

Abstract
Microbial culturomics represents an ongoing revolution in the characterization of the human gut microbiota. By using three culture media containing high salt concentrations (10, 15, and 20% [w/v] NaCl), we attempted an exhaustive exploration of the halophilic microbial diversity of the human gut and isolated strain Marseille‐P2481 (= CSUR P2481 = DSM 103076), a new moderately halophilic bacterium. This bacterium is a Gram‐positive, strictly aerobic, spore‐forming rod that is motile by use of a flagellum and exhibits catalase, but not oxidase activity. Strain Marseille‐P2481 was cultivated in media containing up to 20% (w/v) NaCl, with optimal growth being obtained at 37°C, pH 7.0–8.0, and 7.5% [w/v] NaCl). The major fatty acids were 12‐methyl‐tetradecanoic acid and hexadecanoic acid. Its draft genome is 4,548,390 bp long, composed of 11 scaffolds, with a G+C content of 39.8%. It contains 4,335 predicted genes (4,266 protein coding including 89 pseudogenes and 69 RNA genes). Strain Marseille‐P2481 showed 96.57% 16S rRNA sequence similarity with Gracilibacillus alcaliphilus strain SG103T, the phylogenetically closest species with standing in nomenclature. On the basis of its specific features, strain Marseille‐P2481T was classified as type strain of a new species within the genus Gracilibacillus for which the name Gracilibacillus timonensis sp. nov. is formally proposed.
Keywords: Gracilibacillus timonensis, halophilic, human gut flora, microbial culturomics, taxonogenomics
1. INTRODUCTION
One of the most important methods of food preservation in history has been the use of salt (NaCl). Salt has also become an indispensable ingredient of any kitchen. Considered previously as hostile to most forms of life by limiting the growth of certain bacteria, it was demonstrated to favor the emergence and growth of others, mainly halophilic bacteria (Cantrell, Dianese, Fell, Gunde‐Cimerman, & Zalar, 2011). Several recent studies have reported the isolation of new halophilic species from the human gut microflora (Khelaifia et al., 2016; Lagier, Khelaifia, et al., 2015). Therefore, exploring the diversity of halophilic microorganisms in the human gut flora may provide important insights into our understanding of their presence, interactions with the human digestive environment, and their influence on health.
In order to explore the human gut halophilic microbiota, and as part of the ongoing microbial culturomics study in our laboratory (Lagier et al., 2012, 2016), we used high salt‐containing culture media, which enabled us to isolate a new moderately halophilic bacterial strain, Marseille‐P2481, that belongs to the genus Gracilibacillus (Senghor et al., 2017). First proposed by Wainø et al. in 1999 (Wainø, Tindall, Schumann, & Ingvorsen, 1999), the genus Gracilibacillus currently includes 13 species ( http://www.bacterio.net/gracilibacillus.html) with validly published names (Parte, 2014). These are Gram stain‐positive, aerobic, moderately halophilic or halotolerant, motile bacteria. In most species, cells are motile due to peritrichous flagella and form endospores and white colonies (Wainø et al., 1999). Gracilibacillus species were isolated from diverse salty environmental samples, including sea water, salty lakes (Gao et al., 2012; Jeon et al., 2008), soil (Chen et al., 2008; Huo, Xu, Cui, & Wu, 2010), and/or food (Chamroensaksri et al., 2010; Diop et al., 2016).
Using the taxonogenomics approach that includes phenotypic features, proteomic information obtained by matrix‐assisted laser‐desorption/ionization time‐of‐flight mass spectrometry (MALDI‐TOF MS), and analysis of the complete genome sequence (Pagani et al., 2012; Ramasamy et al., 2014; Sentausa & Fournier, 2013), we present here the characterization of a new halophilic species for which we formally propose the name Gracilibacillus timonensis sp. nov. Strain Marseille‐P2481T (= CSUR P2481 = DSM 103076) is the type strain of Gracilibacillus timonensis sp. nov.
2. MATERIALS AND METHODS
2.1. Sample collection and culture conditions
A stool sample was collected from a 10‐year‐old healthy young Senegalese boy living in N'diop (a rural village in the Guinean‐Sudanian zone of Senegal). The patient's parents gave an informed consent, and the study was approved by the National Ethics Committee of Senegal (N° 00.87 MSP/DS/CNERS) and by the local ethics committee of the IFR48 (Marseille, France) under agreement 09‐022. The stool sample was collected immediately after defecation into a sterile plastic container, preserved at −80°C and transported to Marseille until further analysis.
The salinity of the sample was measured using a digital refractometer (Fisher scientific, Illkirch, France) and its pH measured using a pH‐meter (Eutech Instruments, Strasbourg, France).
Strain Marseille‐P2481 was isolated in aerobic conditions, on a home‐made culture medium consisting of Columbia agar enriched with 10% (w/v) NaCl (Sigma‐Aldrich, Saint‐Louis, MO, USA), as previously described (Diop et al., 2016). Briefly, 1 g of stool sample was inoculated into 100 ml of our home‐made liquid medium and incubated aerobically at 37°C. Subcultures were conducted after 1, 3, 7, 10, 15, 20, and 30 days of incubation. Serial dilutions of 10−1 to 10−10 were then performed in the home‐made liquid culture medium and plated on Columbia and Chapman agar plates (Oxoid, Dardilly, France). After 2 days of incubation at 37°C, all apparent colonies were picked and subcultured several times to obtain pure cultures.
2.2. MALDI‐TOF MS strain identification
Briefly, one isolated bacterial colony was picked from chapman culture plate using a pipette tip and spread it as a thin film on a MTP 96 MALDI‐TOF target plate for identification with a Microflex MALDI‐TOF MS spectrometer (Bruker Daltonics, Leipzig, Germany). In total, 12 distinct deposits for strain Marseille‐P2481were done from 12 individual colonies in duplicate. After air‐drying, 2‐μl matrix solution was applied per spot, as previously reported (Lagier, Khelaifia, et al., 2015). All spectra were recorded in positive linear mode for the mass range of 2,000–20,000 Da (parameter settings: ion source 1 (ISI), 20 kV; IS2, 18.5 kV; lens, 7 kV). The obtained protein spectra were compared with those of 2,480 spectra in the Bruker database enriched with our own database (Lagier, Hugon, et al., 2015). The strain was identified at the species level if the MALDI‐TOF MS score was greater than 1.9. If the score was lower than this threshold, the identification was not considered as reliable and the 16S rRNA gene was sequenced.
2.3. 16S rRNA gene sequencing identification
The 16S rRNA gene was amplified using the broad‐range primer pair FD1 and rp2 (Drancourt et al., 2000). The primers were obtained from Eurogentec (Seraing, France). The obtained amplicon was sequenced using the Big Dye Terminator Sequencing kit and the following internal primers: 536f, 536r, 800f, 800r, 1050f, 1050r, 357f, 357r, as previously described (Drancourt, Bollet, & Raoult, 1997; Drancourt et al., 2000). The sequence was then compared with the NCBI database using the BLASTn algorithm ( https://blast.ncbi.nlm.nih.gov/). If the 16S rRNA gene sequence similarity value was greater than 95% and lower 98.65% with the most closely related species with standing in nomenclature, as previously proposed (Kim, Oh, Park, & Chun, 2014; Stackebrandt & Ebers, 2006), the strain was proposed to belong to a new species (Konstantinidis, Ramette, & Tiedje, 2006).
2.4. Phylogenetic analysis
The 16S sequences from the type strains of the species with a validly published name that exhibited the highest BLAST score with our new strain were downloaded from the NCBI ftp server ( ftp://ftp.ncbi.nih.gov/Genome/). Sequences were aligned using the CLUSTALW 2.0 software (Larkin et al., 2007), and phylogenetic inferences were obtained using the neighbor‐joining method and the maximum likelihood method within the MEGA software, version 6 (Tamura, Stecher, Peterson, Filipski, & Kumar, 2013). The evolutionary distances were computed based on the Kimura 2‐parameter model (Kimura, 1980) with 95% of deletion, and bootstraping analysis was performed with 500 replications.
2.5. Morphological observation
To observe the cell morphology, transmission electron microscopy of the strain was performed using a Tecnai G20 Cryo (FEI company, Limeil‐Brevannes, France) at an operating voltage of 60 Kv after negative staining. Gram staining was performed and observed using a photonic microscope Leica DM2500 (Leica Microsystems, Nanterre, France) with a 100X oil‐immersion objective (Atlas & Snyder, 2011). The motility of the strain was assessed by the Hanging Drop method. The slide was examined using a DM1000 photonic microscope (Leica Microsystems) at 40×. Sporulation was tested following a thermic shock at 80°C during 20 min, and the endospore formation was visualized using a Tecnai G20 Cryo transmission electron microscope (FEI company, Limeil‐Brevannes, France) at an operating voltage of 60 Kv after negative staining.
2.6. Atmospheric tests, biochemical, and antimicrobial susceptibility
In order to evaluate the optimal culture conditions, strain Marseille‐P2481 was cultivated on Chapman agar at different temperatures (25, 28, 37, 45 and 56°C) under aerobic conditions, and in anaerobic and microaerophilic atmospheres using GENbag Anaer and GENbag microaer systems (bioMérieux), respectively. The pH (pH 5, 6, 6.5, 7, and 8.5) and salinity (5–20% [w/v] NaCl) conditions were also tested.
Biochemical tests were performed using the API ZYM, API 50 CH, and API 20 NE strips (bioMerieux, Marcy‐l'Etoile, France), according to the manufacturer's instructions. The API ZYM was incubated for 4 hr and the other two strips for 48 hr.
