

Abstract
Aim of the review
In the past decade, increasing research attention investigated the novel therapeutic potential of steroidal cardiac glycosides in cancer treatment. Huachansu and its main active constituent Bufalin have been studied in vitro, in vivo and clinical studies. This review aims to summarize the multi-target and multi-pathway pharmacological effects of Bufalin and Huachansu in the last decade, with the aim of providing a more comprehensive view and highlighting the recently discovered molecular mechanisms.
Results
Huachansu and its major derivative, Bufalin, had been found to possess anti-cancer effects in a variety of cancer cell lines both in vitro and in vivo. The underlying anti-cancer molecular mechanisms mainly involved anti-proliferation, apoptosis induction, anti-metastasis, anti-angiogenesis, epithelial–mesenchymal transition inhibition, anti-inflammation, Na+/K+-ATPase activity targeting, the steroid receptor coactivator family inhibitions, etc. Moreover, the potential side-effects and toxicities of the toad extract, Huachansu, and Bufalin, including hematological, gastrointestinal, mucocutaneous and cardiovascular adverse reactions, were reported in animal studies and clinic trails.
Conclusions
Further research is needed to elucidate the potential drug–drug interactions and multi-target interaction of Bufalin and Huachansu. Large-scale clinical trials are warranted to translate the knowledge of the anticancer actions of Bufalin and Huachansu into clinical applications as effective and safe treatment options for cancer patients in the future.
Keywords: Bufalin, Huachansu, Anti-cancer, Traditional Chinese Medicine
Introduction
Traditional Chinese Medicine (TCM) is a medical practice with more than 2500 years of history in China. It has recently been recognized as a new type of chemotherapy adjuvant that can improve the efficacy of chemotherapy and ameliorate the side effects of cancer chemotherapies. Ever since the launch of chemotherapy for tumor treatment, natural products have become an important source of the development of novel cancer therapies. Although there is no pathological concept of cancer under TCM theory, it is easy to find the descriptions of cancer-like symptoms in the ancient medical documents described as lumps, bumps, and toxins, termed “Chuang, Yong, Zhong and Du (疮痈肿毒)” in Chinese [1]. Therefore, identifying drugs used to treat lumps and bumps, as well as other cancer-related symptoms, such as relief from fever, diarrhea, vomiting, and pain, may link the traditional use of the agent to the modern pathological concept of cancer and may strengthen the pharmacological relevance of TCM to contemporary anti-cancer treatments.
Chansu (CS, Senso in Japanese) is the dried secretion from the skin glands of Bufo bufo gargarizans Cantor or Bufo melanostictus Schneider [2]. According to the principles of TCM theory, CS is commonly used to counteract toxicity, alleviate pain, and induce resuscitation [3, 4]. It can be considered as an anti-infectious agent for pyogenic infection induced unconsciousness and may be related to its anti-inflammatory and anti-microbial effects [3, 5]. In TCM practice, CS is prescribed to patients with “heat and toxins” syndrome, which refers to the modern concepts including acute gastroenteritis, severe vomiting, diarrhea, abdominal pain, high fever, carbuncles, lumps, and bumps [6]. Huachansu (HCS) is an injectable form of the sterilized hot-water extract of CS [7]. It is manufactured by Anhui Jinchan Biochemistry Company Ltd., in Huaibei, China [Chinese Food and Drug Administration, FDA (ISO9002)] and is widely used for inflammatory diseases as well as for the treatment for various types of cancer, including liver, lung, pancreatic, and colorectal cancers in China [8–12].
The molecular basis for the anti-inflammatory effect of HCS is proposed to be the bioactive steroidal cardiac glycosides [13]. Indeed, glycosides isolated from HCS have been shown to possess blood pressure stimulation, respiratory excitation, anti-inflammatory, anesthetic, and anti-neoplastic activities [14]. HCS and its derived single compounds may achieve their anti-inflammatory effects by modulating nuclear factor-κB (NF-κB) signaling and down-regulating inflammatory-related genes such as cyclooxygenases, lipoxygenases, inducible nitric oxide synthase, and thereby decrease nitric oxide and prostaglandin E2 (PGE2) production [9, 11–13]. In cancerous cells, glycosides derived from HCS also exhibit cytostatic and cytotoxic activities, induce cellular apoptosis, inhibits angiogenesis, reverses chemotherapeutic drug resistance, and modulate immune responses. Previous studies suggest that Na+/K+ pump or sodium- and potassium-activated adenosine 5′-triphosphatase (Na+, K+-ATPase) is a potential drug target that contributes to the selective control role of cardiac glycosides in tumor proliferation, but does not affect normal cell growth [10, 15, 16]. Moreover, accumulating evidence reveals the anti-cancer effect of HCS and its derived single compounds in several tumor types in vitro and in vivo.
Furthermore, in the last decade, some studies have proposed new properties and effects of HCS, Chansu and their major active constitutes, bufalin, in the treatment of cancer (Fig. 1). Interestingly, there are an increasing number of studies investigating both in vitro and in vivo experiments in the recent 5 years, indicating an increased awareness of the translational potential of HCS and its derived steroidal cardiac glycosides in animal studies. In this review article, data on the anti-cancer effect of HCS and its major active constitutes bufalin published in the recent 10-years were retrieved from databases including PubMed, MEDLINE, CNKI, and clinicaltrial.gov. This review focuses on the anti-cancer pharmacological effects and mechanisms of action of HCS and bufalin, with emphasis on elaborating the translational potential and future clinical application. This review article also discusses the recent studies on drug delivery and its derivatives.
Fig. 1.

a Increase of publication in number in the recent 10 years; Data was retrieved from PubMed in 5-year interval with keywords of (“Neoplasms”[MeSH] AND “bufalin” OR “huachansu” OR “chansu” OR “chan-su”). b Steady annual citation of our phase II clinical trial since its publication in recent 5 years
The bioactive constituents of Huachansu
The chemical composition and pharmacological activity of HSC have been investigated since the 1980s [7, 14, 15, 17, 18]. HSC contains two primary bioactive chemical components, indole alkaloids (bufotenine, bufotenidine, cinobufotenine, and serotonin), and steroidal cardiac glycosides [7, 14, 15, 18]. Their extraction rate is mainly determined by the extraction method. High performance liquid chromatography (HPLC) quantitative analysis confirmed that the aqueous extract of HSC yield around 20-fold higher serotonin than bufadienolides (75.7 ± 0.1 mg/g and 3.8 ± 0.0 mg/g, respectively), while methanol or ethanol extraction solution contains 5–26 times higher concentrations of bufadienolides, with only trace amounts of serotonin [14]. So far, there are more than 28 steroidal cardiac glycosides identified from HCS [19]. The investigation into the potential use of cardiac glycosides in cancer therapeutic was initiated more than 40 years ago, yet was abandoned due to the toxicities [20]. However, in 1999, Scandinavian oncologist Haux [21] reported that digoxin induced tumor cell apoptosis in a variety of human cancer cell lines at non-toxic concentration. Recent studies demonstrated that steroidal cardiac glycosides are the major anti-neoplasm component of HCS.
Bufalin (PubChem CID: 9547215, Chemical formula C24H34O4, Fig. 2) is a cardiac glycoside and the major active component is attributed to the anti-tumor activity of HCS [12]. Similar to other bufadienolides (such as resibufogenin, cinobufagin, and bufotalin), Bufalin is a cardioactive C-24 steroid characterized by an α-pyrone ring at C-17 [22, 23]. Bufalin exhibits a variety of biological activities. Its structural similarity with digitoxin accounts for both the therapeutic effect as well as the unwanted side-effects such as cardiotonic, blood pressure stimulatory and respiratory stimulatory effects in cancer treatment [20, 24, 25]. Recent studies emerge on purified compounds and cardiac glycosides. Bufalin may represent a promising form of targeted cancer chemotherapy for long-term applications without severe side effects.
Fig. 2.
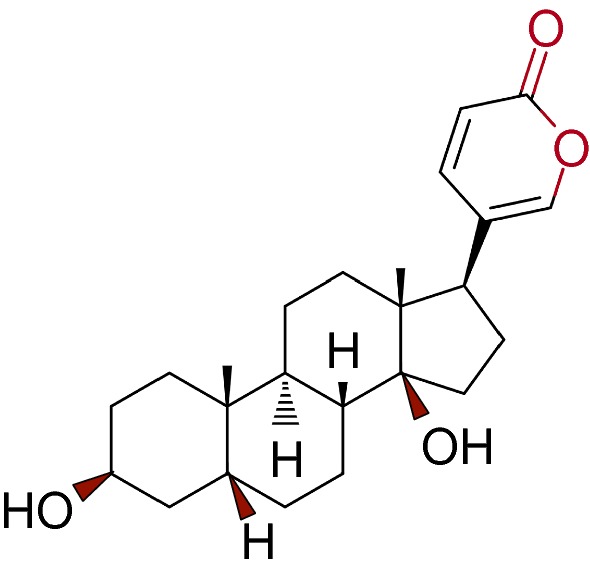
Shows the structure of Bufalin (C24H34O4, Pubchem CID: 9547215). Retrieved from National Center for Biotechnology Information. PubChem Compound Database; CID = 9547215, from https://pubchem.ncbi.nlm.nih.gov/compound/9547215 (accessed 22 Aug 2018)
Cytostatic effects of Huachansu and Bufalin on tumors
It appears that the cytostatic effect of Bufalin and HCS is well demonstrated in a variety of tumors such as breast, colorectal, gastric, lung, liver and bone tumors. The 50% inhibition dose (IC50) among cancer cell lines after 1–3 days of treatment are summarized in Table 1. The in vitro effects of Bufalin and HCS in arresting cell cycle, inducing intrinsic and extrinsic apoptosis are reproducible across studies. Recent studies have confirmed that Bufalin can lead to cell cycle redistribution, at least in part by its role in the inhibition of Na+/K+-ATPase activity, in various types of human cancers [26, 27]. In cancer cells, Bufalin can arrest cell cycle at G0/G1 and G2/M phase, owing to the various dosage of treatment and cell lines [28–30]. The sensitivity to Bufalin treatment varies among cell lines. Currently, there is a lack of in-depth study to predict the exact effect of Bufalin on cell cycle. Moreover, it is worth noting that one article published in 2013 by Clifford and Kaplan suggested that Bufalin and other cardiotonic steroids, including Ouabain and Digitoxin, can inhibit membrane Na+/K+-ATPase activity in multiple cell lines regardless of metastatic potential [31, 32]. Also, human breast tumor cells are more resistant to growth inhibition and apoptosis induction of cardiotonic steroids than non-tumorigenic breast cells [31]. The 50% inhibitory dose with 24-h of Bufalin treatment in metastatic invasive ductal carcinoma MCF10CA1 cell and the invasive estrogen receptor (ER)-negative MDA-MD-231 breast cancer cell is two- and threefold, respectively, the dosage of human noncancerous mammary epithelial cells 184D and 184A1 [31]. Another study on the effect of HCS on normal human lymphocytes was marginal (with more than 60% viability at 48-h) compared with A-549, Jurkat, MCF-7 cancer cells (with less than 10% viability) at a dose of 0.16 mg/mL of CS extract [33]. However, a majority of studies investigating the cytostatic effect of HCS and Bufalin are carried out solely on tumorigenic cells instead of normal non-cancerous cells. Further investigation is warranted to elucidate the pharmacological action of Bufalin treatment.
Table 1.
Growth inhibition of Bufalin (BF) on human cancer cell lines after 24–72H treatment
| Cancer type | Cell line | 24H IC50 | 48H IC50 | 72H IC50 | Refs. |
|---|---|---|---|---|---|
| Breast | MCF-7 | 317.9 ± 1.5 nM | 46.5 ± 1.4 nM | Yan et al. [125] | |
| 3.2 nM | Wang et al. [56] | ||||
| MCF-7/ADR | 100.17 ± 30.16 nmol/L | Wang et al. [126] | |||
| MCF-10A | 465.2 ± 25.9 nM | Clifford et al. [31] | |||
| MCF10CA1 | 635.2 ± 171.8 nM | Clifford et al. [31] | |||
| MDA-MB-231 | 934.1 ± 2.0 nM | 513.3 ± 1.6 nM | Yan et al. [125] | ||
| 263.3 ± 68.24 nmol/L | Wang et al. [126] | ||||
| 936.4 ± 4.9 nM | Clifford et al. [31] | ||||
| 20.0 nM | Song et al. [55] | ||||
| MDA-MB-231-LM3-3 | 16.6 nM | Song et al. [55] | |||
| SUM149PT | 16.6 nM | Song et al. [55] | |||
| SUM159PT | 15.9 nM | Song et al. [55] | |||
| Cervical | Hela | 154 ± 21.5 nM | 37.5 ± 2.15 nM | 11.8 ± 2.13 nM | Pan et al. [127] |
| Colorectal | SW620 | 76.72 ± 6.21 nmol/L | 34.05 ± 4.21 nmol/L | 16.7 ± 6.37 nmol/L | Zhu et al. [128] |
| 287.35 ± 4.34 nM | 57.63 ± 7.80 nM | 20.39 ± 1.95 nM | Zhang et al. [129] | ||
| HCT116 | 82.6 μmol/L | Qiu et al. [130] | |||
| 0.243 μM | 0.024 μM | Wang et al. [131] | |||
| LoVo | 56.778 ± 7.34 nM | 11.48 ± 2.89 nM | 6.64 ± 2.79 nM | Zhang et al. [129] | |
| Endometrial | Ishikawa | 0.7 ng/mL | Takai et al. [132] | ||
| HHUA | 0.5 ng/mL | Takai et al. [132] | |||
| HEC-1B | 0.6 ng/mL | Takai et al. [132] | |||
| Esophageal | Eca-109 | 1.0 μM | Lv et al. [133] | ||
| EC9706 | 1.0 μM | Lv et al. [133] | |||
| TE5 | 2.6 μM | Lv et al. [133] | |||
| TE11 | 4.9 μM | Lv et al. [133] | |||
| Hec2 | 3.8 μM | Lv et al. [133] | |||
| Gallbladder | GBC-SD | 48.12 ± 2.03 nM | 28.23 ± 1.78 nM | Jiang et al. [45] | |
| SGC996 | 125.03 ± 5.16 nM | 102.78 ± 3.21 nM | Jiang et al. [45] | ||
| Gastric | SGC7901 | > 800 nmol/L | Li et al. [134] | ||
| MGC803 | 160 ± 0.87 nmol/L | Li et al. [134] | |||
| Liver | HepG2 | 182.30 ± 13.78 nM | Li et al. [27] | ||
| 307 ± 87 nM | Ozdemir et al. [84] | ||||
| 33.65 nmol/L | Gao et al. [60] | ||||
| 143.2 nM | Miao et al. [49] | ||||
| PLC/PRF/5 | 52.20 ± 14.16 nM | Li et al. [27] | |||
| 157.87 nmol/L | Gao et al. [60] | ||||
| SMMC7721 | 97.74 ± 8.83 nM | Li et al. [27] | |||
| SK-HEP-1 | 110.33 ± 5.32 nM | Tsai et al. [135] | |||
| Leukemia | NB4 | 40 nmol/L | 27 nmol/L | 17 nmol/L | Zhai et al. [136] |
| K562 | 0.0943 μmol/L | Zhai et al. [136] | |||
| K562/VCR | 0.0401 μmol/L | Zhai et al. [136] | |||
| HEL | 0.046 μmol/L | Wang et al. [137] | |||
| Lung | A549 | 4.5 nM | Wang et al. [56] | ||
| 56.14 ± 6.72 nmol/L | 15.57 ± 4.28 nmol/L | Zhu et al. [138] | |||
| 8.15 ± 0.69 nmol/L | Sun et al. [92] | ||||
| 22.00 ± 3.53 nM | 10.20 ± 1.01 nM | Liu et al. [36] | |||
| Melanoma | A375.S2 | 450.38 nM | Hsiao et al. [44] | ||
| Oral | CAL27 | 125 nM | Tsai et al. [139] | ||
| 122.6 nM | Tian et al. [140] | ||||
| Osteosarcoma | U-2OS | 0.297 μM | Zhang et al. [141] | ||
| 8.49 ± 2.1 μg/L | Yin et al. [142] | ||||
| U-2OS/MTX300 | 10.19 ± 1.7 μg/L | Yin et al. [142] | |||
| Saos-2 | 0.318 μΜ | Zhang et al. [141] | |||
| Ovarian | SK-OV-3 | 1.0 ng/mL | Takai et al. [132] | ||
| OMC-3 | 0.6 ng/mL | Takai et al. [132] | |||
| Tongue | SCC-4 | 300 nM | Chou et al. [143] | ||
| Pancreatic | Capan-2 | 159.2 nM | Tian et al. [140] | ||
| Non-cancerous | Breast epithelial cell 184D | 384.6 ± 36.5 nM | Clifford et al. [31] | ||
| Breast epithelial cell 184A | 295.9 ± 10.5 nM | Clifford et al. [31] | |||
| Mouse normal hepatocyte | No cytotoxicity at 10 μM | Song et al. [55] | |||
| HUVECs | 53.1 nmol/L | Qiu et al. [130] | |||
| Het-1Aa | 10.1 μM | Lv et al. [133] | |||
| PBMCsb | 16.5 ± 4.7 ng/mL | Yuan et al. [144] | |||
| Primary normal human endometrial epithelial cells | Little sensitivity to BF from 0.1 to 10 ng/mL | Takai et al. [132] | |||
| PBMCs | 16.5 ± 4.7 ng/mL | Yuan et al. [144] |
aHuman normal esophageal squamous cells
bNormal peripheral blood mononuclear cells (PBMCs) were isolated from three healthy volunteers (32 ± 9 years of age)
In vivo evaluation of Huachansu and its derivative Bufalin on tumor inhibition
To evaluate the anticancer potential of Bufalin in vivo, various xenograft mice models are carried out throughout cancer types such as breast, cervical, colorectal, liver, gallbladder, lung, and pancreatic cancer. The results are very compelling that Bufalin can inhibit xenograft tumor growth, increase sensitivity to chemotherapy, and prolong the survival rate of mice. Most of the studies reported a dosage of 0.1–2 mg/kg intraperitoneally (i.p.) administered at a frequency of once per day 5 days per week to every 3 days for a period ranging from 12 days to 6 weeks in mice xenograft model without inducing significant weight loss or adverse effects, as summarized in Table 2. In the recent 10 years, although in vivo studies have been increasingly used to assess the effect of Bufalin. Most of the studies have used the xenografts models and reported the attenuation of tumor growth rate as well as the reduction of tumor weight. Yet, the lack of orthotropic models in the Bufalin research led to a relative shortage in the evaluation of the systemic effect of Bufalin contributing to not only the tumor itself but to the microenvironment or tumor metastasis. Further studies are warranted to elucidate the related effects.
Table 2.
