

Abstract
The anaerobic gut microbial pathway that converts choline into trimethylamine (TMA) is broadly linked to human disease. Here, we describe the discovery that betaine aldehyde inhibits TMA production from choline by human gut bacterial isolates and a complex gut community. In vitro assays and a crystal structure suggest betaine aldehyde targets the gut microbial enzyme choline TMA-lyase (CutC). In our system, we do not observe activity for the previously reported CutC inhibitor 3,3-dimethylbutanol (DMB). The workflow we establish for identifying and characterizing betaine aldehyde provides a framework for developing additional inhibitors of gut microbial choline metabolism, including therapeutic candidates.
Graphical Abstract
Microorganisms living within the human gastrointestinal (GI) tract (the gut microbiota) produce a diverse repertoire of small molecules, some of which are associated with disease.1,2 Deciphering the roles of these metabolites in disease development and progression has proven challenging. Most methods for manipulating the gut microbiota (antibiotics, probiotics, and prebiotics) alter multiple microbial activities. However, elucidating the mechanisms by which gut microbial metabolites affect host biology requires modulating specific transformations within this community. The development of small molecules that selectively inhibit gut microbial enzymes is a promising but underexplored approach for addressing this challenge.
Anaerobic metabolism of the essential nutrient choline is a prominent, disease-associated gut microbial activity.3 Gut anaerobes use choline as a source of carbon and energy, cleaving its C–N bond to generate trimethylamine (TMA) and acetaldehyde (Figure 1A). TMA is metabolized in the liver to trimethylamine-N-oxide (TMAO) by flavin monooxygenases (FMOs) and excreted.4,5 Elevated levels of plasma TMAO in humans are linked to nonalcoholic fatty liver disease,6,7 cardiovascular disease,8–11 chronic kidney disease,12,13 and type 2 diabetes.14 Additionally, patients with the inherited metabolic disorder trimethylaminuria exhibit decreased FMO3 activity and excrete TMA.15–17 Despite the many connections between this activity and human health, the mechanisms by which choline-degrading gut microbes affect host physiology remain largely uncharacterized.
Figure 1.
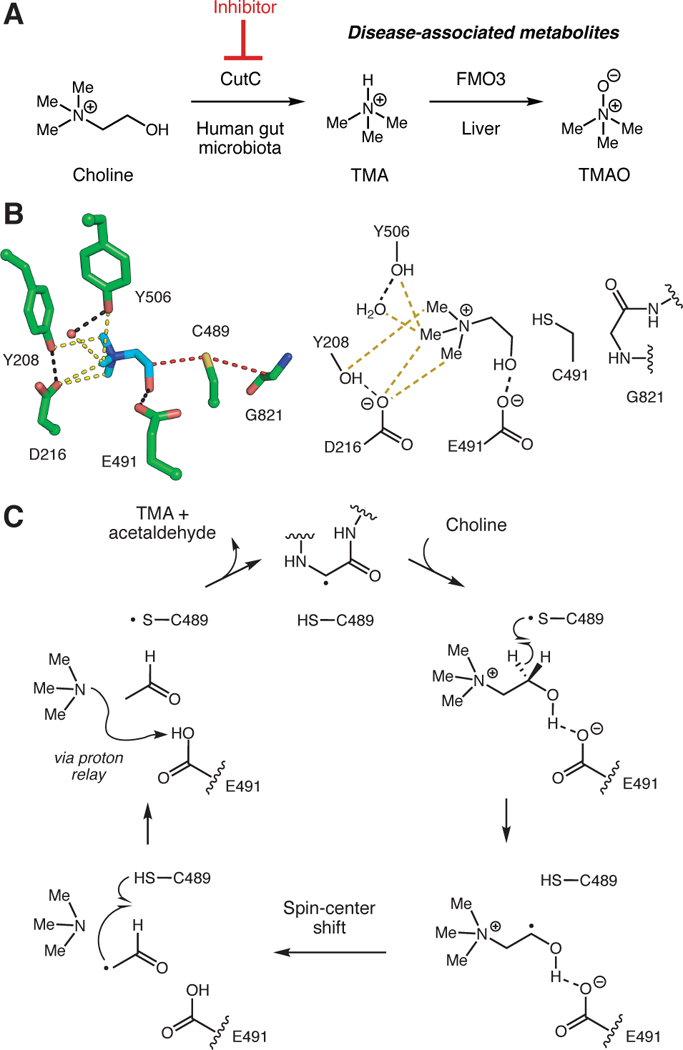
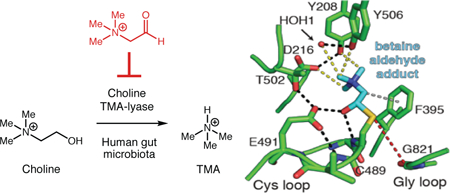
Anaerobic choline metabolism is a disease-linked gut microbial activity. (A) Microbial generation of TMA from choline and its subsequent processing in the human body. (B) Crystal structure of CutC (PBD = 5FAU) and (C) proposed mechanism for choline cleavage.