The antibiotic susceptibility of strain Marseille‐P2481 was determined using the disk diffusion method as previously described (Diop et al., 2016). The following antibiotics were tested: penicillin G (10 μg), amoxicillin (25 μg), ceftriaxone (30 μg), imipenem (10 μg), rifampicin (30 μg), erythromycin (15 μg), gentamicin (500 μg), and metronidazole (4 μg). The results were interpreted using the Scan 1,200 automate (Interscience, Saint Nom la Bretêche, France).
2.7. Fatty acid methyl ester (FAME) analysis by GC/MS
For the FAME analysis, strain Marseille‐P2481 was cultivated on Chapman agar (7.5% NaCl) (Oxoid, Dardilly, France) at 37°C under aerobic atmosphere for 2 days. Cellular fatty acid methyl ester (FAME) analysis was performed by gas chromatography/mass spectrometry (GC/MS). Two samples were prepared with approximately 70 mg of bacterial biomass per tube harvested from several culture plates. FAMEs were prepared as described by Sasser (Sasser, 1990). GC/MS analyses were carried out as previously described (Dione et al., 2016). Briefly, FAMEs were separated using an Elite 5‐MS column and monitored by mass spectrometry (Clarus 500 ‐ SQ 8 S, Perkin Elmer, Courtaboeuf, France). Spectral database search was performed using the MS Search 2.0 software operated with the Standard Reference Database 1A (NIST, Gaithersburg, USA) and the FAMEs mass spectral database (Wiley, Chichester, UK).
2.8. Extraction and genome sequencing
After a pretreatment by lysozyme incubation at 37°C for 2 hr, the DNA of strain Marseille‐P2481 was extracted on the EZ1 biorobot (Qiagen) with EZ1 DNA Tissue kit. The elution volume was 50 μl. The gDNA was quantified by a Qubit assay with the high sensitivity kit (Life Technologies, Carlsbad, CA, USA) to 185 ng/μl.
A MiSeq sequencer and the mate‐pair strategy (Illumina Inc, San Diego, CA, USA) were used to sequence the gDNA. The gDNA was barcoded in order to be mixed with 11 other projects with the Nextera Mate‐Pair sample prep kit (Illumina). The mate‐pair library was prepared with 1.5 μg of gDNA using the Nextera mate‐pair guide. The genomic DNA sample was simultaneously fragmented and tagged with a mate‐pair junction adapter. The pattern of the fragmentation was validated on an Agilent 2100 BioAnalyzer (Agilent Technologies Inc, Santa Clara, CA, USA) with a DNA 7500 labchip. The DNA fragments ranged in size from 1.5 to 11 kb with an optimal size at 5.314 kb. No size selection was performed and 600 ng of tagmented fragments was circularized. The circularized DNA was mechanically sheared with an optimal size at 939 bp on the Covaris device S2 in T6 tubes (Covaris, Woburn, MA, USA). The library profile was visualized on a High Sensitivity Bioanalyzer LabChip (Agilent Technologies Inc, Santa Clara, CA, USA), and the final concentration library was measured at 8.38 nmol/L. The libraries were normalized at 2 nmol/L and pooled. After a denaturation step and dilution at 15 pM, the pool of libraries was loaded. Automated cluster generation and sequencing run were performed in a single 39‐hr run in a 2 × 251 bp.
A total sequencing output of 6.52 Gb was obtained from a 696 K/mm2 cluster density with a cluster passing quality control filters of 95.6% (12,863,388 passing filter paired reads). Within this run, the index representation for strain Marseille‐P2481 was determined to be 9.39%. The 1,207,306 paired reads were trimmed and then assembled.
2.9. Genome annotation and comparison
Prodigal was used for open reading frame (ORF) prediction (Hyatt et al., 2010) with default parameters. Predicted ORFs spanning a sequencing gap region were excluded. Bacterial protein sequences were predicted using BLASTP (E‐value 1e−03, coverage 0.7 and identity percent 30%) against the Clusters of Orthologous Groups (COG) database. If no hit was found, a search against the nr database (Benson et al., 2015) was performed using BLASTP with E‐value of 1e−03, a coverage of 0.7 and an identity percent of 30%. If sequence lengths were smaller than 80 amino acids, we used an E‐value of 1e−05. Pfam conserved domains (PFAM‐A an PFAM‐B domains) were searched on each protein with the HHMscan tool (Finn et al., 2015). RNAmmer (Lagesen et al., 2007) and tRNAScanSE (Lowe & Eddy, 1997) were used to identify ribosomal RNAs and tRNAs, respectively. We predicted lipoprotein signal peptides and the number of transmembrane helices using Phobius (Käll, Krogh, & Sonnhammer, 2004). ORFans were identified if the BLASTP search was negative (E‐value smaller than 1e−03 for ORFs with a sequence size larger than 80 aas or E‐value smaller than 1e−05 for ORFs with sequence length smaller than 80 aas). Artemis (Carver, Harris, Berriman, Parkhill, & McQuillan, 2012) and DNA Plotter (Carver, Thomson, Bleasby, Berriman, & Parkhill, 2009) were used for data management and for visualization of genomic features, respectively. Genomes from members of the genus Gracilibacillus and closely related genera were used for the calculation of AGIOS values. The genome of strain Marseille‐P2481 (EMBL‐EBI accession number FLKH00000000) was compared with those of Gracilibacillus halophilus strain YIM‐C55.5T (APML00000000), G. boraciitolerans strain JCM 21714T (BAVS00000000), G. lacisalsi strain DSM 19029 T (ARIY00000000), G. massiliensis strain Awa‐1T (CZRP00000000), G. kekensis strain K170 T (FRCZ01000001), G. orientalis strain XH‐63 T (FOTR01000001), G. ureilyticus strain MF38 T (FOGL01000001), B. clausii strain KSM‐K16T (AP006627), and B. alcalophilus strain ATCC 27647T (ALPT00000000). Annotation and comparison processes were performed using the multi‐agent software system DAGOBAH (Gouret et al., 2011), which includes Figenix (Gouret et al., 2005) libraries that provide pipeline analysis. We also estimated the degrees of genomic sequence similarity among compared genomes using the following tools: first, we used the MAGI home‐made software (Padmanabhan, Mishra, Raoult, & Fournier, 2013) This software calculates the average genomic identity of orthologous gene sequences (AGIOS) among compared genomes (Ramasamy et al., 2014). It combines the Proteinortho software (Lechner et al., 2011) for detecting orthologous proteins in pairwise genomic comparisons, then retrieves the corresponding genes and determines the mean percentage of nucleotide sequence identity among orthologous ORFs using the Needleman‐Wunsch global alignment algorithm. Second, the digital DNA–DNA hybridization was performed using the GGDC (Genome‐to‐Genome Distance Calculator) analysis via the GGDC web server as previously reported (Klenk, Meier‐Kolthoff, & Göker, 2014). Finally, the average amino acid identity (AAI) was calculated, based on the overall similarity between two genomic datasets of proteins (Rodriguez‐R & Konstantinidis, 2014) available at ( http://enve-omics.ce.gatech.edu/aai/index).
3. RESULTS
3.1. Strain identification and phylogenetic analysis
A MALDI‐TOF MS score of 1.4 was obtained for strain Marseille‐P2481 against our database (Bruker database), suggesting that our isolate was not in the database. The MALDI‐TOF MS spectrum from strain Marseille‐P2481 (Figure 1) was added to our database and a gel view showed the spectral differences between our isolate and other closely related species (Figure 2). The 16S rDNA‐based identification of strain Marseille‐P2481 (EMBL‐EBI accession number LT223702) yielded a 96.57% 16S rRNA gene sequence identity with Gracilibacillus alcaliphilus strain SG103T (GenBank accession number NR_126185), the phylogenetically closest species with a validly published name (Figure 3). As this value was lower than the 98.65% 16S rRNA sequence identity threshold recommended to define a new species without carrying out DNA–DNA hybridization (Kim et al., 2014), strain Marseille‐P2481 was considered as representative of a potential new species within the Gracilibacillus genus.
Figure 1.
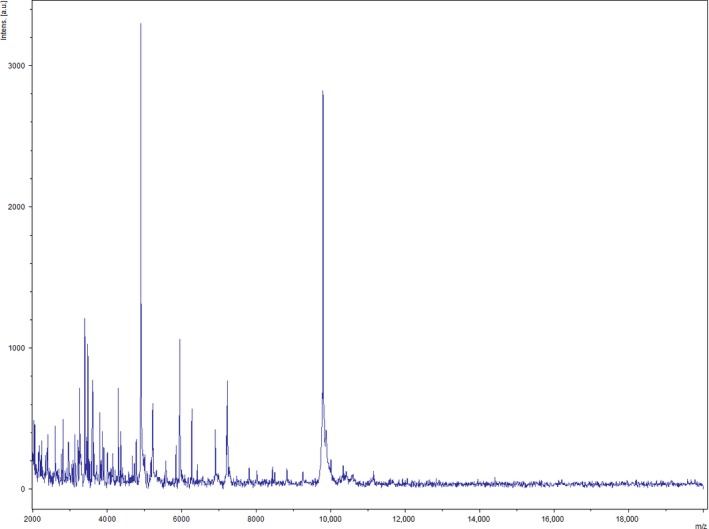
Reference mass spectrum from Gracilibacillus timonensis strain Marseille‐P2481T
Figure 2.
