

Abstract
Geniposide, an iridoid glucoside, is a major component in the fruit of Gardenia jasminoides Ellis (Gardenia fruits). Geniposide has been experimentally proved to possess multiple pharmacological actions involving antioxidative stress, anti-inflammatory, antiapoptosis, antiangiogenesis, antiendoplasmic reticulum stress (ERS), etc. In vitro and in vivo studies have further identified the value of geniposide in a spectrum of preclinical models of diabetes mellitus (DM) and cardiovascular disorders. The antioxidative property of geniposide should be attributed to the result of either the inhibition of numerous pathological processes or the activation of various proteins associated with cell survival or a combination of both. In this review, we will summarize the available knowledge on the antioxidative property and protective effects of geniposide in DM and cardiovascular disease in the literature and discuss antioxidant mechanisms as well as its potential applications in clinic.
1. Introduction
In the early stage of modern medicine development, biologically active compounds extracted from plants have played a crucial role in providing medicines to combat diseases. Medicines extracted from plants continue to account for a significant niche in the treatment of diseases all over the world [1]. Naturally derived compounds have fewer reported advice effects than allopathic medicine, indicating that it could be safer to use for a longer period of time. Geniposide (Figure 1(a)) is one such plant-derived medicine that has been traditionally used as a folk medicine for hundreds of years in Asian countries.
Figure 1.

Chemical structure of geniposide (a) and genipin (b).
Geniposide is one of the natural components extracted with ethanol from the fruit of Gardenia jasminoides Ellis, the concentration of which in Gardenia jasminoides Ellis is 73.44 ± 2.62 mg/g. As a major iridoid glycoside and glucagon-like peptide-1 (GLP-1) receptor agonist, geniposide is supposed to exert various biological effects of the herbs [2–4]. Genipin (Figure 1(b)) is the primary metabolite and aglycon of geniposide. Previous studies have revealed that genipin is transformed from geniposide by the intestinal microflora enzymes in the bowel, which indicates that the major form of geniposide in circulating blood may be genipin [5–7]. Until now, geniposide has been used extensively for the reason that it rarely causes severe toxic effects [8]; meanwhile, it has various pharmacological properties including antioxidative stress [9], anti-inflammatory [5], antiapoptosis [10], antithrombus [11], antiendoplasmic reticulum stress [12], and antifungi [13]. Geniposide exhibits a number of merits as a promising therapeutic agent due to its properties and is currently in preclinical models for a variety of conditions like neurological disorders [14], liver diseases [15], diabetes mellitus (DM) [16], cardiovascular disorders [17], tumors [18], and other conditions.
DM and cardiovascular disease are growing at epidemic proportions giving rise to heavy social, economic, and health burden. Although many efforts and advances have been made, the current therapeutic approaches for the treatment of DM and cardiovascular disease are still far from satisfactory, principally because of serious adverse effects [19]. Hence, some novel therapeutic candidates are now continually being explored to obtain better outcomes. In the last decades, a considerable attention has been paid on functional properties of natural compounds derived from plants for their potential therapeutic purposes. The antioxidative property and potential effects in DM and cardiovascular disease of geniposide make it become an attractive candidate for the treatment, particularly in type2 DM and cardiomyopathy. In this review, we aim to highlight relevant experimental researches performed in terms of pharmacokinetic properties, the antioxidative property, and therapeutic effects of geniposide in DM and cardiovascular disease.
2. Pharmacokinetics
Extensive studies to elucidate the pharmacokinetic properties of geniposide have been performed on different species including rats and humans. Here, we will illustrate its properties according to absorption, distribution, metabolism, and excretion.
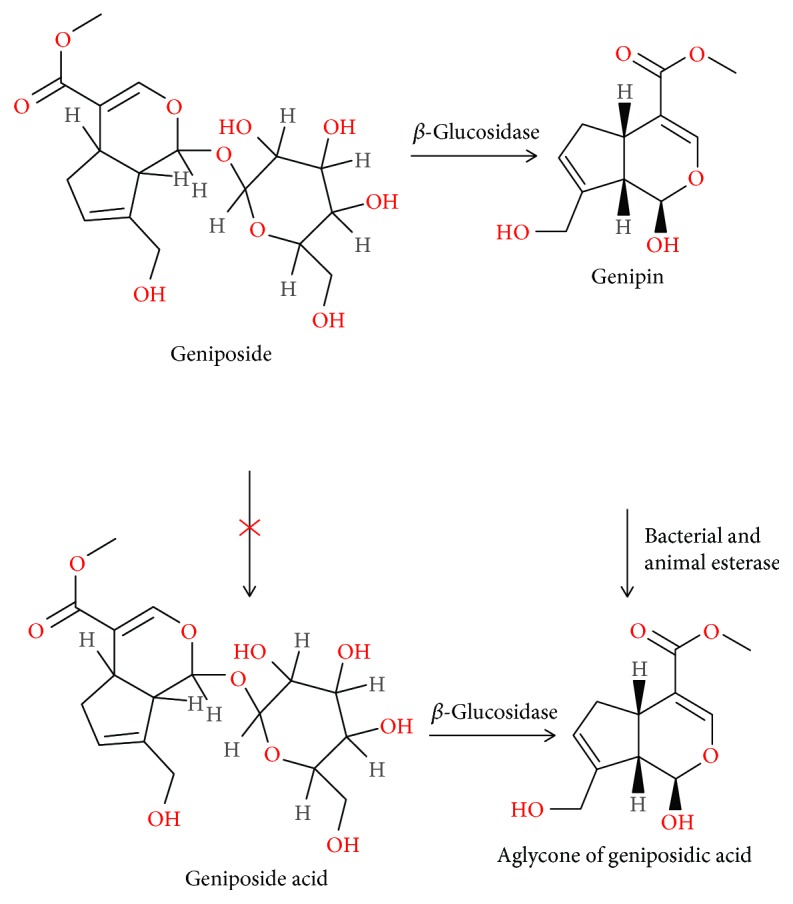
Geniposide exhibited excellent choleretic effects when it was administrated orally or intraduodenally. However, this effect was blocked when injected intraportally. By contrast, genipin, which is the aglycone of geniposide, possessed favorable choleretic effects by either intraportal or intraduodenal administration. Furthermore, in vitro and in vivo studies have demonstrated that geniposide could be transformed to genipin by the intestinal microflora enzymes (β-glucosidase) in the bowel. This indicates that geniposide has no activity as a cholagogue by itself unless it is transformed to another active metabolite, genipin. In the presence of ammonia, genipin could be further transformed to a new nitrogen-containing compound, genipinine. Despite that both geniposide and genipin contain methyl ester, which seem to be hydrolyzed to genoposidic acid, only genipin in fact could be hydrolyzed to the aglycone of genoposidic acid by bacterial and animal esterase [20]. A total of 24 species of human bacteria in intestinal tract could help geniposide transform into genipin (Table 1). Although intestinal bacteria could produce five types of β-glucosidase, it seems that only one fraction from Eubacterium sp. A-44 and Ruminococcus sp. PO1-3 is able to hydrolyze geniposide to genipin. Notably, esterase from Eubacterium sp. A-44 as well as pig liver could not hydrolyze geniposide to geniposide acid directly, but could hydrolyze genipin to geniposide acid [21]. Figure 2 gives a schematic diagram of metabolic progress of geniposide by β-glucosidase and esterase. Yu et al. [22] discovered that the stability of geniposide in different pH K-R solutions did not alter, which revealed that the acidic conditions may have no direct influence on the stability of geniposide. Geniposide was not stable in duodenum perfusions but stable in jejunum, ileum, and colon perfusions, the difference of which may be caused by the intestinal contents. However, after adjusting the pH values of perfusions to 7.0, geniposide was observed to be stable in different regions of the intestine. These two phenomena indicate that the acidic conditions may affect geniposide indirectly through affecting the intestinal contents but not influence the stability of geniposide directly. Adding verapamil, one of the well-known inhibitors of P-glycoprotein (P-gp) could significantly elevate the absorption of geniposide 2.4-fold, but the absorption was not changed by EDTA, one of the typical inhibitors of cell membrane transport channels, which hinted that the transportation of geniposide might be associated with P-gp rather than membrane transport channels. Intriguingly, another traditional medicine, notoginsenoside R1, at 0.1 and 0.2 mg/mL concentrations was able to significantly enhance the intestinal absorption of geniposide (1.424 mg/mL) by 1.7- and 1.4-fold in a fashion similar to that of a P-gp inhibitor [23]. Likewise, P-gp is also the efflux transport of genipin. And the intestinal absorption rate and permeability value of genipin were dramatically enhanced by the genipin/hydroxypropyl-β-cyclodextrin inclusion complex owing to the solubilizing effect and P-gp inhibitory effect of hydroxypropyl-β-cyclodextrin [24].
Table 1.
Amounts of genipin transformed from geniposide by incubation with 24 strains of human intestinal bacteria. Each tube contained 30 mg geniposide.
| Strain | Genipin (mg/tube) | Strain | Genipin (mg/tube) |
|---|---|---|---|
| Klebsiella pneumoniae ATCC 13883 | 3.8 | Bifidobacterium adolescentis | 1.2 |
| Bacteroides fragilis ssp. thetaotus | 2.0 | Lactobacillus acidophilus ATCC 4356 | 6.6 |
| L. brevis II-46 | 1.4 | B. bifidum | 0.3 |
| L. fermentum ATCC 9338 | 6.9 | B. breve S-2 KZ 1287 | 2.4 |
| B. pseudolongum PNC-2-9-G | 1.8 | L. xylosus ATCC 155775 | 6.2 |
| L. plantarum ATCC 14917 | 5.3 | B. longum | 0.9 |
| Clostridium butyricum | 0.7 | Peptostreptococcus anaerobius | 1.4 |
| C. innocuum KZ-633 | 1.2 | Proteus mirabilis S2 | 3.3 |
| C. perfringens To-23 | 0.3 | C. innocuum ES 24-06 | 0.3 |
| P. intermedium | 0.9 | Fusobacterium nucleatum | 3.8 |
| Escherichia coli 0-127 | 5.3 | Ruminococcus sp. PO1-3 | 2.9 |
| Streptococcus faecalis II-136 | 2.5 | Veillonella parvula ssp. parvula | 0.4 |
In the meantime, each individual bacterial suspension was incubated for 24 hours anaerobically in 100 mM phosphate buffer (pH 7.3) at 37°C.
Figure 2.
Schematic diagram of the metabolic process of geniposide by β-glucosidase and esterase [2].
Using a high-performance liquid chromatography (HPLC) with a column-switching system, the simultaneous concentration of geniposide and genipin in mouse plasma after oral administration of 1200 mg/kg hot-water extract Gardenia fruits (1200 mg extract contained 216.4 mg geniposide, while the genipin in extract was a trace amount) was detected after 15 min. The results demonstrated that the concentration of geniposide in plasma peaked (approximately 103.1 μg/mL) at 30 min and reduced gradually. In contrast, the amount of genipin was too small to detect, which seemed to reach a peak (approximate 0.07 μg/mL) at 60 min. This suggested that a large amount of geniposide went into the circulating blood while only a very small amount of genipin could go into the blood after being hydrolyzed for the reason that genipin was so labile a substance that it could easily react with proteins and amino acids [25]. In particular, the oral bioavailability of geniposide in rats was obviously improved when Gardenia jasminoides Ellis was combined with Cortex magnoliae officinalis and Fructus aurantii immaturus. Because of the herb-herb interaction, the pharmacokinetic process of geniposide from Chinese medicinal formula was more complicated and unpredicted. Hence, geniposide in some certain formula should be paid closer attention compared with its single use [26].
In a model of anesthetized rats with intravenous administration [27], geniposide displayed a dose-dependent manner in circulating blood within the dosage range of 10-100 mg/kg. In the dosage of 10 mg/kg, the area under concentration-time curve (AUC) of geniposide in circulating blood was significantly smaller than that in bile, indicating that geniposide was mainly excreted via hepatoenteral circulation. Intriguingly, acupuncture treatment of specific acupuncture points for the liver and bladder channels had no obvious influence on the pharmacokinetics of geniposide in rat blood, bile, and liver although cumulative studies showed that acupuncture or electroacupuncture treatments may interact with drugs in terms of pharmacokinetics [28]. Furthermore, the distribution of geniposide in rats was also studied. After oral administration of geniposide, the maximum concentration (Cmax) emerged at 30 min in the spleen and liver, at 120 min in the kidney and brain. Among all the tissues and organs investigated, kidney was the organ which possessed the highest concentration of geniposide. The AUC0→4h values after the administration of geniposide were according to the order of brain <lung<heart<liver<spleen<kidney [29].
