

Abstract
Autism Spectrum Disorder (ASD) is the most prevalent neurodevelopmental disorder in the United States and often co-presents with sleep problems. Sleep problems in ASD predict the severity of ASD core diagnostic symptoms and have a considerable impact on the quality of life of caregivers. Little is known, however, about the underlying molecular mechanisms of sleep problems in ASD. We investigated the role of Shank3, a high confidence ASD gene candidate, in sleep architecture and regulation. We show that mice lacking exon 21 of Shank3 have problems falling asleep even when sleepy. Using RNA-seq we show that sleep deprivation increases the differences in prefrontal cortex gene expression between mutants and wild types, downregulating circadian transcription factors Per3, Bhlhe41, Hlf, Tef, and Nr1d1. Shank3 mutants also have trouble regulating wheel-running activity in constant darkness. Overall, our study shows that Shank3 is an important modulator of sleep and clock gene expression.
Research organism: Mouse
eLife digest
Autism spectrum disorder (ASD) is the most common neurodevelopmental disorder in the United States. People with ASD tend to have difficulties with communication and social interactions, restricted interests, and may repeat certain behaviors. They also often struggle to fall or stay asleep. Sleep deprivation may exacerbate other symptoms of the disorder. This makes life more difficult for both the person with ASD and their caregivers. Scientists do not yet know what causes sleep difficulties in people with ASD.
Unraveling the complex genetics that underlie ASD may help scientists better understand ASD-related sleep difficulties. One possible genetic culprit for sleep difficulties in ASD is a gene called SHANK3. Patients with an ASD-associated condition called Phelan-McDermid syndrome are often missing the SHANK3 gene. They also often have sleep problems.
Now, Ingiosi, Schoch et al. show that both patients with Phelan-McDermid syndrome and mice with a mutation in the Shank3 gene have problems falling asleep. Using a registry that collects genetic and sleep information on people with Phelan-McDermid syndrome, Ingiosi, Schoch et al. found that people who are missing SHANK3 frequently have trouble falling asleep and wake up many times each night. Mice missing part of the Shank3 gene also had difficulty falling asleep, even after they have been deprived of sleep.
Mice naturally have a daily pattern of sleep and activity. This 24-hour activity cycle is maintained by an internal circadian clock. In mice missing part of Shank3, the circadian clock genes are not turned on correctly. These genes were less active in mice missing Shank3, and this difference worsened with lack of sleep. These mice also ran less on a wheel than typical mice when kept in total darkness, even though the pattern of activity did not change. The experiments suggest that Shank3 controls sleep, likely through its effects on circadian clock genes. Learning more about what causes these sleep problems may help scientists develop ways to improve sleep in people with ASD and Phelan-McDermid syndrome.
Introduction
Autism Spectrum Disorder (ASD) is the most prevalent neurodevelopmental disorder in the United States (diagnosed in 1 in 59 children [Baio et al., 2018]). The core symptoms of ASD include social and communication deficits, restricted interests, and repetitive behaviors (American Psychiatric Association, 2013). In addition, several studies show that individuals with ASD report a variety of co-morbid conditions including sleep problems and altered circadian rhythms (Glickman, 2010). It is estimated that 40–80% of the ASD population experience sleep disorders that do not improve with age (Johnson et al., 2009). More specifically, people with ASD have problems falling and staying asleep (Hodge et al., 2014). A recent study showed that sleep problems co-occur with autistic traits in early childhood and increase over time, suggesting that sleep problems are an essential part of ASD (Verhoeff et al., 2018). Indeed, sleep impairments are a strong predictor of the severity of ASD core symptoms as well as aggression and behavioral issues (Cohen et al., 2014; Tudor et al., 2012). Although a great number of studies documented sleep problems in ASD, little is known about the underlying molecular mechanisms.
To better understand the mechanisms underlying sleep problems in ASD, we need animal models that closely recapitulate sleep phenotypes observed in clinical populations. The study of genetic animal models of ASD, in which a genetic abnormality that is known to be associated with ASD is introduced, has provided valuable insight into the molecular mechanisms underlying ASD (de la Torre-Ubieta et al., 2016). These models include Fragile X syndrome, 16p11.2 deletion syndrome, cortical dysplasia-focal epilepsy (CDFE) syndrome, and mutations in neuroligins, neurexins, and shank genes among others. However, sleep research in animal models of ASD is limited and has not yet revealed the underlying mechanisms of sleep issues associated with ASD. Studies using a fly model of Fragile X syndrome reported an increase in sleep which is in contrast to what is observed in the clinical population (Bushey et al., 2009). The opposite phenotype was reported in a Fragile X mouse model, displaying instead an age-dependent reduction in activity during the light phase (i.e the mouse subjective night) (Boone et al., 2018). Neuroligin 1 knockout mice spend more time asleep (El Helou et al., 2013), but mice with mutations in Neuroligin 2 spend less time asleep and more time awake (Seok et al., 2018). Mice with a missense mutation in Neuroligin 3 show normal sleep behavior (Liu et al., 2017), but rats with a deletion mutation in Neuroligin 3 spend less time in non-rapid eye movement (NREM) sleep than wild type rats (Thomas et al., 2017). Mutant rat models of CDFE syndrome show longer waking periods while the mutant mice show fragmented wakefulness (Thomas et al., 2017). Mice carrying a 16p11.2 deletion syndrome sleep less than wild type mice, but only males are affected (Angelakos et al., 2017). More importantly, issues with sleep onset, the most prominent feature of sleep problems in ASD patients, have not been evaluated in animal models of ASD.
In this study, we examined sleep in Phelan-McDermid syndrome (PMS) patients with SHANK3 mutations and in a mutant mouse with a deletion in Shank3 exon 21 (Shank3ΔC). PMS is a syndromic form of ASD characterized by gene deletions affecting the human chromosomal region 22q13.3 (Phelan and McDermid, 2012), particularly the neuronal structural gene SHANK3. Individuals with PMS have high rates of intellectual disability, difficulties in communication and motor function, and approximately 84% fit the core diagnostic criteria for ASD (Soorya et al., 2013). There is also a high rate of sleep problems in PMS (Bro et al., 2017). Mice with mutations in Shank3 recapitulate multiple features of both ASD and PMS (Bozdagi et al., 2010; Dhamne et al., 2017; Jaramillo et al., 2017; Jaramillo et al., 2016; Kouser et al., 2013; Peça et al., 2011; Speed et al., 2015), including cognitive impairment, deficits in social behavior, and impaired motor coordination. We show that PMS patients have trouble falling and staying asleep similar to what is observed in the general ASD population. We also show that Shank3ΔC mice sleep less than wild type mice when sleep pressure is high, have reduced sleep intensity (using an accepted electroencephalographic (EEG) metric), and have delayed sleep onset following sleep deprivation. To identify molecular mechanisms underlying sleep changes in Shank3ΔC mice, we carried out genome-wide gene expression studies. We previously showed that genome-wide gene expression analysis is a valuable approach to understand the molecular mechanisms underlying the detrimental effects of sleep deprivation (Gerstner et al., 2016; Vecsey et al., 2012). In this study, we found that sleep deprivation sharply increases the differences in gene expression between Shank3ΔC mutants and wild type mice, downregulating circadian transcription factors Per3, Bhlhe41 (Dec2), Hef, Tlf, and Nr1d1 (Rev-erbα). We also show that Shank3ΔC mice are unable to sustain wheel-running activity in constant darkness. Overall, these studies demonstrate that Shank3 is an important modulator of sleep that may exert its effect through the regulation of circadian transcription factors. Our findings may lead to a deeper understanding of the molecular mechanisms underlying sleep problems in ASD. This may one day lead to the development of successful treatments or interventions for this debilitating comorbidity.
Results
Phelan-McDermid syndrome patients have problems falling and staying asleep
Recent studies suggest that sleep problems may be present in a substantial number of PMS patients and may be an important factor for caregivers' well-being (Bro et al., 2017). We obtained genetic and sleep questionnaire data from the Phelan-McDermid Syndrome International Registry (PMSIR) to estimate the frequency and age of onset of sleep problems in PMS individuals carrying a SHANK3 deletion. In parallel, we surveyed the clinical literature to estimate the prevalence of sleep problems in ASD (Andersen et al., 2008; Cotton and Richdale, 2006; Gail Williams et al., 2004; Giannotti et al., 2008; Giannotti et al., 2006; Krakowiak et al., 2008; Liu et al., 2006; Miano et al., 2007; Paavonen et al., 2008; Polimeni et al., 2005; Richdale and Prior, 1995; Tani et al., 2003; Thirumalai et al., 2002; Wiggs and Stores, 2004) and typically developing populations (Anders and Eiben, 1997; Baweja et al., 2013; Bixler et al., 2009; Hysing et al., 2013; Leger et al., 2012; Loessl et al., 2008; Lozoff et al., 1985; Lumeng and Chervin, 2008; Ohayon et al., 2000; Pallesen et al., 2008; Patzold et al., 1998; Sadeh et al., 2000). Figure 1 shows that PMS patients have trouble falling asleep and experience multiple night awakenings starting at about 5 years of age. Those difficulties translate to reduced time asleep particularly during adolescence. Although total sleep time seems to improve in adulthood for PMS patients, that improvement is accompanied by an increase in parasomnias. Problems falling and staying asleep persist regardless of age. The frequency of problems falling and staying asleep in PMS patients is similar to what is observed in the general ASD population and much higher than in typically developing individuals (Figure 1—source data 1).
Figure 1. Increased incidence of sleep problems reported in individuals with Phelan-McDermid syndrome (PMS) compared to typically developing (TD) individuals.

Dashed line indicates median incidence observed in TD (white marker) and ASD (black marker) populations (values from Figure 1—source data 1).
