

Abstract
The distribution of New England salt marsh communities is intrinsically linked to the magnitude, frequency, and duration of tidal inundation. Cordgrass (Spartina alterniflora) exclusively inhabits the frequently flooded lower elevations, whereas a mosaic of marsh hay (Spartina patens), spike grass (Distichlis spicata), and black rush (Juncus gerardi) typically dominate higher elevations. Monitoring plant zonal boundaries in two New England salt marshes revealed that low-marsh cordgrass rapidly moved landward at the expense of higher-marsh species between 1995 and 1998. Plant macrofossils from sediment cores across modern plant community boundaries provided a 2,500-year record of marsh community composition and documented the migration of cordgrass into the high marsh. Isotopic dating revealed that the initiation of cordgrass migration occurred in the late 19th century and continued through the 20th century. The timing of the initiation of cordgrass migration is coincident with an acceleration in the rate of sea-level rise recorded by the New York tide gauge. These results suggest that increased flooding associated with accelerating rates of sea-level rise has stressed high-marsh communities and promoted landward migration of cordgrass. If current rates of sea-level rise continue or increase slightly over the next century, New England salt marshes will be dominated by cordgrass. If climate warming causes sea-level rise rates to increase significantly over the next century, these cordgrass-dominated marshes will likely drown, resulting in extensive losses of coastal wetlands.
Recent studies indicate that both climate warming (1, 2) and increases in the rate of sea-level rise (SLR) in New England (3) over the last 150 years are unprecedented in at least the last 1,000 years. The possibility that emission of greenhouse gases is influencing and will continue to influence global climate and potentially SLR has prompted considerable research into the possible implications for plant and animal communities (4). The distribution of salt-marsh communities is mechanistically linked to the duration of tidal inundation. As a result, coastal wetlands may be particularly sensitive to changes in sea level (5). Wetland loss has been documented in areas of the Mississippi River Delta (6) and Chesapeake Bay (7), where rates of local SLR exceed marsh accretion.
In New England salt marshes, cordgrass (Spartina alterniflora) exclusively dominates daily flooded low-marsh elevations, whereas a mosaic of marsh hay (Spartina patens), spike grass (Distichlis spicata), and black rush (Juncus gerardi) dominates higher marsh elevations (8). Lower species borders are controlled by physical stress tolerance to flooding and soil anoxia, whereas upper species borders are controlled by interspecific plant competition (9, 10). Marsh hay and spike grass are excluded from the low marsh by low substrate oxygen levels. The ability of cordgrass to oxygenate substrates (11) allows this species to dominate frequently flooded lower-marsh elevations, whereas it is competitively excluded from higher elevations where soil oxygen content is higher.
Existing salt marshes in New England developed during the last 4,000 years and have generally kept pace with moderate rates of SLR of about 1 mm/year (12). In the Redfield model of marsh development (12), low marsh dominated by cordgrass accretes vertically and seaward over aggrading mud or sand flats. High marsh, dominated by marsh hay, spike grass, and black rush, exists at about mean high water and accretes vertically and laterally over submerging upland and low-marsh sediments. Several studies have noted a recent reversal of this long-term trend in marsh-community dynamics by documenting sediments dominated by cordgrass remains overlying high-marsh sediments at several sites in southern New England (13–15). Two of these studies (13, 14) attributed the recent migration of cordgrass onto the high-marsh surface to the combined influence of changes in geomorphology associated with the September 21, 1938, New England Hurricane and increased rates of SLR. However, in a southern Rhode Island marsh, the initiation of cordgrass migration predates deposits associated with the 1938 Hurricane (15). Localized nutrient additions can cause a reversal in the competitive relationship between cordgrass and marsh hay resulting in cordgrass migration to higher elevations at the expense of marsh hay and increased cordgrass biomass (16). Therefore, the observations of increased abundance of cordgrass in southern New England may be the result of increased flooding frequency and/or a change in the nutrient-mediated competitiveness between cordgrass and marsh hay. Here we present the results from a combined ecological and stratigraphic study designed to quantify recent changes in vegetation composition at two southern New England salt marshes and test the hypothesis that increased rates of SLR are increasing cordgrass abundance in high-marsh environments.
