

Abstract
Preeclampsia (PE) is termed as an obstetric issue that is characterized by hypertension (≧140/90 mm Hg), together with proteinuria following 20 weeks of pregnancy. Until today, PE still constitutes a severe threat to the lives of both the mothers and fetuses. In the past, long noncoding RNAs (lncRNAs) were considered as the transcriptional noise. However, some investigations have indicated that lncRNAs could be used as innovative indicators in PE. The current review aims to discuss the relationship between lncRNAs and PE in recent years. According to the retrieved data, we concluded that lncRNAs can exert an impact on both the occurrence and development of PE through the changes in the biological functions of trophoblasts, immune regulation, epigenetic regulation, decidualization, and energy metabolism. The mechanisms of lncRNAs in PE will help us to better understand the pathogenesis of PE and help us to find targets for predicting and diagnosing PE in the future.
1. Introduction
Preeclampsia (PE) is termed as an obstetric issue that is characterized by hypertension (≧140/90 mm Hg), together with proteinuria following 20 weeks of pregnancy [1]. The incidence of PE is 5-8%, and it may involve the liver, kidney, cardiovascular system, cerebrovascular system, and blood system [2]. Until today, PE still constitutes a severe threat to the lives of both the mothers and fetuses. Females with PE are prone to an augmented threat of developing cardiovascular illnesses in the future [3]. Besides that, it is held that the underlying causes of PE count on the inflammatory elements, in addition to the damaged endothelial function, vasoactive elements, oxidative stress, and genetic causes [4–8]. Nowadays, the “Two-phase Disorder” [9] statement of PE is quite famous. In the preliminary phase of the embryonic implantation, trophoblasts invade the uterine wall, besides helping with uterine artery remodeling. There is an increase in the diameter of blood vessels, together with the decline in the blood flow resistance, aimed at ensuring a sufficient amount of blood flow [10]. Trophoblast cells are quite pivotal for uterine spiral artery remodeling. In the patients, who have PE, the invasive potential of trophoblasts declines, causing the dysplasia of uterine spiral artery remodeling, further leading to a shallow placental implantation [11]. Nowadays, the effective treatment for PE is still regarded as the termination of pregnancy; nevertheless, it does not constitute a reasonable option in case of too few numbers of gestational weeks, coupled with an immature fetus [12]. There have been carried out several investigations primarily emphasizing the physical and biochemical indicators for the forecast, diagnosis, and management [13]. Nevertheless, no exclusive indicator could be put to use as a standard and productive indicator. Accordingly, it is quite essential to investigate the pathogenesis for the purpose of finding a means of preventing and treating PE [14].
Long noncoding RNAs (lncRNAs) regulate the gene expression at the transcriptional level and the posttranscriptional level [15]. In the past, lncRNAs were considered as the transcriptional noise. At present, there are several evidence showing that lncRNAs are associated with both the occurrence and development of numbers of illnesses, for instance, cancer [16], cardiovascular illnesses [17], and neurological illnesses [18]. Besides that, a few investigations have suggested that lncRNAs take part in numerous cell biological procedures that include cell proliferation, together with cell migration, cell apoptosis, X-chromosome inactivation, gene imprinting, and stem cell transformation [19–22]. LncRNAs contribute by means of a number of mechanisms. They function as scaffolds, signals, and antisense decoys, in addition to participating in the transcriptional disturbance. Single lncRNA less often performs multiple functions [23]. Scaffold lncRNAs bring together proteins and other RNAs for becoming a bigger functional compound; for instance, the telomerase RNA compound was required for the telomere repeat complex [24], and the polycomb repressor complex was needed for the histone regulation [25]. The transcription of a few lncRNAs could manifest the silencing of colocated protein-coding genes [26] or function as an indicator of the upstream transcriptional element. Decoy lncRNAs could function through the combing of a target protein, impeding the protein from functioning as it requires [27] or through combing and separating of small regulatory RNAs, for instance, microRNAs (miRNAs) [28]. In comparison with messenger RNAs (mRNAs), the superiority of targeting lncRNAs indicates that it is capable of causing changes in several downstream signal pathways, accordingly carrying out various functions [29]. The demerits associated with being capable of playing a role in multiple downstream pathways are that it may give rise to several unintended damaging effects. That is why, the role of lncRNAs requires carefully understanding before they could be put to use as a therapeutic indicator. In comparison with a normal pregnancy, there were a number of differentially expressed lncRNAs in the placentas of PE patients, which suggested that these lncRNAs might be associated with PE [30]. Besides that, several investigations have demonstrated that lncRNAs could take part in the progress of PE by impacting on the function of trophoblasts. Accordingly, the current review aims to discuss the relationship between lncRNAs and PE in recent years, besides putting efforts for the discovery of the pathogenesis of PE from a new perspective.
2. LncRNA Microarray Study in PE
Nowadays, people have discovered many lncRNAs when compared with protein-coding genes in the genome [31]. Owing to the development of the next-generation sequencing technology, increasing numbers of lncRNAs have been discovered. There was an age-matched lncRNA microarray study. The empirical group comprised 6 patients having the early-onset PE (onset before 34 weeks) and the control group was constituted by 6 patients with a premature delivery. The investigation discovered that there were 15,646 upregulated and 13,178 downregulated lncRNAs in the placenta tissues of early-onset PE patients [30]. For the purpose of exploring the underlying functions of these differentially expressed lncRNAs, GO analysis was performed, which subsequently revealed that the pathways associated with cell migration were markedly enriched [30]. Another microarray investigation highlighted 738 differently expressed lncRNAs in the placentas of PE in comparison with normal pregnancies [32]. LOC391533, LOC284100, and CEACAMP8 were confirmed to be increased in the placentas of PE, besides being discovered linked to both the lipid metabolism and angiogenesis [32]. Moreover, it was also discovered that there were 163 differentially expressed lncRNAs in late-onset PE (onset over 34 weeks) placentas in comparison with the normal placentas [33]. NONHSAT116812 and NONHSAT145880 both might be utilized as indicators for PE since they were also confirmed in plasma specimens [33].
