

Abstract
l-Ascorbic acid (AsA) and its metabolic precursors give rise to oxalic acid (OxA) found in calcium oxalate crystals in specialized crystal idioblast cells in plants; however, it is not known if AsA and OxA are synthesized within the crystal idioblast cell or transported in from surrounding mesophyll cells. Isolated developing crystal idioblasts from Pistia stratiotes were used to study the pathway of OxA biosynthesis and to determine if idioblasts contain the entire path and are essentially independent in OxA synthesis. Idioblasts were supplied with various 14C-labeled compounds and examined by micro-autoradiography for incorporation of 14C into calcium oxalate crystals. [14C]OxA gave heavy labeling of crystals, indicating the isolated idioblasts are functional in crystal formation. Incubation with [1-14C]AsA also gave heavy labeling of crystals, whereas [6-14C]AsA gave no labeling. Labeled precursors of AsA (l-[1-14C]galactose; d-[1-14C]mannose) also resulted in crystal labeling, as did the ascorbic acid analog, d-[1-14C]erythorbic acid. Intensity of labeling of isolated idioblasts followed the pattern OxA > AsA (erythorbic acid) > l-galactose > d-mannose. Our results demonstrate that P. stratiotes crystal idioblasts synthesize the OxA used for crystal formation, the OxA is derived from the number 1 and 2 carbons of AsA, and the proposed pathway of ascorbic acid synthesis via d-mannose and l-galactose is operational in individual P. stratiotes crystal idioblasts. These results are discussed with respect to fine control of calcium oxalate precipitation and the concept of crystal idioblasts as independent physiological compartments.
Many plants sequester excess calcium taken up from the environment as the insoluble salt, calcium oxalate (Arnott and Pautard, 1970; Franceschi and Horner, 1980; Libert and Franceschi, 1987; Franceschi and Loewus, 1995; Horner and Wagner, 1995). This most often occurs in vacuoles of cells called crystal idioblasts (Foster, 1956), which are specialized for this function. Crystal idioblasts act as high capacity sinks for removal of excess calcium from nearby tissues (Franceschi and Horner, 1979; Borchert, 1985, 1986; Franceschi, 1989; Franceschi and Loewus, 1995). While a role of crystal idioblasts in calcium sequestration has been demonstrated in a number of systems, the physiology of the specialized cells is not well understood. In particular, the source of oxalic acid (OxA) used for precipitation of calcium in crystal idioblasts is not clearly established (Li and Franceschi, 1990).
For many years, OxA was considered a metabolic end product and it was thought that calcium oxalate was formed to maintain low soluble levels of this potentially toxic acid (for review, see Franceschi and Horner, 1980; Libert and Franceschi, 1987; Franceschi and Loewus, 1995). Recent studies, however, have shown OxA is synthesized in response to increased calcium (Keates et al., 2000), which suggests the biosynthetic pathway can be induced by calcium in calcium oxalate accumulating plants. Gycolate, glyoxalate, oxaloacetate, and citrate have all been proposed as precursors of OxA in plants (Hodgkinson, 1977), however the relative contribution to oxalate synthesis through the diverse pathways associated with these compounds has not been determined. After Loewus and colleagues discovered that l-ascorbic acid (AsA) is cleaved between C2 and C3 to yield OxA in plant tissues (for review, see Loewus, 1999), some studies were conducted that indicated AsA could provide the substrate for OxA synthesis used for calcium oxalate formation (Franceschi and Horner, 1979; Franceschi, 1987; Horner et al., 2000) as well as for soluble oxalate accumulated in leaf vacuoles (Wagner, 1981). Wheeler et al. (1998; Smirnoff and Wheeler, 2000) recently found that d-Man and l-Gal were intermediates in the biosynthetic pathway leading to AsA in plants (referred to here as the “Smirnoff-Wheeler pathway”), and Keates et al. (2000) showed that axenic Pistia stratiotes plants were capable of incorporating label from [1-14C]AsA, l-[1-14C]Gal, and d-[1-14C]erythorbic acid (an ascorbic acid analog) into OxA and calcium oxalate crystals. Label from [6-14C]AsA showed little incorporation into OxA or calcium oxalate, consistent with C1 and C2 of AsA giving rise to OxA (Wagner and Loewus, 1973). Keates et al. (2000) also found that label from glycolate and glyoxylate did not give rise to any significant label in OxA or calcium oxalate in the whole plant system. The results showed that AsA is synthesized from l-Gal in P. stratiotes and is a significant substrate for the OxA used in calcium oxalate formation.