Shank3ΔC mice sleep less during the dark phase and show reduced sleep intensity
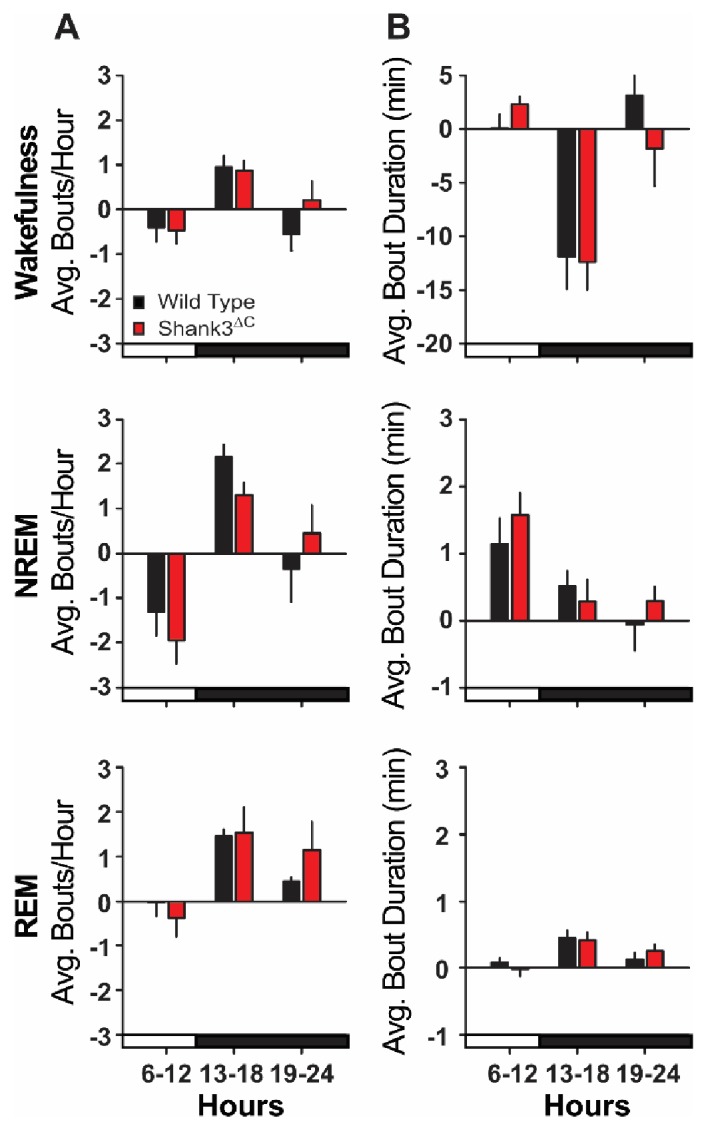
To determine if Shank3ΔC mice have deficits in spontaneous sleep, undisturbed baseline EEG and electromyographic (EMG) recordings were obtained from wild type (WT) and Shank3ΔC mice. There was a significant period x genotype interaction for total sleep time (i.e. total time spent in NREM and rapid eye movement (REM) sleep) during the light (hours 1–12) and dark (hours 13–24) periods (F(1,28) = 5.198, p=0.036). Posthoc pairwise comparisons using Sidak correction showed that Shank3ΔC mice slept less than WT mice during the dark period (p=0.045; Table 1). To determine in which arousal states and hours this effect was most pronounced, we examined hourly time in state data for wakefulness, NREM sleep, and REM sleep. Repeated measures ANOVA over the full 24 hr recording period found significant time x genotype interactions for wakefulness (F(23,345) = 2.419, p<0.0001), NREM sleep (F(23,345) = 2.357, p=0.001), and REM sleep (F(23,345) = 2.175, p=0.002) (Figure 2A). Posthoc comparisons found the most pronounced differences at hour 19 when Shank3ΔC mice spent more time in wakefulness (p<0.0001) and less time NREM (p<0.0001) and REM (p=0.040) sleep (Figure 2A) marking the beginning of an overall trend for increased wakefulness during the last half of the dark period. Analysis of the sleep architecture found time x genotype interactions for bout number during REM sleep (F(3,51) = 2.832, p=0.047) and bout duration during NREM (F(1.749,29.734) = 4.077, p=0.032) and REM (F(1.897,32.243) = 6.536, p=0.005) sleep. Posthoc comparisons showed these effects were driven by differences in the last half of the dark period (hours 19–24) when Shank3ΔC mice had fewer REM bouts (p=0.003; Figure 2—figure supplement 1A) and shorter NREM (p=0.005) and REM (p=0.013) bouts (Figure 2—figure supplement 1B) than WT mice. Overall, these data show that Shank3ΔC mice spend more time awake at the end of the dark phase compared to WT mice under baseline conditions.
Table 1. Shank3ΔC mice sleep less than wild type mice during the dark period.
Repeated measures ANOVA with posthoc pairwise comparisons using Sidak correction. Values are means ± SEM for wild type (n = 10) and Shank3ΔC (n = 10). *p<0.05, difference from wild type.
| Total Sleep Time (%) | Wild Type | Shank3ΔC | p-value |
|---|---|---|---|
| Light Period | 62.90 ± 1.57 | 62.96 ± 1.65 | 0.980 |
| Dark Period | 28.72 ± 2.41 | 21.16 ± 2.54 | 0.045* |
Figure 2. Shank3∆C mice sleep less during the dark period and show altered EEG spectral power under baseline conditions.
The rows represent the vigilance states of wakefulness (top), NREM sleep (middle), and REM sleep (bottom). (A) Time spent in wakefulness and sleep shown as percentage of recording time per hour. Values are means ± SEM. Repeated measures ANOVA with posthoc pairwise comparisons using Sidak correction; *p<0.05, difference from wild type. (B) EEG spectral power normalized as a percentage of the total state-specific EEG power for the light period (left) and dark period (right) fit to smooth curves (solid lines) and expressed with 95% confidence intervals (gray and red shading). Non-overlap of the 95% confidence interval denotes statistically significant differences. The open bars on the x-axis denote the light period and the filled bars denote the dark period of the light:dark cycle. Wild type (n = 10) and Shank3∆C (n = 10) mice.

Figure 2—figure supplement 1. Baseline sleep bout analysis.

Fourier analysis of the EEG indicates that Shank3ΔC mice also sleep differently than WT mice (Figure 2B). Significant genotypic differences in spectral frequencies occur at points of non-overlaping 95% confidence intervals. During NREM sleep, power in the delta frequencies (0.5–4 Hz) was blunted and alpha frequencies (10–15 Hz) were enhanced for Shank3ΔC mice relative to WT during the light and dark periods (Figure 2B). NREM delta power is a measure of synchrony of the neural network and can be a measure of sleep intensity or depth (Achermann and Borbely, 2017). These data indicate that Shank3ΔC mice exhibit disrupted neural connectivity during NREM sleep which may suggest that Shank3ΔC mice sleep less deeply under baseline conditions.
Shank3ΔC mice have abnormal homeostatic responses to sleep deprivation
To investigate sleep homeostasis in Shank3ΔC mice we sleep deprived mutant and WT mice for 5 hr via gentle handling starting at light onset (hour 1). Sleep deprivation (SD) was effective resulting in WT and Shank3ΔC mice spending 97.37 ± 0.76% and 95.08 ± 1.08% time in wakefulness, respectively, during the 5 hr SD period (t(18) = 1.732, p=0.100). Both WT and Shank3ΔC mice showed homeostatic responses to SD (Figure 3—figure supplement 1) with reduced wakefulness and increased time spent in NREM and REM sleep during the recovery phase (Figure 3—source data 2). Sleep was also more consolidated after SD for both WT and Shank3ΔC mice during the light period, and changes in sleep architecture did not statistically differ between WT and Shank3ΔC mice (Figure 3—figure supplement 2). However, there were time x genotype interactions for wakefulness (F(18,288) = 2.025, p=0.009), NREM sleep (F(18,288) = 1.928, p=0.014), and REM sleep (F(18,288) = 1.716, p=0.036) over the SD recovery phase (hours 6–24). This effect was most pronounced near the end of the dark period where Shank3ΔC mice spent more time in wakefulness and less time in NREM and REM sleep compared to WT (Figure 3—figure supplement 1C) which is consistent with the trend seen in the baseline data (Figure 2A). Changes in NREM EEG delta power were similar between WT and Shank3ΔC mice, indicating that mutant mice accumulate sleep pressure similarly to WT mice (Figure 3A). However, Shank3ΔC mice showed a transient enhancement of NREM EEG spectral power (F(1,18) = 12.07, p=0.003) compared to WT mice in the first 2 hr post-SD (hours 6–7). This enhancement was found in the higher frequencies (3.9–19.5 Hz; Figure 3B) and resolved to baseline and WT values during hours 11–12 (F(1,18) = 0.59, p=0.454; Figure 3C). These data suggest that some aspects of the Shank3ΔC homeostatic response to sleep deprivation are abnormal.
Figure 3. Shank3∆C mice take longer to fall asleep and sleep less after sleep deprivation.
(A) Normalized NREM delta (δ; 0.5–4 Hz) power during recovery sleep post-sleep deprivation. The cross-hatched bar on the x-axis denotes the 5 hr sleep deprivation period and the open bar denotes the remaining light period. Repeated measures ANOVA. (B) Normalized NREM spectra for the first 2 hr post-sleep deprivation (hours 6–7; significant from 3.9 to 19.5 Hz). One-way ANOVA with posthoc unpaired t-tests using Benjamini-Hochberg correction. (C) Normalized NREM spectra for the last 2 hr of the light period post-sleep deprivation (hours 11–12). One-way ANOVA. (D) Latency to enter NREM sleep and REM sleep after sleep deprivation. Unpaired t-test. (E) Time in NREM sleep and REM sleep for the first 2 hr post-sleep deprivation (hours 6–7). Unpaired t-test. (F) Time in NREM sleep and REM sleep for the remaining 5 hr of light period post-sleep deprivation (hours 8–12). Unpaired t-test. Values are means ± SEM for wild type (n = 10; black) and Shank3∆C (n = 10; red) mice. *p<0.05, difference from wild type.

Figure 3—figure supplement 1. Shank3∆C mice sleep less than wild type type after sleep deprivation.

Figure 3—figure supplement 2. Bout analysis after sleep deprivation.
Remarkably, Shank3ΔC mice took longer to enter NREM sleep post-SD compared to WT (t(10.44) = −2.31, p=0.043; Figure 3D). As a consequence, Shank3ΔC mice spent less time in NREM sleep compared to WT during the first 2 hr of the recovery phase (t(13.26) = 2.85, p=0.014; Figure 3E). During the subsequent 5 hr of the light period (hours 8–12), Shank3ΔC mice spent more time in NREM sleep compared to WT (t(18) = −2.46; p=0.024; Figure 3F). Overall, our data show that Shank3ΔC mice have difficulties falling asleep despite heightened sleep pressure.
Shank3ΔC mice show downregulation of circadian transcription factors in response to sleep deprivation
We conducted a genome-wide gene expression study to investigate the molecular basis for the Shank3ΔC mouse sleep phenotype. Shank3ΔC and WT adult male mice were subjected to 5 hr of SD starting at light onset and sacrificed immediately post-SD. Additional mice from both genotypes were sacrificed at the same time of day to determine differences in gene expression under homecage conditions (HC). Prefrontal cortex was collected for all animals and subjected to RNA sequencing (RNA-seq, n = 5 per group). As expected, sleep deprivation is the greatest source of variation in the data (principal component 1), followed by the genotype effect (principal component 2; Figure 4A). Furthermore, the difference between genotypes is enhanced after sleep deprivation (Figure 4A) greatly increasing the number of differentially expressed genes between Shank3ΔC and WT mice – starting at 69 genes (HC) and doubling to 134 genes (SD) (Figure 4B, Figure 4—source data 1; false discovery rate (FDR) < 0.1). Most of the differences in gene expression following SD are not present in HC conditions.
