

Abstract
Physiologically-relevant ALK (anaplastic lymphoma kinase) expression was not detected in human and mouse monocytes and macrophages, suggesting that the effects of bioactive compounds on stimulator of interferon genes (STING) activation may not depend on ALK.
Sepsis is a life-threatening condition associated with high lethality. The pathogenesis of sepsis is complex and requires the activation of the innate immune system by microbial pathogens. Part of this activation relies on the recognition of bacteria-derived cyclic dinucleotides (CDN) by the stimulator of interferon genes (STING) transmembrane protein that in turn binds TANK-binding kinase 1 (TBK1) to regulate the transcription of inflammatory genes triggered by the interferon regulatory factor 3 (IRF3). To discover actionable targets for the treatment of sepsis, Zeng et al. (1) recently screened a library of kinase inhibitors for STING-modulating properties and found that some inhibitors, including LDK378 [a Food and Drug Administration (FDA) approved anaplastic lymphoma kinase (ALK) inhibitor also called ceritinib], AP26113 (an FDA approved ALK inhibitor also called brigatinib) and AZD3463 (a preclinical ALK/IGF1R inhibitor) can efficiently block STING-mediated IFNβ release in cells stimulated with CDN. Understandably, they then focused on demonstrating ALK expression in monocytes and macrophages, which are important cells that mediate sepsis.
Surprisingly, they showed abundant expression of ALK in both human and murine monocytes and macrophages. These findings were puzzling as the current knowledge is that ALK is a tyrosine kinase receptor expressed during embryonic development by neurons and muscles, whereas in adults its expression is restricted to different areas of the brain (2–6). In contrast to normal cells, ALK can be readily detected in tumors such as neuroblastoma, ALK-rearranged anaplastic large cell lymphoma (ALCL), non-small cell lung cancer (NSCLC), or inflammatory fibroblastic tumor (IMT) (4, 5). In clinical practice ALK antibodies are considered highly specific for the detection of the oncogenic ALK because tumor cells express ALK more highly than ALK-negative benign cells. Detection of ALK expression in tumors is a critical point for the clinical decision on the use of ALK inhibitors in lung cancer patients, to the extent that the FDA has specifically approved an immunohistochemistry-based test to detect ALK-rearranged NSCLC (7).
We therefore decided to re-examine ALK expression in normal cells, in particular monocytes and macrophages. According to common databases such as The Genotype-Tissue Expression (GTEx) Project (https://www.gtexportal.org/home/gene/ALK) or to the Expression Atlas (http://www.ebi.ac.uk/gxa), ALK expression is overall low and largely confined to few areas in the brain or in the testis. Importantly, tissues rich in monocytes and macrophages such as peripheral blood, spleen, and lung have undetectable ALK expression (Fig. 1A). To directly characterize ALK expression in monocytes and macrophages, we performed Western Blot assays with the rabbit-monoclonal antibody SP8 from Abcam on the same mouse cell lines largely used in Zeng et al. (1), which are immortalized bone-marrow-derived macrophages (iBMDMs) and murine macrophage-like RAW264.7 cells. We used three different protein extraction buffers, all yielding comparable results for ALK expression. The SP8 antibody specifically detected ALK in 293T cells transfected with a plasmid encoding for the mouse full-length Alk receptor as well as the EML4-ALK fusion protein generated by CRISPR/Cas9 from the endogenous mouse Alk gene in lung cancer cells (8)(Fig. 1B). Surprisingly, this antibody did not detect any band corresponding to the mouse ALK receptor in iBMDMs or RAW264 cells (Fig. 1B). We then blotted the same lysates with the F-12 antibody used by Zeng et al. (1). This antibody was raised against an epitope mapping between amino acids 117–145 within the N-terminal extracellular domain of human ALK and recognized the transfected mouse ALK but not the EML4-ALK fusion that contains only the cytoplasmic portion of ALK. Strikingly, it failed to recognize any band consistent with ALK in iBMDMs or RAW264 (Fig. 1B).
Fig. 1. Undetectable ALK expression in monocytes and macrophages.