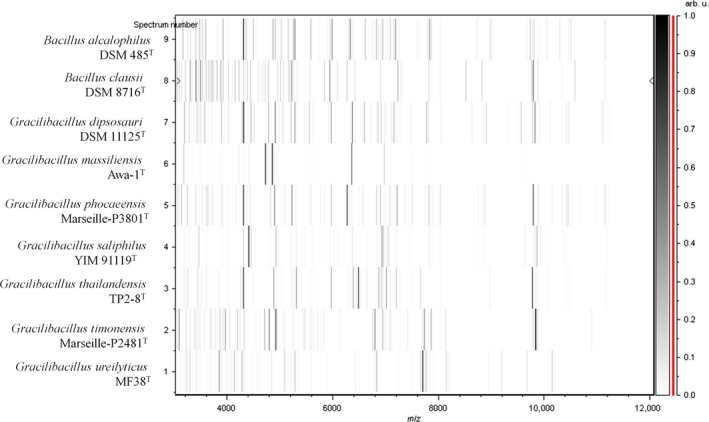
Gel view comparing Gracilibacillus timonensis strain Marseille‐P2481T with other species within the genera Gracilibacillus and Bacillus
Figure 3.
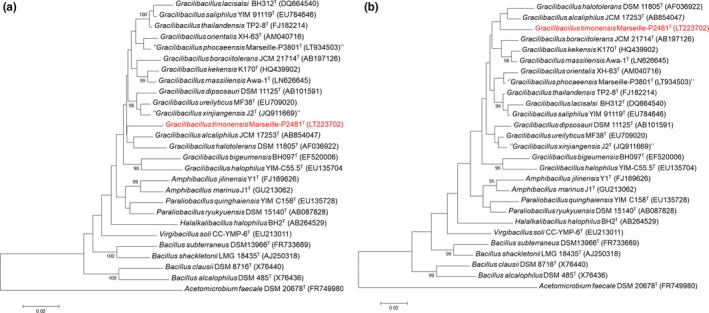
Phylogenetic tree highlighting the position of Gracilibacillus timonensis strain Marseille‐P2481T relative to other closely related species. GenBank accession numbers of each 16S rRNA are indicated after each species name. Sequences were aligned using CLUSTALW, and the evolutionary history was inferred using the Neighbor‐Joining method (a) and the maximum likelihood method (b) with the Kimura 2‐parameter method within MEGA6 software. The percentage of replicate trees in which the associated taxa clustered together in the bootstrap test (500 replicates) is shown next to the branches. The analysis involved 24 nucleotide sequences. All positions with less than 95% site coverage were eliminated. There were a total of 1,404 positions in the final dataset. The scale bar represents a 2% nucleotide sequence divergence
3.2. Physiological and biochemical characteristics
Isolated for the first time in our home‐made halophilic medium with 10% (w/v) NaCl, strain Marseille‐P2481 was able to grow in media containing up to 20% (w/v) NaCl under aerobic conditions with a minimal concentration of growth at 7.5% NaCl, but was also able to grow in anaerobic and microaerophilic atmospheres (at 37°C). After 2 days of growth at 37°C, colonies were creamy orange and circular, with a mean diameter of 0.2 μm. Cells were Gram stain‐positive (Figure 4a), endospore‐forming (Figure 4b), and motile rods with a peritrichous flagellum. Cells were also slightly curved, with mean diameter and length of 0.5 and 1.9 μm, respectively (Figure 4b). Strain Marseille‐P2481 exhibited positive catalase but no oxidase activity. General features and classification of Gracilibacillus timonensis strain Marseille‐P2481T are summarized in Table 1. Using an API ZYM strip, positive results were obtained for esterase, esterase lipase, acid phosphatase, naphtol‐AS‐BI‐phosphohydrolase β‐galactosidase, β‐glucosidase, and α‐glucosidase activities but no reaction was observed for alkaline phosphatase, lipase, Leucine arylamidase, Valine arylamidase, Cystine arylamidase, α‐galactosidase, β‐glucuronidase, trypsin, α‐chymotrypsin, α‐mannosidase, α‐fucosidase, and N‐acetyl‐β‐glucosaminidase. The API 50CH strip revealed that strain Marseille‐P2481 exhibited esculin hydrolysis, but negative reactions were obtained for d‐arabitol, l‐arabitol, d‐glucose, d‐fructose, d‐fucose, d‐galactose, d‐lactose, d‐maltose, d‐ribose, d‐saccharose, d‐lyxose, d‐mannose l‐sorbose, d‐tagatose, d‐turanose, d‐xylose, l‐xylose, d‐arabinose, l‐arabinose, d‐sorbitol, d‐cellobiose, d‐melezitose, d‐melibiose, d‐trehalose, d‐raffinose, l‐rhamnose, d‐adonitol, d‐mannitol, l‐fucose, amygdalin, arbutin, erythritol, dulcitol, gentiobiose, glycerol, glycogen, inositol, inulin, salicin, starch, xylitol, αD‐glucopyranoside, methyl‐βD‐xylopyranoside, methyl‐αD‐mannopyranoside, potassium gluconate, potassium‐2‐ketogluconate potassium‐5‐ketogluconate, N‐acetylglucosamine. Using an API 20NE strip, fermentation of glucose, urease activity, and metabolism of l‐arginine, esculin and 4‐nitrophenyl‐βD‐galactopyrasinoside were positive. In contrast, nitrate and indole production, gelatinase activity and metabolism of d‐glucose, l‐arabinose, d‐mannose, d‐maltose, d‐mannitol, N‐acetyl‐glucosamine, potassium gluconate, capric acid, malic acid, trisodium citrate, and phenylacetic acid were negative. Strain Marseille‐P2481 differed from all other studied members of the genus Gracilibacillus in a combination of negative alkaline phosphatase and nitrate reductase activities but the acidification of d‐fructose (Table 2). The cellular fatty acids from strain Marseille‐P2481 are mainly saturated and the most abundant were 12‐methyl‐tetradecanoic acid, hexadecanoic acid, and 14 methyl‐hexadecanoic acid (45%, 16%, and 14%, respectively). No unsaturated fatty acid was detected (Table 3). Cells are resistant to Penicillin G, amoxicillin, ceftriaxone, and metronidazole, but susceptible to imipenem, rifampicin, gentamicin, and erythromycin.
Figure 4.
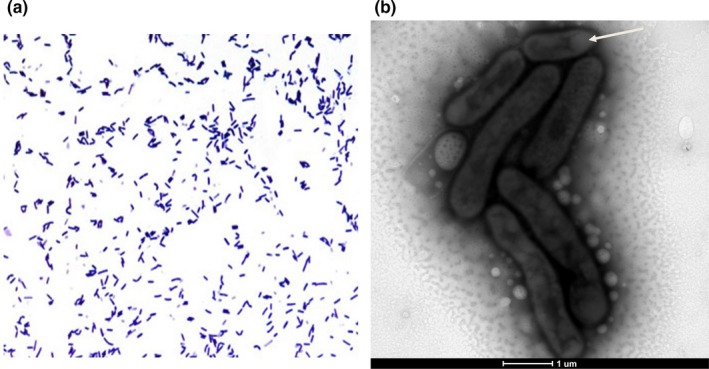
Bacterial morphology. (a) Gram staining of Gracilibacillus timonensis strain Marseille‐P2481T. (b) Transmission electron microscopy showing an endospore in terminal position (arrow). The scale bar represents 1 μm
Table 1.
Classification and general features of Gracilibacillus timonensis strain Marseille‐P2481T according to the MIGS recommendations [23]
| MIGS ID | Property | Term | Evidence codea |
|---|---|---|---|
| Domain: Bacteria | TAS (Woese, Kandler, & Wheelis, 1990) | ||
| Phylum: Firmicutes | TAS (Skerman & Sneath 1980, Murray, 1984, Gibbons and Murray, 1978, Garrity and Holt, 2001) | ||
| Class: Bacilli | TAS (Ludwig, Schleifer, & Whitman 2009) | ||
| Order: Bacillales | TAS (Skerman & Sneath 1980, Prevot, 1953) | ||
| Family: Bacillaceae | TAS (Skerman & Sneath 1980, Fischer, 1895) | ||
| Genus: Gracilibacillus | TAS (Wainø et al., 1999) | ||
| Species: Gracilibacillus timonensis | IDA | ||
| Type strain: Marseille‐P2481T | IDA | ||
| Gram stain | Positive | IDA | |
| Cell shape | Rods | IDA | |
| Motility | Motile | IDA | |
| Sporulation | Spore‐forming | IDA | |
| Temperature (°C) | Mesophile (25‐45) | IDA | |
| Optimum temperature | 37°C | IDA | |
|
pH range: Optimal pH |
6.0–9.0 7.0–8.0 |
IDA | |
| Carbon source | Unknown | IDA | |
| MIGS‐6 | Habitat | Human gut | IDA |
| MIGS‐6.3 | NaCl range: Optimum NaCl |
7.5–20% 7.5% |
IDA |
| MIGS‐22 | Oxygen requirement | Aerobic | IDA |
| MIGS‐15 | Biotic relationship | Free living | IDA |
| MIGS‐14 | Pathogenicity | Unknown | IDA |
Evidence codes: IDA, Inferred from Direct Assay; TAS, Traceable Author Statement (i.e., a direct report exists in the literature). These evidence codes are from the Gene Ontology project (Ashburner et al. 2000).
Table 2.