Figure 4. Sleep deprivation induces a two-fold difference in gene expression between Shank3ΔC and wild type mice.
RNA-seq study of gene expression from prefrontal cortex obtained from adult male Shank3ΔC and wild type mice either under control homecage conditions (HC) or following 5 hr of sleep deprivation (SD). N = 5 mice per group. (A) Principal component analysis of normalized RNA-seq data shows that sleep deprivation is the main source of variance in the data (first principal component, PC1) and genotype is the second (second principal component, PC2). Percent variance explained by each PC is shown on each axis. (B) Venn diagram showing the number of genes differentially expressed at FDR < 0.1 between Shank3ΔC and wild type mice in either control HC conditions or after SD. (C) Heat map of average scaled gene expression for all genes in (B). K-means clustering defined three clusters based on differences in gene expression across all comparisons. Genes belonging to the MAPK pathway and involved in circadian rhythms (see Table 2) are highlighted on the right.

Figure 4—figure supplement 1. Recovery of positive control genes regulated by sleep deprivation.

Clustering of gene expression patterns for all genes differentially expressed between Shank3ΔC and WT mice reveals 3 groups of genes (Figure 4C). Cluster 1 contains genes that are downregulated in mutants versus WT mice under HC conditions. Genes in this cluster are also downregulated in response to SD in Shank3ΔC mice but are generally unaffected by SD in WT mice. SD seems to exacerbate the difference between genotypes, which explains why differential expression of some of the genes in cluster 1 only reach statistical significance after SD. This cluster contains the majority of the genes that are differentially expressed between genotypes. Cluster 2 contains genes that are upregulated in mutants versus WT mice under HC conditions that are also downregulated by SD in WT mice. Cluster 3 contains genes that are normally upregulated by SD. For both clusters 2 and 3, the Shank3ΔC mutation seems to dampen the response to SD.
To better understand the impact of the gene expression at the pathway level, we carried out functional annotation of the transcripts differentially expressed between Shank3ΔC and WT mice (Table 2, Table 2—source data 1). Our results reveal that 1) MAPK/GnRH signaling and circadian rhythm-associated transcripts are downregulated in Shank3ΔC mice and 2) sleep deprivation exacerbates that difference. Circadian transcription factors are particularly affected. For example, while expression of Per3, Hlf, and Tef already differs under homecage conditions, SD leads to additional genotype-specific differences in expression of Nr1d1 and Bhlhe41. All of the above mentioned circadian transcription factors belong to cluster 1 on our heat map (Figure 4C) showing downregulation in response to SD only in Shank3ΔC mice.
Table 2. Results of functional bioinformatics analysis of the genes differentially expressed between Shank3ΔCmice relative to wild type mice.
Functional annotation and clustering analysis was performed using DAVID (https://david.ncifcrf.gov) and functional information was obtained from the following databases: GO (Biological process and Molecular function), KEGG pathways, and Uniprot keywords. Enrichment was performed relative to all transcripts expressed in the mouse prefrontal cortex as defined by our RNA-seq data. Enriched functional terms were clustered at low stringency, to obtain clusters with enrichment score >1.2 (corresponding to an average p-value>0.05). See Table 2—source data 1 for details.
| Shank3ΔC vs. Wild Type | |
|---|---|
| Homecage | Sleep Deprivation |
| Up-regulated | Up-regulated |
| Cholesterol Metabolism: Hmgcr, Insig1 | Potassium Ion Transport: Kcnv1, Kcnk1, Kcnk2 |
| Transcription: Jun, Fosl2, Nfil3, Stat4 | Dephosphorylation: Dusp10, Dusp3, Ptprj |
| Neuron Projection Development: Cntn1, Ntrk2, Reln, Sema3a | |
| Down-regulated | Down-regulated |
| MAPK Signaling: Mapk3 (ERK1), Elk1 | GnRH Signaling: Mapk1 (ERK2), Elk1, Mapk11 (p38) |
| Circadian Rhythms: Per3, Tef, Hlf | Circadian Rhythms: Per3, Nr1d1 (REV-ERBα), Tef, Hlf, Prkab2 (AMPK), Bhlhe41 (DEC2) |
| Transcription: Tef, Hlf | Sodium Ion Transport: Slc6a15, Slc22a4, Slc24a4 |
Wheel-running activity in Shank3ΔC mice is impaired in constant darkness
The results of our RNA-seq analysis revealed a general downregulation of circadian transcription factors in the mutants. These findings suggested that Shank3ΔC mice might also have abnormal circadian rhythms. To test this possibility, we measured wheel-running activity in Shank3ΔC and WT mice. Wheel-running data were collected for 2 weeks under LD (LD weeks 2–3), then mice were released into constant darkness (DD) for 3 weeks (DD weeks 1–3; Figure 5 and Figure 5—figure supplement 1). Alpha, the length of the active phase, was calculated as the time between activity onset and offset. Period length was the time elapsed from the start of an active phase to the start of the subsequent active phase. There was no differences between Shank3ΔC and WT mice for alpha or period length during the DD period (Table 3), but Shank3ΔC mice ran fewer revolutions than WT mice from LD week 3 – DD week 3 (F(1,338) = 30.96; p<0.0001; Table 3, Table 3—source data 1). Some Shank3ΔC mice greatly reduced their running at certain periods of the DD period, which is reflected in the variance for period, alpha, and activity measures (Figure 5, Figure 5—figure supplement 1, and Table 3). Over 5 weeks of continuous LD conditions, however, wheel-running activity for Shank3ΔC mice instead increased (F(1,542) = 85.99, p<0.0001; Supplementary file 1, Source data 1). Overall, these data indicate that constant darkness impairs wheel-running activity of Shank3ΔC mice.
Figure 5. Shank3∆C mice show disruption of running wheel activity in constant darkness.
Representative actograms and periodograms for two wild type and two Shank3∆C mice. Mice were entrained to a 12:12 hr light:dark cycle (LD, 559 ± 4 : 0 ± 0 lux) for two weeks prior to 3 weeks constant darkness (DD, 0 ± 0 lux). Gray shading is representative of the dark period. (A) Actograms for two wild type mice. (B) Corresponding periodograms for wild type mice. (C) Actograms for two Shank3∆C mice. (D) Corresponding periodograms for Shank3∆C mice.

Figure 5—figure supplement 1. Actograms for all wild type and Shank3∆C mice.

Table 3. Shank3∆C mice show reduced wheel-running activity during constant darkness.
Summary of wheel-running behavior measured for each week of 12:12 hr light dark cycle (LD, 559 ± 4 : 0 ± 0 lux) or constant darkness (DD, 0 ± 0 lux) for both wild type and Shank3∆C mice. A linear mixed-effects model was used to estimate the contributions of genotype and the interaction between genotype and time for period, alpha, and wheel running activity for LD week three through DD week 3. There is a significant interaction effect for genotype x time for activity (p < 0.0001; indicated by *) but no significant effect for genotype x time for period (p = 0.15) or genotype x time for alpha (p = 0.88). See Table 3—source data 1 for details. Values are means ± SEM for wild type (n = 8) and Shank3∆C (n = 7) mice. Significance at p < 0.05.
| Wild Type | Shank3∆C | ||
|---|---|---|---|
| Period (h) | |||
| LD Week 2 | 24.0 ± 0.0 | 24.0 ± 0.0 | |
| LD Week 3 | 24.0 ± 0.0 | 23.9 ± 0.1 | |
| DD Week 1 | 23.9 ± 0.1 | 23.7 ± 0.1 | |
| DD Week 2 | 23.7 ± 0.1 | 20.3 ± 3.4 | |
| DD Week 3 | 23.5 ± 0.2 | 25.8 ± 1.5 | |
| Alpha (h) | |||
| LD Week 2 | 10.0 ± 0.4 | 10.7 ± 0.4 | |
| LD Week 3 | 10.3 ± 0.4 | 10.7 ± 0.5 | |
| DD Week 1 | 10.5 ± 0.2 | 9.0 ± 0.4 | |
| DD Week 2 | 10.4 ± 0.4 | 8.2 ± 0.7 | |
| DD Week 3 | 9.0 ± 1.0 | 9.9 ± 1.0 | |
| Activity (rev/day)* | |||
| LD Week 2 | 33985.2 ± 565.9 | 23258.2 ± 861.2 | |
| LD Week 3 | 40306.7 ± 1257.1 | 25365.1 ± 964.0 | |
| DD Week 1 | 37942.6 ± 795.4 | 23027.7 ± 669.6 | |
| DD Week 2 | 38250.7 ± 431.1 | 16820.7 ± 1302.5 | |
| DD Week 3 | 38586.3 ± 231.1 | 6961.5 ± 383.9 |
Discussion
The present study is the first to establish a role for Shank3 in mammalian sleep. We show that both PMS patients and Shank3 mutant mice have trouble falling asleep (Figures 1, 2 and 3). This phenotype is widely observed in the ASD population and until now had not been replicated in animal models. Shank3∆C mice have problems falling asleep after periods of extended wakefulness when sleep pressure is high, such as at the end of the baseline dark period (Figure 2A) or following sleep deprivation (Figure 3D-E). They also have blunted NREM delta power (Figure 2B). However, Shank3∆C mice accumulate sleep pressure (Figure 3A) and show no gross abnormalities in circadian rhythms (Figure 5). This suggests that the primary deficit is in sleep onset.
Our molecular studies show that sleep deprivation increases differences in gene expression between Shank3∆C and WT mice (Figure 4). These differences point to the downregulation of circadian transcription factors and genes involved in the MAPK/GnRH pathways in the mutants (Table 2). Circadian transcription factors affected include Per3, Hlf, Tef, Nr1d1 (REV-ERBα), and Bhlhe41 (DEC2). We therefore investigated the effects of our Shank3 mutation in circadian rhythms. Shank3∆C mice do not show a disruption in circadian rhythmicity, but they do exhibit a large reduction in wheel-running activity in response to constant darkness (Figure 5, Table 3). Shank3∆C mice were reported to have deficits in motor coordination (Kouser et al., 2013); however, under a 12:12 hr LD schedule their wheel-running activity increases over time (Supplementary file 1, Source data 1). Daily rhythms of activity in rodents are linked to circadian oscillations in dopamine release (Feenstra et al., 2000; Menon et al., 2019) in the frontal cortex as well as in the striatum, a motor region with high levels of Shank3 expression (Peça et al., 2011). Together with a reduction in circadian gene expression, this DD-specific activity deficit suggests that the mutant sleep phenotype involves clock gene functions outside of their central time-keeping role. Interestingly, mutations in some of the circadian transcription factors we identified, Per3 and Bhlhe41 (DEC2), indeed lead to deficits in sleep regulation (Archer et al., 2018; He et al., 2009; Hirano et al., 2018). Our results support a role of clock genes in influencing sleep outside of their roles in generating circadian rhythms (Franken, 2013).
