

Abstract
Genome-wide association studies (GWAS) were successful to identify genetic factors robustly associated with lung cancer. This review aims to synthesize the literature in this field and accelerate the translation of GWAS discoveries into results that are closer to clinical applications. A chronological presentation of published GWAS on lung cancer susceptibility, survival and response to treatment is presented. The most important results are tabulated to provide a concise overview in one read. GWAS have reported 45 lung cancer susceptibility loci with varying strength of evidence and highlighted suspected causal genes at each locus. Some genetic risk loci have been refined to more homogeneous subgroups of lung cancer patients in terms of histological subtypes, smoking status, gender, and ethnicity. Overall, these discoveries are an important step for future development of new therapeutic targets and biomarkers to personalize and improve quality of care for patients. GWAS results are on the edge of offering new tools for targeted screening in high-risk individuals, but more research is needed if GWAS are to pay off the investment. Complementary genomic datasets and functional studies are needed to refine the underlying molecular mechanisms of lung cancer preliminarily revealed by GWAS and reach results that are medically actionable.
Keywords: Lung cancer, genome-wide association study, GWAS, susceptibility loci
Introduction
Lung cancer is the leading cause of cancer-related deaths worldwide in both men and women (1, 2). Our molecular understanding of this disease is in progress. Although it has long been recognized that lung cancer runs strongly in families (3–5), the specific genes that are responsible for enhanced risk are just starting to be revealed. Identifying genes responsible for lung cancer before the era of GWAS has been limited. Candidate susceptibility genes coding for enzymes involved in the activation, detoxification and repair of damages caused by tobacco smoke as well as genes in inflammatory and cell-cycle pathways have been extensively studied (6, 7). Many of these candidate gene studies are either preliminary or controversial (8). Rare germline mutations in TP53, RB1 and EGFR have been shown to confer inherited predisposition to lung cancer (9–11). Fine mapping of genome-wide linkage peak on 6q23–25 also identified RGS17 as a predisposing gene (12). With the arrival of GWAS approximately 10 years ago, it became possible to interrogate the human genome more comprehensively for lung cancer susceptibility genes.
Chronological presentation of published GWAS on lung cancer
GWAS have identified genetic factors robustly associated with lung cancer. Tables 1 and 2 provide a chronological presentation of published GWAS on lung cancer in European and Asian populations, respectively, and summarize susceptibility loci identified. During the last decade, GWAS have evolved from finding lung cancer loci per se, to a more refined search strategy focused on specific subgroups of lung cancer patients. Advances in analysis strategies were also achieved moving from single marker analyses to pathway-based and variant prioritization approaches. GWAS have also been performed to find genetic loci associated with lung cancer survival, response to conventional therapies, and multiple sites of cancer. GWAS by environmental exposures and genome-wide epistasis analyses are also emerging. This review aims to provide a concise overview of this literature. All GWAS on lung cancer susceptibility, survival and response to treatment reported in the literature were cumulated and manually curated by the authors. At the time of writing this review, a search on pubmed with the keywords “GWAS” and “lung cancer” was performed to identify any missing GWAS in the field. Finally, GWAS and susceptibility loci on lung cancer were further refined with the GWAS catalogue (13). Please note that we have attempted to include all loci reported in the literature without quality assessment or exclusion criteria based on the magnitude of effects, sample size or other criteria. Lung cancer susceptibility loci were reported based on the interpretation of the authors in the original manuscripts.
Table 1.
Susceptibility loci for lung cancer identified by GWAS in European ancestry populations
| Reference | Studya | Sample size (cases/controls) | Disease/trait | Platform (# SNPs) | Region (size) | Gene | Key SNPs | |
|---|---|---|---|---|---|---|---|---|
| Hung et al. (14) | IARC Toronto EPIC CARET Liverpool HUNT/Tromsø |
1926/2522 330/453 781/1578 764/1515 403/814 235/392 |
Lung cancer | Illumina
HumanHap300 (310,023) |
15q25 (182 kb) |
CHRNA5 CHRNA3 CHRNB4 IREB2 PSMA4 HYKK (AGPHD1) |
rs1051730 rs8034191 rs16969968 (D398N) |
|
| Thorgeirsson et al. (15) | Icelandic
smokers Icelandic smokers Iceland/Spain/The Netherlands |
10995 2950 1024/32244 |
Smoking
quantity Nicotine dependence Lung cancer |
Illumina HumanHap300 (306,207) |
15q24 |
CHRNA5 CHRNA3 CHRNB4 |
rs1051730 | |
| Amos et al. (16) | Texas Texas replication UK |
1154/1137 711/632 2013/3062 |
Non-small cell lung cancer | Illumina HumanHap300 (315,450) |
15q25.1 (88 kb) |
CHRNA5 CHRNA3 PSMA4 HYKK |
rs1051730 rs8034191 rs931794 |
|
| Liu et al. (17) | GELCC | 194/219 | Familial lung cancer | Affymetrix 500K (399,377) or 6.0 (722,376) | 15q24–25.1 (160 kb) |
CHRNA5 CHRNA3 CHRNB4 IREB2 PSMA4 HYKK |
rs8034191 rs1051730 rs16969968 (D398N) rs578776 |
|
| McKay et al. (19) | Central Europe/ Toronto/
HUNT2-Tromso/ CARET EPIC/Szczecin/ CARET2/ Liverpool |
2971/3746 2899/5573 |
Lung cancer | Illumina
HumanHap300 (315,194) |
15q25.1 | rs1051730 | ||
| 5p15.33 |
TERT CLPTM1L |
rs402710 rs2736100 |
||||||
| Wang et al. (18) | British
cohort IARC Texas UK replication |
1952/1438 1989/2625 1154/1137 2448/2983 |
Lung cancer | Illumina HumanHap550 (511,919) |
15q25.1 | rs8042374 | ||
| 6p21.33 (627 kb) |
BAG6 MSH5 |
rs3117582 rs3131379 |
||||||
| 5p15.33 (60 kb) | CLPTM1L | rs401681 | ||||||
| Broderick et al. (20) | GELCAPS phase
1 GELCAPS phase 2 Meta-analysis (GELCAPS, IARC, Texas) |
1952/1438 2465/3005 7560/8205 |
Lung cancer | Illumina
HumanHap550 (511,919) |
15q25.1 (248 kb) | CHRNA3 | rs12914385 rs938682 rs8042374 rs8034191 |
|
| 5p15.33 (60 kb) |
CLPTM1L TERT |
rs4975616 | ||||||
| 6p21.33 |
BAG6 TNXB |
rs3117582 rs1150752 |
||||||
| Landi et al. (21) | NCI (EAGLE, ATBC, PLCO,
CPS-II) Meta-analysis (UK, Central Europe, Texas, DeCODE Genetics, HGF Germany, CARET, HUNT2/Tromso, Canada, France, Estonia) |
5739/5848 13,300/19,666 |
Lung
cancer Adenocarcinoma Squamous cell Small cell |
Illumina (515,922) | 15q25 |
CHRNA3 CHRNA5 HYKK |
rs12914385 rs1051730 rs8034191 |
|
| 5p15 |
TERT CLPTM1L |
rs2736100 rs4635969 rs31489 |
||||||
| 6p21 |
BAG6 APOM |
rs3117582 | ||||||
| Timofeeva et al. (22) | TRICLb Han Chinese |
14900/29485 2338/3077 |
Lung cancer | Illumina HumanHap300 (318,094) + HumanHap550 or 610Quad (217,914) |
15q25 |
CHRNA5 CHRNA3 CHRNB4 HYKK |
rs1051730 rs8034191 rs6495309 rs680244 rs6495306 rs951266 |
|
| 5p15.33 |
TERT CLPTM1L |
rs2736100 rs401681 rs2853677 rs465498 |
||||||
| 6p21–22 |
BAG6 MSH5 |
rs3117582 rs2523546 rs2523571 |
||||||
| Squamous cell carcinoma | 12p13.33 | RAD52 | rs10849605 rs3748522 |
|||||
| 9p21.3 |
CDKN2A CDKN2B ANRIL (CDKN2B-AS1) |
rs1333040 rs1537372 |
||||||
| 2q32.1 | NUP35 | rs11683501 | ||||||
| Wang et al. (23) | MDACC ICR NCI IARC EPIC ICR IARC Toronto |
11348/15861 10246/38295 |
Lung
cancer, Adenocarcinoma, Squamous cell carcinoma |
Illumina 317, 317+240S, 370Duo, 550, 610 or 1M | 13q13.1 |
BRCA2 FRY |
rs11571833 (K3326X) rs56084662 |
|
| 22q12.1 | CHEK2 | rs17879961 (I157T) | ||||||
| 3q28 | TP63 | rs13314271 rs4488809 |
||||||
| McKay, Hung et al. (25) | OncoArrayc | 29863/55586 | Lung cancer | Oncoarray (10,439,017) | 1p31.1 | FUBP1 | rs71658797 | |
| 6q27 | RNASET2 | rs6920364 | ||||||
| 8p21.1 |
EPHX2 CHRNA2 |
rs11780471 | ||||||
| 13q13.1 | BRCA2 | rs11571833 | ||||||
| 15q21.1 | SEMA6D | rs66759488 | ||||||
| 15q25.1 | CHRNA5 | rs55781567 | ||||||
| 19q13.2 | CYP2A6 | rs56113850 | ||||||
| 11245/54619 | Adenocarcinoma | 3q28 | TP63 | rs13080835 | ||||
| 5p15.33 | TERT | rs7705526 | ||||||
| 8p12 | NRG1 | rs4236709 | ||||||
| 9p21.3 |
MTAP CDKN2A |
rs885518 |
||||||
| 10q24.3 | OBFC1 | rs11591710 | ||||||
| 11q23.3 |
MPZL3 AMICA1 |
rs1056562 | ||||||
| 15q21.1 | SECISBP2L | rs77468143 | ||||||
| 20q13.33 | RTEL1 | rs41309931 | ||||||
| 7704/54763 | Squamous cell carcinoma | 6p21.33 | MHC | rs116822326 | ||||
| 12p13.33 | RAD52 | rs7953330 | ||||||
| 22q12.1 | CHEK2 | rs17879961 | ||||||
| Never smokers | ||||||||
| Li et al. (34) | Mayo MDACC Harvard UCLA |
377/377 328/407 92/161 9¼39 |
Lung cancer in never smokers | Illumina HumanHap370 and HumanHap610 (331,918) | 13q31.3 | GPC5 | rs2352028 rs2352029 |
|
| Pathway-based GWAS | ||||||||
| Shi et al. (40) | NCI UK1 Texas UK2 |
5,355/4,344 592/2699 306/1137 1038/933 |
Lung cancer, Squamous cell carcinoma, Adenocarcinoma, small cell | Illumina (19,082) (pathway-based analysis) | 12p13.33 | RAD52 | rs6489769 | |
| Spitz et al. (41) | Texas Mayo |
451/508 303/311 |
NSCLC in never smokers | Illumina (11,737) (pathway-based analysis) | 12q13 |
ACVR1B NR4A1 |
rs12809597 rs2701129 rs1882119 |
|
| Wang et al. (42) | ICR MDACC IARC NCI Toronto HGF Germany |
12160/16838 | Lung cancer | Illumina (826 functional SNPs) (pathway-based analysis) | 6p21.33 |
MSH5 GTF2H4 |
rs3115672 rs114596632 |
|
| 5q14.2 | XRCC4 | rs1056503 rs2035990 |
||||||
| Variant prioritization approaches | ||||||||
| Li et al. (44) | Texas | 1154/1137 | Lung cancer + intermediate phenotype (cigarettes per day) | Illumina HumanHap300 | 15q24–25.1 |
HYKK
CHRNA3 CHRNA5 CHRNB4 |
rs12914385 | |
| 19q13 |
TGFB1 B9D2 |
rs1800469 rs1982072 rs2241714 |
||||||
| 3p26 | rs1444056 rs1403124 |
|||||||
| Poirier et al. (45) | Toronto IARC MDACC HMGU NCI MSH-PMH MEC Harvard |
33¼99 1964/2610 1154/1137 504/484 5699/5815 1073/939 215/225 52¾97 |
Lung cancer + Family history | Illumina HumanHap300, 550 | 10q23.33 | FFAR4 | rs12415204 | |
| 4p15.2 | KCNIP4 | rs1158970 | ||||||
| Brenner et al. (46) | TRICLb ILCCO |
5061 (SQ)/ 6756 (AD)/ 2216 (SCLC)/
33456 625 (SQ)/ 1417 (AD)/ 369 (SCLC)/ 2966 |
Squamous cell carcinoma | Illumina HumanHap300, 550, 610 | 4p15.2 | KCNIP4 | rs6448050 rs9799795 |
|
| Adenocarcinoma | 18q12.1 | GAREM | rs11662168 rs3786309 |
|||||
| Cross-cancer loci | ||||||||
| Hung et al. (55) | GAME-ON / GECCO | 64591/74467 | Cross-cancer | Illumina,
Affymetrix (12,370) (pathway-based analysis) |
12q24 | SH2B3 | rs3184504 (W262R) | |
| 5p15 | TERT | rs2736100 | ||||||
| Fehringer et al. (56) | GAME-ON /
GECCO Europeans Others |
61851/61820 55789/330490 18152/21410 |
Cross-cancer | Illumina, Affymetrix (9,916,564) | 1q22 |
MUC1 ADAM15 THBS3 |
rs1057941 rs4072037 |
|
| 9p21.3 | CDKN2B-AS1 | rs62560775 | ||||||
| 13q13.1 | BRCA2 | rs11571833 | ||||||
Abbreviations: ATBC: Alpha-Tocopherol, Beta-Carotene Cancer Prevention study; CARET: Beta-Carotene and Retinol Efficacy Trial; CPS-II: Cancer Prevention Study II nutrition cohort; EAGLE: Environment and Genetics in Lung Cancer Etiology; EPIC: European Prospective Investigation in Cancer and Nutrition; GAME-ON: The Genetic Associations and Mechanisms in Oncology; GECCO: The Genetic and Epidemiology of Colorectal Cancer Consortium; GELCAPS: Genetic Lung Cancer Predisposition Study; GELCC: Genetic Epidemiology of Lung Cancer Consortium; HGF Germany: Helmholtz-Gemeinschaft Deutscher Forschungszentren Lung Cancer GWAS; HMGU: Germany study; HUNT: Health Study of North-Trondelag; IARC: International Agency for Research on Cancer; ICR: Institute of Cancer Research; ILCCO: International Lung Cancer Consortium; MDACC: MD Anderson Cancer Centre; MEC: Multi-Ethnic Cohort; MSH-PMH: Mount Sinai Hospital and Princess Margaret Hospital in Toronto; NCI: National Cancer Institute; PLCO: Prostate, Lung, Colon, Ovary screening trial; TRICL: Transdisciplinary Research In Cancer of the Lung; UCLA: University of California in Los Angeles.