The metabolic process of geniposide is complicated and remains unclear. Using online liquid chromatography-mass spectrometry (LC-MS) data acquisition and off-line data processing methods involving extracted ion chromatogram, multiple mass defect filters, fragment ion searching, and isotope pattern filtering, a total of thirty-three different metabolites of geniposide were captured in the rat liver, urine, and plasma. Among these metabolites, six were in rat heart, twelve in liver, six in brain, six in lung, three in spleen, twelve in kidney, and four in rat liver microsomes. However, only six of them may exert pharmacological actions. The corresponding metabolic reactions of geniposide involved hydroxylation, hydrolysis, taurine conjugation, demethylation, hydrogenation, decarboxylation, cysteine S-conjugation dehydration, methylation, glucosylation, sulfate conjugation, and their composite reactions [24]. Additionally, the metabolism of geniposide in immune tissues of rats with adjuvant arthritis was also investigated. Rats with adjuvant arthritis were divided into three groups and given geniposide intragastrically at 120, 66, and 33 mg/kg, respectively. Subsequently, the biotransformation of geniposide in different organs was detected and a total of four major metabolites were identified, namely G1, G2, G3, and G4. The distribution of geniposide and its metabolites is shown in Table 2 [26]. Taking plasma for example, after oral administration of geniposide, the parameters in pharmacokinetics of rats with or without adjuvant arthritis exhibited great differences. AUC0→∞, the mean residence time (MRT), the clearance from the total body (CL), and the apparent volume of distribution in adjuvant arthritis groups were significantly higher than those in the normal group, suggesting that the pharmacokinetics of geniposide was altered under the condition of adjuvant arthritis [30].
Table 2.
The distribution of geniposide and its metabolites in rats with adjuvant arthritis.
| Plasma | Spleen | Urine | Liver | Synovium | Mesenteric lymph node | |
|---|---|---|---|---|---|---|
| Geniposide | + | + | + | + | + | + |
| G1 | + | — | + | — | — | — |
| G2 | + | + | + | — | — | + |
| G3 | — | + | — | — | — | — |
| G4 | — | + | + | — | — | — |
G1, G2, G3, and G4 are four major metabolites of geniposide.
Apart from rapid absorption into the circulating blood, wide distribution in tissues and organs, and complicated metabolism, geniposide could be also eliminated rapidly from the plasma within 12 h in rats [29]. Besides, the excretion of geniposide in the human body was also investigated. The Naoxuening injection (its major component is geniposide) was administrated intravenously, and the concentration of geniposide in urine was detected at various points in time. The results showed that geniposide was primarily excreted in the original form via the kidney. At 2 h, 4 h, and 10 h, the excretion level accounted for 52%, 63%, and over 90% of the initiative dosage [31].
Taken together, the pharmacokinetics of geniposide is still at the groping stage. Different experimental subjects, different administration routes, and different disease models may contribute to completely different outcomes. Geniposide predominately exists in the liver and kidney and is excreted in the original form via the kidney, which provides reliable evidence for its dosage regimen and pharmaceutics.
3. Effects of Geniposide on Cytochrome P450 and P-Glycoprotein
Although traditional medicines have become increasingly general all over the world, there are still some crucial possibilities for interaction between these traditional medicines and other allopathic medicines, which tends to steal the spotlight. A great many studies have uncovered that geniposide and genipin could affect the activities of enzymatic proteins and transporters in intestines and liver. Thus, the absorption and degradation of some certain drugs may give rise to altered plasma concentration of these drugs. For example, genipin has been proved to modulate the activity of cytochrome P450 (CYP450) 3A monooxygenase and P-gp [32]. CYP450 isoenzymes, which are mainly distributed in the liver and some extrahepatic tissues, play vital roles in the metabolism of drug oxidation reaction [33]. Inhibiting CYP450 isoenzymes may lead to target organ toxicity because they could increase exposure of these affected drugs. Therefore, the induction of xenobiotics to CYP450 isoenzymes is one of the main causes of therapeutic failure [34]. In human hepatoma HepG2 cells, genipin remarkably inhibited the level of mRNA and protein of CYP3A4 as well as CYP2C19, but significantly induced the expression of CYP2D6. To our knowledge, some traditional Chinese medicines involving Rhizoma coptidis [35] and Lotus leaf [36] have been shown to act as strong inhibitors of CYP2D6 [32]. This signifies that the target organ toxicity will be reduced and therapeutic effects will be enhanced when Gardenia jasminoides is combined with Rhizoma coptidis or Lotus leaf in traditional Chinese formulae. On the other hand, the metabolism of CYP2D6 could inhibit the pharmaceutical activity of some certain ingredients in traditional medicine, such as aconitine [37]. Similarly, in a rat model of nonalcoholic steatohepatitis, geniposide remarkably suppressed the expression of CYP2E1 [38]. Hence, we speculate that the combination with Gardenia jasminoides could expedite the degradation of aconitine and reduce its availability.
P-gp, one of the very famous members in the superfamily of ATP-binding cassette (ABC) transporters, was first characterized in multidrug-resistant Chinese hamster ovary (CHO) cells. P-gp is mainly coded by multidrug resistance gene 1 (MDR1) and plays a crucial part in eliminating xenotoxins against steep concentration gradients [39–41]. P-gp is widely distributed in various tissues and cells including the blood-brain barrier, intestinal epithelium, and tumor cells. The potential functions of P-gp in intestinal absorption as well as prehepatic elimination of a considerable amount of drugs have also been figured out [42, 43]. In doxorubicin-resistant cells, geniposide (50-100 μM) in a dose-dependent manner reversed P-gp-mediated MDR by inhibiting the expression of P-gp and its transport function [44]. Genipin has been also disclosed to be a substrate of P-gp and interact with the drug-binding site of P-gp competitively. Further study showed that the induction of genipin on P-gp by genipin is for the reason that it could strongly elevate the expression of MDR1 protein in hepatoma HepG2 cells via activating the basal P-gp ATPase [32]. From our perspective, the clinical application of genipin or geniposide with other drugs should be paid special attention, meantime the P-gp blockers should be also considered appropriately for the sake of the drug efficiency.
4. Toxicology
At the beginning, Yamano et al. disclosed that the crude extract of gardenia fruit, which was originally used as a kind of dye, could lead to hepatotoxicity in rats in both a time-dependent and dose-dependent manner after acute oral exposure [45]. Notably, intraperitoneal injection of geniposide did not trigger hepatotoxicity in rats but oral administration did. And oral administration of geniposide at a dose of 320 mg/kg or intraperitoneal injection of genipin at a dose of 80 mg/kg had similar effects, both of which could increase the activity of alanine transaminase (ALT) and aspartate aminotransferase (AST). In particular, the geniposide-induced hepatotoxicity was completely suppressed by chloramphenicol, indicating that chloramphenicol may block the hydrolysis of geniposide to genipin via modulating the microenvironment of intestinal microflora. These phenomena proved that it was genipin, not geniposide, that was likely to cause hepatotoxicity [46]. An acute toxicity study showed that the median lethal dose (LD50) of geniposide was 1431.1 mg/kg in rats. When the dose of geniposide in rats was over 574 mg/kg, hepatotoxicity could be triggered and the toxic effect usually emerged at 1-2 days after oral administration. A subchronic toxicity study on rats suggested that oral administration of geniposide at a normal dose of 24.3 mg/kg or less even in the repeated dosing for three months would not give rise to hepatotoxicity [47]. Another subchronic toxicity study revealed that 3-month ingestion of geniposide intake (60 mg/kg/day) did not cause any severe liver and kidney injury in rats [8]. However, in a 13-week toxicity study, some hematological parameters, the percentage of reticulocytes (Retic %), and the weight of some relative organs in male rats administered geniposide (100 mg/kg) significantly increased. In a 26-week toxicity study, the level of ALT, AST, electrolyte including potassium (K+) and sodium (Na+), red blood cell (RBC), white blood cell (WBC), haemoglobin (HGB) levels, Retic %, and the weight of liver and spleen in male rats also significantly increased [48]. The acute hepatotoxicity and nephrotoxicity in both jaundice rats induced by α-naphthylisothiocyanate and healthy rats were also evaluated. Geniposide at the dose of 120 mg/kg could lead to acute liver and kidney injury in rats and displayed a time-toxicity relationship [49]. Compared with the healthy group, the levels of ALT, AST, alkaline phosphatase (ALP), blood urea nitrogen, total bilirubin, and creatinine activity in rats administrated geniposide at the dose of 1.2 g/kg were statistically elevated at 48 h and 72 h [50]. In a rat model after intragastric administration of geniposide (300 mg/kg) for three days, ALT, AST, γ-glutamyltransferase (γ-GT), ALP, and total bile acids in serum were significantly elevated. Meanwhile, geniposide (300 mg/kg) also inhibited the expression of small heterodimer partner (SHP), farnesoid X receptor (FXR), and bile salt export pump (BSEP), suggesting that the dose-related liver injury of geniposide may be related with the abnormal expression of bile acid–regulating genes.
The mechanisms of hepatotoxicity from geniposide may be involved in oxidative stress with accumulation of malondialdehyde (MDA) concentration as well as reduction of total superoxide dismutase activity of livers in rats [47]. Moreover, candidate biomarkers for geniposide overdose-induced hepatotoxicity were investigated using a label-free quantitative proteomics approach. Five biomarkers in serum were identified, of which aldehyde dehydrogenase 1L1 (ALDH1L1), glycogen phosphorylase (PYGL), glycine N-methyltransferase (GNMT), and alanine-glyoxylate aminotransferase 2 (AGXT2) increased at the beginning of liver injury but then declined gradually; in contrast, α-enolase (ENO1) was found to increase consistently in liver injury. Noteworthily, GNMT and PYGL in serum appeared obviously earlier than did ALT and AST, two gold standard biomarkers in liver injury. Hence, these two novel biomarkers could be two indicators to predict early liver injury in the future [51].
5. The Main Properties of Geniposide
5.1. Anti-inflammatory Activity
Inflammation is not only a self-protection mechanism in the body to guarantee removal of the detrimental stimuli but also a healing process to repair damaged tissue, which is usually caused by various factors, like wound, microbial infection, and myocardial ischemia. However, it would be fetal for the human body if immune cells overproduce cytokines to overwhelm pathogens [52]. Thus, some plant-derived natural components with anti-inflammation effects but without obvious side effects will benefit people a lot.
Currently, geniposide has been proved to possess potent anti-inflammatory action under various pathological conditions. At the beginning, geniposide displayed a favorable anti-inflammation effect on paw edema induced by carrageenan. To be more specific, the edema paw volume gradually increased in the rats without administration of geniposide (up to 3 h). But in the group after injection of extraction from Gardenia fruit at doses of 50, 100, 200, and 400 mg/kg, it exhibited inhibiting effects of 10.2%, 25.9%, 28.6%, and 35.8% in acute paw edema at 3 h. Oral administration of geniposide at a dose of 100 mg/kg contributed to a stronger inhibiting effect of 31.7% at 3 h after edema [5]. However, this study just detected the phenotype of geniposide on paw edema but not explored the further molecular mechanisms.
Lipopolysaccharide (LPS) is the main component in the cell wall of Gram-negative bacteria. Recognizing LPS as well as its signal transduction is an important process of the body's own defense response. LPS is also a vital mechanism that causes endotoxic shock, systemic inflammatory response syndrome, and multiple organ failure. Interleukins (ILs) refer to the interaction of leukocytes or immune cells of lymphokines, which are similar cytokines with hematopoietic growth factor (HGF). The coordination and interaction of IL and HGF contribute to the hematopoietic and immunomodulatory functions. ILs display multiple physiological functions in transmitting information, activating and regulating immune cells, and mediating T and B cell activation, proliferation, and differentiation; in the meantime, they play important roles in the inflammatory response [53–55]. Geniposide remarkably suppressed the production of some inflammatory cytokines including IL-6 and IL-1β and tumor necrosis factor-α (TNF-α) in an in vitro model of primary mouse macrophages stimulated by LPS and an in vivo lung injury model induced by LPS through blocking the phosphorylation of inhibitory kappa B (IκBα) protein, p38, p65, extracellular signal-regulated kinase (ERK), c-Jun N-terminal kinase (JNK), and Toll-like receptor 4 (TLR4) [56, 57]. Meantime, in LPS-induced human umbilical vein endothelial cells (HUVECs), geniposide reduced the level of IL-6 and IL-8 effectively and synchronously blocked HUVEC migration and U937 monocyte adhesion to HUVECs, which was involved in the inactivation of ERK, p38MAPK, and nuclear factor-kappa B (NF-κB) signaling pathways [58]. Geniposide also significantly inhibited LPS-induced nitric oxide (NO), prostaglandin E2 (PGE2), and inflammatory cytokines by downregulating NF-κB, MAPK, and activator protein- (AP-) 1 signaling pathways [59].