An interesting question is how can deletion of exon 21 of Shank3 lead to dysregulation of transcriptional and signaling pathways linked to sleep and sleep loss? Exon 21 of Shank3 encodes the homer and cortactin interaction domains of the protein. Homer interacts with metabotropic glutamate receptors (mGluRs) and SHANK3/homer complexes anchor mGluRs to the synapse. Shank3∆C mice show a marked reduction of the major isoforms of SHANK3 as well as an increase of mGluRs at the synapse (Kouser et al., 2013). mGluR signaling activates the MAPK/ERK pathway, a key regulator of activity-dependent transcription and synaptic plasticity in mature neurons (Thomas and Huganir, 2004). So the role of SHANK3 at the synaptic membrane explains the observed regulation of MAPK pathway genes (Table 2). However, it is not yet clear how SHANK3 regulates expression of circadian transcription factors within the nucleus. One possibility might include the role of SHANK3 in Wnt signaling. SHANK3 modulates Wnt-mediated transcriptional regulation by regulating internalization of Wnt receptor Frizzled-2 (Harris et al., 2016) and nuclear translocation of the Wnt ligand beta-catenin (Qin et al., 2018). The Wnt pathway kinase GSK3β phosphorylates circadian transcription factors PER2 (Iitaka et al., 2005), CRY2 (Harada et al., 2005), and REV-ERBα (Yin et al., 2006); however, this mechanism modulates circadian period length which is not altered in Shank3ΔC mice. A second potential mechanism is nuclear translocation of SHANK3 itself. SHANK3 is known to undergo synaptic-nuclear shuttling in response to neuronal activity and interact with nuclear ribonucleoproteins and components of the RNA Pol II mediator complex (Grabrucker et al., 2014). Deletion of the C-terminus leads to nuclear accumulation of SHANK3 and alterations in gene expression (Cochoy et al., 2015; Grabrucker et al., 2014). Thus, mutations in exon 21 of Shank3 could lead to deficits in transcriptional regulation in response to sleep deprivation through direct regulation of transcription in the nucleus. Yeast two-hybrid data show that SHANK3 can directly bind the circadian transcription factors REV-ERBα (encoded by Nr1d1) and DEC1 (encoded by Bhlhe40), a close paralog of DEC2 (Bhlhe41) (Sakai et al., 2011). Future studies of the effects of sleep on SHANK3 nuclear translocation will provide new insights into the non-synaptic function of shank proteins and their role in sleep and circadian rhythms.
Materials and methods
Key resources table.
| Reagent type (species) or resource |
Designation | Source or reference |
Identifiers |
|---|---|---|---|
| Gene (Mus musculus) | Shank3; SH3 and multiple ankyrin repeat domains 3 | NA | ENTREZ_ID: 58234 |
| Gene (Mus musculus) | Per3; period circadian clock 3 | NA | ENTREZ_ID: 18628 |
| Gene (Mus musculus) | Hlf; hepatic leukemia factor | NA | ENTREZ_ID: 217082 |
| Gene (Mus musculus) | Tef; thyrotroph embryonic factor | NA | ENTREZ_ID: 21685 |
| Gene (Mus musculus) | Nr1d1; nuclear receptor subfamily 1, group D, member 1; | NA | ENTREZ_ID: 217166 |
| Gene (Mus musculus) | Bhlhe41; basic helix-loop-helix family, member e41; | NA | ENTREZ_ID: 79362 |
| Strain, strain background (Mus musculus) | Shank3ΔC; Shank3tm1.1Pfw/J | Kouser et al., 2013; The Jackson Laboratory | RRID:IMSR_JAX:018398 |
| Commercial assay or kit | RNAeasy kit | Qiagen | 74104 |
| Commercial assay or kit | High Sensitivity RNA Analysis Kit | Advanced Analytical Technologies | DNF-472 |
| Commercial assay or kit | High Sensitivity NGS Fragment Analysis Kit | Advanced Analytical Technologies | DNF-474 |
| Commercial assay or kit | KAPA Library Quantification Kit | Kapabiosystems | KK4824 |
| Commercial assay or kit | TruSeq Stranded mRNA Library Prep Kit | Illumina | RS-122–2101 |
| Software, algorithm | HiSeq Control Software version (v. 2.2.68) | Illumina | 15073358 Rev A |
| Software, algorithm | bcl2fastq (v. 2.17.1.14) | R/Bioconductor | SCR_015058 |
| Software, algorithm | Rsubread (v. 1.26.1) | R/Bioconductor | SCR_016945 |
| Software, algorithm | NOISeq (v.2.22.1) | R/Bioconductor | SCR_003002 |
| Software, algorithm | RUVSeq (v. 1.12.0) | R/Bioconductor | SCR_006263 |
| Software, algorithm | edgeR (v. 3.20.9) | R/Bioconductor | SCR_012802 |
| Software, algorithm | R package nlme (v. 3.5.0) | CRAN | SCR_015655 |
| Software, algorithm | R package mgcf (v. 3.5.0) | CRAN | |
| Software, algorithm | R package pheatmap (v. 1.0.10) | CRAN | SCR_016418 |
| Software, algorithm | Database for annotation, visualization, and integrated discovery (v 6.7) | DAVID; https://david.ncifcrf.gov | |
| Software, algorithm | VitalRecorder | Kissei Comtec | 3, 0, 0, 0 |
| Software, algorithm | SleepSign for Animal | Kissei Comtec | 3, 0, 0, 812 |
| Software, algorithm | Wheel manager | Med Associates Inc | SOF-860 |
| Software, algorithm | ClockLab (v. 3.5.0) | Actimetrics | 6.0.50 |
Sleep questionnaire study
Sleep questionnaire data for PMS patients was obtained through a data access agreement with the PMSIR. The PMSIR contains demographic, clinical, and genetic data on PMS patients. Parents and caregivers are able to create a profile on behalf of the patient, enter demographic data, complete questionnaires on symptoms and development, and upload files such as genetic test reports and other medical records. Trained genetic counselors curate all genetic data to ensure each patient has as complete and accurate ‘genetic profile’ insofar as possible. Researchers may apply for data exports containing de-identified clinical data, developmental data, and genetic data. Once approved, the appropriate search is performed in the database and the results are provided to the researcher. Data in this study corresponds to a database export performed on 12/1/2016 containing the results of a sleep questionnaire completed by caregivers, as well as biographic and genetic information of PMS individuals. Difficulty falling asleep is defined as needing more than an hour to fall asleep. Multiple night awakenings is defined as more than two awakenings. Reduced sleep time is defined as less than 6 hr per night. Parasomnias are defined as abnormal movements, behaviors, emotions, perceptions, and dreams that occur while falling asleep or sleeping. Presence of sleep apnea is defined as having received a diagnosis of sleep apnea. Only individuals with a genetic counselor-confirmed deletion of the SHANK3 gene were included in the analysis. The final dataset included 176 individuals – 78 males and 98 females (age range 1–39 years).
Animals
Heterozygous Shank3+/∆C mice were obtained from Dr. Paul Worley at Johns Hopkins University. Shank3+/∆C breeding pairs were established to obtain wild type (WT) and Shank3∆C littermates. Mice were housed in standard cages at 24 ± 1°C on a 12:12 hr light:dark cycle (unless otherwise specified) with food and water ad libitum. All experimental procedures were approved by the Institutional Animal Care and Use Committee of Washington State University and conducted in accordance with National Research Council guidelines and regulations for experiments in live animals.
Assessment of sleep-wake behavior
Surgical procedures
Adult male mice (12-weeks-old) were stereotaxically implanted with electroencephalographic (EEG) and electromyographic (EMG) electrodes under isoflurane anesthesia according to previously published methods (Frank et al., 2002). Briefly, four stainless steel screws (BC-002MPU188, Bellcan International Corp, Hialeah, FL) were placed contralaterally over frontal (2) and parietal (2) cortices, and 2 EMG electrodes were inserted in the nuchal muscles. Mice were allowed 5 days of recovery from surgery prior to habituation to the recording environment.
Experimental design
Six days after surgery, wild type (WT; n = 10) and Shank3∆C (n = 10) littermates were connected to a lightweight, flexible tether and allowed 5 days to habituate to the tether and recording environment. After habituation, mice underwent 24 hr undisturbed baseline EEG and EMG recording starting at light onset (hour 1). The next day, mice were sleep deprived for 5 hr (hours 1–5) via gentle handling beginning at light onset according to previously published methods (Halassa et al., 2009; Vecsey et al., 2012; Ingiosi et al., 2015). Mice were then allowed 19 hr undisturbed recovery sleep (hours 6–12 of the light period and hours 13–24 of the dark period).
EEG/EMG data acquisition and analysis
EEG and EMG data were collected with Grass 7 polygraph hardware (Natus Medical Incorporated, Pleasanton, CA) via a light-weight, counterbalanced cable, amplified, and digitized at 200 Hz using VitalRecorder acquisition software (SleepSign for Animal, Kissei Comtec Co., LTD, Nagano, Japan). EEG and EMG data were high- and low-pass filtered at 0.3 and 100 Hz and 10 and 100 Hz, respectively.
Wakefulness and sleep states were determined by visual inspection of the EEG waveform, EMG activity, and fast Fourier transform (FFT) analysis by an experimenter blinded to the genotype. Data were scored as wakefulness, NREM sleep, or REM sleep with 4 s resolution using SleepSign for Animal as previously described (Halassa et al., 2009).
