

Abstract
Human-induced pluripotent stem cells (hiPSC) can be differentiated to cardiomyocytes at high efficiency and are increasingly used to study cardiac disease in a human context. This review evaluated 38 studies on hypertrophic (HCM) and dilated cardiomyopathy (DCM) of different genetic causes asking to which extent published data allow the definition of an in vitro HCM/DCM hiPSC-CM phenotype. The data are put in context with the prevailing hypotheses on HCM/DCM dysfunction and pathophysiology. Relatively consistent findings in HCM not reported in DCM were larger cell size (156 ± 85%, n = 15), more nuclear localization of nuclear factor of activated T cells (NFAT; 175 ± 65%, n = 3), and higher β-myosin heavy chain gene expression levels (500 ± 547%, n = 8) than respective controls. Conversely, DCM lines showed consistently less force development than controls (47 ± 23%, n = 9), while HCM forces scattered without clear trend. Both HCM and DCM lines often showed sarcomere disorganization, higher NPPA/NPPB expression levels, and arrhythmic beating behaviour. The data have to be taken with the caveat that reporting frequencies of the various parameters (e.g. cell size, NFAT expression) differ widely between HCM and DCM lines, in which data scatter is large and that only 9/38 studies used isogenic controls. Taken together, the current data provide interesting suggestions for disease-specific phenotypes in HCM/DCM hiPSC-CM but indicate that the field is still in its early days. Systematic, quantitative comparisons and robust, high content assays are warranted to advance the field.
Keywords: hiPSC, Disease modelling, Cardiomyopathy, Quantitative phenotypes
Introduction
The seminal discovery of means to reprogram human somatic cells into embryonic stem cell-like induced pluripotent stem cells (hiPSC; [89]) opened the possibility to generate patient- and disease-specific hiPSC lines and study disease mechanisms in an individualized and human context. An underlying assumption is that human diseases can be studied in hiPSC-derived differentiated cells cultured in vitro or, in other words, that such cells exhibit disease-specific phenotypes. Indeed, soon after the discovery by Yamanaka and colleagues, the first papers appeared that reported specific abnormalities in the function of patient-derived hiPSC derivatives compared to unrelated genetically normal controls. In the cardiac field, the first examples were longer action potentials in hiPSC-cardiomyocytes (hiPSC-CM) from patients with genetically determined long QT syndrome 1 (LQT1 [68] or LQT2 [40]) and larger cells with a higher degree of sarcomeric organization and preferential localization of NFATc4 in the nucleus in hiPSC-CM from a patient with Leopard syndrome [11]. In the meantime, most genetically determined cardiac diseases have been studied in hiPSC-CM and generally revealed some phenotypic abnormalities that have been described before in native cardiomyocytes from patients with the respective disease. However, it was soon realized that hiPSC-CM are relatively immature cells (for review, see [105]) that exhibit large phenotypic heterogeneity, e.g. in terms of action potential width and shape [68], cell size and sarcomeric organization. Reasons include variability of the original somatic cells used for reprogramming (e.g. skin cells with mosaic mutations or variable levels of epigenetic modifications [53], the reprogramming procedure itself [46], differentiation protocols with less than 100% efficiency, a varying level of maturity in hiPSC-CM in culture as well as methodological issues such as the difficulty to measure action potentials in small cells by patch clamping [38]). The recent introduction of transcription activator-like effector nuclease-mediated gene correction (TALEN) or CRIPSR/Cas9-based methods for gene editing has increased the level of trust in the conclusion that the observed phenotypes were indeed the consequence of the suspected gene mutation [4]. The reader is referred to several excellent reviews on this subject (e.g. [8, 70, 107]).
Hypertrophic and dilated cardiomyopathy—clinical phenotype and pathophysiology
This review will concentrate on the question to which extent a specific “cardiomyopathy phenotype” exists, which can be studied in hiPSC-CM in the dish. It restricts itself to hypertrophic cardiomyopathy (HCM) and dilated cardiomyopathy (DCM), because they are the two most common and clinically relevant cardiomyopathies, often have a defined genetic cause, have been most often studied in hiPSC-CM and present with relatively clearly defined and partially opposing clinical phenotypes (Table 1). The key morphological features of HCM are thickened left ventricular (LV) walls in the absence of apparent hemodynamic reason (e.g. aortic stenosis, severe hypertension). Hypertrophy preferentially affects the interventricular septum, whose thickness is commonly used as inclusion criterion for patients in clinical studies. HCM is generally associated with a normal or rather small LV cavity, preserved LV systolic contractile function and early diastolic dysfunction [37, 66, 102]. Most patients develop various degrees of LV obstruction [60]. Histomorphological signs of HCM are myocardial disarray and increased fibrosis. DCM in contrast is characterized by LV systolic dysfunction, dilation of LV cavities and normal wall thickness. While HCM is the prototypic genetic cardiomyopathy (likely disease-causing mutations can be found by cardiac gene panel, exome or whole genome sequencing in approximately 32–70% of cases [2, 16, 58, 78]), DCM is classified as a mixed cardiomyopathy, which is familial in ~ 20–35% [25, 60], and a recent whole exome sequencing identified mutations in only 12% of cases [58]. The majority of DCM cases are caused by (mainly viral or parasitic) infection, toxins such as alcohol or anti-tumour agents and mitochondrial disorders.
Table 1.
Clinical, morphological and functional characteristics of patients with hypertrophic (HCM) or dilated cardiomyopathy (DCM)
| HCM | DCM | |
|---|---|---|
| Symptoms and biomarkers | Arrhythmias and sudden cardiac death | Dyspnoe (initially exercise-induced) |
| Atrial fibrillation | Heart failure | |
| Exercise-induced dyspnoe | Arrhythmias and sudden cardiac death | |
| Heart failure | Atrial fibrillation | |
| Increased serum BNP levels | Increased serum BNP levels | |
| Morphology | LV hypertrophy ± outflow tract obstruction | LV chamber dilatation |
| Cardiac myocyte hypertrophy (width) | Cardiac myocyte hypertrophy (length) | |
| Myofiber/myocardial disarray | ||
| Fibrosis | Fibrosis | |
| Function | Diastolic dysfunction (pre-hypertrophy stage) | Systolic dysfunction |
| Hypercontractility (inconsistent) | Diastolic dysfunction | |
| Systolic dysfunction (late stage) | ||
| Energy depletion (early stage) | Energy depletion (early stage) |
Parameters distinguishing between HCM and DCM are marked in italics. Note overlap of many parameters
While the partially opposing clinical pictures of HCM and DCM allow a relatively straightforward clinical differentiation, overlaps between the two types of cardiomyopathies exist. Both HCM and DCM exhibit increased serum levels of brain natriuretic peptide and cardiac fibrosis, and HCM patients can develop severe systolic dysfunction requiring heart transplantation. Both can lead to life-threatening ventricular arrhythmias [60] and are accompanied by an increased rate of atrial fibrillation [81, 106] and dilation of the left atrium [36].
Despite the discovery of numerous mutations in genes that underlie HCM and DCM, our understanding of the pathomechanisms leading from the mutation to the phenotype remains incomplete. Reasons are not only the diversity of mutations causing similar clinical pictures, particularly in DCM, the incomplete and highly variable penetrance of both HCM and DCM, but also the fact that mouse models only partially recapitulate the human phenotype. For example, no single mouse model in which a classical HCM mutation in the gene coding for cardiac myosin-binding protein C (MYBPC3, cMyBPC) or β-myosin heavy chain (MYH7, β-MHC) has been introduced in the heterozygous state develop the pathognomonic septal hypertrophy seen in patients (for review, see [21]). Neither has LV obstruction been observed in any such model. Either homozygous knockout or knockin of the respective gene is lethal (as in the case of α-MHC, the rodent pendant of the dominant myosin isoform [28]) or the animals develop severe LV dysfunction (as in the case of cMyBPC [10, 27, 33, 61, 62, 64, 65, 98]). It is also apparent that the mouse work still did not answer a number of fundamental questions: (1) What is the exact physiological role of the sarcomeric proteins most commonly affected in HCM such as cMyBPC and β-MHC? (2) How do they cooperate to ensure proper systolic and diastolic function? (3) What are the specific consequences of even relatively well-studied gene mutations? (4) How do mutations in numerous sarcomeric and non-sarcomeric genes with diverse function lead to the uniform induction of “autonomous” cardiac hypertrophy and disarray in HCM? These questions have been discussed in recent reviews to which the reader is referred [21, 26, 59, 94]. In any case, the experiences with mouse models thus raise the question to which extent they really reflect the human disease and provide an argument to study HCM and DCM in hiPSC-CM.
Another reason for our limited understanding of HCM/DCM pathophysiology is that access to isolated heart tissue and cells from patients with HCM and DCM is sparse, and only very few studies specifically reported on the in vitro phenotype of these diseases (for review, see [22]). The most commonly used sources for human tissues are septum biopsies acquired during surgical correction of LV outflow tract obstruction by myectomy in the case of HCM and LV tissues obtained during implantation of LV assist devices or heart transplantation in case of DCM. Both tissue sources represent a late stage of the disease, raising the question to which extent the abnormalities observed in comparison to (even rarer) non-failing heart tissue comparators reflect primary defects or secondary compensations or consequences. The highly fibrotic texture of the terminally diseased tissue imposes a further challenge to such studies as enzymatic isolation of cardiomyocytes requires harsher conditions, introducing a systematic error.
Prevailing in vitro phenotypes of HCM and DCM
Despite the limitations discussed above, some observations prevail and have led to hypotheses that can be tested in hiPSC-CM.
Abnormal myofilament calcium sensitivity
The relation between intracellular Ca2+ concentrations and force development of the myofilaments is a highly regulated biological constant with half-maximal force development (of skinned myofibers) at a pCa of ~ 5.8 (~ 1.6 μM). Numerous studies reported increased Ca2+ sensitivity on HCM [3, 13, 19, 23, 67, 95, 96] and decreased in DCM [19, 20, 56]. The shift in the pCa/force relation leads to more force development at lower Ca2+ concentrations in case of HCM and less force development in DCM. Importantly, the increased Ca2+ sensitivity in HCM also predicts delayed relaxation in the descending part of the intracellular Ca2+ transient. Both consequences are well compatible with the predominant clinical phenotypes of preserved systolic function and diastolic dysfunction in HCM and systolic dysfunction in DCM. Increased Ca2+ sensitivity in HCM would even predict LV hypercontractility at rest, and indeed, a study in 36 mutation carriers found significantly increased LV ejection fraction by echocardiography compared to 36 healthy controls [36]. This observation forms the basis of novel therapeutic concepts to reduce myosin activity by small molecules to treat HCM [30]. However, it is also clear that increased Ca2+ sensitivity in HCM is not a universal finding. Several studies reported HCM mutations to be associated with either no change [33, 101] or decreased Ca2+ sensitivity [88] in various experimental systems, suggesting mutation-specific differences. Of note, even the knockout of a protein such as cMyBPC [13, 33, 48] has been associated with different effects on myofilament Ca2+ sensitivity. The latter emphasizes the importance of the respective experimental context and supports the notion that altered myofilament Ca2+ sensitivity cannot be the sole unifying mechanism underlying HCM or DCM.