Monitoring the boundary between modern cordgrass and the high-marsh community in two representative marshes in Narragansett Bay (Rumstick Cove in Barrington, Rhode Island, and Nag Creek on Prudence Island, Rhode Island; Fig. 1) from 1995 to 1999 revealed increasing dominance of cordgrass at the expense of marsh hay (Fig. 2). At each site we established sixteen 1-m2 monitoring quadrats on the cordgrass/marsh hay (low marsh/high marsh) border in September 1995. These quadrats were initially positioned on the seaward border of the marsh hay zone, including the landward border of the cordgrass zone, and marked with numbered poly(vinly chloride) (PVC) stakes. Vegetation cover in these plots was quantified during the autumns of 1995–1999 by placing a 1-m2 grid of 5 × 5 cm cells over each plot and scoring each cell for the presence of cordgrass and marsh hay. During this time period none of the monitoring quadrats were disturbed by floating wrack, which can lead to rapid plant species shifts (9). At both sites, the cordgrass/marsh hay border dramatically migrated shoreward during this short period (Fig. 2). Cordgrass cover in these border plots increased more than 5-fold at both sites, whereas marsh hay cover decreased by 16% at Rumstick Cove and 40% at Nag Creek. These data demonstrate that cordgrass is currently moving rapidly shoreward in these marshes, displacing marsh hay.
Figure 1.

(a) Map showing the location of the Prudence Island and Rumstick Cove in Narragansett Bay, RI. (b) Map of Nag Creek (Prudence Island) study site with core transect shown in red on the southeastern edge of Nag Pond and a blow-up of study site with core locations (indicated by numbers) and surface vegetation mapped. (c) Map of Rumstick Cove study site with core transect to the east of the tidal creek in the northern portion of the marsh and a blow-up of study site with core locations (indicated by numbers) and surface vegetation mapped.
Figure 2.

Plant cover in monitoring quadrats originally placed on the seaward border of the marsh hay vegetation zone at our two New England salt marsh study sites. For each site the data represent mean cover data from 16 quadrats ± SE. These data were log transformed and analyzed by study site and plant species with repeated-measure ANOVA. At both sites, cordgrass significantly increased (P < 0.001), whereas marsh hay significantly decreased (P < 0.01) in the plots, indicating shoreward movement of marsh vegetation.
Long-term records of community composition are required to examine possible influences of a warming climate and SLR on ecosystems. Salt-marsh peat can provide a record of vegetation dating back several thousand years (8, 12). To document salt-marsh community composition over the last several thousand years and test the hypothesis that cordgrass has migrated landward in response to recent increases in the rate of SLR, we recovered a series sediment cores (7.5 cm diameter) by using a vibracore rig (17) from across the modern cordgrass/high marsh transition at these sites (Fig. 1). Cores were split and described in the laboratory, and fossil roots and rhizomes were identified to species by using the key developed by Niering et al. (18).
The remains of salt-marsh vegetation preserved within the peat at both sites reveal a time-transgressive transition from marsh hay or spike grass to cordgrass (Fig. 3). We used Cs-137 (19) and Pb-210 (20) methods to provide age control in cores at both sites. The presence of Cs-137 preserved in the sediments is associated with nuclear weapon testing, and its initiation is interpreted to correspond to ≈1954. We use the initiation of Cs-137 preserved in these cores (Fig. 4) to determine the stratigraphic interval that was deposited in about 1954 (Fig. 3).
Figure 3.