Although there is mounting evidence that AsA is the major substrate for synthesis of OxA used in crystal idioblasts, it remains to be determined where the pathway is located relative to oxalate precipitation. Because idioblasts can form masses of crystals within a period of an hour (Franceschi, 1989) it seems likely that OxA is produced within the idioblast directly rather than being imported from surrounding cells (Li and Franceschi, 1990). This is a central question to our understanding the biology of this high capacity calcium accumulation mechanism since coordination of oxalate synthesis with calcium accumulation and precipitation is likely a key regulatory step in crystal idioblast development. This study uses isolated developing crystal idioblasts from P. stratiotes to study the localization of the biosynthetic pathway of OxA synthesis relative to calcium oxalate-forming crystal idioblasts. Disruption of symplasmic connections with adjacent mesophyll cells ensures that OxA produced by mesophyll cells cannot be transferred to crystal idioblasts. Thus, exposure of the idioblasts to various 14C-labeled potential substrates has allowed us to determine if an OxA synthesis pathway is present in the idioblasts and, if so, what is the major precursor to OxA synthesis and what portion of the biosynthetic pathway is present. After incubation of isolated idioblasts with labeled compounds, cells were examined by microautoradiography to look for incorporation of label into calcium oxalate crystals. Our findings are consistent with comparable studies on whole plants (Keates et al., 2000) in showing that AsA is a major precursor of OxA biosynthesis for calcium oxalate formation. Moreover, we demonstrate that the biosynthetic pathways for both AsA and OxA reside within the developing idioblasts.
RESULTS
Crystal Idioblast Cells Are Abundant
A brief description of the calcium oxalate crystal idioblasts formed in our system is necessary for proper interpretation of the micro-autoradiographs. P. stratiotes has two types of calcium oxalate crystal idioblasts; large elongate cells containing bundles of needle-shaped raphide crystals and smaller compact cells with a single multi-faceted druse crystal (Fig. 1). The crystals are formed within the vacuole of these specialized cells. In rapidly expanding leaves of the shoot tips, crystal idioblasts of a broad range of developmental stages occur (Fig. 1A).
Figure 1.
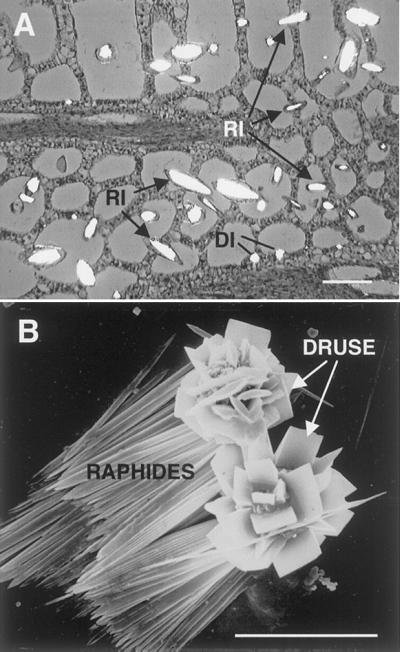
General features of calcium oxalate idioblasts and crystals in P. stratiotes. A, Section through developing leaf viewed with partially crossed polarizing filters. Crystal idioblasts are abundant and appear as bright cells in various stages of development. Raphide idioblasts (RI) become elongated as they grow, whereas druse idioblasts (DI) are smaller and compact. Bar = 30 μm. B, Scanning electron microscopy of isolated crystals. Druse crystals are multifaceted spherical conglomerates, whereas raphides occur as bundles of needle-shaped crystals. Bar = 23 μm.