Abnormal actin-myosin sliding velocity
Another parameter of sarcomere function is the unloaded sliding velocity of thin filaments on immobilized S1-myosin. Several studies indicate that HCM mutations are associated with increased sliding velocity [44, 45, 88] and DCM with decreased sliding velocity [1, 79], and for a review, see [24].
Altered maximal force development
Interestingly, both HCM and DCM mutations were found to generally associate with decreased maximal force development [95, 101], but normal or even higher force output and increased force redevelopment have also been reported [48, 92, 102].
Increased Ca2+-independent cross-bridge cycling in HCM
Mutations (or full deletion) of MYBPC3 or cardiac troponin T (TNNT2, TnT) have been associated with a shallow pCa-force relationship (lower Hill coefficient) and residual force development at very low or nominal absence of Ca2+ [3, 56, 75]. In the case of cMyBPC, the effect may be explained by mutations (or its absence) disturbing its normal role in stabilizing the super-relaxed, inactive state (SRX) of myosin heads [63]. The concept implies that one of the abnormalities in HCM is incomplete arrest of crossbridge cycling in diastole, which could participate in diastolic dysfunction and increased energy expenditure.
Decreased energetic efficiency
Many HCM mutations lead to decreased energetic efficiency of crossbridge cycling, i.e. inefficient usage of ATP to fuel contraction [14, 45, 67, 102]. The phenomenon in a general sense indicates less-than-normal functioning of the sarcomeres harbouring mutated sarcomere proteins and may relate to the partial loss of the myosin SRX state in the case of cMyBPC. In any case, it may well contribute to the decreased phosphocreatine/ATP ratio observed in patients with HCM even in the prehypertrophic state [17]. Energy starvation is not specific to HCM. In fact, it has been shown already in 1992 as a common feature of patients with heart failure due to non-ischemic DCM [71]. However, it is possible that the more diverse causes of DCM include both decreased energetic efficiency of the myofilaments with higher energy expenditure and decreased efficiency of mitochondrial energy generation like in Barth syndrome [39].
Allelic imbalance of β-MHC as a cause of myocardial disarray
Early work (in skeletal muscle fibres from β-MHC-expressing soleus muscle) showed a high variability of myofilament Ca2+ sensitivity between individual muscle fibres [47]. This observation was later related to marked cell-to-cell differences in the expression of the mutated β-MHC in cardiomyocytes and marked differences in individual Ca2+ sensitivity [49]. The interesting phenomenon could well contribute to another hallmark of HCM, myocardial disarray, by individual cardiomyocyte developing different degrees of contractile force. It is not clear whether allelic imbalance is restricted to β-MHC mutations.
HCM and DCM phenotypes in hiPSC-cardiomyocytes
By searching PubMed (keywords: hiPSC cardiomyocytes and hypertrophic cardiomyopathy or dilated cardiomyopathy), we identified 38 original papers reporting phenotypes of hiPSC-CM either derived from hiPSC lines of patients with HCM/DCM (or related syndromes) or from hiPSC lines in which a HCM or DCM mutation had been genetically introduced (Tables 2 and 3). Initial studies compared the phenotype of disease-related hiPSC-CM to unrelated genetically healthy controls; more recent studies used TALEN or CRISPR/Cas9 gene editing approaches to correct a mutation in a patient-specific line or introduce it into a wild-type line, allowing comparison under isogenic conditions. While most studies validated the absence of off-target effects only at the predicted top-10 sites, one TALEN-based study performed whole exome sequencing and reported in two corrected clones 318 and 1331 de-novo indel mutations, respectively, close to possible off-target sites. The significance of this finding is unclear.
Table 2.
Studies reporting cardiomyocyte phenotypes in hiPSC-CM from patients with genetically determined HCM or syndromes associated with HCM phenotypes or from hiPSC lines in which mutations had been introduced
| Mutation | Disease | Peak force | T1 | T2 | Cell size | Disarray | Other phenotypes | CRISPR/TALEN Ctr. | Karyotype Ctr. | Reference |
|---|---|---|---|---|---|---|---|---|---|---|
|
MYH7
Het p.Arg663His |
HCM | n.d. | n.d. | n.d. | + 60% | n.d. | Multinucleation 50 vs. 20%, mRNA of NPPA, NPPB, MYH7 up, higher nuclear NFAT, Ca2+-induced arrhythmias, DAD, higher basal [Ca2+], Iso-induced DAD | No | No | [50] |
|
MYBPC3
3 Pts, 3 Ctr p.Gly999-Gln1004del, 2 w/o identified mut |
HCM | n.d. | n.d. | n.d. | + 20% | + 50–100% | NPPA, TNNT2 up, MYBPC3–20% in mut, stronger hypertrophic, disarray and NFAT response to ET1, contractile abnormalities linked to disarray | No | No | [90] |
|
MYBPC3
Exon 25, 3 pts. |
HCM | n.d. | n.d. | n.d. | + 50–100% | n.d. | No further hypertrophic response to stimuli | No | Yes | [18] |
|
MYH7
Het p.Arg442Gly |
HCM | n.d. | n.d. | n.d. | + 15% | + 200% | Higher nuclear NFAT; arrhythmias + 300%, APD prolongation + 60%, resting [Ca2+] up 20%, ICa and INa up | No | Yes | [32] |
|
MYBPC3
Het c.2373dupG |
HCM | − 50% | n.d. | n.d. | +/− | n.d. | cMyBPC haploinsufficiency | No | No | [7] |
|
GAA
Hom del in exon 18 and CpHet c.1441delT/c.2237G>A 2 lines each |
Pompe | +/− or − 60% | +/− | +/− | n.d. | n.d. | Glycogen accumulation, glycan processing abnormality, but normal autophagic flux | No | Yes | [77] |
| GAA CpHet c.796C>T/ c.1316T>A, 3 clones from pt., 1 clone from ctr |
Pompe | n.d. | n.d. | n.d. | no | n.d. | Glycogen accumulation, no functional data | No | No | [82] |
|
ALPK3
Hom p.W1264X |
DCM/HCM | n.d. | n.d. | n.d. | n.d. | + 230% | Irregular Ca2+ transients + 400%, MEA FP + 100% | No | Yes | [73] |
|
BRAF
Het p.Thr599Arg |
HCM Syndromic |
0 to + 40% | − 30% | − 30% | n.d. | n.d. | Less negative FFR, increased Iso-sensitivity, mRNA of NPPA + 300% and SERCA2a + 40% (ns) | No | No | [12] |
|
BRAF
Het p.Thr599Arg Het p.Gln257Arg |
HCM Syndromic | n.d. | n.d. | n.d. | + 300% | + 260% | mRNA of NPPA, NPPB, MYH7 up, PLN down, higher Ca2+ transients and store, calcium arrhythmias, “fibroblast” profibrotic phenotype | No | Yes | [41] |
|
FXN
GAA triplet repeat |
Friedreich’s ataxia | n.d. | n.d. | n.d. | n.d. | n.d. | ROS, unusual iron responses | No | No | [51] |
|
MYBPC3
Het p.Gln1061X (n = 2) or TPM1 Het p.Asp175Asn (n = 2) |
HCM | n.d. | n.d. | n.d. | + 200% (M), not clear in T | n.d. | More multinucleation (40 vs 20%), Ca2+ arrhythmias (T, not M), more DAD in M, not T, APD high in T, RMP − 75, NPPA +/−, NPPB, MYH7 and many others up, more in M than T, cMyBPC +/−, TPM up in T | No | Yes | [72] |
|
MYH7
Het p.Glu848Gly |
HCM | − 54% | n.d. | n.d. | n.d. | Yes, not quantified |
Skinned myofiber from hiPSC-CM: Fmax 8.2 vs. 18.6 mN/mm2 (adult 110), KAct + 62%, increased Ca2+ sensitivity | No | No | [74] |
|
MYH7
Het p.Val698Ala |
HCM | n.d. | n.d. | n.d. | n.d. | n.d. | n.d. | N.d. | Yes | [80] |
|
GLA
Hemizygote c.919+4G >A |
Fabry | n.d. | n.d. | n.d. | n.d. | n.d. | Gal act. down, GB3 accumulation, low beating rate, arrhythmias | No | [15] | |
|
LAMP2
Het c.129– 130insAT) Het c.64+1G>A |
Danon | n.d. | n.d. | n.d. | n.d. | n.d. | Mitochondrial abnormalities, decreased autophagic flux | No | Yes | [34] |
|
MYBPC3
Het c.1358_1359insC |
HCM | n.d. | n.d. | n.d. | + 65% | n.d. | cMyBPC haploinsufficiency, BNP, MYH7 and others up | Corrected by gene therapy | No | [76] |
|
PRKAG2
Het Arg302Gln |
HCM + WPW | n.d. | n.d. | n.d. | + 10–30% | n.d. | MDP, APD +/−, If +/−, AP irregularity, RR scatter + 500% | Yes | Yes | [5] |
|
SCO2
Hom c.577G>A CpHet c.418G>A/c.17Ins19 |
HCM syndrome | +/− (??) | n.d. | n.d. | n.d. | n.d. | Mitochondrial abnormalities, no Iso or Ca2+ response, DAD, arrhythmic response to Iso | No | Yes | [31] |
|
MT-RNR2
m.2336T>C |
Mitochondrial HCM | n.d. | n.d. | n.d. | + 30% | n.d. | NPPA, NPPB, NFAT up, slightly increased intracellular calcium, SR store, reduced ICa, APD prolonged, arrhythmias, RMP − 55, upstroke 5–10 v/s, DAD | No | Yes | [52] |
|
MYL3
Het c.170C-A, Exac 0.0001154, introduced 170C-g and MYBPC3 Het p.Val321Met |
HCM-associated VUS | n.d. | n.d. | n.d. | +/− (also in mut) | n.d. | No phenotype detected in VUS, mean cell size 1800 μm2, NPPA and MYH7 up in the two diseased, contraction and rel velocity slightly up, arrhythmias, good stats | Yes | Yes | [57] |
|
TNNT2
Het p.Ile79Asn |
HCM | + 75% | n.d. | + 40% | +/− | yes | Sarcomere length +/− (1.8 μm), smaller caffeine transient, higher Ca2+ buffering, shorter APD, Ca2+ beat to beat instability, triangulation, NCX-sensitive | Yes | Yes | [100] |
| MYH7 and MYH6; Het/Hom p.Arg453Cys, frameshift KO, +MYH6 frameshift | HCM | − 20% (het), − 70% (hom) − 80% (KO) | + 20% | +/− or + 10% (+MYH6 fs) | + 50% | yes | NPPB up, multinucleation, basal and max. respiration up, ATP production up, Ca2+ transient irregularities, nifedipine- and ranolazine-sensitive; MYH7/MYH6 ratio up | Yes | Yes | [69] |
ANP/BNP atrial/brain natriuretic peptides, AP action potential, APD action potential duration, ALPK3 α-kinase 3, BRAF B-Raf proto-oncogene, serine/threonine kinase, cMyBPC cardiac myosin-binding protein C, CpHet compound heterozygous, CRISPR clustered regularly interspaced short palindromic repeats/Cas9 gene correction, Ctr control, DAD delayed after depolarizations, Del deletion, Disarray abnormal sarcomeric organization, ET1 endothelin 1, FFR force-frequency relation, Fmax maximal force development, FXN frataxin gene, GAA α-glycosidase gene, GLA α-galactosidase A, GB3 glycosphingolipids, Het heterozygous, Hom homozygous, ICa L-type Ca2+ current, INa Na+ current, Iso isoprenaline, KAct rate constant reflecting crossbridge turnover rate, LAMP2 lysosome-associated membrane protein 2 gene, MEA FP multielectrode array field potentials, MT-RNR2 mitochondrially encoded 16S RNA gene, Mut mutation, MYBPC3 cardiac myosin-binding protein C gene/mRNA, MYL3 myosin light chain 3 (MLC1v) gene, MYH6/MYH7 gene or mRNA of α-/β-myosin heavy chain, NCX sodium-calcium exchanger, NFAT nuclear factor of activated T cells, NPPA atrial natriuretic peptide mRNA, NPPB brain natriuretic peptide mRNA, PLN phospholamban gene/mRNA, Pt(s) patient(s), RMP resting membrane potential, ROS reactive oxygen species, RR scatter beat-to-beat irregularity, SERCA2a sarcoplasmic reticulum ATPase, SCO2 cytochrome c oxidase assembly protein gene, T1 time to peak force, T2 time from peak to relaxation, TALEN transcription activator-like effector nuclease-mediated gene correction, TNNT2 cardiac troponin T gene, TnT cardiac troponin T, TPM1 α-tropomyosin, VUS variant of unknown significance
Table 3.
