolates from 30 countries maps the regional dissemination and evolution of B. pseudomallei across the globe.
- 233.Rhodes KA & Schweizer HP Antibiotic resistance in Burkholderia species. Drug Resist. Updat 28, 82–90 (2016). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 234.Bugrysheva JV et al. Antibiotic resistance markers in Burkholderia pseudomallei strain Bp 1651 identified by genome sequence analysis. Antimicrob. Agents Chemother 61, e00010–17 (2017). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 235.Chantratita N et al. Antimicrobial resistance to ceftazidime involving loss of penicillin-binding protein 3 in Burkholderia pseudomallei. Proc. Natl Acad. Sci. USA 108, 17165–17170 (2011). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 236.Randall LB, Dobos K, Papp-Wallace KM, Bonomo RA & Schweizer HP Membrane-bound PenA beta-lactamase of Burkholderia pseudomallei. Antimicrob. Agents Chemother 60, 1509–1514 (2015). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 237.Price EP et al. Whole-genome sequences of Burkholderia pseudomallei isolates exhibiting decreased meropenem susceptibility. Genome Announc. 5, e00053–17 (2017). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 238.Podnecky NL, Wuthiekanun V, Peacock SJ & Schweizer HP The BpeEF-OprC efflux pump is responsible for widespread trimethoprim resistance in clinical and environmental Burkholderia pseudomallei isolates. Antimicrob. Agents Chemother 57, 4381–4386(2013). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 239.Sirijant N, Sermswan RW & Wongratanacheewin S Burkholderia pseudomallei resistance to antibiotics in biofilm-induced conditions is related to efflux pumps. J. Med. Microbiol 65, 1296–1306 (2016). [DOI] [PubMed] [Google Scholar]
- 240.Kager LM, van der Poll T & Wiersinga WJ The coagulation system in melioidosis: from pathogenesis to new treatment strategies. Expert Rev. Anti-Infective Ther 12, 993–1002 (2014). [DOI] [PubMed] [Google Scholar]
- 241.LaRosa SP et al. Decreased protein C, protein S, and antithrombin levels are predictive of poor outcome in Gram-negative sepsis caused by Burkholderia pseudomallei. Int. J. Infect. Dis 10, 25–31 (2006). [DOI] [PubMed] [Google Scholar]
- 242.Wiersinga WJ et al. Activation of coagulation with concurrent impairment of anticoagulant mechanisms correlates with a poor outcome in severe melioidosis. J. Thromb. Haemost 6, 32–39 (2008). [DOI] [PubMed] [Google Scholar]
- 243.Kager LM et al. Overexpression of activated protein C is detrimental during severe experimental gram-negative sepsis (melioidosis). Crit. Care Med 41, e266–274 (2013). [DOI] [PubMed] [Google Scholar]
- 244.Kager LM et al. Plasminogen activator inhibitor type I contributes to protective immunity during experimental Gram-negative sepsis (melioidosis). J. Thromb. Haemost 9, 2020–2028 (2011). [DOI] [PubMed] [Google Scholar]
- 245.Kager LM et al. Endogenous alpha2-antiplasmin is protective during severe gram-negative sepsis (melioidosis). Am. J. Respir. Crit. Care Med 188, 967–975 (2013). [DOI] [PubMed] [Google Scholar]
- 246.Currie BJ Melioidosis: The 2014 Revised RDH Guideline. Northern Territory Dis. Control Bull 21, 4–8(2014). [Google Scholar]
- 247.Rachlin A et al. Investigation of recurrent melioidosis in Lao People’s Democratic Republic by multilocus sequence typing. Am. J. Trop. Med. Hyg 94, 1208–1211 (2016). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 248.Tauran PM et al. Emergence of melioidosis in Indonesia. Am. J. Trop. Med. Hyg 93, 1160–1163 (2015). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 249.Rothe C et al. Clinical Cases in Tropical Medicine (Saunders Ltd., 2014). [Google Scholar]
- 250.Lafontaine ER, Balder R, Michel F & Hogan RJ Characterization of an autotransporter adhesin protein shared by Burkholderia mallei and Burkholderia pseudomallei. BMC Microbiol. 14, 92 (2014). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 251.DeShazer D, Brett PJ, Carlyon R & Woods DE Mutagenesis of Burkholderia pseudomallei with Tn5-OT182: isolation of motility mutants and molecular characterization of the flagellin structural gene. J. Bacteriol 179, 2116–2125 (1997). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 252.Chua KL, Chan YY & Gan YH Flagella are virulence determinants of Burkholderia pseudomallei. Infect. Immun 71, 1622–1629 (2003). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 253.Muangsombut V et al. Inactivation of Burkholderia pseudomallei bsaQ results in decreased invasion efficiency and delayed escape of bacteria from endocytic vesicles. Arch. Microbiol 190, 623–631 (2008). [DOI] [PubMed] [Google Scholar]