Apart from being against the inflammation induced by LPS, geniposide also displays a favorable anti-inflammatory effect in the process of ischemia/reperfusion (I/R) injury in cerebral tissue through the activation of TLR4. To our knowledge, once the microglia are activated after I/R, it will become hypertrophic and proliferate rapidly, then migrate to the inflammatory region where they will produce a mass of proinflammatory and neurotoxic mediators, subsequently causing neuron injury [60]. Geniposide protected neurons from I/R injury in rats and mitigate oxygen-glucose deprivation-induced activation of microglial cells through decreasing various inflammatory cytokines, such as IL-1β, TNF-α, IL-6, IL-8, and IL-10, and blocking upstream TLR4, NF-κB, and MAPK pathways [61]. TLR4 is a type of protein involving the detection of endotoxins (such as LPS) produced by Gram-negative bacteria and signals through MyD88, subsequently giving rise to the downstream activation of NF-κB and MAPK signaling cascade [62–64]. Therefore, TLR4 could be a promising drug target of geniposide to fight against inflammation in a wide range of cell types. A schematic diagram of mechanisms for the anti-inflammatory effect of geniposide is shown in Figure 3. Additionally, geniposide owned in vitro and in vivo anti-inflammation and antiviral action in the infection of pandemic A/Jiangsu/1/2009 (H1N1) influenza virus. Geniposide (20 mg/kg/day) had similar clinical effects with the neuraminidase inhibitor (peramivir) (30 mg/kg/day) in pandemic A/Jiangsu/1/2009 (H1N1) influenza virus-induced Madin-Darby canine kidney (MDCK) cell injury by suppressing IL-4, IL-6 and IL-10, TNF-α, and interferon-γ (IFN-γ) [65].
Figure 3.

Schematic diagram of mechanisms for anti-inflammatory effects of geniposide. Geniposide inhibits the TLR4-mediated signaling cascades in inflammation induced by LPS. LPS: lipopolysaccharide; TLR4: Toll-like receptor 4; ERK: extracellular signal-regulated kinase; NF-κB: nuclear factor-kappa B; AP-1: activator protein-1; iNOS: inducible nitric oxide synthesis; IL: interleukin; NO: nitric oxide; IFN-γ: interferon-γ; TNF-α: tumor necrosis factor-α; MAPK: mitogen-activated protein kinase.
In fact, genipin similarly possesses a topical, potent, anti-inflammatory effect. Genipin was observed to exhibit higher inhibitory activity on inflammation (49.1% vs. 31.7%) at a lower dose (50 mg/kg) than geniposide at a higher dose (100 mg/kg) in the rat paw edema model [5]. Also, genipin in a concentration-dependent manner suppressed the production of NO and inducible nitric oxide synthesis (iNOS) in murine macrophage cells stimulated by LPS and IFN-γ, which helped to block NF-κB activation [6].
5.2. Antioxidant Activity
Oxidative stress generally occurs under an imbalanced redox status which encompasses either dysfunction of the antioxidant system or excessive production of reactive oxygen species (ROS). Dysregulated oxidative stress is appearing as an obvious causal factor in aging body and some age-related diseases [66, 67]. Cumulative evidence discloses that geniposide could upregulate endogenous antioxidative enzymes to be a promising strategy for delaying the process of cell injury (Figure 4).
Figure 4.

Schematic diagram of mechanisms for antioxidant activity of geniposide. Geniposide activates the GLP-1R-mediated signaling cascades in oxidative stress induced by H2O2, SIN-1, I/R, and high-fat food. ROS: reactive oxygen species; GLP-1R: glucagon-like peptide-1 receptor; CuZn-SOD: copper- and zinc-containing superoxide dismutase; SIN-1: 3-morpholinosydnonimine hydrochloride; PKA: protein kinase A; MAPK: mitogen-activated protein kinase; PPARα: peroxisome proliferator-activated receptor-α; PI3K: phosphatidylinositol-3 kinase; HO-1: heme oxygenase-1; Nrf2: nuclear factor-E2-related factor 2.
GLP-1 receptor is a member of the class B1 family of seven-transmembrane-spanning receptors, belonging to heterotrimeric G protein-coupled receptors (GPCRs) [68]. The signal transduction of the GLP-1 receptor is mainly mediated through the calcium and cyclic AMP (cAMP)/protein kinase A (PKA) signaling cascades [69]. The mechanisms associated with the neuroprotection of geniposide in PC12 cells against oxidative damage are involved in three signaling pathways. First, geniposide increased the expression of heme oxygenase-1 (HO-1) via the cAMP/PKA/cAMP response element-binding protein (CREB) signaling pathway [70]. Second, geniposide upregulated the expression of some antioxidative proteins involving HO-1 and Bcl-2 through activating the transcription factor of p90RSK via the MAPK signaling pathway [71]. Third, geniposide enhanced the phosphorylation of Akt473, Akt308, GSK-3β, and PDK1 in the phosphatidylinositol-3 kinase (PI3K) signaling pathway [72]. Furthermore, in Alzheimer's disease mice, geniposide improved learning and memory ability decreased by reducing oxidative stress as well as mitochondrial dysfunction via alleviating the mitochondrial oxidative damage and elevating the activity of cytochrome c oxidase and the mitochondrial membrane potential [68].
In addition, geniposide and genipin also conferred significant hepatoprotection by decreasing oxidative stress. On the one hand, geniposide potentiated the expression of HO-1 and attenuated cytochrome c protein, tBid expression, and caspase-3 activity [38]. On the other hand, geniposide also protected against hepatic steatosis in high fat diet-induced rats by its antioxidant actions and its regulation on adipocytokine release. In detail, geniposide increased the expression of peroxisome proliferator-activated receptor-α (PPARα) in addition to suppressing the expression of CYP2E1, giving rise to elevated superoxide dismutase and reduced MDA in liver [38]. Also, geniposide upregulated the level of some main antioxidant enzymes including glutathione peroxidase (GPx), hepatic lipid peroxidation (LPO), glutathione-S-transferase (GST), glutathione (GSH), and copper- and zinc-containing superoxide dismutase (CuZn-SOD) and thus protected against alcohol-induced oxidative stress injury in liver [9].
6. Preclinical Evidence for the Therapeutic Potential of Geniposide on Cardiovascular Disease and DM
Accumulating evidence demonstrated that geniposide exerted potential therapeutic benefits through modulating various genes and proteins associated with oxidative stress and inflammation. In this section, we mainly focused on their therapeutic potential in cardiovascular disease, and the details are summarized in Table 3.
Table 3.
Preclinical evidence of therapeutic potential from geniposide.
| System | Disorder | Model | Dose | Administration route | Pharmacological action | Reference |
|---|---|---|---|---|---|---|
| Endocrine system | Type 2 DM | Spontaneously obese Type 2 diabetic TSOD mice |
0.1% and 0.3% | Intragastric administration | Reducing the body weight and visceral fat accumulation and alleviating abnormal lipid metabolism and intrahepatic lipid accumulation | [16] |
| Type 2 DM | Free fatty acid-treated HepG2 cells | 10, 50, 100 μM | Incubation | Blocking the accumulation of intracellular lipid resulted from the free fatty acid treatment and enhanced the PPARα expression | [16] | |
| Type 2 DM | HepG2 cell model of insulin resistance | 15.63-125 mg/L | Incubation | Promoting autophagy and inhibiting insulin resistance in the HepG2 cells, which may be associated with the dynamic regulation of the P62/NF-κB/GLUT-4 pathway | [123] | |
| Type 2 DM | Palmitate-treated INS-1 cells | 1, 10, 100 μM | Incubation | Relieving β-cell apoptosis via activation of the GLP-1 receptor | [132] | |
| Type 2 DM | Cultured mouse islets after challenge with diabetic stimuli | 20 μM | Incubation | Promoting β regeneration and survival via regulating β-catenin/TCF7L2 pathway | [132] | |
| Type 2 DM | HepG2 cells | 1, 10, 100 μM | Incubation | Suppressing hepatic glucose production via AMPK signaling pathway | [137] | |
| Type 1or 2 DM | High glucose-induced glucotoxic insulinoma cells | 10 μM | Incubation | Improving β-cell function and increasing the proliferation of β-cells exposed to prolonged hyperglycemia | [128] | |
| Type 1 DM | Rat pancreatic islets | 10 μM | Incubation | Stimulating insulin secretion in pancreatic β-cells by regulating GLP-1 receptor/cAMP signaling and Ca2+ channels | [133] | |
| Type 1 DM | Rat pancreatic INS-1 cells | 10 μM | Incubation | Enhancing glucose uptake via activating AMPK in pancreatic β cells | [138] | |
| Diabetic nephropathy | Diabetic rats induced by streptozotocin | 50, 100 mg/kg/d | Intragastric administration | Suppressing NF-κB mediated inflammation response | [135] | |
|
| ||||||
| Cardiovascular system | Myocardium I/R | Hypoxia/reoxygenation-induced H9c2 Cells | 10, 20, 40, 80 μM | Incubation | Enhancing mitochondrial function via the GLP-1 receptor mediated the PI3K/Akt signaling pathway | [10] |
| Atherosclerosis | ApoE−/− mice fed a high-cholesterol diet | 100 mg/kg/d | Oral administration | Increasing proliferation of smooth muscle cells and suppressing inflammation | [94] | |
| Atherosclerosis | ApoE−/− mice fed a high-cholesterol diet | 100 mg/kg/d | Oral administration | Decreasing the dendritic cells numbers and inhibiting dendritic cell maturation in bone marrow and infiltration into lesions | [99] | |
| Atherosclerosis | ApoE−/− mice fed a high-cholesterol diet | 100 mg/kg/d | Oral administration | Regulating lipid and promoting the number and function of Treg cells | [103] | |
| Cardiac hypertrophy | Mice with transverse aortic constriction | 50 mg/kg/d | Oral administration | Activating the GLP-1 receptor/AMPKα pathway, inhibiting ERS and oxidative stress | [17] | |
| Cardiac fibrosis | Mice induced by isoprenaline | 50 mg/kg/d | Oral administration | Suppressing oxidative stress, ERS, and acetylated Smad3 in a SIRT1-dependent manner and inhibiting the phosphorylated-Samd3 pathway independent of SIRT1 activation | [78] | |
| Obesity-related cardiac injury | Mice induced by high-fat food | 50 mg/kg/d | Oral administration | Alleviating inflammation in a SIRT1-dependent manner and inhibiting cardiomyocyte apoptosis in a AMPKα-dependent manner | [92] | |
| Cerebral I/R injury | OGD/R-induced SH-SY5Y cells | 10 μM | Incubation | Inhibition of endoplasmic reticulum stress and autophagy | [108] | |
| Ischemic stroke. | tMCAO rats | 75 mg/kg/d | Intraperitoneal injections | Protecting neurons against post-ischemic neurovascular injury through the activation of GluN2A/AKT/ERK pathways | [115] | |
6.1. Cardiovascular Disease
6.1.1. Cardiac Fibrosis
Cardiac fibrosis is a common pathological alteration in myocardial infarction (MI), hypertension, dilated cardiomyopathy, and DM [73]. In the development of cardiac fibrosis, overproduction and deposition of collagen and extracellular matrix (ECM) will increase the stiffness of vessel, meanwhile deteriorating diastolic function, contributing to heart failure eventually [74]. Studies demonstrated that oxidative stress and ERS display close and critical relationships with cardiac fibrosis. On the one hand, cardiac oxidative stress could enhance the ability of collagen synthesis and inhibit collagen degradation in cardiac fibroblasts [75]. On the other hand, oxidative stress triggers accumulation of ECM in the heart of rats with diabetic cardiomyopathy by activating genes implicated with cardiac fibrosis, like TGF-β, fibronectin, and connective tissue growth factor (CTGF), apart from activating the NF-κB signaling pathway [76]. Sirtuin 1 deacetylase (SIRT1) is one of the mammalian homologs of the yeast silent information regulator 2 protein, which in essence is a nicotinamide adenine dinucleotide NAD(+)-dependent deacetylase [77]. A potent agonist of SIRT1, resveratrol, was found to inhibit the production of ROS in cardiomyocytes by activating SIRT1 and enhancing mitochondrial biogenesis [78]. Our previous study unveiled that geniposide could alleviate isoprenaline-induced cardiac dysfunction and fibrosis in the mouse model with cardiac fibrosis. An in vitro experiment disclosed that geniposide also suppressed the transformation of cardiac fibroblasts into myofibroblasts stimulated by TGF-β. The mechanism of anti-cardiac fibrosis from geniposide is associated with the inhibition of the acetylated- (ac-) Smad3 pathway, oxidative stress, and ERS in a SIRT1-dependent manner; meanwhile, geniposide could also reduce phosphorylated- (p-) Smad3 level by inactivating the type II TGF-β receptor independent of SIRT1 activation [78] (Figure 5).
Figure 5.

Schematic diagram of mechanisms in cardiac fibrosis and cardiac hypertrophy of geniposide. TGF-β: transforming growth factor-β; AngII: angiotensin II; Tβ R: TGF-β receptor; ERS: endoplasmic reticulum stress; GLP-1R: glucagon-like peptide-1 receptor; AMPKα: AMP-activated protein kinase α; ERK: extracellular signal-regulated kinase; JNK: c-Jun NH2-terminal kinase; P: phosphorylated; ac: acetylated.