Studies reporting cardiomyocyte phenotypes in hiPSC-CM from patients with genetically determined DCM or syndromes associated with DCM phenotypes or from hiPSC lines in which mutations had been introduced
| Mutation | Disease | Peak force | T1 | T2 | Cell size | Disarray | Other phenotypes | CRISPR/TALEN Ctr. | Karyotype Ctr. | Reference |
|---|---|---|---|---|---|---|---|---|---|---|
|
TNNT2
Het p.Arg173Trp |
DCM | − 80% (AFM) | n.d. | n.d. | +/− | Yes | Desensitized NE response of rate, RMP − 40 mV, APD +/−, smaller Ca2+ and caffeine transient (~2 s), TTP and TTD caffeine + 100% (WT 0.5/1.2 s), rescue by SERCA OE, metoprolol rescues disorganization | No | Yes | [87] |
|
LMNA
Het p.Arg225X (n = 3), another fs mut |
DCM | n.d. | n.d. | n.d. | n.d. | n.d. | Nuclear abnormalities, apoptosis,MEK-inhibition-sensitive, ERK up | No | Yes | [84] |
|
DES
Het p.Ala285Val, + 43 stop/gain mut |
DCM | n.d. | n.d. | n.d. | n.d. | n.d. | Morphological abnormalities + 700%, aggregates, peak Ca2+ transient +/−, − dF/dt − 40%, abnormal Iso response | No | No | [91] |
|
TAZ
Het c.517delG Het c.328T>C |
Barth | − 50–70% | n.d. | n.d. | n.d. | Yes | Decreased mito ox rate, immature cardiolipin, reduced ATP content, excess ROS, tafazzin-sensitive, MitoTempo-sensitive | Yes | Yes | [99] |
|
TNNT2
p.Arg173Trp |
DCM | − 60% (TFM) | n.d. | n.d. | n.d. | n.d. | ISO response down, TTP/TTD +/− (~1 s), cAMP response to Iso − 50%, rate − 60%, PDE2,3,5 mRNA several folds up, rescue of cAMP and force by FSK + IBMX | No | No | [103] |
|
TTN
p.Ser14450fsX4 |
DCM | n.d. | n.d. | n.d. | n.d. | Yes | MYH6, MYH7, ACTC1–50%, rescue by exon skipping | No | No | [29] |
|
DMD
Dp427m |
DMD | n.d. | n.d. | n.d. | n.d. | n.d. | More apoptosis, almost complete lack of MYL2, MYL3 and TPM1 mRNA, ICa − 40%, resting [Ca2+] + 40%, beneficial effects by Poloxamer 188 | No | [54] | |
|
TTN
3 diff. Truncating mut, CRISPR induction |
DCM | − 60–80% (EHT) | n.d. | n.d. | n.d. | + 400% | Stronger phenotype on stiffer posts, rate − 50%, lower SL (1.6 vs. 1,75 μm), lower MYH7/MYH6 ratio, beneficial effect of VEGF | No | No | [35] |
|
PLN
Het p.Arg14del (R14del) |
DCM | − 60–90% | n.d. | up | n.d. | n.d. | Resting [Ca2+] + 30%, irregular Ca2+ transient + 500%, caffeine transient + 75%, RMP − 49 vs. − 58 mV, NPPA/NPPB + 300–900%, MYH7/MYH6 + 60% | Off-target effects | Yes | [43] |
|
RBM20
p.Arg636Ser 2 clones each WT/mut |
DCM | n.d. | n.d. | n.d. | n.d. | Yes | Increased sarcomere length (WT 1.4 μm), increased Ca2+ peak, AUC, decay time at much lower rate | Yes | [104] | |
|
TNNT2
p.Arg173Trp 2 clones each |
DCM | (−) Inconsistent | n.d. | n.d. | n.d. | Yes | Lower rate of cells beat, myosin ATPase − 20%, beneficial effects of omecamtiv | No | [9] | |
|
RBM20
p.Ser635Ala 2 clones each |
DCM | − 40% (EHT) | +/− (100 μm2) | Yes | Actinin periodicity − 80%, lower resting [Ca2+], peak [Ca2+] + 50%, TTP/TTD + 60/150%, duration + 150%, all at 70% lower rate, normal Iso force response, lower length-stress response, TTN + MYH7 exon exclusion | No | [86] | |||
|
BAG3
KO induced in WT, 2 lines MYBPC3 KO |
DCM | − 50% (MTF) | n.d. | n.d. | n.d. | Yes | Stronger disarray increased by bortezomib, normal bortezomib-response in MYBPC3-KO | Yes | Yes | [42] |
|
DMD
3 different mut |
DMD | − 30–70% (EHT) | n.d. | n.d. | n.d. | n.d. | Correction of phenotype by CRISPR | Yes | No | [55] |
|
DMPK
CTG repeats in, 4 × 6 clones analysed |
DM1 | Rundown | n.d. | n.d. | n.d. | n.d. | Nuclear RNA CUG foci, nuclear size + 30%, irregularity, MDP − 52 vs. − 60 mV, APD − 20%, AP amplitude − 20%, upstroke velocity down, force rundown (AFM); altered splicing of MBNL1, MBNL2, TNNT2, SCN5A; fetal SCN5A isoforms, α-MHC and TnT protein down | No | Yes | [85] |
ACTC1 α-cardiac actin gene, α-MHC α-myosin heavy chain, AFM atomic force microscopy, ANP/BNP atrial/brain natriuretic peptide, AP action potential, APD action potential duration, AUC area under the curve, BAG3 BCL2-associated athanogene 3, cAMP cyclic adenosine monophosphate, CRISPR clustered regularly interspaced short palindromic repeats/Cas 9 gene correction, DES desmin gene, Disarray abnormal sarcomeric organization, DM1 myotonic dystrophy type 1, DMD Duchenne muscular dystrophy, DMD dystrophin gene, DMPK dystrophia myotonica protein kinase, EHT engineered heart tissue, ERK extracellular signal-regulated kinase, an important MAPK, Fs frameshift, FSK forskolin, IBMX isobutylmethylxanthine, ICa L-type Ca2+ current, Iso isoprenaline, KO knockout, LMNA lamin A/C gene, MBNL muscle blind-like, MDP maximal diastolic potential, MEK mitogen-activated protein (MAPK) kinase kinase, Mut mutation, MYBPC3 cardiac myosin-binding protein C (cMyBPC) gene, MYH6/MYH7 mRNA of α-/β-myosin heavy chain, MYL2 regulatory myosin light chain gene, ventricular isoform (MLC2v), MYL3 essential myosin light chain gene (MLC1v), NPPA atrial natriuretic peptide gene/mRNA, NPPB brain natriuretic peptide gene/mRNA, NE norepinephrine, PDE phosphodiesterase, PLN phospholamban gene, RBM20 RNA binding motif protein 20 gene, RMP resting membrane potential, ROS reactive oxygen species, SCN5A sodium voltage-gated channel alpha subunit 5 (Nav1.5) gene, SERCA OE sarcoplasmic reticulum ATPase overexpression, SL sarcomere length, T1 time to peak force, T2 time from peak to relaxation, TALEN transcription activator-like effector nuclease-mediated gene correction, TAZ tafazzin gene, TFM traction force microscopy, TNNT2 cardiac troponin T gene/mRNA, TnT cardiac troponin T, TPM1 tropomyosin gene, TTD time-to-decay, TTN titin gene, TTP time-to-peak, VEGF vascular endothelial growth factor, WT wild type
The initial analysis of the papers concentrated on abnormalities in contractile function, based on the hypothesis formulated by Davis and Molkentin that differences between HCM, DCM and wild type (WT) should primarily result in a different tension-time integral of the contraction peak, i.e. the area under the curve of an averaged contraction peak [19]. The hypothesis corroborates the idea that HCM mutations lead to increased, DCM mutations to decreased myofilament Ca2+ sensitivity. As elegantly shown in mouse models with different cardiac troponin C mutations (and in examples of hiPSC-CM), this should lead to higher peak force and prolonged relaxation (T2) in case of HCM and lower peak force and an abbreviated contraction peak (both contraction [T1] and relaxation time [T2]). Unfortunately, contraction kinetics were only studied in a small minority of cases (Tables 2 and 3). Only two studies of a DCM mutation (heterozygous phospholamban (PLN) R14del and truncating titin (TTN) mutation) showed a representative contraction peak, which indicated prolonged relaxation in one case [43] and lower T1 and T2 in the other [35]. Statistics were not provided. Two papers on HCM mutations (TNNT2, MYH7) reported statistically evaluated data on T1 and T2, showing no alteration or the expected increase in time of relaxation [69, 100]. Thus, clearly, more work has to be done to decide whether or not HCM/DCM mutations have a systematic effect on contractile kinetics in hiPSC-CM.
Many studies reported peak force, size of intracellular Ca2+ transients, sarcomere structure and gene expression (Tables 2 and 3, Fig. 1). Interestingly, while almost all studies on HCM lines reported cell sizes in 2D culture, only two did in case of DCM lines (Fig. 1). Similar differences in reporting frequency were observed with regard to multinucleation, nuclear NFAT, contraction kinetics (only HCM), ANP/BNP (NPPA/NPPB) and rhythmicity (more in HCM) or Ca2+ transient kinetics (only DCM). Reasons are unknown, but a reporting bias appears likely.
Fig. 1.