The choline utilization (cut) gene cluster encodes the enzymes involved in anaerobic microbial choline metabolism, including the glycyl radical enzyme choline TMA-lyase (CutC) and its corresponding radical SAM activating protein (CutD).18–20 CutD postranslationally modifies CutC, installing a radical on a conserved glycine residue and enabling catalysis of the key C–N bond cleavage reaction that generates TMA. We gained insights into the mechanism of this transformation from structural studies and mutagenesis (Figure 1B,C).20 The glycyl radical of CutC is proposed to abstract a hydrogen atom from an adjacent cysteine (Cys489), generating a thiyl radical intermediate that initiates the reaction with choline. Within the polar active site of CutC, choline is oriented such that its C1 hydroxyl engages in hydrogen bonding with Glu491. The primary substrate-specific contacts are an array of CH—O hydrogen bonds between the polarized methyl groups of the trimethylammonium moiety and the side-chain oxygen atoms of Tyr208, Tyr506, Asp216, and a bound water molecule. We propose the thiyl radical abstracts the pro-S hydrogen atom from C1 of choline. Subsequent deprotonation of the C1-OH by Glu491, followed by C–N bond cleavage via a ‘spin-center shift’ would eliminate trimethylamine. The resulting product-based radical could re-abstract the hydrogen atom from Cys489, reforming the thiyl radical and generating acetaldehyde. Proton transfer from Glu491 to Asp216 via Thr502 could reset the CutC active site for the next round of catalysis.
These observations informed our search for small molecule inhibitors of CutC. We identified a series of choline analogs that would preserve many of the key interactions with CutC active site residues, including the CH—O hydrogen bonds. These molecules were tested for inhibition of anaerobic choline metabolism by the cut gene cluster-containing human gut isolate Escherichia coli MS 200–1 (Figure 2A). Cultures were incubated anaerobically in a rich medium containing 1 mM of (trimethyl-d9)-choline (d9-choline) and three concentrations (1, 5, and 10 mM) of compound. Production of d9-TMA was analyzed by LC-MS/MS. Thiocholine (2) and betaine aldehyde (9) emerged as promising inhibitors. Notably, we observed no inhibition with DMB (12), a previously reported CutC inhibitor, and only modest inhibition with its oxidized analog 3,3-dimethylbutyric aldehyde (DMBA) (13).21
Figure 2.
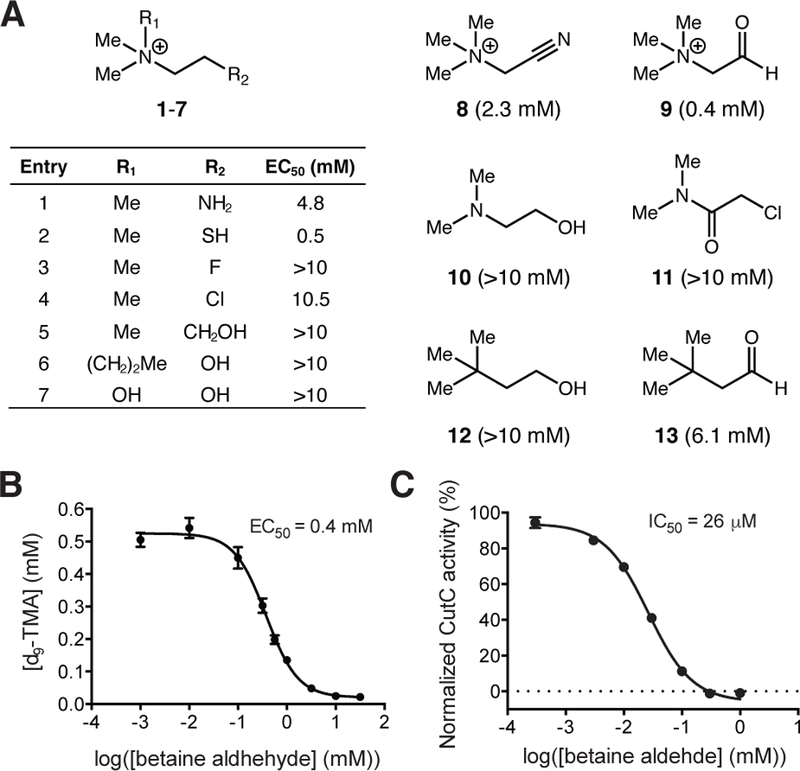
Betaine aldehyde inhibits TMA formation in E. coli and in vitro. (A) Choline analogs were tested for inhibition of anaerobic choline metabolism by E. coli MS 200–1. Bacteria were incubated in BHI medium with 1 mM d9-choline and 1, 5, or 10 mM of inhibitor at 37 °C. Production of d9-TMA was measured by LC-MS/MS after 3.5 hours. Dose-response curves were fit using nonlinear regression to determine EC50 values. (B) Betaine aldehyde inhibits anaerobic d9-TMA formation in E. coli MS 200–1 in a dose-dependent manner. Bacteria were incubated