3. LncRNA RNA Sequencing (RNA-Seq) Study in PE
RNA-Seq is a better method than microarray to discover new lncRNAs [34, 35]. Liu S. et al. found some lncRNAs associated with PE, together with discovering that the jak-stat pathway was related to the etiology of PE [36]. Jing Tong et al. performed another RNA-seq study who collected decidual tissues from normal pregnancy (n=3), early-onset PE patients (n=3), and late-onset PE females (n=3), and the results showed that there were 32 aberrant lncRNAs between early-onset PE and normal pregnancy, 53 differentially expressed lncRNAs between late-onset PE and normal pregnancy, and 32 differentially expressed lncRNAs between early-onset PE and late-onset PE, demonstrating that the pathogenesis of early-onset PE was different from that of late-onset PE [37].
4. LncRNAs Affect Cellular Functions of Trophoblast Cells
The deficiency of uterine spiral artery remodeling is the early cause of PE. The capabilities of trophoblast migration and invasion have a close association with epithelial-mesenchymal transition (EMT), which is related to the placental development [38]. The decreased trophoblast proliferation, migration, invasion, and stimulated apoptosis constitute the pivotal reasons leading to PE [39]. LncRNAs are capable of impacting both the occurrence and development of PE through the modification of these functions of trophoblast cells. Some lncRNAs were discovered as increasing in PE (Table 1). The increased RPAIN inhibited the trophoblast invasion, together with inducing the apoptosis through regulating C1q, accordingly promoting the aggravation of PE [40]. Moreover, increased lncRNA uc.187 in the placentas of PE patients promoted the progression of PE not just through the reduction of cell proliferation and invasion but also induced cell apoptosis [41]. The upregulation of lncRNA CCAT1 was carried out in PE, which had the ability to induce the progression of PE by decreasing the level of CDK4 [42]. There was an increase in the expression of lncRNA PRNCR1 in PE as well [43]. As discovered, the mechanism of lncRNA PRNCR1 involved the regulation of the Mitogen-activated protein kinase (MAPK) signaling pathway [43]. There was observed an increase in lncRNA DLX6-AS1 in the placentas of PE patients, leading to PE through the manipulation of the miR-376c/GADD45A expression [44]. The SPRY4-IT1 upregulation substantially decreased cell migration and proliferation, meanwhile still increasing cell apoptosis [45]. LncRNA STOX2-IT3 might decrease the differentiation and invasion of trophoblasts through the regulation of STOX2, followed by making a contribution towards PE [46]. LINC-HELLP was associated with the familial HELLP in Dutch [47]. LINC-HELLP was discovered to be linked to trophoblast proliferation and invasion, and LINC-HELLP mutation could attenuate the differentiation of trophoblasts [48]. LncRNA uc003fir was upexpressed in the vessels of PE placentas in comparison with the normal ones through the regulation of “HIF1-α–lncRNA uc003fir–CCL5” axis, which resulted into the development of PE [49].
Table 1.
Increased lncRNAs related to cellular functions of trophoblast cells in PE.
| LncRNA name | Functions related to PE | Regulating gene/signal pathway | Reference |
|---|---|---|---|
| RPAIN | Inhibits invasion and promotes cell apoptosis | C1q | [40] |
| uc.187 | Inhibits cell proliferation, invasion and increases cell apoptosis | unknown | [41] |
| CCAT1 | Inhibits cell proliferation | CDK4 | [42] |
| PRNCR1 | Reduces cell viability | MAPK signaling pathway | [43] |
| DLX6-AS1 | Decreases cell proliferation, migration and invasion | miR-376c/GADD45A axis | [44] |
| SPRY4-IT1 | Decreases cell migration and proliferation, increases cell apoptosis | unknown | [45] |
| STOX2-IT3 | Decreases cell invasion | STOX2 | [46] |
| HELLP | Reduces cell differentiation, proliferation and invasion | YBX1, PCBP1, PCBP2, RPS6 and RPL7 | [47, 48] |
| uc003fir | Reduces cell proliferation and migration | HIF1-α–lncRNA uc003fir–CCL5 | [49] |
| CEACAMP8 | unknown | unknown | [32] |
| LOC391533 | unknown | unknown | [32] |
| LOC284100 | unknown | unknown | [32] |
MAPK, Mitogen-activated protein kinase.