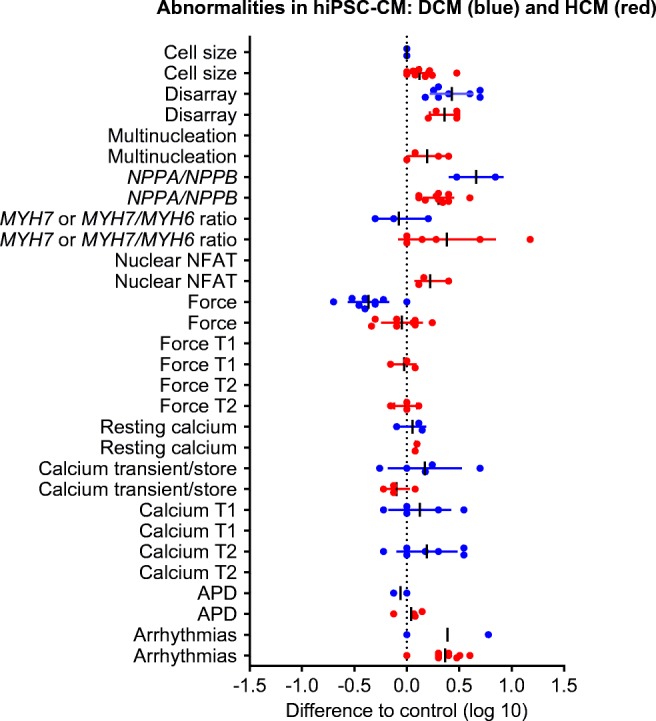
Published abnormalities of structure, gene expression or function of HCM/DCM-derived hiPSC-CM (HCM in red, DCM in blue). The data were extracted from the studies summarized in the Tables and are expressed as fold (log scale) of the control used in the respective study (either healthy control or gene edited isogenic line). More detail is provided in Tables 1 and 2. Each dot indicates one study. Lack of dots for certain parameters (e.g. force T1/T2 for DCM) indicates that none of the studies has reported these parameters. Abbreviations used: NPPA/NPPB atrial/brain natriuretic peptide (mRNA or protein concentration/positivity), MYH7/MYH6 β/α-myosin heavy chain gene expression, T1 time to peak of contraction or (calcium T1) of calcium transient peak, T2 time from contraction peak or (calcium T2) of calcium transient peak to relaxation/decay, APD action potential duration. The line calcium transient/store combines data on the peak calcium transient under baseline or caffeine-induced conditions
Figure 1 summarizes the data from all studies in which functional data were reported in a quantitative manner and presents them compared to the respective controls (log scale; n = 16 HCM, 14 DCM). Three abnormalities appeared to be relatively consistent in both HCM and DCM—sarcomeric disarray (274 ± 81%, n = 6 HCM; 298 ± 146%, n = 8 DCM), increased NPPA or NPPB gene expression (284 ± 249%, n = 11 HCM; 500%, n = 2) and arrhythmic behaviour (327 ± 164%, n = 12 HCM; 350%, n = 2 DCM). HCM lines showed an increase in cell size (156 ± 85%, n = 15; DCM +/−), in MYH7 gene expression (or the ratio of MYH7/MYH6 (500 ± 547%, n = 8; DCM +/− or reduction) and nuclear accumulation of the transcription factor NFAT (175 ± 65%, n = 3; DCM not determined). The most consistent abnormality in DCM lines was lower peak force development compared to the respective control (47 ± 23%, n = 9; HCM +/− with variability).
Besides the reported disease-associated abnormalities in function, structure or gene expression, it is apparent that absolute values varied largely. For example, reported cell surface area in 2D ranged from 100 μm2 [86] to > 2000 [57, 76], with reported cell volumes from 5.8 [100] to 120 μm3 [69]. Both volume data appear extremely low compared to the 95 μm3 in erythrocytes (mean corpuscular volume; Wikipedia). Besides differences in methods (e.g. time of culture in 2D, surface patterning), issues with the imaging technique and calculations may explain the scatter. In any case, hiPSC-CM are largely smaller than their native adult counterparts for which volumes of 15,000–40,000 μm3 have been reported [6]. It is not quite clear why size comparisons by patch clamp (membrane capacitance) indicate much smaller differences between hiPSC-CM and native human atrial or ventricular cardiomyocytes (e.g. 31–47 pF in hiPSC-CM compared to 74/89 pF in right atrial/LV myocytes [38]). The capacitance data are consistent across different studies (e.g. 60 pF in hiPSC-CM [93], 27 pF in hiPSC-CM [52, 54], ~ 60 pF in human atrial cardiomyocytes [97]). Possibly, the ratio between membrane capacitance and cell volume, which varies between species and the developmental stage (pF/pl = 4–9 [83]), is unusually high in hiPSC-CM. Action potential duration (APD90) at 37 °C varied from 240 ms [5] to 710 ms [43]. Again, it is likely that not only biological differences between hiPSC lines and influences of cell culture conditions and CM maturity but also technical issues explain the large variation. We have shown recently that the sharp microelectrode technique provides more reliable action potential data than patch clamping of single cells [38]. In this study, patch clamp-recorded APD90 in isolated hiPSC-CM amounted only to 119 ± 17 ms (human atrial cardiomyocytes 220 ms, human LV cardiomyocytes 434 ms), while those in intact hiPSC-CM or 3D engineered heart tissue were 271 ms (human right atrial tissue 317 ms, LV tissue 334 ms).
Conclusion
The present overview on published reports on the phenotype of HCM/DCM-derived hiPSC-CM allows some preliminary conclusions. (1) The most consistent and to a certain degree differentiating phenotype of hiPSC-CM appears to be decreased force production in DCM, correlating well with the dominant clinical presentation of the disease. (2) HCM lines appear not to exhibit consistent alterations in force development but show increased CM size, nuclear NFAT and increased MYH7 or MYH7/MYH6 ratio. Given the paucity of measurements of these parameters in DCM, it is not possible at present to decide whether these parameters allow a distinction between HCM/DCM phenotypes. (3) Sarcomere disorganization is a common finding in all disease lines and does not appear to allow differentiation between the clinically opposing phenotypes. (4) Overall, the analysis indicates that hiPSC-based disease modelling of cardiomyopathies is still in its early days. Suggestions for a basal set of parameters to be analysed in future studies are given in Table 4. More statistical rigor and robust high content methods are necessary to uncover potentially meaningful but discrete abnormalities of cardiac function in these cells. In this respect, it is interesting to note that only one study evaluated myofilament Ca2+ sensitivity in skinned fibres [74], yet myofilament Ca2+ sensitivity is one of the most commonly studied parameters in HCM/DCM-related human or animal specimens.
Table 4.
Suggestions for a basal set of parameters to be analyzed and reported in hiPSC-CM studies
| Parameter | Comment |
|---|---|
| Karyotype | Karyotype problems are frequent and increase with passage number. Karyotype checks in iPSC should be done < 5–10 passages before analysis. |
| Cardiomyocyte yield | The percent of TnT- or actinin-positive cells (e.g. by FACS) per batch evaluated should be presented as mean ± SD. |
| Number of batches | The number of cells/derivatives (n = x) and the number of differentiation runs the cells were derived from (N = y) in a given experiment need to be reported. |
| Blinding procedures | Given the variability of cells and readouts, procedures should be established and described that allow investigator-blinded assessments. |
| Age of cardiomyocytes | Many parameters change over time of culture in 2D or 3D, therefore the age of cells at time of analysis should be presented (mean ± SD). |
| Expression of disease gene alleles | In case of defined mutations, the relative expression of mutant and wild-type alleles should be determined to get an idea of mechanism. |
| Gene expression | Transcript levels should be reported as a set of standard genes, not only selected examples. |
| Indicators of cardiomyocyte maturity | Absolute transcript levels of α-/β-MHC (+their ratio) in comparison with human heart levels give a good initial indication of maturity. |
| Cell size | High n-numbers and information on cell density are mandatory. Volume data (e.g. FACS) may be more informative and precise than surface measurements in 2D. |
| Force and force kinetics | Given the strong dependence of force and force kinetics on beating rate, temperature and pH, these parameters need to be controlled (e.g. by electrical pacing) and reported. |
Acknowledgements
The work of the authors is supported by the DZHK (German Centre for Cardiovascular Research), the German Ministry of Research Education (BMBF), the German Research Foundation (DFG) and the European Research Council (ERC AG IndivuHeart).
Compliance with ethical standards
Conflict of interest
T.E. is co-founder of EHT Technologies GmbH, a university spin-off providing equipment for EHT generation and analysis.
Footnotes
This article is part of the special issue on Sarcomeric Mutations in Pflügers Archiv – European Journal of Physiology
Contributor Information
Thomas Eschenhagen, Phone: +49-40-7410-52180, Email: t.eschenhagen@uke.de.
Lucie Carrier, Phone: +49-40-7410-57208, Email: l.carrier@uke.de.