The EEG was subjected to fast Fourier transform analysis to produce power spectra between 0–20 Hz with 0.781 Hz resolution. Delta (δ) was defined as 0.5–4 Hz, theta (θ) as 5–9 Hz, and alpha (α) as 10–15 Hz. For genotypic comparisons of light and dark period spectral data, each spectral bin was expressed as a percentage of the sum of total state-specific EEG power (0–20 Hz) of the baseline 12 hr light period and 12 hr dark period, respectively (Baracchi and Opp, 2008). The relation between EEG spectral power and frequency under baseline conditions was fit to smooth curves and 95% confidence intervals for the respective light and dark periods. The analysis was performed via Generalized Additive Models (GAM) (Hastie and Tibshirani, 1990) using the R package mgcf (v. 3.5.0). For hourly NREM delta power analysis after sleep deprivation, spectral values within the delta band for each hour were normalized to the average NREM delta band value of the last 4 hr of the baseline light period (hours 9–12) and expressed as a percentage (Franken et al., 2001). Changes in individual NREM EEG spectral bins (0–20 Hz) after sleep deprivation were normalized to corresponding baseline spectral bins obtained from mean values from the last 4 hr of the baseline light period (hours 9–12). EEG epochs containing artifacts were excluded from spectral analyses.
Statistical analyses were performed using SPSS for Windows (IBM Corporation, Armonk, NY) and R statistical language. Data are presented as means ± standard error of the mean (SEM). If an individual hour bin did not contain a full hour of data due to technical issues (e.g. removal of recording tether), that hour was excluded from analysis for the corresponding individual animal. A general linear model for repeated measures (RM) using time (hours or period) as the repeated measure and genotype (WT vs. Shank3∆C) or treatment (undisturbed baseline vs. sleep deprivation) as the between subjects factor was used when multiple measurements were made over time (i.e. total sleep time, time in stage, bout number, bout duration, hourly NREM delta power). Repeated measures were tested for sphericity, and a Greenhouse-Geisser correction was applied when appropriate. Posthoc pairwise comparisons using Sidak corrections were performed when there were significant interaction effects or main effects of genotype or treatment. Baseline time in state and bout data RM comparisons were made over all time intervals during the full 24 hr recording period (hours 1–24). For sleep deprivation experiments, time in state and bout data RM comparisons were made over all time intervals during the full recovery period (hours 6–24). Hourly NREM delta power RM comparisons post-sleep deprivation were made over hours 6–12 of the recovery phase. Comparisons of normalized NREM EEG spectral power changes after sleep deprivation were made using one-way ANOVA with total spectral power as the dependent variable and genotype as the between subjects factor. If the one-way ANOVA yielded a significant result, unpaired Student’s t-tests with Benjamini-Hochberg correction were then used for comparisons of individual NREM EEG spectral frequency bins with genotype as the grouping variable. Unpaired Student’s t-tests with genotype as the grouping variable were used to analyze latency to state and total time in state post-sleep deprivation. Latency to NREM or REM sleep after sleep deprivation was defined as time elapsed from release to recovery sleep to the first bout of NREM sleep (bout ≥28 s or seven consecutive epochs) or REM sleep (bout ≥16 s or four consecutive epochs).
Genome-wide gene expression
RNA isolation
Adult male (8–10 week-old) Shank3∆C mice and WT littermates were divided into two groups: homecage controls (WT n = 5; Shank3∆Cn = 5) and sleep deprived (WT n = 5; Shank3∆Cn = 5). All mice were individually housed. Homecage control mice were left undisturbed and sacrificed 5 hr after light onset (hour 6). Mice in the sleep deprived group were sleep deprived for 5 hr via gentle handling starting at light onset (hour 1) and then sacrificed upon completion of sleep deprivation (hour 6) without allowing for any recovery sleep. Mice were sacrificed by live cervical dislocation (alternating between homecage controls and sleep deprived mice), decapitated, and prefrontal cortex was swiftly dissected on a cold block. Tissue was flash frozen in liquid nitrogen and stored at −80°C until processing (Poplawski et al., 2016; Tudor et al., 2016). To avoid day-specific effects on gene expression, this protocol was repeated over a 5 day period, and one sample from each genotype + treatment group was collected per day. Tissue was collected within the first 15 min of the hour. All tissue samples were later homogenized in Qiazol buffer (Qiagen, Hilden, Germany) using a TissueLyser (Qiagen) and all RNA was extracted using the Qiagen RNAeasy kit (Qiagen) on the same day.
RNA-seq library preparation and sequencing
The integrity of total RNA was assessed using Fragment Analyzer (Advanced Analytical Technologies, Inc, Ankeny, IA) with the High Sensitivity RNA Analysis Kit (Advanced Analytical Technologies, Inc). RNA Quality Numbers (RQNs) from 1 to 10 were assigned to each sample to indicate its integrity or quality. ‘10’ stands for a perfect RNA sample without any degradation, whereas ‘1’ marks a completely degraded sample. All RNA samples had RQNs ranging from 8.8 to 10 and were used for RNA library preparation with the TruSeq Stranded mRNA Library Prep Kit (Illumina, San Diego, CA). Briefly, mRNA was isolated from 2.5 µg of total RNA using poly-T oligo attached to magnetic beads and then subjected to fragmentation, followed by cDNA synthesis, dA-tailing, adaptor ligation, and PCR enrichment. The sizes of RNA libraries were assessed by Fragment Analyzer with the High Sensitivity NGS Fragment Analysis Kit (Advanced Analytical Technologies, Inc). The concentrations of RNA libraries were measured by StepOnePlus Real-Time PCR System (ThermoFisher Scientific, San Jose, CA) with the KAPA Library Quantification Kit (Kapabiosystems, Wilmington, MA). The libraries were diluted to 2 nM with Tris buffer (10 mM Tris-HCl, pH 8.5) and denatured with 0.1 N NaOH. Eighteen pM libraries were clustered in a high-output flow cell using HiSeq Cluster Kit v4 on a cBot (Illumina). After cluster generation, the flow cell was loaded onto HiSeq 2500 for sequencing using HiSeq SBS kit v4 (Illumina). DNA was sequenced from both ends (paired-end) with a read length of 100 bp.
RNA-seq data analysis
HiSeq Control Software version 2.2.68 was used for base calling. The raw bcl files were converted to fastq files using the software program bcl2fastq2.17.1.14. Adaptors were trimmed from the fastq files during the conversion. The average sequencing depth for all samples was 52 million read pairs. Sequenced reads were mapped with GSNAP (parameter = -N 1 m 7 -A sam --nofails) to mm10. On average 84% of the sequenced reads mapped uniquely to the mm10 genome. All statistical analyses were performed using open source software freely available through the R/Bioconductor project (Gentleman et al., 2004). The count matrix was obtained using featureCounts (parameters = isPairedEnd = TRUE, requireBothEndsMapped = TRUE, annot.inbuilt = ‘mm10’) from the package Rsubread (v. 1.26.1) under Bioconductor (version 3.6) with R (v. 3.4.1) using mm10 built-in annotations (Mus_musculus.GRCm38.90) with ENTREZ ID as row names. Gene counts were filtered using a proportion test (counts per million cutoff of 1), as implemented in the NOISeq package (v.2.22.1) (Tarazona et al., 2015). Removal of unwanted variation (RUV) normalization was performed using RUVSeq (v. 1.12.0) (Risso et al., 2014; Peixoto et al., 2015) after the data was normalized by Trimmed Mean of M-values (TMM) (Robinson and Oshlack, 2010) using edgeR (v. 3.20.9) (Robinson et al., 2010). Specifically, RUVs (with k = 5) was used after defining groups based on both genotype and treatment and using a list of 2677 negative control genes obtained as genes with an adjusted p-value>0.9 in the comparison between sleep deprivation and controls in microarray data available through GEO (GSE78215) (obtained from Gerstner et al., 2016). Analogously, a list of 579 positive control genes obtained from the same study were used to evaluate the ability of our RNA-seq study to replicate known differences in gene expression. These positive controls were genes found to be differentially expressed (FDR < 0.005) by 5 hr of sleep deprivation relative to homecage controls in WT mice in four independent studies using two different microarray platforms and two different brain regions (Additional file 2 from Gerstner et al., 2016). Figure 4—figure supplement 1 shows that our RNA-seq study is able to recover 78% of the positive controls as differentially expressed when performing the same comparison done in the previous studies (HC5 vs SD5 in WT), while being able to detect many more differences in gene expression. This is a very high level of concordance considering these controls were obtained using a different technology and data were collected from experiments in four independent laboratories. Therefore, we are confident in the ability of our study to detect differences in gene expression that can be independently validated.
Differential expression analysis was performed using edgeR (v. 3.20.9) with a factorial design that included genotype (WT or Shank3∆C) and treatment (homecage control or sleep deprivation). We specified the following contrasts: wild type sleep deprived vs. wild type homecage controls (WTSD vs. WTHC), Shank3∆C sleep deprived vs. Shank3∆C homecage controls (S3SD vs. S3HC), Shank3∆C homecage controls vs. wild type homecage controls (S3HC vs. WTHC), and Shank3∆C sleep deprived vs. wild type sleep deprived (S3SD vs. WTSD). To gain insight on the different effects of sleep deprivation between wild type and mutant mice, we considered the union of the differentially expressed genes between genotypes both in homecage controls and sleep deprived mice. We clustered the genes using k-means (k = 3) and the samples using hierarchical clustering based on the selected genes. The resulting ordered genes and samples were visualized as a heatmap using the pheatmap package (v. 1.0.10). The R code used for RNA-seq data analysis is publicly available at https://github.com/drighelli/peixoto (Righelli et al., 2019; copy archived at https://github.com/elifesciences-publications/peixoto).
Functional annotation analysis of genes differentially expressed at FDR < 0.1 was based on Ensembl Gene IDs and performed using the database for annotation, visualization, and integrated discovery v 6.7 (DAVID, https://david.ncifcrf.gov). The following functional categories were used: GO Biological Process, GO Molecular Function, Uniprot keywords, and KEGG pathways. Enrichment was determined relative to all genes expressed in the mouse prefrontal cortex based on our data. We used an enrichment p-value (EASE) cutoff <0.1 for individual functional terms. Enriched functional terms were clustered at low stringency, to obtain clusters with an enrichment score >1.2 (corresponding to an average EASE >0.05).
Assessment of circadian rhythms
Experimental design
Adult male (9-week-old) wild type (n = 8) and Shank3∆C (n = 7) littermate mice were individually housed with a low-profile wireless running wheel (Med Associates Inc, Fairfax, VT) and kept under 12:12 hr light:dark (LD) cycle for 3 weeks (including 1 week of habituation) followed by 3 weeks of complete darkness (DD). Two Shank3∆C mice from a starting n of 9 were excluded from the experiments because they failed run on the wheel for the duration of the experiment. The experiment was repeated with an independent cohort of 11-week-old male wild type (n = 8) and Shank3∆C (n = 8) littermate mice kept under a 12:12 hr light:dark (LD) cycle for one week of habituation and then 5 weeks of data collection. One Shank3∆C mouse from a starting n of 9 was excluded from the second experiment because it failed to run on the wheel for the duration of the experiment. Daily checks were made to verify that each running wheel was transmitting data and that no light was detected in the room during constant darkness conditions.