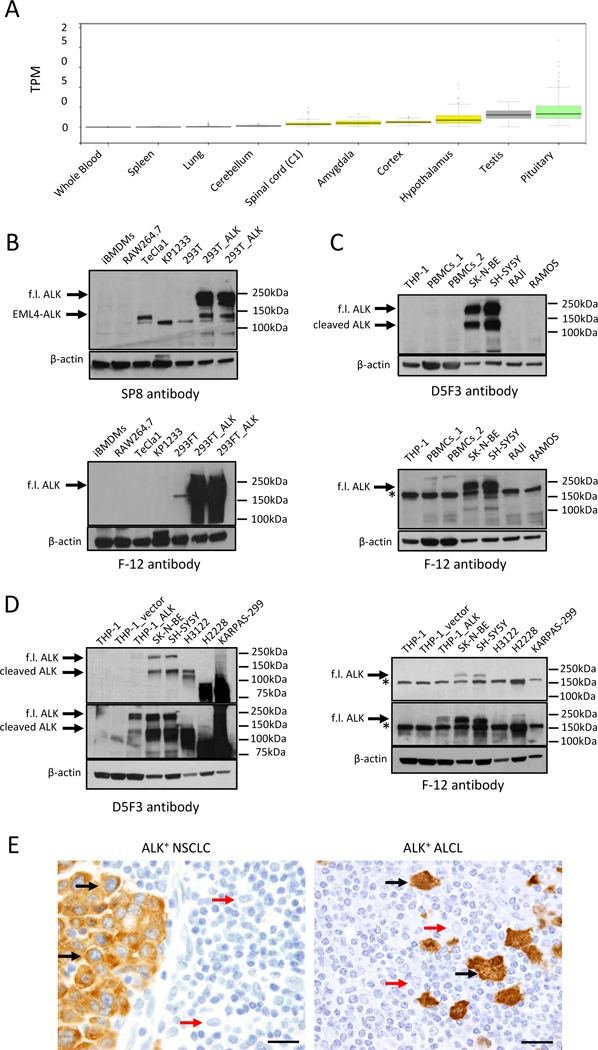
(A) ALK mRNA expression in tissues according to the Genotype-Tissue Expression (GTEx) project. (B) ALK expression in murine cell lines [top: SP8 anti-ALK antibody (1:1000); bottom: F12 anti-ALK antibody ( 1:100)]. Positive controls: 293T cells transfected with mouse Alk (293T_ALK) and EML4-ALK-expressing TeCla1 cells. Negative controls: untransfected 293T and KP1233 murine KRASG12D-driven NSCLC cells. Arrows indicate the full-length (f.l.) ALK or the EML4-ALK fusion. (C) ALK expression in human cell lines [top: D5F3 anti-ALK antibody (1:1000); bottom: F-12 anti-ALK antibody (1:100)]. Positive controls: human neuroblastoma cell lines SK-N-BE and SH-SY5Y. Negative controls: human B-lymphoma Raji and Ramos cell lines. Arrows indicate the full-length (f.l.) ALK or the cleaved ALK. Asterisk indicates the non-specific band detected by the F-12 antibody. (D) ALK expression in human cell lines [left: D5F3 anti-ALK (1:1000); right: F-12 anti-ALK antibody (1:100)]. Positive control: THP-1 cells transduced with human ALK receptor (THP-1_ALK). Negative control: THP-1 cells transduced with empty retroviral vector (THP-1_vector). H3122 and H2228 NSCLC cell lines carry the EML4-ALK (ex13-ex20) and the EML4-ALK (ex6-ex20) rearrangements that generate fusion proteins of 130 kDa and 90 kDa, respectively. Karpas-299 is an NPM-ALK-rearranged cell line that generates a fusion protein of 80 kDa. Positive controls: SK-N-BE and SH-SY5Y. Arrows indicate the full-length (f.l.) ALK or the cleaved ALK. Asterisk indicates the non-specific band detected by the F-12 antibody. Top panels: short exposure; Bottom panels: long exposure (E) Immunohistochemistry performed on an ALK-rearranged NSCLC case (D5F3 anti-ALK antibody) and on an ALCL case (4C5B8 anti-ALK antibody). Tumor cells shown by black arrows; macrophages shown by red arrows. Scale bar indicates 50 µm.