Differential characteristics of Gracilibacillus timonensis strain Marseille‐P2481T and other closely related members of the genus Gracilibacillus
| Properties | G. timonensis | G. saliphilus | G. bigeumensis | G. halophilus | G. boraciitolerans | G. kekensis | G. halotolerans | G. alcaliphilus |
|---|---|---|---|---|---|---|---|---|
| Cell diameter (μm) | 0.5–0.8 | 0.7–0. 9 | 0.3–0.5 | 0.3–0.5 | 0.5–0. 9 | 0.2–1.05 | 0.4–0.6 | 0.5–0.7 |
| Pigmentation | Creamy orange | Creamy white | Creamy | White | Dirty white | Creamy white | Creamy white | Creamy white |
| Oxygen requirement | Aerobic | Aerobic | Aerobic | Aerobic | Aerobic | Aerobic | Aerobic | Aerobic |
| Gram stain | + | + | + | + | + | + | + | + |
| Salt requirement | + | + | + | + | + | + | + | + |
| Motility | + | + | + | + | + | + | + | + |
| Sporulation | + | + | + | + | + | + | + | + |
| Indole | − | − | − | − | − | − | − | − |
| Production of | ||||||||
| Alkaline phosphate | − | + | + | + | + | NA | + | − |
| Catalase | + | + | + | + | + | NA | + | + |
| Oxidase | − | + | + | + | + | − | + | − |
| Nitrate reductase | − | + | − | + | − | − | + | + |
| Urease | + | + | − | − | − | − | + | + |
| β‐galactosidase | + | + | + | + | + | NA | − | NA |
| α‐galactosidase | − | − | − | − | + | NA | + | − |
| N‐acetyl‐glucosamine | − | + | − | − | NA | NA | NA | + |
| l‐arabinose | − | + | + | − | + | + | + | + |
| Ribose | − | + | − | + | + | + | + | + |
| d‐mannose | − | + | + | − | + | + | − | − |
| d‐mannitol | − | + | + | + | + | + | + | + |
| d‐glucose | + | + | + | + | + | + | + | + |
| d‐fructose | − | + | + | + | + | + | + | + |
| d‐maltose | − | + | + | − | + | + | − | + |
| d‐lactose | − | + | + | − | + | + | − | + |
| DNA G+C content (mol %) | 39.8 | 40.1 | 37.9 | 42.3 | 35.8 | 35.8 | 38 | 41.3 |
| Habitat | Human gut | Salt lake | Solar saltern soil | Salty soil | Soil | Salty lake | Saline soil | Fermentation liquor for dyeing |
NA, no data available.
G. timonensis strain Marseille‐P2481T , G. Gracilibacillus bigeumensis strain BH097T (Kim et al., 2012), G. halophilus strain YIM‐C55.5T (Chen et al., 2008), G. boraciitolerans strain T‐16XT (Ahmed et al., 2007), G. saliphilus strain YIM91119T (Tang et al., 2009), G. kekensis strain K170T (Gao et al., 2012), G. halotolerans strain NNT (Wainø et al., 1999), G. alcaliphilus strain SG103T (Hirota, Hanaoka, Nodasaka, & Yumoto, 2014).
Table 3.
Total cellular fatty acid composition of Gracilibacillus timonensis strain Marseille‐P2481T
| Fatty acids | IUPAC name | Mean relative %a |
|---|---|---|
| 15:0 anteiso | 12‐methyl‐tetradecanoic acid | 45.4 ± 1.5 |
| 16:0 | Hexadecanoic acid | 15.6 ± 1.1 |
| 17:0 anteiso | 14‐methyl‐Hexadecanoic acid | 13.9 ± 0.6 |
| 15:0 iso | 13‐methyl‐tetradecanoic acid | 10.3 ± 0.6 |
| 17:0 iso | 15‐methyl‐Hexadecanoic acid | 5.8 ± 1.0 |
| 16:0 iso | 13‐methyl‐Pentadecanoic acid | 3.4 ± 0.4 |
| 18:0 | Octadecanoic acid | 1.2 ± 0.1 |
| 15:0 | Pentadecanoic acid | 1.1 ± 0.2 |
| 14:0 iso | 12‐methyl‐Tridecanoic acid | 1.1 ± 0.1 |
| 17:0 | Heptadecanoic acid | 1.1 ± 0.1 |
| 14:0 | Tetradecanoic acid | TR |
| 10:0 | Decanoic acid | TR |
| 12:0 | Dodecanoic acid | TR |
| 13:0 anteiso | 10‐methyl‐Dodecanoic acid | TR |
| 13:0 iso | 11‐methyl‐Dodecanoic acid | TR |
Mean peak area percentage calculated from the analysis of FAMEs in 2 sample preparations ± standard deviation (n = 3); TR= trace amounts < 1%.
3.3. Genome properties
The genome is 4,548,390 bp long with a 39.8% G+C content. It is composed of 11 scaffolds (composed of 12 contigs). Of the 4,335 predicted genes, 4,266 were protein‐coding genes and 69 were RNAs (4 complete 16S rRNA, 6 complete 5S rRNA gene, 2 complete and 2 partiel 23S rRNA, and 51 tRNA genes, as well as additional 4 other rRNAs). A total of 3,043 genes (70.24%) were assigned a putative function (by COGs or BLAST against nr). A total of 214 genes were identified as ORFans (6.94%). The remaining genes were annotated as hypothetical proteins (861 genes => 19.92%). The genome statistics are presented in Table 4, and the distribution of genes into COGs functional categories is summarized in Table 5.
Table 4.
Nucleotide content and gene count of the genome
| Attribute | Value | % of totala |
|---|---|---|
| Size (bp) | 4,548,390 | 100% |
| G+C content (bp) | 1,808,751 | 39.8% |
| Coding region (bp) | 3,844,022 | 85.07% |
| Total genes | 4,395 | 100% |
| RNA genes | 63 | 1.76% |
| Protein‐coding genes | 4,332 | 98.23% |
| Genes with function prediction | 3,043 | 68.95% |
| Genes assigned to COGs | 2,797 | 63.94% |
| Genes with peptide signals | 474 | 11.20% |
| Genes with transmembrane helices | 1,191 | 27.68% |
The total is based on either the size of the genome in base pairs or the total number of protein‐coding genes in the annotated genome.
Table 5.
Number of genes associated with the 25 general COG functional categories
| Code | Value | % value | Description |
|---|---|---|---|
| J | 212 | 4.89 | Translation |
| A | 0 | 0 | RNA processing and modification |
| K | 266 | 6.14 | Transcription |
| L | 103 | 2.37 | Replication, recombination, and repair |
| B | 1 | 0.02 | Chromatin structure and dynamics |
| D | 52 | 1.20 | Cell cycle control, mitosis, and meiosis |
| Y | 0 | 0 | Nuclear structure |
| V | 98 | 2.26 | Defense mechanisms |
| T | 154 | 3.46 | Signal transduction mechanisms |
| M | 147 | 3.39 | Cell wall/membrane biogenesis |
| N | 49 | 1.13 | Cell motility |
| Z | 0 | 0 | Cytoskeleton |
| W | 3 | 0.06 | Extracellular structures |
| U | 30 | 0.69 | Intracellular trafficking and secretion |
| O | 107 | 2.46 | Posttranslational modification, protein turnover, chaperones |
| X | 57 | 1.31 | Mobilome: prophages, transposons |
| C | 113 | 2.60 | Energy production and conversion |
| G | 478 | 11.03 | Carbohydrate transport and metabolism |
| E | 201 | 4.63 | Amino acid transport and metabolism |
| F | 100 | 2.30 | Nucleotide transport and metabolism |
| H | 138 | 3.18 | Coenzyme transport and metabolism |
| I | 94 | 2.16 | Lipid transport and metabolism |
| P | 192 | 4.43 | Inorganic ion transport and metabolism |
| Q | 66 | 1.52 | Secondary metabolites biosynthesis, transport, and catabolism |
| R | 288 | 6.64 | General function prediction only |
| S | 212 | 4.89 | Function unknown |
| ‐ | 1,535 | 35.43 | Not in COGs |
3.4. Comparative genomics
The draft genome sequence structure of strain Marseille‐P2481 is summarized in Figure 5. It is smaller than those of G. orientalis (4.54 and 4.61 Mb, respectively), but larger than those of G. halophilus, G. boraciitolerans, G. kekensis, G. ureilyticus, G. massiliensis, B. alcalophilus, G. lacisalsi, and B. clausii (3.03, 3.65, 3.93, 4.07, 4.21, 4.37, 4.41 and 4.52 Mb, respectively). The G+C content of strain Marseille‐P2481 is smaller than those of B. clausii (39.8 and 44.75%, respectively), but larger than those of G. boraciitolerans, G. kekensis, G. massiliensis, G. orientalis, G. lacisalsi, B. alcalophilus, G. ureilyticus, and G. halophilus (35.8, 36.0, 36.1, 36.3, 36.8, 37.4, 37.5, and 37.9%, respectively). The gene content of strain Marseille‐P2481 is smaller than those of G. orientalis, B. clausii, and G. boraciitolerans (4,335, 4,350, 4,441, and 4,510 genes, respectively), but larger than those of G. halophilus, G. kekensis, G. massiliensis, B. alcalophilus, G. ureilyticus, and G. lacisalsi, (2,999, 3,842, 3,887, 3,973, 4,066, and 4,290 genes, respectively). The gene distribution into COG categories was similar among all compared genomes (Figure 6). In addition, the AGIOS analysis showed that strain Marseille‐P2481 shared 2,103, 2,112, 2,004, 2,027, 1,461, 1,982, 1,695, 1,539, and 1,578 orthologous proteins with G. lacisalsi, G. orientalis, G. massiliensis, G. kekensis, G. boraciitolerans, G. ureilyticus, G. halophilus, B. alcalophilus, and B. clausii, respectively (Table 6). When comparing strain Marseille‐P2481 to other species, AGIOS values were 69.8, 70.0, 71.0, 71.8, 72.0, 72.1, and 72.3% with G. halophilus, G. ureilyticus, G. boraciitolerans, G. kekensis, G. massiliensis, G. orientalis, and G. lacisalsi, respectively (Table 6), but ranged from 62.9% to 64.5% with B. clausii and B. alcalophilus, respectively (Table 6). In addition, dDDH values relatedness of strain Marseille‐P2481 and the compared closest species varied between 19.1 and 28.67% and were 20.5, 19.8, 21.6, 20.1, 19.1, 21.4, 19.3, 23.6, and 28.67% for G. lacisalsi, G. orientalis, G. massiliensis, G. kekensis, G. boraciitolerans, G. ureilyticus, G. halophilus, B. alcalophilus, and B. clausii, respectively (Table 7). Finally, AAI values relatedness between strain Marseille‐P2481, G. lacisalsi, G. orientalis, G. massiliensis, G. kekensis, G. boraciitolerans, G. ureilyticus, and G. halophilus were 68.72, 68.19, 68.18, 67.90, 68.08, 64.69, and 64.37%, respectively, but were lower when compared with B. alcalophilus and B. clausii, with 51.72 and 50.73%, respectively (Table 8). These dDDH and AAI values were less than the 70% and 95–96% threshold values for species demarcation, respectively (Chun et al., 2018; Klappenbach et al., 2007; Meier‐Kolthoff, Auch, Klenk, & Göker, 2013; Richter & Rosselló‐Móra, 2009; Rodriguez‐R & Konstantinidis, 2014).