- 254.Burtnick MN et al. Burkholderia pseudomallei type III secretion system mutants exhibit delayed vacuolar escape phenotypes in RAW 264.7 murine macrophages. Infect. Immun 76, 2991–3000 (2008). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 255.Pilatz S et al. Identification of Burkholderia pseudomallei genes required for the intracellular life cycle and in vivo virulence. Infect. Immun 74, 3576–3586 (2006). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 256.Warawa J & Woods DH Type III secretion system cluster 3 is required for maximal virulence of Burkholderia pseudomallei in a hamster infection model. FEMS Microbiol. Lett 242, 101–10 8 (2005). [DOI] [PubMed] [Google Scholar]
- 257.Vanaporn M, Vattanaviboon P, Thongboonkerd V & Korbsrisate S The rpoE operon regulates heat stress response in Burkholderia pseudomallei. FEMS Microbiol. Lett 284, 191–196 (2008). [DOI] [PubMed] [Google Scholar]
- 258.Utaisincharoen P, Arjcharoen S, Umposuwan K, Tungpradabkul S & Sirisinha S Burkholderia pseudomallei RpoS regulates multinucleated giant cell formation and inducible nitric oxide synthase expression in mouse macrophage cell line (RAW 264.7). Microb. Pathog 40, 184–189 (2006). [DOI] [PubMed] [Google Scholar]
- 259.Leiman PG et al. Type VI secretion apparatus and phage tail-associated protein complexes share a common evolutionary origin. Proc. Natl Acad. Sci. USA 106, 4154–4159 (2009). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 260.Easier M, Pilhofer M, Henderson GP, Jensen GJ & Mekalanos JJ Type VI secretion requires a dynamic contractile phage tail-like structure. Nature 483, 182–186 (2012). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 261.Burtnick MN et al. The cluster 1 type VI secretion system is a major virulence determinant in Burkholderia pseudomallei. Infect. Immun 79, 1512–1525 (2011). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 262.Chieng S, Mohamed R & Nathan S Transcriptome analysis of Burkholderia pseudomallei T6SS identifies Hcp1 as a potential serodiagnostic marker. Microb. Pathog 79, 47–56 (2015). [DOI] [PubMed] [Google Scholar]
- 263.Toesca IJ, French CT & Miller JF The Type VI secretion system spike protein VgrG5 mediates membrane fusion during intercellular spread by pseudomallei group Burkholderia species. Infect. Immun 82, 1436–1444 (2014). [DOI] [PMC free article] [PubMed] [Google Scholar]; This is an important basic science research paper from the past decade of B. pseudomallei research.
- 264.Tan KS et al. Suppression of host innate immune response by Burkholderia pseudomallei through the virulence factor TssM. J. Immunol 184, 5160–5171 (2010). [DOI] [PubMed] [Google Scholar]
- 265.Reckseidler-Zenteno SL et al. Characterization of the type III capsular polysaccharide produced by Burkholderia pseudomallei. J. Med. Microbiol 59, 1403–1414 (2010). [DOI] [PubMed] [Google Scholar]
- 266.Woodman MH, Worth RG & Wooten RM Capsule influences the deposition of critical complement C3 levels required for the killing of Burkholderia pseudomallei via NADPH-oxidase induction by human neutrophils. PLoS ONE 7, e52276 (2012). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 267.Mongkolrob R, Taweechaisupapong S & Tungpradabkul S Correlation between biofilm production, antibiotic susceptibility and exopolysaccharide composition in Burkholderia pseudomallei bpsl, ppk, and rpoS mutant strains. Microbiol. Immunol 59, 653–663 (2015). [DOI] [PubMed] [Google Scholar]
- 268.Norris MH, Schweizer HP & Tuanyok A Structural diversity of Burkholderia pseudomallei lipopolysaccharides affects innate immune signaling. PLOS Negl Trop. Dis 11, e0005571 (2017). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 269.Ulrich RL et al. Role of quorum sensing in the pathogenicity of Burkholderia pseudomallei. J. Med. Microbiol 53, 1053–1064 (2004). [DOI] [PubMed] [Google Scholar]
- 270.Chan YY & Chua KL The Burkholderia pseudomallei BpeAB-OprB efflux pump: expression and impact on quorum sensing and virulence. J. Bacteriol 187, 4707–4719 (2005). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 271.Cruz A et al. A Burkholderia pseudomallei toxin inhibits helicase activity of translation factor eIF4A. Science 334, 821–824 (2011). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 272.Walsh MJ, Dodd JH & Hautbergue GM Ribosome-inactivating proteins: potent poisons and molecular tools. Virulence 4, 774–784 (2013). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 273.Chantratita N et al. Biological relevance of colony morphology and phenotypic switching by Burkholderia pseudomallei. J. Bacteriol 189, 807–817 (2007). [DOI] [PMC free article] [PubMed] [Google Scholar]