Isolated Idioblasts Can Produce Calcium Oxalate in Vitro
Micro-autoradiography, as used here, allows for determination of the presence and relative amount of label associated with insoluble products such as calcium oxalate. Cells or cell components that have incorporated 14C from supplied radioactive substrates have silver grains over them, which in our micrographs appear as red particles. Exposure to [14C]OxA demonstrates that isolated crystal idioblasts of P. stratiotes are metabolically active and are capable of taking up labeled compounds and can continue to produce calcium oxalate. Isolated raphide and druse crystal idioblasts accumulated large amounts of label from [14C]OxA into crystals as indicated by the intensity of labeling (Fig. 2A). There is no label associated the mesophyll protoplasts, which do not produce calcium oxalate. Some raphide crystal idioblasts break during post-labeling processing, releasing crystals from the bundles, and often these crystals are heavily labeled (Fig. 2, A, C, and D). This clearly shows that the label is present as calcium oxalate. Control protoplasts unexposed to radioisotope have no silver grains associated with them (Fig. 2B), a further demonstration that the silver grains are due to radioactivity and not to some physio-chemical interaction between protoplasts or crystals and the photosensitive emulsion.
Figure 2.

Micro-autoradiography of incorporation of radioactive compounds of the ascorbic acid biosynthesis pathway into specific protoplast types from P. stratiotes shoots. Reflected/transmitted confocal images with red dots indicating location of radioactive label. Bars are in micrometers. A, [14C]OxA leads to heavy labeling of druse (DI) and developing raphide (RI) idioblasts. Some free crystals (C) are also labeled, whereas mesophyll protoplasts (M) are unlabeled. B, Incubated in buffer alone with no radioactivity. Red patches seen are light scattering from trapped air bubbles. C, Incubation with l-[1-14C]ascorbic acid. Raphide (RI) and druse (DI) idioblasts, plus some free crystals are heavily labeled, whereas mesophyll (M) remain unlabeled. D, Single raphide crystal from cells incubated in l-[1-14C]ascorbic acid. Note discrete labeling along the length of the crystal. E, Exposure to l-[6-14C]ascorbic acid. Note lack of incorporation of label into idioblasts. F, d-[1-14C] erythorbic acid is incorporated into raphide idioblasts but not mesophyll cells. The focal plane at the emulsion layer is at one end of idioblast showing heavy labeling. Label around idioblast is from scatter of β-particles away from crystal bundle. G, Label from l-[1-14C]Gal is incorporated into developing raphide idioblasts but not mesophyll cells.
Crystal Idioblasts Synthesize OxA from C1 and C2 of AsA
Exposure of protoplasts to [1-14C]AsA resulted in very heavy labeling of crystal idioblasts, but not mesophyll protoplasts (Fig. 2C). Label was also seen in individual crystals released from idioblasts during processing (Fig. 2, C and D). When the protoplasts were incubated with [6-14C]AsA, there was no incorporation of label into crystal idioblasts or mesophyll protoplasts (Fig. 2E). These results show that crystal idioblasts have the metabolic machinery to synthesize OxA from AsA, carbons 1 and 2 of AsA are used for this synthesis, and the OxA produced from AsA is used for crystal formation.
Keates et al. (2000) found that intact P. stratiotes plants oxidatively cleaved d-erythorbic acid, the carbon-5 diasteriomer of AsA, to produce OxA. Isolated idioblasts were also found to be able to use d-[1-14C]erythorbic acid very effectively, giving heavy labeling of crystals and crystal bundles (Fig. 2F). This is interesting in that it indicates a process that converts d-erythorbic acid to AsA prior to cleavage, or a mechanism for carbon 2/carbon 3 cleavage that accepts d-erythorbic acid as well as AsA to form OxA and a four carbon fragment.
