

Abstract
Osteoporosis, femoral head necrosis, and congenital bone defects are orthopedic disorders characterized by reduced bone generation and insufficient bone mass. Bone regenerative therapy primarily relies on the bone marrow mesenchymal stem cells (BMSCs) and their ability to differentiate osteogenically. Icariin (ICA) is the active ingredient of Herba epimedii, a common herb used in traditional Chinese medicine (TCM) formulations, and can effectively enhance BMSC proliferation and osteogenesis. However, the underlying mechanism of ICA action in BMSCs is not completely clear. In this review, we provide an overview of the studies on the role and mechanism of action of ICA in BMSCs, to provide greater insights into its potential clinical use in bone regeneration.
1. Introduction
Conventional treatments for orthopedic disorders like osteoporosis, femoral head necrosis, bone defects, and nonunion disorders [1–7] have poor clinical efficacy due to their inability to ameliorate the loss in bone mass. Therefore, the current focus of treating bone disorders is tissue regeneration using bone marrow mesenchymal stem cells (BMSCs) [7, 8]. A number of studies have investigated the effects of various drugs, mechanical stress, physical stimuli, and scaffolds on BMSCs [9–11], in order to clinically translate its regenerative capacity [12]. Traditional Chinese medicine (TCM) has also garnered considerable interest in recent years due to its minimal toxicity [13]. According to the principles of TCM, bone function is closely associated with the balance of kidney yin and yang. Herbs such as Herba epimedii, Fructus psoralea, Drynaria fortunei, and Radix dipsaci are known to invigorate the kidney and restore the balance and are therefore frequently used to treat bone disease. Currently, Herba epimedii is the most deeply studied among those herbs in the bone regeneration field [14]. Icariin (ICA) is the main active ingredient of Herba epimedii, which has been used in TCM formulations to strengthen the muscles and bones [15]. Although Herba epimedii is still used to treat orthopedic disorders, its mechanism of action remains unclear. Several studies have analyzed the effects of ICA in BMSCs and other cells and provided new insights into its therapeutic role in orthopedic disorders. In this review, we have summarized the recent findings on the role and mechanism of action of ICA in BMSCs.
2. Basic Properties of ICA
Herba epimedii (Yin Yang Huo in Chinese) is the dried leaf of Epimedium brevicornum Maxim as recorded in the Chinese Pharmacopoeia [16–18], as well as the 400-year-old Chinese medical classic Shennong Ben Cao Jing. It nourishes the kidney and significantly reinforces yang [18]. More than 20 flavonoids have been identified by chemical reaction and spectral analysis and isolated from Herba epimedii by systematical separation technology [17]. ICA (C33H40O15, molecular weight: 676.66, Figure 1) [17] is one of the primary active constituents which is also considered as the chemical marker for quality control components of Herba epimedii [18]. Specifically, the contents of ICA and the total flavonoids are no less than 0.5% and 5.0% of the components, respectively [16]. The main isolation methods for ICA include water boiling extraction, ethanol reflux extraction [19–22], and an ultrasonic-assisted ethanol extraction method developed by Zhang et al. [21]. The optimum ultrasonic-assisted extraction conditions were determined by an orthogonal test as follows: 50% (v/v) ethanol solution, 30 ml/g liquid-solid ratio, ultrasonic duration for 30 minutes, 50°C extraction temperature, and extraction for 3 times. Compared with the traditional water boiling extraction method, this kind of method has higher efficiency. In addition, microwave, high pressure, and vacuum reflux extraction methods have also been tested [17, 21, 23, 24]. Herba epimedii is widely used for the treatment of osteoporosis in China [18, 25, 26], and results from clinical trials [27, 28] show similar antiosteoporotic effects of its flavonoid extracts, as well as those of Epimedium total flavone capsules. The effects of ICA have largely been studied in animal or in vitro models [2, 4, 5, 9, 29, 30], and its potential clinical applications are rarely reported.
Figure 1.
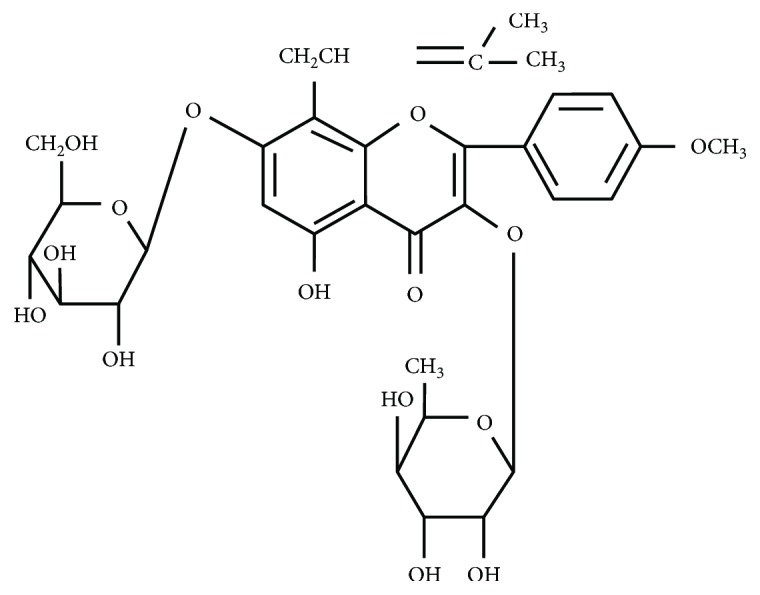
Chemical structure of ICA.
3. Role of BMSCs in Bone Regenerative Therapy
The common culture method of mouse BMSCs was as described in several studies [31–34]. Bone marrow is extracted from the femur and tibia of mice using an aseptic technique. The bone marrow is then cultured in vitro and subcultured to the third passage. As for human BMSCs, the proximal femur or posterior iliac crest is the common part from which to extract human bone marrow [30, 32, 33, 35]. The bone marrow is cultured in vitro and usually cultured to the third passage [33]. It should be noted that the cell phenotype identification is important in BMSC culture processes [32]. The sorted mouse CD29+Sca-1+CD45−CD11b− BMSCs and human CD146+STRO-1+CD45− BMSCs are cultured for 1-2 weeks to reach 80%-85% confluence [32, 33]. Then, first-passage BMSCs are detached and seeded in culture flasks for enrichment of cell populations.
The bone is a kind of mineralized connective tissue which exhibits four types of cells: osteoclasts, osteoblasts, osteocytes, and bone lining cells. Osteoblasts, which comprise 4-6% of the total bone cells, are located along the bone surface and are widely known for their role in bone formation [36]. The osteocytes accounting for 90-95% of the total bone cells are located within lacunae surrounded by a mineralized bone matrix wherein they exhibit a dendritic morphology [37]. The morphology of embedded osteocytes varies independently of bone types. Osteoclasts derive from mononuclear cells of the hematopoietic stem lineage which are multinucleated and terminally differentiated. Though it exhibits an inert appearance, bone tissue is constantly resorbed by osteoclasts and reformed by osteoblasts in a highly dynamic way. The process of bone remodeling is greatly complicated which is in a cycle comprising three stages: (1) initiation of bone resorption by osteoclasts, (2) transition between resorption and reformation, and (3) formation of new bone by osteoblasts. This bone remodeling process requires coordinated actions of osteocytes, osteoclasts, osteoblasts, and bone lining cells which together form the temporary anatomical structure called the basic multicellular unit [38, 39]. Osteoblasts are the main functional cells of bone formation, which are mainly responsible for the synthesis, secretion, and mineralization of the bone matrix. Osteoblasts can produce extracellular matrix proteins and mineralization regulators [38], during which period it undergoes significant proliferation and differentiation. Osteoclasts are the only cells with bone resorption function. Many cytokines such as interleukin-6 (IL-6), tumor necrosis factor-α (TNF-α), and cathepsin K can provide signals for osteoclast differentiation and bone resorption, promote the recruitment of osteoclast precursors, and drive osteoclast differentiation and bone resorption [40–43]. The osteoblasts and the differentiation of osteoclasts are regulated by many signaling pathways, among which the bone morphogenetic protein-drosophila mothers against decapentaplegic protein (BMP-Smad) signaling is important [44]. The bone mass can be increased by promoting the directional differentiation of BMSCs into osteoblasts, which is driven by the runt-related transcription factor 2 (Runx2) and Osterix (Osx) [36, 38, 45, 46]. Since osteocytes are derived from the BMSC lineage through osteoblast differentiation [38], so finding potential drugs prompting the differentiation of BMSCs into osteoblasts may be a promising strategy for bone regeneration.
Hematopoietic stem cells (HSCs) and BMSCs are the two pluripotent cell types found in the bone matrix [47, 48]. BMSCs were first isolated from the adult bone marrow [49] and can differentiate into the adipocytes, chondrocytes, osteoblasts, and myoblasts [29, 50, 51]. Therefore, BMSCs are a highly promising therapeutic option for cardiovascular, orthopedic, and joint degenerative diseases [52–57]. Several studies have examined the ability of BMSCs to improve bone formation and prevent bone loss and necrosis, in addition to ameliorating congenital bone defects and osteoporosis [58, 59]. However, the engrafted BMSCs have poor survival and a low rate of differentiation at the site of transplantation, which significantly reduce the efficacy of BMSC-based regenerative therapy. Therefore, it is essential to develop new drugs to enhance BMSC proliferation and differentiation.