In vivo evaluation of Bufalin in mouse tumor models
| Cancer type | Drugs | Animal | Tumor models | Transplantation | Treatment | Results | Refs. | Year |
|---|---|---|---|---|---|---|---|---|
| Bone | Bufalin | Athymic nude mice; male; 7–8 weeks old | Rat breast sarcocarcinoma Walker 256 cell | Inoculate 2 × 105 cell into the intramedullary space of the mouse femur | 1 day after tumor inoculation, given by i.p. 0.5, 1, 1.5, or 2 mg/kg/day for 21 days | Cancer-induced pain relief; 50% reduction in bone tissue injury | Ji et al. [83] | 2017 |
| Cervical | Bufalin | BALB/c nude mice; female; 4–5 weeks old | Human cervical squamous cell carcinoma Siha cell | Subcutaneously inoculate 3 × 106 cell | After reaching an average tumor volume of 100 mm3, given by i.p. Bufalin 10 mg/kg ± paclitaxel (10 mg/kg per 4 days) for 32 days | Bufalin synergizes with paclitaxel to inhibit tumor growth without apparent loss of body weight | Liu et al. [80] | 2016 |
| Breast | Bufalin-NP | SCID mice; female; 6–7 weeks old | Human breast MDA-MB-231-LM3.3 cell | Inject 0.75 × 106 cell into one of the second mammary fat pads | 6 days after tumor injections, given by i.v. Bufalin-NP (1.5 mg/kg) three times per week for 20 days | Bufalin sensitizes cancer cells to MK-2206 and blocks tumor growth | Wang et al. [56] | 2014 |
| Bufalin-BCS-NP | Balb/c nude mice; female; 4 weeks old | Human breast MCF-7 cell | Subcutaneously implant 1 × 107 cell into the right axilla skin | 7–10 days after tumor inoculation, (1) given by i.p. BF-BCS-NPs (1 mg/kg BF equivalent in PBS) or free BF (1 mg/kg in ethanol) every 2 days for 20 days; (2) give by i.v. BF-BCS-NPs (5 mg/kg BF equivalent in PBS) at D1, D3 | Free BF suppresses tumor growth accompanying with a significantly decreased body weight. BF-BCS-NPs suppresses tumor growth without apparent loss of body weight | Tian et al. [104] | 2014 | |
| 3-Phospho-bufalin | Nude mice; female; 4–6 weeks old | Human breast MDA-MB-231-LM3-3 cell | Inject 0.75 × 106 cell into one of the second mammary fat pads | 14 days after injection, given by s.c. phospho-BF (0.75 mg/kg per dose, 3 doses per week) for 3 weeks | 2.4 times reduction in tumor weight; no cardiotoxicity observed | Song et al. [55] | 2015 | |
| Bufalin | Athymic nude mice; female; 5 weeks old | Human breast MDA-MB-231 cell | Subcutaneously inject 5 × 106 into both dorsal flank regions | 4 weeks after injection, (1) given by intra-tumoral injection BF 10 μL (1 mM, in 0.9% normal saline) to the left flank for 4 weeks; (2) 10 μL volume of normal saline to the right flank for 4 weeks | Significantly enhances breast cancer xenograft growth; promote the inflammatory response | Chen et al. [38] | 2017 | |
| Colorectal | Bufalin-loaded mPEG-PLGA-PLL-cRGD nanoparticles (BNPs) | Athymic nude mice; female; 4–6 weeks old | Human colon SW620 cell | Inject 5 × 106 cell into the dorsal subcutaneous space | Bufalin-loaded mPEG-PLGA-PLL-cRGD nanoparticles (BNPs) containing 1 mg/kg bufalin, given by injection through the vena caudalis once a day for 14 days | Suppresses tumor growth; bufalin-loaded NPs significantly enhanced treatment efficacy compared to that of a bufalin water solution | Yin et al. [105] | 2012 |
| Bufalin | Male athymic nude mice | HCT116-luc-vector and HCT116-luc-miR-497 | Inject 1 × 106 cells intravenously via the tail vein | 1 week after injection, given by Bufalin for 1 mg/kg via the tail vein (three times a week) for 5 weeks | Inhibited colorectal cancer metastasis; improved life of survival. Improved physiological characteristics in terms of body weight, skin roughness, mental status, and survival rate | Qiu et al. [130] | 2014 | |
| Bufalin-loaded pluronic polyetherimide nanoparticles | Male athymic nude mice | HCT116 | Injecte 1 × 106 cells intravenously via the tail vein. | 2 weeks after injection, (1) given by of Bufalin 1 mg/kg; (2) given by 20 mg/kg of Bufalin-loaded pluronic PEI nanoparticles via the tail vein (0.2 mL per mouse, three times per week) for 3 weeks | Inhibited colorectal cancer metastasis. Improved quality of life and physiological characteristics in terms of body weight, skin roughness, and mental status | Hu et al. [93] | 2014 | |
| Bufalin | BALB/c mice; male; 5–6 weeks old | HCT116 | Inject 2 × 1010 cells s.c. into the right axillary region. Tumor mode of the second generation: 2 weeks after injection, harvest the s.c. xenogra tumors, cut into pieces (1.5 mm in diameter), implant into the axillary region s.c. Orthotopic xenogra model: harvest third generation s.c. tumors and cut into pieces (1.5 mm in diameter) | NS group (treated with 0.2 mL normal saline); 5-Fu group (treated with 5-FU, 25 mg/kg); low Bufalin group (0.5 mg/kg); medium Bufalin group (1.0 mg/kg); high Bufalin group (1.5 mg/kg). NS, 5-FU, and Bufalin were administrated by intraperitoneal injection, once per day from day 15 to day 21 (12 mice in each group) | Inhibit cell growth. Lower tumor volume. Prolong survival time | Wang et al. [131] | 2015 | |
| Bufalin-DOX | Athymic nude mice (BALB/c-nu/nu) of 6–8 weeks | HCT8/ADR | Inject 1 × 107 cells s.c. under the shoulder in the nude mice | Mice were randomized into six groups (6 in each group) when the tumor volumes reached 150–200 mm3: control; BF (0.1 mg/kg, i.p., q3d × 5); DOX (0.1 mg/kg, i.p., q3d × 5); DOX (0.5 mg/kg, i.p., q3d × 5); DOX (1.0 mg/kg, i.p., q3d × 5); DOX (0.1 mg/kg, i.p., q3d × 5) plus BF (0.1 mg/kg, i.p., q3d × 5, given 1 h before DOX administration) | BF remarkable increased the effect of DOX against the ABCB1 resistant HCT8/ADR colorectal cell xenografts in nude mice | Yuan et al. [145] | 2015 | |
| Bufalin | Male nude mice (BALB/c nu/nu, 5-week-old) | HCT116 | Inject 2 × 106 cells into the subcutaneous tissues. | 2 weeks after injection, (1) given by cisplatin (10 mg/kg body weight) i.p. every 3 days for 4 weeks.; (2) given by bufalin (1 mg/kg body weight) i.p. every 3 days for 4 weeks | Inhibition of tumor growth and tumor tissue weights are greater with the combination of cisplatin and bufalin than with cisplatin alone. Tumors treated with the combination of cisplatin and bufalin showed more cell vacuolization and nuclear shrinkage than with cisplatin alone | Sun et al. [146] | 2017 | |
| Gallbladder | Bufalin | Male athymic nude mice (5 week-old) | GBC-SD | Xenograft model: Inoculate 1 × 106 GBC-SD cells into the left axillary region | 24 h postinoculation, given by PBS i.p. and bufalin with (0.1, 0.2, and 0.4 mg/kg) i.p. every 2 days for up to 20 days | Suppression of tumor growth | Jiang et al. [45] | 2014 |
| Liver | Bufalin | BALB/c nu/nu mice (18–20 g, 5 week-old) | HCCLM3 | Inject 5 × 106 cells s.c. into the upper left flank region of nude mice. Revmove the tumors when reached approximately 1 cm in length (approximately 4 weeks after injection) and mince into small pieces of equal volume (1.5–2 × 2 × 2 mm3), then transplant into the livers of 24 nude mice | From day 8 to 38, given by1 mg/kg Bufalin; 1.5 mg/kg Bufalin; 100 mg/kg LY294002 and saline i.p. thrice weekly, respectively | 1.5 mg/kg Bufalin decreased the sizes and qualities of orthotopic transplanted tumors as well as pulmonary metastasis. Orthotopic transplanted tumor tissues were necrotic and the apoptotic cell number was markedly higher in 1.5 mg/kg Bufalin group. Inhibition of AKT/GSK3β/β-catenin/E-cadherin signaling pathways | Zhang et al. [72] | 2014 |
| Bufalin–sorafenib | BALBc nu/nu mice (6 week-old) | Human HCC cell lines SMMC7721 | Inoculate 5 × 106 cells s.c. into the abdominal intraderma | Control group: inject with the vehicle i.p. (5 days/week, 2 weeks). Experimental group: (1) injection of 1 mg/kg bufalin i.p. (5 days/week, 2 weeks); (2) oral uptake of 30 mg/kg/day sorafenib (5 days/week, 2 weeks); (3) the combination of both injections of bufalin i.p. and oral uptake of sorafenib (5 days/week, 2 weeks) | Inhibit blood vessel formation in the intradermal tumors, manifested by the vessel numbers and branches and attenuate tumor weight in nude mice with the combination treatment | Wang et al. [64] | 2016 | |
| Bufalin | BALBc nu/nu mice (6 week-old) | Human HCC cell lines SMMC7721 | Inoculate 5 × 106 cells s.c. into the right flank | Tumor size was measured every 4 days after the treatment. Tumor-bearing mice were sacrificed after 16 days of treatment, and the tumor weight was evaluated | Combination treatment inhibits tumor growth and tumor angiogenesis in vivo. The combination treatment group showed more reduced microvessel density than any other group |
Wang et al. [64] | 2016 | |
| Bufalin | BALBc nu/nu mice (6 week-old) | SMMC7721-GFP | Inoculate 5 × 106 cells s.c. into the right flanks. 4 weeks later, cut the non-necrotic tumor tissue into 1 mm3 pieces and orthotopically implanted into the liver. In addition, inject 2 × 106 via mouse tail veins | (1) Inject 1 mg/kg bufalin i.p. (5 days/week for six weeks); (2) Inject PBS i.p. (5 days/week for 6 weeks) | Bufalin reduced liver/lung metastases. Bufalin inhibited invasion through EMT | Wang et al. [76] | 2016 | |
| Myeloma | Bufalin-MK2206 | BALB-c nu/nu female mice (4–6 week-old); NOD-SCID female mice (4–6 week-old) | MOPC315; H929 | Inject 2 × 107 MOPC315 cells s.c. in the right flanks of the BALB-c nu/nu mice. Inject 1 × 107 cells H929 cells s.c. in the right hind leg of NOD/SCID mice | Mice bearing MOPC315 MM tumors were treated with bufalin (1 mg/kg; intraperitoneally) daily in the presence and/or absence of MK2206 (120 mg/kg orally) for 10 days. Mice injected with H929 MM cells were treated with 1 mg/kg bufalin daily with or without 120 mg/kg MK2206 for 12 days | Bufalin combined with MK2206 blocked MM tumor growth, decreased tumor cell proliferation and increased the percentage of apoptotic cells | Xiang et al. [147] | 2017 |
| Osteosarcoma | Bufalin | BALB/c nude mice; female; 6 week-old | U2OS/MTX300 | Inject 5.6 × 106 cells s.c. into the axilla of the mice | 10 days after injection, (1) control group: 100 mL of vehicle i.p.; MTX group: MTX (250 mg/kg) with calcium leucovorin rescue (24 mg/kg at 16, 20, or 24 h after MTX) i.p. per week; (3) low Bufalin group: 0.75 mg/kg i.p.; (4) High Bufalin group:1.5 mg/kg i.p. | Bufalin inhibited tumor growth | Xie et al. [86] | 2012 |
| Pancreas | Bufalin | BALB/c nu/nu mice; male; 4-week-old | Mia PaCa-2 | Inject 6 × 106 cells s.c. into the back of mice | When tumors reached the size of 100 mm3: (1) vehicle alone (control); (2) Bufalin (0.1 mg/kg, for 10 days); (3) Gemcitabine (125 mg/kg, 3 times/week for 2 weeks); (4) Bufalin and Gemcitabine in combination | Bufalin potentiates the anti-tumor effect of gemcitabine in vivo. Combination treatment with gemcitabine signicantly reduced the tumor volume and cell proliferation activity | Chen et al. [148] | 2012 |
| Bufalin | BALBc nu/nu mice; female; 6-week-old | MiaPaCa2/GEM | Inoculated 2 × 105 cells into the right flanks of mice | Given by (1) injections of 1.5 mg/kg bufalin (5 days/week) for 4 weeks; (2) injections with vehicle (20 μL saline) for 4 weeks. In addition, MiaPaCa2/GEM cells (2 × 106) was given to one of another two groups pre-treated with bufalin via the tail veins for 6 weeks | Inhibit pancreatic tumor growth | Wang et al. [82] | 2016 | |
| Bufalin | BALBc nude mice; male; 5-week-old) | BxPC3-luc2 | Injected 1 × 107 cells s.c. into the left buttock of mice | 7 days after inoculation, control group: inject saline i.p.; Bufalin groups: inject 1 mg/kg and 2 mg/kg for 14 days; Positive control: DDP 2 mg/kg every other day i.p. for 14 days | Bufalin treatment inhibits tumor growth | Liu et al. [149] | 2016 | |
| Lung | Bufalin | BALB/c nu/nu mice; male; 6–8 week-old | NCI-H460 | Inject 1.3 × 107 cells s.c. into flank of each mouse | When tumor volume exceeded 100 mm3, given by (1) vehicle (0.1% DMSO); (2) Bufalin: 0.1, 0.2, or 0.4 mg/kg for 14 days | Bufalin suppresses tumor growth | Wu et al. [150] | 2017 |
NP nanoparticles, BSC biotin modified chitosan, s.c. subcutaneously, i.p. intraperitoneally
The acute toxicity measured as 50% lethal dose values for Bufalin in mice when administered intraperitoneally was approximately 2.2 mg/kg, as first reported in1960 by Okada et al. [34] by Wang et al. in 2003 [35] and by Liu et al. in 2016 [36], consistently. Bufalin showed the lowest toxicity when injected intraperitoneally among principle active components, including cinobufagin and resibufogenin, isolated from CS [34]. When administered intravenously (i.v.) to rabbits, Bufalin induced an elevation of blood pressure accompanied by marked respiratory excitation. Yet, although Bufalin is structurally slimier to digitoxin that both of them possess the same aglycone except for the lactone ring in 17-position, digitoxin does not induce respiratory excitation induced by the i.v. administration of Bufalin was not observed with digitoxin intervention at a dosage as high as 0.2 mg/kg via i.v. injection [34]. A study in 1995 examined the toxicity and teratogenicity of CS using a single i.p. injection into maternal mice during the embryonic organogenesis [37]. At doses below 50 mg (dry weight)/kg body weight, no detectable changes were observed in both maternal mice and fetuses; however, at a dose of 50 mg/kg, adverse effects were found with a reduction in maternal mice body weight, liver, and kidney structural abnormalities, and an increased number of resorbed and dead fetuses [37].
It is worth noting that most studies have reported a significant reduction in tumor growth rate and tumor weight after i.p. injection of Bufalin. An article reported a contradictory result in a breast cancer xenograft model with an intra-tumoral injection of Bufalin in 2017 [38]. The results showed that intra-tumoral injection of 10 μL of 1 μM Bufalin, three times a week for 4 weeks, equivalent to 3.87 ng per mice, significantly enhanced tumor growth by promoting inflammatory response compared with intratumoral injection of saline via increasing expression of COX-2/IL-8, promoting p-65 NF-κB translocation and modulating mitogen-activated protein kinase (MAPK) pathways [38]. This contradictory result may be due to the difference in not only the route of administration but also the difference in the equivalent dose among studies.
As previously reported, for example, Han, et al. conducted a study in 2007 and reported an anti-tumor effect of Bufalin with i.p. injection of 0.5, 1.0 or 1.5 mg/kg/day for 10 days in mice hepatocellular carcinoma (HCC) xenograft model with a body weight around 18–20 g, which is equivalent to an i.p. injection of 10 to 30 mg/mice of Bufalin throughout the course of treatment [39]. Other studies, as summarized in Table 2, reported anti-tumor effect with i.p. administration of doses ranging from 0.5 to 1.5 mg/kg at a frequency of daily injection for 10 days, every other day injection for 20 days or once every 3 days for 4 weeks. There appears to be more than a 1000-fold difference between the anti-tumor effect reported by i.p. injection and the pro-inflammatory or tumor-promoting effects upon intra-tumoral injection. A variation in doses is also found. According to the material and safety data sheet, the median lethal dose of Bufalin administered to mice by i.v. injection was 0.74 mg/kg [38]. It is plausible that the contradictory results on the tumor suppressing or the tumor-promoting effects of Bufalin may be due to the variation among different cancer cell lines, different route of administration, or different doses used. In future research, more in-depth study using the same treatment protocol is needed to better assess the role of Bufalin in cancer treatment.
Molecular mechanisms of anti-tumor activity
The cell cycle arresting, intrinsic (also known as mitochondria-mediated) and extrinsic (also known as receptor-mediated) apoptosis-inducing effects of Bufalin at concentrations ranging from 1 nM to 10 μΜ, such as intracellular reactive oxygen species (ROS) production, caspase-dependent apoptosis, modulating MAPK signaling cascade, inhibiting NF-κB signaling, are well not only well established among various cancer cell lines [29, 40–53]. The steroid receptor coactivator (SRC) family, including SRC-1, SRC-2, and SRC-3, is involved in molecular and physiological processes across diseases by activating nuclear receptors and other transcriptional factors, such as NF-κB. SRCs are frequently overexpressed in malignancies and are associated with cancer cell proliferation, invasion, and metastasis [54–56]. Previous studies suggested that Bufalin promotes SRC-3 protein degradation in breast, gastric and lung cancer [36, 55–58]. Moreover, Bufalin suppressed triple negative breast cancer proliferation at a nanomolar concentration (~ 3–5 nM), which the concentration of which digoxin required to inhibit SRC-3/SRC-1 is greater than 200 nM [56]. The effective concentration of Bufalin to inhibit SRCs is within the on cardiac toxicity concentration reported in patients plasma (~ 9 nM) [12]. Furthermore, the water soluble prodrug of Bufalin, 3-phospho-bufalin, inhibits the growth of orthotopic triple negative breast cancer [55]. All in all, Bufalin is a potent and safe steroid receptor coactivators inhibitor.
Although the exact mechanism of the anti-cancer effect of Βufalin has hitherto been unknown. It is worth noting that several novel mechanisms contributing to the anti-cancer effects of Bufalin are proposed and increasingly studied in the recent 10 years, including sensitization of TRAIL-mediated apoptosis, autophagic cell death induction, reversing chemotherapy drug resistance, suppressing cellular invasion, migration, and anti-adhesion, inhibiting epithelial–mesenchymal transition, as shown in Table 3. A schematic summary of the molecular mechanism of Bufalin-mediated anti-cancer effect is shown in Fig. 3.