6.1.2. Cardiac Hypertrophy
Cardiac hypertrophy is the response of the heart to multifarious intrinsic and extrinsic stimuli imposing enhanced biomechanical stress, which can deteriorate to heart failure and eventually cause high mortality [79]. One of the mechanisms contributing to cardiac hypertrophy is oxidative stress and ERS. Once oxidative stress and ERS in the myocardium are initiated, branches of certain protein responses will elevate ROS and bring about apoptosis, resulting in the process of cardiac hypertrophy [80]. AMPKα as a vital regulator of cardiac homeostasis can block the synthesis of proteins by antagonizing mTOR kinase, which has been disclosed to act as an intrinsic suppressor in oxidative stress and ERS [81]. Additionally, activating AMPKα also prevented the heart from I/R injury via the inhibition of oxidative stress ad ERS [82]. Our previous study uncovered that, in the mouse model with transverse aortic constriction (TAC), geniposide remarkably blocked the hypertrophic response by increasing the left ventricular internal diastolic diameter and fractional shortening and decreasing the ejection fraction. Activation of AMPKα and suppression of mTOR, ERK, and ERS were observed in hypertrophic hearts and angiotensin II- (AngII-) induced H9c2 cardiomyocytes that were treated with geniposide. However, all of these alterations were counteracted after AMPKα was blocked by compound C or the GLP-1 receptor was knocked down, implying that the protective roles of geniposide on cardiac hypertrophy were mediated by the GLP-1 receptor/AMPKα signaling pathway [17] (Figure 5).
6.1.3. Myocardium I/R
Myocardium I/R injury is one main cause of morbidity and mortality implicated with ischemia heart disease, the typical pathological alterations of which were cardiomyocyte apoptosis and overproduction of ROS [83]. It is universally recognized that unbalanced and high steady-state levels of ROS and nitrogen species (RNS) are responsible for cytotoxicity in the heart, which gives rise to contractile dysfunction and cardiomyocyte death [84]. Ischemia is featured by a virtual lack of O2 and other critical substrates. Long-term ischemia could accumulate various metabolites including lactate, which further induces a cascade of events involving excess ROS/RNS generation, disruption of Ca2+ homeostasis, and loss of nucleotide homeostasis [85]. Mitochondria not only are the main enzymatic source of ROS but also were major target organelles for ROS-mediated damage in cells [86]. In MI, excess ROS could trigger mitochondrial DNA (mtDNA) damage, which further gives rise to dysregulation of respiratory chain complex enzymes and mtDNA-encoded genes, eventually resulting in the occurrence of cardiac remodelling and heart failure [87].
The previous study showed that geniposide was able to block the oxidative stress and apoptosis of H9c2 cells challenged with hypoxia/reoxygenation by improving mitochondrial function, as shown by improving mitochondrial morphological alterations, relieving mitochondrial oxidative stress, decreasing mitochondrial calcium overload, and maintaining mitochondrial membrane potential. These antioxidative effects of geniposide on mitochondria in I/R injury were partly due to activation of the GLP-1 receptor and downstream PI3K/Akt signaling pathway [10].
6.1.4. Obesity-Related Cardiac Injury
Obesity, a risk factor for cardiovascular disease, triggers structural and functional alterations in the myocardium [88]. Long-term obesity could predispose an individual to various cardiac complications, involving cardiac dysfunction, cardiac energy metabolism disturbance and remodelling, and even heart failure [89]. The driving forces in obesity-related cardiac injury involve an excess supply of saturated fatty acids, fibrosis, hypertrophy, myocardial inflammation, loss of cardiomyocytes, and epigenetics [90–92]. Our previous study demonstrated that oral administration of geniposide could inhibit obesity and obesity-induced cardiac injury. To be more specific, we found that geniposide alleviated myocardial inflammation in high-fat diet- (HFD-) induced mice in a SIRT1-dependent manner; meanwhile, geniposide combated cardiomyocyte apoptosis in an AMPKα-dependent manner. In particular, the activation of SIRT1 and AMPKα in the myocardium was mediated by the GLP-1R [92]. Our study hints that geniposide possesses the potential to be a clinical candidate for the treatment of obesity and obesity-related cardiac injury.
6.1.5. Atherosclerosis
Atherosclerosis is featured by vascular oxidative stress, inflammation, endothelial dysfunction, and formation of lipids in the vessel intima, which is also followed by proliferation and migration of vascular smooth muscle cells, as well as endothelial activation initiated by various inflammatory molecules, and finally causes the buildup of lipid plaque [93]. Studies demonstrated that geniposide combined with another herb, baicalin, could obviously attenuate the formation of atherosclerotic plaque in high-lipid diet-treated mice through increasing the proliferation of smooth muscle cells and suppressing oxidative stress and inflammation, the underlying mechanisms of which may be involved in inhibiting Wnt1 and enhancing the ratio of Wnt1/dickkopf-related protein 1 (DKK1) [94]. Dendritic cells, a type of professional antigen-presenting cells implicated with innate and adaptive immune responses, are also proved to modulate the immune status in atherosclerosis [95, 96]. During the process of atherosclerosis, a great many dendritic cells are discovered inside the adventitial layer of blood vessels and atherosclerotic plaques [97]. On the contrary, the absence of dendritic cells or inhibition of the maturation of dendritic cells could alleviate atherosclerotic lesions by reducing the accumulation of monocytes and macrophages [98, 99]. In ApoE−/− mice fed a high-cholesterol diet, either geniposide or baicalin or a combined administration of the two drugs can regress the process of atherosclerosis through lowering the concentration of low-density lipoprotein (LDL) and total cholesterol (TC) in serum. Interestingly, either geniposide or baicalin alone could reduce the number of CD83-expressing and CD11C dendritic cells in the bone marrow, but the CD83 expression in atherosclerotic lesions was not decreased. However, when combining geniposide and baicalin, the expression, maturation, mobilization, and infiltration of CD83 and CD11C in both atherosclerotic lesions and bone marrow were inhibited [99]. We speculate that the drug-drug interactions may improve the pharmaceutical effects of geniposide on atherosclerosis, but the specific mechanism needs further exploring. Apart from dendritic cells, other immunocytes such as Th1 and Th2 cells are also captured in atherosclerotic plaque, which are highly associated with the process of atherosclerosis. It has been reported that Th2 cells contribute to a beneficial response while Th1 cells have the opposite effects in atherosclerosis [100]. One of the subtypes from T helper cells, CD4+CD25+Foxp3+ regulatory T cells, were identified to exclusively repress both Th1 and Th2 cells and therefore inhibited the development of atherosclerotic lesions [101]. Furthermore, Treg cells that specifically express the forkhead/winged helix transcription factor (Foxp3) display potential anti-inflammatory effects by secreting IL-10 and TGF-β1 [102]. Similarly, pretreatment with geniposide or baicalin helped the ApoE-/- mice with high-cholesterol diet produce more splenic Treg cells, TGF-β1, and IL-10, in addition to notably decreasing serum LDL and TC. At the same time, the levels of Foxp3 and Foxp3+ regulatory T cells in atherosclerotic lesions were significantly enhanced by geniposide and baicalin [103]. Taken together, the three studies above demonstrate that geniposide is both an immunoregulatory agent and a lipid-regulation agent in protecting against atherosclerosis and combination with baicalin will improve the efficacy of geniposide (Figure 6).
Figure 6.

Schematic diagram of mechanisms of geniposide in atherosclerotic. ROS: reactive oxide species; Th cells: T helper cells; IL: interleukin.
6.1.6. Ischemic Stroke
Ischemic stroke, accounting for about 85% of all stroke cases, correlates with high morbidity and mortality globally [104]. Ischemic stroke will further deteriorate to cerebral I/R injury by sudden recovery of the blood supply unless managed appropriately, finally causing cerebral microcirculatory damage and death of neurons [105]. Autophagy activation, ERS, and oxidative stress play significant roles in the development of cerebral I/R injury [106, 107]. Treatment with a combination of geniposide and tauroursodeoxycholic acid could counteract the elevation in cellular ROS levels, the activation of oxygen-glucose deprivation-reoxygenation- (OGD/R-) induced ERS, and autophagy in SH-SY5Y cells with hypoxia-reoxygenation injury [108]. On the other hand, inflammatory events that occur between the blood-endothelium interface of the cerebral capillaries also underlie the resultant ischemic tissue damage [109]. However, these inflammatory events in cerebral ischemia could block geniposide via activating the P2Y14 receptor. In the cellular model of OGD-induced cerebral ischemia, the level of P2Y14 receptor and its downstream signaling pathways involving RAF-1, MEK1/2, and ERK1/2 were all downregulated by geniposide. At the same time, the production of IL-1β, IL-8, and MCP-1 in brain microvascular endothelial cells (BMECs) were also suppressed. These results indicated that anti-inflammatory and antioxidative effects of geniposide in cerebral ischemia depend on the expression of the P2Y14 receptor and its downstream ERK1/2 signaling pathways [110].
Calcium-permeable ionotropic N-methyl-D-aspartate receptor- (NMDAR-) mediated Ca2+ overload, glutamatergic overexcitation, and oxidative stress are believed to be vital factors in regulating the development of ischemic stroke [111]. Assuming that NMDAR overexcitation could disturb the equilibrium between inhibition and excitation in I/R injury, restoring this balance by intervening in neural transmission in the post-ischemic brain is likely to be a selective and useful strategy [112, 113], GluN2A and GluN2B are the major subtypes of NMDAR in the hippocampus, which are, respectively, responsible for neuronal survival and death following I/R [114]. Notably, geniposide could decrease the infarct volume in transient middle cerebral artery occlusion (tMCAO) rats in a dose-dependent manner. A medium level of geniposide applied in vivo effectively prevented post-stroke injury by regulating blood brain barrier leakage/hemorrhage and neuronal loss/apoptosis, meanwhile enhancing the expression of GluN2A, but not GluN2B. However, when applying the GluN2A antagonist, NVP, the therapeutic effects in MCAO rats could be blocked, implying that GluN2A is a potential target of geniposide. Whereafter, activation of GluN2A led to a remarkable increase in network excitation, which further activated prosurvival signals involving pAKT, pERK, and PSD-95 in ischemia. In brief, the GluN2A/AKT/ERK axis was essential for the neuroprotective effects of geniposide in post-ischemic neurovascular injury [115].
6.1.7. Hypertension
Moreover, the antihypertension effects of Gardenia jasminoides were also investigated in the L-NG-nitroarginine- (L-NNA-) induced hypertension mouse model. The Gardenia jasminoides-treated mice displayed lower diastolic blood pressure, systolic blood pressure, and mean blood pressure compared with the hypertensive mice without administration of Gardenia jasminoides. Molecular biological assays showed that the level of NO in multiple tissues including the serum, heart, kidney, stomach, and liver of Gardenia jasminoides-treated mice were higher, but the malondialdehyde contents were lower. In the hypertensive mice treated with Gardenia jasminoides, a higher mRNA expression of neuronal NOS, HO-1, Bax, caspase-3, -8, and -9, and endothelial NOS and a lower mRNA expression of receptor activity-modifying protein (RAMP), adrenomedullin, IL-1β, TNF-α, Bcl-2, inducible NOS, monocyte chemoattractant protein-1 (MCP-1), NF-κB, and matrix metalloproteinase- (MMP-) 2 and -9 were found, indicating that the antihypertension effects of Gardenia jasminoides may be implicated with its antiapoptosis and antioxidative stress. But the active component in Gardenia jasminoides (geniposide, genipin, crocin 1, and crocin 2) for antihypertension is not yet clear [116].
6.2. Diabetes Mellitus
Diabetes mellitus (DM) is a serious, chronic disease which occurs either when the body does not use the insulin produced by itself effectively meaning insulin resistance (type 2 DM) or when the pancreas cannot produce sufficient insulin meaning insulin deficiency (type 1 DM). Over the past few decades, both the number of the prevalence and cases of DM have been steadily growing [117]. Geniposide displays potent effects against DM through multiple mechanisms (Figure 7).
Figure 7.

Schematic diagram of mechanisms of geniposide in diabetes mellitus. TCF7L2: T-cell factor 7-like 2; GLP-1R: glucagon-like peptide-1 receptor; PKA: protein kinase A; cAMP: cyclic adenosine monophosphate; Kv channels: voltage-gated K channels; JAK2: tyrosine kinase 2; P: phosphorylated.
6.2.1. Type 2 DM
Previous study showed that the metabolic syndrome increased the risk for DM independently of other risk factors. With the accumulation of visceral fat, many symptoms of metabolic syndrome such as abnormal glucose/lipid metabolism and insulin resistance are induced, which eventually give rise to the onset of ischemic cerebrovascular disease [118]. In spontaneously obese type 2 DM TSOD mice, geniposide reduced the body weight and visceral fat accumulation and alleviated abnormal lipid metabolism as well as intrahepatic lipid accumulation. Meanwhile, it also exerted good therapeutic effects on hyperinsulinemia and abnormal glucose tolerance. To further explore the direct function of geniposide on the liver, genipin was used in free fatty acid-treated mice. Genipin not merely blocked the accumulation of intracellular lipid which resulted from the free fatty acid treatment but also enhanced the intracellular fatty acid oxidation-related ge HepG2 fatty liver modene (PPARα) expression at the same time. Taken together, these results demonstrated that geniposide possessed antiobesity and antioxidative effects, insulin resistance-alleviating effects, and abnormal lipid metabolism-alleviating effects, but it is genipin that showed a direct effect on liver through enhancing the expression of PPARα [16].