BMSC osteogenesis is the key step in bone regeneration and is affected by several factors including hormones, growth factors, environmental factors, and microRNAs [11]. BMSCs not only give rise to bone tissues but can also differentiate into adipose cells or osteoblasts [60]. Under physiological conditions, a dynamic balance exists between the osteogenic and the adipogenic potential of BMSCs [61–63] and is primarily regulated by Runx2 and the peroxisomal proliferator-activated receptor gamma (PPARγ) [30, 64]. Runx2 is regulated by BMP-2 and is a key modulator of osteogenic differentiation, whereas PPARγ promotes adipogenesis and inhibits osteogenesis [65–67]. Both signaling pathways concurrently regulate different cytokines to determine the fate of BMSC differentiation [68, 69]. The extracellular signal-regulated kinase-mitogen-activated protein kinase (ERK-MAPK) signaling pathway is also a key player in regulating BMSC differentiation [70], whereas the platelet-derived growth factor (PDGF) pathway is an essential proosteogenic pathway [71]. Cao et al. showed that Notch and BMP-9/Smad signaling synergistically enhanced osteogenic differentiation [72], and Li et al. found that miR-21 directly acted on Smad7 in the Smad7-Smad1/5/8-Runx2 pathway to modulate osteogenic differentiation [73]. Long et al. demonstrated that miR-139-5p regulated osteogenic differentiation of BMSCs via the Wnt/β-catenin pathway [74]. Furthermore, the transforming growth factor-β/bone morphogenetic protein (TGF-β/BMP) [73], phosphatidylinositol 3-kinase/protein kinase B/glycogen synthase kinase-3 (PI3K/Akt/GSK-3) [9], extracellular regulated kinase (ERK), PI3K/Akt [75], and insulin-like growth factor 1 (IGF1) [76] pathways also play important roles in osteogenic differentiation and bone formation (Figure 2). Since the two differentiation pathways are competing [68, 69], interregulatory, and interconvertible, certain growth factors can be used to promote the osteogenic differentiation of BMSCs ex vivo for bone tissue engineering.
Figure 2.

Signaling pathways involved in osteogenesis, adipogenesis, and migration regulation of BMSCs.
Migration of BMSCs to the site of bone defect is a critical step in the treatment of orthopedic disorders [77]. Previous studies [78–81] have shown that the C-X-C motif chemokine ligand 12/C-X-C chemokine receptor type 4 (CXCL12/CXCR4) axis modulates BMSC homing and promotes angiogenesis, and the BMP-2/Smads/Runx2/Osterix axis modulates BMSC osteoblastic differentiation. The crosstalk between these two signaling axes is mediated by CXCR4, which modulates the migration [82] and osteogenic differentiation of BMSCs. Some studies have demonstrated that BMSC migration can also occur via the BMP-Smad1/5/8-twist-related protein 1/activating transcription factor 4 (Twist1/Atf4) [83, 84] and CXCR4/stromal-derived factor 1 (SDF-1) [85–87] axes and the Smad pathway [88].
4. Mechanisms of BMSC Regulation by ICA
4.1. BMSC Proliferation and Osteogenesis Promoted by ICA
ICA has multiple pharmacological activities, including hormone-like, antitumor, immunomodulatory, and antioxidative effects [89–94]. Studies [95, 96] show that ICA-mediated osteogenesis is associated with its hormone-like function. It can induce BMP-2 and BMP-4 mRNA expression in osteoblasts and significantly upregulates Osx at low doses [97, 98]. In addition, ICA facilitates bone formation by inducing proosteoblastic genes like Osx, Runx2, alkaline phosphatase (ALP), and collagen type I. It also inhibits bone resorption by regulating the osteoprotegerin/receptor activator of nuclear factor-κb ligand (OPG/RANKL) signaling in the osteoclasts [99]. Zhang et al. found that ICA inhibits the adipogenic differentiation of BMSCs and promotes osteoblastic differentiation [100]. Fan et al. found that ICA promoted not only BMSC proliferation in vitro in a dose-dependent manner but also osteoblastic differentiation at very low doses (10−9 M to 10−6 M). However, a higher concentration of 10−5 M was toxic and suppressed osteoblastic differentiation [30]. Using a rat model of bone fracture, Cao et al. [2] showed that intragastric administration of ICA significantly increased osteotylus formation and accelerated bone healing within 5 months of treatment. These findings demonstrate that ICA administration following bone fracture can accelerate mineralization and osteogenesis and significantly improve bone healing. Therefore, ICA can also be an alternative treatment for postmenopausal osteoporosis and bone fracture.
The imbalance between BMSC adipogenesis and osteogenesis is considered the primary cause of femoral head necrosis [101]. The two processes are normally at an equilibrium under physiological conditions, which can be disrupted by external factors such as steroids and alcohol. Huang et al. [4] showed that ICA can effectively prevent femoral head necrosis, improve prednisolone-induced BMSC proliferation, enhance osteoblastic differentiation, and inhibit adipogenic differentiation. In addition, low concentrations of ICA (10−9 M to 10−5 M) significantly increased BMSC proliferation, especially at 10−6 M [4, 30].
Sun et al. [29] found that ICA restored the balance between osteogenic and adipogenic differentiation of mesenchymal stem cells in patients with osteonecrosis of the femoral head via ATP-binding cassette subfamily B member 1- (ABCB1-) promoter demethylation. In addition, ICA inhibited the differentiation of mesenchymal stem cells into adipocytes by inhibiting PPARγ, CCAAT/enhancer binding protein α (C/EBPα), and fatty acid-binding protein 4 (FABP4) mRNA via the Notch signaling pathway [102]. Zheng et al. [103] also found that daily oral administration of ICA (0.3 mg/g) to osteoprotegerin knockout male mice for 8 weeks increased the trabecular bone volume and trabecular number, indicating an important role of osteoprotegerin in ICA-mediated osteogenic effects. In addition, osteocalcin and osteopontin also mediate ICA-induced osteogenic differentiation by increasing ALP activity and collagen type I levels [104]. These results [29, 102–104] indicated that ICA plays an important role in bone synthesis and metabolism. Furthermore, ICA significantly promoted bone healing by increasing BMSC proliferation and osteoblastic differentiation in a New Zealand rabbit model of bone defect [105]. ICA can also induce BMSC osteoblastic differentiation under various pathological conditions such as osteoporosis [5] and bone necrosis [29]. Estrogen and epigenetic regulation are currently the research focus of ICA-induced osteogenesis under pathological conditions [5, 29]. Sun et al. showed that ICA improved BMSC viability and osteoblastic differentiation by upregulating ABCB1, indicating a demethylating function as well [29]. In addition to promoting osteogenic differentiation of BMSCs, ICA can also promote bone regeneration by promoting angiogenesis [106], since vascularization is a key step in bone regeneration which recruits the BMSCs and essential factors to the site of trauma [107].
Icaritin is a biologically active metabolite of ICA [108] and can be easily extracted from various sources. Wu et al. [12] showed positive effects of icaritin on BMSC osteoblastic differentiation in vitro. It promotes osteogenic differentiation and inhibits adipogenic differentiation of BMSCs by inactivating GSK-3β and suppressing PPARγ expression [102, 109, 110]. In addition, the BMPs (BMP-2, BMP-4, and BMP-7) and the MAPK/ERK pathway are also involved in icaritin-mediated osteogenic differentiation [12, 110].
Taken together, ICA promotes BMSC proliferation and osteoblastic differentiation and inhibits adipogenic differentiation, indicating its potential as a bone regenerative drug.
4.2. Signaling Pathways Involved in ICA-Mediated BMSC Proliferation and Osteogenesis
The MAPK pathway consists of the ERK, p38 kinase (p38), and Jun amino-terminal kinases/stress-activated protein kinase (JNK) factors. It regulates essential cellular functions, such as growth, proliferation, differentiation, and apoptosis. In addition, MAPKs also mediate the biological functions of ICA [111], indicating a possible role in BMSC proliferation as well. Qin et al. [112] found that ICA-induced rat BMSC proliferation in vitro was positively correlated with ERK levels and p38 phosphorylation and significantly upregulated Elk1 and c-Myc, the transcription factors downstream of the MAPK pathway.
A study using BMSCs extracted from SD rat bone marrow showed that 0.05-2.0 mg/l ICA significantly facilitated BMSC proliferation by activating the Wnt/β-catenin pathway [113]. Ye et al. [114] found that low doses of ICA (10−8 M to 10−6 M) promoted the proliferation and osteoblastic differentiation of rat adipose-derived stem cells (ADSCs), and 10−7 M ICA significantly upregulated RhoA (ras homolog gene family, member A) and p-MYPT1 (a ROCK or Rho-associated protein kinase substrate). This indicates that ICA promotes rat ADSC proliferation and osteoblastic differentiation by activating the RhoA-transcriptional coactivator with the PDZ-binding motif (TAZ) signaling pathway. Furthermore, Zhai et al. [115] showed the involvement of the PI3K/Akt/eNOS/NO/cGMP/PKG signaling pathway in the ICA-mediated osteogenesis of BMSCs. As already mentioned, any imbalance between BMSC osteoblastic and adipogenic differentiation impairs bone stability and leads to bone loss and increased bone marrow adipogenesis [116], resulting in osteoporosis and bone necrosis [117, 118]. TAZ is a β-catenin-like transcriptional coactivator involved in modulating this balance [119, 120]. It activates Runx2-mediated transcription to regulate BMSC differentiation and stimulate osteoblastic differentiation and also interacts with PPARγ to suppress adipogenic differentiation. Furthermore, Wei et al. [121] demonstrated that ICA promotes BMSC proliferation and osteogenesis via activation of the estrogen receptor (ER)α-Wnt/β-catenin signaling pathway. There is considerable ambiguity regarding the interaction between TAZ and Wnt/β-catenin. One study [122] indicated an upstream regulatory role of TAZ, while another study [123] showed that TAZ lies downstream of the Wnt/β-catenin cascade. Nevertheless, TAZ is an important regulator of ICA-mediated BMSC osteoblastic differentiation.