Table 3.
Summary on the pharmacological action of Huachansu extract (HCS) and Bufalin (BF) reported in recent 10 years
| Cancer type | Study type | Pharmacological action | Pathways involved | Refs. | Year |
|---|---|---|---|---|---|
| Bladder | In vitro | Growth inhibition (50–300 nM, 24H); G2/M arrest; apoptosis induction (T24, EJ) [BF] | Bax/Bcl-2 ratio Fas, DR4, DR5, TRAIL, PARP cleavage↑; pro-caspase-3, -8, -9, Bcl-xL, Bid, XIAP, cIAP-1, cIAP-2↓ | Hong et al. [43] | 2012 |
| In vitro | Growth inhibition (50–200 nM, 24H); G0/G1 arrest; mitochondrial/apoptosis (T24) [BF] | ROS production, cytochrome c, Apaf-1, AIF, caspase-3, -7, -9, Bax↑; ΔΨm, cyclin D, CDK4, cyclin E, CDK2, phospho-Rb, phospho-AKT Bcl-2↓ | Huang et al. [29] | 2012 | |
| In vitro | Migration and invasion inhibition (5–100 nM, 24H) (T24) [BF] | TIMP-1, -2, phospho-ERK↑; claudin-2, -3, -4, MMP-2, -9, Active-MMP-2, -9↓ | Hong et al. [151] | 2013 | |
| In vitro | Sensitization of TRAIL-mediated apoptosis (2.5–10 nM, 24H) (T24) [BF] | Caspase-9, pro-caspase-9, DR5, degradation of poly (ADP-ribose) polymerase↑ | Kang et al. [152] | 2017 | |
| Bone | In vivo | Relieves cancer-induced pain and bone destruction (Walker 256 cells) [BF] | OPG↑; RANKL, serum TRACP5b, ICTP, PINP↓ | Ji et al. [83] | 2017 |
| Breast | In vitro | Sensitization of TRAIL-mediated apoptosis (0.02–0.25 μM, 24H) (MCF-7, MDA-MB-231) [BF] | Mcl-1, Bcl-xL, p-STAT3↓ | Dong et al. [153] | 2011 |
| In vitro | Growth inhibition (50 nM, 24H); apoptosis induction; enhanced TRAIL-induced apoptosis (MCF-7, MDA-MB-231) [BF] | DR4, DR5, p-ERK, p-JNK, p-p38, cleaved-PARP↑; pro-caspase-8, Cbl-b↓ | Yan et al. [125] | 2012 | |
| In vitro and in vivo | Growth inhibition (1–5 nM, 24H); promote coactivators’ protein degradation (MCF-7) [BF] | SRC-3 mRNA expression, SRC-3 protein degradation↑; intrinsic transcriptional activities of SRC-1 and SRC-3↓ | Wang et al. [56] | 2014 | |
| In vitro | Sensitization of TRAIL-mediated apoptosis (50 nmol/L, 24H); redistributing death receptors in lipid rafts (MCF-7, MDA-MB-231) [BF] | DR4, DR5, cleaved-PARP, cleaved-caspase-8↑; caspase-8↓ | Yan et al. [154] | 2014 | |
| In vitro and in vivo | Growth inhibition (100 ng/mL, 8H); mitochondrial/apoptosis (MCF-7) [BF, BF-BCS-NPs] | Intracellular ROS, BAX, cleaved-caspase-3↑; ΔΨm, Bcl-2, survivin↓ | Tian et al. [104] | 2014 | |
| In vitro | Growth inhibition; apoptosis (10–50 nmol/L, 48H) (MCF-7/ADR, MDA-MB-231)[BF] | PARP cleavage, miR-155-5p expression↑; DNMT1, DNMT3a, FOXO3a↓ | Wang et al. [126] | 2016 | |
| In vitro and in vivo | Growth inhibition (12–200 nM) (LM3-3) [BF, phospho-BFs] | Synergizes with Gefitinib; SRC-3↓ | Song et al. [55] | 2015 | |
| In vitro | Growth inhibition (4–8 nM, 48H) (MDA-MB-231); enhanced HDAC inhibitors induced apoptosis [BF] | ↑;SRC-3p-Akt, Bcl-2↓ | Zou et al. [155] | 2016 | |
| In vitro and in vivo | Promote inflammatory response (0.001–1 μM, 12H) (MDA-MB-231) [BF] | p65 translocation; PKC-induced COX-2 and IL-8, PGE2, p-JNK, p-p38, p-ERK, TPA-induced MMP-3 protein and mRNA expression↑ | Chen et al. [38] | 2017 | |
| Cervical | In vitro and in vivo | Growth inhibition (0.05–0.2 μM, 24H); G2/M arrest; apoptosis; migration and invasion inhibition (0.01–0.04 μM, 24H) (Siha, Hela) [BF] | BAX, P21, p27, E-cadherin, GSK3β↑;Bcl-2, Bcl-xL, cyclin A, cyclin B1, CDK2, MMP-9, SNAIL1, integrin α2, integrin β5, FAK, p-FAK(Tyr397), p-GSK3β(Ser389), AKT1, p-AKT(Ser473)↓ | Liu et al. [80] | 2016 |
| In vitro | Growth inhibition (0–50 nM, 24H); apoptosis induction (Hela) [BF] | HSP27, vimentin, HNRPK↓ | Pan et al. [127] | 2012 | |
| Colorectal | In vitro | Growth inhibition (25–100 nM, 48H); G2/M arrest; autophagy induction (HT-29, Caco-2) [BF] | LC3-II, ROS, Atg-5, Beclin-1, p-JNK2↑ | Xie et al. [59] | 2011 |
| In vitro | Growth inhibition (20, 80 nmol/L, 24H); G2/M arrest; apoptosis (SW620) [BF] | PARP cleavage, cleaved-caspase-3, BAX/BCL-2 ratio↑;p-Stat3, p-ERK, livin↓ | Zhu et al. [128] | 2012 | |
| In vitro | Growth inhibition (100 nM, 9H); mitotic arrest; G2/M arrest (HT-29, HCT-116) [BF] | p-H3↑;HIF-1α, NF-κB, Plk1 expression↓ | Xie et al. [156] | 2013 | |
| In vitro and in vivo | Anti-migration and anti-metastasis (3.12–50 nM, 12H) (HCT116), ex vivo micro vessel sprouting (HUVECs) [BF] | miR-497 expression↑; VEGFA expression↓ | Qiu et al. [130] | 2014 | |
| In vitro and in vivo | Growth inhibition (0.03–3 μΜ, 24–48H); G2/M arrest; apoptosis (HT116) [BF] | PTEN, Bad, caspase-3 phosphorylation, caspase-3 cleavage↑; p-PTEN, p-AKT↓ | Wang et al. [131] | 2015 | |
| In vitro | Anti-proliferation and anti-migration (10–50 nM, 48H); induce apoptosis; G2/M arrest (LoVo, SW620) [BF] | Cyclin B1, p–cdc2, p21, cleaved-PARP, Bax, cleaved-caspase-7, -9, E-cadherin↑; Bcl-2, N-cadherin, β-catenin, CPSF4, hTERT↓; inhibit hTERT by down-regulating CPSF4 | Zhang et al. [129] | 2016 | |
| In vitro and in vivo | Suppress growth of cisplatin-resistant cell rather than sensitive-one; reverse ABCB1-mediated multi-drug resistance (5–20 nM, 48H) (LoVo/ADR, HCT8/ADR, HCT8/ABCB1) [BF] | ATPase activity of ABCB1↑; ABCB1↓ | Yuan et al. [145] | 2015 | |
| In vitro and in vivo | Reverse cisplatin drug resistance (5 nM, 48H) (HCT116, LoVo HCT116-STSCscis, LoVo-STSCscis) [BF] | CD133, CD44, OCT4, SOX2, NANOG, ABCG2 ↓ | Sun et al. [146] | 2017 | |
| Gallbladder | In vitro | Apoptosis induction (25–200 nmol/L, 48H); S-phase arrest; mitochondrial dysfunction induction (GBC-SD, SGC996) [BF] | Cleaved-caspase-3, -9, cleaved-PARP ↑; Cyclin A, Cyclin B1, Cyclin D1, CDK1, NF-κB, Bcl-2, ΔΨm↓ | Jiang et al. [45] | 2014 |
| Gastric | In vitro | Growth inhibition (100, 200 nmol/L, 24H); G2/M phase arrest; intrinsic apoptosis induction (SGC7901, MGC803) [BF] | SPARC antagonizes bufalin-induced apoptosis; p-Src, p-Akt, p-ERK↑; ΔΨm, cyclin B1, cyclin A↓ | Li et al. [134] | 2015 |
| In vitro | Growth inhibition (25–200 nmol/L, 48H); apoptosis induction (SGC7901, MGC803)[BF] | Bax, cleaved-PARP↑; Bcl-2, pro-caspase-3, miR-298 suppressed apoptosis↓ | Zhao et al. [157] | 2015 | |
| In vitro | Proliferation inhibition (100 nM, 48H); cisplatin-sensitization; apoptosis induction (SGC7901, MKN-45, BGC823) [BF] | p-AKT, p-GSK3β, p-mTOR, p-4EBP1, p-S6 K↓ | Zhao et al. [158] | 2016 | |
| In vitro | G0/G1 phase arrest (50, 80 nmol/L, 48Η); caspase and mitochondrial-mediated apoptosis induction; ER stress induction; protective autophagy activation (SGC7901, BGC823) [BF] | Cleaved-caspase-3, cleaved-PARP, Bax/Bcl-2, CHOP, p-eIF2a, p-JNK, LC3-II, Atg5, Beclin-1↑; LC3-I, p62↓ | Zhao et al. [159] | 2017 | |
| Glioma | In vitro | Growth inhibition (20–80 nM, 24H); mitochondria-mediated apoptosis induction; autophagy activation (U87MG, LN229) [BF] | ROS production, Bax, cytosolic cytochrome c, cleaved-PARP, cleaved-caspase-3, -4, LC3-II, p-AMPK, p-ACC, ATF6f, p-PERK, p-IRE1α, p-eIF2α, GRP78, GRP994, CHOP↑; Bcl-2, p-mTOR, p-4EBP1, p-p70S6K, PERK↓ | Shen et al. [160] | 2014 |
| In vitro | Proliferation inhibition (5–40 μM, 24H); cancer stem cell-like phenotypes inhibition; apoptosis induction (U251, U87) [BF] | miR-203↑;OCT4, SOX2, SPARC↓ | Liu et al. [161] | 2017 | |
| HCC | In vitro | Growth inhibition (0.1–1 μΜ, 2H) (HepG2, PLC/PRF/5, SMMC7721) [BF] | p-ERK, p-Akt, ATP1A3↑; FoxO3a↓ | Li et al. [27] | 2011 |
| In vitro | Growth inhibition (0.001–0.1 μmol/L, 24–72H); induction of fas- and mitochondria-mediated apoptosis (HepG2) [BF] | Bax, cytochrome c, cleaved-caspase-3, -9, PARP cleavage ↑; ΔΨm, Bcl-2, pro-caspase-3, -9, -10, Bid ↓ | Qi et al. [162] | 2011 | |
| In vitro | Anti-proliferation (50–250 nM, 48Η); enhances the anti-cancer effects of Sorafenib (6.25lM) (PLC/PRF/5, HepG2) [BF] | Enhanced apoptotic cell death in combination with Sorafenib; p-Akt↑;p-ERK↓ | Gao et al. [60] | 2012 | |
| In vitro | Growth inhibition (50–100 nM, 24H); G2/M arrest; autophagy induction (SK-HEP-1)[CS] | Chk1, Wee1, LC3-II, Atg5, Atg7, Atg12, Beclin-1↑; Cyclin A, cyclin B, CDK1, p-CDK1(Thr161), Cdc25c, p-CDC25c(Ser198), p-Akt(Ser308), p-AKT(Ser473), p-mTOR(Ser2481), AKT kinase activity↓ | Tsai et al. [135] | 2012 | |
| In vitro | Growth inhibition (10–100 nM, 48H); autophagy induction; apoptosis induction (HepG2) [BF] | p-APMK, Beclin-1, LC3-II, p-p70S6K↑; p-mTOR, p62↓ | Miao et al. [49] | 2013 | |
| In vitro | Growth inhibition; G2/M arrest (0.04 μM, 4–12H); autophagy induction (Huh7, Hep3B, HA22T) [BF] | TNF, BECN-1, MAPK, ATG8↑; Bcl-2, Bid↓ | Hsu et al. [30] | 2013 | |
| In vitro | Growth inhibition; anti-migration (10, 100 nmol/L, 48H); anti-invasion; anti-adhesion (HCCLM3, HepG2) [BF] | GSK3β, E-cadherin↑; p-Akt, p-GSK3β, MMP-9, -2, β-catenin nuclear translocation↓ | Qiu et al. [71] | 2013 | |
| In vitro | Growth inhibition (5, 10 nM, 48H); anti-migration and invasion (SK-Hep1) [BF] | PI3K, p-Akt, NF-κB translocation, MMP-2, -9, FAK, Rho A, VEGF, MEKK3, MKK7, uPA↓ | Chen et al. [68] | 2013 | |
| In vitro | Growth inhibition (50–100 nmol/L, 48H); apoptosis induction; ER stress induction; autophagy induction (Huh-7, HepG2) [BF] | Beclin-1, p-JNK1, p-JNK2, IRE1, ATG5, LC3-I, LC3-II↑; p62↓ | Hu et al. [163] | 2014 | |
| In vitro | Reverse multidrug resistance (1 nM, 48H); G0/G1 phase arrest (BEL-7402/5-FU) [BF] | drug efflux pump activity, TS, MRP1, Bcl-xL/Bax ratio ↓ | Gu et al. [164] | 2014 | |
| In vivo | Orthotopic growth inhibition, anti-metastasis (1 mg/kg and 1.5 mg/kg) (HCCLM3-R) [BF] | p-Akt, GSK3β, E-cadherin↑; p-GSK3β, β-catenin, MMP-9, -2↓ | Zhang et al. [72] | 2014 | |
| In vitro | Reverse Sorafenib resistance (50–200 nM, 48H); synergies with sorafenib to induce apoptosis (HepG2, HepG2-Sora, Huh7, Huh7-Sora) [BF] | IRE1, CHOP, P-eIF2α↑; p-Akt↓ | Zhai et al. [165] | 2015 | |
| In vitro | Anti-invasion and metastasis (0.085 μg/mL, 72H) (BEL-7402) [BF] | E-cadherin, ALB↑; β-catenin, p-GSK-3β Ser9, MMP-7, COX-2, Cyclin D1, AFP↓ | Gai et al. [66] | 2015 | |
| In vitro and in vivo | Synergies anti-angiogenic effect of sorafenib (2.5–10 nM, 48H); anti-migration; S-phase arrest (HUVECs, SMMC7721) [BF] | p-ERK↑; VEGF, p-Akt, p-mTOR↓ | Wang et al. [64] | 2016 | |
| In vitro and in vivo | Inhibits TGF-β1 induced EMT and invasion (10 nM, 24–72H) (SMMC7721) [BF] | E-cadherin↑; N-cadherin, Vimentin, Snail, HIF-1α↓ | Wang et al. [76] | 2016 | |
| In vitro | Growth inhibition (40–200 nmol/L, 24–48H); apoptosis induction (Hep3B)[BF] | Apolipoprotein E (APOE) knockdown induced Na+/K+-ATPase, caveolin, PI3K/AKT/GSK3b and apoptosis signal cascades↑;Cyclin D1, Cdc25c, Cdc2↓ | Liu et al. [166] | 2016 | |
| In vitro | Growth inhibition (0.04 μg/mL, 72H); S- and G2-phase arrest; apoptosis induction; anti-migration and invasion; adhesion inhibition (HCC-LM3) [BF] | E-cadherin↑; β-catenin, MMP-2, -9, VEGF↓ | Sheng et al. [67] | 2016 | |
| In vitro | Growth inhibition (0.1 mg/mL, 24H); apoptosis induction (HepG2, HLE) [HCS] | Bcl-2↑;ΔΨm, Bax, Bid, cytochrome c↓ | Xia et al. [167] | 2017 | |
| Leukemia | In vitro | Vitamin D-induced cell differentiation enhancing (7.5, 10, 12.5 nM, 24H); VDR transactivation activity enhancing (HL-60, THP-1, U937) [BF] | 1,25(OH)2D3-induced CYP24A1, CD14, CAMP, PTGS1, CD11b, CDKN1A expression, nuclear VDR expression, histone acetylation and VDR recruitment to the CYP24A1 promoter, Erk MAP kinase activation↑ | Amano et al. [168] | 2009 |
| In vitro | Proliferation inhibition (5–80 nmol/L, 12–72H); apoptosis induction (NB4) [BF] | Synergized with PD98059; caspase-3 activation↑; survivin expression↓ | Zhu et al. [169] | 2012 | |
| In vitro | Reverse multidrug resistance (0.001–0.1 μM, 48H); S-phase arrest (K562, K562/VCR) [BF] | Bax↑; MRP1, Bcl-xL↓ | Zhai et al. [136] | 2014 | |
| In vitro | Growth inhibition (0.01–0.5 μmol/L, 48H); apoptosis induction; cell cycle arrest (HEL) [BF] | WT1 gene methylation, DNMT3a, DNMT3b protein↑; WT1 mRNA expression↓ | Wang et al. [137] | 2017 | |
| Lung | In vitro | Growth inhibition (0.1 μM, 24–48H); apoptosis induction; ROS-dependent mitochondrial dysfunction (ASTC-a-1) [BF] | ROS production, Bax translocation from cytosol to mitochondria, caspase-3 activation↑ | Sun et al. [50] | 2011 |
| In vitro | Growth inhibition (2.5–10 μM, 48–72H); apoptosis induction; G1-phase arrest (A549) [BF] | Cyto C (Cytosol)/Cyto C (mitochondrial), cleaved-caspase-3, cleaved-PARP, p53, p21Waf↑; Bcl-2/Bax ratio, Cyclin D1, COX-2, p-VEGFR2, p-VEGFR1, p-EGFR, p-Akt, p-p38MAPK, p-NKκB, p-ERK1/2↓ | Jiang et al. [46] | 2010 | |
| In vitro | Growth inhibition (20–100 nM, 48–72H); apoptosis induction (A549) [BF] | Bax, cleaved-caspase-3↑; Bcl-2, livin, p-Akt↓ | Zhu et al. [138] | 2012 | |
| In vitro | Growth inhibition (20 nM, 72H); reverses HGF-resistance to EGFR-TKIs; apoptosis induction (HCC827, PC-9, H1975) [BF] | Cleaved-caspase-3, -9, cleaved-PARP↑; blockage of Met/PI3k/Akt pathway | Kang et al. [170] | 2013 | |
| In vitro | Growth inhibition (1–4 μM, 24H); induce DNA condensation (NCI-H460) [BF] | DNA-PK, BRCA1, 14-3-3-σ, MDC1, MGMT, P53↓ | Wu et al. [70] | 2014 | |
| In vitro | Growth inhibition (2–8 ng/mL, 48H); apoptosis induction (A549) [BF] | Cleaved-caspase-3, cytosol cytochrome c↑; ΔΨm↓ | Ding et al. [41] | 2014 | |
| In vitro | TGF-β induced epithelial-to-mesenchymal transition and migration inhibition (50 nM, 24H) (A549) [BF] | TGF-β receptor, I and II TGF-β induced E-cadherin, p-Smad2, p-Smad3↓ | Zhao et al. [79] | 2015 | |
| In vitro | Invasion and migration inhibition (25–100 nM, 24–48H); cell adhesion inhibition (NCI-H460) [BF] | RhoA, MMP-2, -9 expression, TIMP1 expression↑; MMP-2, -9 activity, NF-κB, PKC, GRB2, p-AKT, p-ERK, p-P38, p-JNK1/2, ROCK1, FAK, TIMP2↓ | Wu et al. [70] | 2015 | |
| In vitro | Growth inhibition (5–20 nmol/L, 72H); cellular proteasome activity inhibition; ubiquitinated proteins accumulation (A549) [BF211] | PSMB6 β1↓ | Sun et al. [92] | 2016 | |
| In vitro | Growth inhibition (1–2 nM, 48H); enhanced HDAC inhibitors induced apoptosis (A549) [BF] | SRC-3p-Akt, Bcl-2↓ | Zou et al. [155] | 2016 | |
| In vitro and in vivo | Growth inhibition (1–4 μM, 12–48H); cell morphological changes induction; DNA condensation; apoptosis induction (H460) [BF] | Cytochrome C, Apaf-1, active caspase-3, FasL/CD95, FasL, AIF, Endo G, caspase-9 activity, GADD153 mRNA expression, ROS production↑; pro-caspase-3, Bcl-2, GRP78 mRNA expression, ΔΨm↓ | Wu et al. [150] | 2017 | |
| In vitro and in vivo | Growth inhibition (12.5–50 nM, 1–12H); apoptosis induction (A549) [BF] | Cleaved-caspase-3, cleaved-PARP, Caspase-3 activity↑; p-Src↓ | Liu et al. [36] | 2016 | |
| In vitro | Inhibits gefitinib resistant cell migration and invasion (2.5–10 nM, 24–48H) (NCI-H460) [BF] | p-p38, p65↑; SOS-1, MMP-2, RhoA, N-Cadherin, E-Cadherin↓ | Huang et al. [74] | 2016 | |
