

Introduction
The neuregulins
Neuregulins, a family of EGF-like signaling molecules, are involved in cell-cell crosstalk and play an important role in development, maintenance and repair of the nervous system, heart, breast and other organs. Independent studies described a ligand for the oncogene ErbB2 (neu, Her2) and factors that stimulated proliferation of Schwann cells, as well as synthesis of receptors for acetylcholine by muscle. These ligands and factors are essentially products of the same gene, referred to by Marchionni M. as neuregulin (NRG-1) [1]. Besides NRG-1 gene, there are other genes that encode related proteins, such as NRG-2 (Don-1, NTAK), NRG-3 and NRG-4. There are few studies that describe NRG-2, −3 and −4 and a complete analysis of their function remains a challenge. However, NRG-2 was recently reported as a component of stress granules (SG), microscopically visible aggregates of translationally stalled messenger ribonucleoprotein complexes that are formed in response to direct stress conditions [2]. Furthermore, it was shown that NRG-2, secreted from astrocytes, bound to ErbB3 on neurons and promoted neuronal survival [3]. NRG-3 was shown to be associated with schizophrenia (SZ) in a Chinese population [4], with bipolar disorder (BD) [5] and with the risk and age of onset of Alzheimer disease (AD) [6]. In addition, NRG-3 has a key function in promoting early mammary morphogenesis and is involved in breast cancer (BC) [7]. NRG-4 expression was decreased in human inflammatory bowel disease samples and mouse models of colitis, suggesting that activation of ErbB4 is altered [8]. An interesting study [9] showed that NRG-4 overexpression prevents high fat diet-induced weight gain and fatty liver and reduces obesity-induced chronic inflammation.
Considering the increasing interest and research focused on NRG-1 in the past years, this review summarizes what is currently known about NRG-1 and its impact on health and disease as well as its current and potential use(s) as a CNS anti-inflammatory agent against inducers of brain inflammation and injury as well as in the treatment of various neurological disorders.
Neuregulin-1: Structure, function and signaling pathways:
NRG-1 gene encodes 21 exons located on human chromosome 8p22–21. Alternative splicing of more than 30 exons in different parts of the NRG-1 transcript generates more than 30 isoforms that can be grouped into six types (I-VI) [10–12] (Figure 1). They are synthesized as transmembrane precursors consisting of either an immunoglobulin-like (Ig) domain or cysteine-rich domain (CRD), an EGF-like domain, a transmembrane domain and a cytoplasmic tail [13]. NRG-1 types I and II are released from the cell surface after protease-mediated proteolytic cleavage and may function as paracrine signals [14]. NRG-1 type III remains tethered to the cell membrane after cleavage and acts as a juxtacrine signal [15]. Full-length type IV spans 1.8 kb and encodes a putative protein of 590 amino acids with a predicted molecular mass of approximately 66 kDa [15]. Further-more, there are two major classes of NRG-1 identified as α and β isoforms. NRG-1α and NRG-1β contain related, but structurally distinct EGF-like domains composed of a common amino terminal segment followed by α or β variant sequences [16]. NRG-1β isoforms predominate in the nervous system while α isoforms are common in mesenchymal cells and are critically important for breast development [14]. Type I NRGα and β isoforms are the predominant isoforms expressed in early embryogenesis, whereas type II and type III are undetectable until the mid-gestation stage [17]. Type I or Heregulin is an acetylcholine receptor activator, type II is a Glial Growth Factor, type III is a sensory and motor neuron-derived factor and type IV is involved in neuronal activity regulation. Functions of types V and VI are not well known and their investigation remains a challenge. Types I, II and III of NRG-1 express in human peripheral blood in addition to neurons, while types IV and V express specifically in the brain [18].
Figure 1:
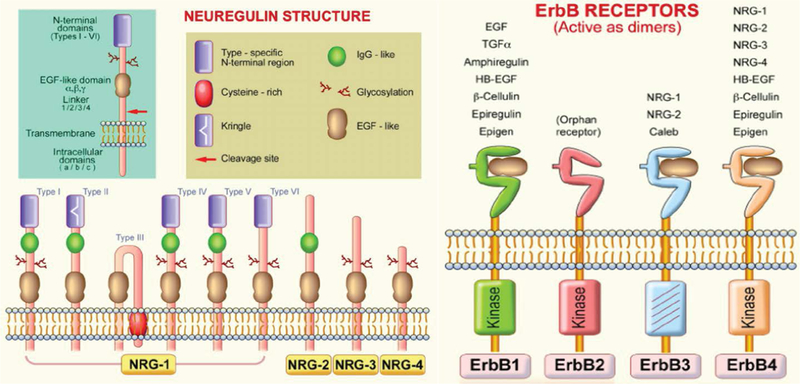
Neuregulin (NRG) and ErbB receptor structures. NRG gene products share a structural characteristic for the extracellular epidermal growth factor (EGF) domain and it differentiates this subfamily from other members of the EGF family. All EGF family members are ligands of ErbB receptors, although with varying affinities, with ErbB3 and ErbB4 being specific NRG-binding receptors. NRG-1 isoforms have been classified in types I-VI on the basis of differences in the NH2-terminal distal region [19].
Neuregulins transmit their signals to target cells by interacting with transmembrane tyrosine kinase receptors of the ErbB (HER) family (Figure 1) [19]. This family includes four members of the epidermal growth factor receptor (EGF-R, ErbB1, ErbB2, ErbB3, and ErbB4). Receptor-ligand interaction induces the heterodimerization of receptor monomers, which in turn results in the activation of intracellular signaling cascades and the induction of cellular responses. In vivo, functional NRG-1 receptors are heterodimers composed of ErbB2 with either an ErbB3, or ErbB4 molecule [20]. This interaction activates various signaling pathways including the mitogen-activated protein (MAP) kinases and phosphatidyl-inositol-3-kinase PI3-K/Akt survival pathways [21,22]. NRG-1 effects are associated with the regulation of both canonical and alternative NF-kB signaling pathways [23]. Increased cardiac mechanical stress leads to upregulation of NRG-1. Interestingly, integrins, a family of receptors expressed on cardiomyocytes, act as sensors for mechanical stress and can non-canonically bind NRG-1 [24]. NRG-1 regulates cell maintenance, differentiation, proliferation, migration, and survival or apoptosis in neuronal and non-neuronal cell types [11].
Many cell types express NRG-1, including microglial and neuronal cells, in response to several stimuli such as tissue injury, blood brain barrier (BBB) dysfunction and neuronal cell damage [25]. NRG-1 has anti-inflammatory properties. For example, recombinant human NRG- 1 decreases the production of superoxide and nitrite by stimulating mouse brain N9 microglial cells [26]. Furthermore, NRG-1 treatment in a cerebral ischemia model induced a decrease in microglial activation and IL-1 mRNA expression. NRG-1 reduced the expression of inflammatory genes associated with leukocyte migration and activation [27]. In contrast, early NRG-1 mutations induced defects in the developing nervous system and embryos died in utero due to cardiovascular failure.
Neuregulin in Invertebrate and Vertebrate Models
In Drosophila central nervous system (CNS), neuronal-glia interactions are necessary for the formation of the longitudinal axon trajectories. The protein Vein, found in drosophila, is similar in structure to NRG and is produced by neurons and leads to the survival of glia [28]. In Caenorhabditis elegans, a simple version of the NRG-ErbB is found, with a single receptor, the product of the let-23 gene and a single ligand encoded by lin-3 [1]. An interesting study developed in fish showed that NRG-1 is found in the axolotl (Ambystoma mexicanum) peripheral nervous system and blastema and is capable of regeneration of amputated limbs, to the point of digit formation [29]. In a zebrafish model, which is hypoxia tolerant, NRG-1 plays a role in establishing the cardiac nervous plexus, while inadequate innervation leads to deficits in cardiac maturation, function and survival [30]. Although flies, worms and fish have been used to understand the functions of simple conserved signaling systems, like the EGF/ErbB pathway, studies in invertebrates have been limited and challenging. Analysis of functions are limited due to the diversification of their corresponding ligands and receptor molecules, unlike in higher organisms [31]. NRG-1 is highly expressed in the developing brain and remains expressed in the adult nervous system [32]. Studies in heterozygous NRG-1 knockout mice showed that the animals exhibit impairments in prepulse inhibition and working memory [33]. However, an interesting study using transgenic mice overexpressing NRG-1 showed that aberrant hyper-signals of NRG-1 also disrupt various cognitive and behavioral processes, but further experiments are necessary to clarify these observations [34]. In a rat model, hypothalamic NRG-1 was transported to the posterior pituitary as the β form. Thus, NRG-1β may function via the regulation of prolactin expression through a paracrine mechanism [35–38]. In addition, a NRG-1 study in non-human primate rhesus monkey assessed the differential expression of NRG-1 and its ErbB receptors in gastrointestinal structures, and found higher expression levels in stomach and small intestine [35]. All these models assessed the complex involvement of NRG-1 in development and in adult life, although many aspects need further investigation.
Neuregulin-1 in Disease
Cardiovascular disorders
During embryogenesis, NRG-1 expression leads to cardiomyocyte proliferation. Studies found that there is an interaction between hyaluronan and Erb signaling during endocardium development [39]. NRG and Notch1 are necessary for the development of the atrioventricular conduction system [40]. In the adult heart, NRG-1 is located in endothelial cells, playing an important role in regulation of survival, hypertrophy, proliferation and interaction between cardiomyocytes [41]. Furthermore, NRG-1 provides protection against apoptosis, myofibrillar disarray, anthracycline-induced cardiomyopathy, and scar formation [42]. These data underline the importance of NRG-1 in cardiovascular development, as well as in cardiac maintenance during the adult life.