Kammerer et al. [124] reported that the ERα signaling pathway transcriptionally regulates Runx2, while Cai et al. [125] showed that the Wnt/β-catenin pathway directly targeted Runx2 to promote osteoblastic differentiation and the calcification of vascular smooth muscle cells. Both studies indicated a close association of the ERα and Wnt/β-catenin signaling pathways with the Runx2 expression. Another study [126] found that ICA stimulated BMSC osteoblastic differentiation by upregulating TAZ and the downstream osteogenic genes, and blocking the aforementioned pathways abrogated ICA-induced TAZ upregulation (Figure 3). These findings point to a TAZ/ERα/Wnt/β-catenin axis that mediates ICA-induced BMSC osteoblastic differentiation. In one study, Wu et al. [104] demonstrated the involvement of the JNK pathway in the osteoblastic differentiation of BMSCs or periodontal ligament stem cells [127, 128].
Figure 3.

Signaling pathways involved in ICA-mediated BMSC proliferation, osteogenesis, angiogenesis, and migration. ICA promotes proliferation of BMSCs through signaling pathways such as MAPK, ERK, p38, Wnt/β-catenin, RhoA-TAZ, and ERα-Wnt/β-catenin. ICA promotes BMSC osteogenesis via signaling pathways such as RhoA-TAZ, JNK, Wnt/β-catenin, ERα-Wnt/β-catenin, and PI3K/Akt/eNOS/NO/cGMP/PKG. ICA promotes BMSC angiogenesis via PI3K/Akt/eNOS, EGF-EGFR, PI3K, and ERK1/2 signaling pathways. ICA promotes migration of BMSCs through the MAPK signaling pathway.
Multiple signaling pathways, including the BMP, nitric oxide (NO), MAPK, and Wnt/β-catenin pathways, are likely activated in the osteoblasts due to the estrogen-like properties of ICA and ICA-induced estrogen production [90, 129, 130]. Shi et al. [131] showed that ICA promoted osteogenesis in rat cranial osteoblasts and in an in vivo rat model of bone growth by activating the cAMP/PKA/CREB signaling pathway.
4.3. ICA Promotes BMSC Migration and Angiogenesis
ICA not only activates endothelial angiogenesis in vitro but also directly stimulates angiogenesis in vivo, through the PI3K/Akt/eNOS-dependent signaling pathways [106]. ICA can activate the epidermal growth factor-epidermal growth factor receptor (EGF-EGFR) pathway to promote endothelial NOS synthesis, thereby facilitating vascular regeneration [132]. In addition, ICA can directly stimulate angiogenesis by activating various angiogenic factors like ERK, PI3K, and Akt [133, 134]. An in vitro study by Liu et al. [135] showed that ICA upregulated angiogenesis-related genes like vascular endothelial growth factor (VEGF) and fibroblast growth factors (FGF). In addition, ICA upregulated brain-derived neurotrophic factor (BDNF) and VEGF via the PI3K and ERK1/2 signaling pathways [134], which in turn promoted the angiogenic differentiation of BMSCs. Furthermore, Jiao et al. [136] found that ICA enhanced the migratory ability of BMSCs in vitro and in vivo, via the MAPK signaling pathway.
In summary, the primary effects of ICA in BMSCs are to promote proliferation and osteogenesis and are mediated by multiple signaling pathways including the MAPK/ERK/p38, Wnt/β-catenin, PI3K/Akt/eNOS/NO/cGMP/PKG, RhoA-TAZ, and ERα-Wnt/β-catenin pathways. In addition, ICA can also act on osteoblasts through the BMP/Runx2, NO, MAPK, Wnt/β-catenin, cAMP/PKA/CREB, and JNK pathways. Furthermore, ICA promotes angiogenesis via the PI3K, ERK1/2, and EGF-EGFR pathways and BMSC migration via the MAPK pathway. The angiogenic effect of ICA is favorable for osteogenesis, although their exact relationship as well as that between angiogenesis and migration still needs to be elucidated.
5. Prospects
ICA can significantly promote BMSC proliferation and osteoblastic differentiation and inhibit adipogenic differentiation, making it a reliable option for bone regenerative therapy. Mechanistic studies show that multiple signaling pathways mediate these processes, indicating the potential of multiple therapeutic targets. Above all, ICA could be made into a liquid state at suitable concentration in the future and be applied in bone regeneration. Besides, the evidence indicated that the optimal concentration for ICA which can perform better effects in BMSCs is 1 μM [4, 30], while others reported 0.1 μM [114, 121, 126]. However, further studies are needed to figure out both a safe and an effective concentration of ICA [14]. For patients with fractures, bone defects, nonunion disorders, and osteonecrosis of the femoral head, a mixture of ICA and autologous BMSCs can be injected locally into the lesion to facilitate bone regeneration. For patients with osteoporosis, ICA can be delivered through intravenous administration. Since ALP (an early marker of osteogenic differentiation) levels peak on the 14th day of the in vitro BMSC culture with ICA [121, 126, 135], it is reasonable to consider 14 days of intravenous ICA administration for the treatment regimen. ICA also promotes the regeneration of periodontal tissue [137], peripheral nerves [138], neural stem cells [139], and endometrium [140], although the optimal concentration of ICA differs across tissues.
There are still several questions that need to be addressed in future studies. For example, although the pathways involved in BMSC migration are well-known, the mechanism(s) underlying ICA-mediated BMSC migration remain to be elucidated. At present, it is not clear whether there is a synergistic or antagonistic crosstalk or upstream and downstream relationship among the signaling pathways involved in ICA-mediated osteogenic differentiation of BMSCs. Most studies on BMSC osteogenesis and migration have been carried out under normoxic conditions. The oxygen levels in ischemic lesions, such as in femoral head necrosis, can be less than 1% [141], and severe hypoxia affects the osteogenic differentiation and migration of BMSCs in vivo and in vitro [142, 143]. Therefore, it is necessary to simulate the hypoxic conditions in the in vitro studies.
In conclusion, a better understanding of the role and mechanism of action of ICA in BMSCs can provide new therapeutic strategies for various orthopedic disorders such as osteoporosis, femoral head necrosis, and bone defects.
Acknowledgments
This study was supported by the 6th National Famous and Experienced Chinese Medicine Experts' Academic Inheritance Project funded by the Chinese Government's State Administration of Traditional Chinese Medicine (No. 292017), the National Natural Science Foundation of China (No. 81472103), the “HuangheYingcai” Project of Wuhan City of China, and the Health and Family Planning Commission of Wuhan Municipality Research Foundation (WX18M01).
Abbreviations
- BMSCs:
Bone marrow mesenchymal stem cells
- ICA:
Icariin
- IL-6:
Interleukin-6
- TNF-1:
Tumor necrosis factor
- PPARγ:
Peroxisomal proliferator-activated receptor gamma
- Runx2:
Runt-related transcription factor 2
- BMP-2:
Bone morphogenetic protein-2
- ERK-MAPK:
Extracellular signal-regulated kinase-mitogen-activated protein kinase
- ALP:
Alkaline phosphatase
- OPG/RANKL:
Osteoprotegerin/receptor activator of nuclear factor-κb ligand
- C/EBPα:
CCAAT/enhancer-binding protein α
- FABP4:
Fatty acid-binding protein 4
- PDGF:
Platelet-derived growth factor
- CXCL12:
C-X-C motif chemokine ligand 12
- CXCR4:
C-X-C chemokine receptor type 4
- SDF-1:
Stromal-derived factor 1
- Smad:
Drosophila mothers against decapentaplegic protein
- Twist1:
Twist-related protein 1, also known as class A basic helix-loop-helix protein 38 (bHLHa38)
- Atf4:
Activating transcription factor 4
- TGF-β/BMP:
Transforming growth factor-β/bone morphogenetic protein
- PI3K/Akt/GSK-3:
Phosphatidylinositol 3-kinase/protein kinase B/glycogen synthase kinase-3
- ERK:
Extracellular signal-regulated kinase
- IGF1:
Insulin-like growth factor 1
- Osx:
Osterix
- EGF-EGFR:
Epidermal growth factor-epidermal growth factor receptor
- NO:
Nitric oxide
- VEGF:
Vascular endothelial growth factor
- FGF:
Fibroblast growth factors
- MAPK:
Mitogen-activated protein kinase
- p38:
p38 kinase
- JNK:
Jun amino-terminal kinases/stress-activated protein kinase
- RhoA:
Ras homolog gene family, member A
- p-MYPT1:
A ROCK (Rho-associated protein kinase) substrate molecule
- ERα:
Estrogen receptor α
- PI3K/Akt/eNOS/NO/cGMP/PKG:
Phosphatidylinositol 3-kinase/protein kinase B/endothelial nitric oxide synthase/nitric oxide/cyclic guanosine monophosphate/protein kinase-G
- cAMP/PKA/CREB:
cAMP/protein kinase A/cAMP response element-binding protein
- GSK-3:
Glycogen synthase kinase-3
- ABCB1:
ATP-binding cassette subfamily B member 1
- TAZ:
Transcriptional coactivator with PDZ-binding motif
- BDNF:
Brain-derived neurotrophic factor.