| In vitro | Growth inhibition (25–100 nM, 6–48H); apoptosis induction (H1975) [BF] | Mcl-1 protein degradation, cleaved-PARP, cleaved-caspase-3, Bax, Bak↑; p-GSK-3β, Bcl-1, Bcl-2, Bcl-xL↓ | Kang et al. [171] | 2017 | |
| In vitro | Growth inhibition (1–100nmoL/L, 24–72H); apoptosis induction; S-phase arrest (A549) [BF] | Caspase-3↑ | Zhang et al. [172] | 2017 | |
| Melanoma | In vitro | Growth inhibition (150–550 nM, 24–48H); apoptosis induction (A375.S2) [BF] | ROS production, intracellular Ca2+ production, NO formation; cleaved-caspase-3, -8, -9, cytochrome c, AIF, Endo G, Bax, Fas, FasL, GRP78; ΔΨm, Bcl-xL↓ | Hsiao et al. [44] | 2012 |
| Myeloma | In vitro | Growth inhibition (20 nM, 48H); apoptosis induction; G2/M arrest (U929, U266) [BF] | Chemosensitivity↑; PARP1↓ | Huang et al. [173] | 2013 |
| In vitro and in vivo | Growth inhibition (12 nM, 12–48H); synergistic with MK2206 surpassed bortezomib resistance (U929, U266) [BF] | p-Akt, p-mTOR↑; p-P70, IL-6 secretion↓ | Xiang et al. [147] | 2017 | |
| Oral | In vitro | Growth inhibition (125 nM, 24H); G0/G1 phase arrest; apoptosis induction (CAL27)[BF] | Cytochrome c, Apaf-1, AIF, cleaved-caspase-3, -9↑; p-Akt, Cyclin D1, p-Bad↓ | Tsai et al. [139] | 2012 |
| In vitro | Growth inhibition (50–150 nM, 24–48H); apoptosis induction (CAL27) [BF] | Intracellular ROS accumulation, p-JNK, p-p38, p-c-Jun↑; hTERT expression ↓ | Tian et al. [140] | 2015 | |
| Osteosarcoma | In vitro | Growth inhibition (25–50 nM, 24–48H); anti-invasion and migration (U2OS) [BF] | SOS-1, JNK1/2, ERK1/2, p-38, MMP-7, -9 enzyme activity↓ | Chueh et al. [69] | 2011 |
| In vitro and in vivo | Growth inhibition (25 nM, 24H); apoptosis induction; G2/M arrest (U2OS, U2OS/MTX300) [BF] | Cleaved-PARP↑; Hsp27, p-Akt, P65↓ | Xie et al. [86] | 2012 | |
| In vitro | Inhibition of differentiation and proliferation (10 μM, 72Η) (hMG63-derived cancer stem cell) [BF] | CD133↓ | Chang et al. [174] | 2014 | |
| In vitro | Growth inhibition (10–40 nM, 24H) (MG-63) [BF] | ROS production, mitochondrial membrane hyperpolarization, Apaf-1, cleaved-PARP, cleaved-caspase-3, -7, -9↑; Bcl-2/Bax ratio↓ | Wang et al. [51] | 2014 | |
| In vitro | Inhibition of differentiation and proliferation (10 μΜ, 8 days) (Primary osteosarcoma stem cell C1OS) [BF] | Cleaved-caspase-3, miR-148a↑; ALDH1, hTERT, Nanog. CD133, Notch, Bmi-1↓ | Chang et al. [175] | 2015 | |
| In vitro | Growth inhibition (10–50 μg/L, 6–24H); apoptosis induction (U-2OS) [BF] | Cytosol/mitochondrial cytochrome c, cleaved-PARP, cleaved-caspase-3, -9, Bax↑; ΔΨm, PARP, Bcl-2↓ | Chen et al. [40] | 2016 | |
| In vitro | Growth inhibition (0.05–10 μM, 24H); apoptosis induction (U-2OS, Saos-2) [BF] | ROS production, BBC3↑; miR221↓ | Zhang et al. [141] | 2016 | |
| In vitro | Growth inhibition (200 nM, 6–48H); apoptosis induction (U-2OS) [BF] | Ca2+ release, caspase-3, -8, -9 activity, cytochrome c, Fas-L, cleaved-PARP, Calpain 1, ATF-6α, GRP-78, caspase-4↑; ΔΨm, Bcl-2, Bcl-xL↓ | Lee et al. [176] | 2017 | |
| Ovarian | In vitro | Growth inhibition (1 ng/mL, 48H); G0/G1 arrest; apoptosis induction [BF] (SK-OV-3, OMC-3) | p21, cleaved-caspase-9↑; Cyclin A, Cyclin D3, Bcl-2, Bcl-xL↓ | Takai et al. [132] | 2008 |
| In vitro | Growth inhibition (1–100 ng/mL, 48H); apoptosis induction (SKOV3, ES-2) [BF] | miR-183 downregulation enhanced bufalin-induced growth inhibition and apoptosis | Chen et al. [177] | 2016 | |
| Pancreatic | In vitro and in vivo | Growth inhibition (0.001–0.1 μM, 48H); apoptosis induction; enhance sensitivity to gemcitabine (Bx-PC3, MiaPaCa2, Panc-1) [BF] | ASK1, p-JNK, cleaved-caspase-3↑; Ki-67, Bcl-2↓ | Chen et al. [148] | 2012 |
| In vitro | Growth inhibition (50–200 nM,48H); G2/M arrest; apoptosis induction; enhanced gemcitabine chemosensitivity (Panc-1, CFPAC-1) [BF] | Bax, P21↑; Bcl-2, pro-caspase-3, -9, CyclinB1, CDK1↓ | Li et al. [47] | 2014 | |
| In vitro | Growth inhibition (50–150 nM, 24–48H); mitochondria-dependent apoptosis induction; (Capan-2)[BF] | p-JNK, p-p38, p-c-Jun, intracellular ROS accumulation↑; hTERT expression↓ | Tian et al. [140] | 2015 | |
| In vitro and in vivo | Cancer stem cells formation inhibition (50 nM, 24H) (MiaPaCa2/GEM) [BF] | CD24 expression, ESA expression, PTCH1, PTCH2, Gli1↓ | Wang et al. [82] | 2016 | |
| In vitro and in vivo | Growth inhibition (0.1–10 μM, 24H); S-phase arrest (SW1990, BxPc3) [BF] | c-Myc, NF-κB↓ | Liu et al. [149] | 2016 | |
| Prostate | In vitro | Growth inhibition (0.1–10 μM, 24H); apoptosis induction (LNCaP, PC3, DU145) [BF] | Caspase-3 activity, caspase-9 activity, intracellular Ca2+↑ | Yeh et al. [178] | 2003 |
| In vitro | Growth inhibition (15 μM, 24H); apoptosis induction (PC3) [BF] | miR-181a induction enhanced bufalin-induced growth inhibition and apoptosis; miR-181a expression, caspase-3 activity↑; Bcl-2↓ | Zhai et al. [179]. | 2013 | |
| Renal | In vitro | Induce new high density glycogen-microtubule structures formation (1–20 nM, 0–4.5H) (ACHN) [BF] | K−ATPase-induced ERK1/2 phosphorylation↑ | Fridman E et al. [180] | 2012 |
| Tongue | In vitro | Growth inhibition (100–500 nM, 48H); G2/M arrest; mitochondria-dependent apoptosis (SCC-4) [BF] | Ca2+, NO production, DR5 expression, caspase-9 expression↑; Bcl-2, Bid, calpain 1, ATF-6β, ΔΨm, ROS production↓ | Chou et al. [143] | 2017 |
Fig. 3.

A schematic summary of the molecular mechanisms of Bufalin-mediated anti-cancer effect
Induce cell death other than apoptosis
In contrast to the apoptosis-inducing activity of Bufalin, well-documented in various cancer cell lines, 100 nM of Bufalin does not induce caspase-dependent apoptosis in human colorectal caco-2 and HT-22 cell lines [59]. Instead, Bufalin induces cell death by triggering autophagy, possibly through a ROS- and JNK-dependent pathways in colorectal cancer cells, followed by increased expression of ATG5 and Beclin-1 without significant induction of apoptosis, PARP cleavage, and caspase-3 cleavage [59]. It may be plausible that Bufalin enhances the radiosensitivity of colorectal cancer through ROS-mediated autophagy, which deserves further investigation [59].
In addition to inducing apoptosis-dependent cell death, laboratory studies have suggested that Bufalin (0.1–1 μM) inhibits Na+/K+-ATPase activity, a crucial ion pump, and transducer ligand receptor. It suppresses cancer cell proliferation and possesses synergistic effect with Sorafenib against liver cancer [60, 61]. Changes in Na+/K+-ATPase activity play an important role in cell survival and function, and high expression of Na+/K+-ATPase subunit α is associated with poor overall survival in liver cancer patients [27]. Moreover, compared with adjacent tissues, the expression of Na+/K+-ATPase subunit α1 was significantly elevated in liver cancer tissues, suggesting that the specific targeting of Na+/K+-ATPase can have therapeutic effects on cancer cells without noticeable effect on normal cells [27]. The role of Na+/K+-ATPase in mediating cytotoxicity of Bufalin should be highlighted, and further studies are warranted.
Metastasis inhibition (angiogenesis, MMPs, EMT, others)
Angiogenesis
Hyperactive angiogenesis is a hallmark of cancer cells. It not only provides oxygen and extra nutrient, maintains the high proliferation rate of cancer cells, but also promotes local and distant metastasis of malignant cells [62]. Bufalin had been reported to inhibit cancer cell migration and invasion in liver cancer and lung cancer cell lines by down-regulating vascular endothelial growth factor (VEGF), which plays a major role in tumor angiogenesis [46, 63, 64]. Anti-angiogenic drugs, such as Sorafenib, may be complicated by relatively easy-acquired drug resistance, rapid onset of relapse after discontinuation, and the potential for tumor metastasis. A previous study revealed that Bufalin enhances the anti-angiogenic effect of Sorafenib via AKT/VEGF signaling [64]. The co-administration of Bufalin and Sorafenib provides a novel therapeutic option for patients with advanced HCC and is worthy for further in-depth investigations [63, 64].
MMPs
Matrix metalloproteinases (MMPs) are a class of enzymes that degrade extracellular matrix proteins and are thought to play important roles in cell proliferation, migration, angiogenesis, and etc. In cancer cells, especially those with high metastatic potentials, MMPs, in particular, MMP-2 and MMP-9, are highly expressed [65]. MMP-2 and MMP-9 primarily control cancer cell motility, and down-regulation of these proteins was reported across studies to reduce cancer metastasis [66, 67]. Bufalin was found to down-regulate MMPs in human liver cancer, lung cancer, osteosarcoma, and etc. [68–70]. In human HCC, Bufalin inhibits metastasis both in vitro and in vivo through pathways including the Akt/GSK3β signaling, E-cadherin/β-catenin nuclear translocation [66, 71–73]. In lung cancer cells, Bufalin (25–10 nM) was observed to suppress MMP-2 and MMP-9 expression, which were mediated through multiple pathways, including p38 MAPK, c-Jun N-terminal kinase (JNK), extracellular signal-regulated kinase1/2 (Erk1/2), focal adhesion kinase (FAK), Rho-associated protein kinase 1 (ROCK1) and NF-κB [69, 70, 74]. Bufalin is a broad inhibitor of various inflammatory signaling pathways in cancer. It was noticed that in human hepatoma cells, the expression of the inflammatory protein COX-2 is down-regulated and is associated with the inhibition of cell invasion and migration [66].
EMT
The epithelial–mesenchymal transition (EMT) is a process in which epithelial cells malignant phenotypes to become mesenchymal stem cells that promote metastasis. Bufalin was found to inhibit EMT in cervical, colorectal, hepatic, lung cancer by downregulating mesenchymal markers and upregulating epithelial markers [66, 67, 71, 75–80]. Bufalin can attenuate transforming growth factor-β (TGF-β)-induced upregulation of N-cadherin, vimentin, and Snail while downregulating E-cadherin by targeting hypoxia-inducible factor-1α (HIF-1α) [76]. In addition, the regulating of motility and invasiveness of cancer cells may be related to the PI3K/AKT/mTOR pathway [76, 81], Wnt/β-catenin pathway [66, 77], Hedgehog signaling pathway [67, 82], and etc.
Others
Bufalin is reported to act as a natural anti-inflammatory small molecule that upregulates osteoprotegerin and down-regulates receptor activator of NF-κB ligand (RANKL), ameliorating cannabinoid 2 receptor (CB2)-mediated cancer-induced pain and bone destruction [83]. Bufalin also down-regulated several metastasis-related genes, such as Rho-associated (Rho A), integrins and FAK [70, 80, 84, 85]. A recent study revealed that bufalin combined with sorafenib synergistically inhibited liver cancer cell migration by targeting mTOR/Akt/VEGF signaling and affecting tumor vascular microenvironment [63, 64].
Omics approach in the study of antineoplastic effects of Bufalin and Huachansu
Few genomics and proteomics approaches are used to study the anti-cancer effects of Bufalin or HCS. Using a comparative proteomics approach, Xie et al. [86, 87] identified that Bufalin modifies 24 differentially expressed protein, particularly, the expression of proteins involve in cell metabolism, apoptosis, and cytoskeleton structure. Among them, the heat shock protein 27 (Hsp27) decreased remarkably, and its down-regulation played a critical role in bufalin-induced apoptosis in osteosarcoma cells [86, 87]. Another study used two quantitative proteomics methods, isobaric tags for relative and absolute quantification (iTRAQ)-based and label-free proteomic analysis, to study the target-related proteins of Bufalin in human A549 lung cancer cells [88]. The number of the differentially expressed protein commonly found in the two methods is 45 proteins, suggesting that the involvement of oxidative stress and the fibronectin-related pathways are important pathways for the anti-cancer effect of Bufalin [88]. Wu et al. [89] reported in 2014 that Bufalin modulates about 165-apoptosis-related genes in human lung cancer CNI-H460 cells using Affymetrix GeneChip. These results provide a deeper understanding of the anti-proliferative and cytotoxic mechanisms of Bufalin at the genetic level in gene assays [89]. Although a small number of proteomics studies and gene chip arrays have studied the target-related proteins or genes involved in the anti-cancer effect of Bufalin, which can serve as a paradigm for further studies of the molecular basis of Bufalin against various types of tumors, there is a lack of transcriptome analysis and other high-throughput screening in examining or predicting cell response to Bufalin treatment at both cytotoxic level and non-cytotoxic level. There is a need for high-throughput analysis to better associate the change of mRNA levels correlated with protein–protein interactions or protein–DNA interactions [90]. Further work is warranted on the molecular mechanism of Bufalin and HCS against various tumors.
Drug delivery and its derivatives
Bufalin participates in complex cell-signal transduction pathways, which contribute to its suppressive effect of tumor progression in various cancer types. However, its structural similarity to digitoxin, digoxin and other cardiac glycosides accounts for the toxic effects at a high dosage. Previous studies suggest that Na+/K+-ATPase is a potential drug target that contributes to the emerging role of cardiac glycosides in selectively controlling tumor proliferation, but does not affect normal cell growth [36]. In the recent years, emerging newly discovered HCS derivatives have been shown to process anti-tumor effects, such as BF211, have been shown to inhibit colorectal, gastric, and lung cancer [91, 92].
In the process of seeking to improve the anti-cancer properties of Bufalin, several drug carriers have been synthesized with Bufalin, such as carbon-based nanomaterials, including liposomes and polymeric microspheres, folate receptor-targeted supramolecules, or [18F]fluoroethyl conjugates [93–101]. Structurally modified and synthetic Bufalin-loaded nanoparticles are designed to promote tumor-specific drug release and cytotoxicity, increase cellular uptake, and improve bio-distribution at the tumor site [99, 102, 103]. Some studies have reported that internalization of the Bufalin nanoparticles may at least partially contribute to the increased intracellular uptake of Bufalin in cancer cells [85, 93, 95, 102, 104–106]. However, there is still a need for better therapeutic materials with good drug solubility, well binding affinity, high tumor-specific targeting, low systemic toxicity and rapid clearance.