Many studies have shown the protective effects of NRG-1 in heart disease. Administration of NRG-1 improved cardiac function via SERCA2a (Sarco/Endoplasmic Reticulum Ca2+ ATPase 2a) and cMLCK (Cardiac Myosin Light Chain Kinase) in a rat heart failure model [43] and significantly enhanced cardiac function in patients with chronic heart failure [44]. Furthermore, NRG-1β treatment leads to iPSC (induced pluripotent cell) differentiation into ventricular-like cardiac cells, which are of preserving cardiac function and tissue viability when transplanted in a mouse model of acute myocardial infarction [45]. NRG-1 and ErbB receptors are expressed in ex vivo skeletal muscle vascular endothelial cells; the addition of NRG-1 induces angiogenesis [46]. Further-more, elevated expression of NRG-1 increased the number of microvessels formed in the ischemic myocardium [47]. Although NRG-1 plays an important role in cardiac development and in tissue recovery following heart ischemia, potentially including it in adjunctive therapy in the treatment of cardiac diseases remains a challenge.
Cancer
In head and neck cancer, tumor cells control the amount of cell-surface NRG-1 available for cleavage and ErbB3 activation [48]. Constitutively active forms of HER/ErbB receptors have been reported in tumors and their activation may occur through their overexpression in the plasma membrane or through their structural alterations [49]. In the invasive mucinous adenocarcinoma of the lung, molecular lesions of the NRG-1 gene leads to aberrant activation of ErbB2/ErbB3 signaling [50]. NRG-1 treatment induced cancer stem cell (CSC) characteristics in breast cancer (BC) cell lines in vitro [51]. Furthermore, NRG-1 produced by BC cells can enhance transendothelial migration and intravasation, due to induction of jagged-1 (JAG1), an important component of the Notch pathway [52]. In BC, NRG-1-dependent activation of HER3 induces primary resistance to trastuzumab in HER2-[53] overexpressing breast cancer cells [54]. In gastric tumors, fibroblasts secreted NRG- 1, which in turn increased the stem cells self-renewal and its overexpression was correlated with gastric cancer prognosis [55]. NRG-1 was demonstrated to be a prominent growth factor during cultivation of spheroids in vitro from non-small cell lung cancer specimens and pleural effusion [53]. Another study showed that elevated levels of ErbB3 on ovarian cancer cells and corresponding ligand NRG-1 in the omentum allowed for tumor cell metastasis and growth in the omentum [56]. Although it has been demonstrated that NRG-1 induces tumor growth, stem cell characteristics, metastasis and chemo resistance, an engineered bivalent NRG-1β (NN) could protect against cardiotoxicity in cancer patients treated with doxorubicin [57]. Studies on circumstances under which NRG-1 functions to enhance regeneration and tissue repair and enhancement of tumor cell growth are urgently needed to elucidate the multidimensional aspects of its functional role in health and disease.
Neuromuscular disease and development
Neuromuscular disorders refer to diseases of the nerves that control muscles and communication between nerves and muscles [58]. Myelin plays an essential role in accurate neuromuscular communication and conduction of electrical impulses by axons in both the central nervous system (CNS) and peripheral nervous system (PNS). PNS myelin development depends on axonal signals which are provided by NRG1/ErbB receptors [59]. Similarly, NRG-1 plays a significant role in Hirchsprung disease, in which mutations in NRG-1 gene can lead to absence of ganglion cells along the intestines, leading to obstruction of bowel function [60]. These ganglion cells are part of the enteric nervous system (ENS), for which studies have shown that NRG-1 also plays an important role in [61,62].
During embryonic development, the formation of muscular tissue is carried out through a process known as myogenesis. Recently, it was shown that NRG-1β plays a protective role to improve impaired myogenesis through PPARγ activation and transcriptional activity of NF-κB [63]. This was the first insight into the function of NRG-1β in regulating PPARγ/NF-κB signaling during myogenesis using an in vitro whole serum-based sepsis model.
Understanding of this process is essential for designing future approaches for the treatment of skeletal muscle diseases and the prevention of or recovery from muscle loss in situations such as cachexia, muscle wasting, sarcopenia and other neuromuscular degenerative diseases.
Neurological disorders
Cerebrovascular diseases:
Stroke, transient ischemic attack and intracerebral hemorrhage are cerebro-vascular diseases which affect blood supply to the brain [15]. The lack of blood flow during a stroke leads to a complex pathophysiological response, resulting in neural injury. The mechanisms that lead to neural cell loss and injury include cytotoxicity, free radical release and inflammatory changes [18]. The inflammatory response enhances the ischemic injury, but also promotes recovery. Following a stroke, immune cells release proinflammatory cytokines and free radicals which increase the inflammatory response and contribute to the neuronal injury post ischemia [18]. Treatment failure may be due to the intricate pathophysiological response to ischemic injury. Targeting multiple molecules in this pathway remains a treatment challenge.
During ischemic brain injury, NRG-1 provides neuro-protection. This effect, demonstrated by in vitro and in vivo models, is achieved through NRG-1’s stimulation of the PI3-kinase/Akt pathway. Oxygen glucose deprivation (OGD) is an in vitro method used to stimulate neuronal cell injury and imitates the pattern that occurs following ischemic injury in vivo [64]. Rat B35 neurons subjected to (OGD)/reoxygenation were treated with NRG-1 and showed significant increases in neuronal survival [65]. Different concentrations of NRG-1 added to neuronal culture at reoxygenation induced protection against OGD-induced neuronal apoptosis [66]. These data show that NRG-1 reduced cell death following OGD in a dose-dependent manner, by activating ErbB4 receptors and GABAergic transmission [23]. Injury to oligodendrocyte progenitor cells (OPCs) caused by hypoxia plays a crucial role in white matter injury. A study observed NRG-1’s potential to influence the survival of OPCs damaged by OGD. OPCs were exposed to OGD, then treated with NRG-1 and the results showed that NRG-1 inhibited OGD-induced apoptosis and increased the survival rate of OPCs [67].
In vivo, NRG-1’s neuroprotective capabilities were tested via permanent middle cerebral artery occlusion (pMCAO) in an ischemic rat model. NRG-1 treatment prior to pMCAO significantly decreased infarction volume. MK-801, a N-methyl-D-aspartate (NMDA) receptor antagonist, shows similar outcomes to NRG −1 treatment, reducing neuronal death [67]. Co-administration of NRG-1 and MK-801 showed synergistic effects on the reduction of infarct following pMCAO. These findings indicate that NRG-1 has the capability of inhibiting neuronal damage in rat pMCAO models and that combination treatment of NRG-1 with MK-801 enhance the neuroprotective effects. In transient middle cerebral artery occlusion (tMCAO) model, NRG-1 given prior or13.5 hours after the onset of ischemia in rats reduced cortical damage and improved neurological outcomes [68]. Current treatment of stroke relies on TPA, which is focused on reducing infarct and inflammation associated with the infarct.
Brain trauma:
Traumatic brain injury (TBI) physical symptoms include loss of consciousness, headache, fatigue and dizziness. The cognitive symptoms include mood swings, memory or concentration problems, and feelings of depression or anxiousness. Earlier studies showed that administration of NRG-1 prior to brain trauma in a mouse model improved retention of spatial memory, suggesting that NRG-1 may improve some functional outcomes after brain injury [69]. Often, BBB disruption occurs following TBI, leading to brain edema, inflammation, and neuronal death. BBB compromised integrity exposes the brain tissue to substances that are present in blood vessels and it disrupts the ionic gradients necessary for proper neuronal function, which may lead to acute and long-term CNS dysfunction [70]. Studies suggest that NRG-1 has beneficial effects on endothelial permeability and BBB permeability following experimental trauma in mice and may have neuroprotective potential during CNS injury [71]. Along with injuring neurons, brain trauma also damages the axons, often resulting in diffuse axonal injury. Axonal injury is one of the most important pathological features following TBI [41]. NRG-1 plays an integral role in axon physiology by influencing the survival and migration of Schwann cell precursors as well as remyelination and functional recovery after injury [42]. In an animal model of spinal cord injury, exogenously administered NRG-1 enhanced the proliferation and differentiation of spinal neural precursor cells into oligodendrocytes, enhanced axonal preservation, and decreased astrogliosis and tissue degeneration [43]. Using a severe sciatic nerve injury model, a study has shown that in NRG-1 mutant mice, axons remyelination was impaired for 2 months after the nerve transection and anastomosis. However, by 3 months post-injury, the axons returned to normal. These data suggest that NRG-1 promotes nerve repair in the early phases of injury [72]. NRG-1 also induced bone pain-like behavior in rats by induction of and activation of ErbB2, Akt-1 and p38MAPK, which could be blocked by ErbB2 inhibitor [73]. The mechanisms involved in nerve repair in the late phases of injury remain to be investigated.
Neuropsychiatric disorders:
Neuropsychiatric Disorders (ND) affects around 450 million people worldwide. Those inflicted by these diseases tend to have memory, pattern separation, and reward processing problems [74–76]. ND impairs executive functioning, emotional regulation, and social facilitation. These neurobehavioral functions occur because of circuits that reside in the prefrontal cortex [77]. By 2020, it is predicted that ND will be the second highest cause of global disease burden [78].
Studies are recently investigating the pivotal role of NRG-1 and inflammation in the development of the cognitive disorders. NRG-1 plays an important role in brain plasticity and development. Moreover, NRG-1 has been found in multiple regions of the adult brain and is involved in the regulation of neurotransmission [79]. ND occurs in individuals with neurodevelopment problems and limited plasticity. In vivo studies using mouse models have focused on increasing and decreasing the NRG-1 and ErbB network levels. Results showed many behavioral deficits that suggested modified signaling intensity, which corresponds to a pathophysiological mechanism of psychosis and ND [80]. Inflammation can be defined as a part of the immune response that occurs after a type of physical injury or microbial invasion [81]. Research suggests that inflammation may play an important role in cognitive disorders and needs to be studied in greater detail in ND patients [82].