Disclosure
The funders had no role in the decision to publish or in the preparation of the manuscript.
Conflicts of Interest
The authors declare that they have no competing interests.
Authors' Contributions
Aofei Yang and Chaochao Yu contributed equally to this manuscript. Qilin Lu and Hao Li helped illustrate the figures. Zhanghua Li and Chengjian He contributed to the conception and helped revise the manuscript.
References
- 1.Algarni A. D., Al Moallem H. M. Clinical and radiological outcomes of extracorporeal shock wave therapy in early-stage femoral head osteonecrosis. Advances in Orthopedics. 2018;2018:6. doi: 10.1155/2018/7410246.7410246 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 2.Cao H., Zhang Y., Qian W., et al. Effect of icariin on fracture healing in an ovariectomized rat model of osteoporosis. Experimental and Therapeutic Medicine. 2017;13(5):2399–2404. doi: 10.3892/etm.2017.4233. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 3.Dailiana Z. H., Stefanou N., Khaldi L., et al. Vascular endothelial growth factor for the treatment of femoral head osteonecrosis: an experimental study in canines. World Journal of Orthopedics. 2018;9(9):120–129. doi: 10.5312/wjo.v9.i9.120. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 4.Huang Z., Cheng C., Cao B., et al. Icariin protects against glucocorticoid-induced osteonecrosis of the femoral head in rats. Cellular Physiology and Biochemistry. 2018;47(2):694–706. doi: 10.1159/000490023. [DOI] [PubMed] [Google Scholar]
- 5.Luo Z., Liu M., Sun L., Rui F. Icariin recovers the osteogenic differentiation and bone formation of bone marrow stromal cells from a rat model of estrogen deficiency-induced osteoporosis. Molecular Medicine Reports. 2015;12(1):382–388. doi: 10.3892/mmr.2015.3369. [DOI] [PubMed] [Google Scholar]
- 6.Moya-Angeler J., Gianakos A. L., Villa J. C., Ni A., Lane J. M. Current concepts on osteonecrosis of the femoral head. World Journal of Orthopedics. 2015;6(8):590–601. doi: 10.5312/wjo.v6.i8.590. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 7.Sanghani-Kerai A., McCreary D., Lancashire H., Osagie L., Coathup M., Blunn G. Stem cell interventions for bone healing: fractures and osteoporosis. Current Stem Cell Research & Therapy. 2018;13(5):369–377. doi: 10.2174/1574888X13666180410160511. [DOI] [PubMed] [Google Scholar]
- 8.Jin Y. Z., Lee J. H. Mesenchymal stem cell therapy for bone regeneration. Clinics in Orthopedic Surgery. 2018;10(3):271–278. doi: 10.4055/cios.2018.10.3.271. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 9.Song F., Jiang D., Wang T., et al. Mechanical stress regulates osteogenesis and adipogenesis of rat mesenchymal stem cells through PI3K/Akt/GSK-3β/β-catenin signaling pathway. BioMed Research International. 2017;2017:10. doi: 10.1155/2017/6027402.6027402 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 10.Heydari Asl S., Hosseinpoor H., Parivar K., Hayati Roodbari N., Hanaee-Ahvaz H. Physical stimulation and scaffold composition efficiently support osteogenic differentiation of mesenchymal stem cells. Tissue & Cell. 2018;50:1–7. doi: 10.1016/j.tice.2017.11.001. [DOI] [PubMed] [Google Scholar]
- 11.Yan Z., Guo Y., Wang Y., Li Y., Wang J. MicroRNA profiles of BMSCs induced into osteoblasts with osteoinductive medium. Experimental and Therapeutic Medicine. 2018;15(3):2589–2596. doi: 10.3892/etm.2018.5723. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 12.Wu T., Shu T., Kang L., et al. Icaritin, a novel plant-derived osteoinductive agent, enhances the osteogenic differentiation of human bone marrow- and human adipose tissue-derived mesenchymal stem cells. International Journal of Molecular Medicine. 2017;39(4):984–992. doi: 10.3892/ijmm.2017.2906. [DOI] [PubMed] [Google Scholar]
- 13.Lin A. X., Chan G., Hu Y., et al. Internationalization of traditional Chinese medicine: current international market, internationalization challenges and prospective suggestions. Chinese Medicine. 2018;13(1):p. 9. doi: 10.1186/s13020-018-0167-z. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 14.Wang Z., Wang D., Yang D., Zhen W., Zhang J., Peng S. The effect of icariin on bone metabolism and its potential clinical application. Osteoporosis International. 2018;29(3):535–544. doi: 10.1007/s00198-017-4255-1. [DOI] [PubMed] [Google Scholar]
- 15.Liu Y., Zuo H., Liu X., Xiong J., Pei X. The antiosteoporosis effect of icariin in ovariectomized rats: a systematic review and meta-analysis. Cellular and Molecular Biology. 2017;63(11):124–131. doi: 10.14715/cmb/2017.63.11.22. [DOI] [PubMed] [Google Scholar]
- 16.Huang H., Liang M., Zhang X., Zhang C., Shen Z., Zhang W. Simultaneous determination of nine flavonoids and qualitative evaluation of Herba Epimedii by high performance liquid chromatography with ultraviolet detection. Journal of Separation Science. 2007;30(18):3207–3213. doi: 10.1002/jssc.200700262. [DOI] [PubMed] [Google Scholar]
- 17.Zhou Z., Luo J., Wang J., Li L., Kong L. Simultaneous enrichment and separation of flavonoids from Herba Epimedii by macroporous resins coupled with preparative chromatographic method. Natural Product Research. 2015;29(2):185–188. doi: 10.1080/14786419.2014.964704. [DOI] [PubMed] [Google Scholar]
- 18.Chen M., Wu J., Luo Q., et al. The anticancer properties of Herba Epimedii and its main bioactive componentsicariin and icariside II. Nutrients. 2016;8(9) doi: 10.3390/nu8090563. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 19.Wang N., Huang X., Wang X., Zhang Y., Wu R., Shou D. Pipette tip solid-phase extraction and high-performance liquid chromatography for the determination of flavonoids from Epimedii herba in rat serum and application of the technique to pharmacokinetic studies. Journal of Chromatography B. 2015;990:64–72. doi: 10.1016/j.jchromb.2015.03.012. [DOI] [PubMed] [Google Scholar]
- 20.Sun X., Li Q., Zhang J., et al. The reason leading to the increase of icariin in Herba Epimedii by heating process. Journal of Pharmaceutical and Biomedical Analysis. 2018;149:525–531. doi: 10.1016/j.jpba.2017.11.050. [DOI] [PubMed] [Google Scholar]
- 21.Zhang H. F., Yang T. S., Li Z. Z., Wang Y. Simultaneous extraction of epimedin A, B, C and icariin from Herba Epimedii by ultrasonic technique. Ultrasonics Sonochemistry. 2008;15(4):376–385. doi: 10.1016/j.ultsonch.2007.09.002. [DOI] [PubMed] [Google Scholar]
- 22.Wang N., Xin H., Zhang Q., et al. Carbon nanotube-polymer composite for effervescent pipette tip solid phase microextraction of alkaloids and flavonoids from Epimedii herba in biological samples. Talanta. 2017;162:10–18. doi: 10.1016/j.talanta.2016.09.059. [DOI] [PubMed] [Google Scholar]
- 23.Chen X. J., Ji H., Zhang Q. W., et al. A rapid method for simultaneous determination of 15 flavonoids in Epimedium using pressurized liquid extraction and ultra-performance liquid chromatography. Journal of Pharmaceutical and Biomedical Analysis. 2008;46(2):226–235. doi: 10.1016/j.jpba.2007.09.016. [DOI] [PubMed] [Google Scholar]
- 24.Li H. B., Chen F. Separation and purification of epimedin A, B, C, and icariin from the medicinal herb Epimedium brevicornum Maxim by dual-mode HSCCC. Journal of Chromatographic Science. 2009;47(5):337–340. doi: 10.1093/chromsci/47.5.337. [DOI] [PubMed] [Google Scholar]
- 25.Xie F., Wu C. F., Lai W. P., et al. The osteoprotective effect of Herba Epimedii (HEP) extract in vivo and in vitro. Evidence-Based Complementary and Alternative Medicine. 2005;2(3):361. doi: 10.1093/ecam/neh101. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 26.Wang L., Li Y., Guo Y., et al. Herba Epimedii: an ancient Chinese herbal medicine in the prevention and treatment of osteoporosis. Current Pharmaceutical Design. 2016;22(3):328–349. doi: 10.2174/1381612822666151112145907. [DOI] [PubMed] [Google Scholar]
- 27.Zhang G., Qin L., Shi Y. Epimedium-derived phytoestrogen flavonoids exert beneficial effect on preventing bone loss in late postmenopausal women: a 24-month randomized, double-blind and placebo-controlled trial. Journal of Bone and Mineral Research. 2007;22(7):1072–1079. doi: 10.1359/jbmr.070405. [DOI] [PubMed] [Google Scholar]