Crystal Idioblasts Can Synthesize AsA Used for Oxalate Production
Because d-Man and l-Gal are precursors of AsA (Smirnoff and Wheeler, 2000), we used 14C-labeled forms of these compounds to determine if crystal idioblasts are capable of synthesizing their own AsA. Exposure of idioblasts to l-[1-14C]Gal resulted in labeled crystal idioblasts and labeled crystals released from these idioblasts (Fig. 2G). Mesophyll protoplasts were unlabeled. d-[1-14C]Man also gave rise to idioblast labeling but labeling was quite low (not shown). According to the scheme for AsA biosynthesis set forth by Smirnoff and Wheeler (2000), there are eight enzymic steps between d-Man and AsA. The fact that even a small amount of 14C from d-[1-14C]Man was incorporated into idioblast calcium oxalate crystals provides supporting evidence for the Smirnoff-Wheeler pathway operating in idioblasts. We found in general that labeling diminished as compounds further removed from OxA were tested. Thus, intensity of labeling of isolated idioblasts (OxA > AsA > l-Gal > d-Man) followed a pattern consistent with the view that the entire pathway of AsA synthesis is present in crystal idioblasts as well as the processes involving metabolism of AsA to OxA.
Glycolic Acid Is Not a Major Oxalate Precursor in Crystal Idioblasts
Glycolic and glyoxylic acids have been proposed as OxA precursors, but whole plant studies with P. stratiotes indicate they are secondary to AsA (Keates et al., 2000). Calcium [1-14C]glycolate labeled crystal idioblasts less effectively than [14C]OxA or [1-14C]AsA (Fig. 3A). In contrast, [1-14C]glyoxylic acid labeled mesophyll as well as crystal cells strongly (Fig. 3B). In both cases, labeling of idioblasts when it occurred appeared to be heavily associated with the idioblast periphery. Labeling patterns with glyoxylic acid in particular suggests these α-hydroxy acids are converted into amino acids and then incorporated into protein in both cell types since mesophyll cells do not produce calcium oxalate. This interpretation is consistent with labeling patterns seen in sections of whole tissues exposed to these compounds (Keates et al., 2000). It is important to point out that in the present experiments, small soluble molecules are lost during subsequent protoplast processing so remaining 14C must reflect incorporation into polymeric structures such as calcium oxalate, protein, nucleic acids, lipids, and/or polysaccharides.
Figure 3.

Micro-autoradiography of incorporation of radioactive compounds into developing P. stratiotes leaf protoplasts. Bars are in micrometers. A, Label from [14C]glycolic acid is primarily associated with the cytoplasmic periphery of some raphide idioblasts (RI) and mesophyll (M) protoplasts. B, Exposure of protoplasts to [14C]glyoxylic acid results in very heavy labeling of mesophyll protoplasts and the periphery of some raphide idioblasts. C, Heavy labeling of raphide idioblasts incubated in l-[1-14C]ascorbic acid. D, Idioblasts incubated with l-[1-14C]ascorbic acid in the presence of 1 mm OxA. Note the decrease in incorporation of label into the raphide idioblast and that heaviest incorporation is at the ends of the crystals where maximum growth is occurring.
Exogenous Oxalate Inhibits Incorporation of 14C into Crystals from [1-14C] AsA
When 1 mm OxA was included in the incubation medium along with [1-14C]AsA, there was a noticeable reduction in the amount of 14C in the idioblasts (Fig. 3D) as compared with cells incubated with AsA alone (Fig. 3C). A similar reduction in idioblast labeling was seen when OxA was supplied along with l-[1-14C]Gal (not shown). This may be due to feedback regulation of the pathway between AsA and OxA such that the amount of OxA produced is not greater than that required for crystal precipitation.
CONCLUSIONS
Crystal idioblasts are cells that are specialized for calcium oxalate formation and they require large amounts of OxA to precipitate the excess tissue calcium they accumulate. We show here that this cell type is self sufficient in OxA biosynthesis and thus does not require import of OxA from adjacent mesophyll cells. Results also clearly demonstrate that AsA is a major, if not the major, carbon source for OxA used for crystal formation. The labeling experiments show that crystal idioblasts contain the pathway for synthesis of OxA from carbons 1 and 2 of AsA. Incorporation of label into crystals from d-[1-14C]Man and l-[1-14C]Gal also demonstrate that the pathway for AsA synthesis is present in the idioblasts. Thus, other than the need for basic carbon and nitrogen substrates (i.e. Suc and amino acids), crystal idioblasts in P. stratiotes leaves are independent of adjoining cells for the critical processes leading to AsA and OxA synthesis. Crystal idioblasts, then, are enriched not only in the components needed for rapid uptake and sequestration of calcium (P.A. Nakata, T.A. Kostman, and V.R. Franceschi, unpublished data), but also are fully capable of producing the OxA needed to complete this important physiological process.
