

The Global Deal for Nature sets an ambitious agenda to protect our biosphere through ecosystem conservation and land restoration.
Abstract
The Global Deal for Nature (GDN) is a time-bound, science-driven plan to save the diversity and abundance of life on Earth. Pairing the GDN and the Paris Climate Agreement would avoid catastrophic climate change, conserve species, and secure essential ecosystem services. New findings give urgency to this union: Less than half of the terrestrial realm is intact, yet conserving all native ecosystems—coupled with energy transition measures—will be required to remain below a 1.5°C rise in average global temperature. The GDN targets 30% of Earth to be formally protected and an additional 20% designated as climate stabilization areas, by 2030, to stay below 1.5°C. We highlight the 67% of terrestrial ecoregions that can meet 30% protection, thereby reducing extinction threats and carbon emissions from natural reservoirs. Freshwater and marine targets included here extend the GDN to all realms and provide a pathway to ensuring a more livable biosphere.
INTRODUCTION
Nature conservation efforts, like climate change policies, are being reassessed in the midst of a planetary emergency (1). Climate concerns rightly prompted the 2015 Paris Agreement, which has facilitated coordinated global action not only among governments but also among companies, cities, and citizens. Research since then suggests that efforts to stabilize the climate and avoid the undesirable outcomes of >1.5°C warming will require a rapid reduction in land conversion and a moratorium by about 2035 (2). The most logical path to avoid the approaching crisis is maintaining and restoring at least 50% of the Earth’s land area as intact natural ecosystems, in combination with energy transition measures (2, 3). Those measures by themselves will likely be insufficient and must be augmented by restoration to create negative emissions to offset the likely clearing and release of greenhouse gases that will occur until a 2035 moratorium can be reached.
Natural ecosystems are key to maintaining human prosperity in a warming world (4, 5), and 65% of Paris Agreement signatories have committed to restoring or conserving ecosystems (6). Intact forests, and especially tropical forests, sequester twice as much carbon as planted monocultures (7, 8). These findings make forest conservation a critical approach to combat global warming. Because about two-thirds of all species on Earth are found in natural forests, maintaining intact forest is vital to prevent mass extinction (9). However, carbon sequestration and storage extends far beyond rainforests: Peatlands, tundra, mangroves, and ancient grasslands are also important carbon storehouses and conserve distinct assemblages of plants and animals. Further, the importance of intact habitats extends to the freshwater and marine realms, with studies pointing to least disturbed wetlands and coastal habitats being superior in their ability to store carbon when compared with more disturbed sites (10, 11).
Opportunities to address both climate change and the extinction crisis are time bound. Climate models show that we are approaching a tipping point: If current trends in habitat conversion and emissions do not peak by 2030, then it will become impossible to remain below 1.5°C (2, 12, 13). Similarly, if current land conversion rates, poaching of large animals, and other threats are not markedly slowed or halted in the next 10 years, “points of no return” will be reached for multiple ecosystems and species (13). It has become clear that beyond 1.5°C, the biology of the planet becomes gravely threatened because ecosystems literally begin to unravel (12, 14). Degradation of the natural environment also diminishes quality of life, threatens public health, and triggers human displacement because of lost access to clean drinking water, reduced irrigation of important subsistence crops, and exacerbation of climate-related storm and drought events (15). These occurrences will become increasingly worse without substantial action over the next few years. Additionally, human migrations, triggered by climate change–induced droughts and sea-level rise in combination with extreme weather events, could displace more than 100 million people by 2050, mostly in the southern hemisphere (12, 13).
A companion pact to the Paris Agreement—a Global Deal for Nature (GDN)—could help ensure that climate targets are met while preventing species extinctions and the rapid erosion of biodiversity and ecosystem services in the terrestrial, freshwater, and marine realms. The concept of a GDN as a policy mechanism emerged from an earlier study restricted to protecting biodiversity in the terrestrial realm (16). We expand that perspective to the freshwater and marine realms while simultaneously lending support to an alternative pathway to remaining below 1.5°C that relies heavily on aggressive conservation of remaining habitats. This approach not only safeguards biodiversity but also is the cheapest and fastest alternative for addressing climate change and is not beholden to developing carbon removal technologies unlikely to be effective or to scale in the time-bound nature of the current twin crises (4). Here, we offer a policy framework based on scientific guidelines that could pair nature and climate deals, be mutually reinforcing, and recommend time-bound milestones and targets. We identify specific threats and drivers of biodiversity loss, and discuss costs of implementation of a GDN. Finally, we introduce breakthrough technologies for monitoring progress.
SCIENTIFIC GUIDANCE FOR A GDN
Atmospheric sciences, Earth sciences, and remote sensing provide the main scientific and technological basis supporting the Paris Climate Agreement. The science of conservation biology underpins the GDN and is based on five fundamental goals: (1) represent all native ecosystem types and successional stages across their natural range of variation—or “representation”; (2) maintain viable populations of all native species in natural patterns of abundance and distribution—or “saving species”; (3) maintain ecological function and ecosystem services; (4) maximize carbon sequestration by natural ecosystems; and (5) address environmental change to maintain evolutionary processes and adapt to the impacts of climate change (17).
A practical application of many previous researchers has been to determine the spatial dimensions required to support goals 1, 2, 3, and 5 within large biogeographic units such as ecoregions—ecosystems of regional extent, terrestrial, freshwater, or marine—or regions holding concentrations of endemic species (biodiversity hotspots), the latter of which are typically delineated as clusters of ecoregions (16, 18). Carbon sequestration is a more recent concern.
Protected areas are the cornerstone of biodiversity conservation, and studies document that well-managed reserves are far more effective in safeguarding biodiversity than are other forms of land use (19). Studies across ecoregions and other large units show that to achieve the five goals of conservation biology requires in the range of 25 to 75% of land or water under some form of conservation management (20–22). Several efforts have identified a middle ground of 50% protected to ensure conservation of biodiversity and ecosystem services upon which humanity depends (16, 20, 23, 24). Exactly how much should be protected to maintain the diversity of life and secure the benefits nature provides is an empirical question that is best determined per biogeographic unit—from the ground up—to better inform global numerical targets. This finer spatial scale delineation adds efficiency by accounting for ecoregion-level patterns of endemism, beta-diversity, and connectivity requirements to maintain viable populations of area-sensitive species (16).
To ensure representation of native ecosystem types, goal 1, terrestrial ecoregions have been a widely used ecological classification system for conservation planning for nearly three decades (25). The rationale is that a global map of ecoregions can serve as the framework to ensure creation of networks of protected areas that represent the widest array of habitats and, by extension, conserve the widest range of species and their unique adaptations to their environments. A recent global review tested the distribution of more than 200 million species records of plants, animals, and fungi against the map of terrestrial ecoregions (Fig. 1A) and revealed sharp, statistically significant discontinuities in species ranges across boundaries (25). Thus, ecoregions effectively represent similar clusters of not only habitat types but also species—underpinning analyses to address goals 1 and 2 of the GDN (25).
Fig. 1. The world’s 846 terrestrial ecoregions and depiction of 30% protection by the 2030 milestone.
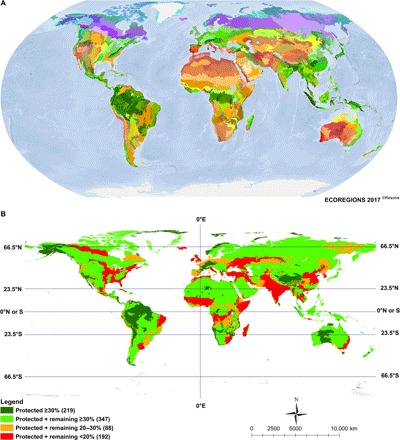
(A) The 846 terrestrial ecoregions. (B) Levels of protection by 2030.
To ensure that global protection efforts reinforce conserving viable populations (goal 2), many studies over the past two decades provide digital distributional data on well-known vertebrate taxa, some plants, a few invertebrate groups, and their levels of endangerment [e.g. (26)]. Many of these studies mapped distributions of taxa with limited ranges (endemics) or highly threatened species. Studies vary in spatial scale from identifying species limited to a single site, to endemics confined to narrow ranges, or to larger aggregations of rare or threatened species, and have been amalgamated within frameworks such as Key Biodiversity Areas (KBAs) or larger Biodiversity Hotspots (18, 27). At the other extreme are wide-ranging species that occur at low densities and often require extensive areas complemented with use management to maintain viable populations. Wide-ranging or area-sensitive large mammalian herbivores and carnivores require stepping stone or contiguous habitat between reserves to allow migrations, seasonal movements, and gene flow (28). Connectivity among reserves also becomes vital for ensuring species persistence in a changing world and for fostering ecosystem resilience (goals 3 and 5) (29).
The principles described above apply to the ocean as well. Marine ecoregions, defined for both coastal and pelagic provinces, delimit regions with distinct species assemblages often characterized by regional endemics (30, 31). Marine protected areas (MPAs), and in particular fully protected marine reserves, have proven much more effective than other actions (e.g., fisheries management) in protecting and restoring biodiversity, increasing yields in adjacent fisheries, and enhancing ecosystem resilience (32). In addition, management of fisheries through either marine reserves or effort control produces identical yields under a reasonable set of assumptions (33), providing support for half of the ocean closed to fishing. On average, species richness is 21% higher and biomass of fish is six times greater within marine reserves than in adjacent unprotected areas (24). Marine reserves help restore the complexity of ecosystems through a chain of ecological effects (trophic cascades) once the abundance of large animals recovers sufficiently (34). Although marine reserves work best for species with limited movement ranges, large reserves can also protect large predators that conduct transoceanic migrations not only if reserves capture reproduction and nursery habitats but also through genetic selection (35). Marine reserves may not be immune to the effects of climate change, but reserves with complex ecosystems are more resilient than unprotected areas (36). Studies suggest that, on average, more than 30% of the ocean should be protected to achieve a series of environmental and socioeconomic objectives (22).
The current global protected area targets agreed by the Convention on Biological Diversity under what is called “Aichi Target 11” set coverage targets for the year 2020—17% in the terrestrial realm and 10% in the marine realm. These are interim measures that are politically driven but not science based and are widely viewed in the scientific literature as inadequate to avoid extinctions or halt the erosion of biodiversity (20). Compounding the problem is that only about half of the 14.9% of the terrestrial realm currently covered by protected areas is also connected (37). Significantly increasing the percent of connected protected area networks under a GDN will thus be essential to achieve representation while also ensuring that viable populations are conserved and ecological and evolutionary processes are maintained (37).
A growing body of research documenting the inherent interconnection between carbon sequestration and biodiversity lends further support for a proposal to pair a GDN with the Paris Accord. Carbon-rich ecosystems, by definition, sequester (both store and pump) the most carbon from the atmosphere. This carbon sequestration service is key to climate stabilization and to climate change mitigation. It is no coincidence that some of the most carbon-rich ecosystems on land—natural forests—also harbor high levels of biodiversity. Evolution has generated carbon-rich forests by packing in long-lived trees that also feed stable soil carbon storage pools. This packing effect is made possible by high levels of coexistence among diverse species and growth forms, and this coexistence has been made possible by the biotic interactions that generate competition and defense. It is the very pests, pathogens, pollinators, decomposers, and predators that comprise a tropical forest that generated the carbon-rich growth forms (in both wood and soil) that take the carbon out of the atmosphere.
Even in the world’s widespread savannas, carbon sequestration is enhanced by biodiversity. Herbivores are key to plant growth as well as soil carbon sequestration. Predators are key to maintaining herbivore balance with primary production. Too many herbivores result in lower carbon storage, but too few predators result in overinvestment in plant biomass, which leads to off-scale fires and losses to the atmosphere. In the ocean realm, from coral reef to blue water, biodiversity is part and parcel to the flux of atmospheric carbon to stored carbonates and deep ocean sediments. Without biodiversity, this system shuts down as well. Studies across multiple realms demonstrate a loss of carbon potential as the biodiverse systems are degraded or destroyed (5, 10, 11).