Potential cardiotoxic property of Bufalin and the side-effects of toad extract and HCS in clinical studies
As a cardioactive steroid, Bufalin has a variety of biological activities, such as cardiotonic, blood pressure stimulation, and etc. At high dosages, cardioactive steroids can cause cardiac arrhythmias and exhibit cardiotoxicity. The concentration of CS (400 ng/mL) extract and Bufalin alone can induce myocardial cell arrest within a few seconds after administration by altering intracellular calcium storage in cardiomyocytes and possibly acts on sites other than the Na+/K+-ATPase [107]. Koh et al. [108] reported that zebrafish larvae responded to 100 µM Bufalin and showed a decrease in heart rates, early depolarization and polymorphic arrhythmia-like changes. Another study found that the addition of Bufalin to guinea pig papillary muscle (0.4 µmol/L) or atrium (225 nmol/L) preparations can lead to arrhythmias [109]. Further study on the effect of ethanol extract of Chinese toad venom (EET) on mice showed that 5 mg/kg of EET caused liver toxicity cardiomyocytes injuries [110–112]. In a Pilot study of HCS in patients with HCC, non-small cell lung cancer, or pancreatic cancer showed that side effects that may have been related to HCS were hematologic (thrombocytopenia and leukopenia), gastrointestinal (loss of appetite, constipation and diarrhea), mucocutaneous (dental ulcers and rashes), and cardiovascular (premature ventricular contraction and hypertension) in nature. Others include myalgia, dyspnea and dizziness [12]. However, when using HCS, no dose-limiting toxicity (DLT) was observed at doses up to 8-times higher than the usual dose used in China (conventional doses = 20–25 mL; highest dose = 162 mL). To the best of our knowledge, the typical therapeutic doses of HCS have not found significant side-effects in clinical studies. However, it has been reported that HCS can reduce the side-effects of chemotherapy and radiotherapy in gastric cancer, lung cancer, colorectal cancer, and etc. [9, 11, 113–116]. Due to the potential toxic effects of other cardiac glycosides, careful clinical evaluation should be performed prior to Bufalin administration.
Clinical trials
Huachansu injection is a sterile hot-aqueous extract of dried toad skin, is approved by the Chinese FDA for use at oncology clinics in China. Since the 1970s, various clinical studies in China have demonstrated the anti-cancer properties of HCS, with a total response rate of 10% and 16% for patients with advanced liver and lung cancer, respectively. In a previous clinical study, the quality and consistency of three separate lots of HCS were evaluated [12]. It was found that and levels of bufalin and resibufogenin in HCS were 18.0–19.5 ng/mL, and 17.7–19.0 ng/mL, respectively [12]. The variation between three lots of HCS was remarkably close, with less than 10% variation. In the detection of bufadienolides concentration after i.v. infusion of HCS in human plasma specimens, the levels of Bufalin, cinobufagin, cinobufotalin, and resibufogenin in the used in a phase I clinical trial was 14.3 ± 0.03, 3.35 ± 0.1, 21.5 ± 0.22, and 24.5 ± 2.18 ng/mL, respectively [12]. As for Bufalin, which studies have shown is the major bufadienolide with the most pronounced anti-cancer activity, was further evaluated its concentration in plasma by liquid chromatography with mass spectrometry (LC/MS/MS) and found a maximal plasma levels at the end of the 2-h infusion which is proportional to the amount of drug administered (0.81–3.38 ng/mL) [12]. In a clinical phase I study of HCS in patients with HCC, non-small cell lung cancer, or pancreatic cancer published in 2009 and found that HCS is partially effective on cancer patients [12]. Only mild adverse events were observed, with doses up to five times the conventional clinical dose, which is approximately 15 mL of drug per meter squared of body mass (mL/m2) [12]. Furthermore, a phase II randomized controlled trial (RCT) of patients with advanced pancreatic adenocarcinoma (PaCa) treated with HCS in 2012 reported no clinical benefit with the addition of HCS to gemcitabine, which is similar to over 30 previously published well-designed RCTs in patients with advanced PaCa evaluating gemcitabine in combination with other cytotoxic or biologic agents [117]. However, the lack of efficacy observed in these trials does not preclude the possible efficacy of HCS in other solid malignancies. There is an increase in related publications later (Fig. 1a), indicating an increasing research interest on the potential use of HCS and its derived steroidal cardiac glycosides in cancer therapy. However, the report on the outcome of any RCT published after the phase II clinical trial is not yet available.
Discussion and future perspectives
At present, TCM is practiced all over the world. Natural products have long been an important source of cancer treatment. CS has been used for thousands of years in the practice of TCM in aqueous extract form for the treatment of cancer. HCS has been used either alone or in combination with chemotherapeutic agents. Since the phase I study of HCS in patients with liver, lung, or pancreatic cancer in 2009, there is an increase in related publications, indicating a great interest is brought on the potential use of HCS and Bufalin in human. HCS and Bufalin exhibit a wide range of biological effects in cancer, including inhibition of cell proliferation, induction of cell apoptosis, disruption of the cell cycle, inhibition of metastasis, reversing multi-drug resistance to chemotherapeutic agents, and regulation of the immune response. HCS and Bufalin also have multiple other effects such as respiratory excitation, anti-inflammation and analgesics [83, 118].
Cancer is characterized by inflammation, and the chronic inflammatory microenvironment is often associated with malignant progression. The anti-inflammatory effects of Bufalin may at least in part contribute to its anti-cancer effect targeting the tumor microenvironment. The involvement of various inflammatory mediators such as COX-2 and PGE2 may be produced not only by cancer cells, but also by the surrounding immune cells, fibroblasts, and endothelial cells. The production of inflammatory mediators may create a positive feedback loop to further promote cancer proliferation, angiogenesis, invasion, and metastasis. Although most studies reported the anti-inflammatory effect of Bufalin, in breast cancer, Bufalin was reported to promote inflammatory response, accompanied by increased COX-2/IL-8 expression and enhanced tumor growth in vivo [38]. The role of Bufalin on NF-κB signaling and pro-inflammatory mediators, such as COX-2, inducible nitric oxide synthase (iNOS), tumor necrosis factor alpha (TNF-α), interleukin 1 beta (IL-1β), and interleukin 6 (IL-6) are studied across diseases, including rheumatoid arthritis [119, 120], asthma [118], bacterial infection [121], and chronic inflammatory disease [122]. It is worth highlighting that Bufalin may also possess chemo-preventive potential in skin cancer when administered topically in a rodent model [123]. Future research may extend to other types of human diseases. Research on the interaction between stromal cells and tumor cells may not only broaden the potential use of Bufalin and HCS, but also helps to understand the tumor immune microenvironment, thereby providing long-lasting repression against cancer growth for future cancer treatment.
Previous studies found that the expression of Na+/K+-ATPase subunit α1 is prognostic in liver cancer patients and is significantly elevated in cancer tissue compared with adjacent tissues [27, 60]. Recent studies of cardiac glycosides suggest Bufalin may potentially represent a promising form of targeted cancer chemotherapy on Na+/K+-ATPase subunit α1 that can be safely used for long periods without severe side effects [27, 60]. Furthermore, the results indicated that the down-regulation of Na+/K+-ATPase α1 can inhibit cell proliferation, migration, and invasion in vitro, and inhibit tumorigenesis in vivo [26, 27, 124]. These increase the likelihood of selecting patients with high Na+/K+-ATPase subunit α1, may improve the therapeutic efficacy of Bufalin and HCS. It is worth noting that since June 2015, another Na+/K+-ATPase inhibitor, RX108, is structurally similar to Bufalin and is undergoing phase I clinical trials to evaluate the safety, tolerability, pharmacokinetics, and pharmacodynamic properties in patients with locally advanced or metastatic solid tumors in Australia. However, as of July 2018, there were no recent reports for phase-I development. This study, if successfully completed, may provide valuable information about the safety of targeting Na+/K+-ATPase.
In the process of identifying active anti-cancer ingredients in Traditional Chinese Medicine, some encouraging results have been achieved and we deepened our understanding of the underlying mechanism of malignant behavior. With the growing awareness of the tumor microenvironment and the wholistic immunomodulation, it is possible that agents with systemic effects may be a double-edged sword. In the identification of novel therapeutic agents, agents that target multiple signaling pathways may have a stronger anti-cancer effect, but inevitably carry more adverse effects, such as gastrointestinal burden and hematological toxicities. Studies combining conventional cancer treatments (such as chemotherapy and radiotherapy) with TCM or derived components should be interpreted with caution, as drug interactions caused by combination therapies may have a negative impact on conventional cancer treatments, reducing their effectiveness. It is also essential to further study the underlying pathophysiological mechanisms of anti-cancer effects and adverse reactions. Inevitably, there is a great need to discover long-term effects, drug use sequences, drug–drug interactions, and individual choices further for different drugs, aimed at improving patients’ quality of life and prolonging survival.
Concluding remarks
Vigorous studies and discoveries have revealed the anti-tumor properties of Bufalin and HCS as potentially multi-targeted agents for cancer treatment. The results indicated that Bufalin inhibits tumor progression by inhibiting cancer cell proliferation via both apoptosis dependent or independent pathways, as well as inhibiting metastasis via repressing cell motility and angiogenesis. Many animal studies and clinical studies conducted over the past decade have revealed their clinical potentials, however, in the current study, long-term toxicological studies on different individuals, pharmacodynamics studies on various dosages and administration routes have been relatively lacking. Further research is needed to elucidate potential drug-drug interactions and multi-target interactions of Bufalin and HCS. Large-scale clinical trials are warranted to translate the knowledge for anti-cancer effects of Bufalin and HCS into clinical applications as effective and safe treatment options for future cancer patients. Overall, this review highlights the advances in the study of Bufalin and HCS as emerging anticancer agents over the past decade and may shed light on the future direction of Bufalin and HCS as novel anticancer agents for clinical applications.
Highlights
Huachansu and Bufalin possess anti-cancer effects both in vitro and in vivo.
The multi-target and multi-pathway pharmacological actions are promising.
Potential drug–drug interactions and multi-target interaction lacks studies.
Further large-scale clinical trials are warranted.
Authors’ contributions
Conceived and designed the study: C-SC, JW, JC, LC, ZC, ZM. Collected the literature, wrote the manuscript: C-SC, JW, JC, K-TK, TJ, HG, LC. Collected and drew figures: C-SC, JW, JC. Revised the manuscript: C-SC, JW, JC, TJ, HG, ZC, ZM. All authors read and approved the final manuscript.
Acknowledgements
The authors deeply appreciate Dr. Xiaoyan Zhu, Dr. Yawen Geng, and Dr. Ye Li for their advices and constructive comments for this review paper in preparation.
Competing interests
The authors declare that they have no competing interests.
Availability of data and materials
Not applicable.
Consent for publication
Not applicable.
Ethics approval and consent to participate
Not applicable.
Funding
The work was supported by National Natural Science Foundation of China (NSFC) [Grant No. 81573753].
Publisher’s Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Abbreviations
- TCM
Traditional Chinese Medicine
- CS
Chansu
- HCS
Huachansu
- NF-B
nuclear Factor-κB
- PGE2
prostaglandin E2
- Na+, K+-ATPase
Na+/K+ pump or sodium- and potassium-activated adenosine 5′-triphosphatase
- HPLC
high-pressure liquid chromatography
- IC50
the 50% inhibition dose
- ER
estrogen receptor
- i.p.
intraperitoneal
- i.v.
intravenous
- COX-2
cyclooxygenase-2
- IL-8
interleukin-8
- MAPK
mitogen-activated protein kinase
- HCC
hepatocellular carcinoma
- ROS
reactive oxygen species
- SRC
steroid receptor coactivator
- TRAIL
tumor necrosis factor related apoptosis-inducing ligand
- JNK
c-Jun N-terminal kinase
- ATG5
autophagy-related gene 5
- PARP
poly-(ADP-ribose) polymerase
- MMPs
matrix metalloproteinases
- EMT
epithelial–mesenchymal transition
- VEGF
vascular endothelial growth factor
- AKT
protein kinase B
- GSK3β
glycogen synthase kinase 3 beta
- Erk1/2
extracellular signal-regulated kinase1/2
- FAK
focal adhesion kinase
- ROCK1
Rho-associated protein kinase 1
- TGF-β
transforming growth factor-β
- HIF-1α
hypoxia-inducible factor-1α
- PI3K
phosphoinositide 3-kinase
- mTOR
mammalian target of rapamycin
- RANKL
receptor activator of nuclear factor-κ B ligand
- CB2
cannabinoid 2 receptor
- Rho A
Rho-associated
- Hsp27
heat shock protein 27
- iTRAQ
isobaric tags for relative and absolute quantification
- EET
ethanol extract of Chinese toad venom
- DLT
dose-limiting toxicity
- LC/MS
liquid chromatography with mass spectrometry
- RCT
randomized controlled trial
- iNOS
inducible nitric oxide synthase
- TNF-α
tumor necrosis factor alpha
- IL-6
interleukin 6
- PaCa
pancreatic adenocarcinoma
- RCT
randomized controlled trial
Contributor Information
Chien-shan Cheng, Email: natcheng@qq.com.
Jiaqiang Wang, Email: jiaqiangxiong@126.com.
Jie Chen, Email: jiechen0128@connect.hku.hk.
Kuei Ting Kuo, Email: alice20051026@gmail.com.
Jian Tang, Email: 18211230034@fudan.edu.cn.
Huifeng Gao, Email: huifeng_gao@hotmail.com.
Lianyu Chen, Phone: (86)21-64175590, Email: lianyu_chen@hotmail.com.
Zhen Chen, Phone: (86)21-64175590, Email: cz120@mail.sh.cn.
Zhiqiang Meng, Phone: (86)21-64175590, Email: mengzhq@yeah.net.
References
- 1.Cho WC. Evidence-based anticancer materia medica. Hong Kong SAR: Springer; 2011. pp. 390–406. [Google Scholar]
- 2.Huang K. Anesthetic and muscle-relaxing herbs: Chan Su. In: The pharmacology of Chinese herbs. 2nd edn. Boca Raton: CRC Press; 1999: 544.