In contrast, the downregulation of some lncRNAs was also confirmed in PE (Table 2). LncRNA MALAT-1 was substantially reduced in the placentas of PE patients, inducing cell proliferation, meanwhile decreasing the cell cycle and cell apoptosis [50]. In addition to that, lncRNA‐ATB was manifested to be associated with the incidence of PE by means of the manipulation of the procedure of the cell invasion, coupled with the development of the endothelial vessels [51]. The decline in small nucleolar RNA host gene 5 (SNHG5) in the placentas of PE women indicated that SNHG5 induced not only cell proliferation, but also cell invasion, and migration through the manipulation of the miR‐26a‐5p/N‐cadherin axis [52]. The expression of lncRNA maternally expressed gene 3 (MEG3) in PE placentas was decreased as well [53]. The downregulation of lncRNA MEG3 had the potential of inhibiting cell migration, inducing cell apoptosis, and increasing the expressions of NF-κB, Caspase 3, and Bax, further causing the dysplasia of uterine spiral artery [53]. LncRNA TUG1 was decreased in the placental tissues of PE females in comparison with the levels in normal ones. Moreover, the downregulation of TUG1 resulted into the inhibitory functions, which attenuated cell proliferation, migration, and invasion together with increasing the apoptosis in trophoblast cells, and further having a role in spiral artery remodeling by means of increasing Ezh2 and decreasing RND3 level in PE [54]. LncRNA RP11-465L10.10 was decreased in the placentas of females in early-onset PE by targeting MMP9 [30]. In addition, functional investigations revealed that HOXA11-AS was capable of increasing Ezh2 and lysine-specific demethylase 1 (LSD1) and regulating RND3 level in the nucleus, while in the cytoplasm, HOXA11-AS regulated the HOXA7 level via sponged miR-15b-5p, which changed cell proliferation [55]. H19 was decreased in the early-onset severe PE through the regulation of miR-675/Nodal Modulator 1 (NOMO) axis and Nodal signal pathway, and H19 was capable of increasing cell proliferation [56]. The decreased level of EGFR Antisense RNA1 (EGFR-AS1) in PE had the potential to decrease the level of EGFR in trophoblast cells and the phosphorylation levels of downstream proteins in JAK/STAT signal pathway, followed by inducing the occurrence of PE [57]. The level of linc00473 was decreased in the placental tissues of severe PE women, linc00473 downregulation in trophoblast cells substantially reduced cell proliferation, together with increasing cell apoptosis, while linc00473 upregulation increased cell proliferation through the attenuation of tissue factor pathway inhibitor 2 (TFPI2) by combing to LSD1 [58]. LncRNA PVT1 downregulation substantially decreased cell proliferation; besides inducing the cell cycle as well as cell apoptosis, the mechanism of PVT1 was through the regulation of angiopoietin-like 4 (ANGPTL4) as well as Ezh2 [59]. The downregulation of MVIH in PE decreased not only cell proliferation, but also cell migration, invasion and angiogenesis in trophoblast cell lines by targeting Jun-B protein [60].
Table 2.
Decreased lncRNAs related to cellular functions of trophoblast cells in PE.
| LncRNA name | Functions related to PE | Regulating gene/signal pathway | Reference |
|---|---|---|---|
| MALAT-1 | Increases proliferation, decreases apoptosis | unknown | [50] |
| ATB | Increases migration, proliferation, tube-formation of HTR-8/SVneo cells |
unknown | [51] |
| SNHG5 | Promotes cell proliferation, invasion and migration | miR‐26a‐5p/N‐cadherin axis | [52] |
| MEG3 | Reduces apoptosis and promotes migration | unknown | [53] |
| TUG1 | Increases proliferation, migration, and invasion and reduces apoptosis | Ezh2, RND3 | [54] |
| RP11-465L10.10 | Increases cell migration and motility | MMP9 | [30] |
| HOXA11-AS | Increases cell proliferation and migration | Ezh2, Lsd1, RND3 miR-15b-5p/HOXA7 axis |
[55] |
| H19 | Increases cell proliferation | miR-675/NOMO1 Nodal signaling | [56] |
| EGFR-AS1 | Increases cell proliferation | JAK/STAT signaling pathway | [57] |
| linc00473 | Increases cell proliferation and reduces cell apoptosis | TFPI2, LSD1 | [58] |
| PVT1 |
Increases cell proliferation and reduces cell apoptosis | ANGPTL4, EZH2 |
[59] |
| MVIH | Promotes cell growth, migration, invasion and angiogenesis | Jun-B | [60] |
SNHG5, small nucleolar RNA host gene 5; MEG3, maternally expressed gene 3; NOMO1, Nodal Modulator 1; EGFR-AS1, EGFR Antisense RNA1; TFPI2, tissue factor pathway inhibitor 2; LSD1, lysine-specific demethylase 1; ANGPTL4, angiopoietin-like 4.
5. LncRNAs Affect Immune Response in PE
The expression level of dendritic cells (DCs) for the production of T regulatory (Treg) cells in PE patients substantially declined [61]. Lnc-DC just appeared in DCs, and functional experiments have indicated that the downregulation of lnc-DC damaged the differentiation of monocytes into DCs, accordingly performing the attenuation of the production of Treg cells by DCs. Lnc-DC could increase the phosphorylation of tyrosine-705 in the cytoplasm, followed by impacting the transcription of downstream genes and stimulating the differentiation as well as the development of DCs [62]. In addition, lnc-DCs stimulated the development of decidual DCs in PE females, besides contributing to a rise in Th1 cells, leading to the etiology of PE [63]. MALAT1-upregulated mesenchymal stem cells (MSCs) induced M2 macrophage polarization and the function was regulated by MALAT1-produced IDO level, which suggested that MALAT1 might induce the immunosuppressive potential of MSCs in vivo [64].