(Left) Stratigraphic cross-section of Nag Creek Marsh transect. (Right) Stratigraphic crosssection of Rumstick Cove Marsh transect. Both Right and Left are based on the close proximity of the cores and the consistent stratigraphy we have interpolated between cores. We are unable to discern the continuity of the cordgrass unit below the marsh hay unit in core RUM2 with the cordgrass unit evident in core RUM1, so we have included a question mark between cores RUM1 and RUM2 to illustrate this uncertainty. Radiocarbon dates and maximum and minimum calibrated calendar age ranges at two standard deviations are displayed for two samples from NC7. White arrows represent the interval at which Cs-137 becomes detectable and is inferred to have been deposited in 1954. Black arrows represent the interval deposited in 1900 based on accretion rates derived from Pb-210 activity. The age of the oldest and most seaward transition from marsh hay or spike grass peat to cordgrass was estimated by extending the accretion rate derived from Pb-210 to this horizon in cores NC1 and RUM1. The extrapolation of Pb-210 accretion rates to this contact indicates that the initiation of cordgrass migration occurred in the late 19th century (1876 in NC1 and 1870 in RUM1).
Figure 4.

Cs-137 and Pb-210 profiles from Nag Creek and Rumstick Marsh. The initial rise in Cs-137 corresponds to 1954 and the peak in Cs-137 activity corresponds to 1963. Unsupported Pb-210 profiles are plotted on a logarithmic scale with respect to Pb-210 activity. S, accretion rate calculated by using Eq. 1.
Pb-210 activity within sediment can be used to estimate accretion rates. Pb-210 is a product of the uranium-decay series where Ra-226 within the crust decays to Rn-222. A fraction of this Rn-222 enters the atmosphere, where it in turn decays to Pb-210. Pb-210 quickly precipitates out of the atmosphere, is deposited at the surface, and decays with a half-life of 22.3 years. If a rate of accretion is constant, the decay process results in an exponential decrease in Pb-210 activity with depth that can be used to estimate sediment accretion rates and therefore sediment age back about 100–150 years (20). The activity of Pb-210 samples where the curve becomes asymptotic with respect to Pb-210 activity is assumed to be the supported Pb-210 level; that is, the amount of Pb-210 produced from the decay of Rn-222 within the sediment column and not deposited from the atmosphere. These supported Pb-210 values are averaged and subtracted from the values obtained above, resulting in an unsupported Pb-210 profile (from atmospheric deposition). An accretion rate can then be calculated by using the relationship:
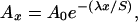 |
1 |
where S is the sedimentation rate, Ao is the unsupported activity of Pb-210 in the modern surface sediments, Ax is the unsupported activity of Pb-210 at depth x, λ is the Pb-210 decay constant (−0.03114 yr−1), and x is depth.
To define an additional dated horizon we calculate the approximate depth of the horizon deposited in 1900 by determining accretion rates based on the Pb-210 activity of the sediments (Fig. 4) and multiplying the accretion rate by 98 years (the number of years of sediment accumulation between 1900 and 1998, the year of sample collection). Two 1-cm3 samples from 230.5–231.5 cm and 85–86 cm in Nag Creek core 7 (NC7) were radiocarbon dated and yielded the ages 2,420 ± 35 years Before Present (2,339–2,761 cal years B.P.) and 740 ± 30 years B.P (702–771 cal years B.P.), respectively.
Cordgrass has migrated landward replacing marsh hay at Nag Creek, resulting in a wedge of peat at the surface dominated by cordgrass remains (Fig. 3). The wedge of cordgrass peat tapers landward from a thickness of 60 cm at the marine edge of the marsh. Marsh-hay-dominated peat extends to a depth of over 2 m. The transition from marsh-hay peat to cordgrass peat in NC1 occurs 11 cm below the 1900 horizon. If the accretion rate of 4.9 mm/year derived from the Pb-210 profile in the upper part of this core is extended to the cordgrass/marsh-hay transition (Fig. 4), the date of the transition is ≈1876 (Fig. 3). The transition to cordgrass in NC8 occurs 3.5 cm above the 1954 horizon and indicates the change to cordgrass in core NC8 likely occurred around 1960.