in BHI medium with 1 mM d9-choline and inhibitor (1 μM to 30 mM) at 37 °C. Production of d9-TMA was measured by LC-MS/MS after 3.5 hours. The data was fit using nonlinear regression. Error bars represent the mean ± SD of three replicates. (C) Betaine aldehyde inhibits the activity of CutC in vitro. Betaine aldehyde (0.30 μM to 1 mM) was added immediately before choline. The activity of CutC was coupled to that of NADH-dependent yeast alcohol dehydrogenase and the absorbance of NADH at 340 nm was recorded over time. The data was fit using nonlinear regression. Error bars represent the mean ± SD of three replicates.
We chose to further examine the activity and mechanism of inhibition of betaine aldehyde. This compound exhibited an EC50 value of 0.4 mM in the whole cell assay (Figure 2B). To assess whether the reduction in TMA resulted from inhibition of CutC, we performed in vitro assays. Recombinant wild-type CutC and CutD from D. alaskensis G20 were expressed and purified.19,20 Using a 7-point dilution series, the IC50 of betaine aldehyde for CutC was determined to be 26 ± 2 μM (Figure 2C). Thus, betaine aldehyde’s ability to inhibit TMA production by E. coli may arise from direct targeting of CutC. DMB did not inhibit CutC, even at the highest concentrations tested (Figure S1).
To gain information about how betaine aldehyde inhibits CutC, we performed X-ray crystallographic studies on the unactivated enzyme-inhibitor complex. After pre-incubating wild-type CutC with 10 mM of betaine aldehyde, we obtained a homogenous solution that was crystalized using conditions similar to those reported previously.20 The resulting structure (Table S1–2) was determined to 2.35-Å resolution and showed a large continuous patch of density around Cys489. We modeled a thiohemiacetal adduct of betaine aldehyde at Cys489 (Figure 3A) and validated this assignment through composite omit electron density maps (Figure 3B). This betaine aldehyde adduct interacts with CutC active site residues in a similar manner to choline, with the thiohemiacetal alcohol hydrogen bonding with Glu491 (2.5 Å) and the backbone amide of Cys489 (2.7 Å) (Figure 3C). The trimethylammonium moiety is shifted slightly away from the rear of the active site, where residues Asp216, Tyr208, Tyr506, and a water molecule make CH–O hydrogen bonds to the methyl groups, slightly altering the distances from those observed in the choline-bound structure (Table S3). Also, the α-carbon of betaine aldehyde is shifted closer to Phe395 (3.5 Å vs. 3.8 Å), a residue thought to make a cation-π-like interaction with the substrate. Overall, betaine aldehyde appears to be well accommodated in this active site. The observed covalent interaction could explain betaine aldehyde’s ability to inhibit CutC and the apparent decrease in IC50 upon preincubation of inhibitor with the enzyme (Figure S2). Alternatively, inhibition could result from a long dwell time of this molecule in the active site due to these favorable interactions.
Figure 3.