- 3.Xu R, Xie HQ, Deng LL, Zhang JX, Yang FM, Liu JH, Hao XJ, Zhang YH. A new bufadienolide with cytotoxic activity from the Chinese traditional drug Ch’an Su. Chin J Nat Med. 2014;12(8):623–627. doi: 10.1016/S1875-5364(14)60095-9. [DOI] [PubMed] [Google Scholar]
- 4.Sakurai K, Yoshii E, Hashimoto H, Kubo K. Gas liquid chromatography of steroids of Ch’an Su. II. Reinvestigation on the determination of bufadienolides. Chem Pharm Bull. 1968;16(6):1140–1143. doi: 10.1248/cpb.16.1140. [DOI] [PubMed] [Google Scholar]
- 5.Shimizu Y, Morishita S. Metabolism and disposition of kyushin, a drug containing senso (ch’an su) Am J Chin Med. 1996;24(3–4):289–303. doi: 10.1142/S0192415X96000359. [DOI] [PubMed] [Google Scholar]
- 6.Chang ZF, Jia DX, Bare J. Chinese materia medica. Beijing: Peoples Medical Publishing House; 2014. [Google Scholar]
- 7.Su YH, Huang XQ, Zhang DZ, Zhang YN, Xie JM, Linh CQ. HPLC separation and determination of bufadienolide in cinobufacini injection. Chin Tradit Pat Med. 2003;25:24–27. [Google Scholar]
- 8.Qin TJ, Zhao XH, Yun J, Zhang LX, Ruan ZP, Pan BR. Efficacy and safety of gemcitabine-oxaliplatin combined with Huachansu in patients with advanced gallbladder carcinoma. World J Gastroenterol. 2008;14(33):5210–5216. doi: 10.3748/wjg.14.5210. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 9.Xie X, Huang X, Li J, Lv X, Huang J, Tang S, Sun Y. Efficacy and safety of Huachansu combined with chemotherapy in advanced gastric cancer: a meta-analysis. Med Hypotheses. 2013;81(2):243–250. doi: 10.1016/j.mehy.2013.04.038. [DOI] [PubMed] [Google Scholar]
- 10.Yin JH, Zhu XY, Shi WD, Liu LM. Huachansu injection inhibits metastasis of pancreatic cancer in mice model of human tumor xenograft. BMC Complement Altern Med. 2014;14:483. doi: 10.1186/1472-6882-14-483. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 11.Zhou B, Wu F, Yuan L, Miao Z, Zhu S. Is huachansu beneficial in treating advanced non-small-cell lung cancer? Evidence from a meta-analysis of its efficacy combined with chemotherapy. Evid Based Complement Alternat Med. 2015;2015:408145. doi: 10.1155/2015/408145. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 12.Meng Z, Yang P, Shen Y, Bei W, Zhang Y, Ge Y, Newman RA, Cohen L, Liu L, Thornton B, et al. Pilot study of huachansu in patients with hepatocellular carcinoma, nonsmall-cell lung cancer, or pancreatic cancer. Cancer. 2009;115(22):5309–5318. doi: 10.1002/cncr.24602. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 13.Wang L, Raju U, Milas L, Molkentine D, Zhang Z, Yang P, Cohen L, Meng Z, Liao Z. Huachansu, containing cardiac glycosides, enhances radiosensitivity of human lung cancer cells. Anticancer Res. 2011;31(6):2141–2148. [PubMed] [Google Scholar]
- 14.Lee HJ, Koung FP, Kwon KR, Kang DI, Cohen L, Yang PY, Yoo HS. Comparative analysis of the bufonis venenum by using TLC, HPLC, and LC–MS for different extraction methods. J Pharmacopunct. 2012;15(4):52–65. doi: 10.3831/KPI.2012.15.012. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 15.Dai LP, Wang ZM, Gao HM, Jiang X, Ding GZ. Determination of bufothionine in skin of Bufo bufo gargarizans and Huachansu injection. Zhongguo Zhong Yao Za Zhi. 2007;32(3):224–226. [PubMed] [Google Scholar]
- 16.Yang T, Shi R, Chang L, Tang K, Chen K, Yu G, Tian Y, Guo Y, He W, Song X, et al. Huachansu suppresses human bladder cancer cell growth through the Fas/Fasl and TNF-alpha/TNFR1 pathway in vitro and in vivo. J Exp Clin Cancer Res. 2015;34:21. doi: 10.1186/s13046-015-0134-9. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 17.Toma S, Hirai Y, Sugimoto C, Shoji M, Oguni Y, Morishita S, Ito C, Horie M. Metabolic fate of bufalin and cinobufagin. Yakugaku Zasshi. 1991;111(11):687–694. doi: 10.1248/yakushi1947.111.11_687. [DOI] [PubMed] [Google Scholar]
- 18.Yang Z, Luo H, Wang H, Hou H. Preparative isolation of bufalin and cinobufagin from Chinese traditional medicine ChanSu. J Chromatogr Sci. 2008;46(1):81–85. doi: 10.1093/chromsci/46.1.81. [DOI] [PubMed] [Google Scholar]
- 19.Su YH, Nu X. Evaluation of pharmacodynamic effect of pharmaceutical agents of Chan Su. J Beijing Univ of TCM. 2001;24:51–54. [Google Scholar]
- 20.Calderon-Montano JM, Burgos-Moron E, Orta ML, Maldonado-Navas D, Garcia-Dominguez I, Lopez-Lazaro M. Evaluating the cancer therapeutic potential of cardiac glycosides. Biomed Res Int. 2014;2014:794930. doi: 10.1155/2014/794930. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 21.Haux J. Digitoxin is a potential anticancer agent for several types of cancer. Med Hypotheses. 1999;53(6):543–548. doi: 10.1054/mehy.1999.0985. [DOI] [PubMed] [Google Scholar]
- 22.Kamano Y, Nogawa T, Yamashita A, Hayashi M, Inoue M, Drasar P, Pettit GR. Isolation and structure of a 20,21-epoxybufenolide series from “Ch’an Su”. J Nat Prod. 2002;65(7):1001–1005. doi: 10.1021/np0200360. [DOI] [PubMed] [Google Scholar]
- 23.Takai N, Kira N, Ishii T, Yoshida T, Nishida M, Nishida Y, Nasu K, Narahara H. Bufalin, a traditional oriental medicine, induces apoptosis in human cancer cells. Asian Pac J Cancer Prev. 2012;13(1):399–402. doi: 10.7314/APJCP.2012.13.1.399. [DOI] [PubMed] [Google Scholar]
- 24.Datta P, Dasgupta A. Interactions between drugs and Asian medicine: displacement of digitoxin from protein binding site by bufalin, the constituent of Chinese medicines Chan Su and Lu-Shen-Wan. Ther Drug Monit. 2000;22(2):155–159. doi: 10.1097/00007691-200004000-00002. [DOI] [PubMed] [Google Scholar]
- 25.Dasgupta A, Emerson L. Neutralization of cardiac toxins oleandrin, oleandrigenin, bufalin, and cinobufotalin by digibind: monitoring the effect by measuring free digitoxin concentrations. Life Sci. 1998;63(9):781–788. doi: 10.1016/S0024-3205(98)00333-6. [DOI] [PubMed] [Google Scholar]
- 26.Huang H, Zhang W. Bufalin induced apoptosis of bladder carcinoma cells through the inactivation of Na+K+-ATPase. Oncol Lett. 2018;16(3):3826–3832. doi: 10.3892/ol.2018.9142. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 27.Li H, Wang P, Gao Y, Zhu X, Liu L, Cohen L, Meng Z, Yang P. Na+/K+-ATPase alpha3 mediates sensitivity of hepatocellular carcinoma cells to bufalin. Oncol Rep. 2011;25(3):825–830. doi: 10.3892/or.2010.1120. [DOI] [PubMed] [Google Scholar]
- 28.Nasu K, Nishida M, Ueda T, Takai N, Bing S, Narahara H, Miyakawa I. Bufalin induces apoptosis and the G0/G1 cell cycle arrest of endometriotic stromal cells: a promising agent for the treatment of endometriosis. Mol Hum Reprod. 2005;11(11):817–823. doi: 10.1093/molehr/gah249. [DOI] [PubMed] [Google Scholar]
- 29.Huang WW, Yang JS, Pai SJ, Wu PP, Chang SJ, Chueh FS, Fan MJ, Chiou SM, Kuo HM, Yeh CC, et al. Bufalin induces G0/G1 phase arrest through inhibiting the levels of cyclin D, cyclin E, CDK2 and CDK4, and triggers apoptosis via mitochondrial signaling pathway in T24 human bladder cancer cells. Mutat Res. 2012;732(1–2):26–33. doi: 10.1016/j.mrfmmm.2011.09.010. [DOI] [PubMed] [Google Scholar]
- 30.Hsu CM, Tsai Y, Wan L, Tsai FJ. Bufalin induces G2/M phase arrest and triggers autophagy via the TNF, JNK, BECN-1 and ATG8 pathway in human hepatoma cells. Int J Oncol. 2013;43(1):338–348. doi: 10.3892/ijo.2013.1942. [DOI] [PubMed] [Google Scholar]
- 31.Clifford RJ, Kaplan JH. Human breast tumor cells are more resistant to cardiac glycoside toxicity than non-tumorigenic breast cells. PLoS ONE. 2013;8(12):e84306. doi: 10.1371/journal.pone.0084306. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 32.Laursen M, Gregersen JL, Yatime L, Nissen P, Fedosova NU. Structures and characterization of digoxin- and bufalin-bound Na+, K+-ATPase compared with the ouabain-bound complex. Proc Natl Acad Sci USA. 2015;112(6):1755–1760. doi: 10.1073/pnas.1422997112. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 33.Lee S, Lee Y, Choi YJ, Han KS, Chung HW. Cyto-/genotoxic effects of the ethanol extract of Chan Su, a traditional Chinese medicine, in human cancer cell lines. J Ethnopharmacol. 2014;152(2):372–376. doi: 10.1016/j.jep.2014.01.023. [DOI] [PubMed] [Google Scholar]
- 34.Okada M, Suga T, et al. Pharmacology of the principles isolated from Senso (Ch’an Su) the dried venom of the Chinese toad (IV) Asian Med J. 1960;3(4):155–160. [Google Scholar]
- 35.Wang FZ, Gong H. Pharmacological study and clinical use of ChanSu. Forum Tradit Chin Med. 2003;18:50–52. [Google Scholar]
- 36.Liu M, Feng LX, Sun P, Liu W, Wu WY, Jiang BH, Yang M, Hu LH, Guo DA, Liu X. A novel bufalin derivative exhibited stronger apoptosis-inducing effect than bufalin in A549 lung cancer cells and lower acute toxicity in mice. PLoS ONE. 2016;11(7):e0159789. doi: 10.1371/journal.pone.0159789. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 37.Chan WY, Ng TB, Yeung HW. Examination for toxicity of a Chinese drug, the toad glandular secretory product Chan su, in pregnant mice and embryos. Biol Neonate. 1995;67(5):376–380. doi: 10.1159/000244188. [DOI] [PubMed] [Google Scholar]
- 38.Chen HT, Sun D, Peng YC, Kao PH, Wu YL. Novel augmentation by bufalin of protein kinase C-induced cyclooxygenase-2 and IL-8 production in human breast cancer cells. Innate Immun. 2017;23(1):54–66. doi: 10.1177/1753425916676347. [DOI] [PubMed] [Google Scholar]
- 39.Han KQ, Huang G, Gu W, Su YH, Huang XQ, Ling CQ. Anti-tumor activities and apoptosis-regulated mechanisms of bufalin on the orthotopic transplantation tumor model of human hepatocellular carcinoma in nude mice. World J Gastroenterol. 2007;13(24):3374–3379. doi: 10.3748/wjg.v13.i24.3374. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 40.Chen Y, Li M, Li Z, Gao P, Zhou X, Zhang J. Bufalin induces apoptosis in the U2OS human osteosarcoma cell line via triggering the mitochondrial pathway. Mol Med Rep. 2016;13(1):817–822. doi: 10.3892/mmr.2015.4583. [DOI] [PubMed] [Google Scholar]
- 41.Ding DW, Zhang YH, Huang XE, An Q, Zhang X. Bufalin induces mitochondrial pathway-mediated apoptosis in lung adenocarcinoma cells. Asian Pac J Cancer Prev. 2014;15(23):10495–10500. doi: 10.7314/APJCP.2014.15.23.10495. [DOI] [PubMed] [Google Scholar]
- 42.Efuet ET, Ding XP, Cartwright C, Pan Y, Cohen L, Yang P. Huachansu mediates cell death in non-Hodgkin’s lymphoma by induction of caspase-3 and inhibition of MAP kinase. Int J Oncol. 2015;47(2):592–600. doi: 10.3892/ijo.2015.3044. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 43.Hong SH, Choi YH. Bufalin induces apoptosis through activation of both the intrinsic and extrinsic pathways in human bladder cancer cells. Oncol Rep. 2012;27(1):114–120. doi: 10.3892/or.2011.1451. [DOI] [PubMed] [Google Scholar]
- 44.Hsiao YP, Yu CS, Yu CC, Yang JS, Chiang JH, Lu CC, Huang HY, Tang NY, Yang JH, Huang AC, et al. Triggering apoptotic death of human malignant melanoma a375.s2 cells by bufalin: involvement of caspase cascade-dependent and independent mitochondrial signaling pathways. Evid Based Complement Alternat Med. 2012;2012:591241. doi: 10.1155/2012/591241. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 45.Jiang L, Zhao MN, Liu TY, Wu XS, Weng H, Ding Q, Shu YJ, Bao RF, Li ML, Mu JS, et al. Bufalin induces cell cycle arrest and apoptosis in gallbladder carcinoma cells. Tumour Biol. 2014;35(11):10931–10941. doi: 10.1007/s13277-014-1911-3. [DOI] [PubMed] [Google Scholar]
- 46.Jiang Y, Zhang Y, Luan J, Duan H, Zhang F, Yagasaki K, Zhang G. Effects of bufalin on the proliferation of human lung cancer cells and its molecular mechanisms of action. Cytotechnology. 2010;62(6):573–583. doi: 10.1007/s10616-010-9310-0. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 47.Li M, Yu X, Guo H, Sun L, Wang A, Liu Q, Wang X, Li J. Bufalin exerts antitumor effects by inducing cell cycle arrest and triggering apoptosis in pancreatic cancer cells. Tumour Biol. 2014;35(3):2461–2471. doi: 10.1007/s13277-013-1326-6. [DOI] [PubMed] [Google Scholar]
- 48.Masuda Y, Kawazoe N, Nakajo S, Yoshida T, Kuroiwa Y, Nakaya K. Bufalin induces apoptosis and influences the expression of apoptosis-related genes in human leukemia cells. Leuk Res. 1995;19(8):549–556. doi: 10.1016/0145-2126(95)00031-I. [DOI] [PubMed] [Google Scholar]
- 49.Miao Q, Bi LL, Li X, Miao S, Zhang J, Zhang S, Yang Q, Xie YH, Zhang J, Wang SW. Anticancer effects of bufalin on human hepatocellular carcinoma HepG2 cells: roles of apoptosis and autophagy. Int J Mol Sci. 2013;14(1):1370–1382. doi: 10.3390/ijms14011370. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 50.Sun L, Chen T, Wang X, Chen Y, Wei X. Bufalin induces reactive oxygen species dependent bax translocation and apoptosis in ASTC-a-1 cells. Evid Based Complement Alternat Med. 2011;2011:249090. doi: 10.1093/ecam/nep082. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 51.Wang D, Bi Z. Bufalin inhibited the growth of human osteosarcoma MG-63 cells via down-regulation of Bcl-2/Bax and triggering of the mitochondrial pathway. Tumour Biol. 2014;35(5):4885–4890. doi: 10.1007/s13277-014-1640-7. [DOI] [PubMed] [Google Scholar]
- 52.Watabe M, Kawazoe N, Masuda Y, Nakajo S, Nakaya K. Bcl-2 protein inhibits bufalin-induced apoptosis through inhibition of mitogen-activated protein kinase activation in human leukemia U937 cells. Cancer Res. 1997;57(15):3097–3100. [PubMed] [Google Scholar]
- 53.Yin PH, Liu X, Qiu YY, Cai JF, Qin JM, Zhu HR, Li Q. Anti-tumor activity and apoptosis-regulation mechanisms of bufalin in various cancers: new hope for cancer patients. Asian Pac J Cancer Prev. 2012;13(11):5339–5343. doi: 10.7314/APJCP.2012.13.11.5339. [DOI] [PubMed] [Google Scholar]
- 54.Arnaud-Batista FJ, Costa GT, Oliveira IM, Costa PP, Santos CF, Fonteles MC, Uchoa DE, Silveira ER, Cardi BA, Carvalho KM, et al. Natriuretic effect of bufalin in isolated rat kidneys involves activation of the Na+–K+-ATPase-Src kinase pathway. Am J Physiol Renal Physiol. 2012;302(8):F959–F966. doi: 10.1152/ajprenal.00130.2011. [DOI] [PubMed] [Google Scholar]
- 55.Song X, Zhang C, Zhao M, Chen H, Liu X, Chen J, Lonard DM, Qin L, Xu J, Wang X, et al. Steroid receptor coactivator-3 (SRC-3/AIB1) as a novel therapeutic target in triple negative breast cancer and its inhibition with a phospho-bufalin prodrug. PLoS ONE. 2015;10(10):e0140011. doi: 10.1371/journal.pone.0140011. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 56.Wang Y, Lonard DM, Yu Y, Chow DC, Palzkill TG, Wang J, Qi R, Matzuk AJ, Song X, Madoux F, et al. Bufalin is a potent small-molecule inhibitor of the steroid receptor coactivators SRC-3 and SRC-1. Cancer Res. 2014;74(5):1506–1517. doi: 10.1158/0008-5472.CAN-13-2939. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 57.Calderon-Montano JM, Burgos-Moron E, Orta ML, Garcia-Dominguez I, Maldonado-Navas D, Lopez-Lazaro M. Bufalin is a steroid receptor coactivator inhibitor-letter. Cancer Res. 2015;75(6):1156. doi: 10.1158/0008-5472.CAN-14-0398. [DOI] [PubMed] [Google Scholar]
- 58.Lonard DM, Xu J, O’Malley BW. Bufalin is a steroid receptor coactivator inhibitor-response. Cancer Res. 2015;75(6):1157. doi: 10.1158/0008-5472.CAN-14-2648. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 59.Xie CM, Chan WY, Yu S, Zhao J, Cheng CH. Bufalin induces autophagy-mediated cell death in human colon cancer cells through reactive oxygen species generation and JNK activation. Free Radic Biol Med. 2011;51(7):1365–1375. doi: 10.1016/j.freeradbiomed.2011.06.016. [DOI] [PubMed] [Google Scholar]
- 60.Gao Y, Li HX, Xu LT, Wang P, Xu LY, Cohen L, Yang PY, Gu K, Meng ZQ. Bufalin enhances the anti-proliferative effect of sorafenib on human hepatocellular carcinoma cells through downregulation of ERK. Mol Biol Rep. 2012;39(2):1683–1689. doi: 10.1007/s11033-011-0908-x. [DOI] [PubMed] [Google Scholar]
- 61.Wang H, Zhang C, Chi H, Meng Z. Synergistic anticancer effects of bufalin and sorafenib by regulating apoptosis associated proteins. Mol Med Rep. 2018;17(6):8101–8110. doi: 10.3892/mmr.2018.8927. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 62.Nishida N, Yano H, Nishida T, Kamura T, Kojiro M. Angiogenesis in cancer. Vasc Health Risk Manag. 2006;2(3):213–219. doi: 10.2147/vhrm.2006.2.3.213. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 63.Wang H, Zhang C, Chi H, Meng Z. Synergistic anti-hepatoma effect of bufalin combined with sorafenib via mediating the tumor vascular microenvironment by targeting mTOR/VEGF signaling. Int J Oncol. 2018;52(6):2051–2060. doi: 10.3892/ijo.2018.4351. [DOI] [PubMed] [Google Scholar]
- 64.Wang H, Zhang C, Ning Z, Xu L, Zhu X, Meng Z. Bufalin enhances anti-angiogenic effect of sorafenib via AKT/VEGF signaling. Int J Oncol. 2016;48(3):1229–1241. doi: 10.3892/ijo.2016.3326. [DOI] [PubMed] [Google Scholar]
- 65.Deryugina EI, Quigley JP. Matrix metalloproteinases and tumor metastasis. Cancer Metastasis Rev. 2006;25(1):9–34. doi: 10.1007/s10555-006-7886-9. [DOI] [PubMed] [Google Scholar]