Italic indicates replication cohorts.
TRICL includes the following cohorts: MDACC, Liverpool, ICR, Toronto, IARC (Central Europe, CARET, Estonia, France, HUNT2/Tromso), DeCODE Genetics, HGF Germany, Harvard, NCI (EAGLE, ATBC, PLCO, CPS-II).
OncoArray includes the following cohorts: CARET, PLCO, MEC, NELCS (New England Lung Cancer Study), Harvard, MDACC, Tampa (Tampa Lung Cancer Study), BioVU (Vanderbilt 2), LCRI-DOD, TLC (Total Lung Cancer: Molecular Epidemiology of Lung Cancer Survival), Canada (Canadian screening study), MSH-PMH, CAPUA (Cancer de Pulmon en Asturias), ATBC, NSHDC (Northern Sweden Health and Disease Cohort), MDCS (The Malmö Diet and Cancer Study), EPIC, Liverpool, Norway (Norway Lung Cancer Study), EAGLE, Nijmegen (The Nijmegen Lung Cancer Study), NICCC-LCA (Clalit National Israeli Cancer Control Center- lung cancer study), L2 (The IARC L2 Study), Copenhagen (Copenhagen lung cancer study), Germany, ReSoLuCENT.
Table 2.
Susceptibility loci for lung cancer identified by GWAS in Asian ancestry populations
| Reference | Studya | Sample size (cases/controls) | Disease/trait | Platform (# SNPs) | Region (size) | Gene | Key SNPs | |
|---|---|---|---|---|---|---|---|---|
| Yoon et al. (26) | Korea Korea replication |
621/1541 804/1470 |
NSCLC | Affymetrix 5.0 (246,758) | 3q29 | C3orf21 | rs2131877 rs10433328 rs952481 rs4677657 |
|
| 5p15 |
TERT CLPTM1L |
rs2736100 rs402710 rs401681 |
||||||
| Miki et al. (27) | Japanese Japanese Korean |
1004/1900 525/7678 569/1470 |
Lung adenocarcinoma | Illumina HumanHap610-Quad and HumanHap550 (432,024) | 5p15 | TERT | rs2736100 | |
| 3q28 | TP63 | rs10937405 rs4488809 rs9816619 rs4600802 |
||||||
| Hu et al. (28) | Han Chinese (Nanjing, Beijing,
Shanghai) 1st stage replication 2nd stage replication |
2331/3077 2283/2243 4030/4166 |
Lung cancer | Affymetrix 6.0 (591,370) | 3q28 | TP63 | rs4488809 rs10937405 |
|
| 5p15.33 |
TERT CLPTM1L |
rs465498 rs2736100 |
||||||
| 13q12.12 |
MIPEP TNFRSF19 |
rs753955 | ||||||
| 22q12.2 |
MTMR3 HORMAD2 LIF |
rs17728461 rs36600 |
||||||
| Dong et al. (29) | Chinese
(GWAS) Chinese (stage 1) Chinese (stage 2) |
2331/3077 2283/2243 5153/5240 |
Lung cancer | Affymetrix 6.0 (591,370) | 10p14 | GATA3 | rs1663689 |
|
| 5q32 |
PPP2R2B STK32A DPYSL3 |
rs2895680 | ||||||
| 20q13.2 | CYP24A1 | rs4809957 rs2296239 |
||||||
| 5q31.1 |
IL3 CSF2 P4HA2 SLC22A5 ACSL6 |
rs247008 | ||||||
| 1p36.32 |
AJAP1 NPHP4 |
rs9439519 | ||||||
| Shiraishi et al. (32) | Japanese 1st validation 2nd validation |
1695/5333 2955/7036 1379/1166 |
Adenocarcinoma | Illumina OmniExpress & Omni1-Quad (538,166) | 5p15.33 | TERT | rs2736100 rs2853677 |
|
| 3q28 | TP63 | rs10937405 | ||||||
| 17q24.3 | BPTF | rs7216064 | ||||||
| 6p21.3 | BTNL2 | rs3817963 | ||||||
| Dong et al. (30) | Han
Chinese Replication 1 Replication 2 |
833/3094 822/2243 140¼166 |
Squamous cell carcinoma | Affymetrix 6.0 (570,009) | 12q23.1 |
NR1H4 SLC17A8 |
rs12296850 | |
| Jin et al. (31) | Han
Chinese Replication 1 Replication 2 |
1341/1982 1115/1246 3584/3669 |
Lung cancer | Illumina HumanExome (72,423) | 6p21.33 |
PRRC2A
(BAT2) FKBPL |
rs9469031 (P515L) rs200847762 (P137L) |
|
| 20q11.21 | BPIFB1 | rs6141383 (V284M) | ||||||
| 6p22.2 | HIST1H1E | rs2298090 (L152R) | ||||||
| Never smokers | ||||||||
| Hsiung et al. (35) | GELAC (Han
Chinese) GELAC (replication) CAMSCH SNU SWHS WHLCS KNUH KUMC GEL-S NJLCS |
584/585 610/560 287/287 259/293 209/213 207/207 121/119 95/87 193/546 203/203 |
Lung adenocarcinoma in never-smoking Asian females | Illumina HumanCNV370-Duo and HumanHap610 Quad (457,504) | 5p15.33 |
CLPTM1L TERT |
rs2736100 | |
| Lan et al. (36) | Female Lung cancer Consortium in Asia | 5510/4544 1099/2913 |
Lung cancer in never smokers | Illumina (512,226) | 10q25.2 | VTI1A | rs7086803 rs11196080 |
|
| 6q22.2 | ROS1, DCBLD1 | rs9387478 | ||||||
| 6p21.32 | HLA class II region | rs2395185 | ||||||
| 5p15.33 | TERT | rs2736100 | ||||||
| 3q28 | TP63 | rs4488809 | ||||||
| 17q24.3 | BPTF | rs7216064 | ||||||
| Wang et al. (37) | Female Lung cancer Consortium in Asia | 6877/6277 5878/7046 |
Lung cancer in never smokers | Illumina (7,564,751) | 6p21.1 |
FOXP4 FOXP4-AS1 |
rs7741164 | |
| 9p21.3 |
CDKN2B CDKN2B-AS1 |
rs72658409 | ||||||
| 12q13.13 | ACVR1B | rs116101143 | ||||||
| Ahn et al. (38) | Korean | 446/497 434/1000 |
NSCLC in never smokers | Affymetrix 6.0 (474,503) | 18p11.22 |
FAM38B (PIEZO2) APCDD1 NAPG |
rs11080466 rs11663246 |
|
| Kim et al. (39) | Korean Replication 1 Replication 2 |
285/1455 29¾95 546/744 |
Lung cancer in never smoker women | Affymetrix 5.0 (331,088) | 2p16.3 | NRXN1 | rs10187911 | |
| Genome-wide epistasis | ||||||||
| Chu et al. (52) | Han
Chinese Replication 1 Replication 2 |
2331/3077 1534/1489 2512/2449 |
Lung cancer | Affymetrix 6.0 (591,370) (epistatis) | 2q32.2 |
HIBCH INPP1 PMS1 STAT1 |
rs2562796 rs16832404 |
|
| Cross-cancer loci | ||||||||
| Jin et al. (54) | Han
Chinese Han Chinese |
5368/4006 9001/11436 |
Cross-cancer | Affymetrix 6.0 | 6p21.1 | LRFN2 | rs2494938 | |
| 7p15.3 |
SP4 DNAH11 |
rs2285947 | ||||||
Abbreviations: CAMSCH: Chinese Academy of Medical Sciences Cancer Hospital Study; GELAC: Genetic Epidemiological Study of Lung Adenocarcinoma; GEL-S: Genes and Environment in Lung Cancer, Singapore study; KNUH: Kyungpook National University Hospital Study; KUMC: Korea University Medical Center Study; NJLCS: Nanjing Lung Cancer Study; SNU: Seoul National University study; SWHS: Shanghai Women’s Health Cohort Study; WHLCS: Wuhan Lung Cancer Study.
Italic indicates replication cohorts.
GWAS on lung cancer susceptibility
GWAS in European populations.
The first GWAS on lung cancer were reported in 2008. Three independent studies identified a susceptibility locus on chromosome 15q. Hung et al. (14) found two SNPs strongly associated with lung cancer on chromosome 15q25. Further genotyping in this region revealed many SNPs in tight linkage disequilibrium (LD) showing evidence of association. Six genes are located in this region including three nicotinic acetylcholine receptor subunits (CHRNA5, CHRNA3, and CHRNB4). Interestingly, no appreciable variation in the risk was found across smoking categories or histological subtypes of lung cancer. In a second GWAS, a SNP within the CHRNA3 gene was strongly associated with smoking quantity and nicotine dependence (15). The same SNP was also strongly associated with lung cancer. The results suggest that the variant on chromosome 15q25 confers risk of lung cancer through its effect on tobacco addiction. In contrast, a third study showed weak evidence that the 15q25 locus influences smoking behavior and is mostly directly associated with lung cancer (16). However, it should be emphasized that the later GWAS was conducted in cases and controls matched on smoking status, thus limiting variation between the two groups and the power to detect any smoking association. Further analyses from the same study suggest that SNPs and smoking have independent effects on risk. Together, these three studies unequivocally support the 15q25 locus as harboring susceptibility variants for lung cancer or smoking behavior.