Autophagy is an intracellular process, which could sequester damaged or senescent organelles and proteins in autophagosomes to recycle their degradation products [119, 120]. Insulin and intracellular molecules involving mTOR can inhibit the process of autophagy, whereas glucagon as a type of counterregulatory hormone of insulin can activate autophagy, implying that autophagy may be implicated with the natural history of DM via its involvement in either organelle function or hormone activity [121]. As a key complex connected with autophagosomal membranes, p62/SQSTM1 (P62) could swallow cytoplasmic content for subsequent degradation. Hyperglycemia enhances the p62/PKC interaction and autophagy, mediating the activation of NF-κB and increasing the expression of NADPH oxidase 4 [122]. In addition, glucose transporter 4 (GLUT-4) is a vital medium in transport of extracellular glucose to insulin-sensitive cells, the translocation of which to the insulin-sensitive cell membrane is modulated by insulin. In a HepG2 cell model of insulin resistance, geniposide treatment initially potentiated the expression of GLUT-4, whereafter it decreased its expression in time to its nadir at 8 h. In the meantime, the level of autophagy was significantly enhanced by geniposide through modulating the expression of LC3 and P62. Also, the NF-κB signaling pathway was blocked by both the direct effect of geniposide and the activation of autophagy, resulting in alleviation of insulin resistance in HepG2 cells eventually [123].
As a result of loss of β-cell mass or defects in insulin secretion, type 2 DM is also connected with the increased level of β-cell apoptosis, which makes pancreatic β cells cannot compensate for peripheral insulin resistance [124]. The emerging evidence uncovers that lipotoxicity may undertake an essential role in pancreatic β-cell apoptosis and oxidative stress in type 2 DM. A high level of plasma free fatty acids is cytotoxic to pancreatic β cells, which activates apoptotic and oxidative signaling pathways to initiate mitochondrial perturbation and causes oxidative stress [125]. Hence, assuming that a drug can preserve β-cell mass by inhibiting lipotoxicity, it may be a promising candidate against the development of DM. Liu et al. found that pretreatment of pancreatic INS-1 cells with geniposide for 7 h relieved β-cell apoptosis induced by palmitate, but at 18 h, this effect disappeared. Additionally, in palmitate-treated INS-1 cells, geniposide also attenuated the impairment of GLP-1R by increasing the phosphorylation of Foxo1 and AKT and enhanced the expression of PDX-1, indicating that GLP-1R was a key part in alleviating lipotoxicity-induced β-cell apoptosis by geniposide [126]. When pancreatic β-cells are chronically exposed to high glucose concentration for an extended time, glucose-stimulated insulin secretion in β-cells would be impaired and β-cell apoptosis also increased, which is so-called glucotoxicity-related apoptosis [127]. However, the glucose-induced impairment of insulin secretion and injury of the pancreas islet could be obviously reverted by the inclusion of 1 or 10 μM of geniposide. Induction of AMPK by geniposide was involved in promoting the use of nutrient stores for energy production and antagonizing pancreatic β-cell injury [128]. Intriguingly, the effects of geniposide on acute glucose-stimulated insulin secretion in INS-1 cells depended on the concentration of glucose. Geniposide could significantly enhance insulin secretion in response to the acute stimulation of low or moderately high concentrations of glucose and promoted glucose uptake as well as the production of intracellular ATP. But in INS-1 cells stimulated acutely by high concentrations of glucose, these protective effects disappeared. This may be because of the capacity to preserve pancreatic β-cells from exhaustion and damage caused by prolonged and oversecretion of insulin as well as glucotoxicity under high glucose burden [129].
Apart from inhibiting β-cell apoptosis, another therapeutic strategy for DM is to promote new β-cell formation. The study showed that proliferation of preexisting β-cells can generate new β-cells [130]. Hence, facultative progenitors in regenerating pancreatic ducts could be a promising source of β-cells [131]. By activating T-cell factor 7-like 2 (TCF7L2), a crucial transcription factor in Wnt/β-catenin signaling and having vital roles in β-cell survival and regeneration, geniposide remarkably increased β-cell survival. Geniposide could trigger duct cell differentiation through enhancing TCF7L2 expression and activating the JAK2/STAT3 pathway in exocrine cells isolated from the mouse pancreas. The two mechanisms of geniposide promoted its regeneration in cultured mouse islets after challenge with diabetic stimuli [132].
6.2.2. Type 1 DM
The role of geniposide in type 1 DM was also investigated. In rat pancreatic islets, geniposide enhanced insulin secretion via activating the GLP-1R as well as its downstream adenylyl cyclase (AC)/cAMP signaling pathway. However, inhibition of PKA blocked the insulinotropic effect from geniposide. On the other hand, geniposide also suppressed voltage-dependent potassium (Kv) channels (Ca2+ channels) and prolonged the action potential duration, but this effect could be reversed by inhibiting PKA or GLP-1R. Collectively, these results demonstrated that geniposide acutely improved the ability of insulin secretion in pancreatic β-cells through activating GLP-1R/cAMP signaling and Ca2+ channels were also involved in this process [133]. Further research unveiled that geniposide in a time-dependent manner induced phosphorylation of acetyl-CoA carboxylase (ACC), a marker of AMPK activity and enhanced glucose uptake, energy metabolism, and insulin secretion in rat pancreatic INS-1 cells; however, these effects were remedied by compound C (an AMPK inhibitor) [133].
6.2.3. Diabetic Nephropathy
Diabetic nephropathy is one of the most common complications of DM and is also a major factor causing end-stage renal disease, the pathogenesis of which is implicated with immunologic and inflammatory response [134]. Intragastric administration of geniposide ameliorated structural and functional abnormalities of the kidney by suppressing NF-κB-mediated inflammation response in diabetic rats induced by streptozotocin [135].
7. Future Perspective of Geniposide
Over the last several decades, a great number of studies in animal and cell models have uncovered the potential of geniposide for the treatment of a vast array of diseases, ranging from neurological diseases to cardiovascular diseases and DM. However, we still face the following problems to be solved urgently in terms of the clinical application of geniposide. For instance, at present, the studies on geniposide only stay at animal and cell levels and have not been carried out in the human body. Hence, further clinical studies are imperative to determining a conclusive role for geniposide in human therapeutics. Additionally, although the pharmacokinetics of geniposide has made a lot of progress, we need more details on its transformation, monitoring, and metabolism so that its efficacy and safety are guaranteed. Firstly, we need to employ some methods to increase the transformation rate of geniposide into genipin. Yang and his colleagues [136] investigated the hydrolysis of geniposide to genipin by β-glucosidase immobilized by the crosslinking-embedding in an aqueous-organic two-phase system. The optimum reaction pH value of immobilized and free β-glucosidase was 4.5 while the optimum reaction temperature was 55°C and 50°C, respectively. Also, the immobilization decreased the effect of pH variation on the activity of β-glucosidase and enhanced its heat resistance. Secondly, natural medicines often possess relatively low active ingredients, the concentration of which is difficult to detect after entering the peripheral blood. Therefore, to further optimize the monitoring program, we had better take advantage of a variety of methods and combine multiple indicators. Thirdly, few studies focus on the metabolites of geniposide currently; hence, we should further evaluate the structure of its metabolites, metabolic pathways, and pharmacological effects in order to elucidate the true molecular basis of pharmacological actions of geniposide. Fourthly, in order to acquire a relatively high geniposide content with low cost, more plants as well as their parts need to be detected to find the optimal candidate for geniposide preparation. All of these mentioned above will be of great importance for its new drug research and clinical rational use.
8. Conclusion
In this review, we uncover the pharmacokinetics and preclinical values of geniposide, which might be beneficial and referential for clinical applications and future investigations. We discover that the pharmacokinetics of geniposide is complicated and it can be decomposed into multiple metabolites in vivo. Geniposide predominately exists in the liver and kidney and is excreted in the original form via the kidney. Furthermore, geniposide possesses a great body of pharmacological effects involving antioxidative stress, anti-inflammation, and antiapoptosis. Owing to these favorable effects, geniposide serves as a weapon against various disorders, especially cardiovascular disease and DM. Although numerous investigations are exploring the preclinical values of geniposide, we assume that geniposide has more possible targets and molecular pathways in disease treatment, which demands further investigation. In particular, the use of these natural antioxidants like geniposide should not be indiscriminate in that it still lacks clinical proofs at present. Hence, more empirical data pertaining to improvement in clinical application, safety, and efficacy of geniposide are demanded before geniposide could be translated from bench to bedside. Nevertheless, geniposide does seem to be on behalf of the light at the end of the tunnel as a supportive and ideal remedy for allopathic treatment considering its wide range of purported efficacy and the relatively lesser incidence of adverse reactions. Sincerely, we hope this review might provide some proofs for clinical applications and further studies in the future.
Acknowledgments
This work was supported by grants from the National Natural Science Foundation of China (Nos. 81470516 and 81770399), the Key Project of the National Natural Science Foundation (No. 81530012), and the Fundamental Research Funds for the Central Universities of China (2042018kf0121).
Conflicts of Interest
The authors declare that there are no conflicts of interest regarding the publication of this paper.
Authors' Contributions
Ning Li and Ling Li contributed equally to this work.
References
- 1.Prasad S., Tyagi A. K. Traditional medicine: the goldmine for modern drugs. Advanced Techniques in Biology & Medicine. 2015;3(2) doi: 10.4172/2379-1764.1000e108. [DOI] [Google Scholar]
- 2.Chang W. L., Wang H. Y., Shi L. S., Lai J. H., Lin H. C. Immunosuppressive iridoids from the fruits of Gardenia jasminoides. Journal of Natural Products. 2005;68(11):1683–1685. doi: 10.1021/np0580816. [DOI] [PubMed] [Google Scholar]
- 3.Koo H. J., Lee S., Shin K. H., Kim B. C., Lim C. J., Park E. H. Geniposide, an anti-angiogenic compound from the fruits of Gardenia jasminoides. Planta Medica. 2004;70(5):467–469. doi: 10.1055/s-2004-818978. [DOI] [PubMed] [Google Scholar]
- 4.Tsai T. R., Tseng T. Y., Chen C. F., Tsai T. H. Identification and determination of geniposide contained in Gardenia jasminoides and in two preparations of mixed traditional Chinese medicines. Journal of Chromatography A. 2002;961(1):83–88. doi: 10.1016/S0021-9673(02)00365-5. [DOI] [PubMed] [Google Scholar]