- 28.Lu M., Wang L. H., Luo Y. W., et al. Treatment of primary osteoporosis with epimedium total flavone capsule: a multicenter clinical observation on 360 cases. Chinese Journal of Osteoporosis. 2013;19(3):279–282. [Google Scholar]
- 29.Sun Z. B., Wang J. W., Xiao H., et al. Icariin may benefit the mesenchymal stem cells of patients with steroid-associated osteonecrosis by ABCB1-promoter demethylation: a preliminary study. Osteoporosis International. 2015;26(1):187–197. doi: 10.1007/s00198-014-2809-z. [DOI] [PubMed] [Google Scholar]
- 30.Fan J. J., Cao L. G., Wu T., et al. The dose-effect of icariin on the proliferation and osteogenic differentiation of human bone mesenchymal stem cells. Molecules. 2011;16(12):10123–10133. doi: 10.3390/molecules161210123. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 31.Sreejit P., Dilip K. B., Verma R. S. Generation of mesenchymal stem cell lines from murine bone marrow. Cell and Tissue Research. 2012;350(1):55–68. doi: 10.1007/s00441-012-1458-9. [DOI] [PubMed] [Google Scholar]
- 32.Li C. J., Xiao Y., Yang M., et al. Long noncoding RNA Bmncr regulates mesenchymal stem cell fate during skeletal aging. The Journal of Clinical Investigation. 2018;128(12):5251–5266. doi: 10.1172/JCI99044. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 33.Li C. J., Cheng P., Liang M. K., et al. MicroRNA-188 regulates age-related switch between osteoblast and adipocyte differentiation. The Journal of Clinical Investigation. 2015;125(4):1509–1522. doi: 10.1172/JCI77716. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 34.Zhang W., Liu N., Shi H., et al. Upregulation of BMSCs osteogenesis by positively-charged tertiary amines on polymeric implants via charge/iNOS signaling pathway. Scientific Reports. 2015;5(1, article 9369) doi: 10.1038/srep09369. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 35.Blanco J. F., García-Briñon J., Benito-Garzón L., Pescador D., Muntión S., Sánchez-Guijo F. Human bone marrow mesenchymal stromal cells promote bone regeneration in a xenogeneic rabbit model: a preclinical study. Stem Cells International. 2018;2018:10. doi: 10.1155/2018/7089484.7089484 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 36.Capulli M., Paone R., Rucci N. Osteoblast and osteocyte: games without frontiers. Archives of Biochemistry and Biophysics. 2014;561:3–12. doi: 10.1016/j.abb.2014.05.003. [DOI] [PubMed] [Google Scholar]
- 37.Franz-Odendaal T. A., Hall B. K., Witten P. E. Buried alive: how osteoblasts become osteocytes. Developmental Dynamics. 2006;235(1):176–190. doi: 10.1002/dvdy.20603. [DOI] [PubMed] [Google Scholar]
- 38.Florencio-Silva R., Sasso G. R. . S., Sasso-Cerri E., Simões M. J., Cerri P. S. Biology of bone tissue: structure, function, and factors that influence bone cells. BioMed Research International. 2015;2015:17. doi: 10.1155/2015/421746.421746 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 39.Huang S., Eleniste P. P., Wayakanon K., et al. The Rho-GEF Kalirin regulates bone mass and the function of osteoblasts and osteoclasts. Bone. 2014;60:235–245. doi: 10.1016/j.bone.2013.12.023. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 40.Wu Q., Zhou X., Huang D., JI Y., Kang F. IL-6 enhances osteocyte-mediated osteoclastogenesis by promoting JAK2 and RANKL activity in vitro. Cellular Physiology and Biochemistry. 2017;41(4):1360–1369. doi: 10.1159/000465455. [DOI] [PubMed] [Google Scholar]
- 41.Teramachi J., Zhou H., Subler M. A., et al. Increased IL-6 expression in osteoclasts is necessary but not sufficient for the development of Paget’s disease of bone. Journal of Bone and Mineral Research. 2014;29(6):1456–1465. doi: 10.1002/jbmr.2158. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 42.Cheung W. Y., Simmons C. A., You L. Osteocyte apoptosis regulates osteoclast precursor adhesion via osteocytic IL-6 secretion and endothelial ICAM-1 expression. Bone. 2012;50(1):104–110. doi: 10.1016/j.bone.2011.09.052. [DOI] [PubMed] [Google Scholar]
- 43.Feng W., Liu H., Luo T., et al. Combination of IL-6 and sIL-6R differentially regulate varying levels of RANKL-induced osteoclastogenesis through NF-κB, ERK and JNK signaling pathways. Scientific Reports. 2017;7(1, article 41411) doi: 10.1038/srep41411. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 44.Bustamante M., Nogués X., Enjuanes A., et al. COL1A1, ESR1, VDR and TGFB1 polymorphisms and haplotypes in relation to BMD in Spanish postmenopausal women. Osteoporosis International. 2007;18(2):235–243. doi: 10.1007/s00198-006-0225-8. [DOI] [PubMed] [Google Scholar]
- 45.Grigoriadis A. E., Heersche J. N., Aubin J. E. Differentiation of muscle, fat, cartilage, and bone from progenitor cells present in a bone-derived clonal cell population: effect of dexamethasone. The Journal of Cell Biology. 1988;106(6):2139–2151. doi: 10.1083/jcb.106.6.2139. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 46.Ducy P., Zhang R., Geoffroy V., Ridall A. L., Karsenty G. Osf2/Cbfa1: a transcriptional activator of osteoblast differentiation. Cell. 1997;89(5):747–754. doi: 10.1016/S0092-8674(00)80257-3. [DOI] [PubMed] [Google Scholar]
- 47.Wong S. P., Rowley J. E., Redpath A. N., Tilman J. D., Fellous T. G., Johnson J. R. Pericytes, mesenchymal stem cells and their contributions to tissue repair. Pharmacology & Therapeutics. 2015;151:107–120. doi: 10.1016/j.pharmthera.2015.03.006. [DOI] [PubMed] [Google Scholar]
- 48.Majumdar M. K., Thiede M. A., Mosca J. D., Moorman M., Gerson S. L. Phenotypic and functional comparison of cultures of marrow-derived mesenchymal stem cells (MSCs) and stromal cells. Journal of Cellular Physiology. 1998;176(1):57–66. doi: 10.1002/(SICI)1097-4652(199807)176:1<57::AID-JCP7>3.0.CO;2-7. [DOI] [PubMed] [Google Scholar]
- 49.Matsumoto T., Kano K., Kondo D., et al. Mature adipocyte-derived dedifferentiated fat cells exhibit multilineage potential. Journal of Cellular Physiology. 2008;215(1):210–222. doi: 10.1002/jcp.21304. [DOI] [PubMed] [Google Scholar]
- 50.Lu C., Gao S., Xu G. Geraniin inhibits TNF-α-induced impairments of osteogenesis through NF-κB and p38 MAPK signalling pathways in bone marrow stem cells. BMJ. 2017;2(2):47–52. doi: 10.1136/svn-2016-000046. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 51.Pittenger M. F., Mackay A. M., Beck S. C., et al. Multilineage potential of adult human mesenchymal stem cells. Science. 1999;284(5411):143–147. doi: 10.1126/science.284.5411.143. [DOI] [PubMed] [Google Scholar]
- 52.Chen S. L., Fang W. W., Ye F., et al. Effect on left ventricular function of intracoronary transplantation of autologous bone marrow mesenchymal stem cell in patients with acute myocardial infarction. The American Journal of Cardiology. 2004;94(1):92–95. doi: 10.1016/j.amjcard.2004.03.034. [DOI] [PubMed] [Google Scholar]
- 53.Horwitz E. M., Prockop D. J., Fitzpatrick L. A., et al. Transplantability and therapeutic effects of bone marrow-derived mesenchymal cells in children with osteogenesis imperfecta. Nature Medicine. 1999;5(3):309–313. doi: 10.1038/6529. [DOI] [PubMed] [Google Scholar]
- 54.Horwitz E. M., Gordon P. L., Koo W. K. K., et al. Isolated allogeneic bone marrow-derived mesenchymal cells engraft and stimulate growth in children with osteogenesis imperfecta: implications for cell therapy of bone. Proceedings of the National Academy of Sciences of the United States of America. 2002;99(13):8932–8937. doi: 10.1073/pnas.132252399. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 55.Koç O. N., Day J., Nieder M., Gerson S. L., Lazarus H. M., Krivit W. Allogeneic mesenchymal stem cell infusion for treatment of metachromatic leukodystrophy (MLD) and Hurler syndrome (MPS-IH) Bone Marrow Transplantation. 2002;30(4):215–222. doi: 10.1038/sj.bmt.1703650. [DOI] [PubMed] [Google Scholar]
- 56.Strauer B. E., Brehm M., Zeus T., et al. Repair of infarcted myocardium by autologous intracoronary mononuclear bone marrow cell transplantation in humans. Circulation. 2002;106(15):1913–1918. doi: 10.1161/01.CIR.0000034046.87607.1C. [DOI] [PubMed] [Google Scholar]