These findings are important to our understanding of how crystal idioblasts are able to coordinate calcium uptake with crystal precipitation, independent of metabolic events occurring in adjacent mesophyll cells. Crystal idioblasts operate as independent physiological entities with a specialized structure and metabolism that provides a function of importance to the physiology of the surrounding cells and the plant as a whole, that is, removal and storage of excess calcium. In a sense, idioblasts serve a complex function to the plant that is more like a tissue or organ, and are thus like a single-celled organ rather than a specialized cell of a more complex organ. The competition experiment with unlabeled OxA and [1-14C]AsA suggests a feed-back system that allows the idioblast to regulate synthesis of OxA from AsA. This system should be critical to the very highly regulated process of crystal precipitation and to avoiding synthesis of OxA that exceeds the capacity for calcium uptake or transport into the appropriate compartments. The subcellular location of OxA synthesis is not known but the idioblasts are enriched in an unusual plastid type. Given that chloroplasts have high AsA levels, it is possible that crystalloplastids may also accumulate AsA and OxA synthesis may be supported in this compartment. Experiments to test this possibility are contemplated.
In summary, our results complement those of Keates et al. (2000), demonstrating P. stratiotes uses AsA for OxA synthesis and extends them to show this occurs in the crystal idioblasts themselves. The results do not support a major role of glycolate and glyoxylate in OxA synthesis in this cell type. The labeling experiments also indicate the newly discovered AsA biosynthetic pathway (Wheeler et al., 1998; Smirnoff and Wheeler, 2000) is operating in crystal idioblasts as well. The regulation of this pathway relative to regulation of OxA synthesis in the idioblasts will be interesting to explore and could provide important basic information on metabolic regulation of AsA synthesis and turnover in general.
MATERIALS AND METHODS
Generation of Protoplasts
Protoplasts were generated to ensure that there were no symplasmic connections between idioblasts and mesophyll cells. This would guarantee that any labeling observed in the “isolated” idioblast cell type was due to incorporation of the 14C-labeled test compound and not due to secondary transfer of the labeled compound or its products via mesophyll cells.
Pistia stratiotes shoot tips were collected from growth chamber plants and rinsed with tap water. All but the first subtending leaf were removed. Shoot tips were sliced into 1-mm2 pieces (approximately 200 mg) and placed immediately in 5 mL of digestion mix consisting of 2.65% (w/v) Worthington cellulase, 2% (w/v) Pectolyase Y-23, 1% (w/v) bovine serum albumin, 0.4 m mannitol, 10 mm CaCl2, and 5 mm MES [2-(N-morpholino)ethanesulfonic acid], pH 5.8. Tissue and enzyme mix was then placed in 20-mL scintillation vials and incubated 2 h at 37°C in a water bath. After digestion the slurry was gently pipetted in and out of a transfer pipette to break up tissue and release individual protoplasts. Protoplast/digestion media mix was then aliquoted into glass test tubes and spun 2 min at 150g in a clinical centrifuge to gently pellet protoplasts. After centrifugation, supernatant was removed and protoplasts were resuspended in Suc buffer (SB: 0.4 m Suc, 2 mm CaCl2, 5 mm HEPES [4-(2-hydroxyethyl)-1-piperazineethanesulfonic acid], pH 7.0). The resulting suspension was then filtered through a 220-μm nylon mesh into a large plastic centrifuge tube. This filtrate was then pipetted into glass test tubes and spun 2 min at 150g to pellet protoplasts. Following this, we removed as much supernatant as possible without removing suspended protoplasts. The mix was then distributed into small glass test tubes, 230 μL per tube. It was determined that the protoplasts did not need to be separated for this type of experiment. Suspensions used for all experiments contained a mixture of crystal idioblast protoplasts and mesophyll protoplasts. Idioblast protoplasts when dried down onto slides appear as either elongated profiles (raphide idioblasts) or small circular profiles (druse idioblasts), whereas mesophyll protoplasts often appeared as irregular clumps or clusters of clumps when dried down.