- 5.Koo H. J., Lim K. H., Jung H. J., Park E. H. Anti-inflammatory evaluation of gardenia extract, geniposide and genipin. Journal of Ethnopharmacology. 2006;103(3):496–500. doi: 10.1016/j.jep.2005.08.011. [DOI] [PubMed] [Google Scholar]
- 6.Koo H. J., Song Y. S., Kim H. J., et al. Antiinflammatory effects of genipin, an active principle of gardenia. European Journal of Pharmacology. 2004;495(2-3):201–208. doi: 10.1016/j.ejphar.2004.05.031. [DOI] [PubMed] [Google Scholar]
- 7.Wang S. C., Liao H. J., Lee W. C., Huang C. M., Tsai T. H. Using orthogonal array to obtain gradient liquid chromatography conditions of enhanced peak intensity to determine geniposide and genipin with electrospray tandem mass spectrometry. Journal of Chromatography A. 2008;1212(1-2):68–75. doi: 10.1016/j.chroma.2008.10.035. [DOI] [PubMed] [Google Scholar]
- 8.Sato S., Kitamura H., Chino M., Takei Y., Hiruma M., Nomura M. A 13-week oral dose subchronic toxicity study of gardenia yellow containing geniposide in rats. Food and Chemical Toxicology. 2007;45(8):1537–1544. doi: 10.1016/j.fct.2007.02.015. [DOI] [PubMed] [Google Scholar]
- 9.Wang J., Zhang Y., Liu R., Li X., Cui Y., Qu L. Geniposide protects against acute alcohol-induced liver injury in mice via up-regulating the expression of the main antioxidant enzymes. Canadian Journal of Physiology and Pharmacology. 2015;93(4):261–267. doi: 10.1139/cjpp-2014-0536. [DOI] [PubMed] [Google Scholar]
- 10.Jiang Y.-Q., Chang G.-l., Wang Y., Zhang D.-Y., Cao L., Liu J. Geniposide prevents hypoxia/reoxygenation-induced apoptosis in H9c2 cells: improvement of mitochondrial dysfunction and activation of GLP-1R and the PI3K/AKT signaling pathway. Cellular Physiology and Biochemistry. 2016;39(1):407–421. doi: 10.1159/000445634. [DOI] [PubMed] [Google Scholar]
- 11.Zhang H. Y., Liu H., Yang M., Wei S. F. Antithrombotic activities of aqueous extract from Gardenia jasminoides and its main constituent. Pharmaceutical Biology. 2013;51(2):221–225. doi: 10.3109/13880209.2012.717088. [DOI] [PubMed] [Google Scholar]
- 12.Lee H. Y., Lee G. H., Lee M. R., et al. Eucommia ulmoides Oliver extract, aucubin, and geniposide enhance lysosomal activity to regulate ER stress and hepatic lipid accumulation. PLoS One. 2013;8(12, article e81349) doi: 10.1371/journal.pone.0081349. [DOI] [PMC free article] [PubMed] [Google Scholar] [Retracted]
- 13.Lelono R. A., Tachibana S., Itoh K. Isolation of antifungal compounds from Gardenia jasminoides. Pakistan Journal of Biological Sciences. 2009;12(13):949–956. doi: 10.3923/pjbs.2009.949.956. [DOI] [PubMed] [Google Scholar]
- 14.Zhao C., Lv C., Li H., et al. Geniposide protects primary cortical neurons against oligomeric Aβ1-42-induced neurotoxicity through a mitochondrial pathway. PLoS One. 2016;11(4, article e0152551) doi: 10.1371/journal.pone.0152551. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 15.Park J. H., Yoon J., Lee K. Y., Park B. Effects of geniposide on hepatocytes undergoing epithelial–mesenchymal transition in hepatic fibrosis by targeting TGFβ/Smad and ERK-MAPK signaling pathways. Biochimie. 2015;113:26–34. doi: 10.1016/j.biochi.2015.03.015. [DOI] [PubMed] [Google Scholar]
- 16.Kojima K., Shimada T., Nagareda Y., et al. Preventive effect of geniposide on metabolic disease status in spontaneously obese type 2 diabetic mice and free fatty acid-treated HepG2 cells. Biological & Pharmaceutical Bulletin. 2011;34(10):1613–1618. doi: 10.1248/bpb.34.1613. [DOI] [PubMed] [Google Scholar]
- 17.Ma Z. G., Dai J., Zhang W. B., et al. Protection against cardiac hypertrophy by geniposide involves the GLP-1 receptor/AMPKα signalling pathway. British Journal of Pharmacology. 2016;173(9):1502–1516. doi: 10.1111/bph.13449. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 18.Cheng Z., Xu H., Wang X., Liu Z. Lactobacillus raises in vitro anticancer effect of geniposide in HSC‑3 human oral squamous cell carcinoma cells. Experimental and Therapeutic Medicine. 2017;14:4586–4594. doi: 10.3892/etm.2017.5105. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 19.Chang T. T., Chen J. W. Emerging role of chemokine CC motif ligand 4 related mechanisms in diabetes mellitus and cardiovascular disease: friends or foes? Cardiovascular Diabetology. 2016;15(1):p. 117. doi: 10.1186/s12933-016-0439-9. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 20.Kawata Y., Hattori M., Akao T., Kobashi K., Namba T. Formation of nitrogen-containing metabolites from geniposide and gardenoside by human intestinal bacteria. Planta Medica. 1991;57(6):536–542. doi: 10.1055/s-2006-960201. [DOI] [PubMed] [Google Scholar]
- 21.Akao T., Kobashi K., Aburada M. Enzymic studies on the animal and intestinal bacterial metabolism of geniposide. Biological & Pharmaceutical Bulletin. 1994;17(12):1573–1576. doi: 10.1248/bpb.17.1573. [DOI] [PubMed] [Google Scholar]
- 22.Yu D., Zhang Y., Guo L., Zhang Q., Zhu H. Study on the absorption mechanism of geniposide in the Chinese formula Huang-Lian-Jie-Du-Tang in rats. AAPS PharmSciTech. 2017;18(4):1382–1392. doi: 10.1208/s12249-016-0610-3. [DOI] [PubMed] [Google Scholar]
- 23.Chula S., Hang L., Yinying B., Jianning S., Shi R. The effects of notoginsenoside R1 on the intestinal absorption of geniposide by the everted rat gut sac model. Journal of Ethnopharmacology. 2012;142(1):136–143. doi: 10.1016/j.jep.2012.04.027. [DOI] [PubMed] [Google Scholar]
- 24.Li Y., Cai W., Cai Q., Che Y., Zhao B., Zhang J. Comprehensive characterization of the in vitro and in vivo metabolites of geniposide in rats using ultra-high-performance liquid chromatography coupled with linear ion trap–Orbitrap mass spectrometer. Xenobiotica. 2015;46(4):357–368. doi: 10.3109/00498254.2015.1079746. [DOI] [PubMed] [Google Scholar]
- 25.Ueno K., Takeda Y., Iwasaki Y., Yoshizaki F. Simultaneous estimation of geniposide and genipin in mouse plasma using high-performance liquid chromatography. Analytical Sciences. 2001;17(10):1237–1239. doi: 10.2116/analsci.17.1237. [DOI] [PubMed] [Google Scholar]
- 26.Chen J., Wu H., Dai M. M., Li H., Chen J. Y., Hu S. L. Identification and distribution of four metabolites of geniposide in rats with adjuvant arthritis. Fitoterapia. 2014;97:111–121. doi: 10.1016/j.fitote.2014.05.023. [DOI] [PubMed] [Google Scholar]
- 27.Tseng T. Y., Tsai T. H. Measurement of unbound geniposide in blood, liver, brain and bile of anesthetized rats: an application of pharmacokinetic study and its influence on acupuncture. Analytica Chimica Acta. 2004;517(1-2):47–52. doi: 10.1016/j.aca.2004.04.040. [DOI] [Google Scholar]
- 28.Wu M. T., Shaw L. H., Wu Y. T., Tsai T. H. Interaction of acupuncture and electroacupuncture on the pharmacokinetics of aspirin and the effect of brain blood flow in rats. Evidence-Based Complementary and Alternative Medicine. 2013;2013:9. doi: 10.1155/2013/670858.670858 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 29.Wang F., Cao J., Hao J., Liu K. Pharmacokinetics, bioavailability and tissue distribution of geniposide following intravenous and peroral administration to rats. Biopharmaceutics & Drug Disposition. 2014;35(2):97–103. doi: 10.1002/bdd.1869. [DOI] [PubMed] [Google Scholar]
- 30.Li H., Wu H., Shen C., Chen J. Y., Hu S. L., Wu H. Comparative pharmacokinetics study after oral administration of geniposide in normal rats and adjuvant-induced arthritis rats by UPLC-MS/MS. Basic & Clinical Pharmacology & Toxicology. 2013;113(5):294–299. doi: 10.1111/bcpt.12113. [DOI] [PubMed] [Google Scholar]
- 31.Wu X. J., Zhao L. M. Serum pharmacochemistry and pharmarcokinetics study of Naoxuening injection. Traditional Chinese Drug Research & Clinical Pharmacology. 2008;19:376–379. [Google Scholar]
- 32.Gao L. N., Zhang Y., Cui Y. L., Yan K. Evaluation of genipin on human cytochrome P450 isoenzymes and P-glycoprotein in vitro. Fitoterapia. 2014;98:130–136. doi: 10.1016/j.fitote.2014.07.018. [DOI] [PubMed] [Google Scholar]
- 33.Shah R. R., Gaedigk A., LLerena A., Eichelbaum M., Stingl J., Smith R. L. CYP450 genotype and pharmacogenetic association studies: a critical appraisal. Pharmacogenomics. 2016;17(3):259–275. doi: 10.2217/pgs.15.172. [DOI] [PubMed] [Google Scholar]
- 34.Thirumaran R. K., Heck J. W., Hocum B. T. CYP450 genotyping and cumulative drug–gene interactions: an update for precision medicine. Personalized Medicine. 2016;13(1):5–8. doi: 10.2217/pme.15.47. [DOI] [PubMed] [Google Scholar]
- 35.Han Y.-L., Yu H.-L., Li D., et al. In vitro inhibition of Huanglian [Rhizoma coptidis(L.)] and its six active alkaloids on six cytochrome P450 isoforms in human liver microsomes. Phytotherapy Research. 2011;25(11):1660–1665. doi: 10.1002/ptr.3475. [DOI] [PubMed] [Google Scholar]
- 36.Ye L. H., Kong L. T., Yan M. Z., et al. Lotus leaf alkaloid fraction can strongly inhibit CYP2D6 isoenzyme activity. Journal of Ethnopharmacology. 2016;194:913–917. doi: 10.1016/j.jep.2016.10.055. [DOI] [PubMed] [Google Scholar]
- 37.Tang L., Ye L., Lv C., Zheng Z., Gong Y., Liu Z. Involvement of CYP3A4/5 and CYP2D6 in the metabolism of aconitine using human liver microsomes and recombinant CYP450 enzymes. Toxicology Letters. 2011;202(1):47–54. doi: 10.1016/j.toxlet.2011.01.019. [DOI] [PubMed] [Google Scholar]
- 38.Ma T., Huang C., Zong G., et al. Hepatoprotective effects of geniposide in a rat model of nonalcoholic steatohepatitis. Journal of Pharmacy and Pharmacology. 2011;63(4):587–593. doi: 10.1111/j.2042-7158.2011.01256.x. [DOI] [PubMed] [Google Scholar]
- 39.Wang J., Gan C., Sparidans R. W., et al. P-glycoprotein (MDR1/ABCB1) and breast cancer resistance protein (BCRP/ABCG2) affect brain accumulation and intestinal disposition of encorafenib in mice. Pharmacological Research. 2018;129:414–423. doi: 10.1016/j.phrs.2017.11.006. [DOI] [PubMed] [Google Scholar]
- 40.Morad S. A. F., Davis T. S., MacDougall M. R., et al. Role of P-glycoprotein inhibitors in ceramide-based therapeutics for treatment of cancer. Biochemical Pharmacology. 2017;130:21–33. doi: 10.1016/j.bcp.2017.02.002. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 41.Abdallah H. M., al-Abd A. M., el-Dine R. S., el-Halawany A. M. P-glycoprotein inhibitors of natural origin as potential tumor chemo-sensitizers: a review. Journal of Advanced Research. 2015;6(1):45–62. doi: 10.1016/j.jare.2014.11.008. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 42.Gatmaitan Z. C., Arias I. M. Structure and function of P-glycoprotein in normal liver and small intestine. Advances in Pharmacology. 1993;24:77–97. doi: 10.1016/S1054-3589(08)60934-5. [DOI] [PubMed] [Google Scholar]
- 43.Abraham J., Salama N. N., Azab A. K. The role of P-glycoprotein in drug resistance in multiple myeloma. Leukemia & Lymphoma. 2015;56(1):26–33. doi: 10.3109/10428194.2014.907890. [DOI] [PubMed] [Google Scholar]
- 44.Huang H., Zhang X., Huang Z., Zhang Y., Zhou Z. Geniposide reverses multidrug resistance in vitro and in vivo by inhibiting the efflux function and expression of P‑glycoprotein. Experimental and Therapeutic Medicine. 2017;13(2):437–442. doi: 10.3892/etm.2016.4011. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 45.Yamano T., Tsujimoto Y., Noda T., et al. Hepatotoxicity of gardenia yellow color in rats. Toxicology Letters. 1988;44(1-2):177–182. doi: 10.1016/0378-4274(88)90144-0. [DOI] [PubMed] [Google Scholar]
- 46.Yamano T., Tsujimoto Y., Noda T., et al. Hepatotoxicity of geniposide in rats. Food and Chemical Toxicology. 1990;28(7):515–519. doi: 10.1016/0278-6915(90)90122-4. [DOI] [PubMed] [Google Scholar]
- 47.Ding Y., Zhang T., Tao J. S., Zhang L. Y., Shi J. R., Ji G. Potential hepatotoxicity of geniposide, the major iridoid glycoside in dried ripe fruits of Gardenia jasminoides (Zhi-zi) Natural Product Research. 2013;27(10):929–933. doi: 10.1080/14786419.2012.673604. [DOI] [PubMed] [Google Scholar]