- 66.Gai JQ, Sheng X, Qin JM, Sun K, Zhao W, Ni L. The effect and mechanism of bufalin on regulating hepatocellular carcinoma cell invasion and metastasis via Wnt/beta-catenin signaling pathway. Int J Oncol. 2016;48(1):338–348. doi: 10.3892/ijo.2015.3250. [DOI] [PubMed] [Google Scholar]
- 67.Sheng X, Sun X, Sun K, Sui H, Qin J, Li Q. Inhibitory effect of bufalin combined with Hedgehog signaling pathway inhibitors on proliferation and invasion and metastasis of liver cancer cells. Int J Oncol. 2016;49(4):1513–1524. doi: 10.3892/ijo.2016.3667. [DOI] [PubMed] [Google Scholar]
- 68.Chen YY, Lu HF, Hsu SC, Kuo CL, Chang SJ, Lin JJ, Wu PP, Liu JY, Lee CH, Chung JG, et al. Bufalin inhibits migration and invasion in human hepatocellular carcinoma SK-Hep1 cells through the inhibitions of NF-kB and matrix metalloproteinase-2/-9-signaling pathways. Environ Toxicol. 2015;30(1):74–82. doi: 10.1002/tox.21896. [DOI] [PubMed] [Google Scholar]
- 69.Chueh FS, Chen YY, Huang AC, Ho HC, Liao CL, Yang JS, Kuo CL, Chung JG. Bufalin-inhibited migration and invasion in human osteosarcoma U-2 OS cells is carried out by suppression of the matrix metalloproteinase-2, ERK, and JNK signaling pathways. Environ Toxicol. 2014;29(1):21–29. doi: 10.1002/tox.20769. [DOI] [PubMed] [Google Scholar]
- 70.Wu SH, Hsiao YT, Kuo CL, Yu FS, Hsu SC, Wu PP, Chen JC, Hsia TC, Liu HC, Hsu WH, et al. Bufalin inhibits NCI-H460 human lung cancer cell metastasis in vitro by inhibiting MAPKs, MMPs, and NF-kappaB pathways. Am J Chin Med. 2015;43(6):1247–1264. doi: 10.1142/S0192415X15500718. [DOI] [PubMed] [Google Scholar]
- 71.Qiu DZ, Zhang ZJ, Wu WZ, Yang YK. Bufalin, a component in Chansu, inhibits proliferation and invasion of hepatocellular carcinoma cells. BMC Complement Altern Med. 2013;13:185. doi: 10.1186/1472-6882-13-185. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 72.Zhang ZJ, Yang YK, Wu WZ. Bufalin attenuates the stage and metastatic potential of hepatocellular carcinoma in nude mice. J Transl Med. 2014;12:57. doi: 10.1186/1479-5876-12-57. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 73.Yu Z, Feng H, Sun X, Zhuo Y, Li M, Zhou Z, Huang L, Jiang Y, Zhu X, Zhang X, et al. Bufalin suppresses hepatocarcinogenesis by targeting beta-catenin/TCF signaling via cell cycle-related kinase. Sci Rep. 2018;8(1):3891. doi: 10.1038/s41598-018-22113-2. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 74.Huang AC, Yang MD, Hsiao YT, Lin TS, Ma YS, Peng SF, Hsia TC, Cheng YD, Kuo CL, Chung JG. Bufalin inhibits gefitinib resistant NCI-H460 human lung cancer cell migration and invasion in vitro. J Ethnopharmacol. 2016;194:1043–1050. doi: 10.1016/j.jep.2016.11.004. [DOI] [PubMed] [Google Scholar]
- 75.Kang X, Lu P, Cui Y, Wang Y, Zhang Q, Gong Y, Xu Z. Bufalin reverses hepatocyte growth factor-induced resistance to afatinib in H1975 lung cancer cells. Zhonghua Zhong Liu Za Zhi. 2015;37(7):490–496. [PubMed] [Google Scholar]
- 76.Wang H, Zhang C, Xu L, Zang K, Ning Z, Jiang F, Chi H, Zhu X, Meng Z. Bufalin suppresses hepatocellular carcinoma invasion and metastasis by targeting HIF-1alpha via the PI3K/AKT/mTOR pathway. Oncotarget. 2016;7(15):20193–20208. doi: 10.18632/oncotarget.7935. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 77.Wang J, Cai H, Xia Y, Wang S, Xing L, Chen C, Zhang Y, Xu J, Yin P, Jiang Y, et al. Bufalin inhibits gastric cancer invasion and metastasis by down-regulating Wnt/ASCL2 expression. Oncotarget. 2018;9(34):23320–23333. doi: 10.18632/oncotarget.24157. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 78.Wang J, Xia Y, Zuo Q, Chen T. Molecular mechanisms underlying the antimetastatic activity of bufalin. Mol Clin Oncol. 2018;8(5):631–636. doi: 10.3892/mco.2018.1591. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 79.Zhao L, Liu S, Che X, Hou K, Ma Y, Li C, Wen T, Fan Y, Hu X, Liu Y, et al. Bufalin inhibits TGF-beta-induced epithelial-to-mesenchymal transition and migration in human lung cancer A549 cells by downregulating TGF-beta receptors. Int J Mol Med. 2015;36(3):645–652. doi: 10.3892/ijmm.2015.2268. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 80.Liu F, Tong D, Li H, Liu M, Li J, Wang Z, Cheng X. Bufalin enhances antitumor effect of paclitaxel on cervical tumorigenesis via inhibiting the integrin alpha2/beta5/FAK signaling pathway. Oncotarget. 2016;7(8):8896–8907. doi: 10.18632/oncotarget.6840. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 81.Xie J, Lin W, Huang L, Xu N, Xu A, Chen B, Watanabe M, Liu C, Huang P. Bufalin suppresses the proliferation and metastasis of renal cell carcinoma by inhibiting the PI3K/Akt/mTOR signaling pathway. Oncol Lett. 2018;16(3):3867–3873. doi: 10.3892/ol.2018.9111. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 82.Wang H, Ning Z, Li Y, Zhu X, Meng Z. Bufalin suppresses cancer stem-like cells in gemcitabine-resistant pancreatic cancer cells via Hedgehog signaling. Mol Med Rep. 2016;14(3):1907–1914. doi: 10.3892/mmr.2016.5471. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 83.Ji D, Liang Z, Liu G, Zhao G, Fang J. Bufalin attenuates cancer-induced pain and bone destruction in a model of bone cancer. Naunyn Schmiedebergs Arch Pharmacol. 2017;390(12):1211–1219. doi: 10.1007/s00210-017-1419-7. [DOI] [PubMed] [Google Scholar]
- 84.Ozdemir A, Simay YD, Ibisoglu B, Yaren B, Bulbul D, Ark M. Cardiac glycoside-induced cell death and Rho/Rho kinase pathway: implication of different regulation in cancer cell lines. Steroids. 2016;109:29–43. doi: 10.1016/j.steroids.2016.03.015. [DOI] [PubMed] [Google Scholar]
- 85.Li H, Hu S, Pang Y, Li M, Chen L, Liu F, Liu M, Wang Z, Cheng X. Bufalin inhibits glycolysis-induced cell growth and proliferation through the suppression of Integrin beta2/FAK signaling pathway in ovarian cancer. Am J Cancer Res. 2018;8(7):1288–1296. [PMC free article] [PubMed] [Google Scholar]
- 86.Xie XB, Yin JQ, Wen LL, Gao ZH, Zou CY, Wang J, Huang G, Tang QL, Colombo C, He WL, et al. Critical role of heat shock protein 27 in bufalin-induced apoptosis in human osteosarcomas: a proteomic-based research. PLoS ONE. 2012;7(10):e47375. doi: 10.1371/journal.pone.0047375. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 87.Xie XB, Wen LL, Yin JQ, Liao HY, Zou CY, Wang B, Huang G, Shen JN. Proteomics research of bufalin-induced apoptosis in osteosarcoma cell lines. Zhongguo Zhong Yao Za Zhi. 2014;39(14):2739–2743. [PubMed] [Google Scholar]
- 88.Zhang DM, Feng LX, Liu M, Jin WH, Luo J, Nie AY, Zhou Y, Li Y, Wu WY, Jiang BH, et al. Possible target-related proteins and signal network of bufalin in A549 cells suggested by both iTRAQ-based and label-free proteomic analysis. Proteomics. 2016;16(6):935–945. doi: 10.1002/pmic.201500418. [DOI] [PubMed] [Google Scholar]
- 89.Wu SH, Hsiao YT, Chen JC, Lin JH, Hsu SC, Hsia TC, Yang ST, Hsu WH, Chung JG. Bufalin alters gene expressions associated DNA damage, cell cycle, and apoptosis in human lung cancer NCI-H460 cells in vitro. Molecules. 2014;19(5):6047–6057. doi: 10.3390/molecules19056047. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 90.Fell DA. Beyond genomics. Trends Genet. 2001;17(12):680–682. doi: 10.1016/S0168-9525(01)02521-5. [DOI] [PubMed] [Google Scholar]
- 91.Wu XY, Tian F, Su MH, Wu M, Huang Y, Hu LH, Jin L, Zhu XJ. BF211, a derivative of bufalin, enhances the cytocidal effects in multiple myeloma cells by inhibiting the IL-6/JAK2/STAT3 pathway. Int Immunopharmacol. 2018;64:24–32. doi: 10.1016/j.intimp.2018.08.016. [DOI] [PubMed] [Google Scholar]
- 92.Sun P, Feng LX, Zhang DM, Liu M, Liu W, Mi T, Wu WY, Jiang BH, Yang M, Hu LH, et al. Bufalin derivative BF211 inhibits proteasome activity in human lung cancer cells in vitro by inhibiting beta1 subunit expression and disrupting proteasome assembly. Acta Pharmacol Sin. 2016;37(7):908–918. doi: 10.1038/aps.2016.30. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 93.Hu Q, Liang B, Sun Y, Guo XL, Bao YJ, Xie DH, Zhou M, Duan YR, Yin PH, Peng ZH. Preparation of bufalin-loaded pluronic polyetherimide nanoparticles, cellular uptake, distribution, and effect on colorectal cancer. Int J Nanomed. 2014;9:4035–4041. doi: 10.2147/IJN.S64708. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 94.Yang Z, Liu J, Huang Q, Zhang Z, Zhang J, Pan Y, Yang Y, Cheng D. Radiosynthesis and pharmacokinetics of [(18)F]fluoroethyl bufalin in hepatocellular carcinoma-bearing mice. Onco Targets Ther. 2017;10:329–338. doi: 10.2147/OTT.S110281. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 95.Zou A, Zhao X, Handge UA, Garamus VM, Willumeit-Romer R, Yin P. Folate receptor targeted bufalin/beta-cyclodextrin supramolecular inclusion complex for enhanced solubility and anti-tumor efficiency of bufalin. Mater Sci Eng C Mater Biol Appl. 2017;78:609–618. doi: 10.1016/j.msec.2017.04.094. [DOI] [PubMed] [Google Scholar]
- 96.Chen Q, Liu J. Transferrin and folic acid co-modified bufalin-loaded nanoliposomes: preparation, characterization, and application in anticancer activity. Int J Nanomed. 2018;13:6009–6018. doi: 10.2147/IJN.S176012. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 97.Yuan J, Zhou X, Cao W, Bi L, Zhang Y, Yang Q, Wang S. Improved antitumor efficacy and pharmacokinetics of bufalin via PEGylated liposomes. Nanoscale Res Lett. 2017;12(1):585. doi: 10.1186/s11671-017-2346-8. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 98.Li Y, Yuan J, Yang Q, Cao W, Zhou X, Xie Y, Tu H, Zhang Y, Wang S. Immunoliposome co-delivery of bufalin and anti-CD40 antibody adjuvant induces synergetic therapeutic efficacy against melanoma. Int J Nanomed. 2014;9:5683–5700. doi: 10.2147/IJN.S73651. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 99.Shi XJ, Qiu YY, Yu H, Liu C, Yuan YX, Yin PH, Liu T. Increasing the anticancer performance of bufalin (BUF) by introducing an endosome-escaping polymer and tumor-targeting peptide in the design of a polymeric prodrug. Colloids Surf B Biointerfaces. 2018;166:224–234. doi: 10.1016/j.colsurfb.2018.03.024. [DOI] [PubMed] [Google Scholar]
- 100.Liu T, Jia T, Yuan X, Liu C, Sun J, Ni Z, Xu J, Wang X, Yuan Y. Development of octreotide-conjugated polymeric prodrug of bufalin for targeted delivery to somatostatin receptor 2 overexpressing breast cancer in vitro and in vivo. Int J Nanomed. 2016;11:2235–2250. doi: 10.2217/nnm-2016-0234. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 101.Liu T, Yuan X, Jia T, Liu C, Ni Z, Qin Z, Yuan Y. Polymeric prodrug of bufalin for increasing solubility and stability: synthesis and anticancer study in vitro and in vivo. Int J Pharm. 2016;506(1–2):382–393. doi: 10.1016/j.ijpharm.2016.04.041. [DOI] [PubMed] [Google Scholar]
- 102.Zhang H, Huang N, Yang G, Lin Q, Su Y. Bufalin-loaded bovine serum albumin nanoparticles demonstrated improved anti-tumor activity against hepatocellular carcinoma: preparation, characterization, pharmacokinetics and tissue distribution. Oncotarget. 2017;8(38):63311–63323. doi: 10.18632/oncotarget.18800. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 103.Liu Y, Wang P, Sun C, Zhao J, Du Y, Shi F, Feng N. Bioadhesion and enhanced bioavailability by wheat germ agglutinin-grafted lipid nanoparticles for oral delivery of poorly water-soluble drug bufalin. Int J Pharm. 2011;419(1–2):260–265. doi: 10.1016/j.ijpharm.2011.07.019. [DOI] [PubMed] [Google Scholar]
- 104.Tian X, Yin H, Zhang S, Luo Y, Xu K, Ma P, Sui C, Meng F, Liu Y, Jiang Y, et al. Bufalin loaded biotinylated chitosan nanoparticles: an efficient drug delivery system for targeted chemotherapy against breast carcinoma. Eur J Pharm Biopharm. 2014;87(3):445–453. doi: 10.1016/j.ejpb.2014.05.010. [DOI] [PubMed] [Google Scholar]
- 105.Yin P, Wang Y, Qiu Y, Hou L, Liu X, Qin J, Duan Y, Liu P, Qiu M, Li Q. Bufalin-loaded mPEG-PLGA-PLL-cRGD nanoparticles: preparation, cellular uptake, tissue distribution, and anticancer activity. Int J Nanomed. 2012;7:3961–3969. doi: 10.2147/IJN.S32063. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 106.Zhang HQ, Yin ZF, Sheng JY, Jiang ZQ, Wu BY, Su YH. A comparison study of pharmacokinetics between bufalin-loaded bovine serum albumin nanoparticles and bufalin in rats. Zhong Xi Yi Jie He Xue Bao. 2012;10(6):674–680. doi: 10.3736/jcim20120612. [DOI] [PubMed] [Google Scholar]
- 107.Bick RJ, Poindexter BJ, Sweney RR, Dasgupta A. Effects of Chan Su, a traditional Chinese medicine, on the calcium transients of isolated cardiomyocytes: cardiotoxicity due to more than Na, K-ATPase blocking. Life Sci. 2002;72(6):699–709. doi: 10.1016/S0024-3205(02)02302-0. [DOI] [PubMed] [Google Scholar]
- 108.Koh CH, Wu J, Chung YY, Liu Z, Zhang RR, Chong K, Korzh V, Ting S, Oh S, Shim W, et al. Electronic supplementary material Identification of a Na(+)/K(+)-ATPase inhibition-independent proarrhythmic ionic mechanisms of cardiac glycosides. Sci Rep. 2017;7(1):2465. doi: 10.1038/s41598-017-02496-4. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 109.Nesher M, Shpolansky U, Viola N, Dvela M, Buzaglo N, Cohen Ben-Ami H, Rosen H, Lichtstein D. Ouabain attenuates cardiotoxicity induced by other cardiac steroids. Br J Pharmacol. 2010;160(2):346–354. doi: 10.1111/j.1476-5381.2010.00701.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 110.Zhao M, Liu Y, Jin B, Hou K. Inhibition effect of ethanol extract of Chinese toad venom on H22 ascites and solid tumors and its toxicity in mice. J Qilu Oncol. 2005;22:1705–1709. [Google Scholar]
- 111.Zhao M, Liu Y, Hou K, Zhu Z, Tian X. Inhibition effect of ethanol extract of toad venom on S180 ascites and solid tumors and its toxicity in mice. Shanxi Med J. 2006;03:189–192. [Google Scholar]
- 112.Zhao M, Liu Y, Hou K. Inhibition effect of ethanol extract of toad venom on Ehrlich’s ascites carcinomas and its toxicity in mice. Shanxi Med J. 2006;06:479–482. [Google Scholar]
- 113.Zhu LB, Liu QY. Clinical observation of cinobufacin combined with cisplatin through intrapericardial infusion in treatment of malignant pericardial effusion. Chin J Med Guide. 2009;09:1525–1526. [Google Scholar]
- 114.Qi F, Li A, Inagaki Y, Gao J, Li J, Kokudo N, Li XK, Tang W. Chinese herbal medicines as adjuvant treatment during chemo- or radio-therapy for cancer. Biosci Trends. 2010;4(6):297–307. [PubMed] [Google Scholar]
- 115.Wang Z, Qi F, Cui Y, Zhao L, Sun X, Tang W, Cai P. An update on Chinese herbal medicines as adjuvant treatment of anticancer therapeutics. Biosci Trends. 2018;12(3):220–239. doi: 10.5582/bst.2018.01144. [DOI] [PubMed] [Google Scholar]
- 116.Lin Y, Wang Z, Wang Z, Gan P. Clinical observation on cinobufotalin tablets combined with XELOX regimen in chemotherapy after colorectal cancer surgery. Chin Med Mod Distance Educ China. 2017;22:107–109. [Google Scholar]
- 117.Meng Z, Liu F, Shen Y, Yang P, Cohen L, Huo Y, Zhao Q, Ng CS, Chang DZ, Garrett CR. A randomized phase II study of gemcitabine (G) plus the cardiac glycoside huachansu (H) in the treatment of patients with locally advanced (LAPC) or metastatic pancreatic cancer (MPC) J Clin Oncol. 2011;29:4127. doi: 10.1200/jco.2011.29.15_suppl.4127. [DOI] [Google Scholar]
- 118.Zhakeer Z, Hadeer M, Tuerxun Z, Tuerxun K. Bufalin inhibits the inflammatory effects in asthmatic mice through the suppression of nuclear factor-kappa B activity. Pharmacology. 2017;99(3–4):179–187. doi: 10.1159/000450754. [DOI] [PubMed] [Google Scholar]
- 119.Chang YW, Zhao YF, Cao YL, Gu W, Pang J, Zhan HS. Bufalin exerts inhibitory effects on IL-1beta-mediated proliferation and induces apoptosis in human rheumatoid arthritis fibroblast-like synoviocytes. Inflammation. 2014;37(5):1552–1559. doi: 10.1007/s10753-014-9882-5. [DOI] [PubMed] [Google Scholar]
- 120.Rong X, Ni W, Liu Y, Wen J, Qian C, Sun L, Wang J. Bufalin, a bioactive component of the Chinese medicine chansu, inhibits inflammation and invasion of human rheumatoid arthritis fibroblast-like synoviocytes. Inflammation. 2014;37(4):1050–1058. doi: 10.1007/s10753-014-9828-y. [DOI] [PubMed] [Google Scholar]
- 121.Xie S, Spelmink L, Codemo M, Subramanian K, Putsep K, Henriques-Normark B, Olliver M. Cinobufagin modulates human innate immune responses and triggers antibacterial activity. PLoS ONE. 2016;11(8):e0160734. doi: 10.1371/journal.pone.0160734. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 122.Wen L, Huang Y, Xie X, Huang W, Yin J, Lin W, Jia Q, Zeng W. Anti-inflammatory and antinociceptive activities of bufalin in rodents. Mediat Inflamm. 2014;2014:171839. doi: 10.1155/2014/171839. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 123.Tongkao-On W, Gordon-Thomson C, Dixon KM, Song EJ, Luu T, Carter SE, Sequeira VB, Reeve VE, Mason RS. Novel vitamin D compounds and skin cancer prevention. Dermatoendocrinol. 2013;5(1):20–33. doi: 10.4161/derm.23939. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 124.Lan YL, Wang X, Lou JC, Xing JS, Yu ZL, Wang H, Zou S, Ma X, Zhang B. Bufalin inhibits glioblastoma growth by promoting proteasomal degradation of the Na(+)/K(+)-ATPase alpha1 subunit. Biomed Pharmacother. 2018;103:204–215. doi: 10.1016/j.biopha.2018.04.030. [DOI] [PubMed] [Google Scholar]