6. LncRNAs Affect PE by Epigenetic Regulation
LncRNAs had the potential of mediating the gene expression at the epigenetic level [65]. Moreover, the alternative epigenetic mediation of the H19-IGF2 domain in placental tissues was associated with PE, resulting into placental dysplasia in the early pregnancy [66]. LncRNA H19 rs217727 polymorphism was related to a higher chance of having PE [67]. Moreover, not only the CTC, but also TTC and TTT haplotypes had an association with the PE susceptibility [67]. STOX2-IT3-lncRA performs the role of a permissive cis-acting regulatory factor of STOX2 selective splicing [46].
7. LncRNAs Affect Decidualization and Energy Metabolism in PE
There have been numerous investigations suggesting that PE has an association with the dysplasia of decidualization [68]. Moreover, the poor decidualization can make a contribution to the decreased invasive potential of extravillous trophoblasts, dysplasia of the uterine spiral artery, and decreasing the blood flow at the maternal-fetal interface [69]. Placental ischemia results into the increased expression of toxic cytokines in the maternal peripheral blood, further impairing endothelial cells. Glycolysis is considered as quite pivotal for the development of endothelial cells, and decreased glycolysis was capable of contributing to the impaired decidualization [70]. The decreased level of HK2P1 and HK2 might be associated with the occurrence as well as the development of PE by means of the impairment of glycolysis and decidualization [71]. HK2P1 mediated the expression of HK2 by functioning as a competing endogenous RNA (ceRNA) [71]. These findings put forward innovative ideas regarding the etiology of PE, and the new regulating axis, which suggested HK2P1, HK2, and miR-6887-3p, might be put to use as the innovative indicators for PE [71]. Both PGK1 and PGK1P2 constitute a couple of ceRNAs related to miR-330-5p, in addition to being quite important in the mechanism of decidualization by means of the mediation of the angiogenesis and glycolysis [72]. Decreased expressions of PGK1 as well as PGK1P2 in the decidual tissues might give rise to the impaired decidualization and the occurrence of PE [72]. At present, increasing numbers of investigations have suggested that the occurrence of PE is associated with the abnormal energy metabolism. The expression of lncZBTB39-1:2 in placenta has the potential of decreasing the trophoblast activity by means of impacting the energy regulation, which may promote the progress of PE [73].
8. Conclusion
In the current review, we have discussed about the latest research progress of lncRNAs and PE. According to the retrieved data, we concluded that lncRNAs can exert an impact on both the occurrence and development of PE through the changes in the biological functions of trophoblasts, immune regulation, epigenetic regulation, decidualization, and energy metabolism (Figure 1). Even though the functions of many lncRNAs are not clear, the mechanisms of lncRNAs in PE will help us to better understand the pathogenesis of PE and help us to find targets for predicting and diagnosing PE in the future.
Figure 1.
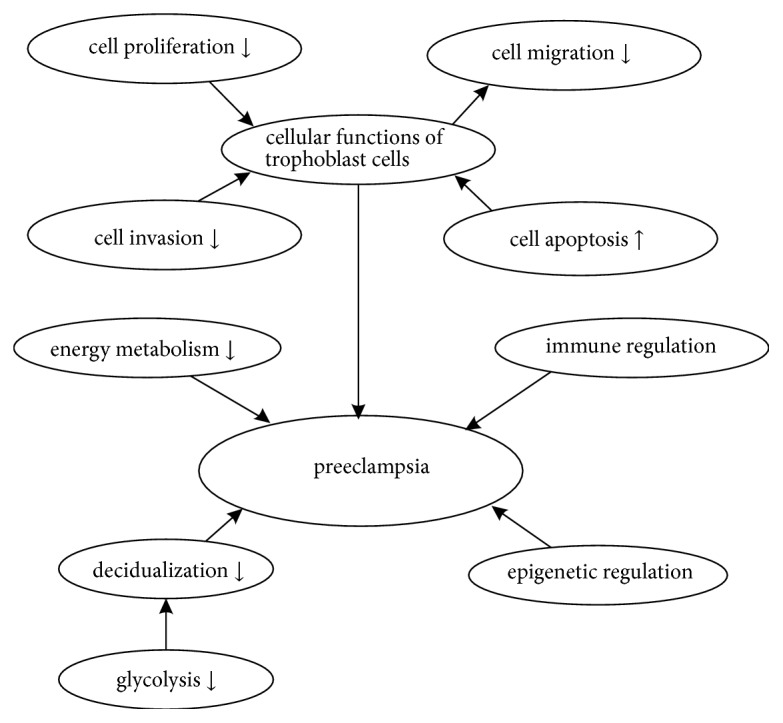
The biological functions of lncRNAs in PE. LncRNAs can exert an impact on both the occurrence and development of PE through the changes in the biological functions of trophoblasts (including cell proliferation, migration, invasion, and apoptosis), immune regulation, epigenetic regulation, decidualization, and energy metabolism, and impaired glycolysis may lead to the decreased decidualization.
Conflicts of Interest
The authors declare that they have no conflicts of interest.