References
- 1.Aksel T, Choe Yu E, Sutton S, Ruppel KM, Spudich JA. Ensemble force changes that result from human cardiac myosin mutations and a small-molecule effector. Cell Rep. 2015;11:910–920. doi: 10.1016/j.celrep.2015.04.006. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 2.Alfares AA, Kelly MA, McDermott G, Funke BH, Lebo MS, Baxter SB, Shen J, McLaughlin HM, Clark EH, Babb LJ, Cox SW, DePalma SR, Ho CY, Seidman JG, Seidman CE, Rehm HL. Results of clinical genetic testing of 2,912 probands with hypertrophic cardiomyopathy: expanded panels offer limited additional sensitivity. Genet Med. 2015;17:880–888. doi: 10.1038/gim.2014.205. [DOI] [PubMed] [Google Scholar]
- 3.Baudenbacher F, Schober T, Pinto JR, Sidorov VY, Hilliard F, Solaro RJ, Potter JD, Knollmann BC. Myofilament Ca2+ sensitization causes susceptibility to cardiac arrhythmia in mice. J Clin Invest. 2008;118:3893–3903. doi: 10.1172/JCI36642. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 4.Bellin M, Casini S, Davis RP, D’Aniello C, Haas J, Ward-van Oostwaard D, Tertoolen LG, Jung CB, Elliott DA, Welling A, Laugwitz KL, Moretti A, Mummery CL. Isogenic human pluripotent stem cell pairs reveal the role of a KCNH2 mutation in long-QT syndrome. EMBO J. 2013;32:3161–3175. doi: 10.1038/emboj.2013.240. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 5.Ben Jehuda R, Eisen B, Shemer Y, Mekies LN, Szantai A, Reiter I, Cui H, Guan K, Haron-Khun S, Freimark D, Sperling SR, Gherghiceanu M, Arad M, Binah O. CRISPR correction of the PRKAG2 gene mutation in the patient's induced pluripotent stem cell-derived cardiomyocytes eliminates electrophysiological and structural abnormalities. Heart Rhythm. 2018;15:267–276. doi: 10.1016/j.hrthm.2017.09.024. [DOI] [PubMed] [Google Scholar]
- 6.Bensley JG, De Matteo R, Harding R, Black MJ. Three-dimensional direct measurement of cardiomyocyte volume, nuclearity, and ploidy in thick histological sections. Sci Rep. 2016;6:23756. doi: 10.1038/srep23756. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 7.Birket MJ, Ribeiro MC, Kosmidis G, Ward D, Leitoguinho AR, van de Pol V, Dambrot C, Devalla HD, Davis RP, Mastroberardino PG, Atsma DE, Passier R, Mummery CL. Contractile defect caused by mutation in MYBPC3 revealed under conditions optimized for human PSC-cardiomyocyte function. Cell Rep. 2015;13:733–745. doi: 10.1016/j.celrep.2015.09.025. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 8.Brandao KO, Tabel VA, Atsma DE, Mummery CL, Davis RP. Human pluripotent stem cell models of cardiac disease: from mechanisms to therapies. Dis Model Mech. 2017;10:1039–1059. doi: 10.1242/dmm.030320. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 9.Broughton KM, Li J, Sarmah E, Warren CM, Lin YH, Henze MP, Sanchez-Freire V, Solaro RJ, Russell B. A myosin activator improves actin assembly and sarcomere function of human-induced pluripotent stem cell-derived cardiomyocytes with a troponin T point mutation. Am J Physiol Heart Circ Physiol. 2016;311:H107–H117. doi: 10.1152/ajpheart.00162.2016. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 10.Carrier L, Knoell R, Vignier N, Keller DI, Bausero P, Prudhon B, Isnard R, Ambroisine ML, Fiszman M, Ross J, Jr, Schwartz K, Chien KR. Asymmetric septal hypertrophy in heterozygous cMyBP-C null mice. Cardiovasc Res. 2004;63:293–304. doi: 10.1016/j.cardiores.2004.04.009. [DOI] [PubMed] [Google Scholar]
- 11.Carvajal-Vergara X, Sevilla A, D'Souza SL, Ang YS, Schaniel C, Lee DF, Yang L, Kaplan AD, Adler ED, Rozov R, Ge Y, Cohen N, Edelmann LJ, Chang B, Waghray A, Su J, Pardo S, Lichtenbelt KD, Tartaglia M, Gelb BD, Lemischka IR. Patient-specific induced pluripotent stem-cell-derived models of LEOPARD syndrome. Nature. 2010;465:808–812. doi: 10.1038/nature09005. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 12.Cashman TJ, Josowitz R, Johnson BV, Gelb BD, Costa KD. Human engineered cardiac tissues created using induced pluripotent stem cells reveal functional characteristics of BRAF-mediated hypertrophic cardiomyopathy. PLoS One. 2016;11:e0146697. doi: 10.1371/journal.pone.0146697. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 13.Cazorla O, Szilagyi S, Vignier N, Salazar G, Kramer E, Vassort G, Carrier L, Lacampagne A. Length and protein kinase A modulations of myocytes in cardiac myosin binding protein C-deficient mice. Cardiovasc Res. 2006;69:370–380. doi: 10.1016/j.cardiores.2005.11.009. [DOI] [PubMed] [Google Scholar]
- 14.Chandra M, Tschirgi ML, Tardiff JC. Increase in tension-dependent ATP consumption induced by cardiac troponin T mutation. Am J Physiol Heart Circ Physiol. 2005;289:H2112–H2119. doi: 10.1152/ajpheart.00571.2005. [DOI] [PubMed] [Google Scholar]
- 15.Chou SJ, Yu WC, Chang YL, Chen WY, Chang WC, Chien Y, Yen JC, Liu YY, Chen SJ, Wang CY, Chen YH, Niu DM, Lin SJ, Chen JW, Chiou SH, Leu HB. Energy utilization of induced pluripotent stem cell-derived cardiomyocyte in Fabry disease. Int J Cardiol. 2017;232:255–263. doi: 10.1016/j.ijcard.2017.01.009. [DOI] [PubMed] [Google Scholar]
- 16.Cirino AL, Lakdawala NK, McDonough B, Conner L, Adler D, Weinfeld M, O’Gara P, Rehm HL, Machini K, Lebo M, Blout C, Green RC, MacRae CA, Seidman CE, Ho CY, MedSeq P (2017) A comparison of whole genome sequencing to multigene panel testing in hypertrophic cardiomyopathy patients. Circ Cardiovasc Genet 10(5):e001768. 10.1161/CIRCGENETICS.117.001768 [DOI] [PMC free article] [PubMed]
- 17.Crilley JG, Boehm EA, Blair E, Rajagopalan B, Blamire AM, Styles P, McKenna WJ, Ostman-Smith I, Clarke K, Watkins H. Hypertrophic cardiomyopathy due to sarcomeric gene mutations is characterized by impaired energy metabolism irrespective of the degree of hypertrophy. J Am Coll Cardiol. 2003;41:1776–1782. doi: 10.1016/S0735-1097(02)03009-7. [DOI] [PubMed] [Google Scholar]
- 18.Dambrot C, Braam SR, Tertoolen LG, Birket M, Atsma DE, Mummery CL. Serum supplemented culture medium masks hypertrophic phenotypes in human pluripotent stem cell derived cardiomyocytes. J Cell Mol Med. 2014;18:1509–1518. doi: 10.1111/jcmm.12356. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 19.Davis J, Davis LC, Correll RN, Makarewich CA, Schwanekamp JA, Moussavi-Harami F, Wang D, York AJ, Wu H, Houser SR, Seidman CE, Seidman JG, Regnier M, Metzger JM, Wu JC, Molkentin JD. A tension-based model distinguishes hypertrophic versus dilated cardiomyopathy. Cell. 2016;165:1147–1159. doi: 10.1016/j.cell.2016.04.002. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 20.Du CK, Morimoto S, Nishii K, Minakami R, Ohta M, Tadano N, Lu QW, Wang YY, Zhan DY, Mochizuki M, Kita S, Miwa Y, Takahashi-Yanaga F, Iwamoto T, Ohtsuki I, Sasaguri T. Knock-in mouse model of dilated cardiomyopathy caused by troponin mutation. Circ Res. 2007;101:185–194. doi: 10.1161/CIRCRESAHA.106.146670. [DOI] [PubMed] [Google Scholar]
- 21.Duncker DJ, Bakkers J, Brundel BJ, Robbins J, Tardiff JC, Carrier L. Animal and in silico models for the study of sarcomeric cardiomyopathies. Cardiovasc Res. 2015;105:439–448. doi: 10.1093/cvr/cvv006. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 22.Eschenhagen T, Mummery C, Knollmann BC. Modelling sarcomeric cardiomyopathies in the dish: from human heart samples to iPSC cardiomyocytes. Cardiovasc Res. 2015;105:424–438. doi: 10.1093/cvr/cvv017. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 23.Fraysse B, Weinberger F, Bardswell SC, Cuello F, Vignier N, Geertz B, Starbatty J, Kramer E, Coirault C, Eschenhagen T, Kentish JC, Avkiran M, Carrier L. Increased myofilament Ca(2+) sensitivity and diastolic dysfunction as early consequences of Mybpc3 mutation in heterozygous knock-in mice. J Mol Cell Cardiol. 2012;52:1299–1307. doi: 10.1016/j.yjmcc.2012.03.009. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 24.Frazier AH, Ramirez-Correa GA, Murphy AM. Molecular mechanisms of sarcomere dysfunction in dilated and hypertrophic cardiomyopathy. Prog Pediatr Cardiol. 2011;31:29–33. doi: 10.1016/j.ppedcard.2010.11.006. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 25.Friedrich FW, Carrier L. Genetics of hypertrophic and dilated cardiomyopathy. Curr Pharm Biotechnol. 2012;13:2467–2476. doi: 10.2174/1389201011208062467. [DOI] [PubMed] [Google Scholar]
- 26.Garfinkel AC, Seidman JG, Seidman CE. Genetic pathogenesis of hypertrophic and dilated cardiomyopathy. Heart Fail Clin. 2018;14:139–146. doi: 10.1016/j.hfc.2017.12.004. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 27.Gedicke-Hornung C, Behrens-Gawlik V, Reischmann S, Geertz B, Stimpel D, Weinberger F, Schlossarek S, Precigout G, Braren I, Eschenhagen T, Mearini G, Lorain S, Voit T, Dreyfus PA, Garcia L, Carrier L. Rescue of cardiomyopathy through U7snRNA-mediated exon skipping in Mybpc3-targeted knock-in mice. EMBO Mol Med. 2013;5:1128–1145. doi: 10.1002/emmm.201202168. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 28.Geisterfer-Lowrance AA, Christe M, Conner DA, Ingwall JS, Schoen FJ, Seidman CE, Seidman JG. A mouse model of familial hypertrophic cardiomyopathy. Science. 1996;272:731–734. doi: 10.1126/science.272.5262.731. [DOI] [PubMed] [Google Scholar]