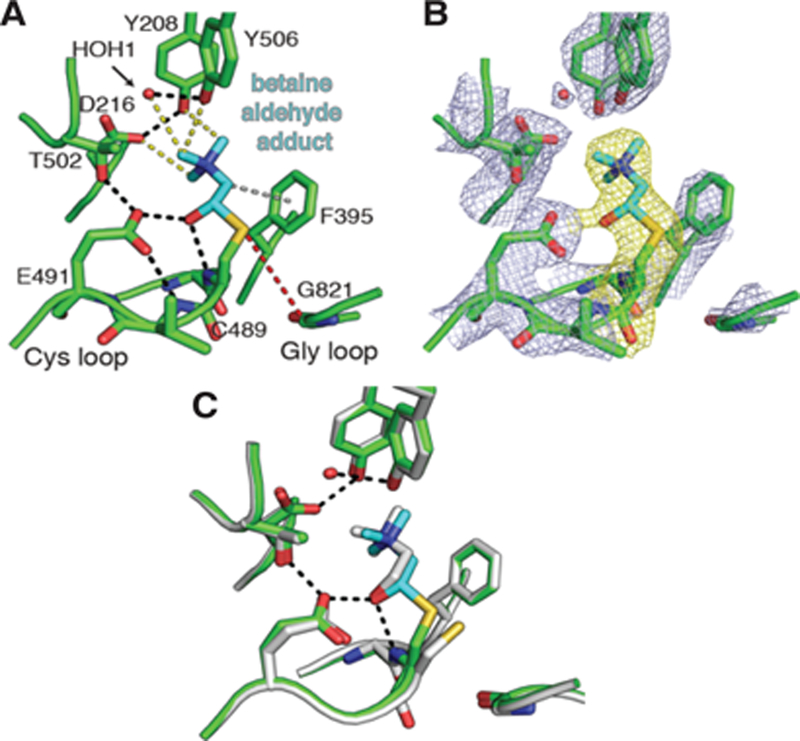
Betaine aldehyde binding to CutC appears to involve a covalent, thiohemiacetal linkage. (A) Crystal structure of unactivated CutC soaked with betaine aldehyde (10 mM) reveals interactions between CutC active site residues and bound inhibitor (cyan). Shown are traditional hydrogen bonds (2.5–3.2 Å, black dashes), CH—O hydrogen bonds (3.2–3.7 Å, yellow dashes), and a cation-π-like interaction (gray dashes). (B) Composite omit 2Fo-Fc density (light blue) contoured at 1.8σ. Density surrounding betaine aldehydemodified Cys493 is highlighted (yellow). (C) Comparison of inhibitor-bound (green/cyan) and choline-bound (grey) CutC structures. Hydrogen bonding interactions with the Cys loop (black dashes) are shown.
CutC is essential for the anaerobic growth of cut gene cluster-containing E. coli when choline is provided as the sole carbon source and fumarate is a terminal electron acceptor.22 We thus explored whether inhibition of CutC by betaine aldehyde would affect anaerobic growth of the choline-metabolizing strain E. coli MS 69–1 in minimal medium containing 30 mM choline as the sole carbon source. Under an N2 atmosphere, 20 mM of betaine aldehyde effectively abolished growth on choline but had no impact on growth in minimal medium containing 30 mM glycerol as the sole carbon source or growth in BHI, a complex medium containing 2 g/L of glucose (Figure 4A), suggesting that betaine aldehyde may specifically target anaerobic choline metabolism. This result, along with previous work in gnotobiotic animal models,22 indicates that anaerobic choline metabolism is non-essential when alternative carbon sources are present and suggests that inhibiting CutC activity may not dramatically alter gut microbiota composition. Finally, as betaine aldehyde is endogenously produced in bacteria as an intermediate in the conversion of choline to glycine betaine, we monitored its stability in cultures grown anaerobically on choline (Figure S3) and BHI media (Figure S4). We did not observe metabolism of d9-betaine aldehyde in either condition.
Figure 4.