To test whether increased tidal inundation or nutrients are responsible for cordgrass migration, we determined the percent organic matter within both cordgrass and marsh-hay peat by loss on ignition (21) analysis of continuous 1-cm3 samples from the upper 50 cm of NC1, NC3, NC7, and NC8. If addition of nutrients causes cordgrass migration the organic content of cordgrass peat should be greater than the underlying marsh-hay peat as a result of enhanced biomass production associated with increases in available nutrients. If an increase in tidal inundation is responsible for cordgrass migration, the organic content of cordgrass peat should be less than the underlying marsh-hay peat because of increased deposition of tidally borne mineral sediment. The 78 cordgrass peat samples have significantly less organic matter (33.5 ± 5.2%) than the 122 marsh-hay peat samples (43.7 ± 6.2%) at a 1% significance level, indicating that increased tidal inundation is likely responsible for cordgrass migration.
The same transgressive trend toward cordgrass dominance is evident at Rumstick Cove, as cordgrass peat has transgressed over high-marsh peat dominated by spike-grass remains in Rumstick cores 1 and 3 (RUM1 and RUM3; Fig. 3). Cordgrass has replaced high-marsh species by migrating onto the high marsh from the lower-marsh elevations at the marine edge and through the development of pannes in the interior of the high marsh. Core RUM3 was taken from the edge of one of these pannes, where cordgrass remains become dominant at 11 cm depth. This panne development is similar to cordgrass-dominated panne development documented in the last 60 years at Barn Island, Connecticut (13). The transition to cordgrass in core RUM1 predates the 1900 horizon derived from Pb-210 data. Extension of the Pb-210 derived accretion rate of 2.3 mm/year (Fig. 4) to this transition yields a date of about 1870.
Marsh Accretion Rates
High-marsh accretion rates in NC7 more than doubled in the last 150 years (Fig. 5a). Based on the C-14-dated samples in NC7, the marsh hay-dominated environment accreted at close to 1 mm/year from about 2,500 years ago to 100 years ago (Fig. 5a). Accretion rates in NC7 in the last 100 years based on Cs-137 and Pb-210 are about 2.5 mm/year (Fig. 5a). In the high-marsh core from Rumstick (RUM2), accretion rates calculated for the last 46 to 100 years based on Cs-137 and Pb-210 profiles are 2.3 and 2.1 mm/year, respectively (Fig. 4). Conversely, cordgrass-dominated areas have been accreting between 2.1 and as much as 5.5 mm/year (Fig. 4).
Figure 5.

(a) Marsh accretion over the last 300 years as recorded by C-14, Pb-210, and Cs-137 dating in core NC7 (green). (b) Changes in annual MSL as recorded by the NYC (1856–1998) and Newport (1931–1998) tide gauges.§ The NYC tide gauge data series is segmented into four distinct intervals of sea-level rise. (c) NH temperature anomaly record (black curve) combined with the instrumental NH temperature record (red curve) (1). (d) Global surface temperature reconstruction from borehole data (blue envelope; ref. 2).
It is important to note that marsh-accretion rates may not necessarily reflect changes in the elevation of the marsh surface. A study from marshes in Louisiana, Florida, and North Carolina indicates that, at least in these southern marshes, subsurface processes such as decomposition and autocompaction can lower the elevation of the marsh surface (22). It has not yet been determined whether subsurface processes are important in counteracting accretion in northern marshes. As a result, the accretion rates presented here should be considered maximum estimates of elevation change of the marsh surface through time.