A GWAS performed in familial lung cancer confirmed the susceptibility locus on 15q24–25.1 (17). A subsequent GWAS identified two newly associated risk loci for lung cancer (18). In this study, 15q25 was again the most strongly associated locus. However, by pooling the results with other studies (14, 16), new cancer risk loci were found. Two intronic SNPs located in different genes (BAG6, previously known as BAT3, and MSH5) and separated by more than 600 kilobases on chromosome 6p21 were significantly associated with lung cancer. The strongest association, aside 15q25 and 6p21, was found on chromosome 5p15 within the CLPTM1L gene. The 5p15 locus was further supported by an expanded GWAS from previous populations (19). Two uncorrelated SNPs in that region were strongly associated with lung cancer. These SNPs are located within or in proximity to two biologically relevant genes namely CLPTM1L and TERT. Together, by the end of 2008, three susceptibility loci for lung cancer were identified, i.e. 15q25, 6p21 and 5p15.
A more extensive follow-up on a previous GWAS (18) further supports the contribution of the three loci (20). The latter study supports the possibility that two independent loci are acting on 15q25. The latter locus was also associated with smoking behavior, with risk alleles correlated with higher tobacco consumption. In contrast, the 5p15 and 6p21 loci were not associated with smoking behavior. However, DNA variants at 5p15 were associated with histological subtypes of lung cancer, with an increased frequency of the risk allele in cases with adenocarcinoma. This observation was subsequently confirmed in a meta-analysis published in 2009 (21). This study provides compelling evidence that the 5p15 susceptibility locus for lung cancer is confined to a more specific subtype of lung cancer, i.e. adenocarcinoma. Particularly intriguing in that study of more than 30,000 subjects is the absence of new genomic regions associated with lung cancer. In 2010, a meta-analysis of 16 GWAS confirmed lung cancer loci on 15q25, 5p15, and 6p21 (22). Again, the association at 5p15 was confined to adenocarcinoma, whereas the 6p21 locus was more strongly associated with squamous cell carcinoma. Stratification by histology identified three loci for squamous cell carcinoma including 12q13.33, 9p21.3, and 2q32.1. In 2014, another GWAS meta-analysis taking advantage of the imputation based on the 1000 Genomes Project was performed (23), which allowed testing for less frequent SNPs not measured in earlier studies. The top nine signals were followed-up and rare genetic variants were associated with squamous cell carcinoma in the BRCA2 gene on 13q13.1 and in CHEK2 on 22q12.1. CHEK2 was previously associated with lung cancer (24), but this was the first time using a GWAS approach. The 3q28 locus was associated with lung adenocarcinoma, which has been previously found in Asian populations (see below). Finally, the latest and largest lung cancer GWAS in individuals of European ancestry was performed in 29,266 cases and 56,450 controls (25). This GWAS highlighted the genetic heterogeneity across histological subtypes of lung cancer and reported novel loci for lung cancer per se (1p31.1, 6q27, 8p21.1, and 15q21.1) and adenocarcinoma (8p12, 10q24.3, 11q23.3, and 20q13.33). Previously reported lung cancer loci were more specifically associated with squamous cell carcinoma in this study including 6p21.33, 12p13.33, and 22q12.1.
GWAS in Asian populations.
Genetic heterogeneity in lung cancer susceptibility is observed between populations of European and Asian descent. For example, the strongest lung cancer susceptibility variants on 15q25 have very low allele frequencies in Asian populations. Similarly, variants on 6p21 found in Europeans are not polymorphic in Asians. Accordingly, GWAS specific for Asian populations were required.
In a Korean population, a GWAS on NSCLC revealed a new locus on chromosome 3q29 (26). This study also confirmed the 5p15 susceptibility locus in Koreans. Other GWAS in Asian populations have followed. Two susceptibility loci were identified in Japanese and Korean populations confirming 5p15 and elucidating a new locus on 3q28 (27). The 5p15 and 3q28 were subsequently confirmed in a larger GWAS in Han Chinese (28). In addition, two new loci on 13q12.12 and 12q12.2 were identified. In the same GWAS, but with an extended validation sample size, five new lung cancer loci were identified including 10p14, 5q32, 20q13.2, 5q31.1, and 1p36.32 (29). A subsequent GWAS specifically for lung squamous cell carcinoma in Han Chinese revealed a new locus on 12q23.1 (30). Using the exome genotyping chip, rare variants on 6p21.33, 20q11.21, and 6p22.2 were also found in the Chinese populations (31). On 6p21.33, two missense variants, one in PRRC1A (also known as BAT2) and the other in FKBPL, were independently associated with the risk of lung cancer, suggesting more than one genetic signal in this region. This study also demonstrated that 6p21.33 is also a susceptibility locus for Asian populations, but with different risk variants. The aforementioned GWAS in Japanese population (27) was later expanded in terms of sample size and SNP coverage to identify a new locus on 17q24.3 (32). The association with lung adenocarcinoma was also confirmed for 5p15, 3q28, and 6p21, but not for 13q12.12 and 22q12.2.
GWAS in never smokers.
Lung cancer in never smokers is known to be a distinct entity (33). The first GWAS on lung cancer in never smokers was reported in 2010 (34). A single locus on chromosome 13q31.3 was identified. The lung cancer-associated SNPs were located in the GPC5 gene and were also associated with mRNA expression levels of this gene in human lung tissues. A subsequent GWAS was performed in never-smoking females from Asia (35). The 5p15 locus was confirmed with an effect size greater than the estimates reported in populations of European background. Interestingly, the 15q25 and 6p21 loci were not associated with lung cancer in this study and no new loci were identified. In a larger GWAS of the same population forming the Female Lung Cancer Consortium in Asia, new susceptibility loci were revealed at 10q25.2 and 6q22.2 (36). The 6p21 was also associated with lung cancer in this study, but significant markers were not in LD with those previously reported, suggesting again more than one independent genetic signal at 6p21. This study in Asian females also confirmed other loci reported before including 5p15, 3q28, and 17q24.3. A recent meta-analysis was reported with an extended sample size of the Female Lung Cancer Consortium in Asia (37). A new locus on 12q13.13 was identified as well as genetic variants not correlated with lung cancer-SNPs previously associated with lung cancer on 6p21.1 and 9p21.3. In never smokers from Korea, a new NSCLC locus on 18p11 was identified (38). The 2p16.3 locus was also suggested in non-smoking Korean women (39). However, previous loci identified in never smoker populations were not replicated in these Korean studies including 5p15 and 13q31.3.
Pathway-based GWAS.
Pathway-based analyses have been used to identify lung cancer loci. Using GWAS data, genes listed under the category of inflammation were evaluated by lung cancer histological subtypes (40). This analysis identified a risk locus on chromosome 12p13.33 harboring the RAD12 gene. A similar approach was used to evaluate SNPs in inflammatory-pathway genes in lifetime never smokers (41). SNPs on chromosome 12q13 in the ACVR1B and NR4A1 genes were associated with lung cancer, particularly in women and those who reported environmental tobacco smoke exposure. Focused on DNA repair genes, a recent study revealed variants in GTF2H4 on 6p21 and in XRCC4 on 5q14.2 associated with lung cancer risk (42). Pathway-based analyses of GWAS data have also identified groups of genes linked by known biological pathways (ABC transporters, VEGF signaling, G1/S check point, and NRAGE signals death through JNK) that were modestly, but coordinately associated with the risk of developing lung cancer (43).
Variant prioritization approaches.
New lung cancer loci were also revealed by incorporating an intermediate phenotype, i.e. smoked cigarettes per day, into the analyses (44). By combining the estimates derived from the case-control analysis and the intermediate phenotype, a stronger signal was observed on 15q25 locus compared to the case-control study alone. Genetic associations with lung cancer were also detected on 19q13 and 3p26, which demonstrated improved power to identify genetic loci by combining different types of data from a single population. Studying cohorts of patients well-characterized for lung cancer may thus be very promising using this approach. A similar approach was used by assigning higher priors to SNPs associated with family history of lung cancer (45). By focusing on SNPs missed by traditional GWAS, this study identified 30 variants that showed evidence of association with lung cancer risk. The strongest associations were found on 10q23.33 and 4p15.2. Biological priors within a Bayesian framework were also applied to histology-specific analyses (46). In this study, the 4p15.2 locus was assigned more specifically to squamous cell carcinoma and a new adenocarcinoma locus was identified on 18q12.1.
GWAS-by-exposure interaction.
Accounting for environmental exposure is challenging owing to the large number of possible factors as well as the level of measurement accuracy that can be achieved for each exposure. Despite these challenges, some genome-wide gene-environment interaction studies are starting to emerge in the field of lung cancer. The first attempt of a genome-wide gene-smoking interaction study identified two SNPs on 14q22.1 and 15q22.32 influencing the risk of lung cancer (47). For asbestos exposure, interacting loci were suggested on 2q34, 7q32.1, and 11q13 (48). A risk locus for asbestos-associated lung cancer was also discovered on 22q13.31 (49). Interacting loci with household air pollution caused by solid fuel burning for heating and cooking were also evaluated in never smoker women from Asia (50). Interestingly, interactions were reported for GWAS-nominated loci previously identified in this population (36), but no new loci reached significance at the genome-wide scale level. Exploratory analyses of gene-occupation interactions in determining lung cancer susceptibility were also performed for 17 established or suspected lung carcinogens and 49 additional occupational agents (51). A large number of gene-environment interactions were reported in that study. However, the results could not be validated in an independent population because of the uniqueness of the dataset with detailed occupational exposure data. So far, results from genome-wide gene-environment studies in lung cancer have been more hypothesis-generating owing to limited sample size and power as well as the lack of appropriate replication sets. To make further progress, extra care will be needed to build large cohorts that are well-characterized for environmental exposures.
Genome-wide epistasis.
The effects of genetic variants on lung cancer are likely to be amplified when multiple variants synergize together. Gene-gene interactions may identify genetic determinants of lung cancer. The first and only genome-wide two-locus interaction analysis performed so far revealed a significant interaction between two SNPs 60 kilobases apart on 2q32.2 (52). Individually, the two interacting SNPs were not significantly associated with the risk of lung cancer. Further investigations of gene-gene interactions will be needed to understand the genetic architecture of lung cancer.
Cross-cancer susceptibility loci.
Large-scale GWAS across cancer sites have been conducted to identify pleiotropic loci. For lung cancer, the first pleiotropic locus was identified on 5p15 (TERT-CLPTM1L) (53). A novel pleiotropic association at 7p15.3 was found in Han Chinese involving lung cancer, non-cardia gastric cancer, and esophageal squamous-cell carcinoma (54). The GAME-ON/GECCO Network on lung, ovary, breast, prostate and colorectal cancer then identified novel pleiotropic associations involving lung cancer on 12q24 (55) and 1q22 (56). Known lung cancer loci were also identified in cross-cancer analyses including 6p21 (54) and 5p15 (55) as well as 9p21.3 and 13q13.1 (56). These loci are particularly promising to reveal shared carcinogenesis mechanisms across multiple cancer sites.
Integration of GWAS on lung cancer susceptibility
Excluding gene-environment loci that are more suggestive at this point, GWAS reported 45 loci associated with lung cancer. Figure 1 shows the chronological and cumulative number of lung cancer susceptibility loci identified. Loci are also listed based on chromosome number in Table 3. Note that these loci are an evolving list. The strength of evidence for association with lung cancer and effect size vary by loci (Figure 2). Evidence supporting some loci is relatively modest and will require validation in independent studies. The magnitude of genetic associations reported in publications also varies within loci. For example, the largest OR for chromosome 15q25 was 7.2 reported in familial form of lung cancer with a relatively modest p value of 1.03 × 10−3 (17). On the other hand, the same locus was reported highly significant (p = 3.08 × 10−103) with an OR of ~1.3 (25), which is an effect size more consistent with most studies on sporadic form of lung cancer. The maximum odds ratio and p value per locus as well as variability between studies are illustrated in Figure 2. Refining susceptibility loci by clinically relevant subgroups is a critical step to reveal functional variants and causative genes (57). Table 3 summarizes the evidence supporting the specificity of lung cancer risk loci by histology, smoking status, gender, ethnicity, and age of onset. For example, convincing evidence supports that the 5p15 locus is specific to lung adenocarcinoma and more strongly associated with never smokers and women (21, 22, 28). Similar genetic association patterns are also emerging for 2p16.3, 5q32, and 6q22. Accordingly, a number of studies have started to delineate the effects of genetic variants in specific subgroups of patients with lung cancer, which is important to reveal the true nature of genetic effects detected in GWAS and narrow the set of genetic variants and genes worthy of functional studies. It is also important to know whether independent variants in the same loci are associated with lung cancer. Convincing evidence supports at least two independent loci on 15q25, 5p15, and 6p21 (Table 3). Two independent loci were also reported at 9p21.3 (37) and 22q12.2 (28), but will require further validation. This knowledge is lacking for other lung cancer loci. Table 3 also provides a glimpse of suspected causal genes at each locus. Further functional and biological analyses will be needed to understand the role of these genes in lung cancer development.