References
- 1.Jim B., Karumanchi S. A. Preeclampsia: pathogenesis, prevention, and long-term complications. Seminars in Nephrology. 2017;37(4):386–397. doi: 10.1016/j.semnephrol.2017.05.011. [DOI] [PubMed] [Google Scholar]
- 2.Duley L. The global impact of pre-eclampsia and eclampsia. Seminars in Perinatology. 2009;33(3):130–137. doi: 10.1053/j.semperi.2009.02.010. [DOI] [PubMed] [Google Scholar]
- 3.Ahmed R., Dunford J., Mehran R., et al. Pre-eclampsia and future cardiovascular risk among women: a review. Journal of the American College of Cardiology. 2014;63(18):1815–1822. doi: 10.1016/j.jacc.2014.02.529. [DOI] [PubMed] [Google Scholar]
- 4.Saftlas A. F., Beydoun H., Triche E. Immunogenetic determinants of preeclampsia and related pregnancy disorders: a systematic review. Obstetrics & Gynecology. 2005;106(1):162–172. doi: 10.1097/01.aog.0000167389.97019.37. [DOI] [PubMed] [Google Scholar]
- 5.Granger J. P., Alexander B. T., Llinas M. T., Bennett W. A., Khalil R. A. Pathophysiology of hypertension during preeclampsia linking placental ischemia with endothelial dysfunction. Hypertension. 2001;38(3):718–722. doi: 10.1161/01.HYP.38.3.718. [DOI] [PubMed] [Google Scholar]
- 6.Rana S., Karumanchi S. A., Levine R. J., et al. Sequential changes in antiangiogenic factors in early pregnancy and risk of developing preeclampsia. Hypertension. 2007;50(1):137–142. doi: 10.1161/HYPERTENSIONAHA.107.087700. [DOI] [PubMed] [Google Scholar]
- 7.Noris M., Todeschini M., Cassis P., et al. L-arginine depletion in preeclampsia orients nitric oxide synthase toward oxidant species. Hypertension. 2004;43(3):614–622. doi: 10.1161/01.hyp.0000116220.39793.c9. [DOI] [PubMed] [Google Scholar]
- 8.Bartha J. L., Romero-Carmona R., Torrejon-Cardoso R., Comino-Delgado R. Insulin, insulin-like growth factor-1, and insulin resistance in women with pregnancy-induced hypertension. American Journal of Obstetrics & Gynecology. 2002;187(3):735–740. doi: 10.1067/mob.2002.126283. [DOI] [PubMed] [Google Scholar]
- 9.Ngene NC., Moodley J. Role of angiogenic factors in the pathogenesis and management of pre-eclampsia. International Journal of Gynecology & Obstetrics. 2002;141(1):5–13. doi: 10.1002/ijgo.12424. [DOI] [PubMed] [Google Scholar]
- 10.Duncan J. R., Tobiasz A. M., Bursac Z., Rios-Doria E. V., Schenone M. H., Mari G. Uterine artery flow velocity waveforms before and after delivery in hypertensive disorders of pregnancy near term. Hypertension in Pregnancy. 2018;37(3):131–136. doi: 10.1080/10641955.2018.1493495. [DOI] [PubMed] [Google Scholar]
- 11.Louwen F., Muschol-Steinmetz C., Reinhard J., Reitter A., Yuan J. A lesson for cancer research: Placental microarray gene analysis in preeclampsia. Oncotarget . 2012;3(8):759–773. doi: 10.18632/oncotarget.595. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 12.Esteve-Valverde E., Ferrer-Oliveras R., Gil-Aliberas N., Baraldès-Farré A., Llurba E., Alijotas-Reig J. Pravastatin for preventing and treating preeclampsia: a systematic review. Obstetrical & Gynecological Survey. 2018;73(1):40–55. doi: 10.1097/OGX.0000000000000522. [DOI] [PubMed] [Google Scholar]
- 13.Park H., Shim S., Cha D., et al. Combined screening for early detection of pre-eclampsia. International Journal of Molecular Sciences. 2015;16(8):17952–17974. doi: 10.3390/ijms160817952. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 14.Bastek J. A., Elovitz M. A. The role and challenges of biomarkers in spontaneous preterm birth and preeclampsia. Fertility and Sterility. 2013;99(4):1117–1123. doi: 10.1016/j.fertnstert.2013.01.104. [DOI] [PubMed] [Google Scholar]
- 15.Klinge C. M. Non-coding RNAs: Long non-coding RNAs and microRNAs in endocrine-related cancers. Endocrine-Related Cancer. 2018;25(4):R259–R282. doi: 10.1530/ERC-17-0548. [DOI] [PubMed] [Google Scholar]
- 16.Wang J., Liu X., Wu H., et al. CREB up-regulates long non-coding RNA, HULC expression through interaction with microRNA-372 in liver cancer. Nucleic Acids Research. 2010;38(16):5366–5383. doi: 10.1093/nar/gkq285. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 17.Leung A., Trac C., Jin W., et al. Novel long noncoding RNAs are regulated by angiotensin II in vascular smooth muscle cells. Circulation Research. 2013;113(3):266–278. doi: 10.1161/CIRCRESAHA.112.300849. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 18.Johnson R. Long non-coding RNAs in Huntington's disease neurodegeneration. Neurobiology of Disease. 2012;46(2):245–254. doi: 10.1016/j.nbd.2011.12.006. [DOI] [PubMed] [Google Scholar]
- 19.Eißmann M., Gutschner T., Hämmerle M., et al. Loss of the abundant nuclear non-coding RNA MALAT1 is compatible with life and development. RNA Biology. 2012;9(8):1076–1087. doi: 10.4161/rna.21089. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 20.Brown C. J., Hendrich B. D., Rupert J. L., et al. The human XIST gene: analysis of a 17 kb inactive X-specific RNA that contains conserved repeats and is highly localized within the nucleus. Cell. 1992;71(3):527–542. doi: 10.1016/0092-8674(92)90520-m. [DOI] [PubMed] [Google Scholar]