The nexus of climate and biodiversity science offers chilling scenarios of the unraveling of biotic systems if temperatures exceed a 1.5°C rise in average global temperature (14). Climate models and vegetation models often underestimate the sensitivity of ecosystems due to idiosyncratic relationships between species. Clear examples include (i) coral polyps rejecting their algal symbionts causing bleaching events, (ii) native bark beetles producing massive tree mortality in coniferous forests, and (iii) debilitating infestation of ticks affecting reindeer, moose, and other cold-climate large ungulates. Historically, we know that individual species were able to move during past climate swings such that ecosystems disassemble and surviving species assemble into novel configurations. However, in the current climate crisis and with reduced connectivity of natural landscapes, species may be unable to move fast enough to track shifting climatic envelopes or at all (38). Tropical rain forests will likely revert to savanna-like vegetation in some species-rich regions, and tropical cloud forests, home to a disproportionate number of endemic species, will be severely compromised by reduction in cloud-borne moisture. These strong feedback loops render the planet impossible to manage biologically. The time-bound targets of the GDN will have the greatest short-term effect on saving species and habitats deemed most sensitive to rapid climate change (39).
PRIORITIES OF A GDN
Our objective is to present scientific guidance for three major themes that should be included in a GDN and a short list of key milestones and targets that could underpin these themes, which would be complementary and, in many cases, reinforcing of the Paris Climate Agreement. These themes are (1) protecting biodiversity, (2) mitigating climate change, and (3) reducing threats to ecosystem intactness and persistence of species. We also propose that the GDN embrace monitoring progress from the ground, or below the sea surface, to space, using powerful new technologies, much of it publicly available.
Theme 1: Protecting biodiversity
We support calls to conserve at least 30% of the Earth’s surface by 2030 (40). This is viewed as a milestone toward the larger end goal of half of the planet protected by 2050, if not sooner, made elsewhere (16). The 30% by 2030 milestone has also been proposed by the International Union for the Conservation of Nature (IUCN) and its member organizations as a critical step for marine conservation (IUCN Resolution: WCC-2016-Res-050-EN).
These global milestones and targets are useful: They are easy to comprehend and help simplify policy and communications strategies. But because biodiversity is unevenly distributed, conservation biologists and planners must be careful to avoid two major risks inherent in a single global percentage: (i) adding more land to reach the global target that is similar to what is already well accounted for at the expense of underrepresented habitats and species, and (ii) the temptation by some governments to protect low-conflict areas that may be lower priority from a biodiversity perspective (41). Adhering to the first goal of biodiversity conservation—representation—greatly reduces these risks. This is the rationale for setting milestones and targets at both levels—global and ecoregional—and shows how the latter helps to operationalize the former to better conserve life on Earth (Table 1).
Table 1. Increased representation by biome of ecoregions achieving 30% protection by 2030.
| GDN category |
Through increased protection only |
Through increased protection and restoration |
|||||||
| Biome name |
No. of ecoregions |
(1) Milestone Reached |
(2) High Potential |
(3) Moderate Potential |
(4) Nature Imperiled |
Ecoregions that can reach 30 × 30 (#) |
Ecoregions that can reach 30 × 30 (%) |
Ecoregions that can reach 30 × 30 (#) |
Ecoregions that can reach 30 × 30 (%) |
| Boreal Forests/Taiga | 26 | 3 | 18 | 2 | 3 | 21 | 81 | 23 | 88 |
| Deserts and Xeric Shrublands |
101 | 17 | 51 | 6 | 27 | 68 | 67 | 74 | 73 |
| Flooded Grasslands and Savannas |
25 | 14 | 3 | 4 | 4 | 17 | 68 | 21 | 84 |
| Mangroves | 20 | 9 | 6 | 1 | 4 | 15 | 75 | 16 | 80 |
| Mediterranean Forests, Woodlands, and Scrub |
40 | 10 | 12 | 10 | 8 | 22 | 55 | 32 | 80 |
| Montane Grasslands and Shrublands |
46 | 22 | 11 | 2 | 11 | 33 | 72 | 35 | 76 |
| Temperate Broadleaf and Mixed Forests |
83 | 16 | 32 | 11 | 24 | 48 | 58 | 59 | 71 |
| Temperate Conifer Forests |
47 | 9 | 21 | 10 | 7 | 30 | 64 | 40 | 85 |
| Temperate Grasslands, Savannas, and Shrublands |
48 | 0 | 20 | 5 | 23 | 20 | 42 | 25 | 52 |
| Tropical and Subtropical Coniferous Forests |
15 | 3 | 11 | 0 | 1 | 14 | 93 | 14 | 93 |
| Tropical and Subtropical Dry Broadleaf Forests |
56 | 6 | 15 | 11 | 24 | 21 | 38 | 32 | 57 |
| Tropical and Subtropical Grasslands, Savannas, and Shrublands |
58 | 13 | 19 | 8 | 18 | 32 | 55 | 40 | 69 |
| Tropical and Subtropical Moist Broadleaf Forests |
230 | 67 | 108 | 18 | 37 | 175 | 76 | 193 | 84 |
| Tundra | 51 | 30 | 20 | 0 | 1 | 50 | 98 | 50 | 98 |
| TOTAL | 846 | 219 | 347 | 88 | 192 | 566 | 67 | 654 | 77 |
To illustrate how biodiversity might be protected under theme 1, we conducted two related assessments: (i) how using the 846 ecoregions as a “conservation template” can achieve increased representation of critical habitats and species in networks of well-connected protected areas, and (ii) how that 30% fraction could be distributed to better protect areas of concentrated terrestrial biodiversity—from narrow-range endemic species in fragmented landscapes to endangered large vertebrate populations ranging across intact primary forests and grasslands. The remaining 20% of intact and semi-intact land is also essential for climate targets to avoid negative impacts on humanity and species populations (15). In addition to expansion of protected areas, we propose climate stabilization areas (CSAs) that would meet the criteria for Other Effective Area-based Conservation Measures (OECMs), recently defined by the Parties to the Convention on Biological Diversity (42). Working in cohort with the Paris Agreement, CSAs would concentrate in habitats like mangroves, tundra, other peatlands, ancient grasslands, and boreal and tropical rainforest biomes that store vast reserves of carbon and other greenhouse gases, and prevent large-scale land cover change. These complementary approaches are detailed below.
To assess how reserve expansion to meet the 30% by 2030 milestone could improve representation, we sorted the world’s 846 terrestrial ecoregions into four categories (Fig. 1B). These categories were created by the intersection of the most recent database of protected areas (43), showing a total of 14.9% of the terrestrial realm with the amount of remaining habitat—totaling 49.9% of natural habitat across the 14 terrestrial biomes—and partitioned by ecoregion. This total sums all intact and semi-intact habitats but excludes permanent ice in Greenland and the highly degraded Sahara ecoregions (table S1). The categories are as follows:
1) Milestone Achieved: ≥30% protected (26% of 846 ecoregions; n = 219);
2) High Potential: less than 30% currently protected, but enough habitat remains outside protected areas to reach 30% by 2030 (41% of 846 ecoregions; n = 347);
3) Moderate Potential: the sum of area protected and habitat remaining outside protected areas is between 20 and 30%. These ecoregions could meet the 2030 target with some restoration (10% of 846 ecoregions; n = 88); and
4) Nature Imperiled: <20% total habitat remaining (protected + unprotected), often much less (23% of 846 ecoregions; n = 192).
Achieving ecoregion milestones of 30% protected by 2030 would greatly improve representation in the global targets (Tables 1 and 2). Of the 347 High Potential (category 2) ecoregions, a median of 47% habitat remains outside protected areas. This provides the possibility for changes in land designation to increase the total number of ecoregions achieving 30% protection from 219 (26% of total ecoregions) to 566 ecoregions (67% of the total). A further 88 ecoregions in category 3 (10% of total) can achieve the 30% by 2030 milestone with some restoration. The 566 ecoregions (categories 1 and 2) that have reached or could reach 30% protected by 2030 are distributed among all of the 14 terrestrial biomes. The most complete representation would fall within the Tundra, Boreal, and Tropical and Subtropical Coniferous biomes (Table 1). The least representation would be in the Tropical and Subtropical Dry Broadleaf Forests and the Temperate Grasslands, Savannas, and Shrublands biomes (Table 1), the latter of which is the center of global food production.
Table 2. Combined milestones and measurable targets for a GDN to better protect biodiversity and biosphere function in the terrestrial, freshwater, and marine realms.