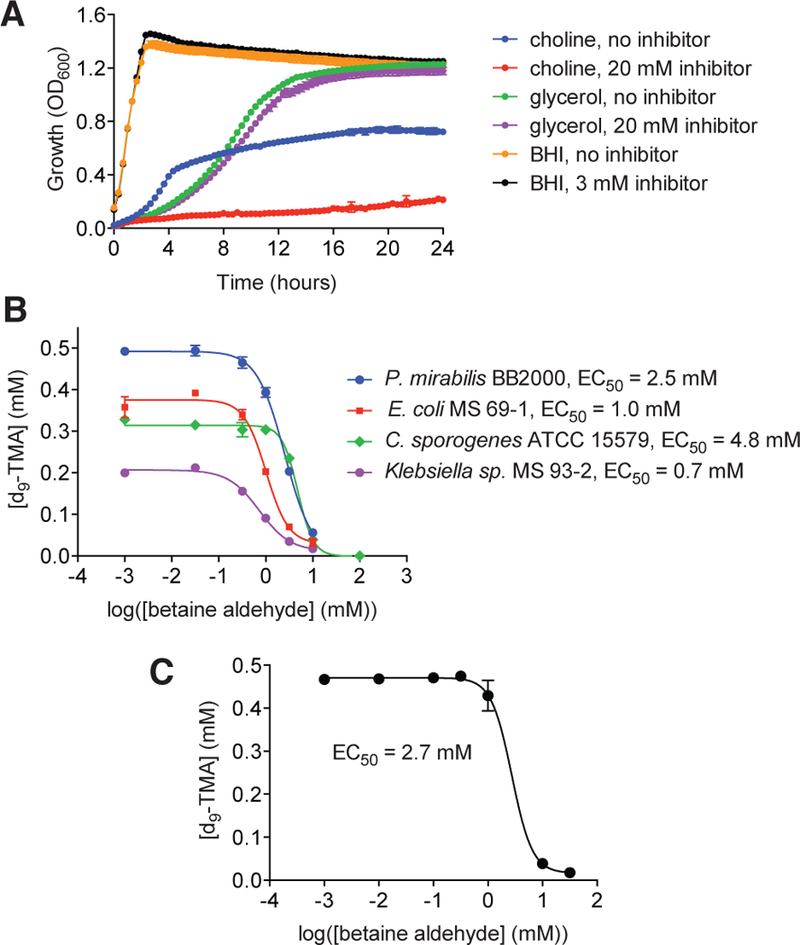
Betaine aldehyde inhibits anaerobic growth only on choline and prevents choline metabolism in both gut bacterial cultures and a human fecal suspension. (A) Growth of E. coli MS 69–1 on minimal media containing 30 mM choline or 30 mM glycerol and on BHI, a rich medium. Cultures were supplemented with either phosphate buffer (no inhibitor) or betaine aldehyde and grown under N2. (B) Betaine aldehyde inhibits d9-TMA formation from d9-choline in cut gene cluster-harboring gut bacteria, including Proteus mirabilis BB2000, Escherichia coli MS 69–1, Clostridium sporogenes ATCC 15579, and Klebsiella sp. MS 93–2. Production of d9-TMA was measured by LC-MS/MS. Error bars represent the mean ± SD of three replicates. (C) Betaine aldehyde inhibits d9-TMA formation from d9-choline by a human fecal suspension. Production of d9-TMA was measured by LC-MS/MS. Error bars represent the mean ± SD of two replicates.
The cut genes are widely distributed among human gut bacteria and likely encode a major pathway for choline utilization in the GI tract.23 We therefore tested whether betaine aldehyde could inhibit TMA production by additional Gram-positive (Clostridium sporogenes ATCC 15579) and Gram-negative (Proteus mirabilis BB2000, Escherichia coli MS 69–1, Klebsiella sp. MS 93–2) choline-metabolizing human gut isolates. Betaine aldehyde was effective against all strains assayed, with EC50 values ranging from 1 to 5 mM (Figure 4B). Finally, betaine aldehyde inhibited the conversion of choline to TMA by a human fecal suspension ex vivo (Figure 4C), confirming its efficacy in a complex setting more reminiscent of the gut microbiota. Overall, these findings demonstrate the feasibility of inhibiting anaerobic choline metabolism across a range of bacteria and in a complex mixture.
Previous work has explored inhibiting microbial choline metabolism as a potential treatment for atherosclerosis.21 The lead compound from this study, DMB (12), attenuated plasma TMAO levels and reduced the size of aortic lesions in mice fed a high-choline diet. Described as a specific inhibitor of TMA production, DMB has been used in additional animal studies.24,25 Unexpectedly, we found DMB did not reduce TMA production from choline by CutC in vitro or in E. coli. This result may be explained by the inability of DMB to engage in critical hydrogen bonding interactions within CutC’s polar active site. As DMB is rapidly metabolized in vivo by liver alcohol dehydrogenase,21 we wondered whether an oxidized metabolite could be responsible for the phenotypes observed in mice. However, the corresponding aldehyde, DMBA (13), was a relatively poor inhibitor of TMA generation by E. coli. Other aldehydes lacking a trimethylammonium moiety were also poor inhibitors of CutC activity in vitro (Figure S5). Using the same P. mirabilis strain employed by Wang et al., we observed minimal inhibition of TMA production by whole cells with DMB and DMBA (5 mM), whereas betaine aldehyde had an EC50 of 74.0 μM (Figure S6). DMB was also ineffective at inhibiting TMA generation by P. mirabilis cell lysates (Figure S7). We therefore conclude that DMB does not directly target anaerobic choline metabolism under these conditions and may not be an appropriate tool compound for studying this microbial activity. These findings also suggest the TMAO-lowering activity of DMB in vivo may not originate from inhibiting CutC.