- 21.Koerner M. V., Pauler F. M., Huang R., Barlow D. P. The function of non-coding RNAs in genomic imprinting. Development. 2009;136(11):1771–1783. doi: 10.1242/dev.030403. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 22.Kim K., Jutooru I., Chadalapaka G., et al. HOTAIR is a negative prognostic factor and exhibits pro-oncogenic activity in pancreatic cancer. Oncogene. 2013;32(13):1616–1625. doi: 10.1038/onc.2012.193. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 23.Geisler S., Coller J. RNA in unexpected places: long non-coding RNA functions in diverse cellular contexts. Nature Reviews Molecular Cell Biology. 2013;14(11):699–712. doi: 10.1038/nrm3679. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 24.Collins K. Physiological assembly and activity of human telomerase complexes. Mechanisms of Ageing and Development. 2008;129(1-2):91–98. doi: 10.1016/j.mad.2007.10.008. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 25.Rinn J. L., Kertesz M., Wang J. K., et al. Functional demarcation of active and silent chromatin domains in human HOX loci by noncoding RNAs. Cell. 2007;129(7):1311–1323. doi: 10.1016/j.cell.2007.05.022. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 26.Mohammad F., Mondal T., Kanduri C. Epigenetics of imprinted long noncoding RNAs. Epigenetics. 2009;4(5):277–286. doi: 10.4161/epi.4.5.9242. [DOI] [PubMed] [Google Scholar]
- 27.Hung T., Wang Y., Lin M. F., et al. Extensive and coordinated transcription of noncoding RNAs within cell-cycle promoters. Nature Genetics. 2011;43(7):621–629. doi: 10.1038/ng.848. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 28.Bayoumi A., Sayed A., Broskova Z., et al. Crosstalk between long noncoding RNAs and MicroRNAs in health and disease. International Journal of Molecular Sciences. 2016;17(3):p. 356. doi: 10.3390/ijms17030356. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 29.Balgkouranidou I., Matthaios D., Karayiannakis A., et al. Prognostic role of APC and RASSF1A promoter methylation status in cell free circulating DNA of operable gastric cancer patients. Mutation Research - Fundamental and Molecular Mechanisms of Mutagenesis. 2015;778:46–51. doi: 10.1016/j.mrfmmm.2015.05.002. [DOI] [PubMed] [Google Scholar]
- 30.Long W., Rui C., Song X., et al. Distinct expression profiles of lncRNAs between early-onset preeclampsia and preterm controls. Clinica Chimica Acta. 2016;463:193–199. doi: 10.1016/j.cca.2016.10.036. [DOI] [PubMed] [Google Scholar]
- 31.Djebali S., Davis C. A., Merkel A. Landscape of transcription in human cells. Nature. 2012;489(7414):101–108. doi: 10.1038/nature11233. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 32.He X., He Y., Xi B., et al. LncRNAs expression in preeclampsia placenta reveals the potential role of LncRNAs contributing to preeclampsia pathogenesis. PLoS ONE. 2013;8(11) doi: 10.1371/journal.pone.0081437. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 33.Wang X., Chen Y., Du L., Li X., Li X., Chen D. Evaluation of circulating placenta-related long noncoding RNAs as potential biomarkers for preeclampsia. Experimental and Therapeutic Medicine. 2018;15(5):4309–4317. doi: 10.3892/etm.2018.5968. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 34.Griffith M., Griffith O. L., Mwenifumbo J., et al. Alternative expression analysis by RNA sequencing. Nature Methods. 2010;7(10):843–847. doi: 10.1038/nmeth.1503. [DOI] [PubMed] [Google Scholar]
- 35.Trapnell C., Williams B. A., Pertea G., et al. Transcript assembly and quantification by RNA-Seq reveals unannotated transcripts and isoform switching during cell differentiation. Nature Biotechnology. 2010;28(5):511–515. doi: 10.1038/nbt.1621. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 36.Liu S., Xie X., Lei H., Zou B., Xie L. Identification of key circRNAs/lncRNAs/miRNAs/mRNAs and pathways in preeclampsia using bioinformatics analysis. Medical Science Monitor. 2019;25:1679–1693. doi: 10.12659/MSM.912801. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 37.Tong J., Zhao W., Lv H., Li W.-P., Chen Z.-J., Zhang C. Transcriptomic profiling in human decidua of severe preeclampsia detected by RNA sequencing. Journal of Cellular Biochemistry. 2018;119(1):607–615. doi: 10.1002/jcb.26221. [DOI] [PubMed] [Google Scholar]
- 38.Kokkinos M. I., Murthi P., Wafai R., Thompson E. W., Newgreen D. F. Cadherins in the human placenta - epithelial-mesenchymal transition (EMT) and placental development. Placenta. 2010;31(9):747–755. doi: 10.1016/j.placenta.2010.06.017. [DOI] [PubMed] [Google Scholar]