| Feature | 2018 Benchmark | Milestone for 2030 | Target outcome for 2050 | References |
| Protecting natural habitats and species | ||||
| Global percent natural habitat protected | ||||
| Global surface protected area coverage for Terrestrial, Freshwater, and Marine Realms |
Under Aichi Biodiversity Target 11 currently: (i) 14.9% of the world’s terrestrial and inland waters, and (ii) Approximately 4% of the global ocean is covered by implemented MPAs but only 2% in fully protected areas |
30% in protected areas: (i) 30% of terrestrial surface (incorporates freshwater), strategically located to better protect biodiversity and biosphere function (ii) at least 30% of each ocean habitat in fully to highly protected MPAs (iii) an additional 20% surface area designated as CSAs |
50% in protected areas composed of: (i) 50% of terrestrial surface (incorporates freshwater), strategically located to better protect biodiversity and biosphere function (ii) networks of fully to highly protected MPAs cover at least 30% of exclusive economic zones and 80% of the high seas |
(24, 40, 43) |
| Biodiversity representation by ecoregions (terrestrial and freshwater) | ||||
| Ecoregion-based representation in global protected area system |
(i) Less than half of the world’s 846 terrestrial ecoregions have at least 17% of their area in protected areas (ii) Only one-third of the 232 marine ecoregions (coastal) have at least 10% of their area protected |
(i) 300 terrestrial ecoregions have reached half protected; 563 terrestrial ecoregions have reached 30% protected (ii) All marine ecoregions have reached at least 30% protected |
(i) 650 terrestrial ecoregions half protected (ii) All marine ecoregions at least half protected |
(16) |
| Priority natural sites and species within ecoregions | ||||
| Alliance for Zero Extinction sites |
56% of 600 AZE sites protected | 100% of 600 sites are effectively conserved including 1-km buffer |
100% of target species have IUCN status improved |
(46) |
| IUCN range rarity of vertebrate species |
Rasterized map of hotspots | 50% of areas identified on map of hotspots of range rarity protected |
100% of areas with hotspots of threatened and small-ranged species protected |
(92) |
| Key Biodiversity Areas | 15,000+ KBAs identified as of 2018; 60% protected in 2018 |
90% of extant and future, formally identified KBAs are protected, including a 1-km buffer around all KBAs |
100% of extant and future, formally identified KBAs are protected, including a 1-km buffer around all KBAs |
(27) |
| High Biodiversity Importance Ecoregions |
455 HBIEs | 11.4% new protected areas added to reach 30% globally |
Addition of all other megafaunal areas overlapping with carbon sinks as CSAs |
Fig. S1H |
| Specific management actions targeting wide-ranging megafauna and large mammal migration routes |
As examples, range collapse and steep declines in populations of African elephants, most rhinoceros species, and many tiger populations. Rampant poaching in many regions; historic migration routes of large mammals under threat from development |
(i) Populations of 10 target species are doubled from 2018 baseline by 2030 (ii) Sport and commercial hunting of endangered megafauna and all trade in live animals and parts are banned (iii) 10 migration hotspots are secured and routes protected as globally recognized corridors |
(i) Populations of 20 target species are doubled (ii) Restoration of relatively intact megafaunal assemblages in 40 priority landscapes (iii) 20 migration hotspots are secured and protected as globally recognized corridors (iv) Achieving the above three targets leads to a delisting of these megafaunal species by the IUCN Red List |
(28, 49, 71) |
| Primary habitats | Combined, old-growth or intact habitats across all biomes cover less than 23% of the Earth’s surface; for some biomes, few large examples remain |
80% of 2018 extant is placed in protected areas or OECMs |
100% of old-growth habitats under protected areas or OECMs |
(15) |
| Stabilizing and restoring ecosystem function | ||||
| CSAs as OECMs with the explicit goals of conserving the carbon storehouses and global forest cover |
(i) Potential CSAs are currently intact and (ii) 2017 forest cover = 11.61 m km2 |
(i) Designated CSAs are 80% intact and 80% conserved through OECMs (ii) International and national protection for all mangrove, coastal marshes, wetlands, seagrass beds, swamp forest, peat forest, peatlands by 2030 and (iii) 80% natural forest cover remains intact globally |
(i) Designated CSAs remain intact as OECMs (ii) Increase in forest cover via Bonn Challenge and other means by 10% |
(3) |
| Indigenous lands | Indigenous peoples’ lands account for 37% of all remaining natural lands on Earth and store >293 gigatons of carbon |
High-priority indigenous lands that self-nominate and are identified as crucial to contributing to 2030 global targets are declared as OECMs with tenure and management financing secure |
All high-priority indigenous lands self-nominated as OECMs receive designation, tenure rights, and support for management effectiveness |
(74) |
| Maintain and restore connectivity of terrestrial protected areas |
7.5% terrestrial protected areas well connected |
20% terrestrial protected areas well connected |
40% protected areas well connected |
(37) |
| Maintain and restore connectivity of inland waters |
More than 800,000 dams and 45,000+ large dams exist; more than half of the world’s rivers blocked by large dams, thousands of smaller dams being planned, 35% of wetlands have been lost since 1970 |
(i) No further planning or building of large- to medium-sized dams on the world’s rivers; concentration of dams on tributaries with existing structures (ii) Maintain two-thirds of all headwaters of the Earth’s major river systems undammed by 2030 through protection and removal of blocking infrastructure (iii) Protect and restore riparian habitats along one-third of all rivers by 2030 (iv) Adequate protection and 1-km buffer zones for all RAMSAR wetlands by 2030 (v) Protection of one-third of the world’s forested upper watersheds by 2030 |
(i) Restoration of 25% of the world’s rivers to free-flowing state by 2050 through removal of dams and barrages (ii) Protection and restoration of riparian habitats along two-thirds of all rivers by 2050 (iii) Adequate protection and 1-km buffer zones for all globally mapped wetlands by 2050 (iv) Protection of one-half of the world’s forested upper watersheds by 2050 |
(57, 93) |
| Maintain and restore connectivity of marine waters |
Scant formal protection of critical marine habitats (reproduction, nurseries) for threatened species and migratory corridors for endangered species of fish, marine mammals, and sea turtles |
(i) Full protection of all critical habitats (reproduction, feeding, and nurseries) for threatened species (ii) Full protection of critical migratory corridors within local networks of MPAs for endangered species of fish, marine mammals, and sea turtles |
(i) Full protection of all critical habitats (reproduction, feeding, and nurseries) for commercial and threatened species (ii) Full protection of critical migratory corridors within local networks of MPAs for commercial and endangered species |
|
One hundred and thirteen ecoregions (13%) have already exceeded 50% protection. Yet, these rarely include the largest ecoregions containing the vast carbon reservoirs. Thus, a critical caveat in the representation approach is that some ecoregions may need much more area under protection to sustain species and processes and avoid biospheric feedback from release of greenhouse gases after conversion (see below discussion on CSAs for Amazon, Congo Basin, Southeast Asia, boreal, and tundra). At the other end of the spectrum, species extinctions will likely be most swift and severe in the 192 Nature Imperiled ecoregions, category 4 (Fig. 1B and table S1). Thus, efforts to bring the Nature Imperiled ecoregions to a protection level of 10% emerges as a clear 2030 restoration and recovery milestone. These ecoregions constitute known centers of endemism but have only a median of 4% protected habitat and 1% remaining outside protected areas. An approach to create a “species safety net”—to ensure the representation of vanishing biota ranging from single-site endemics to intact large mammal assemblages requiring large landscapes—is presented below (and more in greater detail in Fig. 2A).
Fig. 2. Increasing representation of important terrestrial, freshwater, and marine biodiversity sites for global 2030 targets.
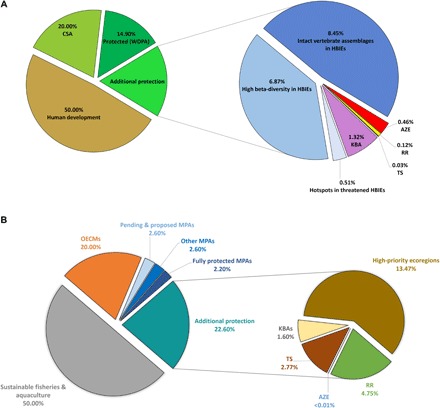
(A) Terrestrial and freshwater biodiversity sites. (B) Marine biodiversity sites. RR, IUCN Sites of Range Rarity; TS, Threatened Species Sites.
Range-restricted and area-sensitive species and conservation of beta-diversity
Saving species and populations is a key feature of a GDN, targeting first the rarest, most range-limited species under the most threat for immediate conservation (44). To ensure that under the 30% by 2030 milestone these additions by ecoregion include irreplaceable sites (e.g., those with narrow endemic taxa found nowhere else), we intersected several widely used fine-scale maps of biodiversity distributions with the ecoregion map and the current database on terrestrial protected areas (45). For each of these overlays, we calculated the area for that theme found in each ecoregion that is currently unprotected but not accounted for by other overlays (to avoid double counting, as the spatial extent of these various mapping efforts depicting species rarity often overlaps considerably). We summed these extensions to present the percent and amount (in km2) added to the global total toward the 30% by 2030 milestone (Fig. 2). These data layers that are included inform milestones and targets (Table 2):
1) The Alliance for Zero Extinction (AZE) sites that are currently unprotected (46). AZE sites pinpoint extreme rarity by seeking to protect those birds, mammals, reptiles, amphibians, cycads, and conifers whose entire global distribution is limited to a single site and ranked as Critically Endangered or Endangered. The total is 928, of which 34% are unprotected (Table 2). All unprotected AZE sites, plus a 1-km buffer surrounding them, are included in the 2030 milestone. These 320 unprotected sites, plus buffer, add 0.46% to the expansion of the global terrestrial protected areas, or 619,490 km2 (Fig. 2A and fig. S1A). In the marine realm, unprotected AZE sites represent 9955 km2 and would contribute less than 0.1% to the expansion of the global MPAs (Fig. 2B).
2) A map of range rarity derived from the IUCN Red List data for the world’s birds, mammals, and amphibians, showing where sites supporting high densities of narrow-range species remain unprotected. The unprotected hotspots of range rarity add 0.12% to the expansion of the global terrestrial protected areas total percent or 162,491 km2, not including AZE sites (Fig. 2A and fig. S1B). In the ocean, conservation priority hotspots (top 5%) for range-restricted species (47) represent 14.7 million km2 and can account for 4.75% of the expansion of the global MPAs (Fig. 2B and fig. S1C).
3) A map of the density of threatened species from the IUCN Red List, showing where concentrations of species threatened with extinction remain unprotected. This layer includes nearly 64,000 species evaluated to date by the IUCN Red List, with almost 20,000 deemed as threatened with extinction. The unprotected polygons could add 0.03% to the expansion of the global terrestrial protected areas total percent or 34,631 km2 excluding overlapping data from the first two layers (Fig. 2A and fig. S1D). Conservation priority hotspots (top 5%) for threatened marine species (47) represent 8.5 million km2 and account for 2.8% of the expansion of the global MPAs (Fig. 2B and fig. S1E).
4) KBAs map globally important sites that meet a number of biodiversity criteria, including the presence of threatened or significant species (27). Though incomplete for some regions of the world (e.g. New Guinea), KBAs are identified by national constituencies using globally standardized criteria and quantitative thresholds and thus are useful to guide the expansion of protected area networks. Currently, approximately 15,000 such sites have been designated, with about 60% within IUCN category I to IV reserves (27). Adding in the unprotected portion of KBAs and a 1-km buffer expands the global terrestrial protected areas total by 1.3% percent or 1,759,341 km2 (Fig. 2A and fig. S1D). Unprotected KBAs in the oceans represent 5.8 million km2 and can contribute 1.6% to the expansion of the global MPAs (Fig. 2B and fig. S1G).
Summing the total area of all of these unprotected sites (plus 1-km buffers) from the first four layers, and removing double counting, adds 1.9% to the existing 14.9% of the terrestrial realm and 9.2% to the existing 7.3% of the marine realm already designated for protection (45). The unexpectedly small area contribution of these irreplaceable terrestrial sites is due to the high spatial overlap present among layers 1 to 4.
5) The remaining contribution to the milestone of 30% by 2030 is drawn from High Biodiversity Importance Ecoregions (HBIEs) that depict 455 of the most biologically distinct ecoregions by biome (fig. S1H). It is a slightly revised and updated map of the most biodiverse ecoregions on Earth (48). This is subdivided into three themes encompassing small fragments or larger landscapes that remain unprotected and not covered in the above four layers:
i) The 36 Biodiversity Hotspots are included in the HBIE layer and contain a combined total of 114 category 3 and category 4 ecoregions, the most threatened categories. Adding the remaining unprotected habitat fragments found in these ecoregions contributes 0.51% or 439,569 km2 to the expansion of the global terrestrial protected areas total (Fig. 2A and fig. S1I).
ii) Biodiversity Hotspots (fig. S1F) represent educated guesses (or sometimes comprehensive lists) by botanists of regions where plant endemism is concentrated globally. These lists are often based on counts of endemics at particular sites—a depiction known as alpha-diversity. Equally important as a conservation metric, but rarely measured, is beta-diversity—the replacement or turnover of the rosters of plant communities with distance or along elevational gradients (fig. S1J). High beta-diversity ecoregions are most common in five biomes: Tropical and Subtropical Moist Broadleaf Forests; Tropical and Subtropical Dry Broadleaf Forests; Mediterranean Climate Shrublands; Tropical and Subtropical Grasslands, Savannas, and Woodlands; and the tropical and subtropical ecoregions found in Desert and Xeric Shrublands. Many of the ecoregions in these five biomes overlap with the biodiversity hotspots; adding in unprotected habitat in some missing ecoregions adds 6.9% to the global total or 9,188,027 km2 (Fig. 2A).
iii) The strongholds of large wide-ranging species that require vast areas to maintain viable populations are another important metric of biodiversity value. HBIEs capture where these large mammals occur by overlaying a map of intact vertebrate assemblages (fig. S1K) (49). Adding in unprotected intact large mammal habitat in some missing ecoregions adds 8.5% to the global total or 11,304,493 km2 (Fig. 2A and fig. S1H).
Conserving the unprotected terrestrial areas selected above would add 17.7% to the existing 14.9% currently protected and exceed the 30% milestone while creating a more representative global system of protected areas that incorporate the major aspects of terrestrial biodiversity (Fig. 2A). Our approach uses what we consider to be the most important biodiversity features, but we recognize that there are multiple pathways to achieve a 30% protected milestone.
Complete protection requires closer to 50% of the Earth’s land surface achieved by combining the above analysis with 20% designated as CSAs (see below section and Table 2).