- 48.Tian J., Yi Y., Zhao Y., et al. Oral chronic toxicity study of geniposide in rats. Journal of Ethnopharmacology. 2018;213:166–175. doi: 10.1016/j.jep.2017.11.008. [DOI] [PubMed] [Google Scholar]
- 49.Cheng S. H., Tang C., Hui-Fang L. I., Wei J. P. Time-toxicity relationship of geniposide on hepatotoxicity and nephrotoxicity in normal rats. Chinese Journal of Experimental Traditional Medical Formulae. 2016;1:162–165. [Google Scholar]
- 50.Cheng S. H., Zhang Y. Y., Hui-Fang L. I., Wei J. P. Acute hepatotoxicity and nephrotoxicity study of geniposide on jaundice rats. Chinese Journal of Experimental Traditional Medical Formulae. 2015;21:174–178. [Google Scholar]
- 51.Wei J., Zhang F., Zhang Y., et al. Proteomic investigation of signatures for geniposide-induced hepatotoxicity. Journal of Proteome Research. 2014;13(12):5724–5733. doi: 10.1021/pr5007119. [DOI] [PubMed] [Google Scholar]
- 52.Takeuchi O., Akira S. Pattern recognition receptors and inflammation. Cell. 2010;140(6):805–820. doi: 10.1016/j.cell.2010.01.022. [DOI] [PubMed] [Google Scholar]
- 53.Merhisoussi F., Kwak B., Magne D., et al. Interleukin-1 plays a major role in vascular inflammation and atherosclerosis in male apolipoprotein E-knockout mice. Cardiovascular Research. 2005;66(3):583–593. doi: 10.1016/j.cardiores.2005.01.008. [DOI] [PubMed] [Google Scholar]
- 54.Iyer S. S., Cheng G. Role of interleukin 10 transcriptional regulation in inflammation and autoimmune disease. Critical Reviews™ in Immunology. 2012;32(1):23–63. doi: 10.1615/critrevimmunol.v32.i1.30. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 55.Dmitrieva O. S., Shilovskiy I. P., Khaitov M. R., Grivennikov S. I. Interleukins 1 and 6 as main mediators of inflammation and cancer. Biochemistry. 2016;81(2):80–90. doi: 10.1134/s0006297916020024. [DOI] [PubMed] [Google Scholar]
- 56.Fu Y., Liu B., Liu J., et al. Geniposide, from Gardenia jasminoides Ellis, inhibits the inflammatory response in the primary mouse macrophages and mouse models. International Immunopharmacology. 2012;14(4):792–798. doi: 10.1016/j.intimp.2012.07.006. [DOI] [PubMed] [Google Scholar]
- 57.Xiaofeng Y., Qinren C., Jingping H., et al. Geniposide, an iridoid glucoside derived from Gardenia jasminoides, protects against lipopolysaccharide-induced acute lung injury in mice. Planta Medica. 2012;78(6):557–564. doi: 10.1055/s-0031-1298212. [DOI] [PubMed] [Google Scholar]
- 58.Liu H. T., He J. L., Li W. M., et al. Geniposide inhibits interleukin-6 and interleukin-8 production in lipopolysaccharide-induced human umbilical vein endothelial cells by blocking p38 and ERK1/2 signaling pathways. Inflammation Research. 2010;59(6):451–461. doi: 10.1007/s00011-009-0118-3. [DOI] [PubMed] [Google Scholar]
- 59.Shi Q., Cao J., Fang L., et al. Geniposide suppresses LPS-induced nitric oxide, PGE2 and inflammatory cytokine by downregulating NF-κB, MAPK and AP-1 signaling pathways in macrophages. International Immunopharmacology. 2014;20(2):298–306. doi: 10.1016/j.intimp.2014.04.004. [DOI] [PubMed] [Google Scholar]
- 60.Smith J. A., Das A., Ray S. K., Banik N. L. Role of pro-inflammatory cytokines released from microglia in neurodegenerative diseases. Brain Research Bulletin. 2012;87(1):10–20. doi: 10.1016/j.brainresbull.2011.10.004. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 61.Wang J., Hou J., Zhang P., Li D., Zhang C., Liu J. Geniposide reduces inflammatory responses of oxygen-glucose deprived rat microglial cells via inhibition of the TLR4 signaling pathway. Neurochemical Research. 2012;37(10):2235–2248. doi: 10.1007/s11064-012-0852-8. [DOI] [PubMed] [Google Scholar]
- 62.Diomede F., Zingariello M., Cavalcanti M. F. X. B., et al. MyD88/ERK/NFkB pathways and pro-inflammatory cytokines release in periodontal ligament stem cells stimulated by Porphyromonas Gingivalis. European Journal of Histochemistry. 2017;61(2) doi: 10.4081/ejh.2017.2791. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 63.da Silveira Cruz-Machado S., Carvalho-Sousa C. E., Tamura E. K., et al. TLR4 and CD14 receptors expressed in rat pineal gland trigger NFKB pathway. Journal of Pineal Research. 2010;49:183–192. doi: 10.1111/j.1600-079x.2010.00785.x. [DOI] [PubMed] [Google Scholar]
- 64.Santos S. H. S., Andrade J. M. O., Fernandes L. R., et al. Oral angiotensin-(1–7) prevented obesity and hepatic inflammation by inhibition of resistin/TLR4/MAPK/NF-κB in rats fed with high-fat diet. Peptides. 2013;46:47–52. doi: 10.1016/j.peptides.2013.05.010. [DOI] [PubMed] [Google Scholar]
- 65.Yunshi Z., Jing Y., Xian Q., Xing L., Xieqin L., Ganzhu F. Geniposide demonstrates anti-inflammatory and antiviral activity against pandemic A/Jiangsu/1/2009 (H1N1) influenza virus infection in vitro and in vivo. Antiviral Therapy. 2017;22(7):599–611. doi: 10.3851/IMP3152. [DOI] [PubMed] [Google Scholar]
- 66.Rani V., Deep G., Singh R. K., Palle K., Yadav U. C. S. Oxidative stress and metabolic disorders: pathogenesis and therapeutic strategies. Life Sciences. 2016;148:183–193. doi: 10.1016/j.lfs.2016.02.002. [DOI] [PubMed] [Google Scholar]
- 67.Zerón H. M., Flores A. P., Chávez A. A. Oxidative stress in preeclampsia, more than enzymes. Revista Latinoamericana de Hipertensión. 2013;8(2):25–28. [Google Scholar]
- 68.Lv C., Liu X., Liu H., Chen T., Zhang W. Geniposide attenuates mitochondrial dysfunction and memory deficits in APP/PS1 transgenic mice. Current Alzheimer Research. 2014;11(6):580–587. doi: 10.2174/1567205011666140618095925. [DOI] [PubMed] [Google Scholar]
- 69.Roger B., Papin J., Vacher P., et al. Adenylyl cyclase 8 is central to glucagon-like peptide 1 signalling and effects of chronically elevated glucose in rat and human pancreatic beta cells. Diabetologia. 2011;54(2):390–402. doi: 10.1007/s00125-010-1955-x. [DOI] [PubMed] [Google Scholar]
- 70.Yin F., Liu J.-h., Zheng X.-x., Guo L.-x. GLP-1 receptor plays a critical role in geniposide-induced expression of heme oxygenase-1 in PC12 cells. Acta Pharmacologica Sinica. 2010;31(5):540–545. doi: 10.1038/aps.2010.28. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 71.Liu J., Yin F., Zheng X., Jing J., Hu Y. Geniposide, a novel agonist for GLP-1 receptor, prevents PC12 cells from oxidative damage via MAP kinase pathway. Neurochemistry International. 2007;51(6-7):361–369. doi: 10.1016/j.neuint.2007.04.021. [DOI] [PubMed] [Google Scholar]
- 72.Liu J., Zheng X., Yin F., et al. Neurotrophic property of geniposide for inducing the neuronal differentiation of PC12 cells. International Journal of Developmental Neuroscience. 2006;24(7):419–424. doi: 10.1016/j.ijdevneu.2006.08.009. [DOI] [PubMed] [Google Scholar]
- 73.Li N., Zhou H., Tang Q. miR-133: a suppressor of cardiac remodeling? Frontiers in Pharmacology. 2018;9:p. 903. doi: 10.3389/fphar.2018.00903. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 74.Li L., Zhao Q., Kong W. Extracellular matrix remodeling and cardiac fibrosis. Matrix Biology. 2018;68-69:490–506. doi: 10.1016/j.matbio.2018.01.013. [DOI] [PubMed] [Google Scholar]
- 75.Zhao W., Zhao T., Chen Y., Ahokas R. A., Sun Y. Oxidative stress mediates cardiac fibrosis by enhancing transforming growth factor-beta1 in hypertensive rats. Molecular and Cellular Biochemistry. 2008;317(1-2):43–50. doi: 10.1007/s11010-008-9803-8. [DOI] [PubMed] [Google Scholar]
- 76.Aragno M., Mastrocola R., Alloatti G., et al. Oxidative stress triggers cardiac fibrosis in the heart of diabetic rats. Endocrinology. 2008;149(1):380–388. doi: 10.1210/en.2007-0877. [DOI] [PubMed] [Google Scholar]
- 77.Heo J., Lim J., Lee S., et al. Sirt1 regulates DNA methylation and differentiation potential of embryonic stem cells by antagonizing Dnmt3l. Cell Reports. 2017;18(8):1930–1945. doi: 10.1016/j.celrep.2017.01.074. [DOI] [PubMed] [Google Scholar]
- 78.Li N., Zhou H., Ma Z.-G., et al. Geniposide alleviates isoproterenol-induced cardiac fibrosis partially via SIRT1 activation in vivo and in vitro. Frontiers in Pharmacology. 2018;9:p. 854. doi: 10.3389/fphar.2018.00854. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 79.Frey N., Olson E. N. Cardiac hypertrophy: the good, the bad, and the ugly. Annual Review of Physiology. 2003;65(1):45–79. doi: 10.1146/annurev.physiol.65.092101.142243. [DOI] [PubMed] [Google Scholar]
- 80.Dickhout J. G., Carlisle R. E., Austin R. C. Interrelationship between cardiac hypertrophy, heart failure, and chronic kidney disease: endoplasmic reticulum stress as a mediator of pathogenesis. Circulation Research. 2011;108(5):629–642. doi: 10.1161/CIRCRESAHA.110.226803. [DOI] [PubMed] [Google Scholar]
- 81.Inoki K., Kim J., Guan K. L. AMPK and mTOR in cellular energy homeostasis and drug targets. Annual Review of Pharmacology and Toxicology. 2012;52(1):381–400. doi: 10.1146/annurev-pharmtox-010611-134537. [DOI] [PubMed] [Google Scholar]
- 82.Tao J., Zhu W., Li Y., et al. Apelin-13 protects the heart against ischemia-reperfusion injury through inhibition of ER-dependent apoptotic pathways in a time-dependent fashion. American Journal of Physiology-Heart and Circulatory Physiology. 2011;301(4):H1471–H1486. doi: 10.1152/ajpheart.00097.2011. [DOI] [PubMed] [Google Scholar]
- 83.Sun L., Fan H., Yang L., Shi L., Liu Y. Tyrosol prevents ischemia/reperfusion-induced cardiac injury in H9c2 cells: involvement of ROS, Hsp70, JNK and ERK, and apoptosis. Molecules. 2015;20(3):3758–3775. doi: 10.3390/molecules20033758. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 84.Tullio F., Angotti C., Perrelli M. G., Penna C., Pagliaro P. Redox balance and cardioprotection. Basic Research in Cardiology. 2013;108(6):p. 392. doi: 10.1007/s00395-013-0392-7. [DOI] [PubMed] [Google Scholar]
- 85.Pagliaro P., Penna C. Redox signalling and cardioprotection: translatability and mechanism. British Journal of Pharmacology. 2015;172(8):1974–1995. doi: 10.1111/bph.12975. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 86.Scialò F., Sriram A., Fernández-Ayala D., et al. Mitochondrial ROS produced via reverse electron transport extend animal lifespan. Cell Metabolism. 2016;23(4):725–734. doi: 10.1016/j.cmet.2016.03.009. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 87.Hori M., Nishida K. Oxidative stress and left ventricular remodelling after myocardial infarction. Cardiovascular Research. 2009;81(3):457–464. doi: 10.1093/cvr/cvn335. [DOI] [PubMed] [Google Scholar]
- 88.Van Gaal L. F., Mertens I. L., De Block C. E. Mechanisms linking obesity with cardiovascular disease. Nature. 2006;444(7121):875–880. doi: 10.1038/nature05487. [DOI] [PubMed] [Google Scholar]
- 89.Lavie C. J., Milani R. V., Ventura H. O. Obesity and cardiovascular disease: risk factor, paradox, and impact of weight loss. Journal of the American College of Cardiology. 2009;53(21):1925–1932. doi: 10.1016/j.jacc.2008.12.068. [DOI] [PubMed] [Google Scholar]
- 90.Wang S., Gu J., Xu Z., et al. Zinc rescues obesity-induced cardiac hypertrophy via stimulating metallothionein to suppress oxidative stress-activated BCL10/CARD9/p38 MAPK pathway. Journal of Cellular and Molecular Medicine. 2017;21(6):1182–1192. doi: 10.1111/jcmm.13050. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 91.Reaven G. M. Insulin resistance: the link between obesity and cardiovascular disease. Medical Clinics of North America. 2011;95(5):875–892. doi: 10.1016/j.mcna.2011.06.002. [DOI] [PubMed] [Google Scholar]
- 92.Ma Z.-G., Kong C.-Y., Song P., Zhang X., Yuan Y.-P., Tang Q.-Z. Geniposide protects against obesity-related cardiac injury through AMPKα- and Sirt1-dependent mechanisms. Oxidative Medicine and Cellular Longevity. 2018;2018:12. doi: 10.1155/2018/6053727.6053727 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 93.Antoniades C., Antonopoulos A., Bendall J., Channon K. Targeting redox signaling in the vascular wall: from basic science to clinical practice. Current Pharmaceutical Design. 2009;15(3):329–342. doi: 10.2174/138161209787354230. [DOI] [PubMed] [Google Scholar]