Data acquisition and analysis
Wheel-running behavior was continuously monitored using Wheel Manager Software and exported into 10 min time bins (Med Associates Inc). ClockLab version 6.0.50 (Actimetrics, Wilmette, IL) was used for analysis. Period, the circadian length in time from activity onset to the next activity onset, was calculated using ClockLab’s chi-squared periodogram analysis function. Alpha, the length of the activity period, was calculated as the time from activity onset to offset (defined by manufacturer defaults). Activity was calculated as the total sum of wheel revolutions per day. A linear mixed-effects model ANOVA was used to estimate the contributions of genotype and the interaction between genotype and time for period, alpha, and wheel running activity for LD week three through DD week three in the first experiment, and LD week 1–5 in the second experiment. The model was fitted using genotype, time, and their interaction as fixed effects, and the individual mice were labeled as a random effect. The analysis was conducted with the R package nlme (v. 3.5.0) and is publicly available at https://github.com/drighelli/peixoto (Righelli et al., 2019; copy archived at https://github.com/elifesciences-publications/peixoto).
Acknowledgements
Data used in the preparation of this article were obtained from the Phelan-McDermid Syndrome International Registry (PMSIR) version dated 12/1/2016.
We thank Dr. Paul Worley for providing the Shank3 mutant mouse line. We thank Dr. Hans Van Dongen and Dr. John Hogenesch for valuable discussion of the results. We thank Dr. Claudia Angelini for valuable discussion of the statistical methods.
Funding Statement
The funders had no role in study design, data collection and interpretation, or the decision to submit the work for publication.
Contributor Information
Marcos G Frank, Email: marcos.frank@wsu.edu.
Lucia Peixoto, Email: lucia.peixoto@wsu.edu.
Huda Y Zoghbi, Texas Children's Hospital, United States.
Eunjoon Kim, Institute for Basic Science, Korea Advanced Institute of Science and Technology, Republic of Korea.
Funding Information
This paper was supported by the following grant:
National Institute of Neurological Disorders and Stroke K01NS104172 to Lucia Peixoto.
Additional information
Competing interests
No competing interests declared.
Author contributions
Conceptualization, Formal analysis, Investigation, Visualization, Methodology, Writing—original draft, Writing—review and editing.
Data curation, Formal analysis, Investigation, Visualization, Writing—review and editing.
Formal analysis, Investigation, Visualization, Methodology, Writing—original draft, Writing—review and editing.
Investigation, Project administration, Writing—review and editing.
Formal analysis, Visualization, Methodology, Writing—review and editing.
Data curation, Formal analysis, Methodology, Writing—review and editing.
Investigation, Methodology.
Conceptualization, Supervision, Visualization, Methodology, Writing—review and editing.
Conceptualization, Resources, Supervision, Methodology, Writing—review and editing.
Conceptualization, Resources, Data curation, Formal analysis, Supervision, Funding acquisition, Investigation, Visualization, Methodology, Writing—original draft, Project administration, Writing—review and editing.
Ethics
Human subjects: PMSIR patient data was obtained de-identified under IRB exemption 15005-Peixoto.
Animal experimentation: This study was performed in strict accordance with the recommendations in the Guide for the Care and Use of Laboratory Animals of the National Institutes of Health. All of the animals were handled according to approved institutional animal care and use committee (IACUC) protocols of Washington State University. Protocol numbers: 04705-001 and 04704-001 IACUC 6155-Peixoto Breeding IACUC 4705-Peixoto experimental IACUC 4581-Frank experimental.
Additional files
Linear mixed-effects ANOVA for LD week 1–5. The model was fitted using genotype, time, and their interaction as fixed effects and the individual mice labeled as random effect. There is a significant effect for genotype x time in activity (p<0.0001), a significant effect of genotype for period (p=0.047), and no significant effect for genotype x time or genotype for alpha. Significance at p<0.05. numDF, numerator degrees of freedom. denDF, denominator degrees of freedom.
Summary of wheel-running behavior measured for each week of 12:12 hr light:dark cycles for both wild type and Shank3∆C mice. A linear mixed-effects ANOVA was used to estimate the contributions of genotype and the interaction between genotype and time for period, alpha, and wheel-running activity for LD week 1 through 5. There is no significant interaction effect for genotype x time for period (p=0.41), but there is a significant effect of genotype (p=0.047, indicated by *). There is no significant interaction effect for genotype x time for alpha (p=0.43), but there is a significant effect of time (p=0.004, indicated by †). There is a significant interaction effect (indicated by vertical bar) for genotype x time for activity (p<0.0001). See Source data 1 for details. Values are means ± SEM for wild type (n = 8) and Shank3∆C (n = 8) mice.
Data availability
Sequencing data have been deposited in GEO under accession code GSE113754. Source data files have been provided for Figures 1-4, Tables 1 and 2, and Supplementary Files 1 and 2. The R code used in this article is available on GitHub (https://github.com/drighelli/peixoto; copy archived at https://github.com/elifesciences-publications/peixoto). The R code used for the statistical analysis of RNA-seq and circadian wheel running data is also available in Source Code file 1.
The following dataset was generated:
Peixoto L, Roser L, Schoch H, Wintler T. 2018. RNA-Seq analysis of Sleep deprivation in wildtype and Shank3 mutant mice. NCBI Gene Expression Omnibus. GSE113754
The following previously published dataset was used:
Peixoto L, Frank M, Gerstner J. 2016. Gene expression linked to sleep homeostasis in murine cortex. NCBI Gene Expression Omnibus. GSE78215
References
- Achermann P, Borbely AA. Sleep homeostasis and models of sleep regulation. In: Kryger M, Roth T, Dement W. C, editors. Principles and Practice of Sleep Medicine. 6th Ed. Philadelphia: Elsevier; 2017. pp. 377–387. [DOI] [Google Scholar]
- American Psychiatric Association . Diagnostic and Statistical Manual of Mental Disorders. 5th Edition. Arlington, VA: American Psychiatric Association; 2013. [Google Scholar]
- Anders TF, Eiben LA. Pediatric sleep disorders: a review of the past 10 years. Journal of the American Academy of Child & Adolescent Psychiatry. 1997;36:9–20. doi: 10.1097/00004583-199701000-00012. [DOI] [PubMed] [Google Scholar]
- Andersen IM, Kaczmarska J, McGrew SG, Malow BA. Melatonin for insomnia in children with autism spectrum disorders. Journal of Child Neurology. 2008;23:482–485. doi: 10.1177/0883073807309783. [DOI] [PubMed] [Google Scholar]
- Angelakos CC, Watson AJ, O'Brien WT, Krainock KS, Nickl-Jockschat T, Abel T. Hyperactivity and male-specific sleep deficits in the 16p11.2 deletion mouse model of autism. Autism Research. 2017;10:572–584. doi: 10.1002/aur.1707. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Archer SN, Schmidt C, Vandewalle G, Dijk DJ. Phenotyping of PER3 variants reveals widespread effects on circadian preference, sleep regulation, and health. Sleep Medicine Reviews. 2018;40:109–126. doi: 10.1016/j.smrv.2017.10.008. [DOI] [PubMed] [Google Scholar]
- Baio J, Wiggins L, Christensen DL, Maenner MJ, Daniels J, Warren Z, Kurzius-Spencer M, Zahorodny W, Robinson C, Rosenberg, White T, Durkin MS, Imm P, Nikolaou L, Yeargin-Allsopp M, Lee L-C, Harrington R, Lopez M, Fitzgerald RT, Hewitt A, Pettygrove S, Constantino JN, Vehorn A, Shenouda J, Hall-Lande J, Van K, Naarden, Braun, Dowling NF. Prevalence of autism spectrum disorder among children aged 8 years — Autism and Developmental Disabilities Monitoring Network, 11 Sites, United States, 2014. MMWR. Surveillance Summaries. 2018;67:1–23. doi: 10.15585/mmwr.ss6706a1. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Baracchi F, Opp MR. Sleep-wake behavior and responses to sleep deprivation of mice lacking both interleukin-1 beta receptor 1 and tumor necrosis factor-alpha receptor 1. Brain, Behavior, and Immunity. 2008;22:982–993. doi: 10.1016/j.bbi.2008.02.001. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Baweja R, Calhoun S, Baweja R, Singareddy R. Sleep problems in children. Minerva Pediatrica. 2013;65:457–472. [PubMed] [Google Scholar]
- Bixler EO, Vgontzas AN, Lin HM, Liao D, Calhoun S, Vela-Bueno A, Fedok F, Vlasic V, Graff G. Sleep disordered breathing in children in a general population sample: prevalence and risk factors. Sleep. 2009;32:731–736. doi: 10.1093/sleep/32.6.731. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Boone CE, Davoudi H, Harrold JB, Foster DJ. Abnormal sleep architecture and hippocampal circuit dysfunction in a mouse model of fragile X syndrome. Neuroscience. 2018;384:275–289. doi: 10.1016/j.neuroscience.2018.05.012. [DOI] [PubMed] [Google Scholar]
- Bozdagi O, Sakurai T, Papapetrou D, Wang X, Dickstein DL, Takahashi N, Kajiwara Y, Yang M, Katz AM, Scattoni ML, Harris MJ, Saxena R, Silverman JL, Crawley JN, Zhou Q, Hof PR, Buxbaum JD. Haploinsufficiency of the autism-associated Shank3 gene leads to deficits in synaptic function, social interaction, and social communication. Molecular Autism. 2010;1:15. doi: 10.1186/2040-2392-1-15. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Bro D, O'Hara R, Primeau M, Hanson-Kahn A, Hallmayer J, Bernstein JA. Sleep disturbances in individuals with Phelan-McDermid syndrome: correlation with caregivers' Sleep quality and daytime functioning. Sleep. 2017;40 doi: 10.1093/sleep/zsw062. [DOI] [PubMed] [Google Scholar]