Figure 1.

Chronological and cumulative lung cancer susceptibility loci identified by GWAS. New loci identified yearly are on a black background. The cumulative number of loci is shown with grey background.
Table 3.
Lung cancer susceptibility loci derived from GWAS, genetic risk specificity by subgroups, and suspected causal genes
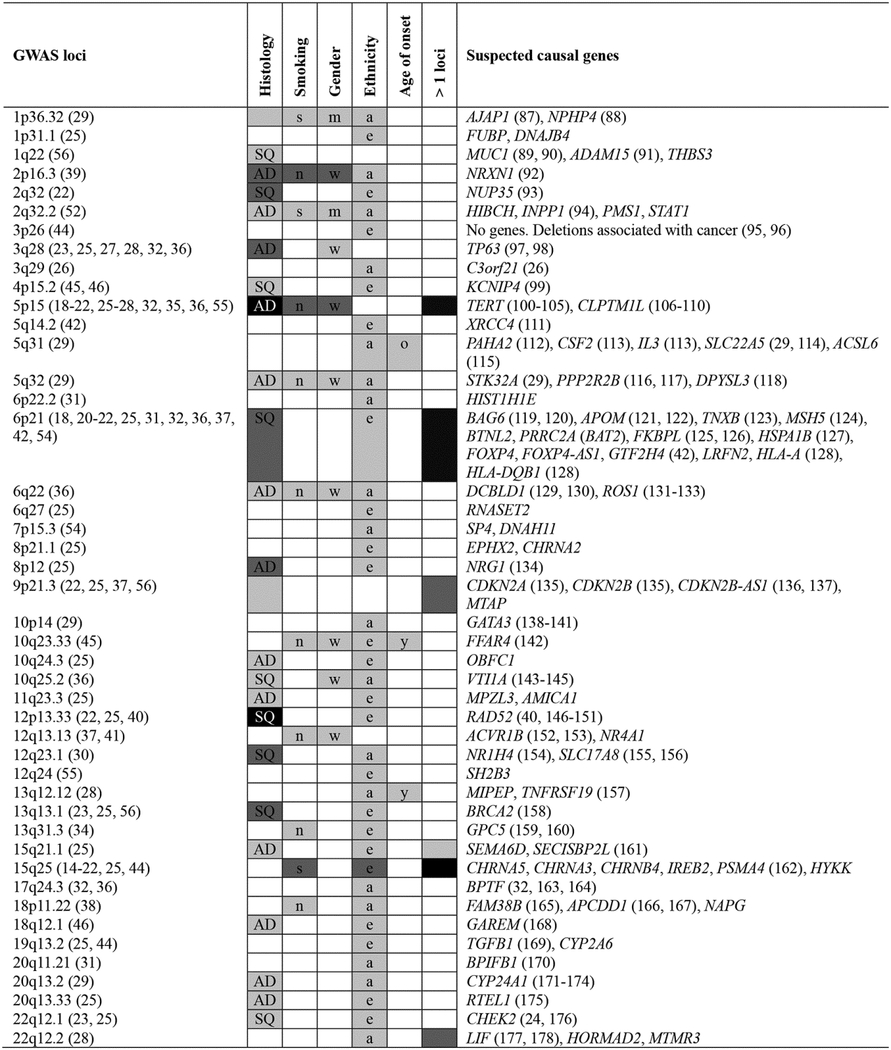
|
Abbreviations: AD: adenocarcinoma; a: Asians; e: Europeans; m: men; n: never-smokers; o: older; s: smokers; SQ: squamous cell carcinoma; w: women; y: younger. The color of the background illustrates the strength of evidence on a black and white scale where black indicates convincing evidence and white indicates no evidence so far. The strength of evidence was assigned based on the content of publications cited in the first column and our best possible judgment and comprehension of each locus considering the number of studies that replicated the associations, the level of statistically significance, and the quality of the studies, e.g. sample size. No evidence (white) does not delineate the lack of associations that have been studied from those not yet examined and also highlights knowledge gaps.
Figure 2.

Magnitude of the genetic associations for the 45 lung cancer susceptibility loci reported in the literature. P values and odds ratio for key SNPs indicated in Tables 1 and 2 were collected from reported studies. The upper and lower panels show p values and odds ratio, respectively. The x-axis shows the 45 lung cancer susceptibility loci ordered based on chromosome position. Bars illustrate the maximum p value (smallest p values) or the maximum OR reported per locus. Tick symbols inside bars are the results of the individual studies. For the upper panel, the y-axis represents p values in -log10 scale. The dashed horizontal line shows the typical GWAS significance threshold, i.e. 5 × 10−8. For the lower panel, the y-axis represents odds ratio. Note that OR lower than 1 were converted into their reciprocal (1/OR) for illustration purpose. OR for locus 3p26 was not available.
GWAS on lung cancer survival
Inter-individual differences in lung cancer survival are observed among lung cancer patients, even among those with the same tumor stage and treatment regimen. The identification of genetic factors associated with lung cancer survival has the potential to guide adjuvant therapy after surgery in early-stage disease, but also to refine prognosis and personalize clinical care in advanced-stages disease. So far, GWAS on lung cancer survival were performed in patients with early-stage NSCLC (58, 59), advanced-stage NSCLC (60–63), and SCLC (64). GWAS were also performed in more specific subgroups of lung cancer patients including never smokers with NSCLC (65) and patients with lung adenocarcinoma (66). Together, these studies have identified 23 loci associated with lung cancer survival (Supplementary Table S1). However, none of these loci was reported in more than one study. The lack of replication may be explained by heterogeneity in treatment regimens. There is also no overlap with GWAS lung cancer susceptibility loci. While the 9p21.3 locus was associated with both susceptibility (22, 37) and survival (62), sentinel SNPs are located more than 1 Mb away from each other, indicating that they are likely not reflecting the same association. GWAS susceptibility loci were specifically evaluated for association with survival in SCLC (67). Briefly, three loci on 20q13.2, 22q12.2, and 5p15 demonstrated some evidence of association with survival. However, none reached genome-wide significance. It should be noted that clinical follow-up of patients are needed to conduct survival analyses and GWAS based on this outcome have thus been performed with much smaller sample sizes compared to studies focused on cancer susceptibility. Larger-scale studies are needed to identify robust lung cancer survival loci.
GWAS on response to lung cancer therapies
Somatic alterations in the tumor genome are known to modulate the response to anticancer therapy. Less is known about the influence of the host genome on treatment response. The effect of germline variants on sensitivity and toxicity to platinum-based chemotherapy have been examined by GWAS. In patients with SCLC, seven loci demonstrated some evidence of association with treatment response in a discovery set, but were not convincingly replicated in a validation set (68). In NSCLC patients, a locus on 21q22.3 was associated with platinum-induced hepatotoxicity (69) and two loci on 2q24.3 and 17p12 were associated with the risk of platinum-induced myelosuppression (70). More and larger studies are needed to effectively delineate chemosensitive patients that will benefit from treatment and nonresponders that may be spared the adverse side effects associated with chemotherapy.
Future directions
Important progress was made to understand host susceptibility to lung cancer using GWAS. This approach is also starting to reveal inherited variants associated with lung cancer survival and response to treatment. During the last decade, progress was driven by enlarging sample sizes, improving methods to genotype and impute SNPs with more comprehensive reference sets (e.g. 1000 Genomes Project), and the creation of large-scale international collaborations and consortia. Additional developments were made by studying patients of different ancestries, never smokers, women, and specific lung cancer histology. Progress was also made by refining results by pathway-based analysis and variant prioritization approaches. The next important step in the field of genomics of lung cancer is to identify the causal genetic variants and genes underpinning GWAS-nominated loci. In addition, the new genomic knowledge must be translated into real benefits for patients. These must be achieved if GWAS are to pay off the huge investment. We foresee different strategies to reach these goals.
eQTL & TWAS.
We need to continue to mine GWAS data using more advanced statistical techniques that leverage other sources of data. So far, expression quantitative trait loci (eQTL) mapping studies in a variety of tissues have been used to extent the functional meaning of GWAS in lung cancer (23, 25, 29, 30, 32, 34, 37, 45, 56, 71). More comprehensive methods of colocalisation of GWAS and eQTL signals were recently developed and must be performed to reveal genetic associations explained by regulatory effects on gene expression (72). The identification of lung cancer-associated genetic variants associated with the expression of specific genes in a disease relevant tissue is an important step forward to understand the molecular mechanisms underpinning GWAS signals. In addition, the relationships between genetic variants, RNA expression levels and lung cancer must be further delineated by causality models and Mendelian randomization approaches (73, 74). Large-scale lung cancer GWAS (25) and lung eQTL (75) are also available to perform the first transcriptome-wide association study (TWAS) in lung cancer. In this approach, the cis genetic component of expression derived from the eQTL dataset is used to impute expression data for cases and controls used in the GWAS. Imputed genome-wide gene expression levels of sample size orders of magnitude larger than any of the transcriptomic datasets generated so far can then be used to identify genes whose expression is significantly associated with the disease. This approach has the potential to elucidate the most likely molecular drivers of lung cancer in GWAS-nominated loci, but also yield molecular drivers of lung cancer outside GWAS loci. GWAS, eQTL and TWAS results will also need to be integrated with genes differentially expressed in lung tumor compared to adjacent non-tumor lung tissues. For example, we have recently derived a robust list of genes differentially expressed in lung tumor from our own transcriptomic dataset (76) as well as two publically available datasets (77, 78). These results identified genes consistently deregulated in lung tumor and revealed important insights about the molecular transitions that occur between normal and tumor lung tissues. Accordingly, very promising research is underway exploiting GWAS and gene expression datasets to identify causal genes and molecular drivers of lung cancer.
Deep molecular profiling and biobanking.
To make further progress, relevant tissues must be profiled beyond gene expression. GWAS variants of lung cancer may not exert their effects through gene regulation, but other molecular phenotypes such as protein expression, protein state, metabolite levels, and epigenetic marks. Accordingly, deep molecular profiling of human lung tissues will be needed to comprehend the molecular impact of inherited variants on lung cancer. Current biobanking activities to collect high-quality and large numbers of well-annotated lung specimens are the essence of this future development.
Exposome.
As depicted in Table 3, the independent contribution of GWAS loci on lung cancer, smoking behavior and nicotine dependence is still not clearly delineated. Larger-scale gene-exposure interaction studies with established environmental risk factors including tobacco smoke and solid fuel burning are warranted. More comprehensive assessment of environmental factors including radon, asbestos, household and outdoor pollution, and occupational agents will be critically important, but at the same time very challenging to measure accurately in large sample size. A well-orchestrated community effort thus seems necessary (79).
Exome and genome sequencing.
New genomic approaches from next-generation of sequencers are also expected to refine GWAS loci and discover new variants unlikely to be found by GWAS. To this effect, whole-exome sequencing in three members of a five-generation family affected by lung cancer has revealed a rare variants in PARK2 resulting in a loss-of-function of this tumor suppressor gene (80). Although rare, the effect size of this mutation was greater than those reported in GWAS. Similarly, exome-sequencing of sporadic and familial cases of lung cancer identified rare deleterious mutations in GWAS-nominated loci located in the CDC147 and DBH (81) genes. Whole-genome sequencing in a family with very high aggregation of lung adenocarcinoma revealed a functional missense variant in the oncogene YAP1 (82) associated with the risk of developing the disease. We expect this type of discoveries using exome and genome sequencing to multiply in a near future.
En route for a genetic risk score.
Identified lung cancer susceptibility loci provide hope to build tools for targeted screening of high-risk individuals. To date, cumulative effects of loci have shown promising results to improve the discriminatory performance of risk prediction models, but not sufficiently to merit clinical implementation (83). For example, a recent report combining GWAS loci demonstrated only small improvement in lung cancer risk prediction in models including basic clinical factors such as age and smoking (84). Interestingly, the best model may not come from considering only the top GWAS loci. A genetic risk score built from seven telomere-length associated genetic variants was associated with lung cancer risk (85). More recently, it was demonstrated that the cumulative effects of susceptibility variants were better predictors when organized in biological pathways (86). These examples demonstrated the variety of strategies that are currently used to develop new clinical tools to predict lung cancer. Such tools are urgently needed to enable earlier diagnosis. The task is challenging and will require major efforts, but seems more realistically feasible with the outcomes of GWAS in hands. We hope that this compendium of lung cancer GWAS loci will facilitate further progress in building a clinically useful genetic risk score.