- 29.Gramlich M, Pane LS, Zhou Q, Chen Z, Murgia M, Schotterl S, Goedel A, Metzger K, Brade T, Parrotta E, Schaller M, Gerull B, Thierfelder L, Aartsma-Rus A, Labeit S, Atherton JJ, McGaughran J, Harvey RP, Sinnecker D, Mann M, Laugwitz KL, Gawaz MP, Moretti A. Antisense-mediated exon skipping: a therapeutic strategy for titin-based dilated cardiomyopathy. EMBO Mol Med. 2015;7:562–576. doi: 10.15252/emmm.201505047. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 30.Green EM, Wakimoto H, Anderson RL, Evanchik MJ, Gorham JM, Harrison BC, Henze M, Kawas R, Oslob JD, Rodriguez HM, Song Y, Wan W, Leinwand LA, Spudich JA, McDowell RS, Seidman JG, Seidman CE. A small-molecule inhibitor of sarcomere contractility suppresses hypertrophic cardiomyopathy in mice. Science. 2016;351:617–621. doi: 10.1126/science.aad3456. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 31.Hallas T, Eisen B, Shemer Y, Ben Jehuda R, Mekies LN, Naor S, Schick R, Eliyahu S, Reiter I, Vlodavsky E, Katz YS, Ounap K, Lorber A, Rodenburg R, Mandel H, Gherghiceanu M, Binah O. Investigating the cardiac pathology of SCO2-mediated hypertrophic cardiomyopathy using patients induced pluripotent stem cell-derived cardiomyocytes. J Cell Mol Med. 2018;22:913–925. doi: 10.1111/jcmm.13392. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 32.Han L, Li Y, Tchao J, Kaplan AD, Lin B, Li Y, Mich-Basso J, Lis A, Hassan N, London B, Bett GC, Tobita K, Rasmusson RL, Yang L. Study familial hypertrophic cardiomyopathy using patient-specific induced pluripotent stem cells. Cardiovasc Res. 2014;104:258–269. doi: 10.1093/cvr/cvu205. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 33.Harris SP, Bartley CR, Hacker TA, McDonald KS, Douglas PS, Greaser ML, Powers PA, Moss RL. Hypertrophic cardiomyopathy in cardiac myosin binding protein-C knockout mice. Circ Res. 2002;90:594–601. doi: 10.1161/01.RES.0000012222.70819.64. [DOI] [PubMed] [Google Scholar]
- 34.Hashem SI, Murphy AN, Divakaruni AS, Klos ML, Nelson BC, Gault EC, Rowland TJ, Perry CN, Gu Y, Dalton ND, Bradford WH, Devaney EJ, Peterson KL, Jones KL, MRG T, Chen J, Chi NC, Adler ED. Impaired mitophagy facilitates mitochondrial damage in Danon disease. J Mol Cell Cardiol. 2017;108:86–94. doi: 10.1016/j.yjmcc.2017.05.007. [DOI] [PubMed] [Google Scholar]
- 35.Hinson JT, Chopra A, Nafissi N, Polacheck WJ, Benson CC, Swist S, Gorham J, Yang L, Schafer S, Sheng CC, Haghighi A, Homsy J, Hubner N, Church G, Cook SA, Linke WA, Chen CS, Seidman JG, Seidman CE (2015) HEART DISEASE. Titin mutations in iPS cells define sarcomere insufficiency as a cause of dilated cardiomyopathy. Science 349:982–986 [DOI] [PMC free article] [PubMed]
- 36.Ho CY, Sweitzer NK, McDonough B, Maron BJ, Casey SA, Seidman JG, Seidman CE, Solomon SD. Assessment of diastolic function with Doppler tissue imaging to predict genotype in preclinical hypertrophic cardiomyopathy. Circulation. 2002;105:2992–2997. doi: 10.1161/01.CIR.0000019070.70491.6D. [DOI] [PubMed] [Google Scholar]
- 37.Ho CY, Carlsen C, Thune JJ, Havndrup O, Bundgaard H, Farrohi F, Rivero J, Cirino AL, Andersen PS, Christiansen M, Maron BJ, Orav EJ, Kober L. Echocardiographic strain imaging to assess early and late consequences of sarcomere mutations in hypertrophic cardiomyopathy. Circ Cardiovasc Genet. 2009;2:314–321. doi: 10.1161/CIRCGENETICS.109.862128. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 38.Horvath A, Lemoine MD, Loser A, Mannhardt I, Flenner F, Uzun AU, Neuber C, Breckwoldt K, Hansen A, Girdauskas E, Reichenspurner H, Willems S, Jost N, Wettwer E, Eschenhagen T, Christ T. Low resting membrane potential and low inward rectifier potassium currents are not inherent features of hiPSC-derived cardiomyocytes. Stem Cell Rep. 2018;10:822–833. doi: 10.1016/j.stemcr.2018.01.012. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 39.Ikon N, Ryan RO. Barth syndrome: connecting cardiolipin to cardiomyopathy. Lipids. 2017;52:99–108. doi: 10.1007/s11745-016-4229-7. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 40.Itzhaki I, Maizels L, Huber I, Zwi-Dantsis L, Caspi O, Winterstern A, Feldman O, Gepstein A, Arbel G, Hammerman H, Boulos M, Gepstein L. Modelling the long QT syndrome with induced pluripotent stem cells. Nature. 2011;471:225–229. doi: 10.1038/nature09747. [DOI] [PubMed] [Google Scholar]
- 41.Josowitz R, Mulero-Navarro S, Rodriguez NA, Falce C, Cohen N, Ullian EM, Weiss LA, Rauen KA, Sobie EA, Gelb BD. Autonomous and non-autonomous defects underlie hypertrophic cardiomyopathy in BRAF-mutant hiPSC-derived cardiomyocytes. Stem Cell Rep. 2016;7:355–369. doi: 10.1016/j.stemcr.2016.07.018. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 42.Judge LM, Perez-Bermejo JA, Truong A, Ribeiro AJ, Yoo JC, Jensen CL, Mandegar MA, Huebsch N, Kaake RM, So PL, Srivastava D, Pruitt BL, Krogan NJ, Conklin BR (2017) A BAG3 chaperone complex maintains cardiomyocyte function during proteotoxic stress. JCI Insight 2(14):94623. 10.1172/jci.insight.94623 [DOI] [PMC free article] [PubMed]
- 43.Karakikes I, Stillitano F, Nonnenmacher M, Tzimas C, Sanoudou D, Termglinchan V, Kong CW, Rushing S, Hansen J, Ceholski D, Kolokathis F, Kremastinos D, Katoulis A, Ren L, Cohen N, Gho JM, Tsiapras D, Vink A, Wu JC, Asselbergs FW, Li RA, Hulot JS, Kranias EG, Hajjar RJ. Correction of human phospholamban R14del mutation associated with cardiomyopathy using targeted nucleases and combination therapy. Nat Commun. 2015;6:6955. doi: 10.1038/ncomms7955. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 44.Kawana M, Sarkar SS, Sutton S, Ruppel KM, Spudich JA. Biophysical properties of human beta-cardiac myosin with converter mutations that cause hypertrophic cardiomyopathy. Sci Adv. 2017;3:e1601959. doi: 10.1126/sciadv.1601959. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 45.Keller DI, Coirault C, Rau T, Cheav T, Weyand M, Amann K, Lecarpentier Y, Richard P, Eschenhagen T, Carrier L. Human homozygous R403W mutant cardiac myosin presents disproportionate enhancement of mechanical and enzymatic properties. J Mol Cell Cardiol. 2004;36:355–362. doi: 10.1016/j.yjmcc.2003.12.006. [DOI] [PubMed] [Google Scholar]
- 46.Kilpinen H, Goncalves A, Leha A, Afzal V, Alasoo K, Ashford S, Bala S, Bensaddek D, Casale FP, Culley OJ, Danecek P, Faulconbridge A, Harrison PW, Kathuria A, McCarthy D, McCarthy SA, Meleckyte R, Memari Y, Moens N, Soares F, Mann A, Streeter I, Agu CA, Alderton A, Nelson R, Harper S, Patel M, White A, Patel SR, Clarke L, Halai R, Kirton CM, Kolb-Kokocinski A, Beales P, Birney E, Danovi D, Lamond AI, Ouwehand WH, Vallier L, Watt FM, Durbin R, Stegle O, Gaffney DJ. Common genetic variation drives molecular heterogeneity in human iPSCs. Nature. 2017;546:370–375. doi: 10.1038/nature22403. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 47.Kirschner SE, Becker E, Antognozzi M, Kubis HP, Francino A, Navarro-Lopez F, Bit-Avragim N, Perrot A, Mirrakhimov MM, Osterziel KJ, McKenna WJ, Brenner B, Kraft T. Hypertrophic cardiomyopathy-related beta-myosin mutations cause highly variable calcium sensitivity with functional imbalances among individual muscle cells. Am J Physiol Heart Circ Physiol. 2005;288:H1242–H1251. doi: 10.1152/ajpheart.00686.2004. [DOI] [PubMed] [Google Scholar]
- 48.Korte FS, McDonald KS, Harris SP, Moss RL. Loaded shortening, power output, and rate of force redevelopment are increased with knockout of cardiac myosin binding protein-C. Circ Res. 2003;93:752–758. doi: 10.1161/01.RES.0000096363.85588.9A. [DOI] [PubMed] [Google Scholar]
- 49.Kraft T, Montag J, Radocaj A, Brenner B. Hypertrophic cardiomyopathy: cell-to-cell imbalance in gene expression and contraction force as trigger for disease phenotype development. Circ Res. 2016;119:992–995. doi: 10.1161/CIRCRESAHA.116.309804. [DOI] [PubMed] [Google Scholar]
- 50.Lan F, Lee AS, Liang P, Sanchez-Freire V, Nguyen PK, Wang L, Han L, Yen M, Wang Y, Sun N, Abilez OJ, Hu S, Ebert AD, Navarrete EG, Simmons CS, Wheeler M, Pruitt B, Lewis R, Yamaguchi Y, Ashley EA, Bers DM, Robbins RC, Longaker MT, Wu JC. Abnormal calcium handling properties underlie familial hypertrophic cardiomyopathy pathology in patient-specific induced pluripotent stem cells. Cell Stem Cell. 2013;12:101–113. doi: 10.1016/j.stem.2012.10.010. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 51.Lee YK, Lau YM, Ng KM, Lai WH, Ho SL, Tse HF, Siu CW, Ho PW. Efficient attenuation of Friedreich’s ataxia (FRDA) cardiomyopathy by modulation of iron homeostasis-human induced pluripotent stem cell (hiPSC) as a drug screening platform for FRDA. Int J Cardiol. 2016;203:964–971. doi: 10.1016/j.ijcard.2015.11.101. [DOI] [PubMed] [Google Scholar]
- 52.Li S, Pan H, Tan C, Sun Y, Song Y, Zhang X, Yang W, Wang X, Li D, Dai Y, Ma Q, Xu C, Zhu X, Kang L, Fu Y, Xu X, Shu J, Zhou N, Han F, Qin D, Huang W, Liu Z, Yan Q. Mitochondrial dysfunctions contribute to hypertrophic cardiomyopathy in patient iPSC-derived cardiomyocytes with MT-RNR2 mutation. Stem Cell Rep. 2018;10:808–821. doi: 10.1016/j.stemcr.2018.01.013. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 53.Liang G, Zhang Y. Genetic and epigenetic variations in iPSCs: potential causes and implications for application. Cell Stem Cell. 2013;13:149–159. doi: 10.1016/j.stem.2013.07.001. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 54.Lin B, Li Y, Han L, Kaplan AD, Ao Y, Kalra S, Bett GC, Rasmusson RL, Denning C, Yang L. Modeling and study of the mechanism of dilated cardiomyopathy using induced pluripotent stem cells derived from individuals with Duchenne muscular dystrophy. Dis Model Mech. 2015;8:457–466. doi: 10.1242/dmm.019505. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 55.Long C, Li H, Tiburcy M, Rodriguez-Caycedo C, Kyrychenko V, Zhou H, Zhang Y, Min YL, Shelton JM, Mammen PPA, Liaw NY, Zimmermann WH, Bassel-Duby R, Schneider JW, Olson EN. Correction of diverse muscular dystrophy mutations in human engineered heart muscle by single-site genome editing. Sci Adv. 2018;4:eaap9004. doi: 10.1126/sciadv.aap9004. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 56.Lynn ML, Tal Grinspan L, Holeman TA, Jimenez J, Strom J, Tardiff JC. The structural basis of alpha-tropomyosin linked (Asp230Asn) familial dilated cardiomyopathy. J Mol Cell Cardiol. 2017;108:127–137. doi: 10.1016/j.yjmcc.2017.06.001. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 57.Ma N, Zhang J, Itzhaki I, Zhang SL, Chen H, Haddad F, Kitani T, Wilson KD, Tian L, Shrestha R, Wu H, Lam CK, Sayed N, Wu JC (2018) Determining the pathogenicity of a genomic variant of uncertain significance using CRISPR/Cas9 and human-induced pluripotent stem cells. Circulation:CIRCULATIONAHA.117.032273 [DOI] [PMC free article] [PubMed]