Wide-ranging megafauna and migration routes
The future survival and recovery of megafauna will, in particular, require additional strategies beyond those above for prioritizing sites that require immediate attention. Many of our Earth’s large terrestrial vertebrate species are suffering range collapse and extirpation of populations and could disappear by the end of this century or sooner if intensive habitat conversion, poaching, and overhunting continue given the slow rates of population recovery characteristic of most megafaunal species (50). Similar sharp declines are being observed in freshwater and marine large vertebrate species that figure prominently as protein sources for billions of people (51). Large mammal migrations, one of the most outstanding natural phenomena on Earth, are disrupted globally as traditional migratory routes are cut off and habitat lost (28). Ultimately, the loss of large herbivores, ecosystem engineers, and top carnivores can produce detrimental effects across entire food webs, termed “trophic cascades” (52).
GDN targets can address maintaining megafaunal populations and landscapes in multiple ways not only through direct protection but also by promoting connectivity (Table 2). The current CBD Aichi Target 11 calls for “well-connected systems of protected areas,” although only a rudimentary definition of “well-connected” and inadequate guidance on how to measure connectivity is provided (37).
Corridors and connectivity
A key target of the GDN would be to reconnect protected areas via corridors along environmental gradients, in riparian networks, and between megafaunal reserves (Table 2). The amount of area required and how this would affect the protection or restoration target should be subject to a major study based on the needs of the most wide-ranging, area-sensitive species. Replanting of native trees or simply allowing degraded forest lands to recover as forest corridors could create an ecological road map guiding where restoration can have the maximum benefits for biodiversity.
Old-growth habitats
Before the industrial revolution, primary or old-growth habitats covered most of Earth’s 846 terrestrial ecoregions. Today, these primary habitats, as represented by unlogged forests, ungrazed deserts, ancient grasslands, and savannas—and in the marine realm, the untrawled seafloor and unfished seamounts—are now remnants. These ancient repositories of rich and vulnerable biodiversity are optimal arenas for life-sustaining processes (53). Clear, time-bound milestones and targets for the above biodiversity features, including targets for old-growth forests and ever-wet forests, are included in the GDN and are drawn from the scientific literature supporting each target (Table 2). Protecting habitats that have low anthropogenic disturbance offers the most cost-effective approach to conserve the largest number of species and also for their climate resilience and should become obvious targets under the GDN (54). As a prime example, tropical forests occur on only 7% of the land area, yet they harbor more than half of the world’s known species and most of these are dependent on primary forest (9, 55). The buffering conditions of many complex “old-growth” habitats also enable local adaptation to climate change for many vulnerable species. These biologically important areas also serve as carbon repositories. For example, ancient grasslands are extremely species rich, including endemics (53), and they store approximately as much carbon globally as forests. Because most grassland carbon is stored belowground, it is a highly secure and reliable carbon sink, especially in the face of fire and other climate-sensitive disturbance factors (56).
The freshwater realm
Freshwater habitats harbor roughly one-third of all vertebrate species and 10% of global biodiversity, yet only cover 1% of the planet (57). Freshwater ecoregions particularly rich in species, endemics, and intact megafauna are priorities for biodiversity conservation, as are those where watersheds are largely intact and waterways flow freely (58). Freshwater ecosystems and biodiversity are among the most threatened on Earth (57). Many of the protection measures identified for terrestrial areas cover required actions for freshwater, and targets for the former benefit the latter. The GDN targets protecting and restoring 30% of the world’s freshwater ecoregions by 2030 as a vital milestone (Table 2).
The marine realm
A meta-analysis of 144 studies showed that, on average, 37% of the ocean should be strongly protected (preferably in fully protected marine reserves) to achieve a number of goals: protect biodiversity, ensure connectivity, avoid species collapse and adverse evolution, ensure sustainable fisheries, and benefit multiple stakeholders (22). More than 80% of these objectives can be met with 50% marine reserve protection (22). With only 2% of the world’s ocean currently in fully to highly protected marine reserves (59), these estimates highlight enormous gaps in marine conservation.
In addition to biodiversity, marine ecosystems are also important carbon sinks. Coastal ecosystem such as mangroves, salt marshes, and seagrass beds sequester astonishing amounts of carbon per hectare and thus play a disproportionately large role in the global capture and storage of carbon (60, 61). Although the spatial extent of the world’s 20 mangrove ecoregion complexes is minute compared to other biomes (300,294 km2 or 0.23%), conservation and restoration of mangroves are vital to climate and marine conservation scenarios, and buffer storm surges and hurricanes.
The ocean also harbors old-growth ecosystems equivalent to primary forests, such as unfished seamounts with deep corals thousands of years old (62). In the coastal zone, some seagrass species such as the Mediterranean Posidonia oceanica have complex belowground organs that help accrete sediment and bury carbon over millennia (63). Preserving the vegetative cover of these coastal habitats is important, but preserving their megafauna is essential, as studies suggest that intact predator populations are critical to maintaining or growing reserves of carbon stored in marine ecosystems (64). For example, tiger sharks in seagrass beds in Australia create a “landscape of fear,” where sea turtles and dugongs preferentially forage in seagrass microhabitats that are low in predation risk. The majority of the habitat, with high predation risk, has greater carbon stocks.
Ongoing efforts are identifying priority areas that will help protect biodiversity at multiple levels (from species to ecosystems), help produce food through fish spillover from fully protected reserves, and help mitigate climate change through the protection and restoration of important carbon sinks. Here, we present general targets and categories that should be prioritized as part of the GDN (Fig. 2B). The current shortfalls in ocean protection are presented in a revised map of the coastal ecoregions and pelagic provinces of the world, alongside the global system of MPAs (Fig. 3, A and B) (30, 31).
Fig. 3. Coastal ecoregions, pelagic provinces, and marine protected areas of the world oceans.
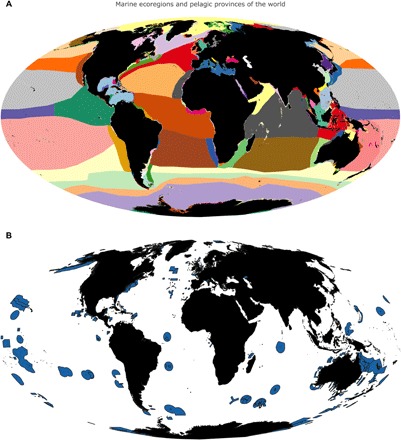
(A) Coastal ecoregions and pelagic provinces. (B) Map of marine protected areas.
Theme 2: Mitigating climate change
Conserving the carbon storehouses: CSAs
Protecting 30% of the Earth as high-priority conservation areas will be essential but insufficient for holding emissions below 1.5°C (2). Carbon storehouses, our terrestrial carbon sink, currently absorb one-quarter of emissions, and the conservation of natural habitats as stabilization areas under the GDN reinforces this role. Natural habitats outside protected areas can also be managed to maintain intactness and enhance conservation of area-sensitive species. These lands can be considered under the term OECMs (43). CSAs can fit under the OECM umbrella and contribute to area-based conservation targets. Simply put, CSAs are areas where conservation of vegetative cover occurs and greenhouse gas emissions are prevented, which can be achieved under various forms of land management. A key target of the GDN would enhance efforts to maintain at least 85% forest cover in critical areas such as the Amazon. Some parts of the planet work as a system and need to be managed for a high proportion of intactness to continue to function as a weather machine for the planet (65, 66).
These essential carbon storehouses could be retained and protected under CSAs. The tundra and boreal biomes, which together comprise 24,162,700 km2 (approximately 18%) of Earth’s terrestrial landmass, are exceptional because of their carbon storage [fig. S2A and (13)]. Examples of forests with high carbon density include the temperate rain forests of the Pacific Northwest United States and Canada, the temperate moist eucalypt forests in southeast Australia, Congo Basin peat swamp forests, and intact forest reserves in Malaysian Borneo [e.g., (67, 68)]. In recent years, more than 60% of global emissions from natural habitats (which accounted for 15% of annual greenhouse gas emissions), stemmed from clearing and fires in just two provinces—Riau province in Sumatra, Indonesia, and Matto Grosso state in Brazil. Protection for these and other “high-biomass forests” that are disproportionately important in climate mitigation could be enabled through their designation as CSAs.
Tundra and boreal biomes, aside from extensive belowground carbon storage, retain the largest areas of intact large mammal assemblages and are home to some of the most extensive and extant large mammal migrations on Earth (fig. S1K) (28, 49). Globally, the world’s intact large mammal assemblages coincide with the carbon storehouses and overlap substantially with the world’s remaining wilderness areas, excepting permanent ice in Greenland and in the heavily degraded, defaunated, and overgrazed ecoregions of the Sahara (fig. S2B) (15). However, preventing the transformation of large vegetated regions is not sufficient to maximize their carbon sequestration role. In some major ecosystems, the presence of large predators and herbivores helps store more carbon (69). For example, many large tropical trees with sizeable contributions to carbon stock rely on large vertebrates for seed dispersal and regeneration (70).
Conservation of tiger habitat provides another example of how CSAs could also help improve species protection and recovery efforts. Tigers are habitat generalists, but today, extant populations are largely restricted to forests, and many of these forested habitats lack formal protection under IUCN reserves. Approximately 434 protected areas fall with the tiger’s current range (fig. S3), not all of which contain breeding tigers, and not a single protected area is large enough to support a viable population over the long term (71). These adjacent reserves must be managed as metapopulations, populations linked by occasional dispersal, and so connectivity, and in many cases restoring forest corridors, is essential. The sum of all protected areas in the tiger range is 495,807 km2 or 42% of the total area of all 76 Tiger Conservation Landscapes combined (fig. S3). Between now and 2030, only a fraction of the remaining habitat outside formally protected areas will be incorporated into expanded or new reserves. Yet, enough habitat remains to achieve the target of a near-tripling the wild tiger population (72), and management of this habitat could be supported under the OECM umbrella as megafaunal landscapes in that part of the breeding tiger population, especially in India, that occurs outside formal protected areas. The payoff for climate stabilization is dramatic: An earlier study showed that forested areas that contain tigers have three times the carbon density compared to forests and degraded lands where tigers have been eradicated (71). Restoration in tiger habitat and other megafaunal landscapes could center on protecting remaining fragments of natural habitat, reconnecting and buffering them by restoring degraded lands, thus aligning with the Bonn Challenge that seeks to restore 350 million ha of forest by 2030 (73). The same rationale could be used to extend protection in high-carbon density habitats for gibbons and other primates, hornbills and other large fruit-eating birds, and fruit-eating bats.
The central role of indigenous lands
Potentially prominent among OECMs are indigenous peoples’ lands, which account for 37% of all remaining natural lands across the Earth, and these lands store >293 gigatons of carbon (74). Although many of these lands meet the definition of a protected area, many others may be appropriately characterized as OECMs. Here, the global policies articulated in the Paris Agreement and the proposed GDN merge with addressing human rights. The direction, insights, rights, and voices of indigenous peoples are essential but rarely published in scientific journals. The GDN could assist indigenous peoples, where requested, to keep lands intact—for hunting areas, protection of traditional lifestyles, or other features—and provide a mechanism to assist these communities with securing tenure rights. Supporting efforts to maintain these lands, many of which are critical to global terrestrial biodiversity conservation, in many cases would result in lower rates of deforestation and better protection of the biodiversity and ecosystem functions upon which these communities depend (75). Furthermore, more than one-third of the carbon identified in community lands across the tropics lies in areas without secure tenure rights. The Amazon Basin, Congo Basin, boreal, tundra, Borneo, and New Guinea ecoregions all store massive amounts of above- and belowground carbon (fig. S2A) and overlap greatly with indigenous lands.