Labeling and Micro-Autoradiography
All labeled compounds were administered at the same specific activity. For each labeled substrate used, the amount of unlabeled “carrier” needed to produce a final concentration of 1 mm substrate species (unlabeled + 5 μCi of label) was prepared in incubation medium. This carrier consisted of SB + 0.1% (w/v) casein hydrolysate. Two-hundred-fifty microliters of incubation solution containing the appropriate carrier substrate was then added to 230 μL of protoplasts (total volume, 480 μL). Eight different substrates were used with the following activities (and hence microliter amounts) added to the protoplast mixture: d-[1-14C]Man, 0.3 μCi μL−1 (16.67 μL); l-[1-14C]Gal, 0.1 μCi μL−1 (50 μL); [1-14C]glyoxylic acid, 0.1 μCi μL−1 (50 μL); [14C]OxA, 0.1 μCi μL−1 (50 μL); [1-14C]AsA, 0.1 μCi μL−1 (50 μL); d-[1-14C ]erythorbic acid, 0.1 μCi μL−1 (50 μL); [6-14C]AsA, 0.1 μCi μL−1 (50 μL); and calcium [1-14C]glycolate, 0.1 μCi μL−1 (50 μL). Protoplasts were then incubated at 20°C for 4 h in the radiolabeled incubation solution. Following incubation, protoplasts were allowed to settle to the bottom of the tube, and a 200-μL aliquot of supernatant was removed and frozen for liquid scintillation analysis. Protoplasts were then rinsed two times with SB, re-suspended in a large volume of SB, spun at 150g to pellet, then supernatant was removed. After a second rinse, protoplasts were fixed 2 h at 4°C by resuspension in 1.25% (v/v) glutaraldehyde in SB.
Following fixation, protoplasts were pelleted, then rinsed as described above, two times with SB, one time with 1:1 water:SB, then three times with distilled water. After the final rinse a small amount of water was left in the tube and the contents were gently resuspended. A drop of the protoplasts/water mix was placed onto gelatin-coated slides and gently spread out to disperse the protoplasts. Slides were left to air dry prior to being coated with a 1:2 dilution of Ilford LP4 Nuclear Tracking Emulsion in water. After dipping slides in the emulsion, the slides were left to dry upright for 2 h then stored in total darkness (3–36 d) at 4°C prior to development. Slides were developed in undiluted Kodak D-19 for 5 min, rinsed in running water for 1 min, then fixed in Kodak Rapid Fixer for 10 min, followed by a 10-min running water rinse. Slides were then stained for 15 s with 0.5% (w/v) aqueous Safranin O, rinsed, dried, and sealed under a coverslip with immersion oil. For light microscopy images, slides were examined and images captured using a laser-scanning confocal microscope (MRC 1024, BioRad, Hercules, CA) operated in a transmitted/reflected mode.
General Structure of Calcium Oxalate Crystals and Idioblasts
P. stratiotes shoot tips were fixed in formaldehyde-acetic acid-alcohol (4% [v/v] formaldehyde, 5% [v/v] acetic acid, and 48% [v/v] ethanol) for 24 h, dehydrated with ethanol, and infiltrated with paraffin. Twelve-micrometer-thick sections were dried onto gelatin coated slides, the paraffin removed, and the tissue was stained with Safranin O and Fast Green. Sections were photographed with partially crossed polarizing filters that cause the birefringent crystals to appear as very bright areas against the darker tissue structure.
For scanning electron microscopy of crystal structure, protoplasts were ruptured by transfer to water, and the contents allowed to settle. After washing, the dense contents were pipetted onto a coverslip, dried, and coated with gold prior to examination on a scanning electron microscope (model S-570, Hitachi, Tokyo).
ACKNOWLEDGMENT
Confocal microscopy was done at the Electron Microscopy Center, Washington State University.
Footnotes
This work was supported by the National Science Foundation (grant no. MCB–9904562 to V.R.F.) and by the College of Agriculture and Home Economics Research Center, Washington State University (project no. 0266 to F.A.L.).
