

Abstract
A case of intergeneric hybridization in the wild between a female bottlenose dolphin (Tursiops truncatus) and a short-beaked common dolphin (Delphinus delphis), considered members of ‘vulnerable’ and ‘endangered’ subpopulations in the Mediterranean, respectively, by the International Union of Conservation of Nature is described in this paper. The birth of the hybrid was registered in the Bay of Algeciras (southern Spain) in August 2016, and the animal has been tracked on frequent trips aboard dolphin-watching platforms. This unique occurrence is the result of an apparent ongoing interaction (10 years) between a female bottlenose dolphin and common dolphins. The calf has a robust body with length similar to Tursiops, while its lateral striping and coloration are typical of Delphinus. It displays the common dolphin’s ‘criss-cross’ pattern. However, the thoracic patch is lighter than in D. delphis and its dorsal area is light grey, with a ‘V’ shape under the dorsal fin. This paper also provides a comprehensive mini-review of hybridizations of T. truncatus with other species.
Introduction
The Bay of Algeciras, located in the south of Spain (Fig 1), hosts an important population of common dolphins (Dephinus delphis) which, since 2003, are considered ‘Endangered’ in the Mediterranean Sea according to the Red List criteria by the International Union of Conservation of Nature (IUCN) [1] and also ‘Vulnerable’ according to the Spanish National Catalogue of Endangered Species [2]. This area has been considered a feeding and breeding ground for this species [3, 4]. Also, it is possible to observe, more sporadically, groups of bottlenose dolphin (Tursiops truncatus), a species which is also considered as a ‘Vulnerable’ Mediterranean subpopulation by the IUCN. Striped dolphins (Stenella coeruleoalba) are occasionally detected (‘Vulnerable’ in the Mediterranean by IUCN) mixing with common dolphin, but the groups are mainly formed by mothers, calves and immature juveniles.
Fig 1. Study area, Bay of Algeciras.
This map has been elaborated using GIS software ArcGIS 10.4.
The three species involved in this study D. delphis, S. coeruleoalba and T. truncatus, included in the clade Delphininae [5], share the Bay of Algeciras in sympatric coexistence. They have a wide geographical distribution (the Atlantic, Pacific and Indian Oceans) and ‘can tolerate lower water temperatures and may occupy higher latitudes’ [6, 7]. Furthermore, it is known from fossil records that these species already belonged to three different genera in the Pleistocene, an epoch after which they may have diverged [8]. T. truncatus populations from the Black Sea (eastern Mediterranean) and Scotland (north-eastern Atlantic Ocean) [9] showed important genetic differences, which also exist between western Mediterranean and the Atlantic Ocean (Galicia and Portugal) ‘supporting evidences of a genetic boundary at the Almeria-Orán front’ [10]. Furthermore, genetic distinctions have been also detected between the Atlantic and Mediterranean stock in S. coeruleoalba and D. delphis species [11–16].
Common and bottlenose dolphins overlap ranges in temperate and tropical waters [17], although aggressive behaviours in bottlenose dolphins towards smaller species in different locations have been described [18]. According to observations in the Bay of Algeciras, mixed groups of both species have never before been recorded locally. However, an ongoing intergeneric interaction between a lone female bottlenose dolphin, commonly known as ‘Billie’ and groups of common dolphins has been observed since 2006.
The bottlenose dolphin was identified among the common dolphins due to morphologic differences in size and coloration: she was bigger than the common dolphins, with a robust body, a falcate dorsal fin, of light grey coloration to darker grey dorsally, and showed a light blaze marking on her sides. A well marked demarcation at the end of the melon, convex flippers and a short and stubby beak [17] were also recognisable features in the individual. The gender (female) was supported by photographic evidence.
Before 2016, Billie was detected three times assisting common dolphin births, leading newborns to the surface and offering alloparental care for only a few minutes after labour and always accompanied by other common dolphins. Allomaternal care often occurs among bottlenose dolphins [19, 20] and has been described in captivity and in the wild [21, 22]. However, it is uncommon to observe a bottlenose dolphin calf adopted by a common dolphin, although it has been previously reported in the northern Adriatic waters [23].
On 11 August 2016, Billie was observed raising and pushing to the surface a neonate (Fig 2A). Visible features such as foetal folds (vertical depigmented lines) were visible on each flank of the individual, as well as a bent dorsal fins and curled flukes, produced by the neonatal posture in the uterus during the gestation period [24]. No other dolphins were present. In view thereof, and taking into account the observations mentioned above, a hybridization was considered. It was possible to compare photographs before and after Billie´s pregnancy, showing weight gain and grosser bodyshape while pregnant (Fig 2B). Billie showed a much slimmer and thinner bodyshape when she was not pregnant (Fig 2C).
Fig 2. Billie raising the newborn to the surface, and comparison of the bodyshape of the pregnant/not pregnant female bottlenose dolphin.

(A) On 11 August the female bottlenose dolphin was observed holding and pushing a newborn to the surface. The newborn showed folded fins and marked foetal folds. (B) Billie pregnant (28 July 2016). (C) Billie not pregnant (27 May 2017).
Cetaceans exhibit surprising karyotype uniformity, suggesting that they have a higher potential to produce hybrid offspring than do other mammals [25–27]. However, the identification of cetacean hybrids in the wild is difficult, and the molecular evidence of wild cetacean hybrids is extremely limited [28]. This is not to say that these hybridizations in mammals are uncommon and do not occur in various marine and terrestrial species [29–31].
Marine mammal hybridizations are very difficult to detect in comparison to other taxonomic groups of animals and plants, although several examples in captivity and in the wild have been reported [32–35]. Intergeneric or interspecific reproduction is more likely to happen between species when their habitat ranges overlap, when different species have the same habits or similar social behaviours. Hybridization can also be a result of particular and unique life histories developed within dolphin groups [36], as had resulted in this particular case, where a single female bottlenose dolphin had been adopted by groups of common dolphins.