Theme 3: Reducing major threats
Land conversion and infrastructure development risk compromising the ability of protected areas and CSAs to protect species and store carbon. Slowing and stopping the clearing of intact natural habitat for agriculture, the dominant form of land use today, is critical as part of the overall strategy to stay below 1.5°C. By increasing intensification and directing cropland expansion to degraded lands, and by reducing food waste, the 2050 world food demand could be met without additional land clearing (76, 77). The total length of paved roads globally is projected to increase by 20 million km (enough to encircle the Earth more than 600 times), and 90% of all new infrastructure is slated for the world’s tropical and subtropical biodiverse ecosystems (78). Infrastructure and energy development projects—major sources of fragmentation and penetration into wilderness areas, protected areas, indigenous territories, and CSAs—should be closely scrutinized (see Table 3 for recommended targets and policies) (79). Proactive approaches are needed to optimize human benefits while limiting harm.
Table 3. Enabling policies, milestones, and targets to reduce major threats and drivers of change.
| Enabling policies to reduce threats and drivers | ||||
| Feature | 2018 Benchmark | Milestone for 2030 | Target outcome for 2050 | References |
| Agricultural expansion |
Cropland covers at least 12% of the planet’s ice-free surface; the expected range of cropland expansion is 123–495 Mha per annum |
(i) Expansion of agro-commodity production and supporting roads and settlements is moved to degraded or previously converted areas such that range of cropland expansion into natural areas is halved from 2020 levels. (ii) Priority biodiversity and biospheric areas are experiencing no net loss of habitat due to agricultural expansion. (iii) Targets established and met for increase in per ha productivity |
No loss of natural habitat for commercial agro-commodity production and sourcing is occurring |
(94–96) |
| Roads | At least 25 million km of new roads projected by 2050 (a 60% increase in the total length of roads over that in 2010); 70% of the world’s forests are less than 1 km from a forest edge |
(i) Transnational transport corridor projects that will affect priority biodiversity and biosphere function target areas are subject to international oversight of strategic road planning that minimizes impacts on biodiversity and biosphere function targets. (ii) Top 50 planned road networks or improvements that would directly affect priority biodiversity and biosphere function habitats and regions are not eligible for international financing. (iii) International financing is predicated on ensuring overpasses and underpasses in engineering designs to ensure integration of social and ecological connectivity |
All transnational transport corridor projects that can affect priority biodiversity and biosphere function target areas are subject to international oversight of strategic road planning that minimizes impacts on biodiversity and biosphere function targets. All planned road networks or improvements that would directly affect priority biodiversity and biosphere function habitats and regions are not eligible for international financing |
(78, 79, 97) |
| Dams, barrages, channelizations |
More than 800,000 dams and 45,000+ large dams exist; more than half the world’s rivers blocked by large dams, thousands of smaller dams being planned |
(i) No further planning or building of large- to medium-sized dams on the world’s rivers (ii) Maintain two-thirds of all headwaters of the Earth’s major river systems undammed by 2030 through protection and removal of blocking infrastructure |
Restoration of 25% of the world’s rivers to free-flowing state by 2050 through removal of dams and barrages |
(57, 93, 98) |
| Overfishing | The global marine catch peaked in 1996 and has been declining since, with more than 30% of fisheries collapsed; more than 1000 species threatened with extinction due to fishing |
(i) Subsidies that contribute to overcapacity and overfishing are eliminated (ii) Global fishing capacity cut in half (iii) Regional Fisheries Management Organizations reformed and made accountable to a new independent global fisheries agency |
(i) End of overfishing (ii) All commercial fisheries management is science based and sustainable and is based on access rights (iii) Sustainable aquaculture based on non-fish feed has replaced half of the marine catch |
(24, 81, 99, 100) |
| Wildlife trade | Overexploitation affects three-fourths of threatened species; wildlife products are legally traded internationally at volumes of an average of 100 million whole organism equivalents per year over the past 10 years |
(i) Sport and commercial hunting of threatened terrestrial, marine, and freshwater animals and parts are banned nationally and internationally (ii) Agreements in place to prohibit international trade/sale/transport for commercial purposes of all wild-caught threatened species |
(i) Global ban in international transport for commercial purposes of all wildlife species and threatened plant taxa (ii) Global legislation and enforcement banning any trade in threatened species (iii) Legal trade volumes considered sustainable for all species |
(9, 100, 101) |
| Invasive species | ~17,000+ invasive species documented |
(i) Solidify gains in the Actions and Milestones of Aichi Target 9 invasive alien species prevented and controlled, namely, “By 2020, invasive alien species and pathways are identified and prioritized, priority species are controlled or eradicated, and measures are in place to manage pathways to prevent their introduction and establishment.” (ii) Control of top plant or animal invasive species in 100 global priority islands |
(i) Solidify gains in the Actions and Milestones of Aichi Target 9 Invasive alien species prevented and controlled, namely, “By 2020, invasive alien species and pathways are identified and prioritized, priority species are controlled or eradicated, and measures are in place to manage pathways to prevent their introduction and establishment.” (ii) Control of top plant or animal invasive species in 200 global priority islands |
(84, 100) |
| Plastics | The amount of plastic making its way into the oceans is predicted to increase from 9 million metric tons in 2015 to 16 million metric tons in 2025 |
Global ban on all nonrecyclable, single-use plastics; recycling of 30% of the world’s plastics To achieve the SDG target to “prevent and significantly reduce marine pollution” by 2025, the world needs to move from our current “linear economy” (make, use, dispose) to a circular economy in which resources do not become waste but instead are recovered and regenerated at the end of each service life. Government should embed the circular economy into national strategies |
Global ban on all single-use plastics; recycling of 50% of the world’s plastics |
(85, 86, 102) |
| Toxins | Current widespread use of ecologically damaging toxins occurs, causing massive declines in global pollinators, invertebrate biomass, and degradation of aquatic ecosystems |
The most ecologically damaging classes of commercial toxins (e.g., certain pesticides, herbicides, nematocides, and fungicides, especially those that kill pollinators, poison freshwaters, and sterilize soils) no longer produced, sold, or used globally |
Global program to monitor and enforce no production, sale, and use of most ecologically damaging toxins, including testing newly developed commercial toxins |
(103) |
| Ozone-depleting chemicals |
The Montreal Protocol on Substances that Deplete the Ozone Layer currently regulates ozone-depleting chemicals |
A global ban on production and use of ozone-depleting chemicals effectively enforced |
(104) | |
On land, hunting by humans imperils 40 to 50% of all threatened bird and mammal species (80). In the marine realm, industrial fishing is the largest hunting operation on the planet and targets more than half of the ocean surface, spanning an area four times that covered by terrestrial agriculture (81). Currently, fishing exploitation rates remain uncontrolled in vast ocean areas, including the high seas. Only a small fraction of the fisheries of the world are managed and science based, and they mostly concern single species targeted by industrial fleets in developed countries (82). In many cases, reducing fishing effort could help increase efficiency and profitability (83). Illegal and unsustainable trade in animals and plants, especially in threatened species, must also be curtailed. Further, resources will be needed to enforce protection as protected areas expand under a GDN. The most commonly invoked intervention to counteract poaching and overhunting is law enforcement patrolling to deter, detect, and punish poachers. Halting illegal and unsustainable trade in animals and plants, in particular of species threatened with extinction and where trade adds to the pressure on that species, is essential (Table 3).
The proliferation of invasive species, pollutants, and toxins is a major driver of species loss, population declines, and habitat degradation around the world (84). The amount of plastic making its way into the oceans is predicted to nearly double in the next decade; allowing this to occur would unleash extremely detrimental impacts on marine species and ecosystems (85). Beyond plastics, widespread use of ecologically damaging toxins is causing massive declines in global pollinators, invertebrate biomass, and degradation of aquatic ecosystems. To achieve the Sustainable Development Goals (SDG) target to prevent and significantly reduce pollution, the world needs to move from our current “linear economy” (make, use, dispose) to a circular economy in which resources do not become waste but instead are recovered and regenerated at the end of each service life (86). A GDN should encourage appropriate regulations, market incentives, and enterprise in areas such as waste management upstream to prevent plastic trash entering the ocean (Table 3). Funding for research, technology, and invasive species management programs in targeted areas can have marked effects in restoring native species populations and ecosystem services.
The conservation biology literature offers extensive analyses and detailed case studies of global threats and drivers of biodiversity loss. We have distilled key papers from this literature to identify clear milestones and targets that reflect their scope and intensity and how to reduce their impact as an integral part of the GDN (Table 3).
DISCUSSION
The Paris Agreement offers a useful template for a GDN because it sets global targets, provides a model for financial support, and supports bottom-up efforts. All nations have signed on to this agreement. But the Paris Agreement is only a half-deal; it will not alone save the diversity of life on Earth or conserve ecosystem services upon which humanity depends. It is also reliant on natural climate solutions that require bolstering outside of the Paris Agreement to ensure that these natural approaches can contribute to its success. Yet, land-based sequestration efforts receive only about 2.5% of climate mitigation dollars (4, 87).
At the same time that climate scientists were arriving at a single numerical target for maintaining Earth’s atmosphere at safe limits, biodiversity scientists identified multiple targets for the required habitats to conserve the rest of life on Earth. But to communicate effectively, as in the Paris Agreement, these many needs could be encompassed within a single target: protect at least half of Earth by 2050 and ensure that these areas are connected (16, 23, 40). The evidence arising since these calls were made clearly demonstrate that while we may be able to afford to wait to formally designate 50% protected in nature reserves, we need to fast-track the protection and restoration of all natural habitat by 2030 (2). A GDN that will ensure that we have at least 50% intact natural habitats by 2030 is the only path that will enable a climate-resilient future and is one that will offer a myriad of other benefits (3, 4). Since the crucial role of intact, diverse systems has also been demonstrated to be essential for carbon storage (8, 15), the GDN will need to emphasize mechanisms for protecting intactness both inside and outside of protected areas (e.g. in CSAs/OECMs) well before 2050.
Tallis and colleagues (3) demonstrated that with existing technologies and large-scale adoption of common conservation approaches (e.g., protected areas, renewable energy, sustainable fisheries management, and regenerative agriculture), it would be possible to advance a desired future of multiple economic and environmental objectives (including 50% of each biome intact, with the exception of temperate grasslands). This spatial coexistence is possible even with the prospects of feeding and supporting the material needs of a growing human population (88). The success of proposals to boost food production while protecting biodiversity will likely depend on our success in addressing human population growth, however, and our willingness to marshal financial resources accordingly (89).
Gross costs for nature conservation measures across half the Earth could be $100 billion per year, but the international community currently spends $4 billion to $10 billion per year on conservation (90). Extending the area-based targets in the post-2020 strategic plan for biodiversity to 30% by 2030 will likely require direct involvement of the private sector. In key sectors—fishing, forestry, agriculture, and insurance—corporations may be able to align their financial returns directly to reaching targets recommended by the GDN. However, the typical approach to conservation planning does not involve the real (net) costs because the direct benefits of conservation and the averted costs of inaction are not included in the calculations. Barbier and colleagues (90) showed that potential direct benefits from biodiversity conservation for various sectors range from increasing annual profits by $53 billion in the seafood industry to $4300 billion in the insurance industry. In addition, marine reserves can provide more economic benefits from tourism than fishing in many locations worldwide (91). Financial investments of even 10 to 20% of potential benefits from biodiversity conservation from three key industries could make up as much of one-third of the commitment needed to implement a GDN. A GDN may appeal to a broader set of nonstate actors, including corporations and local government entities. The solutions could be implemented in ways that have direct positive benefits to local or regional communities and especially indigenous peoples. Land-based jobs, food security, green space, access to wilderness, and ecosystem services are benefits that deliver advantages to rural and urban dwellers alike.