- 125.Yan S, Qu X, Xu C, Zhu Z, Zhang L, Xu L, Song N, Teng Y, Liu Y. Down-regulation of Cbl-b by bufalin results in up-regulation of DR4/DR5 and sensitization of TRAIL-induced apoptosis in breast cancer cells. J Cancer Res Clin Oncol. 2012;138(8):1279–1289. doi: 10.1007/s00432-012-1204-4. [DOI] [PubMed] [Google Scholar]
- 126.Wang Q, Li C, Zhu Z, Teng Y, Che X, Wang Y, Ma Y, Wang Y, Zheng H, Liu Y, et al. miR-155-5p antagonizes the apoptotic effect of bufalin in triple-negative breast cancer cells. Anticancer Drugs. 2016;27(1):9–16. doi: 10.1097/CAD.0000000000000296. [DOI] [PubMed] [Google Scholar]
- 127.Pan S, Wang Y, Feng L, Fan C, Guo D, Liu X, Fan J. Study on proteomics of Hela cell apoptosis in bufalin-induced human cervical carcinoma. Zhongguo Zhong Yao Za Zhi. 2012;37(13):1998–2004. [PubMed] [Google Scholar]
- 128.Zhu Z, Li E, Liu Y, Gao Y, Sun H, Ma G, Wang Z, Liu X, Wang Q, Qu X, et al. Inhibition of Jak-STAT3 pathway enhances bufalin-induced apoptosis in colon cancer SW620 cells. World J Surg Oncol. 2012;10:228. doi: 10.1186/1477-7819-10-228. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 129.Zhang N, Xie Y, Tai Y, Gao Y, Guo W, Yu W, Li J, Feng X, Hao J, Gao Y, et al. Bufalin inhibits hTERT expression and colorectal cancer cell growth by targeting CPSF4. Cell Physiol Biochem. 2016;40(6):1559–1569. doi: 10.1159/000453206. [DOI] [PubMed] [Google Scholar]
- 130.Qiu YY, Hu Q, Tang QF, Feng W, Hu SJ, Liang B, Peng W, Yin PH. MicroRNA-497 and bufalin act synergistically to inhibit colorectal cancer metastasis. Tumour Biol. 2014;35(3):2599–2606. doi: 10.1007/s13277-013-1342-6. [DOI] [PubMed] [Google Scholar]
- 131.Wang J, Chen C, Wang S, Zhang Y, Yin P, Gao Z, Xu J, Feng D, Zuo Q, Zhao R, et al. Bufalin inhibits HCT116 colon cancer cells and its orthotopic xenograft tumor in mice model through genes related to apoptotic and PTEN/AKT pathways. Gastroenterol Res Pract. 2015;2015:457193. doi: 10.1155/2015/457193. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 132.Takai N, Ueda T, Nishida M, Nasu K, Narahara H. Bufalin induces growth inhibition, cell cycle arrest and apoptosis in human endometrial and ovarian cancer cells. Int J Mol Med. 2008;21(5):637–643. [PubMed] [Google Scholar]
- 133.Lv J, Lin S, Peng P, Cai C, Deng J, Wang M, Li X, Lin R, Lin Y, Fang A, et al. Arenobufagin activates p53 to trigger esophageal squamous cell carcinoma cell apoptosis in vitro and in vivo. Onco Targets Ther. 2017;10:1261–1267. doi: 10.2147/OTT.S104767. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 134.Li A, Qu X, Li Z, Qu J, Song N, Ma Y, Liu Y. Secreted protein acidic and rich in cysteine antagonizes bufalin-induced apoptosis in gastric cancer cells. Mol Med Rep. 2015;12(2):2926–2932. doi: 10.3892/mmr.2015.3676. [DOI] [PubMed] [Google Scholar]
- 135.Tsai SC, Yang JS, Peng SF, Lu CC, Chiang JH, Chung JG, Lin MW, Lin JK, Amagaya S, Wai-Shan Chung C, et al. Bufalin increases sensitivity to AKT/mTOR-induced autophagic cell death in SK-HEP-1 human hepatocellular carcinoma cells. Int J Oncol. 2012;41(4):1431–1442. doi: 10.3892/ijo.2012.1579. [DOI] [PubMed] [Google Scholar]
- 136.Zhai X, Lu J, Wang Y, Fang F, Li B, Gu W. Reversal effect of bufalin on multidrug resistance in K562/VCR vincristine-resistant leukemia cell line. J Tradit Chin Med. 2014;34(6):678–683. doi: 10.1016/S0254-6272(15)30082-0. [DOI] [PubMed] [Google Scholar]
- 137.Wang LP, Zhao YN, Sun X, Gao RL. Effects of bufalin on up-regulating methylation of Wilm’s tumor 1 gene in human erythroid leukemic cells. Chin J Integr Med. 2017;23(4):288–294. doi: 10.1007/s11655-017-2404-1. [DOI] [PubMed] [Google Scholar]
- 138.Zhu Z, Sun H, Ma G, Wang Z, Li E, Liu Y, Liu Y. Bufalin induces lung cancer cell apoptosis via the inhibition of PI3K/Akt pathway. Int J Mol Sci. 2012;13(2):2025–2035. doi: 10.3390/ijms13022025. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 139.Tsai SC, Lu CC, Lee CY, Lin YC, Chung JG, Kuo SC, Amagaya S, Chen FN, Chen MY, Chan SF, et al. AKT serine/threonine protein kinase modulates bufalin-triggered intrinsic pathway of apoptosis in CAL 27 human oral cancer cells. Int J Oncol. 2012;41(5):1683–1692. doi: 10.3892/ijo.2012.1605. [DOI] [PubMed] [Google Scholar]
- 140.Tian X, Dai S, Sun J, Jiang S, Sui C, Meng F, Li Y, Fu L, Jiang T, Wang Y, et al. Bufalin induces mitochondria-dependent apoptosis in pancreatic and oral cancer cells by downregulating hTERT expression via activation of the JNK/p38 pathway. Evid Based Complement Alternat Med. 2015;2015:546210. doi: 10.1155/2015/546210. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 141.Zhang J, Sha J, Zhou Y, Han K, Wang Y, Su Y, Yin X, Hu H, Yao Y. Bufalin inhibits proliferation and induces apoptosis in osteosarcoma cells by downregulating MicroRNA-221. Evid Based Complement Alternat Med. 2016;2016:7319464. doi: 10.1155/2016/7319464. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 142.Yin JQ, Shen JN, Su WW, Wang J, Huang G, Jin S, Guo QC, Zou CY, Li HM, Li FB. Bufalin induces apoptosis in human osteosarcoma U-2OS and U-2OS methotrexate300-resistant cell lines. Acta Pharmacol Sin. 2007;28(5):712–720. doi: 10.1111/j.1745-7254.2007.00559.x. [DOI] [PubMed] [Google Scholar]
- 143.Chou HY, Chueh FS, Ma YS, Wu RS, Liao CL, Chu YL, Fan MJ, Huang WW, Chung JG. Bufalin induced apoptosis in SCC4 human tongue cancer cells by decreasing Bcl2 and increasing Bax expression via the mitochondria dependent pathway. Mol Med Rep. 2017;16(6):7959–7966. doi: 10.3892/mmr.2017.7651. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 144.Yuan B, He J, Kisoh K, Hayashi H, Tanaka S, Si N, Zhao HY, Hirano T, Bian B, Takagi N. Effects of active bufadienolide compounds on human cancer cells and CD4+ CD25+ Foxp3+ regulatory T cells in mitogen-activated human peripheral blood mononuclear cells. Oncol Rep. 2016;36(3):1377–1384. doi: 10.3892/or.2016.4946. [DOI] [PubMed] [Google Scholar]
- 145.Yuan ZT, Shi XJ, Yuan YX, Qiu YY, Zou Y, Liu C, Yu H, He X, Xu K, Yin PH. Bufalin reverses ABCB1-mediated drug resistance in colorectal cancer. Oncotarget. 2017;8(29):48012–48026. doi: 10.18632/oncotarget.18225. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 146.Sun J, Xu K, Qiu Y, Gao H, Xu J, Tang Q, Yin P. Bufalin reverses acquired drug resistance by inhibiting stemness in colorectal cancer cells. Oncol Rep. 2017;38(3):1420–1430. doi: 10.3892/or.2017.5826. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 147.Xiang RF, Wang Y, Zhang N, Xu WB, Cao Y, Tong J, Li JM, Wu YL, Yan H. MK2206 enhances the cytocidal effects of bufalin in multiple myeloma by inhibiting the AKT/mTOR pathway. Cell Death Dis. 2017;8(5):e2776. doi: 10.1038/cddis.2017.188. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 148.Chen Y, Guo Q, Zhang B, Kang M, Xie Q, Wu Y. Bufalin enhances the antitumor effect of gemcitabine in pancreatic cancer. Oncol Lett. 2012;4(4):792–798. doi: 10.3892/ol.2012.783. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 149.Liu X, Xiao XY, Shou QY, Yan JF, Chen L, Fu HY, Wang JC. Bufalin inhibits pancreatic cancer by inducing cell cycle arrest via the c-Myc/NF-kappaB pathway. J Ethnopharmacol. 2016;193:538–545. doi: 10.1016/j.jep.2016.09.047. [DOI] [PubMed] [Google Scholar]
- 150.Wu SH, Bau DT, Hsiao YT, Lu KW, Hsia TC, Lien JC, Ko YC, Hsu WH, Yang ST, Huang YP, et al. Bufalin induces apoptosis in vitro and has antitumor activity against human lung cancer xenografts in vivo. Environ Toxicol. 2017;32(4):1305–1317. doi: 10.1002/tox.22325. [DOI] [PubMed] [Google Scholar]
- 151.Hong SH, Kim GY, Chang YC, Moon SK, Kim WJ, Choi YH. Bufalin prevents the migration and invasion of T24 bladder carcinoma cells through the inactivation of matrix metalloproteinases and modulation of tight junctions. Int J Oncol. 2013;42(1):277–286. doi: 10.3892/ijo.2012.1683. [DOI] [PubMed] [Google Scholar]
- 152.Kang KH, Han MH, Jeong JW, Park C, Lee SH, Lee HW, Hong SH, Choi YH, Hong SH. Bufalin sensitizes human bladder carcinoma cells to TRAIL-mediated apoptosis. Oncol Lett. 2017;14(1):853–859. doi: 10.3892/ol.2017.6223. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 153.Dong Y, Yin S, Li J, Jiang C, Ye M, Hu H. Bufadienolide compounds sensitize human breast cancer cells to TRAIL-induced apoptosis via inhibition of STAT3/Mcl-1 pathway. Apoptosis. 2011;16(4):394–403. doi: 10.1007/s10495-011-0573-5. [DOI] [PubMed] [Google Scholar]
- 154.Yan S, Qu X, Xu L, Che X, Ma Y, Zhang L, Teng Y, Zou H, Liu Y. Bufalin enhances TRAIL-induced apoptosis by redistributing death receptors in lipid rafts in breast cancer cells. Anticancer Drugs. 2014;25(6):683–689. doi: 10.1097/CAD.0000000000000095. [DOI] [PubMed] [Google Scholar]
- 155.Zou Z, Luo X, Nie P, Wu B, Zhang T, Wei Y, Wang W, Geng G, Jiang J, Mi Y. Inhibition of SRC-3 enhances sensitivity of human cancer cells to histone deacetylase inhibitors. Biochem Biophys Res Commun. 2016;478(1):227–233. doi: 10.1016/j.bbrc.2016.07.063. [DOI] [PubMed] [Google Scholar]
- 156.Xie CM, Liu XY, Yu S, Cheng CH. Cardiac glycosides block cancer growth through HIF-1alpha- and NF-kappaB-mediated Plk1. Carcinogenesis. 2013;34(8):1870–1880. doi: 10.1093/carcin/bgt136. [DOI] [PubMed] [Google Scholar]
- 157.Zhao H, Zhao D, Tan G, Liu Y, Zhuang L, Liu T. Bufalin promotes apoptosis of gastric cancer by down-regulation of miR-298 targeting bax. Int J Clin Exp Med. 2015;8(3):3420–3428. [PMC free article] [PubMed] [Google Scholar]
- 158.Zhao H, Zhao D, Jin H, Li H, Yang X, Zhuang L, Liu T. Bufalin reverses intrinsic and acquired drug resistance to cisplatin through the AKT signaling pathway in gastric cancer cells. Mol Med Rep. 2016;14(2):1817–1822. doi: 10.3892/mmr.2016.5426. [DOI] [PubMed] [Google Scholar]
- 159.Zhao H, Li Q, Pang J, Jin H, Li H, Yang X. Blocking autophagy enhances the pro-apoptotic effect of bufalin on human gastric cancer cells through endoplasmic reticulum stress. Biol Open. 2017;6(10):1416–1422. doi: 10.1242/bio.026344. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 160.Shen S, Zhang Y, Wang Z, Liu R, Gong X. Bufalin induces the interplay between apoptosis and autophagy in glioma cells through endoplasmic reticulum stress. Int J Biol Sci. 2014;10(2):212–224. doi: 10.7150/ijbs.8056. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 161.Liu T, Wu C, Weng G, Zhao Z, He X, Fu C, Sui Z, Huang SX. Bufalin inhibits cellular proliferation and cancer stem cell-like phenotypes via upregulation of MiR-203 in glioma. Cell Physiol Biochem. 2017;44(2):671–681. doi: 10.1159/000485279. [DOI] [PubMed] [Google Scholar]
- 162.Qi F, Inagaki Y, Gao B, Cui X, Xu H, Kokudo N, Li A, Tang W. Bufalin and cinobufagin induce apoptosis of human hepatocellular carcinoma cells via Fas- and mitochondria-mediated pathways. Cancer Sci. 2011;102(5):951–958. doi: 10.1111/j.1349-7006.2011.01900.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 163.Hu F, Han J, Zhai B, Ming X, Zhuang L, Liu Y, Pan S, Liu T. Blocking autophagy enhances the apoptosis effect of bufalin on human hepatocellular carcinoma cells through endoplasmic reticulum stress and JNK activation. Apoptosis. 2014;19(1):210–223. doi: 10.1007/s10495-013-0914-7. [DOI] [PubMed] [Google Scholar]
- 164.Gu W, Liu L, Fang FF, Huang F, Cheng BB, Li B. Reversal effect of bufalin on multidrug resistance in human hepatocellular carcinoma BEL-7402/5-FU cells. Oncol Rep. 2014;31(1):216–222. doi: 10.3892/or.2013.2817. [DOI] [PubMed] [Google Scholar]
- 165.Zhai B, Hu F, Yan H, Zhao D, Jin X, Fang T, Pan S, Sun X, Xu L. Bufalin reverses resistance to sorafenib by inhibiting Akt activation in hepatocellular carcinoma: the role of endoplasmic reticulum stress. PLoS ONE. 2015;10(9):e0138485. doi: 10.1371/journal.pone.0138485. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 166.Liu M, Feng LX, Sun P, Liu W, Mi T, Lei M, Wu W, Jiang B, Yang M, Hu L, et al. Knockdown of apolipoprotein E enhanced sensitivity of Hep3B cells to cardiac steroids via regulating Na+/K+-ATPase signalosome. Mol Cancer Ther. 2016;15(12):2955–2965. doi: 10.1158/1535-7163.MCT-15-0961. [DOI] [PubMed] [Google Scholar]
- 167.Xia J, Inagaki Y, Gao J, Qi F, Song P, Han G, Sawakami T, Gao B, Luo C, Kokudo N, et al. Combination of cinobufacini and doxorubicin increases apoptosis of hepatocellular carcinoma cells through the Fas- and mitochondria-mediated pathways. Am J Chin Med. 2017;45(7):1537–1556. doi: 10.1142/S0192415X17500835. [DOI] [PubMed] [Google Scholar]
- 168.Amano Y, Cho Y, Matsunawa M, Komiyama K, Makishima M. Increased nuclear expression and transactivation of vitamin D receptor by the cardiotonic steroid bufalin in human myeloid leukemia cells. J Steroid Biochem Mol Biol. 2009;114(3–5):144–151. doi: 10.1016/j.jsbmb.2009.01.022. [DOI] [PubMed] [Google Scholar]
- 169.Zhu Z, Li E, Liu Y, Gao Y, Sun H, Wang Y, Wang Z, Liu X, Wang Q, Liu Y. Bufalin induces the apoptosis of acute promyelocytic leukemia cells via the downregulation of survivin expression. Acta Haematol. 2012;128(3):144–150. doi: 10.1159/000339424. [DOI] [PubMed] [Google Scholar]
- 170.Kang XH, Xu ZY, Gong YB, Wang LF, Wang ZQ, Xu L, Cao F, Liao MJ. Bufalin reverses HGF-induced resistance to EGFR-TKIs in EGFR mutant lung cancer cells via blockage of Met/PI3k/Akt pathway and induction of apoptosis. Evid Based Complement Alternat Med. 2013;2013:243859. doi: 10.1155/2013/243859. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 171.Kang XH, Zhang JH, Zhang QQ, Cui YH, Wang Y, Kou WZ, Miao ZH, Lu P, Wang LF, Xu ZY, et al. Degradation of Mcl-1 through GSK-3beta activation regulates apoptosis induced by bufalin in non-small cell lung cancer H1975 cells. Cell Physiol Biochem. 2017;41(5):2067–2076. doi: 10.1159/000475438. [DOI] [PubMed] [Google Scholar]
- 172.Zhang C, Fu L. Effects of bufalin combined with doxorubicin on the proliferation and apoptosis of human lung cancer cell line A549 in vitro. Zhong Nan Da Xue Xue Bao Yi Xue Ban. 2017;42(7):762–768. doi: 10.11817/j.issn.1672-7347.2017.07.004. [DOI] [PubMed] [Google Scholar]
- 173.Huang H, Cao Y, Wei W, Liu W, Lu SY, Chen YB, Wang Y, Yan H, Wu YL. Targeting poly (ADP-ribose) polymerase partially contributes to bufalin-induced cell death in multiple myeloma cells. PLoS ONE. 2013;8(6):e66130. doi: 10.1371/journal.pone.0066130. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 174.Chang Y, Zhao Y, Zhan H, Wei X, Liu T, Zheng B. Bufalin inhibits the differentiation and proliferation of human osteosarcoma cell line hMG63-derived cancer stem cells. Tumour Biol. 2014;35(2):1075–1082. doi: 10.1007/s13277-013-1143-y. [DOI] [PubMed] [Google Scholar]
- 175.Chang Y, Zhao Y, Gu W, Cao Y, Wang S, Pang J, Shi Y. Bufalin inhibits the differentiation and proliferation of cancer stem cells derived from primary osteosarcoma cells through Mir-148a. Cell Physiol Biochem. 2015;36(3):1186–1196. doi: 10.1159/000430289. [DOI] [PubMed] [Google Scholar]
- 176.Lee CH, Shih YL, Lee MH, Au MK, Chen YL, Lu HF, Chung JG. Bufalin induces apoptosis of human osteosarcoma U-2 OS cells through endoplasmic reticulum stress, caspase- and mitochondria-dependent signaling pathways. Molecules. 2017;22(3):437. doi: 10.3390/molecules22030437. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 177.Chen H, Zhang L, Zhang L, Du J, Wang H, Wang B. MicroRNA-183 correlates cancer prognosis, regulates cancer proliferation and bufalin sensitivity in epithelial ovarian caner. Am J Transl Res. 2016;8(4):1748–1755. [PMC free article] [PubMed] [Google Scholar]
- 178.Yeh JY, Huang WJ, Kan SF, Wang PS. Effects of bufalin and cinobufagin on the proliferation of androgen dependent and independent prostate cancer cells. Prostate. 2003;54(2):112–124. doi: 10.1002/pros.10172. [DOI] [PubMed] [Google Scholar]
- 179.Zhai XF, Fang FF, Liu Q, Meng YB, Guo YY, Chen Z. MiR-181a contributes to bufalin-induced apoptosis in PC-3 prostate cancer cells. BMC Complement Altern Med. 2013;13:325. doi: 10.1186/1472-6882-13-325. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 180.Fridman E, Lichtstein D, Rosen H. Formation of new high density glycogen-microtubule structures is induced by cardiac steroids. J Biol Chem. 2012;287(9):6518–6529. doi: 10.1074/jbc.M111.273698. [DOI] [PMC free article] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.
Data Availability Statement
Not applicable.