- Bushey D, Tononi G, Cirelli C. The Drosophila fragile X mental retardation gene regulates sleep need. Journal of Neuroscience. 2009;29:1948–1961. doi: 10.1523/JNEUROSCI.4830-08.2009. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Cochoy DM, Kolevzon A, Kajiwara Y, Schoen M, Pascual-Lucas M, Lurie S, Buxbaum JD, Boeckers TM, Schmeisser MJ. Phenotypic and functional analysis of SHANK3 stop mutations identified in individuals with ASD and/or ID. Molecular Autism. 2015;6:23. doi: 10.1186/s13229-015-0020-5. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Cohen S, Conduit R, Lockley SW, Rajaratnam SM, Cornish KM. The relationship between sleep and behavior in autism spectrum disorder (ASD): a review. Journal of Neurodevelopmental Disorders. 2014;6:44. doi: 10.1186/1866-1955-6-44. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Cotton S, Richdale A. Brief report: parental descriptions of sleep problems in children with autism, down syndrome, and Prader-Willi syndrome. Research in Developmental Disabilities. 2006;27:151–161. doi: 10.1016/j.ridd.2004.12.003. [DOI] [PubMed] [Google Scholar]
- de la Torre-Ubieta L, Won H, Stein JL, Geschwind DH. Advancing the understanding of autism disease mechanisms through genetics. Nature Medicine. 2016;22:345–361. doi: 10.1038/nm.4071. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Dhamne SC, Silverman JL, Super CE, Lammers SHT, Hameed MQ, Modi ME, Copping NA, Pride MC, Smith DG, Rotenberg A, Crawley JN, Sahin M. Replicable in vivo physiological and behavioral phenotypes of the Shank3B null mutant mouse model of autism. Molecular Autism. 2017;8 doi: 10.1186/s13229-017-0142-z. [DOI] [PMC free article] [PubMed] [Google Scholar]
- El Helou J, Bélanger-Nelson E, Freyburger M, Dorsaz S, Curie T, La Spada F, Gaudreault PO, Beaumont É, Pouliot P, Lesage F, Frank MG, Franken P, Mongrain V. Neuroligin-1 links neuronal activity to sleep-wake regulation. PNAS. 2013;110:9974–9979. doi: 10.1073/pnas.1221381110. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Feenstra MG, Botterblom MH, Mastenbroek S. Dopamine and noradrenaline efflux in the prefrontal cortex in the light and dark period: effects of novelty and handling and comparison to the nucleus accumbens. Neuroscience. 2000;100:741–748. doi: 10.1016/S0306-4522(00)00319-5. [DOI] [PubMed] [Google Scholar]
- Frank MG, Stryker MP, Tecott LH. Sleep and sleep homeostasis in mice lacking the 5-HT2c receptor. Neuropsychopharmacology. 2002;27:869–873. doi: 10.1016/S0893-133X(02)00353-6. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Franken P, Chollet D, Tafti M. The homeostatic regulation of sleep need is under genetic control. The Journal of Neuroscience. 2001;21:2610–2621. doi: 10.1523/JNEUROSCI.21-08-02610.2001. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Franken P. A role for clock genes in sleep homeostasis. Current Opinion in Neurobiology. 2013;23:864–872. doi: 10.1016/j.conb.2013.05.002. [DOI] [PubMed] [Google Scholar]
- Gail Williams P, Sears LL, Allard A. Sleep problems in children with autism. Journal of Sleep Research. 2004;13:265–268. doi: 10.1111/j.1365-2869.2004.00405.x. [DOI] [PubMed] [Google Scholar]
- Gentleman RC, Carey VJ, Bates DM, Bolstad B, Dettling M, Dudoit S, Ellis B, Gautier L, Ge Y, Gentry J, Hornik K, Hothorn T, Huber W, Iacus S, Irizarry R, Leisch F, Li C, Maechler M, Rossini AJ, Sawitzki G, Smith C, Smyth G, Tierney L, Yang JY, Zhang J. Bioconductor: open software development for computational biology and bioinformatics. Genome Biology. 2004;5:R80. doi: 10.1186/gb-2004-5-10-r80. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Gerstner JR, Koberstein JN, Watson AJ, Zapero N, Risso D, Speed TP, Frank MG, Peixoto L. Removal of unwanted variation reveals novel patterns of gene expression linked to sleep homeostasis in murine cortex. BMC Genomics. 2016;17:727. doi: 10.1186/s12864-016-3065-8. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Giannotti F, Cortesi F, Cerquiglini A, Bernabei P. An open-label study of controlled-release melatonin in treatment of sleep disorders in children with autism. Journal of Autism and Developmental Disorders. 2006;36:741–752. doi: 10.1007/s10803-006-0116-z. [DOI] [PubMed] [Google Scholar]
- Giannotti F, Cortesi F, Cerquiglini A, Miraglia D, Vagnoni C, Sebastiani T, Bernabei P. An investigation of sleep characteristics, EEG abnormalities and epilepsy in developmentally regressed and non-regressed children with autism. Journal of Autism and Developmental Disorders. 2008;38:1888–1897. doi: 10.1007/s10803-008-0584-4. [DOI] [PubMed] [Google Scholar]
- Glickman G. Circadian rhythms and sleep in children with autism. Neuroscience & Biobehavioral Reviews. 2010;34:755–768. doi: 10.1016/j.neubiorev.2009.11.017. [DOI] [PubMed] [Google Scholar]
- Grabrucker S, Proepper C, Mangus K, Eckert M, Chhabra R, Schmeisser MJ, Boeckers TM, Grabrucker AM. The PSD protein ProSAP2/Shank3 displays synapto-nuclear shuttling which is deregulated in a schizophrenia-associated mutation. Experimental Neurology. 2014;253:126–137. doi: 10.1016/j.expneurol.2013.12.015. [DOI] [PubMed] [Google Scholar]
- Halassa MM, Florian C, Fellin T, Munoz JR, Lee SY, Abel T, Haydon PG, Frank MG. Astrocytic modulation of sleep homeostasis and cognitive consequences of sleep loss. Neuron. 2009;61:213–219. doi: 10.1016/j.neuron.2008.11.024. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Harada Y, Sakai M, Kurabayashi N, Hirota T, Fukada Y. Ser-557-phosphorylated mCRY2 is degraded upon synergistic phosphorylation by glycogen synthase kinase-3 beta. Journal of Biological Chemistry. 2005;280:31714–31721. doi: 10.1074/jbc.M506225200. [DOI] [PubMed] [Google Scholar]
- Harris KP, Akbergenova Y, Cho RW, Baas-Thomas MS, Littleton JT. Shank modulates postsynaptic wnt signaling to regulate synaptic development. Journal of Neuroscience. 2016;36:5820–5832. doi: 10.1523/JNEUROSCI.4279-15.2016. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Hastie TJ, Tibshirani RJ. Generalized Additive Models (Monographs on Statistics and Applied Probability 43. Chapman and Hall/CRC; 1990. [Google Scholar]
- He Y, Jones CR, Fujiki N, Xu Y, Guo B, Holder JL, Rossner MJ, Nishino S, Fu YH. The transcriptional repressor DEC2 regulates sleep length in mammals. Science. 2009;325:866–870. doi: 10.1126/science.1174443. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Hirano A, Hsu PK, Zhang L, Xing L, McMahon T, Yamazaki M, Ptáček LJ, Fu YH. DEC2 modulates orexin expression and regulates sleep. PNAS. 2018;115:3434–3439. doi: 10.1073/pnas.1801693115. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Hodge D, Carollo TM, Lewin M, Hoffman CD, Sweeney DP. Sleep patterns in children with and without autism spectrum disorders: developmental comparisons. Research in Developmental Disabilities. 2014;35:1631–1638. doi: 10.1016/j.ridd.2014.03.037. [DOI] [PubMed] [Google Scholar]
- Hysing M, Pallesen S, Stormark KM, Lundervold AJ, Sivertsen B. Sleep patterns and insomnia among adolescents: a population-based study. Journal of Sleep Research. 2013;22:549–556. doi: 10.1111/jsr.12055. [DOI] [PubMed] [Google Scholar]
- Iitaka C, Miyazaki K, Akaike T, Ishida N. A role for glycogen synthase kinase-3beta in the mammalian circadian clock. Journal of Biological Chemistry. 2005;280:29397–29402. doi: 10.1074/jbc.M503526200. [DOI] [PubMed] [Google Scholar]
- Ingiosi AM, Raymond RM, Pavlova MN, Opp MR. Selective contributions of neuronal and astroglial interleukin-1 receptor 1 to the regulation of sleep. Brain, Behavior, and Immunity. 2015;48:244–257. doi: 10.1016/j.bbi.2015.03.014. [DOI] [PubMed] [Google Scholar]
- Jaramillo TC, Speed HE, Xuan Z, Reimers JM, Liu S, Powell CM. Altered striatal synaptic function and abnormal behaviour in Shank3 Exon4-9 Deletion Mouse Model of Autism. Autism Research. 2016;9:350–375. doi: 10.1002/aur.1529. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Jaramillo TC, Speed HE, Xuan Z, Reimers JM, Escamilla CO, Weaver TP, Liu S, Filonova I, Powell CM. Novel Shank3 mutant exhibits behaviors with face validity for autism and altered striatal and hippocampal function. Autism Research. 2017;10:42–65. doi: 10.1002/aur.1664. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Johnson KP, Giannotti F, Cortesi F. Sleep patterns in autism spectrum disorders. Child and Adolescent Psychiatric Clinics of North America. 2009;18:917–928. doi: 10.1016/j.chc.2009.04.001. [DOI] [PubMed] [Google Scholar]
- Kouser M, Speed HE, Dewey CM, Reimers JM, Widman AJ, Gupta N, Liu S, Jaramillo TC, Bangash M, Xiao B, Worley PF, Powell CM. Loss of predominant Shank3 isoforms results in hippocampus-dependent impairments in behavior and synaptic transmission. Journal of Neuroscience. 2013;33:18448–18468. doi: 10.1523/JNEUROSCI.3017-13.2013. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Krakowiak P, Goodlin-Jones B, Hertz-Picciotto I, Croen LA, Hansen RL. Sleep problems in children with autism spectrum disorders, developmental delays, and typical development: a population-based study. Journal of Sleep Research. 2008;17:197–206. doi: 10.1111/j.1365-2869.2008.00650.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Leger D, Beck F, Richard JB, Godeau E. Total sleep time severely drops during adolescence. PLOS ONE. 2012;7:e45204. doi: 10.1371/journal.pone.0045204. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Liu X, Hubbard JA, Fabes RA, Adam JB. Sleep disturbances and correlates of children with autism spectrum disorders. Child Psychiatry and Human Development. 2006;37:179–191. doi: 10.1007/s10578-006-0028-3. [DOI] [PubMed] [Google Scholar]