Figure 5.
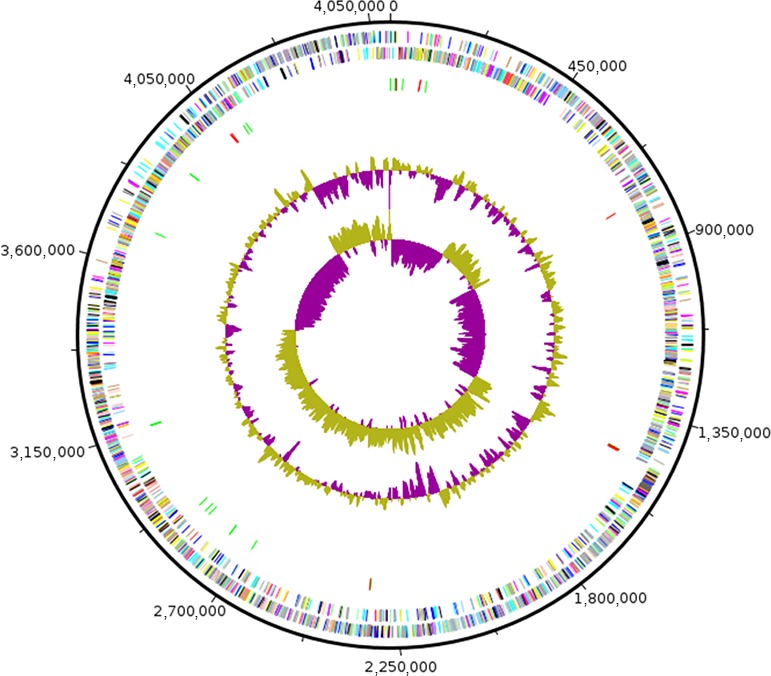
Graphical circular map of the chromosome. From the outside to the center: Genes on the forward strand colored by Clusters of Orthologous Groups of proteins (COG) categories (only genes assigned to COG), genes on the reverse strand colored by COG categories (only gene assigned to COG), RNA genes (tRNAs green, rRNAs red), GC content, and GC skew
Figure 6.
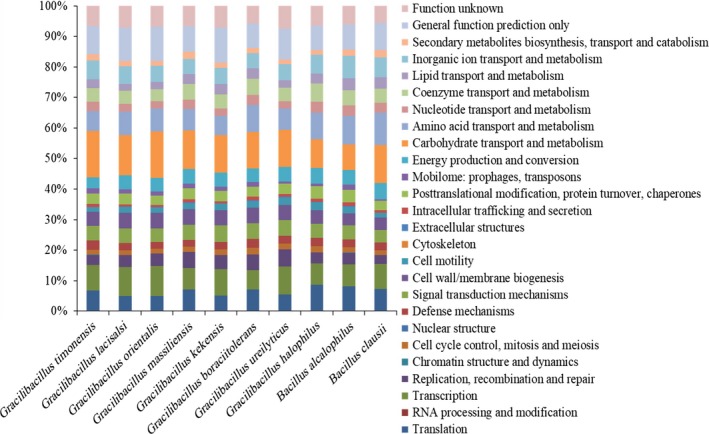
Distribution of functional classes of predicted genes according to the clusters of orthologous groups of proteins of Gracilibacillus timonensis strain Marseille‐P2481T and other compared species
Table 6.
Numbers of orthologous proteins (upper right) and AGIOS values (lower left, %) obtained between compared genomes. The numbers of proteins per genome are indicated in bold
| GT | GL | GO | GM | GK | GB | GU | GH | BA | BC | |
|---|---|---|---|---|---|---|---|---|---|---|
| GT | 4,333 | 2,103 | 2,112 | 2,004 | 2,027 | 1,461 | 1,982 | 1,695 | 1,539 | 1,578 |
| GL | 72.3 | 4,268 | 2,654 | 2,405 | 2,467 | 1,693 | 2,374 | 1,995 | 1,654 | 1,703 |
| GO | 72.1 | 85.2 | 4,313 | 2,370 | 2,412 | 1,686 | 2,318 | 1,940 | 1,656 | 1,710 |
| GM | 72.0 | 77.0 | 77.0 | 3,839 | 2,559 | 1,724 | 2,346 | 1,892 | 1,569 | 1,567 |
| GK | 71.8 | 76.6 | 76.7 | 88.7 | 3,730 | 1,724 | 2,345 | 1,907 | 1,596 | 1,594 |
| GB | 71.0 | 75.2 | 75.2 | 78.1 | 77.9 | 4,587 | 1,612 | 1,408 | 1,166 | 1,151 |
| GU | 70.0 | 72.6 | 72.6 | 72.6 | 72.6 | 71.5 | 4,001 | 1,880 | 1,605 | 1,599 |
| GH | 69.8 | 71.8 | 71.9 | 71.9 | 71.7 | 70.7 | 70.6 | 3,156 | 1,348 | 1,363 |
| BA | 64.5 | 65.6 | 65.4 | 65.6 | 65.6 | 64.7 | 65.1 | 65.1 | 4,269 | 1,532 |
| BC | 62.9 | 63.0 | 62.8 | 62.8 | 62.7 | 62.1 | 62.9 | 63.1 | 66.6 | 4,449 |
GT: Gracilibacillus timonensis Marseille‐P2481; GL: Gracilibacillus lacisalsi DSM 19029; GO: Gracilibacillus orientalis XH‐63; GM: Gracilibacillus massiliensis Awa‐1; GK: Gracilibacillus kekensis K170; GB: Gracilibacillus boraciitolerans JCM 21714; GU: Gracilibacillus ureilyticus MF38; GH: Gracilibacillus halophilus YIM‐C55.5; BA: Bacillus alcalophilus ATCC 27647; BC: Bacillus clausii KSM‐K16.
Table 7.
dDDH values obtained by comparison of all studied genomes
| GL | GO | GM | GK | GB | GU | GH | BA | BC | |
|---|---|---|---|---|---|---|---|---|---|
| GT | 20.5% ± 2.35 | 19.8% ± 2.3 | 21.6% ± 2.35 | 20.1% ± 2.3 | 19.1% ± 2.3 | 21.4% ± 2.35 | 19.3% ± 2.3 | 23.6% ± 2.4 | 28.67% ± 2.4 |
| GL | 29.1% ± 2.4 | 21.0% ± 2.35 | 20.9% ± 2.35 | 20.2% ± 2.3 | 19.4% ± 2.3 | 18.7% ± 2.25 | 18.1% ± 2.25 | 24.4% ± 2.35 | |
| GO | 21.0% ± 2.35 | 20.9% ± 2.35 | 19.9% ± 2.3 | 19.4% ± 2.25 | 18.2% ± 2.25 | 18.4% ± 2.25 | 25.2% ± 2.4 | ||
| GM | 35.4% ± 2.45 | 22.2% ± 2.35 | 19.4% ± 2.3 | 19.1% ± 2.3 | 19.9% ± 2.3 | 31.2% ± 2.5 | |||
| GK | 21.8% ± 2.35 | 19.7% ± 2.3 | 19.2% ± 2.3 | 18.4% ± 2.25 | 29.5% ± 2.45 | ||||
| GB | 18.5% ± 2.25 | 17.4% ± 2.2 | 18.2% ± 2.25 | 33.9% ± 2.5 | |||||
| GU | 16.9% ± 2.2 | 20.9% ± 2.3 | 24.6% ± 2.4 | ||||||
| GH | 27.2% ± 2.4 | 29.8% ± 2.45 | |||||||
| BA | 27.4% ± 2.45 |
GT: Gracilibacillus timonensis Marseille‐P2481; GL: Gracilibacillus lacisalsi DSM 19029; GO: Gracilibacillus orientalis XH‐63; GM: Gracilibacillus massiliensis Awa‐1; GK: Gracilibacillus kekensis K170; GB: Gracilibacillus boraciitolerans JCM 21714; GU: Gracilibacillus ureilyticus MF38; GH: Gracilibacillus halophilus YIM‐C55.5; BA: Bacillus alcalophilus ATCC 27647; BC: Bacillus clausii KSM‐K16.