Complex life has existed on Earth for about 550 million years, and it is now threatened with the sixth mass extinction. If we fail to change course, it will take millions of years for Earth to recover an equivalent spectrum of biodiversity. Future generations of people will live in a biologically impoverished world. Adopting a GDN and the milestones and targets presented here would better allow humanity to develop a vibrant, low-impact economy and conserve intact ecosystems, all while leaving space for nature. Linking the GDN and the Paris Agreement could solve the two major challenges facing the biosphere and all the species within it and result in a return to safe operating space for humanity.
Supplementary Material
Acknowledgments
We thank B. Babbitt, M. Gomera, N. Lapham, S. Pimm, and J. Watson for reviewing drafts of the manuscript. Two anonymous reviewers helped improve the article. Author contributions: E.D., C.V., E.S., T.E.L., D.O., K.B., R.F.N., J.E.M.B., and Y.P.Z. designed the framework of this paper; A.R.J., S.F., J.M., and A.B. conducted data analysis and interpretation of data; E.D., C.V., E.S., D.O., G.P.A., N.D.B., R.F.N., S.F., K.B., T.B., N.H., L.N.J., and E.W. contributed substantially to the writing and editing of the manuscript. Competing interests: There are no financial or competing interests involved in authorship of this paper. Data and materials availability: All data needed to evaluate the conclusions in the paper are present in the paper, the Supplementary Materials, and/or the materials cited herein. Additional data related to this paper may be requested from the authors.
SUPPLEMENTARY MATERIALS
Supplementary material for this article is available at http://advances.sciencemag.org/cgi/content/full/5/4/eaaw2869/DC1
Section S1. Maps of important biodiversity and carbon layers
Section S2. Underlying data for increasing representation of ecoregions by adding unprotected areas of high priority
Section S3. Monitoring progress under the GDN from the ground to below the sea to space
Fig. S1. Maps used to increase representation among terrestrial ecoregions and unprotected sites of biodiversity importance that contribute to the global milestone of 30% protected by 2030.
Fig. S2. Maps showing total terrestrial carbon and overlap with intact large mammal assemblages.
Fig. S3. Overlay of tiger conservation landscapes and protected areas.
Table S1. Underlying data for increasing representation of ecoregions by adding unprotected areas of high priority.
REFERENCES AND NOTES
- 1.Rockström J., Steffen W., Noone K., Persson Å., Chapin F. S. III, Lambin E. F., Lenton T. M., Scheffer M., Folke C., Schellnhuber H. J., Nykvist B., de Wit C. A., Hughes T., van der Leeuw S., Rodhe H., Sörlin S., Snyder P. K., Costanza R., Svedin U., Falkenmark M., Karlberg L., Corell R. W., Fabry V. J., Hansen J., Walker B., Liverman D., Richardson K., Crutzen P., Foley J. A., A safe operating space for humanity. Nature 461, 472–475 (2009). [DOI] [PubMed] [Google Scholar]
- 2.S. Teske, Achieving the Paris Climate Agreement (Springer, 2019).
- 3.Tallis H. M., Hawthorne P. L., Polasky S., Reid J., Beck M. W., Brauman K., Bielicki J. M., Binder S., Burgess M. G., Cassidy E., Clark A., Fargione J., Game E. T., Gerber J., Isbell F., Kiesecker J., McDonald R., Metian M., Molnar J. L., Mueller J. L., O’Connell C., Ovando D., Troell M., Boucher T. M., McPeek B., An attainable global vision for conservation and human well-being. Front. Ecol. Environ. 16, 563–570 (2018). [Google Scholar]
- 4.Griscom B. W., Adams J., Ellis P. W., Houghton R. A., Lomax G., Miteva D. A., Schlesinger W. H., Shoch D., Siikamäki J. V., Smith P., Woodbury P., Zganjar C., Blackman A., Campari J., Conant R. T., Delgado C., Elias P., Gopalakrishna T., Hamsik M. R., Herrero M., Kiesecker J., Landis E., Laestadius L., Leavitt S. M., Minnemeyer S., Polasky S., Potapov P., Putz F. E., Sanderman J., Silvius M., Wollenberg E., Fargione J., Natural climate solutions. Proc. Natl. Acad. Sci. U.S.A. 114, 11645–11650 (2017). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 5.Martin T. G., Watson J. E. M., Intact ecosystems provide best defence against climate change. Nat. Clim. Chang. 6, 122–124 (2016). [Google Scholar]
- 6.Seddon N., Turner B., Berry P., Chausson A., Giradin C. A. J., Grounding nature-based climates solutions in sound biodiversity science. Nat. Clim. Chang. 9, 84–87 (2019). [Google Scholar]
- 7.Baccini A., Walker W., Carvalho L., Farina M., Sulla-Menashe D., Houghton R. A., Tropical forests are a net carbon source based on aboveground measurements of gain and loss. Science 358, 230–234 (2017). [DOI] [PubMed] [Google Scholar]
- 8.Huang Y., Chen Y., Castro-Izaguirre N., Baruffol M., Brezzi M., Lang A., Li Y., Härdtle W., von Oheimb G., Yang X., Liu X., Pei K., Both S., Yang B., Eichenberg D., Assmann T., Bauhus J., Behrens T., Buscot F., Chen X.-Y., Chesters D., Ding B.-Y., Durka W., Erfmeier A., Fang J., Fischer M., Guo L.-D., Guo D., Gutknecht J. L. M., He J.-S., Hector C.-L. H. A., Hönig L., Hu R.-Y., Klein A.-M., Kühn P., Liang Y., Li S., Michalski S., Scherer-Lorenzen M., Schmidt K., Scholten T., Schuldt A., Shi X., Tan M.-Z., Tang Z., Trogisch S., Wang Z., Welk E., Wirth C., Wubet T., Xiang W., Yu M., Yu X.-D., Zhang J., Zhang S., Zhang N., Zhou H.-Z., Zhu C.-D., Zhu L., Bruelheide H., Ma K., Niklaus P. A., Schmid B., Impacts of species richness on productivity in a large-scale subtropical forest experiment. Science 362, 80–83 (2018). [DOI] [PubMed] [Google Scholar]
- 9.J. C. Vié, C. Hilton-Taylor, S. N. Stuart, Wildlife in a Changing World—An Analysis of the 2008 IUCN Red List of Threatened Species (International Union for Conservation of Nature and Natural Resources, 2008).
- 10.Mazurcyzk T., Brooks R. P., Carbon storage dynamics of temperate freshwater wetlands in Pennsylvania. Wetl. Ecol. Manag. 26, 893–914 (2018). [Google Scholar]
- 11.Duarte C. M., Middelburg J. J., Caraco N., Major role of marine vegetation on the oceanic carbon cycle. Biogeosci. Discuss. 1, 659–679 (2004). [Google Scholar]
- 12.Intergovernmental Panel on Climate Change, Global Warming of 1.5°C: An IPCC Special Report on the Impacts of Global Warming of 1.5°C Above Pre-Industrial Levels and Related Global Greenhouse Gas Emission Pathways, in the Context of Strengthening the Global Response to the Threat of Climate (Intergovernmental Panel on Climate Change, 2018).
- 13.Steffen W., Rockström J., Richardson K., Lenton T. M., Folke C., Liverman D., Summerhayes C. P., Barnosky A. D., Cornell S. E., Crucifix M., Donges J. F., Fetzer I., Lade S. J., Scheffer M., Winkelmann R., Schellnhuber R., Schellnhuber H. J., Trajectories of the Earth system in the Anthropocene. Proc. Natl. Acad. Sci. U.S.A. 115, 8252–8259 (2018). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 14.Lovejoy T. E., Hannah L., Avoiding the climate failsafe point. Sci. Adv. 4, eaau9981 (2018). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 15.Watson J. E. M., Evans T., Venter O., Williams B., Tulloch A., Stewart C., Thompson I., Ray J. C., Murray K., Salazar A., McAlpine C., Potapov P., Walston J., Robinson J. G., Painter M., Wilkie D., Filardi C., Laurence W. F., Houghton R. A., Maxwell S., Grantham H., Samper C., Wang S., Laestadius L., Runting R. K., Silva-Chávez G. A., Ervin J., Lindenmayer D., The exceptional value of intact forest ecosystems. Nat. Ecol. Evol. 2, 599–610 (2018). [DOI] [PubMed] [Google Scholar]
- 16.Dinerstein E., Olson D., Joshi A., Vynne C., Burgess N. D., Wikramanayake E., Hahn N., Palminteri S., Hedao P., Noss R., Hansen M., Locke H., Ellis E. C., Jones B., Barber C. V., Hayes R., Kormos C., Martin V., Crist E., Sechrest W., Price L., Baillie J. E. M., Weeden D., Suckling K., Davis C., Sizer N., Moore R., Thau D., Birch T., Potapov P., Turubanova S., Tyukavina A., de Souza N., Pintea L., Brito J. C., Llewellyn O. A., Miller A. G., Patzelt A., Ghazanfar S. A., Timberlake J., Klöser H., Shennan-Farpon Y., Kindt R., Lillesø J.-P. B., van Breugel P., Graudal L., Voge M., Al-Shammari K. F., Saleem M., An ecoregion-based approach to protecting half the terrestrial realm. Bioscience 67, 534–545 (2017). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 17.R. F. Noss, A. Cooperrider, Saving Nature’s Legacy: Protecting and Restoring Biodiversity (Island Press, 1994).
- 18.Myers N., Mittermeier R. A., Mittermeier C. G., da Fonseca G. A. B., Kent J., Biodiversity hotspots for conservation priorities. Nature 403, 853–858 (2000). [DOI] [PubMed] [Google Scholar]
- 19.Gray C. L., Hill S. L. L., Newbold T., Hudson L. N., Boïrger L., Contu S., Hoskins A. J., Ferrier S., Purvis A., Scharlemann J. P. W., Local biodiversity is higher inside than outside terrestrial protected areas worldwide. Nat. Commun. 7, 12306 (2016). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 20.Noss R. F., Dobson A. P., Baldwin R., Beier P., Davis C. R., Dellasala D. A., Francis J., Locke H., Nowak K., Lopez R., Reining C., Trombulak S. C., Tabor G., Bolder thinking for conservation. Conserv. Biol. 26, 1–4 (2012). [DOI] [PubMed] [Google Scholar]
- 21.Pressey R. L., Cowling R. M., Rouget M., Formulating conservation targets for biodiversity pattern and process in the Cape Floristic Region, South Africa. Biol. Conserv. 112, 99–127 (2003). [Google Scholar]
- 22.O’Leary B. C., Winther-Janson M., Bainbridge J. M., Aitken J., Hawkins J. P., Roberts C. M., Effective coverage targets for ocean protection. Conserv. Lett. 9, 398–404 (2016). [Google Scholar]
- 23.E. O. Wilson, Half Earth: Our Planet’s Fight for Life (Liveright Publishing Corporation, ed. 1, 2016).
- 24.E. Sala, K. Rechberger, in From Summits to Solutions: Innovations in Implementing the Sustainable Development Goals, R. Desai, H. Kato, H. Kharas, J. McArhur, Eds. (Brookings Institution Press, 2018), pp. 239–261.