Methods
The study was carried out on board opportunistic dolphin-watching platforms (14 m and 12 m in length), offering trips of 90 min with consistent daily itineraries from August 2016 until May 2017. Once the group of interest (the one including the mother/hybrid pair) was detected, standardized data were gathered (weather permitting), such as date, time, GPS position, structure and group size. Group composition data were collected using a combination of sampling methods. Individual-following protocols [37, 38], focusing on the mother/hybrid pair were applied during the sightings. The 10 m chain rule [39] was also applied: the pair were determined to be together if they were less than 10 m apart. Swimming positions, general behaviour and body-contact events were also gathered when they were displayed, irrespective of the time. Sea surface temperature was measured from the side of the boat, using a digital thermometer with 0.1°C graduations [40]. The mother/hybrid pair and other dolphins were photographed (Nikon DSLR camera, Nikon 70–300 mm lens) for re-identification and also for morphological analyses of the presumed hybrid [41]. In some cases, images were slightly retouched (descriptors: saturation, contrast, exposition, clearness and shades), with Adobe Photoshop Lightroom software, to improve the display of the morphological features described in the text. A plotter map was elaborated using ArcGIS 10.4 software, including coordinates of the mother/hybrid pair.
Tissue sampling by means of biopsy dart was not attempted as it was considered invasive and inappropriate due to the immaturity of the calf [42]. It is well known that newborns obtain temporary immunological protection from maternal antibodies, and the immune system of many mammalian species is not fully developed at birth [43]. Skin swabbing [44] was also considered for genetic analysis, but cautious measures were taken, ensuring that the calf was at least one year of age before the tests were attempted.
Data regarding the group composition of the species involved were collected over 25 weeks (11 August 2016 to 29 May 2017). Sightings were classified according to 19 descriptors. A box plot analysis (using SPSS 15 statistical software IBM, New York, NY, USA), was applied to analyse the behaviour of the mother/hybrid pair, comparing the composition and the frequency they were found in mixed-species groups, separated or alone.
This study was carried out in strict accordance with the cetacean protocol included in the Marine Regulations, 2014 and has been approved by the Committee on the Ethics of Animal Experiments of the Ministry for Education, Heritage, Environment, Energy and Climate Change of Gibraltar.
Results
After the first sighting on 11 August 2016, re-sighting took place on 17 August 2016, after which they were seen on an almost daily basis mixing with ‘nursery groups’ of common dolphins (D. delphis) (Fig 3A and 3B). Data were collected between 17 August 2016 and 4 June 2017. The pair was observed 113 times (57 h 11 min of observation) in a total of 355 sightings. Of these, 104 times (53 h 55 min) the pair was found within nursery groups of common dolphins formed by females and calves [45], twice (1 h 23 min) in mixed nursery groups of common dolphins accompanied by mothers, calves and immature juveniles of striped dolphins (S. coeruleoalba) and in only six sightings (1 h 53 min) was the pair sighted alone, distanced from the common dolphins (minimum 500 m between groups). The pair were detected together less than 10 m apart in 112 sightings (99.1%); 1 occasion (0.83%) was Billie (female T. truncatus) separated from the hybrid by 100 m, both of them accompanied by common dolphins. Sea surface temperature (SST) during these observations was an average of 19.35°C (66.83°F) with minimum of 14°C (57.2°F) and maximum of 26°C (78.8°F). From 2 June 2017 until the end of the year, the hybrid was not sighted again, leading to the reasonable suspicion of death.
Fig 3. Mixed group of mother/hybrid pair with common dolphins.

(A) The mother/hybrid pair and an adult common dolphin spotted on 6 October 2016; (B) Hybrid displaying jumps alongside a common dolphin separated by more than 10 m from the presumed mother on 4 September 2016.
Table 1 presents every sighting recorded during the campaign classified according to the group composition represented by 19 different descriptors, depending whether the mother/hybrid pair was separated, together, alone or mixed with other species.
Table 1. Data regarding species group composition collected over 25 weeks (period 11 August 2016 to 29 May 2017).
Sightings were classified according to 19 descriptors: (A1) Common dolphins only; (A2) Striped dolphins only; (A3) Bottlenose dolphins only; (B1) Common dolphins with mother/hybrid pair together; (B2) Striped dolphins with mother/hybrid pair together; (B3) Bottlenose dolphins with mother/hybrid pair together; (C1) Common dolphins with mother/hybrid pair not together; (C2) Striped dolphins with mother/hybrid pair not together; (C3) Bottlenose dolphins with mother/hybrid pair not together; (D1) Common dolphins with one of the mother/hybrid pair (mother or hybrid); (D2) Striped dolphins with one of the mother/hybrid pair (mother or hybrid); (D3) Bottlenose dolphins with one of the mother/hybrid pair (mother or hybrid); (E1) Mother/hybrid pair only; (E2) Mother only; (E3) Hybrid only; (F1) Common mixed with striped dolphins; (F2) Common mixed with bottlenose dolphins; (F3) Bottlenose mixed with striped dolphins; (F4) Common and striped dolphins mixed with mother/hybrid pair.