In summary, we have identified a small molecule that inhibits the gut bacterial choline-metabolizing enzyme CutC. This compound and the workflow used to characterize its activity will inform future inhibitor development. Although small molecules that specifically disrupt pathogen-host interactions have been discovered,26 the use of such compounds to modulate activities from the human gut microbiota remains largely unexplored. Notable exceptions are gut bacterial β-glucuronidase inhibitors that prevent reactivation of glucuronide-drug conjugates.27 Small molecules that selectively modulate additional gut microbial activities would aid in deciphering microbe-microbe and microbe-host interactions that influence host biology and may provide a novel approach for treating disease.
Supplementary Material
ACKNOWLEDGMENT
We acknowledge support from the Packard Fellowship for Science and Engineering (E.P.B.), the National Science Foundation (EAGER MCB-1650086) (E.P.B.), and a Blavatnik Biomedical Accelerator Award (E.P.B.). S.B. is supported by an HHMI International Student Research Fellowship. M.A.F. is supported in part by the National Science Foundation Graduate Research Fellowship under grant no. 0645960. C.L.D. is a Howard Hughes Medical Institute Investigator. This work is based upon research conducted at the Advanced Photon Source on the North-eastern Collaborative Access Team beamlines, which are supported by award GM103403 from the National Center for Research Resources at the National Institute of Health. The Pilatus 6M detector on 24-ID-C beamline is funded by the Office of Research Infrastructure Programs High-End Instrumentation grant no. S10 RR029205. Use of the Advanced Photon Source is supported by the U.S. Department of Energy, Office of Basic Energy Sciences, under contract no. DE-AC02–06CH11357. We acknowledge Peter Turnbaugh (University of California San Francisco) for supplying the fecal sample.
Footnotes
ASSOCIATED CONTENT
Supporting Information.
The Supporting Information is available free of charge on the ACS Publications website.
Experimental data and procedures, crystallographic data (PDF)
REFERENCES
- (1).Sharon G; Garg N; Debelius J; Knight R; Dorrestein PC; Mazmanian SK Specialized metabolites from the microbiome in health and disease. Cell Metab 2014, 20 (5), 719–730. [DOI] [PMC free article] [PubMed] [Google Scholar]
- (2).Postler TS; Ghosh S Understanding the holobiont: how microbial metabolites affect human health and shape the immune system. Cell Metab 2017, 26 (1), 110–130. [DOI] [PMC free article] [PubMed] [Google Scholar]
- (3).Zeisel SH; Warrier M Trimethylamine N-oxide, the Microbiome, and Heart and Kidney Disease. Annu. Rev. Nutr 2017, 37,157–181. [DOI] [PubMed] [Google Scholar]
- (4).Krueger SK; Williams DE Mammalian flavin-containing monooxygenases: structure/function, genetic polymorphisms and role in drug metabolism. Pharmacol. Ther 2005, 106 (3), 357–387. [DOI] [PMC free article] [PubMed] [Google Scholar]
- (5).Bennett BJ; de Aguiar Vallim TQ; Wang Z; Shih DM; Meng Y; Gregory J; Allayee H; Lee R; Graham M; Crooke R; Edwards PA; Hazen SL; Lusis AJ Trimethylamine-N-oxide, a metabolite associated with atherosclerosis, exhibits complex genetic and dietary regulation. Cell Metab 2013, 17 (1), 49–60. [DOI] [PMC free article] [PubMed] [Google Scholar]