- 39.Fu G., Ye G., Nadeem L., et al. MicroRNA-376c impairs transforming growth factor-β and nodal signaling to promote trophoblast cell proliferation and invasion. Hypertension. 2013;61(4):864–872. doi: 10.1161/HYPERTENSIONAHA.111.203489. [DOI] [PubMed] [Google Scholar]
- 40.Song X., Rui C., Meng L., et al. Long non-coding RNA RPAIN regulates the invasion and apoptosis of trophoblast cell lines via complement protein C1q. Oncotarget . 2017;8(5):7637–7646. doi: 10.18632/oncotarget.13826. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 41.Cao C., Li J., Li J., Liu L., Cheng X., Jia R. Long non-coding RNA Uc.187 is upregulated in preeclampsia and modulates proliferation, apoptosis, and invasion of HTR-8/SVneo trophoblast cells. Journal of Cellular Biochemistry. 2017;118(6):1462–1470. doi: 10.1002/jcb.25805. [DOI] [PubMed] [Google Scholar]
- 42.Li J. L., Li R., Gao Y., et al. LncRNA CCAT1 promotes the progression of preeclampsia by regulating CDK4. European Review for Medical and Pharmacological Sciences. 2018;22(5):1216–1223. doi: 10.26355/eurrev_201803_14461. [DOI] [PubMed] [Google Scholar]
- 43.Jiao S., Wang S. Y., Huang Y. LncRNA PRNCR1 promoted the progression of eclampsia by regulating the MAPK signal pathway. European Review for Medical and Pharmacological Sciences. 2018;22(12):3635–3642. doi: 10.26355/eurrev_201806_15240. [DOI] [PubMed] [Google Scholar]
- 44.Tan Y., Xiao D., Xu Y., Wang C. Long non-coding RNA DLX6-AS1 is upregulated in preeclampsia and modulates migration and invasion of trophoblasts through the miR-376c/GADD45A axis. Experimental Cell Research. 2018;370(2):718–724. doi: 10.1016/j.yexcr.2018.07.039. [DOI] [PubMed] [Google Scholar]
- 45.Zou Y., Jiang Z., Yu X., et al. Upregulation of long noncoding RNA SPRY4-IT1 modulates proliferation, migration, apoptosis, and network formation in trophoblast cells HTR-8SV/neo. PLoS ONE. 2013;8(11):p. e79598. doi: 10.1371/journal.pone.0079598. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 46.Oudejans C. B., Poutsma A., Michel O. J., et al. Noncoding RNA-regulated gain-of-function of STOX2 in Finnish pre-eclamptic families. Scientific Reports. 2016;6(1):p. 32129. doi: 10.1038/srep32129. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 47.van Dijk M., Thulluru H. K., Mulders J., et al. HELLP babies link a novel lincRNA to the trophoblast cell cycle. The Journal of Clinical Investigation. 2012;122(11):4003–4011. doi: 10.1172/jci65171. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 48.van Dijk M., Visser A., Buabeng K. M. L., Poutsma A., van der Schors R. C., Oudejans C. B. M. Mutations within the LINC-HELLP non-coding RNA differentially bind ribosomal and RNA splicing complexes and negatively affect trophoblast differentiation. Human Molecular Genetics. 2015;24(19):5475–5485. doi: 10.1093/hmg/ddv274. [DOI] [PubMed] [Google Scholar]
- 49.Pengjie Z., Xionghui C., Yueming Z., et al. LncRNA uc003fir promotes CCL5 expression and negatively affects proliferation and migration of trophoblast cells in preeclampsia. Pregnancy Hypertension: An International Journal of Women's Cardiovascular Health. 2018;14:90–96. doi: 10.1016/j.preghy.2018.08.449. [DOI] [PubMed] [Google Scholar]
- 50.Chen H., Xu X., Teng J., et al. CXCR4 inhibitor attenuates allergen-induced lung inflammation by down-regulating MMP-9 and ERK1/2. International Journal of Clinical and Experimental Pathology. 2015;8(6):6700–6707. [PMC free article] [PubMed] [Google Scholar]
- 51.Liu X., Chen H., Kong W., et al. Down-regulated long non-coding RNA-ATB in preeclampsia and its effect on suppressing migration, proliferation, and tube formation of trophoblast cells. Placenta. 2017;49:80–87. doi: 10.1016/j.placenta.2016.12.004. [DOI] [PubMed] [Google Scholar]
- 52.Yang Y., Xi L., Ma Y., et al. The lncRNA small nucleolar RNA host gene 5 regulates trophoblast cell proliferation, invasion, and migration via modulating miR-26a-5p/N-cadherin axis. Journal of Cellular Biochemistry. 2019;120(3):3173–3184. doi: 10.1002/jcb.27583. [DOI] [PubMed] [Google Scholar]
- 53.Zhang Y., Zou Y., Wang W., et al. Down-regulated long non-coding RNA MEG3 and its effect on promoting apoptosis and suppressing migration of trophoblast cells. Journal of Cellular Biochemistry. 2015;116(4):542–550. doi: 10.1002/jcb.25004. [DOI] [PubMed] [Google Scholar]
- 54.Xu Y., Ge Z., Zhang E., et al. The lncRNA TUG1 modulates proliferation in trophoblast cells via epigenetic suppression of RND3. Cell Death & Disease. 2017;8(10):p. e3104. doi: 10.1038/cddis.2017.503. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 55.Xu Y., Wu D., Liu J., et al. Downregulated lncRNA HOXA11-AS affects trophoblast cell proliferation and migration by regulating RND3 and HOXA7 expression in PE. Molecular Therapy - Nucleic Acids. 2018;12:195–206. doi: 10.1016/j.omtn.2018.05.007. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 56.Gao W.-L., Liu M., Yang Y., et al. The imprinted H19 gene regulates human placental trophoblast cell proliferation via encoding miR-675 that targets nodal modulator 1 (NOMO1) RNA Biology. 2012;9(7):1002–1010. doi: 10.4161/rna.20807. [DOI] [PubMed] [Google Scholar]