Next, we tested human peripheral blood mononuclear cells (PBMCs) and the human THP-1 cell line also used by Zeng et al. (1) to seek for ALK expression in human monocytes and macrophages. We used the FDA-approved D5F3 antibody for ALK detection (7) and neuroblastoma cells that express the endogenous human ALK as controls. As expected, the D5F3 antibody specifically recognized the ALK protein in neuroblastoma lines with a pattern consisted in two bands of 220–230 kDa and 130–140 kDa as previously reported, where the lower band corresponds to a cleavage product of the ALK receptor (9). In contrast, no bands with such pattern were seen in PBMCs or THP-1 cells even after long exposure of the membrane (Fig. 1C). When the same cell lysates were probed with the F-12 antibody used by Zeng et al. (1), the antibody correctly detected the 220–230 kDa full-length ALK in neuroblastoma cells, but not the 130–140 kDa cleaved ALK protein that lacks amino acids 117–145. Unexpectedly, this antibody also recognized a band of approximately 190–200 kDa that was detected across all human cells (Fig. 1C). However, this band is different from the pattern of ALK expression in neuroblastoma and was also seen in negative controls such as the ALK-negative Burkitt lymphoma cell lines Raji and Ramos (Fig. 1C). In addition, expression data from the Cancer Cell Line Encyclopedia (CCLE) (https://portals.broadinstitute.org/ccle) did not show detectable ALK mRNA in THP-1 cells (Fig. 2). To exclude an unexpected expression or cleavage of ALK in macrophages, we transduced human THP-1 cells with a retroviral vector encoding human ALK. Both the D5F3 and F-12 antibody recognized a band in ALK-transduced THP-1 cells, but not in THP-1 cells transduced with an empty vector, with a pattern similar to neuroblastoma cells. Again, the F-12 antibody recognized a lower band in all cell lines, including ALK-rearranged NSLCL and ALCL, that carry ALK fusions with different molecular weights (Fig. 1D). We concluded that the 190–200 kDa band detected by the F-12 antibody in all human cells is non-specific. Finally, immunohistochemistry failed to detect ALK expression in normal macrophages surrounding ALK-rearranged NCLSCL and ALCL (Fig. 1E).
Fig. 2. Undetectable ALK mRNA expression in Raji and THP-1 cell lines.
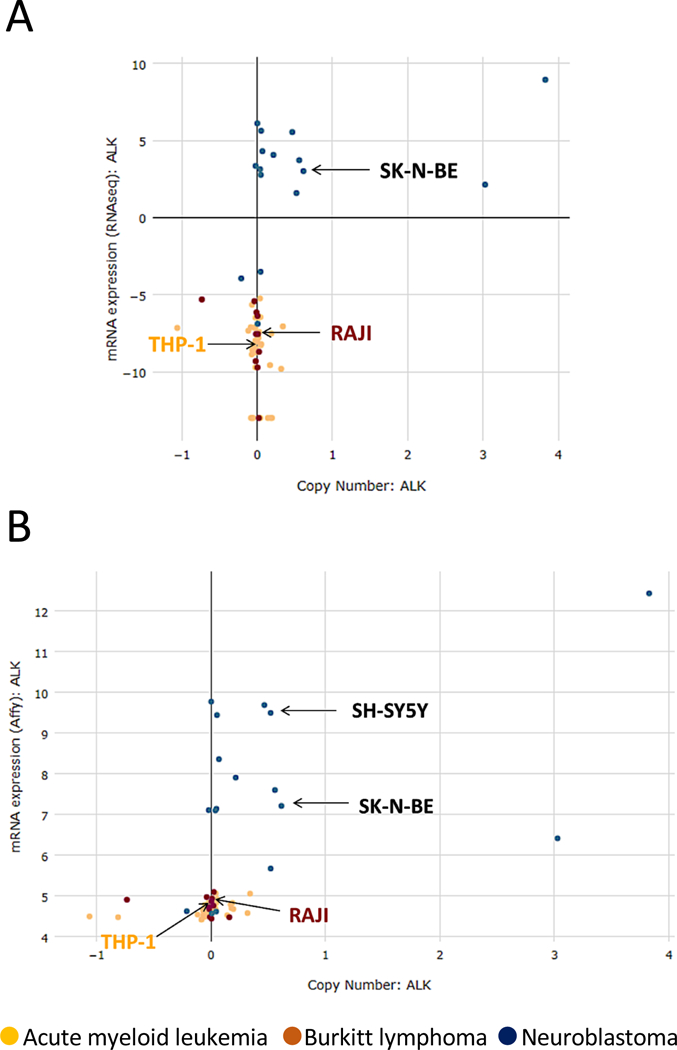
ALK mRNA expression in different tumor cell lines according to two different datasets in the Cancer Cell Line Encyclopedia (CCLE) (https://portals.broadinstitute.org/ccle). (A) RNA sequencing data; (B) Gene expression profiling data. Raji, THP-1 and the ALK-positive neuroblastoma cell lines SH-SY5Y and SK-N-BE are indicated by arrows.