Sea-Level and Climate Changes
The closest tide gauge to the study sites is Newport, RI§, operational since 1931 and located ≈13 km to the south of Nag Creek and 24 km south of Rumstick Cove. A New York City (NYC) tide gauge§, operating 230 km to the west since 1856, is used in this study to represent changes in mean sea level (MSL) for Rhode Island. The validity of the application of this record to Rhode Island is supported by excellent agreement between annual mean sea level recorded at Newport and NYC since 1931 (Fig. 5b). Linear regression of the NYC tide gauge data reveals that MSL has increased at a rate of ≈2.7 mm/year since 1856. Shorter-term variations in the rate of sea level change are also evident. We identify four distinct intervals with different rates of SLR since 1856, and estimate the rate of SLR for these intervals with a series of linear regressions (Fig. 5b). At NYC, MSL rose at a rate of 1 mm/year from 1856 to 1878. This rate is consistent with the rate of SLR determined from geologic evidence over the last 1,500 years for this region (23). No annual mean data are available for the NYC tide gauge data between 1878 and 1893. Between 1893 and 1921, the rate of SLR increased to 2.4 mm/year at NYC. The highest rate of SLR on a multidecadal time scale recorded by the NYC tide gauge is 4.2 mm/year occurring between 1922 and 1960. This is similar to the rate derived from a linear regression of the annual mean Newport tide gauge data from 1931 to 1960, which yielded a SLR rate of 3.8 mm/year. The rate of SLR decreases to 2.7 and 2.6 mm/year between 1961 and 1998 at NYC and Newport, respectively.
Mann et al. (1) published a record of Northern Hemisphere (NH) temperature anomalies since 1400. The instrumental portion of this record shows an increase of close to 1°C in NH temperatures from about 1920 to present (Fig. 5c). Two periods of significant increase in NH temperatures are evident in the Mann et al. (1) temperature record. The first is between 1910 and 1940, when NH temperatures increase by ≈0.4°C. The second significant rise occurs after a period of effectively no net change in NH temperatures between 1940 and 1975. In the latest period of increase, between 1975 and 1998, NH temperatures increased an additional 0.7°C. A global temperature reconstruction derived from borehole temperatures (2) indicates that the average global temperature rose about 0.9°C between 1700 and 2000 (Fig. 5c). About 0.5°C of this increase occurred between 1900 and present (Fig. 5c). If sea-level changes are linked to regional and global temperature fluctuations, a correlation between the MSL record from NYC and temperature records (Fig. 5) would be expected.
The increase in the rate of SLR from 1 mm/year to 2.4 mm/year evident in the NYC tide-gauge data occurs between 1878 and 1893, predates the increase in NH temperature from the Mann et al. (1) reconstruction by 20 or 40 years, and may indicate that a fraction of the increase in the rate of SLR may not be related to warming atmospheric temperatures on these time scales. The greatest increase in MSL on a multidecadal time scale occurs between 1922 and 1960 at a rate of 4.2 mm/year (Fig. 5). This period of greatest increase in MSL recorded by the NYC tide gauge follows the first large increase in NH temperatures in the 20th century (1910–1940) by about a decade. The more moderate rate of SLR of 2.7 mm/year initiating around 1961 lags behind the period of essentially no net change in NH temperatures that began around 1940 by about 20 years (Fig. 5). No increase in the rate of SLR is evident at NYC since the largest and most recent increase in NH temperatures that initiated around 1975 (Fig. 5). If global temperature changes drove sea-level fluctuations in the last 150 years, through thermal expansion and the melting of glaciers, the data from the NYC tide gauge and the Mann et al. (1) NH temperature record may indicate a 10- to 20-year lag in the response of the oceans to atmospheric temperature forcing. Contrary to the Mann et al. record, the global temperature reconstruction from the borehole data (2) indicates a warming of about 0.2°C in the 19th century and is more concomitant with the increase in the rate of SLR in the late 19th century evident in the NYC tide gauge (Fig. 5d). This apparent discrepancy between proxy temperature records needs to be resolved before any tenable connection between the timing of observed SLR and climate warming is drawn.