Table 8.
Average amino acid identity (AAI) values (%) between Gracilibacillus timonensis strain Marseille‐P2481T and other closely related species
| GL | GO | GM | GK | GB | GU | GH | BA | BC | |
|---|---|---|---|---|---|---|---|---|---|
| GT | 68.72 | 68.19 | 68.18 | 67.90 | 68.08 | 64.69 | 64.37 | 51.72 | 50.73 |
| GL | 85.64 | 77.21 | 76.84 | 75.47 | 70.41 | 68.82 | 52.40 | 51.31 | |
| GO | 76.88 | 76.74 | 75.23 | 70.21 | 68.17 | 51.95 | 50.76 | ||
| GM | 90.32 | 79.78 | 70.72 | 68.09 | 52.02 | 50.74 | |||
| GK | 80.04 | 70.55 | 68.19 | 52.31 | 50.83 | ||||
| ²GB | 69.60 | 67.34 | 51.99 | 50.92 | |||||
| GU | 67.03 | 52.53 | 51.16 | ||||||
| GH | 51.53 | 50.77 | |||||||
| BA | 57.85 |
GT: Gracilibacillus timonensis Marseille‐P2481; GL: Gracilibacillus lacisalsi DSM 19029; GO: Gracilibacillus orientalis XH‐63; GM: Gracilibacillus massiliensis Awa‐1; GK: Gracilibacillus kekensis K170; GB: Gracilibacillus boraciitolerans JCM 21714; GU: Gracilibacillus ureilyticus MF38; GH: Gracilibacillus halophilus YIM‐C55.5; BA: Bacillus alcalophilus ATCC 27647; BC: Bacillus clausii KSM‐K16.
4. DISCUSSION
Due to the concept of microbial culturomics, aiming at exploring the diversity of the human microbiota as exhaustively as possible, many new bacterial species have been discovered over the past 5 years (Lagier et al., 2016). This concept is based on the diversification of physicochemical parameters of culture conditions (Lagier et al., 2012, 2016; Lagier, Hugon, et al.,2015) to mimick as closely as possible the entirety of selective constraints that have shaped the human flora. To date, 329 new species have been characterized (Lagier et al., 2017). These new species include 52 species belonging to the order Bacillales, which is one of the most represented bacterial orders (Lagier et al., 2016). Using hypersaline conditions, many hitherto unknown bacteria extremely and or moderately halophilic have been identified in humans, including strain Marseille‐P2481. To the best of our knowledge, this is the first Gracilibacillus species described in the human gut. Whether it is a resident species of the human gut or a transitory species brought by food is as yet unknown. Its phenotypic, phylogenetic, and genomic characteristics suggested that it represents a new species within the genus Gracilibacillus. Members of this genus are generally Gram‐positive bacteria, aerobic, motile, moderately halophile and produce white colonies although G. boraciitolerans forms pink to red colonies (Ahmed, Yokota, & Fujiwara, 2007), and endospore‐forming. However, Gracilibacillus timonensis sp. nov. differs from other Gracilibacillus species in colony color and metabolism of β‐galactosidase, l‐arabinose, and d‐mannitol. In addition, its genomic DNA G + C content differed from those of other Gracilibacillus species, and the dDDH, AAI, and AGIOS values comforted its new species status.
5. CONCLUSION
The moderately halophilic strain Marseille‐P2481 was isolated from a stool sample of a 10‐year‐old healthy Senegalese boy as part of a study of halophilic bacteria from the human gut. Based on its phenotypic, phylogenetic, and genomic characteristics, this strain is proposed to represent a novel species in the genus Gracilibacillus, for which the name Gracilibacillus timonensis sp. nov. is proposed. Strain Marseille‐P2481T is the type strain of Gracilibacillus timonensis sp. nov.
5.1. Description of Gracilibacillus timonensis sp. nov
Gracilibacillus timonensis (ti.mo.nen′sis, N. L adj. masc., timonensis of Timone, the name of the main hospital of Marseille, France, where the type strain was first isolated).
The bacterium is preferentially aerobic but is able to grow in anaerobic and microaerophilic atmospheres at 37°C. Strain Marseille‐P2481T is able to grow in media containing up to 20% (w/v) NaCl, but no growth occurs in the absence of NaCl. The optimal culture conditions are 37°C, pH 7.0‐8.0, and 7.5% (w/v) NaCl. After 48 hr of incubation at 37°C on our home‐made culture medium (7.5% [w/v] NaCl), colonies are creamy orange and circular and have a mean diameter of 0.2 μm. Cells are Gram‐positive, motile rods (with peritrichous flagella) that form endospores rods and are slightly curved, with mean diameter and length of 0.5 and 1.9 μm, respectively.
Using an API ZYM strip, positive results were obtained for esterase, esterase lipase, acid phosphatase, naphtol‐AS‐BI‐phosphohydrolase β‐galactosidase, β‐glucosidase, and α‐glucosidase activities, but no reaction was observed for alkaline phosphatase, lipase, Leucine arylamidase, Valine arylamidase, Cystine arylamidase, α‐galactosidase, β‐glucuronidase, trypsin, α‐chymotrypsin, α‐mannosidase, α‐fucosidase, and N‐acetyl‐β‐glucosaminidase. The API 50CH strip revealed that strain Marseille‐P2481 exhibited esculin hydrolysis, but negative reactions were obtained for d‐arabitol, l‐arabitol, d‐glucose, d‐fructose, d‐fucose, d‐galactose, d‐lactose, d‐maltose, d‐ribose, d‐saccharose, d‐lyxose, d‐mannose l‐sorbose, d‐tagatose, d‐turanose, d‐xylose, l‐xylose, d‐arabinose, l‐arabinose, d‐sorbitol, d‐cellobiose, d‐melezitose, d‐melibiose, d‐trehalose, d‐raffinose, l‐rhamnose, d‐adonitol, d‐mannitol, l‐fucose, amygdalin, arbutin, erythritol, dulcitol, gentiobiose, glycerol, glycogen, inositol, inulin, salicin, starch, xylitol, αD‐glucopyranoside, methyl‐βD‐xylopyranoside, methyl‐αD‐mannopyranoside, potassium gluconate, potassium‐2‐ketogluconate potassium‐5‐ketogluconate, N‐acetylglucosamine. Using an API 20NE strip, fermentation of glucose, urease activity, and metabolism of l‐arginine, esculin and 4‐nitrophenyl‐βD‐galactopyrasinoside were positive. In contrast, nitrate and indole production, gelatinase activity and metabolism of d‐glucose, l‐arabinose, d‐mannose, d‐maltose, d‐mannitol, N‐acetyl‐glucosamine, potassium gluconate, capric acid, malic acid, trisodium citrate, and phenylacetic acid were negative. Cell membrane fatty acids are mainly saturated structures, with 12‐methyl‐tetradecanoic acid (45%) and hexadecanoic acid (16%) being the most abundant. No unsaturated structure was found. The genomic DNA G+C content is 39.8 mol%. The 16S rRNA and genome sequences are deposited in EMBL‐EBI under accession numbers LT223702 and FLKH00000000, respectively. The type strain of Gracilibacillus timonensis is strain Marseille‐P2481T (= CSUR P2481 = DSM 103076).
CONFLICT OF INTEREST
The authors declare no competing interest in relation to this research.
ACKNOWLEDGMENTS
This study was funded by the Méditerranée‐Infection foundation and the French Agence Nationale de la Recherche under reference Investissements d'Avenir Méditerranée Infection 10‐IAHU‐03.