Conclusions
Understanding the genetic factors underlying the development of lung cancer is important to elucidate the etiology of the disease. This genetic knowledge is a prerequisite to develop and improve future clinical strategies for lung cancer management. Discovered loci summarized in this review testify progress made in this field during the last decade. This review also highlights knowledge gaps about causal variants and genes responsible for the underlying genetic associations and proposes some short term solutions to ensure further progress through eQTL, colocalization, causality models and TWAS. The specificity of many lung cancer loci in terms of histological subtypes, gender and ethnicity have been discovered for some loci, but will demand large studies with well-characterized individuals for others. Although smoking and other environmental factors, notably solid fuel burning, are clearly interacting with host factors to cause the disease, the specific variants that come into play are still elusive. Preliminary data provides some clues about inherited variants associated with lung cancer survival and response to treatment, but will require validation in larger-scale studies. On the other hand, robust genetic factors associated with lung cancer derived from GWAS give hope for possible clinical translation. In short term, a genetic risk score to screen high-risk individuals seems realistically achievable and would allow more effective treatments available at earlier stages of the disease. In mid and longer terms, discovering the causal genes underpinning GWAS signals will propel results one step closer to clinical applications by revealing new therapeutic targets and biomarkers to personalize quality of care.
Supplementary Material
Financial support
Y. Bossé holds a Canada Research Chair in Genomics of Heart and Lung Diseases.
Abbreviation lists
- eQTL
expression quantitative trait loci
- GWAS
genome-wide association study
- LD
linkage disequilibrium
- Mb
megabase
- NSCLC
non-small cell lung cancer
- SCLC
small cell lung cancer
- SNP
single nucleotide polymorphism
- TWAS
transcriptome-wide association study
References
- 1.Herbst RS, Heymach JV, Lippman SM. Lung cancer. N Engl J Med. 2008;359:1367–80. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 2.Siegel R, Ma J, Zou Z, Jemal A. Cancer statistics, 2014. CA Cancer J Clin. 2014;64:9–29. [DOI] [PubMed] [Google Scholar]
- 3.Matakidou A, Eisen T, Houlston RS. Systematic review of the relationship between family history and lung cancer risk. Br J Cancer. 2005;93:825–33. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 4.Tokuhata GK, Lilienfeld AM. Familial aggregation of lung cancer in humans. J Natl Cancer Inst. 1963;30:289–312. [PubMed] [Google Scholar]
- 5.Cote ML, Liu M, Bonassi S, Neri M, Schwartz AG, Christiani DC, et al. Increased risk of lung cancer in individuals with a family history of the disease: a pooled analysis from the International Lung Cancer Consortium. Eur J Cancer. 2012;48:1957–68. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 6.Schwartz AG, Prysak GM, Bock CH, Cote ML. The molecular epidemiology of lung cancer. Carcinogenesis. 2007;28:507–18. [DOI] [PubMed] [Google Scholar]
- 7.Yokota J, Shiraishi K, Kohno T. Genetic basis for susceptibility to lung cancer: Recent progress and future directions. Adv Cancer Res. 2010;109:51–72. [DOI] [PubMed] [Google Scholar]
- 8.Li W, Li K, Zhao L, Zou H. DNA repair pathway genes and lung cancer susceptibility: a meta-analysis. Gene. 2014;538:361–5. [DOI] [PubMed] [Google Scholar]
- 9.Hwang SJ, Cheng LS, Lozano G, Amos CI, Gu X, Strong LC. Lung cancer risk in germline p53 mutation carriers: association between an inherited cancer predisposition, cigarette smoking, and cancer risk. Hum Genet. 2003;113:238–43. [DOI] [PubMed] [Google Scholar]
- 10.Kleinerman RA, Tarone RE, Abramson DH, Seddon JM, Li FP, Tucker MA. Hereditary retinoblastoma and risk of lung cancer. J Natl Cancer Inst. 2000;92:2037–9. [DOI] [PubMed] [Google Scholar]
- 11.Bell DW, Gore I, Okimoto RA, Godin-Heymann N, Sordella R, Mulloy R, et al. Inherited susceptibility to lung cancer may be associated with the T790M drug resistance mutation in EGFR. Nat Genet. 2005;37:1315–6. [DOI] [PubMed] [Google Scholar]
- 12.You M, Wang D, Liu P, Vikis H, James M, Lu Y, et al. Fine mapping of chromosome 6q23–25 region in familial lung cancer families reveals RGS17 as a likely candidate gene. Clin Cancer Res. 2009;15:2666–74. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 13.Welter D, MacArthur J, Morales J, Burdett T, Hall P, Junkins H, et al. The NHGRI GWAS Catalog, a curated resource of SNP-trait associations. Nucleic Acids Res. 2014;42:D1001–6. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 14.Hung RJ, McKay JD, Gaborieau V, Boffetta P, Hashibe M, Zaridze D, et al. A susceptibility locus for lung cancer maps to nicotinic acetylcholine receptor subunit genes on 15q25. Nature. 2008;452:633–7. [DOI] [PubMed] [Google Scholar]
- 15.Thorgeirsson TE, Geller F, Sulem P, Rafnar T, Wiste A, Magnusson KP, et al. A variant associated with nicotine dependence, lung cancer and peripheral arterial disease. Nature. 2008;452:638–42. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 16.Amos CI, Wu X, Broderick P, Gorlov IP, Gu J, Eisen T, et al. Genome-wide association scan of tag SNPs identifies a susceptibility locus for lung cancer at 15q25.1. Nat Genet. 2008;40:616–22. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 17.Liu P, Vikis HG, Wang D, Lu Y, Wang Y, Schwartz AG, et al. Familial aggregation of common sequence variants on 15q24–25.1 in lung cancer. J Natl Cancer Inst. 2008;100:1326–30. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 18.Wang Y, Broderick P, Webb E, Wu X, Vijayakrishnan J, Matakidou A, et al. Common 5p15.33 and 6p21.33 variants influence lung cancer risk. Nat Genet. 2008;40:1407–9. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 19.McKay JD, Hung RJ, Gaborieau V, Boffetta P, Chabrier A, Byrnes G, et al. Lung cancer susceptibility locus at 5p15.33. Nat Genet. 2008;40:1404–6. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 20.Broderick P, Wang Y, Vijayakrishnan J, Matakidou A, Spitz MR, Eisen T, et al. Deciphering the impact of common genetic variation on lung cancer risk: a genome-wide association study. Cancer Res. 2009;69:6633–41. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 21.Landi MT, Chatterjee N, Yu K, Goldin LR, Goldstein AM, Rotunno M, et al. A genome-wide association study of lung cancer identifies a region of chromosome 5p15 associated with risk for adenocarcinoma. Am J Hum Genet. 2009;85:679–91. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 22.Timofeeva MN, Hung RJ, Rafnar T, Christiani DC, Field JK, Bickeboller H, et al. Influence of common genetic variation on lung cancer risk: meta-analysis of 14 900 cases and 29 485 controls. Hum Mol Genet. 2012;21:4980–95. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 23.Wang Y, McKay JD, Rafnar T, Wang Z, Timofeeva MN, Broderick P, et al. Rare variants of large effect in BRCA2 and CHEK2 affect risk of lung cancer. Nat Genet. 2014;46:736–41. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 24.Brennan P, McKay J, Moore L, Zaridze D, Mukeria A, Szeszenia-Dabrowska N, et al. Uncommon CHEK2 mis-sense variant and reduced risk of tobacco-related cancers: case control study. Hum Mol Genet. 2007;16:1794–801. [DOI] [PubMed] [Google Scholar]
- 25.McKay JD, Hung RJ, Han Y, Zong X, Carreras-Torres R, Christiani DC, et al. Large scale genetic analysis identifies novel loci and histological variability in susceptibility to lung cancer. Nat Genet. 2017. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 26.Yoon KA, Park JH, Han J, Park S, Lee GK, Han JY, et al. A genome-wide association study reveals susceptibility variants for non-small cell lung cancer in the Korean population. Hum Mol Genet. 2010;19:4948–54. [DOI] [PubMed] [Google Scholar]
- 27.Miki D, Kubo M, Takahashi A, Yoon KA, Kim J, Lee GK, et al. Variation in TP63 is associated with lung adenocarcinoma susceptibility in Japanese and Korean populations. Nat Genet. 2010;42:893–6. [DOI] [PubMed] [Google Scholar]
- 28.Hu Z, Wu C, Shi Y, Guo H, Zhao X, Yin Z, et al. A genome-wide association study identifies two new lung cancer susceptibility loci at 13q12.12 and 22q12.2 in Han Chinese. Nat Genet. 2011;43:792–6. [DOI] [PubMed] [Google Scholar]
- 29.Dong J, Hu Z, Wu C, Guo H, Zhou B, Lv J, et al. Association analyses identify multiple new lung cancer susceptibility loci and their interactions with smoking in the Chinese population. Nat Genet. 2012;44:895–9. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 30.Dong J, Jin G, Wu C, Guo H, Zhou B, Lv J, et al. Genome-wide association study identifies a novel susceptibility locus at 12q23.1 for lung squamous cell carcinoma in han chinese. PLoS Genet. 2013;9:e1003190. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 31.Jin G, Zhu M, Yin R, Shen W, Liu J, Sun J, et al. Low-frequency coding variants at 6p21.33 and 20q11.21 are associated with lung cancer risk in Chinese populations. Am J Hum Genet. 2015;96:832–40. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 32.Shiraishi K, Kunitoh H, Daigo Y, Takahashi A, Goto K, Sakamoto H, et al. A genome-wide association study identifies two new susceptibility loci for lung adenocarcinoma in the Japanese population. Nat Genet. 2012;44:900–3. [DOI] [PubMed] [Google Scholar]
- 33.Sun S, Schiller JH, Gazdar AF. Lung cancer in never smokers--a different disease. Nat Rev Cancer. 2007;7:778–90. [DOI] [PubMed] [Google Scholar]
- 34.Li Y, Sheu CC, Ye Y, de Andrade M, Wang L, Chang SC, et al. Genetic variants and risk of lung cancer in never smokers: a genome-wide association study. Lancet Oncol. 2010;11:321–30. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 35.Hsiung CA, Lan Q, Hong YC, Chen CJ, Hosgood HD, Chang IS, et al. The 5p15.33 locus is associated with risk of lung adenocarcinoma in never-smoking females in Asia. PLoS Genet. 2010;6. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 36.Lan Q, Hsiung CA, Matsuo K, Hong YC, Seow A, Wang Z, et al. Genome-wide association analysis identifies new lung cancer susceptibility loci in never-smoking women in Asia. Nat Genet. 2012;44:1330–5. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 37.Wang Z, Seow WJ, Shiraishi K, Hsiung CA, Matsuo K, Liu J, et al. Meta-analysis of genome-wide association studies identifies multiple lung cancer susceptibility loci in never-smoking Asian women. Hum Mol Genet. 2016;25:620–9. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 38.Ahn MJ, Won HH, Lee J, Lee ST, Sun JM, Park YH, et al. The 18p11.22 locus is associated with never smoker non-small cell lung cancer susceptibility in Korean populations. Hum Genet. 2012;131:365–72. [DOI] [PubMed] [Google Scholar]