- 25.Smith J. R., Letten A. D., Ke P.-J., Anderson C. B., Hendershot J. N., Dhami M. K., Dlott G. A., Grainger T. N., Howard M. E., Morrison B. M. L., Routh D., San Juan P. A., Mooney H. A., Mordecai E. A., Crowther T. W., Daily G. C., A global test of ecoregions. Nat. Ecol. Evol. 2, 1889–1896 (2018). [DOI] [PubMed] [Google Scholar]
- 26.Watson J. E. M., Jones K. R., Fuller R. A., Di Marco M., Segan D. B., Butchart S. H. M., Allan J. R., McDonald-Madden E., Venter O., Persistent disparities between recent rates of habitat conversion and protection and implications for future global conservation targets. Conserv. Lett. 9, 413–421 (2016). [Google Scholar]
- 27.Dellasala D. A., Goldstein M. I., Langhammer P. F., Butchart S. H. M., Brooks T. M., Key biodiversity areas. Encycl. Anthropol. 341–345 (2018). [Google Scholar]
- 28.Harris G., Thirgood S., Hopcraft J. G. C., Cromsigt J. P. G. M., Berger J., Global decline in aggregated migrations of large terrestrial mammals. Endanger. Species Res. 7, 55–76 (2009). [Google Scholar]
- 29.Littlefield C. E., McRae B. H., Michalak J. L., Lawler J. J., Carroll C., Connecting today’s climates to future climate analogs to facilitate movement of species under climate change. Conserv. Biol. 31, 1397–1408 (2017). [DOI] [PubMed] [Google Scholar]
- 30.Spalding M. D., Fox H. E., Allen G. R., Davidson N., Ferdaña Z. A., Finlayson M., Halpern B. S., Jorge M. A., Lombana A., Lourie S. A., Martin K. D., McManus E., Molnar J., Recchia C. A., Robertson J., Marine ecoregions of the world: A bioregionalization of coastal and shelf areas. Bioscience 57, 573–583 (2007). [Google Scholar]
- 31.Spalding M. D., Agostini V. N., Rice J., Grant S. M., Pelagic provinces of the world: A biogeographic classification of the world’s surface pelagic waters. Ocean Coast. Manag. 60, 19–30 (2012). [Google Scholar]
- 32.Sala E., Giakoumi S., No-take marine reserves are the most effective protected areas in the ocean. ICES J. Mar. Sci. 75, 1166–1168 (2018). [Google Scholar]
- 33.Hastings A., Botsford L. W., Equivalence in yield from marine reserves and traditional fisheries management. Science 284, 1537–1538 (1999). [DOI] [PubMed] [Google Scholar]
- 34.Guidetti P., Sala E., Community-wide effects of marine reserves in the Mediterranean Sea. Mar. Ecol. Prog. Ser. 335, 43–56 (2007). [Google Scholar]
- 35.Mee J. A., Otto S. P., Pauly D., Evolution of movement rate increases the effectiveness of marine reserves for the conservation of pelagic fishes. Evol. Appl. 10, 444–461 (2017). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 36.Roberts C. M., O’Leary B. C., McCauley D. J., Cury P. M., Cuarte C. M., Lubchenco J., Pauly D., Sáenz-Arroyo A., Sumaila U. R., Wilson R. W., Worm B., Castilla J. C., Marine reserves can mitigate and promote adaptation to climate change. Proc. Natl. Acad. Sci. U.S.A. 114, 6167–6175 (2017). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 37.Saura S., Bertzky B., Bastin L., Battistella L., Mandrici A., Dubois G., Protected area connectivity: Shortfalls in global targets and country-level priorities. Biol. Conserv. 219, 53–67 (2018). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 38.Loarie S. R., Duffy P. B., Hamilton H., Asner G. P., Field C. B., Ackerly D. D., The velocity of climate change. Nature 462, 1052–1055 (2009). [DOI] [PubMed] [Google Scholar]
- 39.T. E. Lovejoy, in Biodiversity and Climate Change, T. E. Lovejoy, L. Hannah, Eds. (Yale Univ. Press, 2019), pp. 326–331.
- 40.Baillie J., Zhang Y.-P., Space for nature. Science 361, 1051 (2018). [DOI] [PubMed] [Google Scholar]
- 41.Joppa L. N., Pfaff A., High and far: Biases in the location of protected areas. PLOS ONE 4, e8273 (2009). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 42.Dudley N., Jonas H., Nelson F., Parrish J., Phyhälä A., Stolton S., Watson J. E. M., The essential role of other effective area-based conservation measures in achieving big bold conservation targets. Glob. Ecol. Conserv. 15, e00424 (2018). [Google Scholar]
- 43.UNEP-WCMC, IUCN, and NGS, Protected Planet Report 2018 (UNEP-WCMC, IUCN, and NGS, 2018).
- 44.Pimm S. L., Jenkins C. N., Li B. V., How to protect half of earth to ensure it protects sufficient biodiversity. Sci. Adv. 4, eaat2616 (2018). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 45.IUCN, UNEP-WCMC, The World Database on Protected Areas (WDPA) (2018); www.protectedplanet.net.
- 46.Alliance for Zero Extinction (2018); zeroextinction.org.
- 47.Jenkins C. N., Van Houtan K. S., Global and regional priorities for marine biodiversity protection. Biol. Conserv. 204, 333–339 (2016). [Google Scholar]
- 48.Olson D. M., Dinerstein E., The Global 200: Priority ecoregions for global conservation. Ann. Mo. Bot. Gard. 89, 199–224 (2002). [Google Scholar]
- 49.Morrison J. C., Sechrest W., Dinerstein E., Wilcove D. S., Lamoreux J. F., Persistence of large mammal faunas as indicators of global human impacts. J. Mammal. 88, 1363–1380 (2007). [Google Scholar]
- 50.Ripple W. J., Abernethy K., Betts M. G., Chapron G., Dirzo R., Galetti M., Levi T., Lindsey P. A., Macdonald D. W., Machovina B., Newsome T. M., Peres C. A., Wallach A. D., Wolf C., Saving the world’s terrestrial megafauna. Bioscience 66, 807–812 (2016). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 51.Carrizo S. F., Jähnig S. C., Bremerich V., Freyhof J., Harrison I., He F., Langhans S. D., Tockner K., Zarfl C., Darwall W., Freshwater megafauna: Flagships for freshwater biodiversity under threat. Bioscience 67, 919–927 (2017). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 52.Estes J. A., Terborgh J., Brashares J. S., Power M. E., Berger J., Bond W. J., Carpenter S. R., Essington T. E., Holt R. D., Jackson J. B. C., Marquis R. J., Oksanen L., Oksanen T., Paine R. T., Pikitch E. K., Ripple W. J., Sandin S. A., Scheffer M., Schoener T. W., Shurin J. B., Sinclair A. R. E., Soulé M. E., Virtanen R., Wardle D. A., Trophic downgrading of planet earth. Science 333, 301–306 (2011). [DOI] [PubMed] [Google Scholar]
- 53.Bond W. J., Ancient grasslands at risk. Science 351, 120–122 (2016). [DOI] [PubMed] [Google Scholar]
- 54.Betts M. G., Wolf C., Ripple W. J., Phalan B., Millers K. A., Duarte A., Butchart S. H. M., Levi T., Global forest loss disproportionately erodes biodiversity in intact landscapes. Nature 547, 441–444 (2017). [DOI] [PubMed] [Google Scholar]
- 55.Gibson L., Lee T. M., Koh L. P., Brook B. W., Gardner T. A., Barlow J., Peres C. A., Bradshaw C. J. A., Laurance W. F., Lovejoy T. E., Sodhi N. S., Primary forests are irreplaceable for sustaining tropical biodiversity. Nature 478, 378–381 (2011). [DOI] [PubMed] [Google Scholar]
- 56.Dass P., Houlton B. Z., Wang Y., Warlind D., Grasslands may be more reliable carbon sinks than forests in California. Environ. Res. Lett. 13, 074027 (2018). [Google Scholar]
- 57.Strayer D. L., Dudgeon D., Freshwater biodiversity conservation: Recent progress and future challenges. J. North Am. Benthol. Soc. 29, 344–358 (2010). [Google Scholar]
- 58.Abell R., Thieme M. L., Revenga C., Bryer M., Kottelat M., Bogutskaya N., Coad B., Mandrak N., Balderas S. C., Bussing W., Stiassny M. L. J., Skelton P., Allen G. R., Unmack P., Naseka A., Ng R., Sindorf N., Robertson J., Armijo E., Higgins J. V., Heibel T. J., Wikramanayake E., Olson D., López H. L., Reis R. E., Lundberg J. G., Sabaj Pérez M. H., Petry P., Freshwater ecoregions of the world: A new map of biogeographic units for freshwater biodiversity conservation. Bioscience 58, 403–414 (2008). [Google Scholar]
- 59.Sala E., Lubchenco J., Grorud-Colvert K., Novelli C., Roberts C., Rashid S. U., Assessing real progress towards effective ocean protection. Mar. Policy 91, 11–13 (2018). [Google Scholar]
- 60.McLeod E., Chmura G. L., Bouillon S., Salm R., Björk M., Duarte C. M., Lovelock C. E., Schlesinger W. H., Silliman B. R., A blueprint for blue carbon: Toward an improved understanding of the role of vegetated coastal habitats in sequestering CO2. Front. Ecol. Environ. 9, 552–560 (2011). [Google Scholar]
- 61.Fourqurean J. W., Duarte C. M., Kennedy H., Marbà N., Holmer M., Mateo M. A., Apostolaki E. T., Kendrick G. A., Krause-Jensen D., McGlathery K. J., Serrano O., Seagrass ecosystems as a globally significant carbon stock. Nat. Geosci. 5, 505–509 (2012). [Google Scholar]
- 62.S. Samadi, T. A. Schlacher, B. Richer de Forges, Seamount benthos, in Seamounts: Ecology, Fisheries and Conservation, T. J. Pitcher, T. Morato, P. J. B. Hart, M. R. Clark, N. Haggan, R. S. Santos, Eds. (Wiley/Blackwell, 2007), pp. 119–140.
- 63.Mateo M. A., Romero J., Pérez M., Littler M. M., Littler D. S., Dynamics of millenary organic deposits resulting from the growth of the Mediterranean seagrass Posidonia oceanica. Estuar. Coast. Shelf Sci. 44, 103–110 (1997). [Google Scholar]
- 64.Atwood T. B., Connolly R. M., Ritchie E. G., Lovelock C. E., Heithaus M. R., Hays G. C., Fourqurean J. W., Macreadie P. I., Predators help protect carbon stocks in blue carbon ecosystems. Nat. Clim. Chang. 5, 1038–1045 (2015). [Google Scholar]
- 65.Nobre C. A., Sampaio G., Borma L. S., Castilla-Rubio J. C., Silva J. S., Cardoso M., Land-use and climate change risks in the Amazon and the need of a novel sustainable development paradigm. Proc. Natl. Acad. Sci. U.S.A. 113, 10759–10768 (2016). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 66.Lovejoy T. E., Nobre C., Amazon tipping point. Sci. Adv. 4, eaat2340 (2018). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 67.Asner G. P., Brodrick P. G., Philipson C., Vaughn N. R., Martin R. E., Knapp D. E., Heckler J., Evans L. J., Jucker T., Goossens B., Stark D. J., Reynolds G., Ong R., Renneboog N., Kugan F., Coomes D. A., Mapped aboveground carbon stocks to advance forest conservation and recovery in Malaysian Borneo. Biol. Conserv. 217, 289–310 (2018). [Google Scholar]
- 68.Dargie G. C., Lewis S. L., Lawson I. T., Mitchard E. T. A., Page S. E., Bocko Y. E., Ifo S. A., Age, extent and carbon storage of the central Congo Basin peatland complex. Nature 542, 86–90 (2017). [DOI] [PubMed] [Google Scholar]
- 69.Cromsigt J. P., te Beest M., Kerley G. I., Landman M., le Roux E., Smith F. A., Trophic rewilding as a climate change mitigation strategy? Philos. Trans. R. Soc. Lond. B Biol. Sci. 373, 20170440 (2018). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 70.Bello C., Galetti M., Pizo M. A., Magnago L. F. S., Rocha M. R., Lima R. A. F., Peres C. A., Ovaskainen O., Jordano P., Defaunation affects carbon storage in tropical forests. Sci. Adv. 1, e1501105 (2015). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 71.Dinerstein E., Loucks C., Wikramanayake E., Ginsberg J., Sanderson E., Seidensticker J., Forrest J., Bryja G., Heydlauff A., Klenzendorf S., Leimgruber P., Mills J., O’Brien T. G., Shrestha M., Simons R., Songer M., The fate of wild tigers. Bioscience 57, 508–514 (2007). [Google Scholar]
- 72.Joshi A. R., Dinerstein E., Wkramanayake E., Anderson M. L., Olson D., Jones B. S., Seidensticker J., Lumpkin S., Hansen M. C., Sizer N. C., Davis C. L., Palminteri S., Hahn N. R., Tracking changes and preventing loss in critical tiger habitat. Sci. Adv. 2, e1501675 (2016). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 73.UNEP, Bonn Challenge (2011); www.bonnchallenge.org/content/challenge.