- 57.Shen H., Wang Y., Zhang Z., Yang J., Hu S., Shen Z. Mesenchymal stem cells for cardiac regenerative therapy: optimization of cell differentiation strategy. Stem Cells International. 2015;2015:10. doi: 10.1155/2015/524756.524756 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 58.Mok S. K., Chen W. F., Lai W. P., et al. Icariin protects against bone loss induced by oestrogen deficiency and activates oestrogen receptor-dependent osteoblastic functions in UMR 106 cells. British Journal of Pharmacology. 2010;159(4):939–949. doi: 10.1111/j.1476-5381.2009.00593.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 59.Ming L. G., Chen K. M., Xian C. J. Functions and action mechanisms of flavonoids genistein and icariin in regulating bone remodeling. Journal of Cellular Physiology. 2013;228(3):513–521. doi: 10.1002/jcp.24158. [DOI] [PubMed] [Google Scholar]
- 60.Titorencu I., Pruna V., Jinga V. V., Simionescu M. Osteoblast ontogeny and implications for bone pathology: an overview. Cell and Tissue Research. 2014;355(1):23–33. doi: 10.1007/s00441-013-1750-3. [DOI] [PubMed] [Google Scholar]
- 61.Kokabu S., Lowery J. W., Jimi E. Cell fate and differentiation of bone marrow mesenchymal stem cells. Stem Cells International. 2016;2016:7. doi: 10.1155/2016/3753581.3753581 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 62.Yang A., Yu C., You F., He C., Li Z. Mechanisms of Zuogui pill in treating osteoporosis: perspective from bone marrow mesenchymal stem cells. Evidence-Based Complementary and Alternative Medicine. 2018;2018:8. doi: 10.1155/2018/3717391.3717391 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 63.Justesen J., Stenderup K., Ebbesen E. N., Mosekilde L., Steiniche T., Kassem M. Adipocyte tissue volume in bone marrow is increased with aging and in patients with osteoporosis. Biogerontology. 2001;2(3):165–171. doi: 10.1023/A:1011513223894. [DOI] [PubMed] [Google Scholar]
- 64.Miller M., Shuman J. D., Sebastian T., Dauter Z., Johnson P. F. Structural basis for DNA recognition by the basic region leucine zipper transcription factor CCAAT/enhancer-binding protein α. Journal of Biological Chemistry. 2003;278(17):15178–15184. doi: 10.1074/jbc.M300417200. [DOI] [PubMed] [Google Scholar]
- 65.Tontonoz P., Hu E., Spiegelman B. M. Stimulation of adipogenesis in fibroblasts by PPARγ2, a lipid-activated transcription factor. Cell. 1994;79(7):1147–1156. doi: 10.1016/0092-8674(94)90006-X. [DOI] [PubMed] [Google Scholar]
- 66.Peng S., Zhou G., Luk K. D. K., et al. Strontium promotes osteogenic differentiation of mesenchymal stem cells through the Ras/MAPK signaling pathway. Cellular Physiology and Biochemistry. 2009;23(1-3):165–174. doi: 10.1159/000204105. [DOI] [PubMed] [Google Scholar]
- 67.James A. W. Review of signaling pathways governing MSC osteogenic and adipogenic differentiation. Scientifica. 2013;2013:17. doi: 10.1155/2013/684736.684736 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 68.Valenti M. T., Garbin U., Pasini A., et al. Role of ox-PAPCs in the differentiation of mesenchymal stem cells (MSCs) and Runx2 and PPARγ2 expression in MSCs-like of osteoporotic patients. PLoS One. 2011;6(6, article e20363) doi: 10.1371/journal.pone.0020363. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 69.Li X., Cui Q., Kao C., Wang G. J., Balian G. Lovastatin inhibits adipogenic and stimulates osteogenic differentiation by suppressing PPARγ2 and increasing Cbfa1/Runx2 expression in bone marrow mesenchymal cell cultures. Bone. 2003;33(4):652–659. doi: 10.1016/S8756-3282(03)00239-4. [DOI] [PubMed] [Google Scholar]
- 70.Mei Y., Bian C., Li J., et al. miR-21 modulates the ERK-MAPK signaling pathway by regulating SPRY2 expression during human mesenchymal stem cell differentiation. Journal of Cellular Biochemistry. 2013;114(6):1374–1384. doi: 10.1002/jcb.24479. [DOI] [PubMed] [Google Scholar]
- 71.Goff L. A., Boucher S., Ricupero C. L., et al. Differentiating human multipotent mesenchymal stromal cells regulate microRNAs: prediction of microRNA regulation by PDGF during osteogenesis. Experimental Hematology. 2008;36(10):1354–1369.e2. doi: 10.1016/j.exphem.2008.05.004. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 72.Cao J., Wei Y., Lian J., et al. Notch signaling pathway promotes osteogenic differentiation of mesenchymal stem cells by enhancing BMP9/Smad signaling. International Journal of Molecular Medicine. 2017;40(2):378–388. doi: 10.3892/ijmm.2017.3037. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 73.Li X., Guo L., Liu Y., et al. MicroRNA-21 promotes osteogenesis of bone marrow mesenchymal stem cells via the Smad7-Smad1/5/8-Runx2 pathway. Biochemical and Biophysical Research Communications. 2017;493(2):928–933. doi: 10.1016/j.bbrc.2017.09.119. [DOI] [PubMed] [Google Scholar]
- 74.Long H., Sun B., Cheng L., et al. miR-139-5p represses BMSC osteogenesis via targeting Wnt/β-catenin signaling pathway. DNA and Cell Biology. 2017;36(8):715–724. doi: 10.1089/dna.2017.3657. [DOI] [PubMed] [Google Scholar]
- 75.Sen B., Guilluy C., Xie Z., et al. Mechanically induced focal adhesion assembly amplifies anti-adipogenic pathways in mesenchymal stem cells. Stem Cells. 2011;29(11):1829–1836. doi: 10.1002/stem.732. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 76.Tahimic C. G. T., Long R. K., Kubota T., et al. Regulation of ligand and shear stress-induced insulin-like growth factor 1 (IGF1) signaling by the integrin pathway. Journal of Biological Chemistry. 2016;291(15):8140–8149. doi: 10.1074/jbc.M115.693598. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 77.Su P., Tian Y., Yang C., et al. Mesenchymal stem cell migration during bone formation and bone diseases therapy. International Journal of Molecular Sciences. 2018;19(8, article 2343) doi: 10.3390/ijms19082343. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 78.Li Z., Wang W., Xu H., et al. Effects of altered CXCL12/CXCR4 axis on BMP2/Smad/Runx2/Osterix axis and osteogenic gene expressions during osteogenic differentiation of MSCs. American Journal of Translational Research. 2017;9(4):1680–1693. [PMC free article] [PubMed] [Google Scholar]
- 79.Zhang S. J., Song X. Y., He M., Yu S. B. Effect of TGF-β1/SDF-1/CXCR4 signal on BM-MSCs homing in rat heart of ischemia/perfusion injury. European Review for Medical and Pharmacological Sciences. 2016;20(5):899–905. [PubMed] [Google Scholar]
- 80.Abduelmula A., Huang R., Pu Q., Tamamura H., Morosan-Puopolo G., Brand-Saberi B. SDF-1 controls the muscle and blood vessel formation of the somite. The International Journal of Developmental Biology. 2016;60(1-2-3):29–38. doi: 10.1387/ijdb.150132rh. [DOI] [PubMed] [Google Scholar]
- 81.Zhang J. F., Fu W. M., He M. L., et al. MiRNA-20a promotes osteogenic differentiation of human mesenchymal stem cells by co-regulating BMP signaling. RNA Biology. 2011;8(5):829–838. doi: 10.4161/rna.8.5.16043. [DOI] [PubMed] [Google Scholar]
- 82.Sanghani A., Osagie-Clouard L., Samizadeh S., et al. CXCR4 has the potential to enhance bone formation in osteopenic rats. Tissue Engineering. Part A. 2018;24(23-24):1775–1783. doi: 10.1089/ten.tea.2018.0121. [DOI] [PubMed] [Google Scholar]