- Liu JJ, Grace KP, Horner RL, Cortez MA, Shao Y, Jia Z. Neuroligin 3 R451C mutation alters electroencephalography spectral activity in an animal model of autism spectrum disorders. Molecular Brain. 2017;10:10. doi: 10.1186/s13041-017-0290-2. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Loessl B, Valerius G, Kopasz M, Hornyak M, Riemann D, Voderholzer U. Are adolescents chronically sleep-deprived? an investigation of sleep habits of adolescents in the southwest of Germany. Child: Care, Health and Development. 2008;34:549–556. doi: 10.1111/j.1365-2214.2008.00845.x. [DOI] [PubMed] [Google Scholar]
- Lozoff B, Wolf AW, Davis NS. Sleep problems seen in pediatric practice. Pediatrics. 1985;75:477–483. [PubMed] [Google Scholar]
- Lumeng JC, Chervin RD. Epidemiology of pediatric obstructive sleep apnea. Proceedings of the American Thoracic Society. 2008;5:242–252. doi: 10.1513/pats.200708-135MG. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Menon JML, Nolten C, Achterberg EJM, Joosten R, Dematteis M, Feenstra MGP, Drinkenburg WHP, Leenaars CHC. Brain microdialysate monoamines in relation to circadian rhythms, sleep, and sleep deprivation - a systematic review, network Meta-analysis, and new primary data. Journal of Circadian Rhythms. 2019;17:1. doi: 10.5334/jcr.174. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Miano S, Bruni O, Elia M, Trovato A, Smerieri A, Verrillo E, Roccella M, Terzano MG, Ferri R. Sleep in children with autistic spectrum disorder: a questionnaire and polysomnographic study. Sleep Medicine. 2007;9:64–70. doi: 10.1016/j.sleep.2007.01.014. [DOI] [PubMed] [Google Scholar]
- Ohayon MM, Roberts RE, Zulley J, Smirne S, Priest RG. Prevalence and patterns of problematic sleep among older adolescents. Journal of the American Academy of Child & Adolescent Psychiatry. 2000;39:1549–1556. doi: 10.1097/00004583-200012000-00019. [DOI] [PubMed] [Google Scholar]
- Paavonen EJ, Vehkalahti K, Vanhala R, von Wendt L, Nieminen-von Wendt T, Aronen ET. Sleep in children with asperger syndrome. Journal of Autism and Developmental Disorders. 2008;38:41–51. doi: 10.1007/s10803-007-0360-x. [DOI] [PubMed] [Google Scholar]
- Pallesen S, Hetland J, Sivertsen B, Samdal O, Torsheim T, Nordhus IH. Time trends in sleep-onset difficulties among norwegian adolescents: 1983--2005. Scandinavian Journal of Public Health. 2008;36:889–895. doi: 10.1177/1403494808095953. [DOI] [PubMed] [Google Scholar]
- Patzold LM, Richdale AL, Tonge BJ. An investigation into sleep characteristics of children with autism and Asperger's Disorder. Journal of Paediatrics and Child Health. 1998;34:528–533. doi: 10.1046/j.1440-1754.1998.00291.x. [DOI] [PubMed] [Google Scholar]
- Peça J, Feliciano C, Ting JT, Wang W, Wells MF, Venkatraman TN, Lascola CD, Fu Z, Feng G. Shank3 mutant mice display autistic-like behaviours and striatal dysfunction. Nature. 2011;472:437–442. doi: 10.1038/nature09965. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Peixoto L, Risso D, Poplawski SG, Wimmer ME, Speed TP, Wood MA, Abel T. How data analysis affects power, reproducibility and biological insight of RNA-seq studies in complex datasets. Nucleic Acids Research. 2015;43:7664–7674. doi: 10.1093/nar/gkv736. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Phelan K, McDermid HE. The 22q13.3 Deletion Syndrome (Phelan-McDermid Syndrome) Molecular Syndromology. 2012;2:186–201. doi: 10.1159/000334260. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Polimeni MA, Richdale AL, Francis AJ. A survey of sleep problems in autism, Asperger's disorder and typically developing children. Journal of Intellectual Disability Research. 2005;49:260–268. doi: 10.1111/j.1365-2788.2005.00642.x. [DOI] [PubMed] [Google Scholar]
- Poplawski SG, Peixoto L, Porcari GS, Wimmer ME, McNally AG, Mizuno K, Giese KP, Chatterjee S, Koberstein JN, Risso D, Speed TP, Abel T. Contextual fear conditioning induces differential alternative splicing. Neurobiology of Learning and Memory. 2016;134:221–235. doi: 10.1016/j.nlm.2016.07.018. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Qin L, Ma K, Wang ZJ, Hu Z, Matas E, Wei J, Yan Z. Social deficits in Shank3-deficient mouse models of autism are rescued by histone deacetylase (HDAC) inhibition. Nature Neuroscience. 2018;21:564–575. doi: 10.1038/s41593-018-0110-8. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Richdale AL, Prior MR. The sleep/wake rhythm in children with autism. European Child & Adolescent Psychiatry. 1995;4:175–186. doi: 10.1007/BF01980456. [DOI] [PubMed] [Google Scholar]
- Righelli D, Risso D, Roser LG. GitHub; 2019. https://github.com/drighelli/peixoto [DOI] [PMC free article] [PubMed] [Google Scholar]
- Risso D, Ngai J, Speed TP, Dudoit S. Normalization of RNA-seq data using factor analysis of control genes or samples. Nature Biotechnology. 2014;32:896–902. doi: 10.1038/nbt.2931. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Robinson MD, McCarthy DJ, Smyth GK. edgeR: a bioconductor package for differential expression analysis of digital gene expression data. Bioinformatics. 2010;26:139–140. doi: 10.1093/bioinformatics/btp616. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Robinson MD, Oshlack A. A scaling normalization method for differential expression analysis of RNA-seq data. Genome Biology. 2010;11:R25. doi: 10.1186/gb-2010-11-3-r25. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Sadeh A, Raviv A, Gruber R. Sleep patterns and sleep disruptions in school-age children. Developmental Psychology. 2000;36:291–301. doi: 10.1037/0012-1649.36.3.291. [DOI] [PubMed] [Google Scholar]
- Sakai Y, Shaw CA, Dawson BC, Dugas DV, Al-Mohtaseb Z, Hill DE, Zoghbi HY. Protein interactome reveals converging molecular pathways among autism disorders. Science Translational Medicine. 2011;3:86ra49. doi: 10.1126/scitranslmed.3002166. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Seok BS, Cao F, Bélanger-Nelson E, Provost C, Gibbs S, Jia Z, Mongrain V. The effect of Neuroligin-2 absence on sleep architecture and electroencephalographic activity in mice. Molecular Brain. 2018;11:52. doi: 10.1186/s13041-018-0394-3. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Soorya L, Kolevzon A, Zweifach J, Lim T, Dobry Y, Schwartz L, Frank Y, Wang AT, Cai G, Parkhomenko E, Halpern D, Grodberg D, Angarita B, Willner JP, Yang A, Canitano R, Chaplin W, Betancur C, Buxbaum JD. Prospective investigation of autism and genotype-phenotype correlations in 22q13 deletion syndrome and SHANK3 deficiency. Molecular Autism. 2013;4:18. doi: 10.1186/2040-2392-4-18. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Speed HE, Kouser M, Xuan Z, Reimers JM, Ochoa CF, Gupta N, Liu S, Powell CM. Autism-Associated insertion mutation (InsG) of Shank3 exon 21 causes impaired synaptic transmission and behavioral deficits. Journal of Neuroscience. 2015;35:9648–9665. doi: 10.1523/JNEUROSCI.3125-14.2015. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Tani P, Lindberg N, Nieminen-von Wendt T, von Wendt L, Alanko L, Appelberg B, Porkka-Heiskanen T. Insomnia is a frequent finding in adults with asperger syndrome. BMC Psychiatry. 2003;3:12. doi: 10.1186/1471-244X-3-12. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Tarazona S, Furió-Tarí P, Turrà D, Pietro AD, Nueda MJ, Ferrer A, Conesa A. Data quality aware analysis of differential expression in RNA-seq with NOISeq R/Bioc package. Nucleic Acids Research. 2015;13:gkv711. doi: 10.1093/nar/gkv711. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Thirumalai SS, Shubin RA, Robinson R. Rapid eye movement sleep behavior disorder in children with autism. Journal of Child Neurology. 2002;17:173–178. doi: 10.1177/088307380201700304. [DOI] [PubMed] [Google Scholar]
- Thomas AM, Schwartz MD, Saxe MD, Kilduff TS. Sleep/Wake physiology and quantitative electroencephalogram analysis of the Neuroligin-3 knockout rat model of autism spectrum disorder. Sleep. 2017;40 doi: 10.1093/sleep/zsx138. [DOI] [PubMed] [Google Scholar]
- Thomas GM, Huganir RL. MAPK cascade signalling and synaptic plasticity. Nature Reviews Neuroscience. 2004;5:173–183. doi: 10.1038/nrn1346. [DOI] [PubMed] [Google Scholar]
- Tudor ME, Hoffman CD, Sweeney DP. Children with autism: sleep problems and symptom severity. Focus Autism Dev. Disabil. 2012;27:254–262. doi: 10.1177/1088357612457989. [DOI] [Google Scholar]
- Tudor JC, Davis EJ, Peixoto L, Wimmer ME, van Tilborg E, Park AJ, Poplawski SG, Chung CW, Havekes R, Huang J, Gatti E, Pierre P, Abel T. Sleep deprivation impairs memory by attenuating mTORC1-dependent protein synthesis. Science Signaling. 2016;9:ra41. doi: 10.1126/scisignal.aad4949. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Vecsey CG, Peixoto L, Choi JH, Wimmer M, Jaganath D, Hernandez PJ, Blackwell J, Meda K, Park AJ, Hannenhalli S, Abel T. Genomic analysis of sleep deprivation reveals translational regulation in the hippocampus. Physiological Genomics. 2012;44:981–991. doi: 10.1152/physiolgenomics.00084.2012. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Verhoeff ME, Blanken LME, Kocevska D, Mileva-Seitz VR, Jaddoe VWV, White T, Verhulst F, Luijk M, Tiemeier H. The bidirectional association between sleep problems and autism spectrum disorder: a population-based cohort study. Molecular Autism. 2018;9 doi: 10.1186/s13229-018-0194-8. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Wiggs L, Stores G. Sleep patterns and sleep disorders in children with autistic spectrum disorders: insights using parent report and actigraphy. Developmental Medicine & Child Neurology. 2004;46:372–380. doi: 10.1017/S0012162204000611. [DOI] [PubMed] [Google Scholar]
- Yin L, Wang J, Klein PS, Lazar MA. Nuclear receptor Rev-erbalpha is a critical lithium-sensitive component of the circadian clock. Science. 2006;311:1002–1005. doi: 10.1126/science.1121613. [DOI] [PubMed] [Google Scholar]