- 39.Kim JH, Park K, Yim SH, Choi JE, Sung JS, Park JY, et al. Genome-wide association study of lung cancer in Korean non-smoking women. J Korean Med Sci. 2013;28:840–7. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 40.Shi J, Chatterjee N, Rotunno M, Wang Y, Pesatori AC, Consonni D, et al. Inherited variation at chromosome 12p13.33, including RAD52, influences the risk of squamous cell lung carcinoma. Cancer discovery. 2012;2:131–9. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 41.Spitz MR, Gorlov IP, Amos CI, Dong Q, Chen W, Etzel CJ, et al. Variants in inflammation genes are implicated in risk of lung cancer in never smokers exposed to second-hand smoke. Cancer discovery. 2011;1:420–9. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 42.Wang M, Liu H, Liu Z, Yi X, Bickeboller H, Hung RJ, et al. Genetic variant in DNA repair gene GTF2H4 is associated with lung cancer risk: a large-scale analysis of six published GWAS datasets in the TRICL consortium. Carcinogenesis. 2016;37:888–96. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 43.Lee D, Lee GK, Yoon KA, Lee JS. Pathway-based analysis using genome-wide association data from a Korean non-small cell lung cancer study. PLoS One. 2013;8:e65396. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 44.Li Y, Huang J, Amos CI. Genetic association analysis of complex diseases incorporating intermediate phenotype information. PLoS One. 2012;7:e46612. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 45.Poirier JG, Brennan P, McKay JD, Spitz MR, Bickeboller H, Risch A, et al. Informed Genome-Wide Association Analysis With Family History As a Secondary Phenotype Identifies Novel Loci of Lung Cancer. Genet Epidemiol. 2015;39:197–206. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 46.Brenner DR, Amos CI, Brhane Y, Timofeeva MN, Caporaso N, Wang Y, et al. Identification of lung cancer histology-specific variants applying Bayesian framework variant prioritization approaches within the TRICL and ILCCO consortia. Carcinogenesis. 2015;36:1314–26. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 47.Zhang R, Chu M, Zhao Y, Wu C, Guo H, Shi Y, et al. A genome-wide gene-environment interaction analysis for tobacco smoke and lung cancer susceptibility. Carcinogenesis. 2014;35:1528–35. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 48.Wei S, Wang LE, McHugh MK, Han Y, Xiong M, Amos CI, et al. Genome-wide gene-environment interaction analysis for asbestos exposure in lung cancer susceptibility. Carcinogenesis. 2012;33:1531–7. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 49.Liu CY, Stucker I, Chen C, Goodman G, McHugh MK, D’Amelio AM Jr., et al. Genome-wide Gene-Asbestos Exposure Interaction Association Study Identifies a Common Susceptibility Variant on 22q13.31 Associated with Lung Cancer Risk. Cancer Epidemiol Biomarkers Prev. 2015;24:1564–73. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 50.Hosgood HD 3rd, Song M, Hsiung CA, Yin Z, Shu XO, Wang Z, et al. Interactions between household air pollution and GWAS-identified lung cancer susceptibility markers in the Female Lung Cancer Consortium in Asia (FLCCA). Hum Genet. 2015;134:333–41. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 51.Malhotra J, Sartori S, Brennan P, Zaridze D, Szeszenia-Dabrowska N, Swiatkowska B, et al. Effect of occupational exposures on lung cancer susceptibility: a study of gene-environment interaction analysis. Cancer Epidemiol Biomarkers Prev. 2015;24:570–9. [DOI] [PubMed] [Google Scholar]
- 52.Chu M, Zhang R, Zhao Y, Wu C, Guo H, Zhou B, et al. A genome-wide gene-gene interaction analysis identifies an epistatic gene pair for lung cancer susceptibility in Han Chinese. Carcinogenesis. 2014;35:572–7. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 53.Rafnar T, Sulem P, Stacey SN, Geller F, Gudmundsson J, Sigurdsson A, et al. Sequence variants at the TERT-CLPTM1L locus associate with many cancer types. Nat Genet. 2009;41:221–7. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 54.Jin G, Ma H, Wu C, Dai J, Zhang R, Shi Y, et al. Genetic variants at 6p21.1 and 7p15.3 are associated with risk of multiple cancers in Han Chinese. Am J Hum Genet. 2012;91:928–34. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 55.Hung RJ, Ulrich CM, Goode EL, Brhane Y, Muir K, Chan AT, et al. Cross Cancer Genomic Investigation of Inflammation Pathway for Five Common Cancers: Lung, Ovary, Prostate, Breast, and Colorectal Cancer. J Natl Cancer Inst. 2015;107. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 56.Fehringer G, Kraft P, Pharoah PD, Eeles RA, Chatterjee N, Schumacher FR, et al. Cross-Cancer Genome-Wide Analysis of Lung, Ovary, Breast, Prostate, and Colorectal Cancer Reveals Novel Pleiotropic Associations. Cancer Res. 2016;76:5103–14. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 57.Ioannidis JP, Thomas G, Daly MJ. Validating, augmenting and refining genome-wide association signals. Nat Rev Genet. 2009;10:318–29. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 58.Huang YT, Heist RS, Chirieac LR, Lin X, Skaug V, Zienolddiny S, et al. Genome-wide analysis of survival in early-stage non-small-cell lung cancer. J Clin Oncol. 2009;27:2660–7. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 59.Tang S, Pan Y, Wang Y, Hu L, Cao S, Chu M, et al. Genome-wide association study of survival in early-stage non-small cell lung cancer. Annals of surgical oncology. 2015;22:630–5. [DOI] [PubMed] [Google Scholar]
- 60.Sato Y, Yamamoto N, Kunitoh H, Ohe Y, Minami H, Laird NM, et al. Genome-wide association study on overall survival of advanced non-small cell lung cancer patients treated with carboplatin and paclitaxel. Journal of thoracic oncology : official publication of the International Association for the Study of Lung Cancer. 2011;6:132–8. [DOI] [PubMed] [Google Scholar]
- 61.Wu X, Ye Y, Rosell R, Amos CI, Stewart DJ, Hildebrandt MA, et al. Genome-wide association study of survival in non-small cell lung cancer patients receiving platinum-based chemotherapy. J Natl Cancer Inst. 2011;103:817–25. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 62.Hu L, Wu C, Zhao X, Heist R, Su L, Zhao Y, et al. Genome-wide association study of prognosis in advanced non-small cell lung cancer patients receiving platinum-based chemotherapy. Clin Cancer Res. 2012;18:5507–14. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 63.Lee Y, Yoon KA, Joo J, Lee D, Bae K, Han JY, et al. Prognostic implications of genetic variants in advanced non-small cell lung cancer: a genome-wide association study. Carcinogenesis. 2013;34:307–13. [DOI] [PubMed] [Google Scholar]
- 64.Wu C, Xu B, Yuan P, Miao X, Liu Y, Guan Y, et al. Genome-wide interrogation identifies YAP1 variants associated with survival of small-cell lung cancer patients. Cancer Res. 2010;70:9721–9. [DOI] [PubMed] [Google Scholar]
- 65.Wu X, Wang L, Ye Y, Aakre JA, Pu X, Chang GC, et al. Genome-wide association study of genetic predictors of overall survival for non-small cell lung cancer in never smokers. Cancer Res. 2013;73:4028–38. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 66.Galvan A, Colombo F, Frullanti E, Dassano A, Noci S, Wang Y, et al. Germline polymorphisms and survival of lung adenocarcinoma patients: a genome-wide study in two European patient series. International journal of cancer Journal international du cancer. 2015;136:E262–71. [DOI] [PubMed] [Google Scholar]
- 67.Li D, Wei L, Xu B, Yu D, Chang J, Yuan P, et al. Association of GWAS-identified lung cancer susceptibility loci with survival length in patients with small-cell lung cancer treated with platinum-based chemotherapy. PLoS One. 2014;9:e113574. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 68.Wu C, Xu B, Yuan P, Ott J, Guan Y, Liu Y, et al. Genome-wide examination of genetic variants associated with response to platinum-based chemotherapy in patients with small-cell lung cancer. Pharmacogenet Genomics. 2010;20:389–95. [DOI] [PubMed] [Google Scholar]
- 69.Cao S, Wang C, Ma H, Yin R, Zhu M, Shen W, et al. Genome-wide Association Study on Platinum-induced Hepatotoxicity in Non-Small Cell Lung Cancer Patients. Sci Rep. 2015;5:11556. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 70.Cao S, Wang S, Ma H, Tang S, Sun C, Dai J, et al. Genome-wide association study of myelosuppression in non-small-cell lung cancer patients with platinum-based chemotherapy. Pharmacogenomics J. 2016;16:41–6. [DOI] [PubMed] [Google Scholar]
- 71.Nguyen JD, Lamontagne M, Couture C, Conti M, Pare PD, Sin DD, et al. Susceptibility loci for lung cancer are associated with mRNA levels of nearby genes in the lung. Carcinogenesis. 2014;35:2653–9. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 72.Giambartolomei C, Vukcevic D, Schadt EE, Franke L, Hingorani AD, Wallace C, et al. Bayesian test for colocalisation between pairs of genetic association studies using summary statistics. PLoS Genet. 2014;10:e1004383. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 73.Lamontagne M, Timens W, Hao K, Bossé Y, Laviolette M, Steiling K, et al. Genetic regulation of gene expression in the lung identifies CST3 and CD22 as potential causal genes for airflow obstruction. Thorax. 2014;69:997–1004. [DOI] [PubMed] [Google Scholar]
- 74.Zhu Z, Zhang F, Hu H, Bakshi A, Robinson MR, Powell JE, et al. Integration of summary data from GWAS and eQTL studies predicts complex trait gene targets. Nat Genet. 2016;48:481–7. [DOI] [PubMed] [Google Scholar]
- 75.Hao K, Bossé Y, Nickle DC, Pare PD, Postma DS, Laviolette M, et al. Lung eQTLs to Help Reveal the Molecular Underpinnings of Asthma. PLoS Genet. 2012;8:e1003029. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 76.Bossé Y, Sazonova O, Gaudreault N, Bastien N, Conti M, Pagé S, et al. Transcriptomic microenvironment of lung adenocarcinoma. Cancer Epidemiology, Biomarkers & Prevention. 2016. [DOI] [PubMed] [Google Scholar]
- 77.Selamat SA, Chung BS, Girard L, Zhang W, Zhang Y, Campan M, et al. Genome-scale analysis of DNA methylation in lung adenocarcinoma and integration with mRNA expression. Genome Res. 2012;22:1197–211. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 78.Landi MT, Dracheva T, Rotunno M, Figueroa JD, Liu H, Dasgupta A, et al. Gene expression signature of cigarette smoking and its role in lung adenocarcinoma development and survival. PLoS One. 2008;3:e1651. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 79.Alexandrov LB, Ju YS, Haase K, Van Loo P, Martincorena I, Nik-Zainal S, et al. Mutational signatures associated with tobacco smoking in human cancer. Science. 2016;354:618–22. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 80.Xiong D, Wang Y, Kupert E, Simpson C, Pinney SM, Gaba CR, et al. A Recurrent Mutation in PARK2 Is Associated with Familial Lung Cancer. Am J Hum Genet. 2015;96:301–8. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 81.Liu Y, Kheradmand F, Davis CF, Scheurer ME, Wheeler D, Tsavachidis S, et al. Focused Analysis of Exome Sequencing Data for Rare Germline Mutations in Familial and Sporadic Lung Cancer. Journal of thoracic oncology : official publication of the International Association for the Study of Lung Cancer. 2016;11:52–61. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 82.Chen HY, Yu SL, Ho BC, Su KY, Hsu YC, Chang CS, et al. R331W Missense Mutation of Oncogene YAP1 Is a Germline Risk Allele for Lung Adenocarcinoma With Medical Actionability. J Clin Oncol. 2015;33:2303–10. [DOI] [PubMed] [Google Scholar]