- 83.Tan H. B., Giannoudis P. V., Boxall S. A., McGonagle D., Jones E. The systemic influence of platelet-derived growth factors on bone marrow mesenchymal stem cells in fracture patients. BMC Medicine. 2015;13(1):p. 6. doi: 10.1186/s12916-014-0202-6. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 84.Li A., Xia X., Yeh J., et al. PDGF-AA promotes osteogenic differentiation and migration of mesenchymal stem cell by down-regulating PDGFRα and derepressing BMP-Smad1/5/8 signaling. PLoS One. 2014;9(12, article e113785) doi: 10.1371/journal.pone.0113785. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 85.Sanghani-Kerai A., Osagie-Clouard L., Blunn G., Coathup M. The influence of age and osteoporosis on bone marrow stem cells from rats. Bone & Joint Research. 2018;7(4):289–297. doi: 10.1302/2046-3758.74.BJR-2017-0302.R1. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 86.Sanghani-Kerai A., Coathup M., Samazideh S., et al. Osteoporosis and ageing affects the migration of stem cells and this is ameliorated by transfection with CXCR4. Bone & Joint Research. 2017;6(6):358–365. doi: 10.1302/2046-3758.66.BJR-2016-0259.R1. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 87.Ho C. Y., Sanghani A., Hua J., Coathup M., Kalia P., Blunn G. Mesenchymal stem cells with increased stromal cell-derived factor 1 expression enhanced fracture healing. Tissue Engineering Part A. 2015;21(3-4):594–602. doi: 10.1089/ten.tea.2013.0762. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 88.Tang Y., Wu X., Lei W., et al. TGF-β1–induced migration of bone mesenchymal stem cells couples bone resorption with formation. Nature Medicine. 2009;15(7):757–765. doi: 10.1038/nm.1979. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 89.Wang Z., Zhang X., Wang H., Qi L., Lou Y. Neuroprotective effects of icaritin against beta amyloid-induced neurotoxicity in primary cultured rat neuronal cells via estrogen-dependent pathway. Neuroscience. 2007;145(3):911–922. doi: 10.1016/j.neuroscience.2006.12.059. [DOI] [PubMed] [Google Scholar]
- 90.Yang L., Lu D., Guo J., Meng X., Zhang G., Wang F. Icariin from Epimedium brevicornum Maxim promotes the biosynthesis of estrogen by aromatase (CYP19) Journal of Ethnopharmacology. 2013;145(3):715–721. doi: 10.1016/j.jep.2012.11.031. [DOI] [PubMed] [Google Scholar]
- 91.Li S., Dong P., Wang J., et al. Icariin, a natural flavonol glycoside, induces apoptosis in human hepatoma SMMC-7721 cells via a ROS/JNK-dependent mitochondrial pathway. Cancer Letters. 2010;298(2):222–230. doi: 10.1016/j.canlet.2010.07.009. [DOI] [PubMed] [Google Scholar]
- 92.Zhou J., Wu J., Chen X., et al. Icariin and its derivative, ICT, exert anti-inflammatory, anti-tumor effects, and modulate myeloid derived suppressive cells (MDSCs) functions. International Immunopharmacology. 2011;11(7):890–898. doi: 10.1016/j.intimp.2011.01.007. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 93.Guo J., Li F., Wu Q., Gong Q., Lu Y., Shi J. Protective effects of icariin on brain dysfunction induced by lipopolysaccharide in rats. Phytomedicine. 2010;17(12):950–955. doi: 10.1016/j.phymed.2010.03.007. [DOI] [PubMed] [Google Scholar]
- 94.Sze S. C. W., Tong Y., Ng T. B., Cheng C. L. Y., Cheung H. P. Herba Epimedii: anti-oxidative properties and its medical implications. Molecules. 2010;15(11):7861–7870. doi: 10.3390/molecules15117861. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 95.Qin L., Han T., Zhang Q., et al. Antiosteoporotic chemical constituents from Er-Xian decoction, a traditional Chinese herbal formula. Journal of Ethnopharmacology. 2008;118(2):271–279. doi: 10.1016/j.jep.2008.04.009. [DOI] [PubMed] [Google Scholar]
- 96.Ye H. Y., Lou Y. J. Estrogenic effects of two derivatives of icariin on human breast cancer MCF-7 cells. Phytomedicine. 2005;12(10):735–741. doi: 10.1016/j.phymed.2004.10.002. [DOI] [PubMed] [Google Scholar]
- 97.Zhao J., Ohba S., Shinkai M., Chung U. I., Nagamune T. Icariin induces osteogenic differentiation in vitro in a BMP- and Runx2-dependent manner. Biochemical and Biophysical Research Communications. 2008;369(2):444–448. doi: 10.1016/j.bbrc.2008.02.054. [DOI] [PubMed] [Google Scholar]
- 98.Yin X. X., Chen Z. Q., Liu Z. J., Ma Q. J., Dang G. T. Icariine stimulates proliferation and differentiation of human osteoblasts by increasing production of bone morphogenetic protein 2. Chinese Medical Journal. 2007;120(3):204–210. doi: 10.1097/00029330-200702010-00006. [DOI] [PubMed] [Google Scholar]
- 99.Ma X. N., Ge B. F., Chen K. M., et al. Mechanisms of icariin in regulating bone formation of osteoblasts and bone resorption of osteoclasts. Zhongguo Yi Xue Ke Xue Yuan Xue Bao. 2013;35(4):432–438. doi: 10.3881/j.issn.1000-503X.2013.04.014. [DOI] [PubMed] [Google Scholar]
- 100.Zhang S., Feng P., Mo G., et al. Icariin influences adipogenic differentiation of stem cells affected by osteoblast-osteoclast co-culture and clinical research adipogenic. Biomedicine & Pharmacotherapy. 2017;88:436–442. doi: 10.1016/j.biopha.2017.01.050. [DOI] [PubMed] [Google Scholar]
- 101.Li X., Jin L., Cui Q., Wang G. J., Balian G. Steroid effects on osteogenesis through mesenchymal cell gene expression. Osteoporosis International. 2005;16(1):101–108. doi: 10.1007/s00198-004-1649-7. [DOI] [PubMed] [Google Scholar]
- 102.Liu H., Xiong Y., Zhu X., et al. Icariin improves osteoporosis, inhibits the expression of PPARγ, C/EBPα, FABP4 mRNA, N1ICD and jagged1 proteins, and increases Notch2 mRNA in ovariectomized rats. Experimental and Therapeutic Medicine. 2017;13(4):1360–1368. doi: 10.3892/etm.2017.4128. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 103.Zheng D., Peng S., Yang S. H., et al. The beneficial effect of icariin on bone is diminished in osteoprotegerin-deficient mice. Bone. 2012;51(1):85–92. doi: 10.1016/j.bone.2012.04.006. [DOI] [PubMed] [Google Scholar]
- 104.Wu Y., Xia L., Zhou Y., Xu Y., Jiang X. Icariin induces osteogenic differentiation of bone mesenchymal stem cells in a MAPK-dependent manner. Cell Proliferation. 2015;48(3):375–384. doi: 10.1111/cpr.12185. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 105.Fan J., Bi L., Wu T., et al. A combined chitosan/nano-size hydroxyapatite system for the controlled release of icariin. Journal of Materials Science. Materials in Medicine. 2012;23(2):399–407. doi: 10.1007/s10856-011-4491-4. [DOI] [PubMed] [Google Scholar]
- 106.Chung B. H., Kim J. D., Kim C. K., et al. Icariin stimulates angiogenesis by activating the MEK/ERK- and PI3K/Akt/eNOS-dependent signal pathways in human endothelial cells. Biochemical and Biophysical Research Communications. 2008;376(2):404–408. doi: 10.1016/j.bbrc.2008.09.001. [DOI] [PubMed] [Google Scholar]
- 107.Wernike E., Montjovent M. O., Liu Y., et al. VEGF incorporated into calcium phosphate ceramics promotes vascularisation and bone formation in vivo. European Cells & Materials. 2010;19:30–40. doi: 10.22203/ecm.v019a04. [DOI] [PubMed] [Google Scholar]
- 108.Lim R. Z. L., Li L., Yong E. L., Chew N. STAT-3 regulation of CXCR4 is necessary for the prenylflavonoid Icaritin to enhance mesenchymal stem cell proliferation, migration and osteogenic differentiation. Biochimica et Biophysica Acta (BBA) - General Subjects. 2018;1862(7):1680–1692. doi: 10.1016/j.bbagen.2018.04.016. [DOI] [PubMed] [Google Scholar]
- 109.Sheng H., Rui X. F., Sheng C. J., et al. A novel semisynthetic molecule icaritin stimulates osteogenic differentiation and inhibits adipogenesis of mesenchymal stem cells. International Journal of Medical Sciences. 2013;10(6):782–789. doi: 10.7150/ijms.6084. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 110.Luo G., Xu B., Wang W., Wu Y., Li M. Study of the osteogenesis effect of icariside II and icaritin on canine bone marrow mesenchymal stem cells. Journal of Bone and Mineral Metabolism. 2018;36(6):668–678. doi: 10.1007/s00774-017-0889-5. [DOI] [PubMed] [Google Scholar]
- 111.Ding L., Liang X. G., Zhu D. Y., Lou Y. J. Icariin promotes expression of PGC-1α, PPARα, and NRF-1 during cardiomyocyte differentiation of murine embryonic stem cells in vitro. Acta Pharmacologica Sinica. 2007;28(10):1541–1549. doi: 10.1111/j.1745-7254.2007.00648.x. [DOI] [PubMed] [Google Scholar]
- 112.Qin S., Zhou W., Liu S., Chen P., Wu H. Icariin stimulates the proliferation of rat bone mesenchymal stem cells via ERK and p38 MAPK signaling. International Journal of Clinical and Experimental Medicine. 2015;8(5):7125–7133. [PMC free article] [PubMed] [Google Scholar]
- 113.Fu S., Yang L., Hong H., Zhang R. Wnt/β-catenin signaling is involved in the icariin induced proliferation of bone marrow mesenchymal stem cells. Journal of Traditional Chinese Medicine. 2016;36(3):360–368. doi: 10.1016/s0254-6272(16)30050-4. [DOI] [PubMed] [Google Scholar]