- 57.Zhao Z. M., Jiang J. Lowly expressed EGFR-AS1 promotes the progression of preeclampsia by inhibiting the EGFR-JAK/STAT signaling pathway. European Review for Medical and Pharmacological Sciences. 2018;22(19):6190–6197. doi: 10.26355/eurrev_201810_16024. [DOI] [PubMed] [Google Scholar]
- 58.Wu D., Xu Y., Zou Y., et al. Long noncoding RNA 00473 is involved in preeclampsia by LSD1 binding-regulated TFPI2 transcription in trophoblast cells. Molecular Therapy - Nucleic Acids. 2018;12:381–392. doi: 10.1016/j.omtn.2018.05.020. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 59.Xu Y., Lian Y., Zhang Y., et al. The long non-coding RNA PVT1 represses ANGPTL4 transcription through binding with EZH2 in trophoblast cell. Journal of Cellular and Molecular Medicine. 2018;22(2):1272–1282. doi: 10.1111/jcmm.13405. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 60.Zou Y., Li Q., Xu Y., et al. Promotion of trophoblast invasion by lncRNA MVIH through inducing Jun-B. Journal of Cellular and Molecular Medicine. 2018;22(2):1214–1223. doi: 10.1111/jcmm.13400. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 61.Hsu P., Santner-Nanan B., Dahlstrom J. E., et al. Altered decidual DC-SIGN+ antigen-presenting cells and impaired regulatory T-cell induction in preeclampsia. The American Journal of Pathology. 2012;181(6):2149–2160. doi: 10.1016/j.ajpath.2012.08.032. [DOI] [PubMed] [Google Scholar]
- 62.Wang P., Xue Y., Han Y., et al. The STAT3-binding long noncoding RNA Inc-DC controls human dendritic cell differentiation. Science. 2014;344(6181):310–313. doi: 10.1126/science.1251456. [DOI] [PubMed] [Google Scholar]
- 63.Zhang W., Zhou Y., Ding Y. Lnc-DC mediates the over-maturation of decidual dendritic cells and induces the increase in Th1 cells in preeclampsia. American Journal of Reproductive Immunology. 2017;77(6):p. e12647. doi: 10.1111/aji.12647. [DOI] [PubMed] [Google Scholar]
- 64.Li X., Song Y., Liu F., et al. Long non-coding RNA MALAT1 promotes proliferation, angiogenesis, and immunosuppressive properties of mesenchymal stem cells by inducing VEGF and IDO. Journal of Cellular Biochemistry. 2017;118(9):2780–2791. doi: 10.1002/jcb.25927. [DOI] [PubMed] [Google Scholar]
- 65.Mercer T. R., Dinger M. E., Mattick J. S. Long non-coding RNAs: insights into functions. Nature Reviews Genetics. 2009;10(3):155–159. doi: 10.1038/nrg2521. [DOI] [PubMed] [Google Scholar]
- 66.Gao W.-L., Li D., Xiao Z.-X., et al. Detection of global DNA methylation and paternally imprinted H19 gene methylation in preeclamptic placentas. Hypertension Research. 2011;34(5):655–661. doi: 10.1038/hr.2011.9. [DOI] [PubMed] [Google Scholar]
- 67.Harati-Sadegh M., Kohan L., Teimoori B., Salimi S. The long non-coding RNA H19 rs217727 polymorphism is associated with PE susceptibility. Journal of Cellular Biochemistry. 2018;119(7):5473–5480. doi: 10.1002/jcb.26708. [DOI] [PubMed] [Google Scholar]
- 68.Lockwood C. J., Oner C., Uz Y. H., et al. Matrix metalloproteinase 9 (MMP9) expression in preeclamptic decidua and MMP9 induction by tumor necrosis factor alpha and interleukin 1 beta in human first trimester decidual cells. Biology of Reproduction. 2008;78(6):1064–1072. doi: 10.1095/biolreprod.107.063743. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 69.Lash G. E., Ernerudh J. Decidual cytokines and pregnancy complications: focus on spontaneous miscarriage. Journal of Reproductive Immunology. 2015;108:83–89. doi: 10.1016/j.jri.2015.02.003. [DOI] [PubMed] [Google Scholar]
- 70.Tsai J., Chi M. M., Schulte M. B., Moley K. H. The fatty acid beta-oxidation pathway is important for decidualization of endometrial stromal cells in both humans and mice. Biology of Reproduction. 2014;90(2) doi: 10.1095/biolreprod.113.113217. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 71.Lv H., Tong J., Yang J., et al. Dysregulated pseudogene HK2P1 may contribute to preeclampsia as a competing endogenous rna for hexokinase 2 by impairing decidualization. Hypertension. 2018;71(4):648–658. doi: 10.1161/HYPERTENSIONAHA.117.10084. [DOI] [PubMed] [Google Scholar]
- 72.Tong J., Yang J., Lv H., Lv S., Zhang C., Chen Z. Dysfunction of pseudogene PGK1P2 is involved in preeclampsia by acting as a competing endogenous RNA of PGK1. Pregnancy Hypertension: An International Journal of Women's Cardiovascular Health. 2018;13:37–45. doi: 10.1016/j.preghy.2018.05.003. [DOI] [PubMed] [Google Scholar]
- 73.Liu Y., Han TL., Luo X. The metabolic role of LncZBTB39-1:2 in the trophoblast mobility of preeclampsia. Genes and Diseases. 2018;5(3):235–244. doi: 10.1016/j.gendis.2018.04.005. [DOI] [PMC free article] [PubMed] [Google Scholar]