| Week | A1 | A2 | A3 | B1 | B2 | B3 | C1 | C2 | C3 | D1 | D2 | D3 | E1 | E2 | E3 | F1 | F2 | F3 | F4 |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| 1 | 1,286 | 0,143 | 0,571 | 1,286 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| 2 | 2,4286 | 0 | 0 | 2,286 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0,286 | 0 | 0 | 0 | 0 |
| 3 | 2 | 0 | 0,2 | 1,2 | 0 | 0 | 0 | 0 | 0 | 0,2 | 0 | 0 | 0,2 | 0 | 0 | 0,2 | 0 | 0 | 0 |
| 4 | 2,667 | 0 | 0,667 | 1,667 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0,167 | 0 | 0 | 0 | 0 | 0 | 0 |
| 5 | 2,333 | 0 | 0 | 1 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0,667 | 0 | 0 | 0 |
| 6 | 2 | 0 | 0 | 1,143 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0,167 | 1,423 | 0 | 0 | 0 | 0 | 0 |
| 7 | 1,5 | 0 | 0 | 0,667 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0,333 | 0 | 0 | 0 |
| 8 | 1,333 | 0 | 0,167 | 1,333 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0,167 |
| 9 | 2 | 1,1667 | 0,1667 | 0,833 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0,167 | 0 | 0 | 1,167 | 0 | 0 | 0 |
| 10 | 1,833 | 0,333 | 1 | 0,833 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0,167 | 0 | 0 | 0 |
| 11 | 1,333 | 0 | 0,5 | 0,167 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0,167 | 0 | 0 | 0,167 | 0 | 0 | 0,167 |
| 12 | 1,143 | 0 | 0,286 | 0,571 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| 13 | 1 | 0 | 0 | 0,833 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| 14 | 1 | 0 | 0 | 1,2 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| 15 | 1,25 | 0,25 | 0 | 0,75 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| 16 | 1 | 0 | 0 | 0,6 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| 17 | 1 | 0 | 0 | 0,667 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| 18 | 1 | 0 | 0 | 0,25 | 0 | 0 | 0,25 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0,25 | 0 | 0 | 0 |
| 19 | 0,75 | 0,25 | 0 | 1 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| 20 | 0,667 | 0 | 0 | 0,333 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0,333 | 0 | 0 | 0 | 0 | 0 | 0 |
| 21 | 0,571 | 0,714 | 0,143 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0,143 | 0 | 0 | 0 |
| 22 | 1,5 | 0,25 | 0,5 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| 23 | 3 | 0 | 0,25 | 0 | 0 | 0 | 0 | 0 | 0 | 1,5 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| MEAN | 1,5041130 | 0,1350739 | 0,193508 | 0,8095217 | 0 | 0 | 0,0108695 | 0 | 0 | 0,0739130 | 0 | 0 | 0,0522173 | 0,0618695 | 0,0124347 | 0,1345217 | 0 | 0 | 0,014521 |
| SD | 0,6636513 | 0,2836602 | 0,275524 | 0,5628901 | 0 | 0 | 0,052128 | 0 | 0 | 0,3136549 | 0 | 0 | 0,0952823 | 0,2967160 | 0,0596351 | 0,2763548 | 0 | 0 | 0,048113 |
The female bottlenose dolphin was observed showing continuous epimeletic and nurturant behaviour towards the newborn, offering care and protection and exhibiting near-body contact for the first three months of observation. The two main swimming positions for calves and their mothers are defined as ‘echelon position’ (the calf swimming alongside the mother) and ‘infant position’ (the calf swimming under the mother) [19]. The hybrid was observed in the echelon position most of the time; in the infant position on only two occasions, when the pair approached to bow-ride during the study period. At this time, the young calf still showed clear foetal folds [46].
Calves often show rubbing behaviour with their mothers, with particular focus on her head region [19]. Body-contact events such as flipper–belly, flipper–flipper, forehead–belly, head and beak rubbing, blowhole rubbing, back-to-back calf jumping backwards over the mother’s back) and petting were observed between Billie and the hybrid, and were considered typical behaviour displays between a mother and her calf [47, 48]. Mother chasing towards the newborn and vice versa were also detected, which is shown to be indicative of an imprinting period [19]. When the calf´s rostrum was in contact with the mother’s mammary slit area for longer than 2 s [19] it was recognised as a nursing event and was recorded at least four times on 19, 23 and 27 August 2016 and 2 September 2016.
The mother/hybrid pair showed normal breathing and developmental patterns and close swimming positions until 4 September 2016, when at 24 days the neonate was observed breaching and swimming in echelon position alongside an adult common dolphin (Fig 3B). At the time Billie was observed displaying feeding behaviour among other common dolphins. After this event, the calf returned to its mother’s side.
From photographs, morphological features of the presumed hybrid offspring were compared with both common dolphin (D. delphis) and bottlenose dolphin (T. truncatus) (Table 2), and characteristics of both species were confirmed in the neonate. In addition, no mammary slits were observed, and a separation between the genital and anal slits was documented, suggesting the neonate was male [49].
Table 2. Comparison of morphological features between species.
Morphological features of common dolphins (D. delphis), bottlenose dolphin (T. truncatus) (Mediterranean population) and the presumed hybrid, recorded in the Bay of Algeciras, south of Spain.
| Delphinus delphis | Tursiops truncatus | Presumed Hybrid | |
|---|---|---|---|
| Size at birth | 0.76–0.86 m [50] | 0.9–1.3 m, 32 kg [50] | 1.25 m (Estimated) |
| Body shape | Slender [51] | Robust [51] | Robust [51] |
| Beak | Long beak sharply demarcated from the melon [52] | Rounded forehead/ marked creased beak. Short and stubby beak [52] | Rounded forehead/ marked creased, short and stubby beak [35] |
| Body colour pattern | Criss-cross pattern; brownish/black back, ‘V’ shape under dorsal fin; yellowish thoracic patch; light grey posterior patch; white belly. [51, 52] | Light grey to black dorsally and laterally; light belly; light blaze or brush marking sometimes observed on their flanks.[51, 52] | Medium/dark grey back; pale creamy flanks from eye to peduncle; white belly and post-pectoral patches. |
| Dark flipper-to-anus stripe parallel to the lower margin of the cape; dark flipper stripe joining the lip patch on the underside of the beak. [35–51] | Light creamy yellow/greyish stripe from the back of the eye to its posterior flanks. | ||
| Dorsal fin | Tall/moderate falcate | Falcate | Falcate |
| Clear patch sometimes [35, 51] | Dark. [35, 51] | Dark | |
| Mouth-to-flipper stripe | Present | Absent | Present |
According to Whitehead and Mann [53], Tursiops spp. Neonates are 1.1 m, while in common dolphins 0.8 m. The presumed hybrid was less than half the size of the female bottlenose at birth (approximately 1.25 m), and remained in echelon position close to her most of the time, characteristics consistent with a newborn. By the middle of November, the animal was just over half the length of Billie, and therefore considered an infant or calf [19, 38, 40]. The neonate’s beak was short and stubby, with a round melon and robust body, showing more similarities to the bodyshape of a bottlenose dolphin than to that of a common dolphin [24, 40, 51].
Coloration and striping patterns were examined from photographs. A typical ‘criss-cross’ coloration along the neonate’s thorax and flank was detected, corresponding to markings characteristic of common dolphins (Fig 4A). A pale creamy-coloured patch ran from the low melon/rostrum/eye along the thorax, which faded at the light grey posterior flank patch (Fig 4B and 4C). The hybrid had a flipper stripe, which is characteristic of D. delphis, although in this case it was light brown in colour from the anterior insertion of the flipper to the lower jaw and gape (Fig 4D). Two other stripes were identified: one that ran from the caudal canthus of the eye to the anterior insertion of the flipper and a second above the flipper stripe, from the caudal canthus of the eye to its flank. Both were slightly darker in colour (Fig 4C and 4D). Also, it showed a white patch between the dorsal and ventral stripe (Fig 4C and 4D), which has also been observed in other bottlenose dolphins neonates in the bay (Fig 5A–5D). The neonate’s sides were light grey, with a V-shaped pattern on its side under the dorsal fin. The dorsal fin was bigger and wider than in common dolphins and grey, becoming much lighter over time (Fig 4E). The ventral side was white.