- 114.Ye Y., Jing X., Li N., Wu Y., Li B., Xu T. Icariin promotes proliferation and osteogenic differentiation of rat adipose-derived stem cells by activating the RhoA-TAZ signaling pathway. Biomedicine & Pharmacotherapy. 2017;88:384–394. doi: 10.1016/j.biopha.2017.01.075. [DOI] [PubMed] [Google Scholar]
- 115.Zhai Y. K., Guo X. Y., Ge B. F., et al. Icariin stimulates the osteogenic differentiation of rat bone marrow stromal cells via activating the PI3K-AKT-eNOS-NO-cGMP-PKG. Bone. 2014;66:189–198. doi: 10.1016/j.bone.2014.06.016. [DOI] [PubMed] [Google Scholar]
- 116.Xue P., Wu X., Zhou L., et al. IGF1 promotes osteogenic differentiation of mesenchymal stem cells derived from rat bone marrow by increasing TAZ expression. Biochemical and Biophysical Research Communications. 2013;433(2):226–231. doi: 10.1016/j.bbrc.2013.02.088. [DOI] [PubMed] [Google Scholar]
- 117.Astudillo P., Rios S., Pastenes L., Pino A. M., Rodriguez J. P. Increased adipogenesis of osteoporotic human-mesenchymal stem cells (MSCs) characterizes by impaired leptin action. Journal of Cellular Biochemistry. 2008;103(4):1054–1065. doi: 10.1002/jcb.21516. [DOI] [PubMed] [Google Scholar]
- 118.Suh K. T., Kim S. W., Roh H. L., Youn M. S., Jung J. S. Decreased osteogenic differentiation of mesenchymal stem cells in alcohol-induced osteonecrosis. Clinical Orthopaedics and Related Research. 2005;(431):220–225. doi: 10.1097/01.blo.0000150568.16133.3c. [DOI] [PubMed] [Google Scholar]
- 119.Byun M. R., Kim A. R., Hwang J. H., et al. Phorbaketal A stimulates osteoblast differentiation through TAZ mediated Runx2 activation. FEBS Letters. 2012;586(8):1086–1092. doi: 10.1016/j.febslet.2012.03.008. [DOI] [PubMed] [Google Scholar]
- 120.Byun M. R., Lee C. H., Hwang J. H., et al. Phorbaketal A inhibits adipogenic differentiation through the suppression of PPARγ-mediated gene transcription by TAZ. European Journal of Pharmacology. 2013;718(1-3):181–187. doi: 10.1016/j.ejphar.2013.08.035. [DOI] [PubMed] [Google Scholar]
- 121.Wei Q., Zhang J., Hong G. J., et al. Icariin promotes osteogenic differentiation of rat bone marrow stromal cells by activating the ERα-Wnt/β-catenin signaling pathway. Biomedicine & Pharmacotherapy. 2016;84:931–939. doi: 10.1016/j.biopha.2016.09.107. [DOI] [PubMed] [Google Scholar]
- 122.Varelas X., Miller B. W., Sopko R., et al. The Hippo pathway regulates Wnt/β-catenin signaling. Developmental Cell. 2010;18(4):579–591. doi: 10.1016/j.devcel.2010.03.007. [DOI] [PubMed] [Google Scholar]
- 123.Azzolin L., Zanconato F., Bresolin S., et al. Role of TAZ as mediator of Wnt signaling. Cell. 2012;151(7):1443–1456. doi: 10.1016/j.cell.2012.11.027. [DOI] [PubMed] [Google Scholar]
- 124.Kammerer M., Gutzwiller S., Stauffer D., Delhon I., Seltenmeyer Y., Fournier B. Estrogen receptor α (ERα) and estrogen related receptor α (ERRα) are both transcriptional regulators of the Runx2-I isoform. Molecular and Cellular Endocrinology. 2013;369(1-2):150–160. doi: 10.1016/j.mce.2013.01.024. [DOI] [PubMed] [Google Scholar]
- 125.Cai T., Sun D., Duan Y., et al. WNT/β-catenin signaling promotes VSMCs to osteogenic transdifferentiation and calcification through directly modulating Runx2 gene expression. Experimental Cell Research. 2016;345(2):206–217. doi: 10.1016/j.yexcr.2016.06.007. [DOI] [PubMed] [Google Scholar]
- 126.Wei Q., He M., Chen M., et al. Icariin stimulates osteogenic differentiation of rat bone marrow stromal stem cells by increasing TAZ expression. Biomedicine & Pharmacotherapy. 2017;91:581–589. doi: 10.1016/j.biopha.2017.04.019. [DOI] [PubMed] [Google Scholar]
- 127.Wang Y., Li J., Song W., Yu J. Mineral trioxide aggregate upregulates odonto/osteogenic capacity of bone marrow stromal cells from craniofacial bones via JNK and ERK MAPK signalling pathways. Cell Proliferation. 2014;47(3):241–248. doi: 10.1111/cpr.12099. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 128.Yu Y., Mu J., Fan Z., et al. Insulin-like growth factor 1 enhances the proliferation and osteogenic differentiation of human periodontal ligament stem cells via ERK and JNK MAPK pathways. Histochemistry and Cell Biology. 2012;137(4):513–525. doi: 10.1007/s00418-011-0908-x. [DOI] [PubMed] [Google Scholar]
- 129.Zhang Z. B., Yang Q. T. The testosterone mimetic properties of icariin. Asian Journal of Andrology. 2006;8(5):601–605. doi: 10.1111/j.1745-7262.2006.00197.x. [DOI] [PubMed] [Google Scholar]
- 130.Liu J., Ye H., Lou Y. Determination of rat urinary metabolites of icariin in vivo and estrogenic activities of its metabolites on MCF-7 cells. Die Pharmazie. 2005;60(2):120–125. [PubMed] [Google Scholar]
- 131.Shi W., Gao Y., Wang Y., et al. The flavonol glycoside icariin promotes bone formation in growing rats by activating the cAMP signaling pathway in primary cilia of osteoblasts. The Journal of Biological Chemistry. 2017;292(51):20883–20896. doi: 10.1074/jbc.M117.809517. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 132.Liu T., Qin X. C., Li W. R., et al. Effects of icariin and icariside II on eNOS expression and NOS activity in porcine aorta endothelial cells. Beijing Da Xue Xue Bao Yi Xue Ban. 2011;43(4):500–504. [PubMed] [Google Scholar]
- 133.Xu H. B., Huang Z. Q. Icariin enhances endothelial nitric-oxide synthase expression on human endothelial cells in vitro. Vascular Pharmacology. 2007;47(1):18–24. doi: 10.1016/j.vph.2007.03.002. [DOI] [PubMed] [Google Scholar]
- 134.Liu D., Ye Y., Xu L., Yuan W., Zhang Q. Icariin and mesenchymal stem cells synergistically promote angiogenesis and neurogenesis after cerebral ischemia via PI3K and ERK1/2 pathways. Biomedicine & Pharmacotherapy. 2018;108:663–669. doi: 10.1016/j.biopha.2018.09.071. [DOI] [PubMed] [Google Scholar]
- 135.Liu T., Zhang X., Luo Y., Huang Y., Wu G. Slowly delivered icariin/allogeneic bone marrow-derived mesenchymal stem cells to promote the healing of calvarial critical-size bone defects. Stem Cells International. 2016;2016:13. doi: 10.1155/2016/1416047.1416047 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 136.Jiao F., Tang W., Huang H., et al. Icariin promotes the migration of BMSCs in vitro and in vivo via the MAPK signaling pathway. Stem Cells International. 2018;2018:9. doi: 10.1155/2018/2562105.2562105 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 137.Zhang X., Han N., Li G., et al. Local icariin application enhanced periodontal tissue regeneration and relieved local inflammation in a minipig model of periodontitis. International Journal of Oral Science. 2018;10(2):p. 19. doi: 10.1038/s41368-018-0020-3. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 138.Chen B., Niu S. P., Wang Z. Y., et al. Local administration of icariin contributes to peripheral nerve regeneration and functional recovery. Neural Regeneration Research. 2015;10(1):84–89. doi: 10.4103/1673-5374.150711. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 139.Fu X., Li S., Zhou S., Wu Q., Jin F., Shi J. Stimulatory effect of icariin on the proliferation of neural stem cells from rat hippocampus. BMC Complementary and Alternative Medicine. 2018;18(1):p. 34. doi: 10.1186/s12906-018-2095-y. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 140.Le A. W., Wang Z. H., Dai X. Y., et al. An experimental study on the use of icariin for improving thickness of thin endometrium. Genetics and Molecular Research. 2017;16(1) doi: 10.4238/gmr16019126. [DOI] [PubMed] [Google Scholar]
- 141.Lewis J. S., Lee J. A., Underwood J. C. E., Harris A. L., Lewis C. E. Macrophage responses to hypoxia: relevance to disease mechanisms. Journal of Leukocyte Biology. 1999;66(6):889–900. doi: 10.1002/jlb.66.6.889. [DOI] [PubMed] [Google Scholar]
- 142.Xu N., Liu H., Qu F., et al. Hypoxia inhibits the differentiation of mesenchymal stem cells into osteoblasts by activation of Notch signaling. Experimental and Molecular Pathology. 2013;94(1):33–39. doi: 10.1016/j.yexmp.2012.08.003. [DOI] [PubMed] [Google Scholar]
- 143.Benjamin S., Sheyn D., Ben-David S., et al. Oxygenated environment enhances both stem cell survival and osteogenic differentiation. Tissue Engineering Part A. 2013;19(5-6):748–758. doi: 10.1089/ten.tea.2012.0298. [DOI] [PubMed] [Google Scholar]