- 58.Mak TSH, Lee YK, Tang CS, Hai JSH, Ran X, Sham PC, Tse HF. Coverage and diagnostic yield of whole exome sequencing for the evaluation of cases with dilated and hypertrophic cardiomyopathy. Sci Rep. 2018;8:10846. doi: 10.1038/s41598-018-29263-3. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 59.Marian AJ, Braunwald E. Hypertrophic cardiomyopathy: genetics, pathogenesis, clinical manifestations, diagnosis, and therapy. Circ Res. 2017;121:749–770. doi: 10.1161/CIRCRESAHA.117.311059. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 60.Maron BJ, Towbin JA, Thiene G, Antzelevitch C, Corrado D, Arnett D, Moss AJ, Seidman CE, Young JB, American Heart A, Council on Clinical Cardiology HF, Transplantation C, Quality of C, Outcomes R, Functional G, Translational Biology Interdisciplinary Working G, Council on E, and Prevention Contemporary definitions and classification of the cardiomyopathies: an American Heart Association Scientific Statement from the Council on Clinical Cardiology, Heart Failure and Transplantation Committee; Quality of Care and Outcomes Research and Functional Genomics and Translational Biology Interdisciplinary Working Groups; and Council on Epidemiology and Prevention. Circulation. 2006;113:1807–1816. doi: 10.1161/CIRCULATIONAHA.106.174287. [DOI] [PubMed] [Google Scholar]
- 61.McConnell BK, Jones KA, Fatkin D, Arroyo LH, Lee RT, Aristizabal O, Turnbull DH, Georgakopoulos D, Kass D, Bond M, Niimura H, Schoen FJ, Conner D, Fischman DA, Seidman CE, Seidman JG. Dilated cardiomyopathy in homozygous myosin-binding protein-C mutant mice. J Clin Invest. 1999;104:1235–1244. doi: 10.1172/JCI7377. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 62.McConnell BK, Fatkin D, Semsarian C, Jones KA, Georgakopoulos D, Maguire CT, Healey MJ, Mudd JO, Moskowitz IP, Conner DA, Giewat M, Wakimoto H, Berul CI, Schoen FJ, Kass DA, Seidman CE, Seidman JG. Comparison of two murine models of familial hypertrophic cardiomyopathy. Circ Res. 2001;88:383–389. doi: 10.1161/01.RES.88.4.383. [DOI] [PubMed] [Google Scholar]
- 63.McNamara JW, Li A, Smith NJ, Lal S, Graham RM, Kooiker KB, van Dijk SJ, Remedios CGD, Harris SP, Cooke R. Ablation of cardiac myosin binding protein-C disrupts the super-relaxed state of myosin in murine cardiomyocytes. J Mol Cell Cardiol. 2016;94:65–71. doi: 10.1016/j.yjmcc.2016.03.009. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 64.Mearini G, Stimpel D, Kramer E, Geertz B, Braren I, Gedicke-Hornung C, Precigout G, Muller OJ, Katus HA, Eschenhagen T, Voit T, Garcia L, Lorain S, Carrier L. Repair of Mybpc3 mRNA by 5′-trans-splicing in a mouse model of hypertrophic cardiomyopathy. Mol Ther Nucleic Acids. 2013;2:e102. doi: 10.1038/mtna.2013.31. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 65.Mearini G, Stimpel D, Geertz B, Weinberger F, Krämer E, Schlossarek S, Mourot-Filiatre J, Stöhr A, Dutshc A, Wijnker PJM, Braren I, Katus HA, Müller OJ, Voit T, Eschenhagen T, Carrier L. Mybpc3 gene therapy for neonatal cardiomyopathy enables longterm disease prevention in mice. Nat Commun. 2014;5:5515. doi: 10.1038/ncomms6515. [DOI] [PubMed] [Google Scholar]
- 66.Michels M, Soliman OI, Kofflard MJ, Hoedemaekers YM, Dooijes D, Majoor-Krakauer D, ten Cate FJ. Diastolic abnormalities as the first feature of hypertrophic cardiomyopathy in Dutch myosin-binding protein C founder mutations. JACC Cardiovasc Imaging. 2009;2:58–64. doi: 10.1016/j.jcmg.2008.08.003. [DOI] [PubMed] [Google Scholar]
- 67.Montgomery DE, Tardiff JC, Chandra M. Cardiac troponin T mutations: correlation between the type of mutation and the nature of myofilament dysfunction in transgenic mice. J Physiol. 2001;536:583–592. doi: 10.1111/j.1469-7793.2001.0583c.xd. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 68.Moretti A, Bellin M, Welling A, Jung CB, Lam JT, Bott-Flugel L, Dorn T, Goedel A, Hohnke C, Hofmann F, Seyfarth M, Sinnecker D, Schomig A, Laugwitz KL. Patient-specific induced pluripotent stem-cell models for long-QT syndrome. N Engl J Med. 2010;363:1397–1409. doi: 10.1056/NEJMoa0908679. [DOI] [PubMed] [Google Scholar]
- 69.Mosqueira D, Mannhardt I, Bhagwan JR, Lis-Slimak K, Katili P, Scott E, Hassan M, Prondzynski M, Harmer SC, Tinker A, Smith JGW, Carrier L, Williams PM, Gaffney D, Eschenhagen T, Hansen A, Denning C (2018) CRISPR/Cas9 editing in human pluripotent stem cell-cardiomyocytes highlights arrhythmias, hypocontractility, and energy depletion as potential therapeutic targets for hypertrophic cardiomyopathy. Eur Heart J [DOI] [PMC free article] [PubMed]
- 70.Musunuru K, Sheikh F, Gupta RM, Houser SR, Maher KO, Milan DJ, Terzic A, Wu JC, American Heart Association Council on Functional G, Translational B, Council on Cardiovascular Disease in the Y, Council on C, and Stroke N Induced pluripotent stem cells for cardiovascular disease modeling and precision medicine: a scientific statement from the American Heart Association. Circ Genom Precis Med. 2018;11:e000043. doi: 10.1161/HCG.0000000000000043. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 71.Neubauer S, Krahe T, Schindler R, Horn M, Hillenbrand H, Entzeroth C, Mader H, Kromer EP, Riegger GA, Lackner K, et al. 31P magnetic resonance spectroscopy in dilated cardiomyopathy and coronary artery disease. Altered cardiac high-energy phosphate metabolism in heart failure. Circulation. 1992;86:1810–1818. doi: 10.1161/01.CIR.86.6.1810. [DOI] [PubMed] [Google Scholar]
- 72.Ojala M, Prajapati C, Polonen RP, Rajala K, Pekkanen-Mattila M, Rasku J, Larsson K, Aalto-Setala K. Mutation-specific phenotypes in hiPSC-derived cardiomyocytes carrying either myosin-binding protein C or alpha-tropomyosin mutation for hypertrophic cardiomyopathy. Stem Cells Int. 2016;2016:1684792. doi: 10.1155/2016/1684792. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 73.Phelan DG, Anderson DJ, Howden SE, Wong RC, Hickey PF, Pope K, Wilson GR, Pebay A, Davis AM, Petrou S, Elefanty AG, Stanley EG, James PA, Macciocca I, Bahlo M, Cheung MM, Amor DJ, Elliott DA, Lockhart PJ. ALPK3-deficient cardiomyocytes generated from patient-derived induced pluripotent stem cells and mutant human embryonic stem cells display abnormal calcium handling and establish that ALPK3 deficiency underlies familial cardiomyopathy. Eur Heart J. 2016;37:2586–2590. doi: 10.1093/eurheartj/ehw160. [DOI] [PubMed] [Google Scholar]
- 74.Pioner JM, Racca AW, Klaiman JM, Yang KC, Guan X, Pabon L, Muskheli V, Zaunbrecher R, Macadangdang J, Jeong MY, Mack DL, Childers MK, Kim DH, Tesi C, Poggesi C, Murry CE, Regnier M. Isolation and mechanical measurements of myofibrils from human induced pluripotent stem cell-derived cardiomyocytes. Stem Cell Rep. 2016;6:885–896. doi: 10.1016/j.stemcr.2016.04.006. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 75.Pohlmann L, Kroger I, Vignier N, Schlossarek S, Kramer E, Coirault C, Sultan KR, El-Armouche A, Winegrad S, Eschenhagen T, Carrier L. Cardiac myosin-binding protein C is required for complete relaxation in intact myocytes. Circ Res. 2007;101:928–938. doi: 10.1161/CIRCRESAHA.107.158774. [DOI] [PubMed] [Google Scholar]
- 76.Prondzynski M, Kramer E, Laufer SD, Shibamiya A, Pless O, Flenner F, Muller OJ, Munch J, Redwood C, Hansen A, Patten M, Eschenhagen T, Mearini G, Carrier L. Evaluation of MYBPC3 trans-splicing and gene replacement as therapeutic options in human iPSC-derived cardiomyocytes. Mol Ther Nucleic Acids. 2017;7:475–486. doi: 10.1016/j.omtn.2017.05.008. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 77.Raval KK, Tao R, White BE, De Lange WJ, Koonce CH, Yu J, Kishnani PS, Thomson JA, Mosher DF, Ralphe JC, Kamp TJ. Pompe disease results in a Golgi-based glycosylation deficit in human induced pluripotent stem cell-derived cardiomyocytes. J Biol Chem. 2015;290:3121–3136. doi: 10.1074/jbc.M114.628628. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 78.Richard P, Charron P, Carrier L, Ledeuil C, Cheav T, Pichereau C, Benaiche A, Isnard R, Dubourg O, Burban M, Gueffet JP, Millaire A, Desnos M, Schwartz K, Hainque B, Komajda M. Hypertrophic cardiomyopathy: distribution of disease genes, spectrum of mutations and implications for molecular diagnosis strategy. Circulation. 2003;107:2227–2232. doi: 10.1161/01.CIR.0000066323.15244.54. [DOI] [PubMed] [Google Scholar]
- 79.Robinson P, Mirza M, Knott A, Abdulrazzak H, Willott R, Marston S, Watkins H, Redwood C. Alterations in thin filament regulation induced by a human cardiac troponin T mutant that causes dilated cardiomyopathy are distinct from those induced by troponin T mutants that cause hypertrophic cardiomyopathy. J Biol Chem. 2002;277:40710–40716. doi: 10.1074/jbc.M203446200. [DOI] [PubMed] [Google Scholar]
- 80.Ross SB, Fraser ST, Nowak N, Semsarian C. Generation of induced pluripotent stem cells (iPSCs) from a hypertrophic cardiomyopathy patient with the pathogenic variant p.Val698Ala in beta-myosin heavy chain (MYH7) gene. Stem Cell Res. 2017;20:88–90. doi: 10.1016/j.scr.2017.02.015. [DOI] [PubMed] [Google Scholar]