Fig 4. Morphological features as coloration and shape patterns of the presumed hybrid.

(A) muted ‘criss-cross’ pattern typical of common dolphins, photographed 5 November 2016. (B) Creamy yellow patch on its side documented 7 February 2017; (C) light grey posterior flank on 20 February 2017; (D) Striped pattern on 20 February 2017; (E) Comparison between potential hybrid (top right) and common dolphin calf (bottom left). A similar ‘V’ shape and light-coloured dorsal fin can be observed in these dolphins. Documented on 20 February 2017.
Fig 5. Coloration features displayed by D. delphis, T. truncatus, S. coeruleoalba and the hybrid calves.

Notice the similarities of colour patterns between (A) D. delphis, (B) T. truncatus and the hybrid (D). These similarities are absent when comparing features of (C) S. coeruleoalba (bluish-grey dorsally, white to light grey blaze on the flanks, eye-to-anus stripe that runs ventrally [55]) and the hybrid.
Dolphin neonates show lines across their flanks and backs called foetal folds [54]. By 21 December 2016, the hybrid no longer showed foetal folds so it was considered an infant.
Data analysis from Table 1 is represented in Fig 6. Only the descriptors A1, A2, A3, B1, E1 and F1 resulted in valuable information, thus making it possible to compute the median (robust measure of central tendency, independent from the extreme scores). The other variables registered exceptional or no sightings. Groups of exclusively common dolphins (A1) were those most often spotted in the area. When the mother/hybrid pair were detected included in mixed-species groups, this was mostly with nursery groups of common dolphins (B1). A third species (striped dolphin) was located in the mixed groups on an exceptional basis, but they always proved to be mothers, with immature juveniles and calves (F4). The pair was detected alone (E1) on a few occasions, at a distance of more than 500m from the groups of common dolphin.
Fig 6. Box plot analysis exposing medians, quartiles and confidence interval bars (95%) of data reported in Table 1, referred to 19 types (descriptors A1, A2 …F4) of sighting.
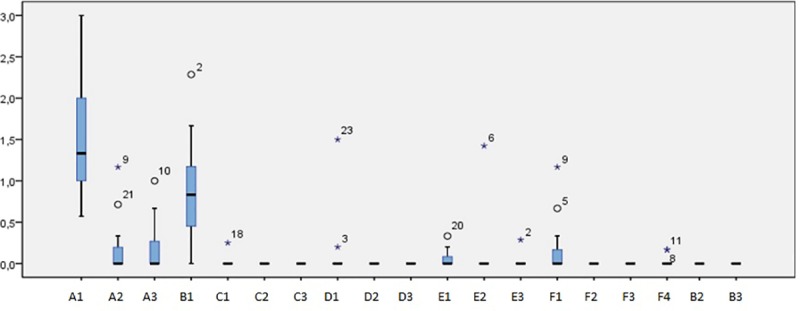
The extreme values (asterisks) are those that were more than three times the interquartile range from Q3. Outliers (white circles) are those that were located between 1.5 and 3 times the interquartile range from Q3. The figure clearly shows the close relationship of Billie and the newborn hybrid with common dolphins (B1), their relationships being completely nil with striped dolphins (A2) and with bottlenose dolphins (B3). The number of sightings of Billie and the newborn hybrid together in the absence of other dolphins also deserves mention (E1).
Discussion
There is little information about hybrids in the wild; therefore, the significance of this potential hybridization is two-fold. First, this event between these species in the wild supports what has been observed in the non-natural conditions of captivity. Second, species such as T. truncatus and D. delphis, with spatially overlapping habitats [17], have rarely been recognised as interbreeding until now. Moreover, this type of intergeneric interaction occurs at a low level, as although the habitats of the two species described overlap, they rarely mix.
On the other hand, hybridization events in Delphinidae in captivity have been reported multiple times, and T. truncatus hybrids have been described interbreeding with several species (Table 3), but this event (Stenella frontalis × T. truncatus) has only been observed once in the wild [56, 57]. In captivity, intergeneric hybridization was produced by a cross between T. truncatus and several other species including Delphinus capensis [35, 58], which resulted in four hybrids. Two of the calves died, but a living fertile female back-crossed with a T.truncatus, and the calf didn’t survive either. In 2018, Gridley, reported multiple intra-generic matings between T. truncatus and Tursiops aduncus producing a health F1 hybrid, which survived to adulthood and also produced back-crossed hybrid offspring [59].
Table 3. Registry table of hybridization between individuals of bottlenose dolphins (T. truncatus) in captivity and in the wild.
| Parental species | Number of hybrids F1 | References | Environment |
|---|---|---|---|
| T. truncatus x Grampus griseus | 3 | [62] | Captivity |
| Globicephala macrorhynchus x T. truncatus | 2 | [58, 63] | Captivity |
| Steno bredanensis x T. truncatus | 1 | [64] | Captivity |
| T. truncatus x Pseudorca. Crassidens | 6 | [58, 65] | Captivity |
| Llagenorhynchus obliquidens x T. truncatus | 1 | [66] | Captivity |
| T. truncatus x G. griseus | 13 | [66, 67, 68, 69] | Captivity |
| T. truncatus x S. frontalis | 1 | [56, 57] | Wild |
| T. truncatus x D. capensis | 4 | [35] | Captivity |
| S. guianensis x T. truncatus | 1 | [70] | Captivity |
| T. truncatus x T. aduncus | 7 | [59] | Captivity |
| T. truncatus x D. delphis | 1 | This paper | Wild |
According to morphological [71, 52] and genetic [72, 73] studies, S. coeruleoalba and Delphinus have a closer phylogenetic relationship, being more closely related to each other than to T. truncatus. Furthermore, the greatest number of interactions between the three species cited have been observed between D. delphis and S. coeruleoalba [74–81]. In fact, D. delphis and S. coeruleoalba coexist in sympatry in three different areas of the Mediterranean, including the Alborán sea [79–81]. In addition, ‘S. coeruleoalba displayed more opportunistic trophic habits compared with D. delphis’ in the north of Spain (Bay of Biscay) [78]. ‘Fission-fusion grouping patterns’ have been described between T. truncatus and D. delphis [82, 83], depending on the distribution and availability of food sources. Furthermore, in the eastern Ionian Sea, when both species coexist in ‘direct sympatry’ [81], habitat partition results [82, 84]. A niche separation has been suggested that might have reduced the direct food-base competition [82] in such species, observing ‘different foraging strategies, with D. delphis feeding in the water column or near the surface and T. truncatus focusing on bottom prey’ [85]. This has been also observed in the Bay of Algeciras.