The main ND that present a correlation with inflammation and NRG-1 are Schizophrenia (SZ), Bipolar Disorder (BD), and Alzheimer’s Disease (AD). Evidence for increased inflammation has been found in a study that included 3,952 adolescents diagnosed with BD, Autism Spectrum Disorder (ASD), and SZ. Proinflammatory markers (PIMs) were measured and showed to be elevated in those diagnosed with ND. However, this study suggested that future research is needed, using a larger population with one specific diagnosis [76].
SZ is an extremely debilitating disorder that has afflicts about 1% of the population [11]. Symptoms of this disorder include a misinterpretation of reality, hallucinations, disordered thinking, and an overall impairment of daily functioning that results from behavior changes. NRG-1 has been one of the most extensively studied genes in SZ [80]. One study conducted in 2017 examined the levels of NRG-1 and EGF in treatment-resistant schizophrenic patients and found decreased or abnormally expressed levels of these proteins [79]. While past research has examined the levels of NRG-1 in the brain [83], future research could look toward understanding the mechanisms regulating NRG-1 expression and function and possible use of NRG-1 as a treatment for ND disorders. NRG-1 has also been used to suppress the induction and the expression of long-term potentiation in the hippocampus. In another mutant mice study, looking at the influence of NRG-1 and ErbB4, researchers observed a trend of loss of function in specific genes that specifically show to be risk factors for SZ [11]. This finding raises the question of a possible disease-related pathophysiological significance, perhaps inflammation. A study conducted on a group of post-mortem human schizophrenic brains have shown differential expression of NRG-1 mRNA and protein in many different regions of the brain, most notably seen in dorsolateral prefrontal cortex and the hippocampus [79]. This differential expression of these pathways suggests that NRG-1 is an interesting target to examine for SZ research.
In BD, individuals have an impairment of multiple cognitive abilities and functions, which corroborates a potential role of neuroinflammation [84]. Macrophages, or microglia, located in the brain, become activated in response to any kind of damage, leading to inflammation [81]. Inflammation is responsible for influencing the shaping process of synaptogenesis (the formation of new synapses) and the overall reduction of neurons [81]. The link between dysfunctions, cognitive impairment, medical comorbidity and premature mortality seen in BD patients can be explained by immune system signaling the microglia [81]. In both BD and SZ studies, NRG-1 deficiency has resulted in behavioral deficits such as working memory impairments in both disorders [74]. There has been much overlap in the diagnosis of BD and SZ because of the similarity they have. Research has found that over 50% of those diagnosed with BD have some other kind of comorbidity [81–84], which makes the differentiation between BD and SZ difficult for clinicians. The biggest difference found between the two ND is in regard to the neuropsychological deficits and the time at which they present themselves: SZ individuals exhibit dysfunction preceding the onset of the illness, which becomes greater during the first few years of being diagnosed, while BD patients show cognitive development pre-morbidly and demonstrate first episodes that are extremely intense [84]. These key differences of the timing of the presentation of symptoms have been likened role of inflammation in BD and SZ [85], along with abnormal sequences and patterns of NRG-1 and other proteins. A study suggested that NRG-1 deficiency regulates microglial cells that are stress induced [74]. Thus, NRG-1 could be used as treatment because of its anti-inflammatory properties.
Alzheimer’s disease:
Studies on Alzheimer’s disease (AD) have shown that neuroinflammation plays a role in this disease [12,86,87], thus NRG-1 is considered a candidate for its treatment [6]. NRG-1 is responsible for creating extremely complex networks of signaling proteins with many differing expressions and divergent functions during the development of an adult nervous system [86]. During inflammation, these proteins alter their function. Incorrect or low expression of these proteins lead to manifestation of AD symptoms [74]. NRG- 1 deficiencies have been shown to provide evidence for the neuropathology of psychosis [85]. In AD brains, ErbB4 immunoreactivity was shown to colocalize with apoptotic Bax signal in apoptotic hippocampal neurons. Thus, the upregulation of immunoreactivity of NRG-1 receptor ErbB4, may be involved in AD progression [32]. Another study showed that NRG-1 upregulated the expression of anti-apoptotic protein Bcl-2, demonstrating the neuroprotective potential of NRG-1 in AD [88]. The soluble NRG-1 plasma levels were significantly higher in mild and moderate AD patients compared to control patients, indicating that NRG-1 could be used as a marker for early diagnosis of AD [89]. Due to its localization in the brain areas relating to auditory and visual hallucinations and delusions, it has been hypothesized that the presence of NRG-1 may interact with specific inflammatory pathological processes that have been shown to be associated with Late Onset Alzheimer’s Dementia (LOAD) [85]. Furthermore, a study suggests that NRG1 plays a role in increasing the genetic risk to positive symptoms of psychosis in a proportion of LOAD families [85]. However, these results are controversial, since a different study [90] found no correlation between NRG- 1 variants and delusions, hallucinations, psychosis, or elation/mania in an AD cohort. Further investigations are required, and large longitudinal patient cohorts should be used.
Neuregulin and Infectious Diseases
Malaria, caused by Plasmodium (with P. falciparum being the deadliest), is a devastating infectious disease causing approximately 207 million cases worldwide and 627,000 deaths a year, mainly in children under 5 years of age living in sub-Saharan Africa, [91]. In the trophozoite-stage, intra erythrocyte (RBC) parasite proteins are transferred to the surface of the RBC where they play a role in adherence to the host’s endothelium [92,93], destruction of RBC’s and release of cytotoxic heme moieties into circulation which together with interactions with host cells [94,95] mediate severity of the disease.
Approximately 1% of P. falciparum infections results in cerebral malaria (CM), a brain encephalopathy associated with malaria especially in children [91]. In adults in South-East Asia, CM accounts for 50% of the malaria deaths, as they not only suffer from encephalitis, but also have multiple organ failure [96]. CM is clinically defined as having P. falciparum parasitemia and unarousable coma, ruling out any other causes, such as meningitis.
Recently there has been an awareness of the burden of varying neurological deficit in survivors [97]. CM-related neurological sequelae are more severe in children than in adults. Children living in Africa with severe neurological sequelae often die within a few months of discharge [98]. In adults, coma depth and duration along with multiple convulsions are independent risk factors for neurological sequelae [98]. These include cranial nerve lesions, neuropathies, and extrapyramidal disorders [99].
The pathogenic mechanisms leading to fatal human CM are multifactorial and involve alterations in cytokine and chemokine expression, localized inflammation and microvascular injury that results in the loss of blood brain barrier (BBB) integrity. Two hypotheses have been proposed to explain dysfunction of the BBB and mortality associated with CM [100]. First, the sequestration hypothesis, proposes that infected red blood cells (iRBCs) bind to endothelial cells and accumulate within brain microvessels [101–103] obstructing blood flow, resulting in low tissue perfusion, compromised oxygenation and brain tissue damage. Second, the immunopatho-logical “cytokine” hypothesis, suggests that an exaggerated host immune response to Plasmodium infection and the adherence of parasitized erythrocytes to endothelial cells promotes the release of pro-inflammatory mediators and other cytotoxic molecules responsible for compromising BBB cellular components, resulting in edema, access of toxins to sensitive brain tissue, neuronal cell damage, coma and death [104–106]. However, pathogenesis of CM has not been shown to be exclusive to any one hypothesis but involves a combination of pathophysiological features, including parasite sequestration in the brain, metabolic disturbances and host inflammatory responses that mediate CM pathogenesis [104–106].
There are pathophysiological similarities between CM, acute ischemia syndrome (AIS) and TBI, including an exaggerated host expression of pro-inflammatory factors. This inflammatory response leads to increased microvascular endothelial activation with upregulation of adhesion molecules, glial activation, focal inflammation and activation of apoptotic pathways, which can eventually lead to brain damage, coma, and death. Rec-ognizing fundamental similarities shared by CM patho-genesis and both AIS and TBI, an experimental cerebral malaria (ECM) model (Plasmodium berghei ANKA in C57BL/6 mice) was used for the first time to investigate the role of NRG-1 in ECM prognosis. The results indicated that ECM was associated with depletion in circulating levels of NRG-1β [107]. Indeed, intravenous infusion of NRG-1 at onset of ECM attenuated ECM mortality by stimulating a robust anti-inflammatory response, while reducing pro-inflammatory factors. Also noted was the reduction of accumulation of infected erythrocytes in brain microvessels, reduction of brain tissue damage, and reduction of total mortality.
We recently showed that NRG-1 protects human brain microvascular endothelial cells (hCMEC/D3) and astrocytes from cell death induced by CXCL10 and heme (a cytotoxic by product released by damaged infected and non-infected erythrocytes) in vitro [21,22] In addition, NRG-1 improved heme-disrupted BBB integrity in an in vitro BBB model consisting of hCMEC/D3 and human astrocytes. In the, cortex and hippocampus of mice with advanced stages of ECM, the ErbB4 protein is inactivated through dephosphorylation, indicating a diffuse distribution in regions of tissue injury. However, exogenous infusion of NRG-1 increased phosphorylated ErbB4 levels in the cortex and hippocampus of infected mice; this subsequently reduced STAT3 activation (a typical pathogenic pathway in CM) and increased AKT activation. These findings were consistent with Lok’s report [108] which showed that NRG-1 increases pAKT in order to enhance in vitro survival of endothelial cells. Overall, it seems that NRG-1 protects against apoptosis and cell death of two major BBB components, human brain microvascular endothelial cells and astrocytes (Figure 2). Thus, NRG-1 attenuates mortality associated with CM through activation of ErbB4/AKT and inactivation of STAT3 signaling pathways. These findings suggest that adjunctively augmenting NRG-1 during ECM therapy may be an effective therapeutic approach to reduce CM-induced CNS tissue injury. To date no other studies with other infectious diseases such as Zika, toxoplasmosis, HIV/AIDS and others with reported deleterious effects on the CNS has been reported. It will be interesting to determine whether NRG will attenuate the CNS injury associated with these diseases and their respective neuropathologies or sequelae.