Diop A, Seck EH, Dubourg G, et al. Genome sequence and description of Gracilibacillus timonensis sp. nov. strain Marseille‐P2481T, a moderate halophilic bacterium isolated from the human gut microflora. MicrobiologyOpen. 2019;8:e638 10.1002/mbo3.638
REFERENCES
- Ahmed, I. , Yokota, A. , & Fujiwara, T. (2007). Gracilibacillus boraciitolerans sp. nov., a highly boron‐tolerant and moderately halotolerant bacterium isolated from soil. International Journal of Systematic and Evolutionary Microbiology, 57, 796–802. 10.1099/ijs.0.64284-0 [DOI] [PubMed] [Google Scholar]
- Ashburner, M. , Ball, C. A. , Blake, J. A. , Botstein, D. , Butler, H. , Cherry, J. M. , … Sherlock, G . (2000). Gene ontology: Tool for the unification of biology. The Gene Ontology Consortium. Nature Genetics, 25, 25–29. 10.1038/75556 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Atlas, R. M. , & Snyder, J. W . (2011). Reagents, Stains, and Media: Bacteriology In Versalovic J., Carroll K., Funke G., Jorgensen J., Landry M. & Warnock D. W. (Eds.), Manual of Clinical Microbiology 10th Ed (pp. 272–303). Washington, DC: ASM Press Wash; 10.1128/9781555816728.ch17 [DOI] [Google Scholar]
- Benson, D. A. , Clark, K. , Karsch‐Mizrachi, I. , Lipman, D. J. , Ostell, J. , & Sayers, E. W. (2015). GenBank. Nucleic Acids Research, 43, D30–D35. 10.1093/nar/gku1216 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Cantrell, S. A. , Dianese, J. C. , Fell, J. , Gunde‐Cimerman, N. , & Zalar, P. (2011). Unusual fungal niches. Mycologia, 103, 1161–1174. 10.3852/11-108 [DOI] [PubMed] [Google Scholar]
- Carver, T. , Harris, S. R. , Berriman, M. , Parkhill, J. , & McQuillan, J. A. (2012). Artemis: An integrated platform for visualization and analysis of high‐throughput sequence‐based experimental data. Bioinformatics, 28, 464–469. 10.1093/bioinformatics/btr703 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Carver, T. , Thomson, N. , Bleasby, A. , Berriman, M. , & Parkhill, J. (2009). DNAPlotter: Circular and linear interactive genome visualization. Bioinformatics, 25, 119–120. 10.1093/bioinformatics/btn578 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Chamroensaksri, N. , Tanasupawat, S. , Akaracharanya, A. , Visessanguan, W. , Kudo, T. , & Itoh, T. (2010). Gracilibacillus thailandensis sp. nov., from fermented fish (pla‐ra). International Journal of Systematic and Evolutionary Microbiology, 60, 944–948. 10.1099/ijs.0.011981-0 [DOI] [PubMed] [Google Scholar]
- Chen, Y.‐G. , Cui, X.‐L. , Zhang, Y.‐Q. , Li, W.‐J. , Wang, Y.‐X. , Xu, L.‐H. , … Jiang, C.‐L. (2008). Gracilibacillus halophilus sp. nov., a moderately halophilic bacterium isolated from saline soil. International Journal of Systematic and Evolutionary Microbiology, 58, 2403–2408. 10.1099/ijs.0.65698-0 [DOI] [PubMed] [Google Scholar]
- Chun, J. , Oren, A. , Ventosa, A. , Christensen, H. , Arahal, D. R. , da Costa, M. S. , … Trujillo, M. E. (2018). Proposed minimal standards for the use of genome data for the taxonomy of prokaryotes. International Journal of Systematic and Evolutionary Microbiology, 68, 461–466. 10.1099/ijsem.0.002516 [DOI] [PubMed] [Google Scholar]
- Dione, N. , Sankar, S. A. , Lagier, J.‐C. , Khelaifia, S. , Michele, C. , Armstrong, N. , … Fournier, P.‐E . (2016). Genome sequence and description of Anaerosalibacter massiliensis sp. nov. New Microbes New Infect, 10, 66–76. 10.1016/j.nmni.2016.01.002 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Diop, A. , Khelaifia, S. , Armstrong, N. , Labas, N. , Fournier, P.‐E. , Raoult, D. , & Million, M. (2016). Microbial culturomics unravels the halophilic microbiota repertoire of table salt: Description of Gracilibacillus massiliensis sp. nov. Microbial Ecology in Health and Disease, 27, 10.3402/mehd.v27.32049 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Drancourt, M. , Bollet, C. , Carlioz, A. , Martelin, R. , Gayral, J.‐P. , & Raoult, D. (2000). 16S ribosomal DNA sequence analysis of a large collection of environmental and clinical unidentifiable bacterial isolates. Journal of Clinical Microbiology, 38, 3623–3630. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Drancourt, M. , Bollet, C. , & Raoult, D. (1997). Stenotrophomonas africana sp. nov., an opportunistic human pathogen in Africa. International Journal of Systematic and Evolutionary Microbiology, 47, 160–163. 10.1099/00207713-47-1-160 [DOI] [PubMed] [Google Scholar]
- Finn, R. D. , Clements, J. , Arndt, W. , Miller, B. L. , Wheeler, T. J. , Schreiber, F. , … Eddy, S. R. (2015). HMMER web server: 2015 update. Nucleic Acids Research, 43, W30–W38. 10.1093/nar/gkv397 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Fischer, A. (1895). Untersuchungen über bakterien. Jahrbücher für Wissenschaftliche Botanik, 27, 1–163. [Google Scholar]
- Gao, M. , Liu, Z.‐Z. , Zhou, Y.‐G. , Liu, H.‐C. , Ma, Y.‐C. , Wang, L. , … Ji, X.‐C. (2012). Gracilibacillus kekensis sp. nov., a moderate halophile isolated from Keke Salt Lake. International Journal of Systematic and Evolutionary Microbiology, 62, 1032–1036. 10.1099/ijs.0.030858-0 [DOI] [PubMed] [Google Scholar]
- Garrity, G. M. , & Holt, J. (2001). The road map to the manual In Garrity G. M., Boone D. R., & Castenholz R. W. (Eds.), Bergey's manual of systematic bacteriology (Vol. 1, 2nd ed., pp. 119–169). New York: Springer‐Verlag. [Google Scholar]
- Gibbons, N. E. , & Murray, R. G. E. (1978). Proposals concerning the higher taxa of Bacteria . International Journal of Systematic and Evolutionary Microbiology, 28, 1–6. 10.1099/00207713-28-1-1 [DOI] [Google Scholar]
- Gouret, P. , Paganini, J. , Dainat, J. , Louati, D. , Darbo, E. , Pontarotti, P. , & Levasseur, A . (2011). Integration of evolutionary biology concepts for functional annotation and automation of complex research in evolution: The multi‐agent software system DAGOBAH In Pontarotti P., (ed.) Evolutionary biology – concepts, biodiversity, macroevolution and genome evolution (pp. 71–87). Berlin, Heidelberg: Springer‐Verlag; 10.1007/978-3-642-20763-1_5 [DOI] [Google Scholar]
- Gouret, P. , Vitiello, V. , Balandraud, N. , Gilles, A. , Pontarotti, P. , & Danchin, E. G. (2005). FIGENIX: Intelligent automation of genomic annotation: Expertise integration in a new software platform. BMC Bioinformatics, 6, 198 10.1186/1471-2105-6-198 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Hirota, K. , Hanaoka, Y. , Nodasaka, Y. , & Yumoto, I. (2014). Gracilibacillus alcaliphilus sp. nov., a facultative alkaliphile isolated from indigo fermentation liquor for dyeing. International Journal of Systematic and Evolutionary Microbiology, 64, 3174–3180. 10.1099/ijs.0.060871-0 [DOI] [PubMed] [Google Scholar]
- Huo, Y.‐Y. , Xu, X.‐W. , Cui, H.‐L. , & Wu, M. (2010). Gracilibacillus ureilyticus sp. nov., a halotolerant bacterium from a saline‐alkaline soil. International Journal of Systematic and Evolutionary Microbiology, 60, 1383–1386. 10.1099/ijs.0.016808-0 [DOI] [PubMed] [Google Scholar]
- Hyatt, D. , Chen, G.‐L. , LoCascio, P. F. , Land, M. L. , Larimer, F. W. , & Hauser, L. J. (2010). Prodigal: Prokaryotic gene recognition and translation initiation site identification. BMC Bioinformatics, 11, 119 10.1186/1471-2105-11-119 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Jeon, C. O. , Lim, J.‐M. , Jang, H. H. , Park, D.‐J. , Xu, L.‐H. , Jiang, C.‐L. , & Kim, C.‐J. (2008). Gracilibacillus lacisalsi sp. nov., a halophilic Gram‐positive bacterium from a salt lake in China. International Journal of Systematic and Evolutionary Microbiology, 58, 2282–2286. 10.1099/ijs.0.65369-0 [DOI] [PubMed] [Google Scholar]
- Käll, L. , Krogh, A. , & Sonnhammer, E. L. (2004). A combined transmembrane topology and signal peptide prediction method. Journal of Molecular Biology, 338, 1027–1036. 10.1016/j.jmb.2004.03.016 [DOI] [PubMed] [Google Scholar]
- Khelaifia, S. , Lagier, J.‐C. , Bibi, F. , Azhar, E. I. , Croce, O. , Padmanabhan, R. , … Raoult, D. (2016). Microbial culturomics to map halophilic bacterium in human gut: Genome sequence and description of Oceanobacillus jeddahense sp. nov. Omics: A Journal of Integrative Biology, 20, 248–258. 10.1089/omi.2016.0004 [DOI] [PubMed] [Google Scholar]
- Kim, P. , Lee, J.‐C. , Park, D.‐J. , Shin, K.‐S. , Kim, J.‐Y. , & Kim, C.‐J. (2012). Gracilibacillus bigeumensis sp. nov., a moderately halophilic bacterium from solar saltern soil. International Journal of Systematic and Evolutionary Microbiology, 62, 1857–1863. 10.1099/ijs.0.034264-0 [DOI] [PubMed] [Google Scholar]
- Kim, M. , Oh, H.‐S. , Park, S.‐C. , & Chun, J. (2014). Towards a taxonomic coherence between average nucleotide identity and 16S rRNA gene sequence similarity for species demarcation of prokaryotes. International Journal of Systematic and Evolutionary Microbiology, 64, 346–351. 10.1099/ijs.0.059774-0 [DOI] [PubMed] [Google Scholar]