Accordingly, the probability of hybridization of D. delphis with S. coeruleoalba was expected to be higher than with T. truncatus. However, interactions between D. delphis and T. truncatus [82, 86] are well known, as are sympatric associations between the species, and according to the sympatry concept [81], ‘the co-occurrence of two or more dolphin species in the same immediate habitat’ [81, 82, 86] can increase the possibility of hybridization. Also to be taken in consideration is the high level of promiscuity of T. truncatus and their potentiality to produce hybrids with up to ten different genera of dephinids (Table 3). All factors mentioned above strongly support that the hybrid described in this paper is the result of at least 10 years of integration of Billie into groups of. D. delphis. This is corroborated in Table 1, which shows that Billie was mixing to a negiglible degree with mothers and sexually immature S. coeruleoalba calves.
Despite the uniqueness of this hybridization, DNA samples from the hybrid were not obtained. Considerations were that the rare, but extremely dangerous experiences during cetacean sampling [87, 88], the death of a common dolphin while being sampled by a dart [89], and, most importantly, the early and delicate developmental stage of the calf, made taking a biopsy too risky.
The Bay of Algeciras is a heavily anthropised area, but it serves as a feeding, nursing and breeding ground for cetaceans, including both common and bottlenose dolphins and future hybrids. Enforcement of the cetacean observation protocols and the introduction of an environmental education plan to minimise the impacts on cetaceans in the Bay of Algeciras are vital. In this regard, conservation measures have already been proposed for this hotspot area for cetaceans facing detrimental threats [4].
Acknowledgments
We would like to thank the entire team of Dolphin Adventure for donating their platforms that made possible to conduct this project. We would also like to thank Jennifer Carlin and Alessia Scuderi for their collaboration in the data processing. Finally, we would like to mention Dr. Rosenfeld (Academic Editor PLOS ONE) and the reviewers of this paper, especially Dr. Duffield (who kindly identified herself), for their valuable recommendations, which improved the contents of this work.
Data Availability
All relevant data are within the paper.
Funding Statement
This research was supported by the University of Seville providing salary to the author JCG-G and Research foundation of University of Seville (project FIUS18/0014) providing salaries to LO-P, supporting with logistics, material and management. RE as Honorary collaborator of the department of Zoology (University of Seville). Dolphin Adventure provided payed personnel [LH, RE, EM] and research platforms. The funding organisations did not have any additional role in the study design, data collection, analysis, decision to publish, or preparation of the manuscript. The specific roles of these authors are articulated in the 'authors contributions' sections.
References
- 1.Bearzi G. Delphinus delphis (Mediterranean subpopulation). In: IUCN Red List of Threatened Species (Version 2011.2. www.iucnredlist.org); 2003. [Google Scholar]
- 2.BOE, 2011. Real Decreto 139/2011, de 4 de febrero, para el desarrollo del listado de Especies Silvestres en Régimen de Protección Especial y del Catálogo Español de Especies Amenazadas.
- 3.Giménez J, Gauffier P, Verborgh P, Esteban R, Jiménez-Torres C, de Stephanis R. The Bay of Algeciras: a feeding and breeding ground for common dolphins? Abstract Book 25th Conference of European Cetacean Society, Cádiz, Spain; March 2011.
- 4.Espada R, Martín E, Haasova L, Olaya-Ponzone L, García-Gómez JC. Permanente presencia del delfín común en la bahía de Algeciras. Hacia un plan de gestión, vigilancia y conservación de la especie. Almoraima. Revista de Estudios Campogibraltareños. 2018; 49. [Google Scholar]
- 5.LeDuc RG, Perrin WF, Dizon AE. Phylogenetic relationships among the delphinid cetaceans based on full cytochrome d sequences. Mar. Mammal Sci. 1999; 15: 619–648. [Google Scholar]
- 6.Forcada J. Distribution. In: Perrin WF, Würsig B, Thewissen JGM, editors. Encyclopedia of Marine Mammals. Academic Press Inc., San Diego; 2009. pp. 316–321. [Google Scholar]
- 7.Leduc RG. Delphinids, Overview In: Perrin WF, Würsig B, Thewissen JGM editors. Encyclopedia of Marine Mammals, 2nd ed. Academic Press, San Diego; 2009. b. pp. 298–302. [Google Scholar]
- 8.do Amaral KB, Amaral AR, Fordyce RE, Moreno IB. Historical biogeography of delphininae dolphins and related taxa (Artiodactyla: Delphinidae). J Mamm Evol. 2016: 1–19. [Google Scholar]
- 9.Natoli A, Birkun A, Aguilar A, Lopez A, Hoelzel AR. Habitat structure and the dispersal of male and female bottlenose dolphins (Tursiops truncatus). Proc. R. Soc. Lond., B, Biol. Sci. 2005; 272(1569): 1217–1226. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 10.Guidarelli G, Colangelo P, de Francesco MC, Nicolosi P, Meloro C, Loy A. Phenotypic Changes Across a Geographic Gradient: The Case of Three Sympatric Dolphin Species. Evol. Biol. 2018; 45(1): 113–125. [Google Scholar]
- 11.Garcia‐Martinez J, Barrio E, Raga JA, Latorre A. Mitochondrial DNA variability of striped dolphins (Stenella coeruleoalba) in the Spanish Mediterranean waters. Mar. Mammal Sci. 1995; 11(2): 183–199. [Google Scholar]
- 12.Garcia‐Martinez J, Moya A, Raga JA, Latorre A. Genetic differentiation in the striped dolphin Stenella coeruleoalba from European waters according to mitochondrial DNA (mtDNA) restriction analysis. Mol. Ecol. 1999; 8(6): 1069–1073. [DOI] [PubMed] [Google Scholar]