Figure 2:
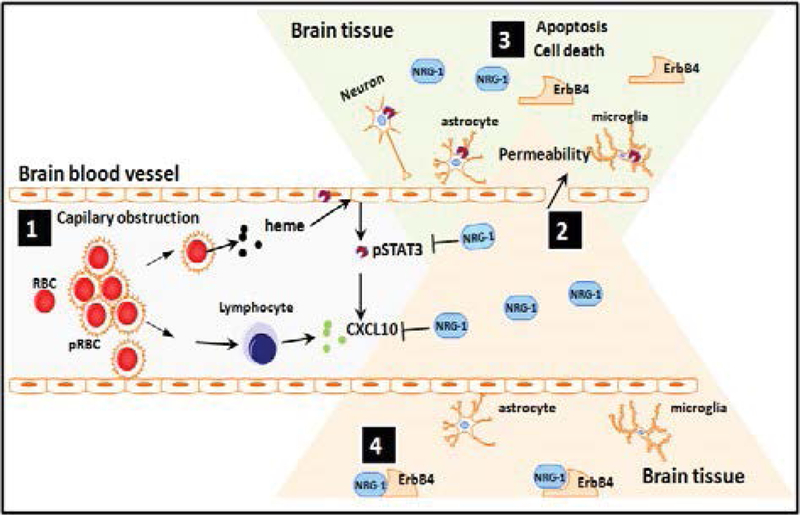
Proposed model demonstrating protective role of NRG1 against cerebral malaria (CM) pathogenesis. During CM pathogenesis, some P. falciparum-infected erythrocytes (pRBC) lyse and release heme while others bind to brain vascular endothelial cells via adhesion molecules (ICAM, VCAM, etc) causing capillary obstruction and activation of endothelial cells as well as release of pro-inflammatory and angiostatic factors such as CXCL10 and activation of STAT3 (1). Downstream of the capillary obstruction, endothelial cells, astrocytes, microglia and neurons located in the penumbra are at risk of undergoing apoptosis which results in increased permeability of BBB (2) due to dead and dying cells in irreversibly damaged brain tissue regions (3). After administration of adequate amounts of NRG-1, cells in the penumbra can be rescued from apoptosis. It is proposed that this might be the basis for effective attenuation of mortality and associated neurological sequelae in patients with CM (4).
Neuregulin: Drug or Target?
Animal studies have shown that myocardial NRG-1 is activated in response to ischemia. Furthermore, when NRG-1β is used as treatment in animal models of cardiac failure, studies have shown that it improves heart function [109]. In an in vitro study, NRG-1β treatment within the period of deoxygenation and glucose deprivation, significantly increased oligodendrocyte type2 astrocytes progenitor’s survival and decreased apoptosis [110]. There is also concern that NRG treatment might exert “off-target” effects by nature of its mitogenic properties as a growth factor, and the well-es-tablished role of ErbB2 as an oncogene in a variety of malignancies. To address this issue, a bivalent version of NRG was engineered to preferentially target ErbB4 homodimer activation, thereby preventing doxorubicin-induced cardiomyocyte death without activating or potentiating cancer cell signaling [76,78]. Further studies are clearly warranted to determine whether this or other modified ligands, or alternative delivery methods are needed to increase the therapeutic window for NRG [111]. When NRG-1 was administered for 10 days in patients with heart failure, it improved cardiac function when patients were evaluated after 30 days (ChiCTR-TRC-00000414) [44]. When NRG-1 was infused for 11 days, cardiac function was improved for 12 weeks (Australian New Zealand Clinical Trials Registry, anzctr.org.au Identifier: ACTRN12607000330448) [112]. Currently, clinical trials that use NRG-1 in heart failure are: NCT01214096, NCT01439789, NCT01439893, NCT01251406, NCT03388593, but no results have been yet posted.
NRG-1 has neuroprotective effects against ischemic stroke. Studies suggest that there is a therapeutic window of opportunity between the onset of ischemia and irreversible neuronal death and that irreversible injury is not complete until about 6 hours after ischemia [113]. However, if a single bolus injection of NRG-1 was administered at either 0, 4 or 12 hours after reperfusion following middle cerebral artery occlusion [113], a significant decrease in the subcortical and cortical lesions in NRG-1 treated rats was shown. These results contribute to NRG-1’s potential use in therapy in focal cerebral ischemia. When NRG-1β was administered, the ischemia-induced apoptosis decreased, the abnormal morphological structures of nerve cells were ameliorated, the integrity of the BBB was restored, and infarct volume was reduced. At the same time, neurological function was significantly recovered [114]. Cimaglermin (NRG-1β) treatment significantly enhanced recovery after stroke, inducing axonal sprouting and synapse formation. These data suggested that Cimaglermin represents a potential candidate for stroke treatment [115].
Because of its critical contribution, BBB integrity is a key potential therapeutic target in the treatment of the acute phase of brain trauma. A study was carried out in which mice were subjected to controlled cortical impact (CCI) under anesthesia and BBB permeability was assessed by measuring Evans blue dye extravasation [86]. NRG-1 was administered intravenously for 10 minutes following trauma and Evans blue dye extravasation decreased by 35% 2 hours post trauma. The results showed that NRG-1 decreases BBB injury in a model of TBI. This data provides evidence that NRG-1 has a beneficial effect on the barrier function of the brain’s microvasculature following trauma. These various studies demonstrate NRG-1’s potential role in the therapeutic strategies for traumatic brain injury. Currently, NRG-1 is undergoing clinical trials for use against TBI (Clinical-Trails.gov NCT01258387 and NCT01944683).
In a rat model of Charcot-Marie-Tooth neuropathy, early NRG-1 treatment promoted Schwann cell differentiation, preserved axons and restored nerve function [116]. In a mouse model of spared nerve injury, NRG-1 inhibited neuropathic pain in a dose-dependent manner. However, in a formalin-induced pain model, NRG-1 was found to aggravate pain. These findings may provide different approaches for neuropathic pain in different injury types [117]. Conversely, monoclonal antibody Herceptin (Trastuzumab) blocked NRG receptor ErbB2 and increased axonal regeneration in a rat model of acute or chronic nerve injury. However, these effects were independent from NRG pathway. These results raise the possibility of using targeted therapies to improve outcomes of peripheral nerve injury [118].
NRG-1 could potentially significantly increase the effectiveness of current anti-malarial therapy against human CM. Despite appropriate anti-malarial treatment, mortality associated with CM remain as high as 30% while up to 20% of survivors experience neurological complications [96]. Thus, anti-malarial drugs are clearly not adequate for saving lives of CM non-survivors and adjunctive therapy is needed. Adjunctive therapy has been proposed based on pathophysiology studies conducted with murine ECM. The purpose of these adjunctive therapies were to shorten coma and decrease fatality as well reduction in the rate and severity of neurological sequelae [98]. Some of these therapies have been deleterious but others are currently being explored, NRG-1 being one of them.
Evidence from genetic, transgenic and post-mortem studies have strongly suggested that altered NRG-1/ ErbB4 signaling is involved in SZ susceptibility. NRG-1/ ErbB4 signaling has interactions with GABAergic, glutamatergic and dopaminergic neurotransmissions that are involved in SZ. Identifying the targets in NRG-1/ErbB4 signaling and interactive pathways will provide opportunity for development of antipsychotics with specific efficacy and fewer side effects [119].
Neuregulin can be used as treatment in other diseases. Neuregulin regulates glucose and lipid homeo-stasis. An engineered fusion protein using NRG-1 and Fc domain of human IgG1 (NRG1-Fc) exhibits extended half-life in circulation and improved potency in receptor signaling. NRG1-Fc treatment lowered blood glucose, improved insulin sensitivity and suppressed food intake in obese mice [120]. In a different study, NRG-1 treatments improved glucose tolerance in Db/Db mice, through inhibition of hepatic gluconeogenesis [121].
Neuregulin Signaling Blockade in Cancer
Studies have shown that in lung cancer animal models, inhibition of NRG-1 signaling leads to reduced tumor growth and enhanced intensity and duration of the response to chemotherapy [122]. Furthermore, clinical trials especially used drugs that targeted the ErbB (HER) receptors. In a Phase IV clinical trial that enrolled patients with advanced HER+ carcinomas, Lumretuzumab (anti-HER3 monoclonal antibody) was used in combination with EGFR-blocking agent Erlotinib or Cetuximab. The toxicity of those drugs was manageable, but there was no evidence of meaningful clinical benefit [123]. However, Duligotuzumab, a dual-action antibody that blocks ligand binding to human HER3 and EGFR, in combination with chemotherapy (Cisplatin-5-Fluorouracil or Carboplatin-Paclitaxel) was used for patients with recurrent or metastatic squamous cell cancer of the head and neck. This treatment combination showed encouraging activity, 67% of the patients confirming partial or complete response [124].
Clinical trials targeting NRG pathways in cancer use MM-121 (antibody targeting ErbB3) in combination with chemotherapy in Heregulin-positive non-small cell lung cancer (NCT02387216). A Phase 2 trial of MM-141 (antibody against Her3 and IGF-1) and Nab-paclitaxel and Gemcitabine in pancreatic cancer is active, but not recruiting yet (NCT02399137). In ovarian cancer, study #NCT01447706 tried to determine whether the combination of MM-121 and paclitaxel has better results than paclitaxel alone. Results showed that overall survival was 13.7 months for Pax + MM141 compared to Pax alone (10.17 months). In cancer, targeting NRG pathway seem to improve survival and prognosis.
Conclusions
Studies have shown the importance of NRG and its corresponding receptors in organ development and maintenance, as well as in many disorders. However, the functions of NRG-1 and the pathways it mediates are not completely understood; Figure 3 shows proposed NRG-1 pathways. In animal models, homozygous deletion of the NRG-1 gene is devastating, leading to death in utero, overexpression of NRG-1 impacts negatively on various cognitive and behavioral processes, suggesting that further investigations are necessary to determine the NRG-1 dose that is ultimately beneficial for patients.