Conclusions and Implications
Cordgrass is currently replacing high-marsh species at both Rumstick and Nag Creek Marshes. The initiation of cordgrass migration at both sites roughly coincides with the increase in the rate of SLR from 1 to 2.4 mm/year recorded at the NYC tide gauge in the late 19th century (Fig. 5). Our documentation of changes from marsh hay or spike grass to cordgrass at both Rumstick Cove and Nag Creek Marshes and similar observations at numerous other sites in the region (13–15) indicate that these changes reflect at least a regional alteration of salt-marsh community composition. The relative increase in mineral matter within the cordgrass-dominated peat indicates that more frequent flooding, not nutrient additions, is responsible for the landward migration of cordgrass. The regional extent of the phenomenon, the increase in tidal-borne sedimentation, and the correspondence in time with increases in sea level measured by tide gauges indicate that a region-wide change in sea level is likely responsible for increasing (i) the frequency with which the marsh surface is flooded and (ii) cordgrass dominance. This recent migration of cordgrass appears to be unprecedented in at least the last 2,500 years, indicating that no other acceleration in SLR of the magnitude and duration of the increase occurring over the last 200 years has taken place in the past few thousand years.
The Intergovernmental Panel on Climate Change (IPCC) projects that global sea level may increase by as much as 88 cm relative to 1990 levels by 2100, with most model scenarios indicating between 30 and 50 cm of SLR by 2100 (ref. 24; http://www.ipcc.ch/). If these latter IPCC projections are correct, the rate of global SLR may be close to 5 mm/year by 2100. With the addition of between 0.5 and 1 mm/year of subsidence resulting from glacio-isostatic adjustment (25–27), SLR rates in southern New England may be over 6 mm/year by 2100. For coastal wetlands to continue to survive, surface elevation must increase at rates equal to or greater than long-term rates of SLR. With rates of SLR in the region over the last 100 to 150 years of close to 3 mm/year, portions of New England salt marshes dominated by marsh hay and spike grass are accreting at between 2.0 and 2.5 mm/year and are experiencing an accretion deficit with respect to changing sea level. The accretion deficit results in increased flooding frequency and lower oxygen levels in the substrate, stressing high-marsh species such as marsh hay and spike grass. Cordgrass, with its ability to oxygenate the substrate, moves up the marsh and replaces marsh hay and spike grass. Accretion rates measured in cordgrass-dominated portions of New England salt marshes are generally between 2 and 6 mm/year over the last 100 years, keeping pace with or surpassing rates of local SLR.
If current rates of SLR persist or increase only slightly over the next century in southern New England, marshes will likely continue to be transformed into cordgrass-dominated wetlands. If current projections are correct and local SLR rates of 6 mm/year or more become a reality in the coming century, cordgrass communities may drown as well, leading to significant loss of coastal wetlands in southern New England and many other regions of the world. In the face of these alterations we need to understand how the productivity of coastal wetlands may change, how changes in plant communities will impact migratory bird and other faunal populations, and how the loss of coastal wetlands will impact other coastal ecosystems.
Acknowledgments
We dedicate this paper to the memory of Bill Niering, whose insights helped stimulate our work. We thank P. Cleary, B. Shuman, J. Butler, N. Emery, and N. Logar, who contributed significantly to this research by assisting with data collection and analysis. In addition we thank three anonymous reviewers and E. Bryant, J. Mitton, L. Buss, and T. Webb III for providing useful comments on this manuscript. This work was supported by a Woods Hole Oceanographic Institute Sea Grant Program grant (to J.D.), a Mellon Foundation grant (to M.D.B.), the Narragansett Bay National Estuarine Research Reserve, and the Rhode Island Sea Grant Program.
Abbreviations
- NYC
New York City
- NH
northern hemisphere
- SLR
sea-level rise
- NCn
Nag Creek core n
- RUMn, Rumstick core n
MSL, mean sea level
Footnotes
This paper was submitted directly (Track II) to the PNAS office.
National Oceanic and Atmospheric Administration, National Ocean Service, Center for Operational Oceanographic Products and Services (2001) Verified Historical Water Level Data, Internet website (http://www.opsd.nos.noaa.gov/data_res.html). (Newport, RI-station 8452660; New York City, NY-station 8518750).