- 13.Valsecchi E, Amos W, Raga JA, Podestà M, Sherwin W. The effects of inbreeding on mortality during a morbillivirus outbreak in the Mediterranean striped dolphin (Stenella coeruleoalba). Anim. Conserv. 2004; 7(2): 139–146. [Google Scholar]
- 14.Gaspari S, Azzellino A, Airoldi S, Hoelzel AR. Social kin associations and genetic structuring of striped dolphin populations (Stenella coeruleoalba) in the Mediterranean Sea. Mol. Ecol. 2007; 16(14): 2922–2933. 10.1111/j.1365-294X.2007.03295.x [DOI] [PubMed] [Google Scholar]
- 15.Bourret VJ, Macé MR, Crouau-Roy B. Genetic variation and population structure of western Mediterranean and northern Atlantic Stenella coeruleoalba populations inferred from microsatellite data. J. Mar. Biol. Assoc. U. K. 2007; 87(1): 265–269. [Google Scholar]
- 16.Natoli A, Canadas A, Vaquero C, Politi E, Fernandez-Navarro P, Hoelzel AR. Conservation genetics of the short-beaked common dolphin (Delphinus delphis) in the Mediterranean Sea and in the eastern North Atlantic Ocean. Conserv. Genet. 2008; 9(6): 1479. [Google Scholar]
- 17.Wells RS, Scott MD. Bottlenose dolphin Tursiops truncatus (Montagu, 1821). Handbook of Marine Mammals: the second book of dolphins and porpoises. 1999; 6: 137–182. [Google Scholar]
- 18.Patterson IAP, Reid RJ, Wilson B, Grellier K, Ross HM, Thompson PM. Evidence for infanticide in bottlenose dolphins: an explanation for violent interactions with harbour porpoises?. Proc R Soc Lond B Biol Sci. 1998; 265(1402):1167–1170. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 19.Mann J, Smuts BB. Natal attraction: allomaternal care and mother–infant separations in wild bottlenose dolphins. Anim Behav. 1998; 55(5):1097–1113. 10.1006/anbe.1997.0637 [DOI] [PubMed] [Google Scholar]
- 20.Howells EM, Reif JS, Bechdel SE, Murdoch ME, Bossart GD, McCulloch SD, et al. A novel case of non-offspring adoption in a free-ranging Atlantic bottlenose dolphin (Tursiops truncatus) inhabiting the Indian River Lagoon, Florida. Aquat Mamm. 2009; 35(1):43. [Google Scholar]
- 21.Tavolga M., Essapian FS. The behavior of the bottle-nosed dolphin (Tursiops truncatus)-mating, pregnancy, parturition and mother-infant behavior. In Anatomical Record. 1955; 122(3):426–426). [Google Scholar]
- 22.Wells RS. The role of long-term study in understanding the social structure of a bottlenose dolphin community. Dolphin Societies: Discoveries and puzzles. 1991; 199–225. [Google Scholar]
- 23.Bearzi G. A ‘remnant’ common dolphin observed in association with bottlenose dolphins in the Kvarneric (northern Adriatic Sea). Eur. Res. Cet. 1996; 10:204. [Google Scholar]
- 24.McBride A, Kritzler H. Observations on Pregnancy, Parturition, and Postnatal Behavior in the Bottlenose Dolphin. J. Mammal. 1951; 32(3): 251–266. 10.2307/1375657 [DOI] [Google Scholar]
- 25.Árnason O. The karyotype of the fin whale.–Hereditas. 1969; 62: 273–284. [DOI] [PubMed] [Google Scholar]
- 26.Árnason O. The role of chromosomal rearrangement in mammalian speciation with special reference to Cetacea and Pinnipedia. Hereditas. 1972; 70: 113–118. [PubMed] [Google Scholar]
- 27.Árnason O. Karyotype stability in marine mammals.-Cytogenet. Cell Genet. 1982; 33: 274–276. [DOI] [PubMed] [Google Scholar]
- 28.Willis PM., Crespi BJ, Dill LM, Baird RW, Hanson MB. Natural hybridization between Dall's porpoises (Phocoenoides dalli) and harbour porpoises (Phocoena phocoena). Can. J. Zool. 2004; 82(5):828–834. [Google Scholar]
- 29.Gray AP. Mammalian hybrids: a check-list with bibliography. Mammalian hybrids: a check-list with bibliography. 2nd ed. 1972. [Google Scholar]
- 30.Arnold ML. Natural hybridization and evolution. Oxford University Press; 1997. [Google Scholar]
- 31.Mallet J. Hybridization as an invasion of the genome. Trends Ecol. Evol. 2005; 20(5):229–237. 10.1016/j.tree.2005.02.010 [DOI] [PubMed] [Google Scholar]
- 32.Árnason Ú, Spilliaert R, Pálsdóttir A, Árnason Á. Molecular identification of hybrids between the two largest whale species, the blue whale (Balaenoptera musculus) and the fin whale (B. physalus). Hereditas. 1991; 115 (2):183–189. [DOI] [PubMed] [Google Scholar]
- 33.Bérubé M. Hybridism In: Perrin WF, Würsig B, Thewissen JGM (eds) Encyclopedia of Marine Mammals, 2nd ed. Academic Press, San Diego, CA: 2009; 588–592. [Google Scholar]
- 34.Crossman C, Barrett-Lennard LG, Taylor EB. Population structure and intergeneric hybridization in harbour porpoises Phocoena phocoena in British Columbia, Canada. Endanger Species Res. 2014; 26:1–12. [Google Scholar]
- 35.Zornetzer HR, Duffield DA. Captive-born bottlenose dolphin× common dolphin (Tursiops truncatus×Delphinus capensis) intergeneric hybrids. Can. J. Zool. 2003; 81(10):1755–1762. [Google Scholar]
- 36.Bérubé M. Hybridism In: Perrin WF, Würsig B, Thewissen JGM, editors. Encyclopedia of Marine Mammals. 2nd ed San Diego: Academic Press; 2002. pp. 596–600. [Google Scholar]
- 37.Mann J. Behavioral sampling methods for cetaceans: a review and critique. Mar. Mamm. Sci. 1999; 15(1):102–122. [Google Scholar]
- 38.Mann J. Cetacean societies: field studies of dolphins and whales. University of Chicago Press; 2000. [Google Scholar]
- 39.Smolker RA, Richards AF, Connor RC, Pepper JW. Sex differences in patterns of association among Indian Ocean bottlenose dolphins. Behav. 1992; 123(1): 38–69. [Google Scholar]
- 40.Schaffar-Delaney A. Female reproductive strategies and mother-calf relationships of common dolphins (Delphinus delphis) in the Hauraki Gulf, New Zealand. Master of Science thesis, Massey University, New Zealand. 2004, 221 pp. Available from: https://mro.massey.ac.nz/handle/10179/12524.