Figure 3:
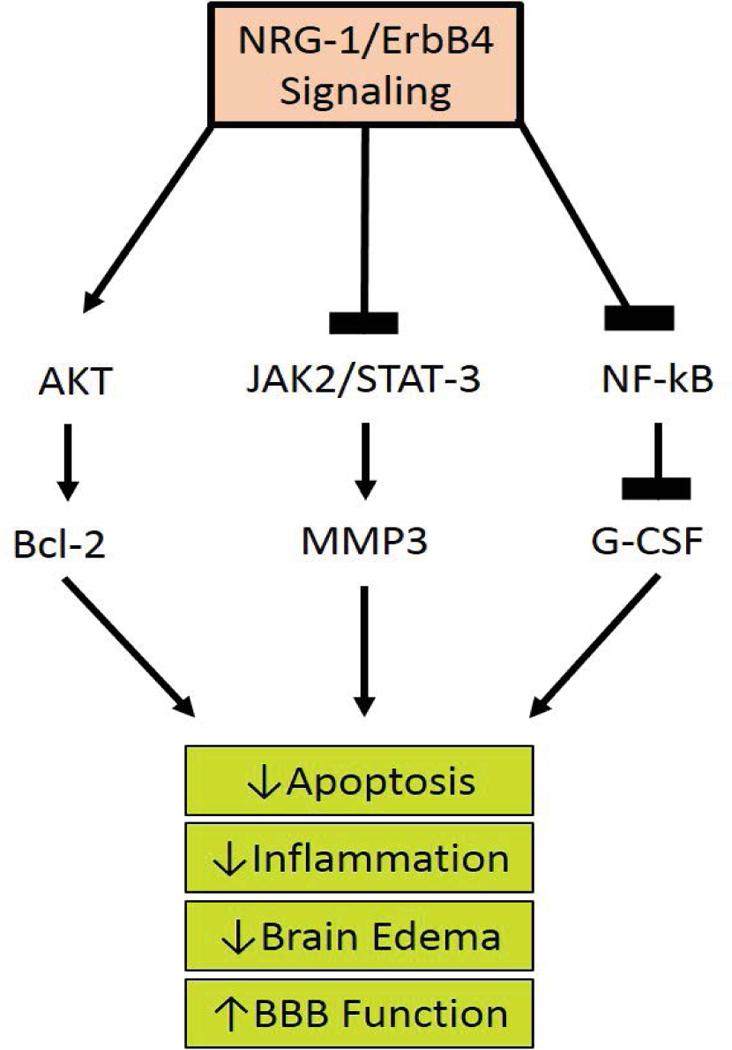
Proposed Neuregulin-1 Pathways. The ligand interaction with ErbB 1,3, and 4 increases their affinity and induces heterodimerization of ErbB1–4, activating the tyrosine kinase domain. This allows the cytoplasmic region of the ErbB to become phosphorylated. The phosphorylated tyrosine residues recruit various adaptors/effectors that induce intracellular signals leading to the survival of the cell by lowering apoptosis, inflammation, edema, and ultimately restoring BBB function.
Although NRG-1 plays a protective role during development and adult life, its effects in disease vary. NRG-1 improves cardiac function in patients with chronic heart failure and increases tissue viability and angiogenesis in animal models of heart ischemia. Furthermore, NRG-1 is neuroprotective during episodes of brain ischemia and injury, as well as in AD. Its elevated plasma levels in patients with mild or moderate AD suggests that NRG-1 can be used as a marker for early diagnosis. However, the role of NRG-1 in cognitive disorders remains to be fully understood. In ECM, we demonstrated that NRG- 1 circulating levels diminished, while its infusion led to a reduced mortality due to a strong anti-inflammatory response and a reduction of parasitic RBC accumulation in microvessels and decreased tissue damage. Furthermore, we showed that NRG-1 protects brain microvascular cells, astrocytes and BBB integrity from heme, a product released by damaged erythrocytes. In addition, we demonstrated that NRG-1 effects on ECM were induced through ErbB4/AKT and STAT3 pathways. Although NRG-1 has protective roles in noninfectious cardiovascular and neurological disorders, as well as in infectious diseases, such as malaria, its role in cancer is completely different. It has been demonstrated that in cancer, NRG-1 induces tumor growth, stem cell self-renewal and chemoresistance.
Due to its multiple functions in health and disease, NRG-1 could be used either as treatment or as a target in clinical trials. Future studies will open new opportunities of research and potentially increase the success of using NRG-1 in treatment regimens.
Acknowledgement
JKS is funded by the National Institute of Neurological Disorders and Stroke NIH/NINDS R01 NS091616 and R21 TW006804 as well as 8G12MD007602 from the National Institute of Minority Health and Health Disparities (NIMHD). The funding bodies had no involvement in the design of the study and collection, analysis, and interpretation of data and in writing the manuscript. The content is solely the responsibility of the authors and does not necessarily represent the official views of the NIMHD or the NIH.
Abbreviations
- AD
Alzheimer Disease
- AIS
Acute Ischemic Stroke
- ASD
Autism Spectrum Disorder
- BBB
Blood Brain Barrier
- BC
Breast Cancer
- BD
Bipolar Disease
- CCI
Controlled Cortical Impact
- CM
Cerebral Malaria
- cMLCK
Cardiac Myosin Light Chain Kinase
- CNS
Central Nervous System
- CRD
Cysteine-rich Domain
- CSC
Cancer Stem Cell
- ECM
Experimental Cerebral Malaria
- EGF
Epidermal Growth Factor
- hCMEC
Human Brain Microvascular Endothelial Cell
- HIV
Human Immunodeficiency Virus
- IG
Immunoglobulin
- IL
Interleukin
- iPSC
Induced Pluripotent Stem Cell
- iRBC
Infected Red Blood Cell
- JAG1
Jagged 1
- KDa
Kilo-Dal-tons
- LOAD
Late Onset Alzheimer’s Dementia
- MAP
Mitogen-activated protein
- ND
Neuropsychiatric Disorders
- NMDA
N-methyl-D-aspartate
- NRG
Neuregulin
- NTAK
Neural and Thymus-Derived Activator for ErbB Kinases
- OGD
Oxygen Glucose Depravation
- OPC
Oligodendrocyte Progenitor Cells
- PI3-K
Phosphatidyl-Inositol-3-Ki-nase
- PIMs
Pro-inflammatory Markers
- pMCAO
Permanent Middle Cerebral Artery Occlusion
- RBC
Red Blood Cell
- RNA
Ribose Nucleic Acid
- SERCA2a
Sarco/Endoplasmic Reticulum Ca2+ ATPase 2a
- SG
Stress Granules
- STAT3
Signal Transducer and Activator of Transcription 3
- SZ
Schizophrenia
- TBI
Traumatic Brain Injury
- USD
United States Dollar
References
- 1.Burden S, Yarden Y (1997) Neuregulins and their recep- tors: a versatile signaling module in organogenesis and on- cogenesis. Neuron 18: 847–855. [DOI] [PubMed] [Google Scholar]
- 2.Kim JA, Jayabalan AK, Kothandan VK, Mariappan R, Kee Y, et al. (2016) Identification of Neuregulin-2 as a novel stress granule component. BMB Rep 49: 449–454. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 3.Nakano N, Kanekiyo K, Nakagawa T, Asahi M, Ide C (2016) NTAK/neuregulin-2 secreted by astrocytes promotes sur- vival and neurite outgrowth of neurons via ErbB3. Neurosci Lett 622: 88–94. [DOI] [PubMed] [Google Scholar]
- 4.Wang Y, Chen JY, Chen ML, Chen CH, Lai IC, et al. (2008) Neuregulin 3 genetic variations and susceptibility to schizo- phrenia in a Chinese population. Biol Psychiatry 64: 1093–1096. [DOI] [PubMed] [Google Scholar]
- 5.Meier S, Strohmaier J, Breuer R, Mattheisen M, Degenhardt F, et al. (2013) Neuregulin 3 is associated with at-tention deficits in schizophrenia and bipolar disorder. Int J Neuropsychopharmacol 16: 549–556. [DOI] [PubMed] [Google Scholar]
- 6.Wang KS, Xu N, Wang L, Aragon L, Ciubuc R, et al. (2014) NRG3 gene is associated with the risk and age at onset of Alzheimer disease. J Neural Transm (Vienna) 121: 183–192. [DOI] [PubMed] [Google Scholar]
- 7.Howard BA (2008) The role of NRG3 in mammary develop- ment. J Mammary Gland Biol Neoplasia 13: 195–203. [DOI] [PubMed] [Google Scholar]
- 8.Bernard JK, McCann SP, Bhardwaj V, Washington MK, Frey MR (2012) Neuregulin-4 is a survival factor for colon epithelial cells both in culture and in vivo. J Biol Chem 287: 39850–39858. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 9.Ma Y, Gao M, Liu D (2016) Preventing high fat diet-induced obesity and improving insulin sensitivity through neuregulin 4 gene transfer. Sci Rep 6: 26242. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 10.Steinthorsdottir V, Stefansson H, Ghosh S, Birgisdottir B, Bjornsdottir S, et al. (2004) Multiple novel transcription initiation sites for NRG1. Gene 342: 97–105. [DOI] [PubMed] [Google Scholar]
- 11.Mei L, Xiong WC (2008) Neuregulin 1 in neural development, synaptic plasticity and schizophrenia. Nat Rev Neurosci 9: 437–452. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 12.Jiang Q, Chen S, Hu C, Huang P, Shen H, et al. (2016) Neuregulin-1 (NRG1) signaling has a preventive role and is altered in the frontal cortex under the pathological conditions of Alzheimer’s disease. Mol Med Rep 14: 2614–2624. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 13.Falls DL (2003) Neuregulins: Functions, forms, and signaling strategies. Exp Cell Res 284: 14–30. [DOI] [PubMed] [Google Scholar]
- 14.Nave KA, Salzer JL (2006) Axonal regulation of myelination by neuregulin 1. Curr Opin Neurobiol 16: 492–500. [DOI] [PubMed] [Google Scholar]
- 15.Tan W, Wang Y, Gold B, Chen J, Dean M, et al. (2007) Molecular cloning of a brain-specific, developmentally regulated neuregulin 1 (NRG1) isoform and identification of a functional promoter variant associated with schizophrenia. J Biol Chem 282: 24343–24351. [DOI] [PubMed] [Google Scholar]
- 16.Wen D, Suggs SV, Karunagaran D, Liu N, Cupples RL, et al. (1994) Structural and functional aspects of the multiplicity of Neu differentiation factors. Mol Cell Biol 14: 1909–1919. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 17.Meyer D, Yamaai T, Garratt A, Riethmacher-Sonnenberg E, Kane D, et al. (1997) Isoform-specific expression and function of neuregulin. Development 124: 3575–3586. [DOI] [PubMed] [Google Scholar]
- 18.Liu X, Bates R, Yin DM, Shen C, Wang F, et al. (2011) Specific regulation of NRG1 isoform expression by neuronal activity. J Neurosci 31: 8491–8501. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 19.Gumà A, Martínez-Redondo V, López-Soldado I, Cantó C, Zorzano A (2010) Emerging role of neuregulin as a modulator of muscle metabolism. Am J Physiol Endocrinol Metab 298: 742–750. [DOI] [PubMed] [Google Scholar]