- 83.Wang X, Oldani MJ, Zhao X, Huang X, Qian D. A review of cancer risk prediction models with genetic variants. Cancer Inform. 2014;13:19–28. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 84.Weissfeld JL, Lin Y, Lin HM, Kurland BF, Wilson DO, Fuhrman CR, et al. Lung Cancer Risk Prediction Using Common SNPs Located in GWAS-Identified Susceptibility Regions. Journal of thoracic oncology : official publication of the International Association for the Study of Lung Cancer. 2015;10:1538–45. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 85.Machiela MJ, Hsiung CA, Shu XO, Seow WJ, Wang Z, Matsuo K, et al. Genetic variants associated with longer telomere length are associated with increased lung cancer risk among never-smoking women in Asia: a report from the female lung cancer consortium in Asia. International journal of cancer Journal international du cancer. 2015;137:311–9. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 86.Qian DC, Han Y, Byun J, Shin HR, Hung RJ, McLaughlin JR, et al. A Novel Pathway-Based Approach Improves Lung Cancer Risk Prediction Using Germline Genetic Variations. Cancer Epidemiol Biomarkers Prev. 2016;25:1208–15. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 87.McDonald JM, Dunlap S, Cogdell D, Dunmire V, Wei Q, Starzinski-Powitz A, et al. The SHREW1 gene, frequently deleted in oligodendrogliomas, functions to inhibit cell adhesion and migration. Cancer biology & therapy. 2006;5:300–4. [DOI] [PubMed] [Google Scholar]
- 88.Habbig S, Bartram MP, Muller RU, Schwarz R, Andriopoulos N, Chen S, et al. NPHP4, a cilia-associated protein, negatively regulates the Hippo pathway. The Journal of cell biology. 2011;193:633–42. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 89.Nath S, Mukherjee P. MUC1: a multifaceted oncoprotein with a key role in cancer progression. Trends Mol Med. 2014;20:332–42. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 90.Kharbanda A, Rajabi H, Jin C, Tchaicha J, Kikuchi E, Wong KK, et al. Targeting the oncogenic MUC1-C protein inhibits mutant EGFR-mediated signaling and survival in non-small cell lung cancer cells. Clin Cancer Res. 2014;20:5423–34. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 91.Kuefer R, Day KC, Kleer CG, Sabel MS, Hofer MD, Varambally S, et al. ADAM15 disintegrin is associated with aggressive prostate and breast cancer disease. Neoplasia. 2006;8:319–29. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 92.Nussbaum J, Xu Q, Payne TJ, Ma JZ, Huang W, Gelernter J, et al. Significant association of the neurexin-1 gene (NRXN1) with nicotine dependence in European- and African-American smokers. Hum Mol Genet. 2008;17:1569–77. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 93.Hawryluk-Gara LA, Platani M, Santarella R, Wozniak RW, Mattaj IW. Nup53 is required for nuclear envelope and nuclear pore complex assembly. Molecular biology of the cell. 2008;19:1753–62. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 94.Li SR, Gyselman VG, Lalude O, Dorudi S, Bustin SA. Transcription of the inositol polyphosphate 1-phosphatase gene (INPP1) is upregulated in human colorectal cancer. Mol Carcinog. 2000;27:322–9. [DOI] [PubMed] [Google Scholar]
- 95.Dasgupta S, Chakraborty SB, Roy A, Roychowdhury S, Panda CK. Differential deletions of chromosome 3p are associated with the development of uterine cervical carcinoma in Indian patients. Molecular pathology : MP. 2003;56:263–9. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 96.Braga E, Senchenko V, Bazov I, Loginov W, Liu J, Ermilova V, et al. Critical tumor-suppressor gene regions on chromosome 3P in major human epithelial malignancies: allelotyping and quantitative real-time PCR. International journal of cancer Journal international du cancer. 2002;100:534–41. [DOI] [PubMed] [Google Scholar]
- 97.Katoh I, Aisaki KI, Kurata SI, Ikawa S, Ikawa Y. p51A (TAp63gamma), a p53 homolog, accumulates in response to DNA damage for cell regulation. Oncogene. 2000;19:3126–30. [DOI] [PubMed] [Google Scholar]
- 98.Flores ER. The roles of p63 in cancer. Cell cycle. 2007;6:300–4. [DOI] [PubMed] [Google Scholar]
- 99.Bonne A, Vreede L, Kuiper RP, Bodmer D, Jansen C, Eleveld M, et al. Mapping of constitutional translocation breakpoints in renal cell cancer patients: identification of KCNIP4 as a candidate gene. Cancer genetics and cytogenetics. 2007;179:11–8. [DOI] [PubMed] [Google Scholar]
- 100.Janknecht R On the road to immortality: hTERT upregulation in cancer cells. FEBS Lett. 2004;564:9–13. [DOI] [PubMed] [Google Scholar]
- 101.Lantuejoul S, Salon C, Soria JC, Brambilla E. Telomerase expression in lung preneoplasia and neoplasia. International journal of cancer Journal international du cancer. 2007;120:1835–41. [DOI] [PubMed] [Google Scholar]
- 102.Mavrogiannou E, Strati A, Stathopoulou A, Tsaroucha EG, Kaklamanis L, Lianidou ES. Real-time RT-PCR quantification of human telomerase reverse transcriptase splice variants in tumor cell lines and non-small cell lung cancer. Clin Chem. 2007;53:53–61. [DOI] [PubMed] [Google Scholar]
- 103.Wu TC, Lin P, Hsu CP, Huang YJ, Chen CY, Chung WC, et al. Loss of telomerase activity may be a potential favorable prognostic marker in lung carcinomas. Lung cancer. 2003;41:163–9. [DOI] [PubMed] [Google Scholar]
- 104.Strazisar M, Mlakar V, Glavac D. The expression of COX-2, hTERT, MDM2, LATS2 and S100A2 in different types of non-small cell lung cancer (NSCLC). Cell Mol Biol Lett. 2009;14:442–56. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 105.Xie M, Chen Q, He S, Li B, Hu C. Silencing of the human TERT gene by RNAi inhibits A549 lung adenocarcinoma cell growth in vitro and in vivo. Oncology reports. 2011;26:1019–27. [DOI] [PubMed] [Google Scholar]
- 106.Yamamoto K, Okamoto A, Isonishi S, Ochiai K, Ohtake Y. A novel gene, CRR9, which was up-regulated in CDDP-resistant ovarian tumor cell line, was associated with apoptosis. Biochemical and biophysical research communications. 2001;280:1148–54. [DOI] [PubMed] [Google Scholar]
- 107.Zienolddiny S, Skaug V, Landvik NE, Ryberg D, Phillips DH, Houlston R, et al. The TERT-CLPTM1L lung cancer susceptibility variant associates with higher DNA adduct formation in the lung. Carcinogenesis. 2009;30:1368–71. [DOI] [PubMed] [Google Scholar]
- 108.James MA, Wen W, Wang Y, Byers LA, Heymach JV, Coombes KR, et al. Functional characterization of CLPTM1L as a lung cancer risk candidate gene in the 5p15.33 locus. PLoS One. 2012;7:e36116. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 109.Ni Z, Tao K, Chen G, Chen Q, Tang J, Luo X, et al. CLPTM1L is overexpressed in lung cancer and associated with apoptosis. PLoS One. 2012;7:e52598. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 110.James MA, Vikis HG, Tate E, Rymaszewski AL, You M. CRR9/CLPTM1L regulates cell survival signaling and is required for Ras transformation and lung tumorigenesis. Cancer Res. 2014;74:1116–27. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 111.Yu H, Zhao H, Wang LE, Han Y, Chen WV, Amos CI, et al. An analysis of single nucleotide polymorphisms of 125 DNA repair genes in the Texas genome-wide association study of lung cancer with a replication for the XRCC4 SNPs. DNA Repair (Amst). 2011;10:398–407. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 112.Teodoro JG, Parker AE, Zhu X, Green MR. p53-mediated inhibition of angiogenesis through up-regulation of a collagen prolyl hydroxylase. Science. 2006;313:968–71. [DOI] [PubMed] [Google Scholar]
- 113.Dougan M, Li D, Neuberg D, Mihm M, Googe P, Wong KK, et al. A dual role for the immune response in a mouse model of inflammation-associated lung cancer. J Clin Invest. 2011;121:2436–46. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 114.Zeller T, Wild P, Szymczak S, Rotival M, Schillert A, Castagne R, et al. Genetics and beyond--the transcriptome of human monocytes and disease susceptibility. PLoS One. 2010;5:e10693. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 115.Chen J, Brunzell DH, Jackson K, van der Vaart A, Ma JZ, Payne TJ, et al. ACSL6 is associated with the number of cigarettes smoked and its expression is altered by chronic nicotine exposure. PLoS One. 2011;6:e28790. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 116.Vazquez A, Kulkarni D, Grochola LF, Bond GL, Barnard N, Toppmeyer D, et al. A genetic variant in a PP2A regulatory subunit encoded by the PPP2R2B gene associates with altered breast cancer risk and recurrence. International journal of cancer Journal international du cancer. 2011;128:2335–43. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 117.Tan J, Lee PL, Li Z, Jiang X, Lim YC, Hooi SC, et al. B55beta-associated PP2A complex controls PDK1-directed myc signaling and modulates rapamycin sensitivity in colorectal cancer. Cancer Cell. 2010;18:459–71. [DOI] [PubMed] [Google Scholar]
- 118.Okita K, Motohashi S, Shinnakasu R, Nagato K, Yamasaki K, Sato Y, et al. A set of genes associated with the interferon-gamma response of lung cancer patients undergoing alpha-galactosylceramide-pulsed dendritic cell therapy. Cancer science. 2010;101:2333–40. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 119.Sasaki T, Gan EC, Wakeham A, Kornbluth S, Mak TW, Okada H. HLA-B-associated transcript 3 (Bat3)/Scythe is essential for p300-mediated acetylation of p53. Genes Dev. 2007;21:848–61. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 120.Grover A, Izzo AA. BAT3 regulates Mycobacterium tuberculosis protein ESAT-6-mediated apoptosis of macrophages. PLoS One. 2012;7:e40836. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 121.Mu QF, Luo GH, Chen LJ, Wei J, Zheng L, Zhang XY, et al. [Apolipoprotein M expression in human colorectal cancer tissues and its clinicopathological relevance]. Zhonghua wei chang wai ke za zhi = Chinese journal of gastrointestinal surgery. 2012;15:855–8. [PubMed] [Google Scholar]
- 122.Luo G, Zhang X, Mu Q, Chen L, Zheng L, Wei J, et al. Expression and localization of apolipoprotein M in human colorectal tissues. Lipids in health and disease. 2010;9:102. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 123.Levy P, Ripoche H, Laurendeau I, Lazar V, Ortonne N, Parfait B, et al. Microarray-based identification of tenascin C and tenascin XB, genes possibly involved in tumorigenesis associated with neurofibromatosis type 1. Clin Cancer Res. 2007;13:398–407. [DOI] [PubMed] [Google Scholar]
- 124.Tompkins JD, Wu X, Her C. MutS homologue hMSH5: role in cisplatin-induced DNA damage response. Molecular cancer. 2012;11:10. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 125.Robson T, James IF. The therapeutic and diagnostic potential of FKBPL; a novel anticancer protein. Drug Discov Today. 2012;17:544–8. [DOI] [PubMed] [Google Scholar]
- 126.McKeen HD, Brennan DJ, Hegarty S, Lanigan F, Jirstrom K, Byrne C, et al. The emerging role of FK506-binding proteins as cancer biomarkers: a focus on FKBPL. Biochem Soc Trans. 2011;39:663–8. [DOI] [PubMed] [Google Scholar]
- 127.Guo H, Deng Q, Wu C, Hu L, Wei S, Xu P, et al. Variations in HSPA1B at 6p21.3 are associated with lung cancer risk and prognosis in Chinese populations. Cancer Res. 2011;71:7576–86. [DOI] [PubMed] [Google Scholar]
- 128.Wang Y, Wei Y, Gaborieau V, Shi J, Han Y, Timofeeva MN, et al. Deciphering associations for lung cancer risk through imputation and analysis of 12,316 cases and 16,831 controls. Eur J Hum Genet. 2015;23:1723–8. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 129.Koshikawa K, Osada H, Kozaki K, Konishi H, Masuda A, Tatematsu Y, et al. Significant up-regulation of a novel gene, CLCP1, in a highly metastatic lung cancer subline as well as in lung cancers in vivo. Oncogene. 2002;21:2822–8. [DOI] [PubMed] [Google Scholar]
- 130.Nagai H, Sugito N, Matsubara H, Tatematsu Y, Hida T, Sekido Y, et al. CLCP1 interacts with semaphorin 4B and regulates motility of lung cancer cells. Oncogene. 2007;26:4025–31. [DOI] [PubMed] [Google Scholar]