- Kimura, M. (1980). A simple method for estimating evolutionary rates of base substitutions through comparative studies of nucleotide sequences. Journal of Molecular Evolution, 16, 111–120. 10.1007/BF01731581 [DOI] [PubMed] [Google Scholar]
- Klappenbach, J. A. , Goris, J. , Vandamme, P. , Coenye, T. , Konstantinidis, K. T. , & Tiedje, J. M. (2007). DNA–DNA hybridization values and their relationship to whole‐genome sequence similarities. International Journal of Systematic and Evolutionary Microbiology, 57, 81–91. 10.1099/ijs.0.64483-0 [DOI] [PubMed] [Google Scholar]
- Klenk, H.‐P. , Meier‐Kolthoff, J. P. , & Göker, M . (2014). Taxonomic use of DNA G+C content and DNA?DNA hybridization in the genomic age. International Journal of Systematic and Evolutionary Microbiology, 64, 352–356. 10.1099/ijs.0.056994-0 [DOI] [PubMed] [Google Scholar]
- Konstantinidis, K. T. , Ramette, A. , & Tiedje, J. M. (2006). The bacterial species definition in the genomic era. Philosophical Transactions of the Royal Society B: Biological Sciences, 361, 1929–1940. 10.1098/rstb.2006.1920 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Lagesen, K. , Hallin, P. , Rødland, E. A. , Stærfeldt, H.‐H. , Rognes, T. , & Ussery, D. W. (2007). RNAmmer: Consistent and rapid annotation of ribosomal RNA genes. Nucleic Acids Research, 35, 3100–3108. 10.1093/nar/gkm160 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Lagier, J.‐C. , Armougom, F. , Million, M. , Hugon, P. , Pagnier, I. , Robert, C. , … Raoult, D. (2012). Microbial culturomics: Paradigm shift in the human gut microbiome study. Clinical Microbiology & Infection, 18, 1185–1193. 10.1111/1469-0691.12023 [DOI] [PubMed] [Google Scholar]
- Lagier, J. C. , Drancourt, M. , Charrel, R. , Bittar, F. , La Scola, B. , Ranque, S. , & Raoult, D. (2017). Many more microbes in humans: Enlarging the microbiome repertoire. Clinical Infectious Diseases, 65, S20–S29. 10.1093/cid/cix404 [DOI] [PubMed] [Google Scholar]
- Lagier, J.‐C. , Hugon, P. , Khelaifia, S. , Fournier, P.‐E. , La Scola, B. , & Raoult, D. (2015). The rebirth of culture in microbiology through the example of culturomics to study human gut microbiota. Clinical Microbiology Reviews, 28, 237–264. 10.1128/CMR.00014-14 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Lagier, J.‐C. , Khelaifia, S. , Alou, M. T. , Ndongo, S. , Dione, N. , Hugon, P. , … Raoult, D. (2016). Culture of previously uncultured members of the human gut microbiota by culturomics. Nature Microbiology, 1, 16203 10.1038/nmicrobiol.2016.203 [DOI] [PubMed] [Google Scholar]
- Lagier, J.‐C. , Khelaifia, S. , Azhar, E. I. , Croce, O. , Bibi, F. , Jiman‐Fatani, A. A. , … Raoult, D. (2015). Genome sequence of Oceanobacillus picturae strain S1, an halophilic bacterium first isolated in human gut. Standards in Genomic Sciences, 10, 10.1186/s40793-015-0081-2 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Larkin, M. A. , Blackshields, G. , Brown, N. P. , Chenna, R. , McGettigan, P. A. , McWilliam, H. , … Higgins, D. G. (2007). Clustal W and Clustal X version 2.0. Bioinformatics, 23, 2947–2948. 10.1093/bioinformatics/btm404 [DOI] [PubMed] [Google Scholar]
- Lechner, M. , Findeiss, S. , Steiner, L. , Marz, M. , Stadler, P. F. , & Prohaska, S. J. (2011). Proteinortho: Detection of (co‐) orthologs in large‐scale analysis. BMC Bioinformatics, 12, 124 10.1186/1471-2105-12-124 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Lowe, T. M. , & Eddy, S. R. (1997). tRNAscan‐SE: A program for improved detection of transfer RNA genes in genomic sequence. Nucleic Acids Research, 25, 955–964. 10.1093/nar/25.5.0955 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Ludwig, W. , Schleifer, K. H. , & Whitman, W. B . (2009). Class I. Bacilli class nov In: De Vos P., Garrity G., Jones D., Krieg N. R., Ludwig W., Rainey F. A., Schleifer K. H. & Whitman W. B. (Eds.), Bergey's manual of systematic bacteriology (Vol. 3, 2nd ed., pp. 19–20). New York: Springer‐Verlag. [Google Scholar]
- Meier‐Kolthoff, J. P. , Auch, A. F. , Klenk, H.‐P. , & Göker, M. (2013). Genome sequence‐based species delimitation with confidence intervals and improved distance functions. BMC Bioinformatics, 14, 60 10.1186/1471-2105-14-60 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Murray, R. G. E . (1984). The higher taxa, or, a place for everything…? In: Holt J. G. (ed.), Bergey's manual of systematic bacteriology (Vol. 1, 1st ed., pp. 31–34). Baltimore: The Williams and Wilkins Co. [Google Scholar]
- Padmanabhan, R. , Mishra, A. K. , Raoult, D. , & Fournier, P.‐E. (2013). Genomics and metagenomics in medical microbiology. Journal of Microbiol Methods, 95, 415–424. 10.1016/j.mimet.2013.10.006 [DOI] [PubMed] [Google Scholar]
- Pagani, I. , Liolios, K. , Jansson, J. , Chen, I.‐M. A. , Smirnova, T. , Nosrat, B. , … Kyrpides, N. C. (2012). The Genomes OnLine Database (GOLD) v. 4: Status of genomic and metagenomic projects and their associated metadata. Nucleic Acids Research, 40, D571–D579. 10.1093/nar/gkr1100 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Parte, A. C. (2014). LPSN—list of prokaryotic names with standing in nomenclature. Nucleic Acids Research, 42, D613–D616. 10.1093/nar/gkt1111 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Ramasamy, D. , Mishra, A. K. , Lagier, J.‐C. , Padhmanabhan, R. , Rossi, M. , Sentausa, E. , … Fournier, P.‐E. (2014). A polyphasic strategy incorporating genomic data for the taxonomic description of novel bacterial species. International Journal of Systematic and Evolutionary Microbiology, 64, 384–391. 10.1099/ijs.0.057091-0 [DOI] [PubMed] [Google Scholar]
- Richter, M. , & Rosselló‐Móra, R. (2009). Shifting the genomic gold standard for the prokaryotic species definition. Proceedings of the National Academy of Sciences, 106, 19126–19131. 10.1073/pnas.0906412106 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Rodriguez‐R, L. M. , & Konstantinidis, K. T. (2014). Bypassing cultivation to identify bacterial species. Microbe, 9, 111–118. [Google Scholar]
- Sasser, M . (1990). Identification of bacteria by gas chromatography of cellular fatty acids. http://natasha.eng.usf.edu/gilbert/courses/Biotransport%20Phenomena/pdf/bacteria_gc_1.pdf (Accessed June 13, 2017)
- Senghor, B. , Seck, E. H. , Khelaifia, S. , Bassène, H. , Sokhna, C. , Fournier, P.‐E. , … Lagier, J.‐C. (2017). Description of “Bacillus dakarensis” sp. nov., “Bacillus sinesaloumensis” sp. nov., “Gracilibacillus timonensis” sp. nov., “Halobacillus massiliensis” sp. nov., “Lentibacillus massiliensis” sp. nov., “Oceanobacillus senegalensis” sp. nov., “Oceanobacillus timonensis” sp. nov., “Virgibacillus dakarensis” sp. nov. and “Virgibacillus marseillensis” sp. nov., nine halophilic new species isolated from human stool. New Microbes and new Infections, 17, 45–51. 10.1016/j.nmni.2017.01.010 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Sentausa, E. , & Fournier, P.‐E. (2013). Advantages and limitations of genomics in prokaryotic taxonomy. Clinical Microbiology & Infection, 19, 790–795. 10.1111/1469-0691.12181 [DOI] [PubMed] [Google Scholar]
- Skerman, V. B. D. , & Sneath, P. H. A. (1980). Approved list of bacterial names. International Journal of Systematic Bacteriology, 30, 225–420. 10.1099/00207713-30-1-225 [DOI] [Google Scholar]
- Stackebrandt, E. , & Ebers, J. (2006). Taxonomic parameters revisited: Tarnished gold standards. Microbiol Today., 33, 152–155. [Google Scholar]
- Tamura, K. , Stecher, G. , Peterson, D. , Filipski, A. , & Kumar, S. (2013). MEGA6: Molecular evolutionary genetics analysis version 6.0. Molecular Biology and Evolution, 30, 2725–2729. 10.1093/molbev/mst197 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Tang, S.‐K. , Wang, Y. , Lou, K. , Mao, P.‐H. , Jin, X. , Jiang, C.‐L. , … Li, W.‐J. (2009). Gracilibacillus saliphilus sp. nov., a moderately halophilic bacterium isolated from a salt lake. International Journal of Systematic and Evolutionary Microbiology, 59, 1620–1624. 10.1099/ijs.0.006569-0 [DOI] [PubMed] [Google Scholar]
- Wainø, M. , Tindall, B. J. , Schumann, P. , & Ingvorsen, K. (1999). Gracilibacillus gen. nov., with description of Gracilibacillus halotolerans gen. nov., sp. nov.; transfer of Bacillus dipsosauri to Gracilibacillus dipsosauri comb. nov., and Bacillus salexigens to the genus Salibacillus gen. nov., as Salibacillus salexigens comb. nov. International Journal of Systematic and Evolutionary Microbiology, 49, 821–831. 10.1099/00207713-49-2-821 [DOI] [PubMed] [Google Scholar]
- Woese, C. R. , Kandler, O. , & Wheelis, M. L . (1990). Towards a natural system of organisms: Proposal for the domains Archaea, Bacteria, and Eukarya . Proceedings of the National Academy of Sciences U.S.A., 87, 4576–4579. 10.1073/pnas.87.12.4576 [DOI] [PMC free article] [PubMed] [Google Scholar]