- 20.Britsch S (2007) The neuregulin-I/ErbB signaling system in development and disease. Adv Anat Embryol Cell Biol 190: 1–65. [PubMed] [Google Scholar]
- 21.Law AJ, Lipska BK, Weickert CS, Hyde TM, Straub RE, et al. (2006) Neuregulin 1 transcripts are differentially expressed in schizophrenia and regulated by 5’ SNPs associated with the disease. Proc Natl Acad Sci U S A 103: 6747–6752. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 22.Liu M, Solomon W, Cespedes JC, Wilson NO, Ford B, et al. (2018) Neuregulin-1 attenuates experimental cerebral malaria (ECM) pathogenesis by regulating ErbB4/AKT/STAT3 signaling. J Neuroinflammation 15: 104. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 23.Simmons Lauren J, Surles-Zeigler Monique C, Yonggang Li, Ford Gregory D, Newman Gale D, et al. (2016) Regulation of inflammatory responses by neuregulin-1 in brain ischemia and microglial cells in vitro involves the NF-kappa B pathway. J Neuroinflammation 13: 237. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 24.Ieguchi K, Fujita M, Ma Z, Davari P, Taniguchi Y, et al. (2010) Direct binding of the EGF-like domain of neuregulin-1 to integrins ({alpha}v{beta}3 and {alpha}6{beta}4) is involved in neuregulin-1/ErbB signaling. J Biol Chem 285: 31388–31398. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 25.Tokita Yoshihito, Keino Hiroomi, Matsui Fumiko, Aono Sachiko, Ishiguro Hiroshi, et al. (2001) Regulation of neuregulin expression in the injured rat brain and cultured astro-cytes. J Neurosci 21: 1257–1264. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 26.Dimayuga FO, Ding Q, Keller JN, Marchionni MA, Seroogy KB, et al. (2003) The neuregulin GGF2 attenuates free radical release from activated microglial cells. J Neuroimmunol 136: 67–74. [DOI] [PubMed] [Google Scholar]
- 27.Wu Limin, Walas Samantha J, Leung Wendy, Lo Eng H, Lok Josephine (2015) Neuregulin-1 and neurovascular protection, in brain neurotrauma. In: Kobeissy FH, Molec- ular, neuropsychological, and rehabilitation aspects CRC Press/Taylor & Francis, Boca Raton (FL). [Google Scholar]
- 28.Hidalgo A, Kinrade EF, Georgiou M (2001) The Drosophila neuregulin vein maintains glial survival during axon guid- ance in the CNS. Dev Cell 1: 679–690. [DOI] [PubMed] [Google Scholar]
- 29.Farkas JE, Freitas PD, Bryant DM, Whited JL, Monaghan JR (2016) Neuregulin-1 signaling is essential for nerve-depen- dent axolotl limb regeneration. Development 143: 2724–2731. [DOI] [PubMed] [Google Scholar]
- 30.Brown D, Samsa LA, Ito C, Ma H, Batres K, et al. (2018) Neuregulin-1 is essential for nerve plexus formation during cardiac maturation. J Cell Mol Med 22: 2007–2017. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 31.Yarden Y (2001) The EGFR family and its ligands in human cancer. signalling mechanisms and therapeutic opportunities. Eur J Cancer 4: S3–S8. [DOI] [PubMed] [Google Scholar]
- 32.Woo RS, Li XM, Tao Y, Carpenter-Hyland E, Huang YZ, et al. (2007) Neuregulin-1 enhances depolarization-induced GABA release. Neuron 54: 599–610. [DOI] [PubMed] [Google Scholar]
- 33.Gerlai R, Pisacane P, Erickson S (2000) Heregulin, but not ErbB2 or ErbB3, heterozygous mutant mice exhibit hyper-activity in multiple behavioral tasks. Behav Brain Res 109: 219–227. [DOI] [PubMed] [Google Scholar]
- 34.Kato T, Kasai Atsushi, Mizuno Makoto, Fengyi Liang, Shintani Nori-hito, et al. (2010) Phenotypic characterization of transgenic mice overexpressing neuregulin-1. PLoS One 5: e14185. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 35.Zhao WJ (2013) The expression and localization of neuregulin-1 (NRG1) in the gastrointestinal system of the rhesus monkey. Folia Histochem Cytobiol 51: 38–44. [DOI] [PubMed] [Google Scholar]
- 36.Zhao WY, Shen Y, Ren S (2011) Endogenous expression of Neuregulin-1 (NRG1) as a potential modulator of prolactin (PRL) secretion in GH3 cells. Cell Tissue Res 344: 313–320. [DOI] [PubMed] [Google Scholar]
- 37.Zhao WJ, Jiang Q, Mei JP, (2015) Neurohypophyseal Neuregulin 1 Is Derived from the Hypothalamus as a Potential Prolactin Modulator. Neuroendocrinology 102: 288–299. [DOI] [PubMed] [Google Scholar]
- 38.Zhao W, Ren SG (2011) Neuregulin-1 (NRG1) is mainly expressed in rat pituitary gonadotroph cells and possibly regulates prolactin (PRL) secretion in a juxtacrine manner. J Neuroendocrinol 23: 1252–1262. [DOI] [PubMed] [Google Scholar]
- 39.Person AD, Klewer SE, Runyan RB (2005) Cell biology of cardiac cushion development. Int Rev Cytol 243: 287–335. [DOI] [PubMed] [Google Scholar]
- 40.Milan DJ, Giokas AC, Serluca FC, Peterson RT, MacRae CA, et al. (2006) Notch1b and neuregulin are required for specification of central cardiac conduction tissue. Development 133: 1125–1132. [DOI] [PubMed] [Google Scholar]
- 41.Yaniz-Galende E, Chen J, Chemaly E, Liang L, Hulot JS, et al. (2012) Stem cell factor gene transfer promotes cardiac repair after myocardial infarction via in situ recruitment and expansion of c-kit+ cells. Circ Res 111: 1434–1445. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 42.Parodi EM, Kuhn B (2014) Signalling between microvascular endothelium and cardiomyocytes through neuregulin. Cardiovasc Res 102: 194–204. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 43.Gu X, Liu X, Xu D, Li X, Yan M, et al. (2010) Cardiac functional improvement in rats with myocardial infarction by up-regulating cardiac myosin light chain kinase with neuregulin. Cardiovasc Res 88: 334–343. [DOI] [PubMed] [Google Scholar]
- 44.Gao R, Zhang J, Cheng L, Wu X, Dong W, et al. (2010) A Phase II, randomized, double-blind, multicenter, based on standard therapy, placebo-controlled study of the efficacy and safety of recombinant human neuregulin-1 in patients with chronic heart failure. J Am Coll Cardiol 55: 1907–1914. [DOI] [PubMed] [Google Scholar]
- 45.Iglesias-GarcIa O, Baumgartner S, Macrí-Pellizzeri L, Rodriguez-Madoz JR, Abizanda G, et al. (2015) Neureg- ulin-1beta induces mature ventricular cardiac differentiation from induced pluripotent stem cells contributing to cardiac tissue repair. Stem Cells Dev 24: 484–496. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 46.Hedhli N, Dobrucki LW, Kalinowski A, Zhuang ZW, Wu X, et al. (2012) Endothelial-derived neuregulin is an important mediator of ischaemia-induced angiogenesis and arteriogenesis. Cardiovasc Res 93: 516–524. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 47.Xiao J, Li B, Zheng Z, Wang M, Peng J, et al. (2012) Therapeutic effects of neuregulin-1 gene transduction in rats with myocardial infarction. Coron Artery Dis 23: 460–468. [DOI] [PubMed] [Google Scholar]
- 48.Zhang Kaihua, Jones Lamont, Lim Sora, Maher Christopher A, Adkinset Douglas, et al. (2014) Loss of Trop2 causes ErbB3 activation through a neuregulin-1-dependent mechanism in the mesenchymal subtype of HNSCC. Oncotarget 5: 9281–9294. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 49.Montero JC, Rodríguez-Barrueco Ruth, Ocaña Alberto, Díaz-Rodríguez Ele-na, Esparís-Ogando Azucena, et al. (2008) Neuregulins and cancer. Clin Cancer Res 14: 3237–3241. [DOI] [PubMed] [Google Scholar]
- 50.Trombetta D, Rossi A, Fabrizio FP, Sparaneo A, Graziano P, et al. (2017) NRG1-ErbB Lost in Translation: A New Paradigm for Lung Cancer? Curr Med Chem 24: 4213–4228. [DOI] [PubMed] [Google Scholar]
- 51.Jeong H, Jeong H, Kim J, Lee Y, Seo JH, et al. (2014) Neuregulin-1 induces cancer stem cell characteristics in breast cancer cell lines. Oncol Rep 32: 1218–1224. [DOI] [PubMed] [Google Scholar]
- 52.Cabrera RM, Cabrera RM, Mao SPH, Surve CR, Condeelis JS, et al. (2018) A novel neuregulin -jagged1 paracrine loop in breast cancer transendothelial migration. Breast Cancer Res 20: 24. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 53.Endo H, Okami J, Okuyama H, Kumagai T, Uchida J, et al. (2013) Spheroid culture of primary lung cancer cells with neuregulin 1/HER3 pathway activation. J Thorac Oncol 8: 131–139. [DOI] [PubMed] [Google Scholar]