- 131.Acquaviva J, Wong R, Charest A. The multifaceted roles of the receptor tyrosine kinase ROS in development and cancer. Biochimica et biophysica acta. 2009;1795:37–52. [DOI] [PubMed] [Google Scholar]
- 132.Takeuchi K, Soda M, Togashi Y, Suzuki R, Sakata S, Hatano S, et al. RET, ROS1 and ALK fusions in lung cancer. Nat Med. 2012;18:378–81. [DOI] [PubMed] [Google Scholar]
- 133.Bergethon K, Shaw AT, Ou SH, Katayama R, Lovly CM, McDonald NT, et al. ROS1 rearrangements define a unique molecular class of lung cancers. J Clin Oncol. 2012;30:863–70. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 134.Fernandez-Cuesta L, Plenker D, Osada H, Sun R, Menon R, Leenders F, et al. CD74-NRG1 fusions in lung adenocarcinoma. Cancer discovery. 2014;4:415–22. [DOI] [PubMed] [Google Scholar]
- 135.Gil J, Peters G. Regulation of the INK4b-ARF-INK4a tumour suppressor locus: all for one or one for all. Nature reviews Molecular cell biology. 2006;7:667–77. [DOI] [PubMed] [Google Scholar]
- 136.Kotake Y, Nakagawa T, Kitagawa K, Suzuki S, Liu N, Kitagawa M, et al. Long non-coding RNA ANRIL is required for the PRC2 recruitment to and silencing of p15(INK4B) tumor suppressor gene. Oncogene. 2011;30:1956–62. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 137.Yu W, Gius D, Onyango P, Muldoon-Jacobs K, Karp J, Feinberg AP, et al. Epigenetic silencing of tumour suppressor gene p15 by its antisense RNA. Nature. 2008;451:202–6. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 138.Enciso-Mora V, Broderick P, Ma Y, Jarrett RF, Hjalgrim H, Hemminki K, et al. A genome-wide association study of Hodgkin’s lymphoma identifies new susceptibility loci at 2p16.1 (REL), 8q24.21 and 10p14 (GATA3). Nat Genet. 2010;42:1126–30. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 139.Maurice D, Hooper J, Lang G, Weston K. c-Myb regulates lineage choice in developing thymocytes via its target gene Gata3. EMBO J. 2007;26:3629–40. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 140.Tkocz D, Crawford NT, Buckley NE, Berry FB, Kennedy RD, Gorski JJ, et al. BRCA1 and GATA3 corepress FOXC1 to inhibit the pathogenesis of basal-like breast cancers. Oncogene. 2012;31:3667–78. [DOI] [PubMed] [Google Scholar]
- 141.Asselin-Labat ML, Sutherland KD, Vaillant F, Gyorki DE, Wu D, Holroyd S, et al. Gata-3 negatively regulates the tumor-initiating capacity of mammary luminal progenitor cells and targets the putative tumor suppressor caspase-14. Mol Cell Biol. 2011;31:4609–22. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 142.Wu Q, Wang H, Zhao X, Shi Y, Jin M, Wan B, et al. Identification of G-protein-coupled receptor 120 as a tumor-promoting receptor that induces angiogenesis and migration in human colorectal carcinoma. Oncogene. 2013;32:5541–50. [DOI] [PubMed] [Google Scholar]
- 143.Bose A, Guilherme A, Huang S, Hubbard AC, Lane CR, Soriano NA, et al. The v-SNARE Vti1a regulates insulin-stimulated glucose transport and Acrp30 secretion in 3T3-L1 adipocytes. The Journal of biological chemistry. 2005;280:36946–51. [DOI] [PubMed] [Google Scholar]
- 144.Petridou ET, Mitsiades N, Gialamas S, Angelopoulos M, Skalkidou A, Dessypris N, et al. Circulating adiponectin levels and expression of adiponectin receptors in relation to lung cancer: two case-control studies. Oncology. 2007;73:261–9. [DOI] [PubMed] [Google Scholar]
- 145.Bass AJ, Lawrence MS, Brace LE, Ramos AH, Drier Y, Cibulskis K, et al. Genomic sequencing of colorectal adenocarcinomas identifies a recurrent VTI1A-TCF7L2 fusion. Nat Genet. 2011;43:964–8. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 146.Mortensen UH, Lisby M, Rothstein R. Rad52. Current biology : CB. 2009;19:R676–7. [DOI] [PubMed] [Google Scholar]
- 147.Danoy P, Michiels S, Dessen P, Pignat C, Boulet T, Monet M, et al. Variants in DNA double-strand break repair and DNA damage-response genes and susceptibility to lung and head and neck cancers. International journal of cancer Journal international du cancer. 2008;123:457–63. [DOI] [PubMed] [Google Scholar]
- 148.Benson FE, Baumann P, West SC. Synergistic actions of Rad51 and Rad52 in recombination and DNA repair. Nature. 1998;391:401–4. [DOI] [PubMed] [Google Scholar]
- 149.Feng Z, Scott SP, Bussen W, Sharma GG, Guo G, Pandita TK, et al. Rad52 inactivation is synthetically lethal with BRCA2 deficiency. Proc Natl Acad Sci U S A. 2011;108:686–91. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 150.Treuner K, Helton R, Barlow C. Loss of Rad52 partially rescues tumorigenesis and T-cell maturation in Atm-deficient mice. Oncogene. 2004;23:4655–61. [DOI] [PubMed] [Google Scholar]
- 151.Lieberman R, Xiong D, James M, Han Y, Amos CI, Wang L, et al. Functional characterization of RAD52 as a lung cancer susceptibility gene in the 12p13.33 locus. Mol Carcinog. 2016;55:953–63. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 152.Su GH, Bansal R, Murphy KM, Montgomery E, Yeo CJ, Hruban RH, et al. ACVR1B (ALK4, activin receptor type 1B) gene mutations in pancreatic carcinoma. Proc Natl Acad Sci U S A. 2001;98:3254–7. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 153.Burdette JE, Jeruss JS, Kurley SJ, Lee EJ, Woodruff TK. Activin A mediates growth inhibition and cell cycle arrest through Smads in human breast cancer cells. Cancer Res. 2005;65:7968–75. [DOI] [PubMed] [Google Scholar]
- 154.Modica S, Murzilli S, Salvatore L, Schmidt DR, Moschetta A. Nuclear bile acid receptor FXR protects against intestinal tumorigenesis. Cancer Res. 2008;68:9589–94. [DOI] [PubMed] [Google Scholar]
- 155.Park JY, Helm JF, Zheng W, Ly QP, Hodul PJ, Centeno BA, et al. Silencing of the candidate tumor suppressor gene solute carrier family 5 member 8 (SLC5A8) in human pancreatic cancer. Pancreas. 2008;36:e32–9. [DOI] [PubMed] [Google Scholar]
- 156.Rafnar T, Vermeulen SH, Sulem P, Thorleifsson G, Aben KK, Witjes JA, et al. European genome-wide association study identifies SLC14A1 as a new urinary bladder cancer susceptibility gene. Hum Mol Genet. 2011;20:4268–81. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 157.Eby MT, Jasmin A, Kumar A, Sharma K, Chaudhary PM. TAJ, a novel member of the tumor necrosis factor receptor family, activates the c-Jun N-terminal kinase pathway and mediates caspase-independent cell death. The Journal of biological chemistry. 2000;275:15336–42. [DOI] [PubMed] [Google Scholar]
- 158.McAllister KA, Bennett LM, Houle CD, Ward T, Malphurs J, Collins NK, et al. Cancer susceptibility of mice with a homozygous deletion in the COOH-terminal domain of the Brca2 gene. Cancer Res. 2002;62:990–4. [PubMed] [Google Scholar]
- 159.De Cat B, David G. Developmental roles of the glypicans. Seminars in cell & developmental biology. 2001;12:117–25. [DOI] [PubMed] [Google Scholar]
- 160.Filmus J Glypicans in growth control and cancer. Glycobiology. 2001;11:19R–23R. [DOI] [PubMed] [Google Scholar]
- 161.Yu CT, Hsia JY, Hseih YC, Su LJ, Lee TC, Ku CF, et al. The novel protein suppressed in lung cancer down-regulated in lung cancer tissues retards cell proliferation and inhibits the oncokinase Aurora-A. Journal of thoracic oncology : official publication of the International Association for the Study of Lung Cancer. 2011;6:988–97. [DOI] [PubMed] [Google Scholar]
- 162.Liu Y, Liu P, Wen W, James MA, Wang Y, Bailey-Wilson JE, et al. Haplotype and cell proliferation analyses of candidate lung cancer susceptibility genes on chromosome 15q24–25.1. Cancer Res. 2009;69:7844–50. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 163.Ruthenburg AJ, Li H, Milne TA, Dewell S, McGinty RK, Yuen M, et al. Recognition of a mononucleosomal histone modification pattern by BPTF via multivalent interactions. Cell. 2011;145:692–706. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 164.Medina PP, Sanchez-Cespedes M. Involvement of the chromatin-remodeling factor BRG1/SMARCA4 in human cancer. Epigenetics : official journal of the DNA Methylation Society. 2008;3:64–8. [DOI] [PubMed] [Google Scholar]
- 165.Coste B, Mathur J, Schmidt M, Earley TJ, Ranade S, Petrus MJ, et al. Piezo1 and Piezo2 are essential components of distinct mechanically activated cation channels. Science. 2010;330:55–60. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 166.Shimomura Y, Agalliu D, Vonica A, Luria V, Wajid M, Baumer A, et al. APCDD1 is a novel Wnt inhibitor mutated in hereditary hypotrichosis simplex. Nature. 2010;464:1043–7. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 167.Takahashi M, Fujita M, Furukawa Y, Hamamoto R, Shimokawa T, Miwa N, et al. Isolation of a novel human gene, APCDD1, as a direct target of the beta-Catenin/T-cell factor 4 complex with probable involvement in colorectal carcinogenesis. Cancer Res. 2002;62:5651–6. [PubMed] [Google Scholar]
- 168.Tashiro K, Tsunematsu T, Okubo H, Ohta T, Sano E, Yamauchi E, et al. GAREM, a novel adaptor protein for growth factor receptor-bound protein 2, contributes to cellular transformation through the activation of extracellular signal-regulated kinase signaling. The Journal of biological chemistry. 2009;284:20206–14. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 169.Park KH, Lo Han SG, Whang YM, Lee HJ, Yoo YD, Lee JW, et al. Single nucleotide polymorphisms of the TGFB1 gene and lung cancer risk in a Korean population. Cancer genetics and cytogenetics. 2006;169:39–44. [DOI] [PubMed] [Google Scholar]
- 170.Liao Q, Zeng Z, Guo X, Li X, Wei F, Zhang W, et al. LPLUNC1 suppresses IL-6-induced nasopharyngeal carcinoma cell proliferation via inhibiting the Stat3 activation. Oncogene. 2014;33:2098–109. [DOI] [PubMed] [Google Scholar]
- 171.Lin R, White JH. The pleiotropic actions of vitamin D. BioEssays : news and reviews in molecular, cellular and developmental biology. 2004;26:21–8. [DOI] [PubMed] [Google Scholar]
- 172.Peehl DM, Skowronski RJ, Leung GK, Wong ST, Stamey TA, Feldman D. Antiproliferative effects of 1,25-dihydroxyvitamin D3 on primary cultures of human prostatic cells. Cancer Res. 1994;54:805–10. [PubMed] [Google Scholar]
- 173.Anderson MG, Nakane M, Ruan X, Kroeger PE, Wu-Wong JR. Expression of VDR and CYP24A1 mRNA in human tumors. Cancer chemotherapy and pharmacology. 2006;57:234–40. [DOI] [PubMed] [Google Scholar]
- 174.Chen G, Kim SH, King AN, Zhao L, Simpson RU, Christensen PJ, et al. CYP24A1 is an independent prognostic marker of survival in patients with lung adenocarcinoma. Clin Cancer Res. 2011;17:817–26. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 175.Wrensch M, Jenkins RB, Chang JS, Yeh RF, Xiao Y, Decker PA, et al. Variants in the CDKN2B and RTEL1 regions are associated with high-grade glioma susceptibility. Nat Genet. 2009;41:905–8. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 176.Cybulski C, Masojc B, Oszutowska D, Jaworowska E, Grodzki T, Waloszczyk P, et al. Constitutional CHEK2 mutations are associated with a decreased risk of lung and laryngeal cancers. Carcinogenesis. 2008;29:762–5. [DOI] [PubMed] [Google Scholar]
- 177.Chen Y, Deng J, Fujimoto J, Kadara H, Men T, Lotan D, et al. Gprc5a deletion enhances the transformed phenotype in normal and malignant lung epithelial cells by eliciting persistent Stat3 signaling induced by autocrine leukemia inhibitory factor. Cancer Res. 2010;70:8917–26. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 178.Niwa H, Ogawa K, Shimosato D, Adachi K. A parallel circuit of LIF signalling pathways maintains pluripotency of mouse ES cells. Nature. 2009;460:118–22. [DOI] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.