- (6).Dumas ME; Barton RH; Toye A; Cloarec O; Blancher C; Rothwell A; Fearnside J; Tatoud R; Blanc V; Lindon JC; Mitchell SC; Holmes E; McCarthy MI; Scott J; Gauguier D; Nicholson JK Metabolic profiling reveals a contribution of gut microbiota to fatty liver phenotype in insulin-resistant mice. Proc. Natl. Acad. Sci. U.S.A 2006, 103 (33), 12511–12516. [DOI] [PMC free article] [PubMed] [Google Scholar]
- (7).Spencer MD; Hamp TJ; Reid RW; Fischer LM; Zeisel SH; Fodor AA Association between composition of the human gastrointestinal microbiome and development of fatty liver with choline deficiency. Gastroenterology 2011, 140 (3), 976–986. [DOI] [PMC free article] [PubMed] [Google Scholar]
- (8).Wang Z; Klipfell E; Bennett BJ; Koeth R; Levison BS; DuGar B; Feldstein AE; Britt EB; Fu X; Chung Y-M; Wu Y; Schauer P; Smith JD; Allayee H; Tang WHW; DiDonato JA; Lusis AJ; Hazen SL Gut flora metabolism of phosphatidylcholine promotes cardiovascular disease. Nature 2011, 472 (7341), 57–63. [DOI] [PMC free article] [PubMed] [Google Scholar]
- (9).Koeth RA; Wang Z; Levison BS; Buffa JA; Org E; Sheehy BT; Britt EB; Fu X; Wu Y; Li L; Smith JD; Di-Donato JA; Chen J; Li H; Wu GD; Lewis JD; Warrier M; Brown JM; Krauss RM; Tang WHW; Bushman FD; Lusis AJ; Hazen SL Intestinal microbiota metabolism of L-carnitine, a nutrient in red meat, promotes atherosclerosis. Nat. Med 2013, 19 (5), 576–585. [DOI] [PMC free article] [PubMed] [Google Scholar]
- (10).Tang WHW; Wang Z; Levison BS; Koeth RA; Britt EB; Fu X; Wu Y; Hazen SL Intestinal microbial metabolism of phosphatidylcholine and cardiovascular risk. N. Engl. J. Med 2013, 368 (17), 1575–1584. [DOI] [PMC free article] [PubMed] [Google Scholar]
- (11).Gregory JC; Buffa JA; Org E; Wang Z; Levison BS; Zhu W; Wagner MA; Bennett BJ; Li L; DiDonato JA; Lusis AJ; Hazen SL Transmission of atherosclerosis susceptibility with gut microbial transplantation. J. Biol. Chem 2015, 290 (9), 5647–5660. [DOI] [PMC free article] [PubMed] [Google Scholar]
- (12).Tang WHW; Wang Z; Kennedy DJ; Wu Y; Buffa JA; Agatisa-Boyle B; Li XS; Levison BS; Hazen SL Gut microbiota-dependent trimethylamine N-oxide (TMAO) pathway contributes to both development of renal insufficiency and mortality risk in chronic kidney disease. Circ. Res 2015, 116 (3), 448–455. [DOI] [PMC free article] [PubMed] [Google Scholar]
- (13).Missailidis C; Hällqvist J; Qureshi AR; Barany P; Heimbürger O; Lindholm B; Stenvinkel P; Bergman P Serum trimethylamine-N-oxide is strongly related to renal function and predicts outcome in chronic kidney disease. PLoS One 2016, 11 (1), e0141738. [DOI] [PMC free article] [PubMed] [Google Scholar]
- (14).Miao J; Ling AV; Manthena PV; Gearing ME; Graham MJ; Crooke RM; Croce KJ; Esquejo RM; Clish CB; Morbid Obesity Study Group; Vicent, D.; Biddinger, S. B. Flavin-containing monooxygenase 3 as a potential player in diabetes-associated atherosclerosis. Nat. Commun 2015, 6, 6498. [DOI] [PMC free article] [PubMed] [Google Scholar]
- (15).Mitchell SC; Smith RL Trimethylaminuria: the fish malodor syndrome. Drug Metab. Dispos 2001, 29 (4), 517–521. [PubMed] [Google Scholar]
- (16).Dolphin CT; Janmohamed A; Smith RL; Shephard EA; Phillips IR Missense mutation in flavin-containing monooxygenase 3 gene, FMO3, underlies fish-odour syndrome. Nat. Genet 1997, 17 (4), 491–494. [DOI] [PubMed] [Google Scholar]
- (17).Christodoulou J Trimethylaminuria: an under‐recognised and socially debilitating metabolic disorder. J. Paediatr. Child Health 2012, 48 (3), E153–E155. [DOI] [PubMed] [Google Scholar]
- (18).Craciun S; Balskus EP Microbial conversion of choline to trimethylamine requires a glycyl radical enzyme. Proc. Natl. Acad. Sci. U.S.A 2012, 109 (52), 21307–21312. [DOI] [PMC free article] [PubMed] [Google Scholar]