- 54.Yang L, Yang L, Li Y, Shen E, Cao F, et al. (2017) NRG1-dependent activation of HER3 induces primary resistance to trastuzumab in HER2-overexpressing breast cancer cells. Int J Oncol 51: 1553–1562. [DOI] [PubMed] [Google Scholar]
- 55.Han ME, Kim HJ, Shin DH, Hwang SH, Kang CD, et al. (2015) Overexpression of NRG1 promotes progression of gastric cancer by regulating the self-renewal of cancer stem cells. J Gastroenterol 50: 645–656. [DOI] [PubMed] [Google Scholar]
- 56.Pradeep S, Kim SW, Wu SY, Nishimura M, Chaluvally-Raghavan P, et al. (2014) Hematogenous metastasis of ovarian cancer: rethinking mode of spread. Cancer Cell 26: 77–91. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 57.Jay SM, Murthy AC, Hawkins JF, Wortzel JR, Steinhauser ML, et al. (2013) An engineered bivalent neuregulin pro- tects against doxorubicin-induced cardiotoxicity with reduced proneoplastic potential. Circulation 128: 152–161. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 58.Crespo V, James ML (2016) Neuromuscular Disease in the Neurointensive Care Unit. Anesthesiol Clin 34: 601–619. [DOI] [PubMed] [Google Scholar]
- 59.Birchmeier C, Bennett DL (2016) Neuregulin/ErbB Signaling in Developmental Myelin Formation and Nerve Repair. Curr Top Dev Biol 116: 45–64. [DOI] [PubMed] [Google Scholar]
- 60.Gunadi, Budi NYP, Sethi R, Fauzi AR, Kalim AS, et al. (2018) NRG1 variant effects in patients with Hirschsprung disease. BMC Pediatr 18: 292. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 61.Garcia-Barcelo MM, Tang CS, Ngan ES, Lui VC, Chen Y, et al. (2009) Genome-wide association study identifies NRG1 as a susceptibility locus for Hirschsprung’s disease. Proc Natl Acad Sci U S A 106: 2694–2699. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 62.Tang CS, Ngan ES, Tang WK, So MT, Cheng G, et al. (2012) Mutations in the NRG1 gene are associated with Hirschsprung disease. Hum Genet 131: 67–76. [DOI] [PubMed] [Google Scholar]
- 63.Liu L, Liu X, Bai Y, Tang N, Li J, et al. (2018) Neuregulin-1beta modulates myogenesis in septic mouse serum-treated C2C12 myotubes in vitro through PPARgam- ma/NF-kappaB signaling. Mol Biol Rep [DOI] [PubMed] [Google Scholar]
- 64.Bruno VM, Goldberg MP, Dugan LL, Giffard RG, Choi DW (1994) Neuroprotective effect of hypothermia in cortical cultures exposed to oxygen-glucose deprivation or excitatory amino acids. J Neurochem 63: 1398–1406. [DOI] [PubMed] [Google Scholar]
- 65.Croslan DR, Schoell MC, Ford GD, Pulliam JV, Gates A, et al. (2008) Neuroprotective effects of neuregulin-1 on B35 neuronal cells following ischemia. Brain Res 1210: 39–47. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 66.Guan YF, Wu CY, Fang YY, Zeng YN, Luo ZY, et al. (2015) Neuregulin 1 protects against ischemic brain injury via ErbB4 receptors by increasing GABAergic transmission. Neuroscience 307: 151–159. [DOI] [PubMed] [Google Scholar]
- 67.Li Y, Xu Z, Ford GD, Croslan DR, Cairobe T, et al. (2007) Neuroprotection by neuregulin-1 in a rat model of permanent focal cerebral ischemia. Brain Res 1184: 277–283. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 68.Guo WP, Wang J, Li RX, Peng YW (2006) Neuroprotective effects of neuregulin-1 in rat models of focal cerebral ischemia. Brain Res 1087: 180–185. [DOI] [PubMed] [Google Scholar]
- 69.Lok J, Wang H, Murata Y, Zhu HH, Qin T, et al. (2007) Effect of neuregulin-1 on histopathological and functional outcome after controlled cortical impact in mice. J Neurotrauma 24: 1817–1822. [DOI] [PubMed] [Google Scholar]
- 70.Zlokovic BV (2008) The blood-brain barrier in health and chronic neurodegenerative disorders. Neuron 57: 178–201. [DOI] [PubMed] [Google Scholar]
- 71.Lok J, Zhao S, Leung W, Seo JH, Navaratna D, et al. (2012) Neuregulin-1 effects on endothelial and blood-brain-barrier permeability after experimental injury. Transl Stroke Res 3: S119–S124. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 72.Fricker FR, Antunes-Martins A, Galino J, Paramsothy R, La Russa F, et al. (2013) Axonal neuregulin 1 is a rate limiting but not essential factor for nerve remyelination. Brain 136: 2279–2297. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 73.Jiang J, Zhang J, Yao P, Wu X, Li K (2014) Activation of spinal neuregulin 1-ErbB2 signaling pathway in a rat model of cancer-induced bone pain. Int J Oncol 45: 235–244. [DOI] [PubMed] [Google Scholar]
- 74.Clarke DJ, Sarkissian L, Todd SM, Suraev AS, Bahceci D, et al. (2018) NRG1 deficiency modulates the behavioural effects of prenatal stress in mice. Prog Neuropsychophar-macol Biol Psychiatry 88: 86–95. [DOI] [PubMed] [Google Scholar]
- 75.Mitchell RH, Goldstein BI (2014) Inflammation in children and adolescents with neuropsychiatric disorders: A systematic review. J Am Acad Child Adolesc Psychiatry 53: 274–296. [DOI] [PubMed] [Google Scholar]
- 76.Yun S, Reynolds RP, Masiulis I, Eisch AJ (2016) Re-evaluating the link between neuropsychiatric disorders and dys-regulated adult neurogenesis. Nat Med 22: 1239–1247. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 77.Murray CJ, Murray CJ, Vos T, Lozano R, Naghavi M, et al. (2012) Disability-adjusted life years (DALYs) for 291 diseases and injuries in 21 regions, 1990–2010: A systematic analysis for the global burden of disease study 2010. Lancet 380: 2197–2223. [DOI] [PubMed] [Google Scholar]
- 78.Taber KH, Hurley RA, Yudofsky SC (2010) Diagnosis and treatment of neuropsychiatric disorders. Annu Rev Med 61: 121–133. [DOI] [PubMed] [Google Scholar]
- 79.Mostaid MS, Lee TT, Chana G, Sundram S, Shannon Weickert C, et al. (2017) Elevated peripheral expression of neuregulin-1 (NRG1) mRNA isoforms in clozapine-treated schizophrenia patients. Transl Psychiatry 7: 1280. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 80.Mostaid MS, Lloyd D, Liberg B, Sundram S, Pereira A, et al. (2016) Neuregulin-1 and schizophrenia in the genome-wide association study era. Neurosci Biobehav Rev 68: 387–409. [DOI] [PubMed] [Google Scholar]
- 81.Stertz L, Magalhaes PV, Kapczinski F (2013) Is bipolar dis- order an inflammatory condition? The relevance of microg- lial activation. Curr Opin Psychiatry 26: 19–26. [DOI] [PubMed] [Google Scholar]
- 82.Idrizi R, Malcolm P, Weickert CS, Zavitsanou K, Sundram Suresh, et al. (2016) Striatal but not frontal cortical up-regulation of the epidermal growth factor receptor in rats exposed to immune activation in utero and cannabinoid treatment in adolescence. Psychiatry Res 240: 260–264. [DOI] [PubMed] [Google Scholar]
- 83.Dantzer R, O’Connor JC, Freund GG, Johnson RW, Kelley KW (2008) From inflammation to sickness and depression: when the immune system subjugates the brain. Nat Rev Neurosci 9: 46–56. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 84.Lewandowski KE, Cohen BM, Ongur D (2011) Evolution of neuropsychological dysfunction during the course of schizophrenia and bipolar disorder. Psychol Med 41: 225–241. [DOI] [PubMed] [Google Scholar]
- 85.Go RC, Perry RT, Wiener H, Bassett SS, Blacker D, et al. (2005) Neuregulin-1 polymorphism in late onset Alzheimer’s disease families with psychoses. Am J Med Genet B Neuropsychiatr Genet 139B: 28–32. [DOI] [PubMed] [Google Scholar]
- 86.Theoharides TC, Zhang B, Conti P (2011) Decreased mitochondrial function and increased brain inflammation in bipolar disorder and other neuropsychiatric diseases. J Clin Psychopharmacol 31: 685–687. [DOI] [PubMed] [Google Scholar]
- 87.Yang Zhai, Jiang Qiong, Chen Shuang-Xi, Hu Cheng-Liang, Shen Hui-Fan, et al. (2016) Differential changes in Neuregulin-1 signaling in major brain regions in a lipopoly-saccharide-induced neuroinflammation mouse model. Mol Med Rep 14: 790–796. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 88.Ryu J, Yu HN, Cho H, Kim HS, Baik TK, et al. (2012) Neuregulin-1 exerts protective effects against neurotoxicities induced by C-terminal fragments of APP via ErbB4 receptor. J Pharmacol Sci 119: 73–81. [DOI] [PubMed] [Google Scholar]
- 89.Chang KA, Shin KY, Nam E, Lee YB, Moon C, et al. (2016) Plasma soluble neuregulin-1 as a diagnostic biomarker for Alzheimer’s disease. Neurochem Int 97: 1–7. [DOI] [PubMed] [Google Scholar]