LITERATURE CITED
- Arnott HJ, Pautard FGE. Calcification in plants. In: Shraer H, editor. Biological Calcification: Cellular and Molecular Aspects. New York: Appleton-Century-Crofts; 1970. pp. 375–446. [Google Scholar]
- Borchert R. Calcium induced patterns of calcium oxalate crystals in isolated leaflets of Gleditsia triacanthos L. and Albizia julibrissin Durazz. Planta. 1985;165:301–310. doi: 10.1007/BF00392226. [DOI] [PubMed] [Google Scholar]
- Borchert R. Calcium acetate induces calcium uptake and formation of calcium oxalate crystals in isolated leaflets of Gleditsia triacanthos L. Planta. 1986;168:571–578. doi: 10.1007/BF00392278. [DOI] [PubMed] [Google Scholar]
- Foster AS. Plant idioblasts: remarkable examples of cell specialization. Protoplasma. 1956;46:184–193. [Google Scholar]
- Franceschi VR. Oxalic acid metabolism and calcium oxalate formation in Lemna minor L. Plant Cell Environ. 1987;10:397–406. [Google Scholar]
- Franceschi VR. Calcium oxalate formation is a rapid and reversible process in Lemna minor L. Protoplasma. 1989;148:130–139. [Google Scholar]
- Franceschi VR, Horner HT., Jr Use of Psychotria punctata callus in study of calcium oxalate crystal idioblast formation. Z Pflanzenphysiol. 1979;92:61–75. [Google Scholar]
- Franceschi VR, Horner HT., Jr Calcium oxalate crystals in plants. Bot Rev. 1980;46:361–427. [Google Scholar]
- Franceschi VR, Loewus FA. Oxalate biosynthesis and function in plants. In: Khan SR, editor. Calcium Oxalate in Biological Systems. Boca Raton, FL: CRC Press; 1995. pp. 113–130. [Google Scholar]
- Hodgkinson A. Oxalic Acid in Biology and Medicine. London: Academic Press; 1977. [Google Scholar]
- Horner HT, Kausch AP, Wagner BL. Ascorbic acid: a precursor of oxalate in crystal idioblasts of Yucca torreyi in liquid root culture. Intl J Plant Sci. 2000;161:861–868. [Google Scholar]
- Horner HT, Wagner BL. Calcium formation in higher plants. In: Khan SR, editor. Calcium Oxalate in Biological Systems. Boca Raton, FL: CRC Press; 1995. pp. 53–72. [Google Scholar]
- Keates SE, Tarlyn NM, Loewus FA, Franceschi VR. l-Ascorbic acid and l-galactose are sources for oxalic acid and calcium oxalate in Pistia stratiotes. Phytochemistry. 2000;53:433–440. doi: 10.1016/s0031-9422(99)00448-3. [DOI] [PubMed] [Google Scholar]
- Li X, Franceschi VR. Distribution of peroxisomes and glycolate metabolism in relation to calcium oxalate formation in Lemna minor L. Eur J Cell Biol. 1990;51:9–16. [PubMed] [Google Scholar]
- Libert B, Franceschi VR. Oxalate in plant crops. J Agric Food Chem. 1987;35:926–938. [Google Scholar]
- Loewus FA. Biosynthesis and metabolism of ascorbic acid in plants and of analogs of ascorbic acid in fungi. Phytochemistry. 1999;52:193–210. [Google Scholar]
- Smirnoff N, Wheeler GL. Ascorbic acid in plants: biosynthesis and function. CRC Crit Rev Plant Sci. 2000;19:267–290. doi: 10.1080/10409230008984166. [DOI] [PubMed] [Google Scholar]
- Wagner G, Loewus FA. The biosynthesis of (+)-tartaric acid in Pelargonium crispum. Plant Physiol. 1973;52:651–654. doi: 10.1104/pp.52.6.651. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Wagner GJ. Vacuolar deposition of ascorbate-derived oxalic acid in barley. Plant Physiol. 1981;67:591–593. doi: 10.1104/pp.67.3.591. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Wheeler GL, Jones MA, Smirnoff N. The biosynthetic pathway of vitamin C in higher plants. Nature. 1998;393:365–369. doi: 10.1038/30728. [DOI] [PubMed] [Google Scholar]