Of note, Zeng et al. (1) performed both the library compound screening as well as all the experiments with LDK378 or AP26113 with an inhibitor concentration of 10 µM. However, drug concentration is critical for selective ALK inhibition. For example, LDK378 is selective for ALK with an IC50 of 200 pM but at higher concentrations inhibits other tyrosine kinases such as IGF-1R (IC50 of 8 nM), with broad inhibition of several tyrosine kinases at concentrations above 100 nM (10). Likewise, AP26113 has an IC50 of 370 pM for ALK and of 24.9 nM for IGF-1R (11). Therefore, when used at 10 µM [such as in the experiments performed by Zeng et al. (1)] it is likely that these compounds block multiple tyrosine kinases.
Taken together, our findings indicate that mouse and human monocytes and macrophages do not express detectable amounts of the ALK receptor. Thus, the interesting findings that LDK378 blocks CDN-induced STING activation in vitro, as well as polymicrobial sepsis or LPS-induced endotoxemia in vivo, may be explained by the activity of this compound against other kinase(s) different than ALK.
Supplementary Material
ACKNOWLEDGEMENTS
We would like to thank Dr. Chiara Ambrogio for the helpful discussion and critical reading of the manuscript. We would like to thank Dr. K. Fitzgerald (Medical School, UMass) for the THP-1, RAW264.7, and iBMDMs cell lines and Dr. T. Jacks (MIT) for the KP1233 cell line. The work has been supported by the R01 CA196703, D2017–026 V Foundation, and Translational Research Program (TRP) grants to RC.
Footnotes
Supplementary Materials
Materials and Methods
REFERENCES
- 1.Zeng L et al. , ALK is a therapeutic target for lethal sepsis. Science translational medicine 9, (2017). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 2.Iwahara T et al. , Molecular characterization of ALK, a receptor tyrosine kinase expressed specifically in the nervous system. Oncogene 14, 439–449 (1997). [DOI] [PubMed] [Google Scholar]
- 3.Morris SW et al. , ALK, the chromosome 2 gene locus altered by the t(2;5) in non-Hodgkin’s lymphoma, encodes a novel neural receptor tyrosine kinase that is highly related to leukocyte tyrosine kinase (LTK). Oncogene 14, 2175–2188 (1997). [DOI] [PubMed] [Google Scholar]
- 4.Chiarle R, Voena C, Ambrogio C, Piva R, Inghirami G, The anaplastic lymphoma kinase in the pathogenesis of cancer. Nature reviews. Cancer 8, 11–23 (2008). [DOI] [PubMed] [Google Scholar]
- 5.Hallberg B, Palmer RH, Mechanistic insight into ALK receptor tyrosine kinase in human cancer biology. Nature reviews. Cancer 13, 685–700 (2013). [DOI] [PubMed] [Google Scholar]
- 6.Vernersson E et al. , Characterization of the expression of the ALK receptor tyrosine kinase in mice. Gene expression patterns : GEP 6, 448–461 (2006). [DOI] [PubMed] [Google Scholar]
- 7.Conde E, Hernandez S, Prieto M, Martinez R, Lopez-Rios F, Profile of Ventana ALK (D5F3) companion diagnostic assay for non-small-cell lung carcinomas. Expert review of molecular diagnostics 16, 707–713 (2016). [DOI] [PubMed] [Google Scholar]
- 8.Blasco RB et al. , Simple and rapid in vivo generation of chromosomal rearrangements using CRISPR/Cas9 technology. Cell reports 9, 1219–1227 (2014). [DOI] [PubMed] [Google Scholar]
- 9.Moog-Lutz C et al. , Activation and inhibition of anaplastic lymphoma kinase receptor tyrosine kinase by monoclonal antibodies and absence of agonist activity of pleiotrophin. The Journal of biological chemistry 280, 26039–26048 (2005). [DOI] [PubMed] [Google Scholar]
- 10.Marsilje TH et al. , Synthesis, structure-activity relationships, and in vivo efficacy of the novel potent and selective anaplastic lymphoma kinase (ALK) inhibitor 5-chloro-N2-(2-isopropoxy-5-methyl-4-(piperidin-4-yl)phenyl)-N4-(2-(isopropylsulf onyl)phenyl)pyrimidine-2,4-diamine (LDK378) currently in phase 1 and phase 2 clinical trials. J Med Chem 56, 5675–5690 (2013). [DOI] [PubMed] [Google Scholar]
- 11.Huang WS et al. , Discovery of Brigatinib (AP26113), a Phosphine Oxide-Containing, Potent, Orally Active Inhibitor of Anaplastic Lymphoma Kinase. J Med Chem 59, 4948–4964 (2016). [DOI] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.