- 90.Middle F, Pritchard AL, Handoko H, Haque S, Holder R, et al. (2010) No association between neuregulin 1 and psy-chotic symptoms in Alzheimer’s disease patients. J Alzhei-mers Dis 20: 561–567. [DOI] [PubMed] [Google Scholar]
- 91.Fitzsimons DW (2013) World Health Organization. Acta Med Port 26: 186–187. [PubMed] [Google Scholar]
- 92.Kraemer SM, Smith JD (2006) A family affair: Var genes, PfEMP1 binding, and malaria disease. Curr Opin Microbiol 9: 374–380. [DOI] [PubMed] [Google Scholar]
- 93.Chan JA, Howell KB, Reiling L, Ataide R, Mackintosh CL, et al. (2012) Targets of antibodies against Plasmodium falciparum-infected erythrocytes in malaria immunity. J Clin Invest 122: 3227–3238. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 94.Pain A, Ferguson DJ, Kai O, Urban BC, Lowe B, et al. (2001) Platelet-mediated clumping of Plasmodium falciparum-infected erythrocytes is a common adhesive pheno-type and is associated with severe malaria. Proc Natl Acad Sci U S A 98: 1805–1810. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 95.Rowe JA, Claessens A, Corrigan RA, Arman M (2009) Adhesion of Plasmodium falciparum-infected erythrocytes to human cells: Molecular mechanisms and therapeutic implications. Expert Rev Mol Med 11: e16. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 96.Idro R, Jenkins NE, Newton CR (2005) Pathogenesis, clinical features, and neurological outcome of cerebral malaria. Lancet Neurol 4: 827–840. [DOI] [PubMed] [Google Scholar]
- 97.Birbeck GL, Molyneux ME, Kaplan PW, Seydel KB, Chimalizeni YF, et al. (2010) Blantyre Malaria Project Epilepsy Study (BMPES) of neurological outcomes in retinopathy-positive paediatric cerebral malaria survivors: A pro- spective cohort study. Lancet Neurol 9: 1173–1181. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 98.Gay F, Zougbédé S, N’dilimabaka N, Rebollo A, Mazier D, et al. (2012) Cerebral malaria: What is known and what is on research. Rev Neurol (Paris) 168: 239–256. [DOI] [PubMed] [Google Scholar]
- 99.Kochar DK, Shubhakaran, Kumawat BL, Kochar SK, Hal-wai M, et al. (2002) Cerebral malaria in Indian adults: A prospective study of 441 patients from Bikaner, north-west India. J Assoc Physicians India 50: 234–241. [PubMed] [Google Scholar]
- 100.Storm J, Craig AG (2014) Pathogenesis of cerebral ma-laria--inflammation and cytoadherence. Front Cell Infect Microbiol 4: 100. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 101.Ponsford MJ, Medana IM, Prapansilp P, Hien TT, Lee SJ, et al. (2012) Sequestration and microvascular congestion are associated with coma in human cerebral malaria. J Infect Dis 205: 663–671. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 102.White NJ, Turner GD, Day NP, Dondorp AM (2013) Lethal malaria: Marchiafava and Bignami were right. J Infect Dis 208: 192–198. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 103.Fairhurst RM, Bess CD, Krause MA (2012) Abnormal PfEMP1/knob display on Plasmodium falciparum-infected erythrocytes containing hemoglobin variants: Fresh insights into malaria pathogenesis and protection. Microbes Infect 14: 851–862. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 104.Clark IA, Alleva LM (2009) Is human malarial coma caused, or merely deepened, by sequestration? Trends Parasitol 25: 314–318. [DOI] [PubMed] [Google Scholar]
- 105.Clark IA, Rockett KA (1994) Sequestration, cytokines, and malaria pathology. Int J Parasitol 24: 165–166. [DOI] [PubMed] [Google Scholar]
- 106.Clark IA, Rockett KA (1994) The cytokine theory of human cerebral malaria. Parasitol Today 10: 410–412. [DOI] [PubMed] [Google Scholar]
- 107.Solomon W, Wilson NO, Anderson L, Pitts S, Patrickson J, et al. (2014) Neuregulin-1 attenuates mortality associated with experimental cerebral malaria. J Neuroinflammation 11: 9. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 108.Lok J, Sardi S Pablo, Guo Shuzhen, Besancon Elaine, Ha Duy M, et al. (2009) Neuregulin-1 signaling in brain endothelial cells. J Cereb Blood Flow Metab 29: 39–43. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 109.Sawyer DB, Caggiano A (2011) Neuregulin-1beta for the treatment of systolic heart failure. J Mol Cell Cardiol 51: 501–505. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 110.Linying Z, Wei W, Minxia W, Wenmin Z, Liangcheng Z (2014) Neuroprotective effects of neuregulin-1 ss on oligodendrocyte type 2 astrocyte progenitors following oxy- gen and glucose deprivation. Pediatr Neurol 50: 357–362. [DOI] [PubMed] [Google Scholar]
- 111.Galindo CL, Ryzhov S, Sawyer DB (2014) Neuregulin as a heart failure therapy and mediator of reverse remodeling. Curr Heart Fail Rep 11: 40–49. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 112.Jabbour A, Hayward CS, Keogh AM, Kotlyar E, McCrohon JA, et al. (2011) Parenteral administration of recombinant human neuregulin-1 to patients with stable chronic heart failure produces favourable acute and chronic haemodynamic responses. Eur J Heart Fail 13: 83–92. [DOI] [PubMed] [Google Scholar]
- 113.Xu Z, Croslan DR, Harris AE, Ford GD, Ford BD (2006) Extended therapeutic window and functional recovery after intraarterial administration of neuregulin-1 after focal ischemic stroke. J Cereb Blood Flow Metab 26: 527–535. [DOI] [PubMed] [Google Scholar]
- 114.Ji Y, Teng L, Zhang R, Sun J, Guo Y (2017) NRG-1beta exerts neuroprotective effects against ischemia reperfusion-induced injury in rats through the JNK signaling path- way. Neuroscience 362: 13–24. [DOI] [PubMed] [Google Scholar]
- 115.Iaci JF (2016) An optimized dosing regimen of cimaglermin (neuregulin 1beta3, glial growth factor 2) enhances molecular markers of neuroplasticity and functional recovery after permanent ischemic stroke in rats. J Neurosci Res 94: 253–265. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 116.Martini R (2014) Neuregulin-1 alleviates Charcot-Marie-Tooth disease in rats. Nat Med 20: 984–985. [DOI] [PubMed] [Google Scholar]
- 117.Dai DW, Xu Z, Chen X, Yuan L, Zhang AJ, et al. (2014) Distinct roles of neuregulin in different models of neuro- pathic pain. Neurol Sci 35: 531–536. [DOI] [PubMed] [Google Scholar]
- 118.Hendry JM, Alvarez-Veronesi MC, Placheta E, Zhang JJ, Gordon T, et al. (2016) ErbB2 blockade with Herceptin (trastuzumab) enhances peripheral nerve regeneration after repair of acute or chronic peripheral nerve injury. Ann Neurol 80: 112–126. [DOI] [PubMed] [Google Scholar]
- 119.Deng C, Pan B, Engel M, Huang XF (2013) Neuregulin-1 signalling and antipsychotic treatment: Potential therapeutic targets in a schizophrenia candidate signalling pathway. Psychopharmacology (Berl) 226: 201–215. [DOI] [PubMed] [Google Scholar]
- 120.Zhang P, Kuang H, He Y, Idiga SO, Li S, et al. (2018) NRG1-Fc improves metabolic health via dual hepatic and central action. JCI Insight 3. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 121.Ennequin G, Boisseau Nathalie, Caillaud Kevin, Chavanelle Vivien, Etienne Monique, et al. (2015) Neuregulin 1 Improves Glucose Tolerance in db/db Mice. PLoS One 10: e0130568. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 122.Hegde GV, de la Cruz CC, Chiu C, Alag N, Schaefer G, et al. (2013) Blocking NRG1 and other ligand-mediated Her4 signaling enhances the magnitude and duration of the chemotherapeutic response of non-small cell lung cancer. Sci Transl Med 5: 171. [DOI] [PubMed] [Google Scholar]
- 123.Meulendijks D, Jacob W, Voest EE, Mau-Sorensen M, Martinez-Garcia M, et al. (2017) Phase Ib Study of Lumretuzumab Plus Cetuximab or Erlotinib in Solid Tumor Patients and Evaluation of HER3 and Heregulin as Potential Biomarkers of Clinical Activity. Clin Cancer Res 23: 5406–5415. [DOI] [PubMed] [Google Scholar]
- 124.Jimeno A, Machiels JP, Wirth L, Specenier P, Seiwert TY, et al. (2016) Phase Ib study of duligotuzumab (MEH- D7945A) plus cisplatin/5-fluorouracil or carboplatin/paclitaxel for first-line treatment of recurrent/metastatic squa- mous cell carcinoma of the head and neck. Cancer 122: 3803–3811. [DOI] [PubMed] [Google Scholar]