- 74.Garnett S. T., Burgess N. D., Fa J. E., Fernández-Llamazares Á., Molnár Z., Robinson C. J., Watson J. E. M., Zander K. K., Austin B., Brondizio E. S., Collier N. F., Duncan T., Ellis E., Geyle H., Jackson M. V., Jonas H., Malmer P., McGowan B., Sivongxay A., Leiper I., A spatial overview of the global importance of Indigenous lands for conservation. Nat. Sustain. 1, 369–374 (2018). [Google Scholar]
- 75.Blackman A., Corral L., Lima E. S., Asner G. P., Titling indigenous communities protects forests in the Peruvian Amazon. Proc. Natl. Acad. Sci. U.S.A. 114, 4123–4128 (2017). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 76.Mauser W., Klepper G., Zabel F., Delzeit R., Hank T., Putzenlechner B., Calzadilla A., Global biomass production potentials exceed expected future demand without the need for cropland expansion. Nat. Commun. 6, 8946 (2015). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 77.Erb K.-H., Kastner T., Plutzar C., Bais A. L. S., Carvalhais N., Fetzel T., Gingrich S., Haberl H., Lauk C., Niedertscheider M., Pongratz J., Thurner M., Luyssaert S., Unexpectedly large impact of forest management and grazing on global vegetation biomass. Nature 553, 73–76 (2018). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 78.Laurance W. F., Clements G. R., Sloan S., O’Connell C. S., Mueller N. D., Goosem M., Venter O., Edwards D. P., Phalan B., Balmford A., Van Der Ree R., Arrea I. B., A global strategy for road building. Nature 513, 229–232 (2014). [DOI] [PubMed] [Google Scholar]
- 79.Laurance W. F., Conservation and the global infrastructure tsunami: Disclose, debate, delay! Trends Ecol. Evol. 33, 568–571 (2018). [DOI] [PubMed] [Google Scholar]
- 80.Tilman D., Clark M., Williams D. R., Kimmel K., Polasky S., Packer C., Future threats to biodiversity and pathways to their prevention. Nature 546, 73–81 (2017). [DOI] [PubMed] [Google Scholar]
- 81.Kroodsma D. A., Mayorga J., Hochberg T., Miller N. A., Boerder K., Ferretti F., Wilson A., Bergman B., White T. D., Block B. A., Woods P., Sullivan B., Costello C., Worm B., Tracking the global footprint of fisheries. Science 359, 904–908 (2018). [DOI] [PubMed] [Google Scholar]
- 82.Mora C., Myers R. A., Coll M., Libralato S., Pitcher T. J., Sumaila R. U., Zeller D., Watson R., Gaston K. J., Worm B., Management effectiveness of the world’s marine fisheries. PLOS Biol. 7, e1000131 (2009). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 83.World Bank, The Sunken Billions Revisited: Progress and Challenges in Global Marine Fisheries (World Bank, 2017).
- 84.Seebens H., Blackburn T. M., Dyer E. E., Genovesi P., Hulme P. E., Jeschke J. M., Pagad S., Pyšek P., Winter M., Arianoutsou M., Vacher S., Blasius B., Brundu G., Capinha C., Celesti-Grapow L., Dawson W., Dullinger S., Fuentes N., Jäger H., Kartesz J., Kenis M., Kreft H., Kühn I., Lenzner B., Kiebhold A., Mosena A., Moser D., Nishino M., Pearman D., Pergl J., Rabitsch W., Rojas-Sandoval J., Roques A., Rorke S., Rossinelli S., Roy H. E., Scalera R., Schindler S., Štajerová K., Tokarska-Guzik B., Van Kleunen M., Walker K., Weigelt P., Yamanaka T., Essl F., No saturation in the accumulation of alien species worldwide. Nat. Commun. 8, 14435 (2017). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 85.Jambeck J. R., Geyer R., Wilcox C., Siegler T. R., Perryman M., Andrady A., Narayan R., Law K. L., Plastic waste inputs from land into the ocean. Science 347, 768–771 (2015). [DOI] [PubMed] [Google Scholar]
- 86.Geng Y., Sarkis J., Ulgiati S., Zhang P., Measuring China’s circular economy. Science 339, 1526–1527 (2013). [DOI] [PubMed] [Google Scholar]
- 87.B. Buchner, M. Stadelmann, J. Wilkinson, F. Mazza, A. Rosenberg, D. Abramskiehn, Global Landscape of Climate Finance 2014 (Climate Policy Initiative, 2014).
- 88.Foley J. A., Ramankutty N., Brauman K. A., Cassidy E. S., Gerber J. S., Johnston M., Mueller N. D., O’Connell C., Ray D. K., West P. C., Balzer C., Bennett E. M., Carpenter S. R., Hill J., Monfreda C., Polasky S., Rockström J., Sheehan J., Siebert S., Tilman D., Zaks D. P. M., Solutions for a cultivated planet. Nature 478, 337–342 (2011). [DOI] [PubMed] [Google Scholar]
- 89.Crist E., Mora C., Engelman R., The interaction of human population, food production, and biodiversity protection. Science 356, 260–264 (2017). [DOI] [PubMed] [Google Scholar]
- 90.Barbier E. B., Burgess J. C., Dean T. J., How to pay for saving biodiversity. Science 360, 486–488 (2018). [DOI] [PubMed] [Google Scholar]
- 91.Sala E., Costello C., De Bourbon Parme J., Fiorese M., Heal G., Kelleher K., Moffitt R., Morgan L., Plunkett J., Rechberger K. D., Rosenberg A. A., Sumaila R., Fish banks: An economic model to scale marine conservation. Mar. Policy 73, 154–161 (2016). [Google Scholar]
- 92.Pimm S. L., Jenkins C. N., Abell R., Brooks T. M., Gittleman J. L., Joppa L. N., Raven P. H., Roberts C. M., Sexton J. O., The biodiversity of species and their rates of extinction, distribution, and protection. Science 344, 1246752 (2014). [DOI] [PubMed] [Google Scholar]
- 93.Couto T. B. A., Olden J. D., Global proliferation of small hydropower plants – science and policy. Front. Ecol. Environ. 16, 91–100 (2018). [Google Scholar]
- 94.Erb K. H., Lauk C., Kastner T., Mayer A., Theurl M. C., Haberl H., Exploring the biophysical option space for feeding the world without deforestation. Nat. Commun. 7, 11382 (2016). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 95.Phalan B., Balmford A., Green R. E., Scharlemann J. P. W., Minimising the harm to biodiversity of producing more food globally. Food Policy 36, S62–S71 (2011). [Google Scholar]
- 96.Veach V., Moilanen A., Di Minin E., Threats from urban expansion, agricultural transformation and forest loss on global conservation priority areas. PLOS ONE 12, e0188397 (2017). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 97.Laurance W. F., Arrea I. B., Roads to riches or ruin? Science 358, 442–444 (2017). [DOI] [PubMed] [Google Scholar]
- 98.Lehner B., Liermann C. R., Revenga C., Vörömsmarty C., Fekete B., Crouzet P., Döll P., Endejan M., Frenken K., Magome J., Nilsson C., Robertson J. C., Rödel R., Sindorf N., Wisser D., High-resolution mapping of the world’s reservoirs and dams for sustainable river-flow management. Front. Ecol. Environ. 9, 494–502 (2011). [Google Scholar]
- 99.Pauly D., Zeller D., Catch reconstructions reveal that global marine fisheries catches are higher than reported and declining. Nat. Commun. 7, 10244 (2016). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 100.Maxwell S. L., Fuller R. A., Brooks T. M., Watson J. E. M., Biodiversity: The ravages of guns, nets and bulldozers. Nature 536, 143–145 (2016). [DOI] [PubMed] [Google Scholar]
- 101.Harfoot M., Glaser S. A. M., Tittensor D. P., Britten G. L., McLardy C., Malsch K., Burgess N. D., Unveiling the patterns and trends in 40 years of global trade in CITES-listed wildlife. Biol. Conserv. 223, 47–57 (2018). [Google Scholar]
- 102.Dauvergne P., Why is the global governance of plastic failing the oceans? Glob. Environ. Chang. 51, 22–31 (2018). [Google Scholar]
- 103.Rhodes C. J., Pollinator decline—An ecological calamity in the making? Sci. Prog. 101, 121–160 (2018). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 104.Lunt M. F., Park S., Li S., Henne S., Manning A. J., Ganesan A. L., Simpson I. J., Blake D. R., Liang Q., O’Doherty S., Harth C. M., Mühle J., Salameh P. K., Weiss R. F., Krummel P. B., Fraser P. J., Prinn R. G., Reimann S., Rigby M., Continued emissions of the ozone-depleting substance carbon tetrachloride from eastern Asia. Geophys. Res. Lett. 45, 11423–11430 (2018). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 105.M. Santoro, GlobBiomass—Global datasets of forest biomass. PANGAEA (2018).
- 106.Bouvet A., Mermoz S., Le Toan T., Villard L., Mathieu R., Naidoo L., Asner G. P., An above-ground biomass map of African savannahs and woodlands at 25 m resolution derived from ALOS PALSAR. Remote Sens. Environ. 206, 156–173 (2018). [Google Scholar]
- 107.Xia J., Liu S., Liang S., Chen Y., Xu W., Yuan W., Spatio-temporal patterns and climate variables controlling of biomass carbon stock of global grassland ecosystems from 1982 to 2006. Remote Sens. 6, 1783–1802 (2014). [Google Scholar]
- 108.S. A. Spawn, T. J. Lark, H. K. Gibbs, A New Synthetic Global Biomass Carbon Map for the Year 2010. Am. Geophys. Union Fall Meet. Abstr. (2017).
- 109.Hengl T., Mendes de Jesus J., Heuvelink G. B. M., Ruiperez Gonzalez M., Kilibarda M., Blagotić A., Shangguan W., Wright M. N., Geng X., Bauer-Marschallinger B., Guevara M. A., Vargas R., MacMillan R. A., Batjes N. H., Leenaars J. G. B., Ribeiro E., Wheeler I., Mantel S., Kempen B., SoilGrids250m: Global gridded soil information based on machine learning. PLOS ONE 12, e0169748 (2017). [DOI] [PMC free article] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.
Supplementary Materials
Supplementary material for this article is available at http://advances.sciencemag.org/cgi/content/full/5/4/eaaw2869/DC1
Section S1. Maps of important biodiversity and carbon layers
Section S2. Underlying data for increasing representation of ecoregions by adding unprotected areas of high priority
Section S3. Monitoring progress under the GDN from the ground to below the sea to space
Fig. S1. Maps used to increase representation among terrestrial ecoregions and unprotected sites of biodiversity importance that contribute to the global milestone of 30% protected by 2030.
Fig. S2. Maps showing total terrestrial carbon and overlap with intact large mammal assemblages.
Fig. S3. Overlay of tiger conservation landscapes and protected areas.
Table S1. Underlying data for increasing representation of ecoregions by adding unprotected areas of high priority.