- (19).Craciun S; Marks JA; Balskus EP Characterization of choline trimethylamine-lyase expands the chemistry of glycyl radical enzymes. ACS Chem. Biol 2014, 9 (7), 1408–1413. [DOI] [PubMed] [Google Scholar]
- (20).Bodea S; Funk MA; Balskus EP; Drennan CL Molecular basis of C-N bond cleavage by the glycyl radical enzyme choline trimethylamine-lyase. Cell Chem. Biol 2016, 23 (10), 1206–1216. [DOI] [PMC free article] [PubMed] [Google Scholar]
- (21).Wang Z; Roberts AB; Buffa JA; Levison BS; Zhu W; Org E; Gu X; Huang Y; Zamanian-Daryoush M; Culley MK; DiDonato AJ; Fu X; Hazen JE; Krajcik D; DiDonato JA; Lusis AJ; Hazen SL Non-lethal inhibition of gut microbial trimethylamine production for the treatment of atherosclerosis. Cell 2015, 163 (7), 1585–1595. [DOI] [PMC free article] [PubMed] [Google Scholar]
- (22).Romano KA; Martínez-del Campo A; Kasahara K; Chittim CL; Vivas EI; Amador-Noguez D; Balskus EP; Rey FE Metabolic, epigenetic, and transgenerational effects of gut bacterial choline consumption. Cell Host Microbe 2017, 22 (3), 279–290. [DOI] [PMC free article] [PubMed] [Google Scholar]
- (23).Martínez-del Campo A; Craciun S; Hamer HA; Haiser HJ; Turnbaugh PJ; Balskus EP Characterization and detection of a widely distributed gene cluster that predicts anaerobic choline utilization by human gut bacteria. mBio 2015, 6, e00042–15. [DOI] [PMC free article] [PubMed] [Google Scholar]
- (24).Chen K; Zheng X; Feng M;, Li D; Zhang H Gut microbiota-dependent metabolite trimethylamine N-oxide contributes to cardiac dysfunction in western diet-induced obese mice. Front. Physiol 2017, 8, 139. [DOI] [PMC free article] [PubMed] [Google Scholar]
- (25).Chilloux J; Brial F; Everard A; Smyth D; Zhang L; Plovier H; Myridakis A; Hoyles L; Fuchs JE; Blancher C; Gencer S; Martinez-Gili L; Fearnside JF; Barton RH; Neves AL; Rothwell AR; Gerard C; Calderari S; Boulange CL; Patel S; Scott J; Glen RC; Gooderham NJ; Nicholson JK; Gauguier D; Liu PP; Cani PD ; Dumas ME Microbiome inhibition of IRAK-4 by trimethylamine mediates metabolic and immune benefits in high-fat-diet-induced insulin resistance. bioRxiv 277434; doi: 10.1101/277434. [DOI]
- (26).Anthouard R; DiRita VJ Chemical biology applied to the study of bacterial pathogens. Infect. Immun 2015, 83 (2), 456–469. [DOI] [PMC free article] [PubMed] [Google Scholar]
- (27).Wallace BD; Wang H; Lane KT; Scott JE; Orans J; Koo JS; Venkatesh M; Jobin C; Yeh L-A; Mani S; Redinbo MR Alleviating cancer drug toxicity by inhibiting a bacterial enzyme. Science 2010, 330 (6005), 831–835. [DOI] [PMC free article] [PubMed] [Google Scholar]
- (28).While this manuscript was in revision, another paper reporting the development of inhibitors of anaerobic choline metabolism appeared: Wallace BD; Wang H; Lane KT; Scott JE; Orans J; Koo JS; Venkatesh M; Jobin C; Yeh L-A; Mani S; Redinbo MR Alleviating cancer drug toxicity by inhibiting a bacterial enzyme. Science 2010, 330 (6005), 831–835. Roberts, A. B.; Gu, X.; Buffa, J. A.; Hurd, A. G.; Wang, Z.; Zhu, W.; Gupta, N.; Skye, S. M.; Cody, D. B.; Levison, B. S.; Barrington, W. T.; Russell, M. W.; Reed, J. M.; Duzan, A.; Lang, J. M.; Fu, X,; Li, L.; Myers, A. J.; Rachakonda, S.; DiDonato, J. A.; Brown, J. M.; Gogonea, V.; Lusis, A. J.; Garcia-Garcia, J. C.; Hazen, S. L. Nat. Med. 2018, 24, 1407–1417. [DOI] [PMC free article] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.