References
- 1.Mann M E, Bradley R S, Hughes M K. Nature (London) 1998;392:779–787. [Google Scholar]
- 2.Pollack H N, Huang S, Shen P Y. Science. 1998;282:279–281. doi: 10.1126/science.282.5387.279. [DOI] [PubMed] [Google Scholar]
- 3.Gehrels W R. Quat Res. 1999;52:350–359. [Google Scholar]
- 4.Karieva P, Lawton J H, Huey R. Biotic Interactions and Global Change. Sunderland, MA: Sinauer; 1993. [Google Scholar]
- 5.Nuttle W K, Brinson M M, Cahoon D, Callaway J C, Christian R R, Chmura G L, Conner W H, Day R H, Ford M, Grace J, et al. Eos Trans Am Geophys Union. 1997;78:260–261. [Google Scholar]
- 6.Baumann R H, Day J W, Jr, Miller C A. Science. 1984;224:1093–1095. doi: 10.1126/science.224.4653.1093. [DOI] [PubMed] [Google Scholar]
- 7.Wray R D, Leatherman S P, Nicholls R J. J Coastal Res. 1995;11:1195–1203. [Google Scholar]
- 8.Niering W A, Warren R S. BioScience. 1980;30:301–307. [Google Scholar]
- 9.Bertness M D, Ellison A M. Ecol Monogr. 1987;57:129–147. [Google Scholar]
- 10.Bertness M D. Ecology. 1991;72:138–148. [Google Scholar]
- 11.Teal J M, Kanwisher J W. J Exp Bot. 1966;17:355–361. [Google Scholar]
- 12.Redfield A C. Science. 1965;147:50–55. doi: 10.1126/science.147.3653.50. [DOI] [PubMed] [Google Scholar]
- 13.Warren R S, Niering W A. Ecology. 1993;74:96–103. [Google Scholar]
- 14.Orson R A, Howes B L. Estuarine Coastal Shelf Sci. 1992;35:453–471. [Google Scholar]
- 15.Donnelly J P, Webb T, III, Prell W L. Curr Topics Wetland Biogeochem. 1999;3:152–160. [Google Scholar]
- 16.Levine J, Brewer S J, Bertness M D. J Ecol. 1998;86:285–292. [Google Scholar]
- 17.Lanesky D E, Logan B W, Brown R G, Hine A C. J Sediment Petrol. 1979;49:654–657. [Google Scholar]
- 18.Niering W A, Warren R S, Weymouth C G. Conn Arbor Bull. 1977;22:12. [Google Scholar]
- 19.Pennington W, Cambray R S, Fisher E M. Nature (London) 1973;242:324–326. doi: 10.1038/242324a0. [DOI] [PubMed] [Google Scholar]
- 20.Faure G. Principles of Isotope Geology. 2nd Ed. New York: Wiley; 1986. pp. 374–377. [Google Scholar]
- 21.Dean W E. J Sediment Petrol. 1974;44:242–248. [Google Scholar]
- 22.Cahoon, Donald R, Reed, Denise J, Day J W., Jr Mar Geol. 1995;128:1–9. [Google Scholar]
- 23.Redfield A C, Rubin M. Proc Natl Acad Sci USA. 1962;48:1728–1735. doi: 10.1073/pnas.48.10.1728. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 24.Intergovernmental Panel on Climate Change. Climate Change 2001: The Scientific Basis. New York: Cambridge Univ. Press; 2001. Chapter 11, p. 892. [Google Scholar]
- 25.Davis J L, Mitrovica J X. Nature (London) 1996;379:331–333. [Google Scholar]
- 26.Peltier W R. Geophys Res Lett. 1996;23:717–720. [Google Scholar]
- 27.Donnelly JP. Geores Forum. 1998;3–4:393–400. [Google Scholar]