- 81.Rowin EJ, Maron MS, Maron BJ. Response by Rowin et al to letter regarding article, “Clinical profile and consequences of atrial fibrillation in hypertrophic cardiomyopathy”. Circulation. 2018;137:2541–2542. doi: 10.1161/CIRCULATIONAHA.118.034857. [DOI] [PubMed] [Google Scholar]
- 82.Sato Y, Kobayashi H, Higuchi T, Shimada Y, Era T, Kimura S, Eto Y, Ida H, Ohashi T. Disease modeling and lentiviral gene transfer in patient-specific induced pluripotent stem cells from late-onset Pompe disease patient. Mol Ther Methods Clin Dev. 2015;2:15023. doi: 10.1038/mtm.2015.23. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 83.Satoh H, Delbridge LM, Blatter LA, Bers DM. Surface:volume relationship in cardiac myocytes studied with confocal microscopy and membrane capacitance measurements: species-dependence and developmental effects. Biophys J. 1996;70:1494–1504. doi: 10.1016/S0006-3495(96)79711-4. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 84.Siu CW, Lee YK, Ho JC, Lai WH, Chan YC, Ng KM, Wong LY, Au KW, Lau YM, Zhang J, Lay KW, Colman A, Tse HF. Modeling of lamin A/C mutation premature cardiac aging using patient-specific induced pluripotent stem cells. Aging (Albany NY) 2012;4:803–822. doi: 10.18632/aging.100503. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 85.Spitalieri P, Talarico RV, Caioli S, Murdocca M, Serafino A, Girasole M, Dinarelli S, Longo G, Pucci S, Botta A, Novelli G, Zona C, Mango R, Sangiuolo F. Modelling the pathogenesis of myotonic dystrophy type 1 cardiac phenotype through human iPSC-derived cardiomyocytes. J Mol Cell Cardiol. 2018;118:95–109. doi: 10.1016/j.yjmcc.2018.03.012. [DOI] [PubMed] [Google Scholar]
- 86.Streckfuss-Bomeke K, Tiburcy M, Fomin A, Luo X, Li W, Fischer C, Ozcelik C, Perrot A, Sossalla S, Haas J, Vidal RO, Rebs S, Khadjeh S, Meder B, Bonn S, Linke WA, Zimmermann WH, Hasenfuss G, Guan K. Severe DCM phenotype of patient harboring RBM20 mutation S635A can be modeled by patient-specific induced pluripotent stem cell-derived cardiomyocytes. J Mol Cell Cardiol. 2017;113:9–21. doi: 10.1016/j.yjmcc.2017.09.008. [DOI] [PubMed] [Google Scholar]
- 87.Sun N, Yazawa M, Liu J, Han L, Sanchez-Freire V, Abilez OJ, Navarrete EG, Hu S, Wang L, Lee A, Pavlovic A, Lin S, Chen R, Hajjar RJ, Snyder MP, Dolmetsch RE, Butte MJ, Ashley EA, Longaker MT, Robbins RC, Wu JC. Patient-specific induced pluripotent stem cells as a model for familial dilated cardiomyopathy. Sci Transl Med. 2012;4:130ra147. doi: 10.1126/scitranslmed.3003552. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 88.Sweeney HL, Feng HS, Yang Z, Watkins H. Functional analyses of troponin T mutations that cause hypertrophic cardiomyopathy: insights into disease pathogenesis and troponin function. Proc Natl Acad Sci U S A. 1998;95:14406–14410. doi: 10.1073/pnas.95.24.14406. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 89.Takahashi K, Tanabe K, Ohnuki M, Narita M, Ichisaka T, Tomoda K, Yamanaka S. Induction of pluripotent stem cells from adult human fibroblasts by defined factors. Cell. 2007;131:861–872. doi: 10.1016/j.cell.2007.11.019. [DOI] [PubMed] [Google Scholar]
- 90.Tanaka A, Yuasa S, Mearini G, Egashira T, Seki T, Kodaira M, Kusumoto D, Kuroda Y, Okata S, Suzuki T, Inohara T, Arimura T, Makino S, Kimura K, Kimura A, Furukawa T, Carrier L, Node K, Fukuda K (2014) Endothelin-1 induces myofibrillar disarray and contractile vector variability in hypertrophic cardiomyopathy-induced pluripotent stem cell-derived cardiomyocytes. J Am Heart Assoc 3(6):e001263. 10.1161/JAHA.114.001263 [DOI] [PMC free article] [PubMed]
- 91.Tse HF, Ho JC, Choi SW, Lee YK, Butler AW, Ng KM, Siu CW, Simpson MA, Lai WH, Chan YC, Au KW, Zhang J, Lay KW, Esteban MA, Nicholls JM, Colman A, Sham PC. Patient-specific induced-pluripotent stem cells-derived cardiomyocytes recapitulate the pathogenic phenotypes of dilated cardiomyopathy due to a novel DES mutation identified by whole exome sequencing. Hum Mol Genet. 2013;22:1395–1403. doi: 10.1093/hmg/dds556. [DOI] [PubMed] [Google Scholar]
- 92.Ujfalusi Z, Vera CD, Mijailovich SM, Svicevic M, Yu EC, Kawana M, Ruppel KM, Spudich JA, Geeves MA, Leinwand LA. Dilated cardiomyopathy myosin mutants have reduced force-generating capacity. J Biol Chem. 2018;293:9017–9029. doi: 10.1074/jbc.RA118.001938. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 93.Vaidyanathan R, Markandeya YS, Kamp TJ, Makielski JC, January CT, Eckhardt LL. IK1-enhanced human-induced pluripotent stem cell-derived cardiomyocytes: an improved cardiomyocyte model to investigate inherited arrhythmia syndromes. Am J Physiol Heart Circ Physiol. 2016;310:H1611–H1621. doi: 10.1152/ajpheart.00481.2015. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 94.van der Velden J, Ho CY, Tardiff JC, Olivotto I, Knollmann BC, Carrier L. Research priorities in sarcomeric cardiomyopathies. Cardiovasc Res. 2015;105:449–456. doi: 10.1093/cvr/cvv019. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 95.van Dijk SJ, Dooijes D, Dos Remedios C, Michels M, Lamers JM, Winegrad S, Schlossarek S, Carrier L, Ten Cate FJ, Stienen GJ, van der Velden J. Cardiac myosin-binding protein C mutations and hypertrophic cardiomyopathy: haploinsufficiency, deranged phosphorylation, and cardiomyocyte dysfunction. Circulation. 2009;119:1473–1483. doi: 10.1161/CIRCULATIONAHA.108.838672. [DOI] [PubMed] [Google Scholar]
- 96.van Dijk SJ, Paalberends ER, Najafi A, Michels M, Sadayappan S, Carrier L, Boontje NM, Kuster DW, van Slegtenhorst M, Dooijes D, Dos Remedios C, Ten Cate FJ, Stienen GJ, van der Velden J. Contractile dysfunction irrespective of the mutant protein in human hypertrophic cardiomyopathy with normal systolic function. Circ Heart Fail. 2012;5:36–46. doi: 10.1161/CIRCHEARTFAILURE.111.963702. [DOI] [PubMed] [Google Scholar]
- 97.Vandecasteele G, Eschenhagen T, Fischmeister R. Role of the NO-cGMP pathway in the muscarinic regulation of the L-type Ca2+ current in human atrial myocytes. J Physiol. 1998;506(Pt 3):653–663. doi: 10.1111/j.1469-7793.1998.653bv.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 98.Vignier N, Schlossarek S, Fraysse B, Mearini G, Kramer E, Pointu H, Mougenot N, Guiard J, Reimer R, Hohenberg H, Schwartz K, Vernet M, Eschenhagen T, Carrier L. Nonsense-mediated mRNA decay and ubiquitin-proteasome system regulate cardiac myosin-binding protein C mutant levels in cardiomyopathic mice. Circ Res. 2009;105:239–248. doi: 10.1161/CIRCRESAHA.109.201251. [DOI] [PubMed] [Google Scholar]
- 99.Wang G, McCain ML, Yang L, He A, Pasqualini FS, Agarwal A, Yuan H, Jiang D, Zhang D, Zangi L, Geva J, Roberts AE, Ma Q, Ding J, Chen J, Wang DZ, Li K, Wang J, Wanders RJ, Kulik W, Vaz FM, Laflamme MA, Murry CE, Chien KR, Kelley RI, Church GM, Parker KK, Pu WT. Modeling the mitochondrial cardiomyopathy of Barth syndrome with induced pluripotent stem cell and heart-on-chip technologies. Nat Med. 2014;20:616–623. doi: 10.1038/nm.3545. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 100.Wang L, Kim K, Parikh S, Cadar AG, Bersell KR, He H, Pinto JR, Kryshtal DO, Knollmann BC. Hypertrophic cardiomyopathy-linked mutation in troponin T causes myofibrillar disarray and pro-arrhythmic action potential changes in human iPSC cardiomyocytes. J Mol Cell Cardiol. 2018;114:320–327. doi: 10.1016/j.yjmcc.2017.12.002. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 101.Witjas-Paalberends ER, Piroddi N, Stam K, van Dijk SJ, Oliviera VS, Ferrara C, Scellini B, Hazebroek M, ten Cate FJ, van Slegtenhorst M, dos Remedios C, Niessen HW, Tesi C, Stienen GJ, Heymans S, Michels M, Poggesi C, van der Velden J. Mutations in MYH7 reduce the force generating capacity of sarcomeres in human familial hypertrophic cardiomyopathy. Cardiovasc Res. 2013;99:432–441. doi: 10.1093/cvr/cvt119. [DOI] [PubMed] [Google Scholar]
- 102.Witjas-Paalberends ER, Guclu A, Germans T, Knaapen P, Harms HJ, Vermeer AM, Christiaans I, Wilde AA, Dos Remedios C, Lammertsma AA, van Rossum AC, Stienen GJ, van Slegtenhorst M, Schinkel AF, Michels M, Ho CY, Poggesi C, van der Velden J. Gene-specific increase in the energetic cost of contraction in hypertrophic cardiomyopathy caused by thick filament mutations. Cardiovasc Res. 2014;103:248–257. doi: 10.1093/cvr/cvu127. [DOI] [PubMed] [Google Scholar]
- 103.Wu H, Lee J, Vincent LG, Wang Q, Gu M, Lan F, Churko JM, Sallam KI, Matsa E, Sharma A, Gold JD, Engler AJ, Xiang YK, Bers DM, Wu JC. Epigenetic regulation of phosphodiesterases 2A and 3A underlies compromised beta-adrenergic signaling in an iPSC model of dilated cardiomyopathy. Cell Stem Cell. 2015;17:89–100. doi: 10.1016/j.stem.2015.04.020. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 104.Wyles SP, Li X, Hrstka SC, Reyes S, Oommen S, Beraldi R, Edwards J, Terzic A, Olson TM, Nelson TJ. Modeling structural and functional deficiencies of RBM20 familial dilated cardiomyopathy using human induced pluripotent stem cells. Hum Mol Genet. 2016;25:254–265. doi: 10.1093/hmg/ddv468. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 105.Yang X, Pabon L, Murry CE. Engineering adolescence: maturation of human pluripotent stem cell-derived cardiomyocytes. Circ Res. 2014;114:511–523. doi: 10.1161/CIRCRESAHA.114.300558. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 106.Yeung C, Enriquez A, Suarez-Fuster L, Baranchuk A (2018) Atrial fibrillation in patients with inherited cardiomyopathies. Europace. 10.1093/europace/euy064 [DOI] [PubMed]
- 107.Yoshida Y, Yamanaka S. Induced pluripotent stem cells 10 years later: for cardiac applications. Circ Res. 2017;120:1958–1968. doi: 10.1161/CIRCRESAHA.117.311080. [DOI] [PubMed] [Google Scholar]