- 94.Wang B., Liao P. P., Liu L. H., Fang X., Li W., Guan S. M. Baicalin and geniposide inhibit the development of atherosclerosis by increasing Wnt1 and inhibiting dickkopf-related protein-1 expression. Journal of Geriatric Cardiology. 2016;13(10):846–854. doi: 10.11909/j.issn.1671-5411.2016.10.013. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 95.Koltsova E. K., Ley K. How dendritic cells shape atherosclerosis. Trends in Immunology. 2011;32(11):540–547. doi: 10.1016/j.it.2011.07.001. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 96.Steinman R. M., Hemmi H. Dendritic cells: translating innate to adaptive immunity. Current Topics in Microbiology and Immunology. 2006;311:17–58. doi: 10.1007/3-540-32636-7_2. [DOI] [PubMed] [Google Scholar]
- 97.Paulson K. E., Zhu S. N., Chen M., Nurmohamed S., Jongstra-Bilen J., Cybulsky M. I. Resident intimal dendritic cells accumulate lipid and contribute to the initiation of atherosclerosis. Circulation Research. 2010;106(2):383–390. doi: 10.1161/CIRCRESAHA.109.210781. [DOI] [PubMed] [Google Scholar]
- 98.Wu H., Gower R. M., Wang H., et al. Functional role of CD11c+ monocytes in atherogenesis associated with hypercholesterolemia. Circulation. 2009;119(20):2708–2717. doi: 10.1161/CIRCULATIONAHA.108.823740. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 99.Takeda M., Yamashita T., Sasaki N., et al. Oral administration of an active form of vitamin D3 (calcitriol) decreases atherosclerosis in mice by inducing regulatory T cells and immature dendritic cells with tolerogenic functions. Arteriosclerosis, Thrombosis, and Vascular Biology. 2010;30(12):2495–2503. doi: 10.1161/ATVBAHA.110.215459. [DOI] [PubMed] [Google Scholar]
- 100.Meng X., Zhang K., Li J., et al. Statins induce the accumulation of regulatory T cells in atherosclerotic plaque. Molecular Medicine. 2012;18:598–605. doi: 10.2119/molmed.2011.00471. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 101.Pandiyan P., Zheng L., Ishihara S., Reed J., Lenardo M. J. CD4+CD25+Foxp3+ regulatory T cells induce cytokine deprivation–mediated apoptosis of effector CD4+ T cells. Nature Immunology. 2007;8(12):1353–1362. doi: 10.1038/ni1536. [DOI] [PubMed] [Google Scholar]
- 102.Ait-Oufella H., Taleb S., Mallat Z., Tedgui A. Cytokine network and T cell immunity in atherosclerosis. Seminars in Immunopathology. 2009;31(1):23–33. doi: 10.1007/s00281-009-0143-x. [DOI] [PubMed] [Google Scholar]
- 103.Liao P., Liu L., Wang B., Li W., Fang X., Guan S. Baicalin and geniposide attenuate atherosclerosis involving lipids regulation and immunoregulation in ApoE−/− mice. European Journal of Pharmacology. 2014;740:488–495. doi: 10.1016/j.ejphar.2014.06.039. [DOI] [PubMed] [Google Scholar]
- 104.Lu Y., Zhang J., Ma B., et al. Glycine attenuates cerebral ischemia/reperfusion injury by inhibiting neuronal apoptosis in mice. Neurochemistry International. 2012;61(5):649–658. doi: 10.1016/j.neuint.2012.07.005. [DOI] [PubMed] [Google Scholar]
- 105.Wahlgren N., Moreira T., Michel P., et al. Mechanical thrombectomy in acute ischemic stroke: consensus statement by ESO-Karolinska Stroke Update 2014/2015, supported by ESO, ESMINT, ESNR and EAN. International Journal of Stroke. 2016;11(1):134–147. doi: 10.1177/1747493015609778. [DOI] [PubMed] [Google Scholar]
- 106.Yuan Y., Guo Q., Ye Z., Pingping X., Wang N., Song Z. Ischemic postconditioning protects brain from ischemia/reperfusion injury by attenuating endoplasmic reticulum stress-induced apoptosis through PI3K-Akt pathway. Brain Research. 2011;1367:85–93. doi: 10.1016/j.brainres.2010.10.017. [DOI] [PubMed] [Google Scholar]
- 107.Zhang X., Yan H., Yuan Y., et al. Cerebral ischemia-reperfusion-induced autophagy protects against neuronal injury by mitochondrial clearance. Autophagy. 2013;9(9):1321–1333. doi: 10.4161/auto.25132. [DOI] [PubMed] [Google Scholar]
- 108.Cheng F., Ma C., Sun L., et al. Synergistic neuroprotective effects of geniposide and ursodeoxycholic acid in hypoxia-reoxygenation injury in SH-SY5Y cells. Experimental and Therapeutic Medicine. 2017;15(1):320–326. doi: 10.3892/etm.2017.5395. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 109.Huang J., Upadhyay U. M., Tamargo R. J. Inflammation in stroke and focal cerebral ischemia. Surgical Neurology. 2006;66(3):232–245. doi: 10.1016/j.surneu.2005.12.028. [DOI] [PubMed] [Google Scholar]
- 110.Li F., Li W., Li X., et al. Geniposide attenuates inflammatory response by suppressing P2Y14 receptor and downstream ERK1/2 signaling pathway in oxygen and glucose deprivation-induced brain microvascular endothelial cells. Journal of Ethnopharmacology. 2016;185:77–86. doi: 10.1016/j.jep.2016.03.025. [DOI] [PubMed] [Google Scholar]
- 111.Lai T. W., Shyu W. C., Wang Y. T. Stroke intervention pathways: NMDA receptors and beyond. Trends in Molecular Medicine. 2011;17(5):266–275. doi: 10.1016/j.molmed.2010.12.008. [DOI] [PubMed] [Google Scholar]
- 112.Calabresi P., Centonze D., Pisani A., Cupini L. M., Bernardi G. Synaptic plasticity in the ischaemic brain. The Lancet Neurology. 2003;2(10):622–629. doi: 10.1016/S1474-4422(03)00532-5. [DOI] [PubMed] [Google Scholar]
- 113.Hardingham G. E., Bading H. Synaptic versus extrasynaptic NMDA receptor signalling: implications for neurodegenerative disorders. Nature Reviews Neuroscience. 2010;11(10):682–696. doi: 10.1038/nrn2911. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 114.Yao W., Ji F., Chen Z., et al. Glycine exerts dual roles in ischemic injury through distinct mechanisms. Stroke. 2012;43(8):2212–2220. doi: 10.1161/STROKEAHA.111.645994. [DOI] [PubMed] [Google Scholar]
- 115.Huang B., Chen P., Huang L., et al. Geniposide attenuates post-ischaemic neurovascular damage via GluN2A/AKT/ ERK-dependent mechanism. Cellular Physiology and Biochemistry. 2017;43(2):705–716. doi: 10.1159/000480657. [DOI] [PubMed] [Google Scholar]
- 116.Chen S., Sun P., Zhao X., et al. Gardenia jasminoides has therapeutic effects on L-NNA-induced hypertension in vivo. Molecular Medicine Reports. 2017;15(6):4360–4373. doi: 10.3892/mmr.2017.6542. [DOI] [PubMed] [Google Scholar]
- 117.World Health Organization. Global Report on Diabetes. World Health Organization; 2016. [Google Scholar]
- 118.Lorenzo C., Okoloise M., Williams K., Stern M. P., Haffner S. M., San Antonio Heart Study The metabolic syndrome as predictor of type 2 diabetes: the San Antonio heart study. Diabetes Care. 2003;26(11):3153–3159. doi: 10.2337/diacare.26.11.3153. [DOI] [PubMed] [Google Scholar]
- 119.Levine B., Kroemer G. Autophagy in the pathogenesis of disease. Cell. 2008;132(1):27–42. doi: 10.1016/j.cell.2007.12.018. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 120.Rovira J., Ramírez-Bajo M. J., Banon-Maneus E., et al. mTOR inhibition: reduced insulin secretion and sensitivity in a rat model of metabolic syndrome. Transplantation direct. 2016;2 doi: 10.1097/txd.0000000000000576. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 121.Kim Y. C., Guan K. L. mTOR: a pharmacologic target for autophagy regulation. The Journal of Clinical Investigation. 2015;125(1):25–32. doi: 10.1172/JCI73939. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 122.Xi G., Shen X., Wai C., Vilas C. K., Clemmons D. R. Hyperglycemia stimulates p62/PKCζ interaction, which mediates NF-κB activation, increased Nox4 expression, and inflammatory cytokine activation in vascular smooth muscle. The FASEB Journal. 2015;29(12):4772–4782. doi: 10.1096/fj.15-275453. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 123.Jiang H., Ma Y., Yan J., Liu J., Li L. Geniposide promotes autophagy to inhibit insulin resistance in HepG2 cells via P62/NF‑κB/GLUT‑4. Molecular Medicine Reports. 2017;16(5):7237–7244. doi: 10.3892/mmr.2017.7503. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 124.Ahrén B. Type 2 diabetes, insulin secretion and β-cell mass. Current Molecular Medicine. 2005;5(3):275–286. doi: 10.2174/1566524053766004. [DOI] [PubMed] [Google Scholar]
- 125.Poitout V., Briaud I., Kelpe C., Hagman D. Gluco-lipotoxicity of the pancreatic beta cell. Annales d'Endocrinologie. 2004;65(1):37–41. doi: 10.1016/S0003-4266(04)95628-4. [DOI] [PubMed] [Google Scholar]
- 126.Liu J., Yin F., Xiao H., Guo L., Gao X. Glucagon-like peptide 1 receptor plays an essential role in geniposide attenuating lipotoxicity-induced β-cell apoptosis. Toxicology in Vitro. 2012;26(7):1093–1097. doi: 10.1016/j.tiv.2012.07.004. [DOI] [PubMed] [Google Scholar]
- 127.Nyblom H. K., Thorn K., Ahmed M., Bergsten P. Mitochondrial protein patterns correlating with impaired insulin secretion from INS-1E cells exposed to elevated glucose concentrations. Proteomics. 2006;6(19):5193–5198. doi: 10.1002/pmic.200600137. [DOI] [PubMed] [Google Scholar]
- 128.Guo L. X., Liu J. H., Zheng X. X., Yin Z. Y., Kosaraju J., Tam K. Y. Geniposide improves insulin production and reduces apoptosis in high glucose-induced glucotoxic insulinoma cells. European Journal of Pharmaceutical Sciences. 2017;110:70–76. doi: 10.1016/j.ejps.2017.03.038. [DOI] [PubMed] [Google Scholar]
- 129.Liu J., Guo L., Yin F., Zhang Y., Liu Z., Wang Y. Geniposide regulates glucose-stimulated insulin secretion possibly through controlling glucose metabolism in INS-1 cells. PLoS One. 2013;8(10, article e78315) doi: 10.1371/journal.pone.0078315. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 130.Dor Y., Brown J., Martinez O. I., Melton D. A. Adult pancreatic β-cells are formed by self-duplication rather than stem-cell differentiation. Nature. 2004;429(6987):41–46. doi: 10.1038/nature02520. [DOI] [PubMed] [Google Scholar]
- 131.Bonner-Weir S., Weir G. C. New sources of pancreatic β-cells. Nature Biotechnology. 2005;23(7):857–861. doi: 10.1038/nbt1115. [DOI] [PubMed] [Google Scholar]
- 132.Yao D. D., Yang L., Wang Y., et al. Geniposide promotes beta-cell regeneration and survival through regulating β-catenin /TCF7L2 pathway. Cell Death & Disease. 2015;6(5, article e1746) doi: 10.1038/cddis.2015.107. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 133.Zhang Y., Ding Y., Zhong X., et al. Geniposide acutely stimulates insulin secretion in pancreatic β-cells by regulating GLP-1 receptor/cAMP signaling and ion channels. Molecular and Cellular Endocrinology. 2016;430:89–96. doi: 10.1016/j.mce.2016.04.020. [DOI] [PubMed] [Google Scholar]
- 134.Gnudi L., Coward R. J. M., Long D. A. Diabetic nephropathy: perspective on novel molecular mechanisms. Trends in Endocrinology & Metabolism. 2016;27(11):820–830. doi: 10.1016/j.tem.2016.07.002. [DOI] [PubMed] [Google Scholar]
- 135.Hu X., Zhang X., Jin G., Shi Z., Sun W., Chen F. Geniposide reduces development of streptozotocin-induced diabetic nephropathy via regulating NF-κB signaling pathways. Fundamental & Clinical Pharmacology. 2017;31(1):54–63. doi: 10.1111/fcp.12231. [DOI] [PubMed] [Google Scholar]
- 136.Yang Y. S., Zhang T., Yu S. C., et al. Transformation of geniposide into genipin by immobilized β-glucosidase in a two-phase aqueous-organic system. Molecules. 2011;16(5):4295–4304. doi: 10.3390/molecules16054295. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 137.Guo L., Zheng X., Liu J., Yin Z. Geniposide suppresses hepatic glucose production via AMPK in HepG2 cells. Biological & Pharmaceutical Bulletin. 2016;39(4):484–491. doi: 10.1248/bpb.b15-00591. [DOI] [PubMed] [Google Scholar]
- 138.Hao Y., Liu C., Yin F., Zhang Y., Liu J. 5′-AMP-activated protein kinase plays an essential role in geniposide-regulated glucose-stimulated insulin secretion in rat pancreatic INS-1 β cells. Journal of Natural Medicines. 2017;71(1):123–130. doi: 10.1007/s11418-016-1038-5. [DOI] [PubMed] [Google Scholar]