- 41.Würsig B, Jefferson TA. Methods of photo-identification for small cetaceans. In: Hammond PS, Mizroch SA, Donovan GP editors. Individual recognition of cetaceans: Use of photo identification and other techniques to estimate population parameters. Report of the International Whale Commission, Special Issue 12 1990. pp. 43–51. [Google Scholar]
- 42.Frère CH, Krzyszczyk E, Patterson EM, Hunter S, Ginsburg A, Mann J. Thar She Blows! A Novel Method for DNA Collection from Cetacean Blow. PLoS ONE. 2010; 5(8): e12299 10.1371/journal.pone.0012299 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 43.Ross PS, de Swart RL, Visser IK, Vedder LJ, Murk W, Bowen W D, et al. Relative immunocompetence of the newborn harbour seal, Phoca vitulina. Vet. Immunol. Immunopathol. 1994; 42(3–4): 331–348. [DOI] [PubMed] [Google Scholar]
- 44.Harlin AD, Würsig B, Baker CS, Markowitz TM. Skin swabbing for genetic analysis: application to dusky dolphins (Lagenorhynchus obscurus). Mar. Mamm. Sci. 1999; 15: 409–425. [Google Scholar]
- 45.Neumann DR. Activity budget of free-ranging common dolphins (Delphinus delphis) in the northwestern Bay of Plenty, New Zealand. Aquat Mamm. 2001; 27(2):121–136. [Google Scholar]
- 46.Barbara JM. Behavioral development in wild bottlenose dolphin newborns (Tursiops sp.). Behaviour. 1999; 136(5):529–566. [Google Scholar]
- 47.Cockcroft VG, Sauer W. Observed and inferred epimeletic (nurturant) behaviour in bottlenose dolphins. Aquat Mamm. 1990; 16(1):31–32. [Google Scholar]
- 48.Weinpress M. Maternal and Alloparental Discipline in Atlantic Spotted Dolphins (Stenella frontalis) in the Bahamas. Master of Science. Florida Atlantic University. 2013.Available from: https://www.researchgate.net/publication/319149315. [Google Scholar]
- 49.Neumann DR, Russell K, Orams MB, Baker CS, Duignan P. Identifying sexually mature, male short-beaked common dolphins (Delphinus delphis) at sea, based on the presence of a postanal hump. Aquat Mamm. 2002; 28(2):181–187. [Google Scholar]
- 50.Reidenberg JS, Laitman JT. Cetacean prenatal development. In: Perrin WF, Würsig B, Thewissen JGM, editors. Encyclopedia of Marine Mammals (Second Edition); 2009. pp. 220–230. [Google Scholar]
- 51.Carwardine M. Mark Carwardine's Guide to Whale Watching In Britain and Europe. Bloomsbury Publishing; 2016. [Google Scholar]
- 52.Perrin WF. Common dolphins: Delphinus delphis and D. capensis. In: Perrin WF, Würsig B, Thewissen JGM, editors. Encyclopedia of Marine Mammals (Second Edition); 2009. pp. 255–259. [Google Scholar]
- 53.Whitehead H, Mann J. Female reproductive strategies of cetaceans. In: Mann J, Connor RC, Tyack PL, Whitehead H, editors. Cetacean societies: Field studies of dolphins and whales: 2000. pp. 219–246. [Google Scholar]
- 54.Mann J,Smuts B. Behavioral development in wild bottlenose dolphin newborns (Tursiops sp.). Behaviour, 1999; 136: 529–566. [Google Scholar]
- 55.Archer II, Frederick I. Striped Dolphin: Stenella coeruleoalba. In: Perrin WF, Würsig B, Thewissen JGM, editors. Encyclopedia of Marine Mammals (Second Edition); pp. 1127–1129. [Google Scholar]
- 56.Herzing DL, Johnson CM. Interspecific interactions between Atlantic spotted dolphins (Stenella frontalis) and bottlenose dolphins (Tursiops truncatus) in the Bahamas, 1985–1995. Aquat Mamm. 1997; 23:85–100. [Google Scholar]
- 57.Herzing DL, Moewe K, Brunnick BJ. Interspecies interactions between Atlantic spotted dolphins, Stenella frontalis, and bottlenose dolphins, Tursiops truncatus, on Great Bahama Bank, Bahamas. Aquat Mamm. 2003; 29(3):335–341. [Google Scholar]
- 58.Duffield DA. Examples of captive hybridization and a genetic point of view. Page 421. In: Evans PGH, Parsons ECM, editors. World Marine Mammal Science Conference, vol 12. 1998.
- 59.Gridley T, Elwen SH, Harris G, Moore DM, Hoelzel AR, Lampen F. Hybridization in bottlenose dolphins—A case study of Tursiops aduncus× T. truncatus hybrids and successful backcross hybridization events. PloS one. 2018; 13(9): e0201722 10.1371/journal.pone.0201722 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 60.Do Nascimento Schaurich M, Vieira Lopes FR, de Oliveira LR. Revisão sobre o fenômeno da hibridização em cetáceos e pinípedes. Neotrop. Bio. & Cons. 2012; 7(3). [Google Scholar]
- 61.Crossman CA, Taylor EB, Barrett‐Lennard LG. Hybridization in the Cetacea: widespread occurrence and associated morphological, behavioral, and ecological factors. Ecol Evol. 2016. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 62.Fraser FC. Three anomalous dolphins from Blacksod Bay, Ireland. Proc. Roy. Irish Acad. 1940; 45 (Sect. B): 413–462. [Google Scholar]
- 63.Antrim JE, Cornell LH. Globicephala–Tursiops hybrid In Fourth Biennial Conference on the Biology of Marine Mammals, San Francisco, Calif; 1981, pp. 14–18. [Google Scholar]
- 64.Dohl TP, Norris KS, Kang I. Un híbrido de marsopa: Tursiops × Steno. Diario de Mammalogía. 1974; 55 (1): 217–221. [PubMed] [Google Scholar]
- 65.Nishiwaki M, Tobayama T. Morphological study on the hybrid between Tursiops and Pseudorca. Sci. Rep. Whales Res. Inst. Tokyo. 1982; 34:109–121. [Google Scholar]
- 66.Miyazaki N, Hirosaki Y, Kinuta T, Omura H. Osteological study of a hybrid between Tursiops truncatus and Grampus griseus. Bull. Natl. Sci. Mus. Ser. A (Zool.). 1992; 18: 79–94. [Google Scholar]
- 67.Sezaki K, Hirosaki Y, Wataba S, Hashinomoto K. Eletrophoretic characters of the hybrids between two dolphin Tursiops truncatus and Grampus griseus. Bulletin of Japanese Society of Science Fishered. 1984; 50(10):1771–1776. 10.2331/suisan.50.1771. [DOI] [Google Scholar]
- 68.Sylvestre JP, Tasaka S. On the intergeneric hybrids in cetaceans. Aquat Mamm. 1985; 11 (3): 101–108 [Google Scholar]
- 69.Shimura E, Numachi K, Sezaki K, Hirosaki Y, Watabe S, Hashimoto K. Biochemical evidence of hybrid formation between the two species of dolphin Tursiops truncatus and Grampus griseus. Bull.Jpn.Soc.Sci.Fish. 1986; 52:725–730. [Google Scholar]
- 70.Caballero S, Baker CS. Captive‐born intergeneric hybrid of a Guiana and bottlenose dolphin: Sotalia guianensis× Tursiops truncatus. Zoo Biology. 2010; 29(5): 647–657. 10.1002/zoo.20299 [DOI] [PubMed] [Google Scholar]
- 71.Perrin WF, Mitchell ED, Mead JG, Caldwell DK, Caldwell MC, Van Bree PJH, Dawbin WH. Revision of the spotted dolphins, Stenella spp. Mar. Mammal Sci. 1987; 3(2): 99–170. [Google Scholar]
- 72.Amaral AR, Sequeira M, Martínez-Cedeira J, Coelho MM. New insights on population genetic structure of Delphinus delphis from the northeast Atlantic and phylogenetic relationships within the genus inferred from two mitochondrial markers. Mar. Biol. 2007; 151(5): 1967–1976. [Google Scholar]
- 73.Amaral AR, Jackson JA, Möller LM, Beheregaray LB, Coelho MM. Species tree of a recent radiation: the subfamily Delphininae (Cetacea, Mammalia). Mol. Phylogenet. Evol. 2012; 64(1): 243–253. 10.1016/j.ympev.2012.04.004 [DOI] [PubMed] [Google Scholar]
- 74.Au DW, Perryman WL, Perrin WF. Dolphin distribution and the relationship to environmental features in the eastern tropical Pacific. SWFC. 1979. [Google Scholar]
- 75.Au DW, Perryman WL. Dolphin habitats in the eastern tropical Pacific. Fish Bull. 1985; 83(4): 623–644. [Google Scholar]
- 76.Polacheck T. Relative abundance, distribution and inter‐specific relationship of cetacean schools in the eastern tropical Pacific. Mar. Mammal Sci. 1987; 3(1), 54–77. [Google Scholar]
- 77.Forcada J, Aguilar A, Hammond PS, Pastor X, Aguilar R. Distribution and numbers of striped dolphins in the western Mediterranean Sea after the 1990 epizootic outbreak. Mar. Mammal Sci. 1994; 10(2): 137–150. [Google Scholar]
- 78.Das K, Lepoint G, Loizeau V, Debacker V, Dauby P, Bouquegneau JM. Tuna and dolphin associations in the North-East Atlantic: evidence of different ecological niches from stable isotope and heavy metal measurements. Mar Pollut Bull. 2000; 40(2): 102–109. [Google Scholar]
- 79.Garcia S, Knouse D, Sagarminaga R, Cañadas A. An insight on the biological significance of mixed groups of common dolphins (Delphinus delphis) and striped dolphins (Stenella coeruleoalba) in the Alboran Sea. European Research on Cetaceans. 2000; 14: 135–137. [Google Scholar]
- 80.Frantzis A, Herzing DL. Mixed-species associations of striped dolphins (Stenella coeruleoalba), short-beaked common dolphins (Delphinus delphis), and Risso's dolphins (Grampus griseus) in the Gulf of Corinth (Greece, Mediterranean Sea). Aquat Mamm. 2002; 28(2): 188–197. [Google Scholar]
- 81.Bearzi M. Dolphin sympatric ecology. Mar. Biol. Res. 2005; 1(3): 165–175. [Google Scholar]
- 82.Bruno S, Politi E, Bearzi G. Social organisation of a common dolphin community in the eastern Ionian Sea: evidence of a fluid fission-fusion society. European Research on Cetaceans. 2004; 15: 49–51. [Google Scholar]
- 83.Connor RC, Wells RS, Mann J, Read AJ. The bottlenose dolphin. Cetacean societies. 2000; 91–125. [Google Scholar]
- 84.Politi E, Airoldi S, Natoli A, Frantzis A. Unexpected prevalence of common dolphins over sympatric bottlenose dolphins in eastern Ionian Sea inshore waters. European Research on Cetaceans. 1998; 12: 120. [Google Scholar]
- 85.Ferretti S, Bearzi G, Politi E. Comparing behavior of inshore bottlenose and common dolphins in the eastern Ionian Sea through focal group surfacing pattern analysis. European Research on Cetaceans. 1998; 12: 209. [Google Scholar]
- 86.Bearzi G. A ‘remnant’common dolphin observed in association with bottlenose dolphins in the Kvarneric (northern Adriatic Sea). European Research on Cetaceans. 1997; 10: 204. [Google Scholar]
- 87.Weinrich MT, Lambertsen RH, Baker CS, Schilling MR, Belt CR. Behavioural response of humpback whales (Megaptera novaeangliae) in the southern gulf of Maine to biopsy sampling. In: Hoelzel AR editor. Genetic ecology of whales and dolphins: Incorporating the proceedings of the workshop on the genetic analysis of cetacean populations. Report of the International Whaling Commission, Special Issue 13 1991. pp. 91–97. [Google Scholar]
- 88.Noren DP, Mocklin JA. Review of cetacean biopsy techniques: Factors contributing to successful sample collection and physiological and behavioural impacts. Mar. Mamm. Sci. 2012; 28(1):154–199. [Google Scholar]
- 89.Bearzi G. First report of a common dolphin (Delphinus delphis) death following penetration of a biopsy dart. J. Cetacean Res. Manag. 2000; 2(3): 217–221. [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.
Data Availability Statement
All relevant data are within the paper.