

Abstract
In plants, sugars are required to sustain growth and regulate gene expression. A large set of genes are either up- or down-regulated by sugars; however, whether there is a common mechanism and signal transduction pathway for differential and coordinated sugar regulation remain unclear. In the present study, the rice (Oryza sativa cv Tainan 5) cell culture was used as a model system to address this question. Sucrose and glucose both played dual functions in gene regulation as exemplified by the up-regulation of growth-related genes and down-regulation of stress-related genes. Sugar coordinately but differentially activated or repressed gene expression, and nuclear run-on transcription and mRNA half-life analyses revealed regulation of both the transcription rate and mRNA stability. Although coordinately regulated by sugars, these growth- and stress-related genes were up-regulated or down-regulated through hexokinase-dependent and/or hexokinase-independent pathways. We also found that the sugar signal transduction pathway may overlap the glycolytic pathway for gene repression. α-Amylase and the stress-related genes identified in this study were coordinately expressed under sugar starvation, suggesting a convergence of the nutritional and environmental stress signal transduction pathways. Together, our studies provide a new insight into the complex signal transduction network and mechanisms of sugar regulation of growth and stress-related genes in plants.
In plants, sugars do not only function as metabolic resources and structural constituents of cells, they also act as important regulators of various processes associated with plant growth and development. A variety of genes, whose products are involved in diverse metabolic pathways and cellular functions, are either induced or repressed depending upon the availability of soluble sugars. In general, sugars favor the expression of enzymes in connection with biosynthesis, use, and storage of reserves (including starch, lipid, and proteins), while repressing the expression of enzymes involved in photosynthesis and reserve mobilization (Koch, 1996). A large and specific set of genes has been reported to be positively regulated by sugars. Examples include: (a) genes that encode storage proteins, e.g. patatin in potato and sporamin in sweet potato (Hattori et al., 1990; Jefferson et al., 1990); (b) genes that encode proteins related to starch biosynthesis, e.g. ADP-Glc pyrophosphorylase (Müller- Röber et al., 1990); (c) genes that encode defense proteins, e.g. proteinase inhibitor II in potato (Kim et al., 1991); and (d) genes that encode proteins for Suc metabolism, e.g. invertase and Suc synthase (Sus1) (Koch et al., 1992; Roitsch et al., 1995). In contrast, a variety of genes are negatively regulated by sugars, and their expression is induced by sugar deprivation, e.g. sugar represses expression of α-amylase genes in rice (Oryza sativa cv Tainan 5) suspension cells and germinating embryos (Yu et al., 1991, 1996); endopeptidase, Suc synthase (Sh1), and Asn synthase genes in maize root tips (Koch et al., 1992; James et al., 1993; Chevalier et al., 1995); and malate synthase and isocitrate lyase genes in cucumber cotyledon and suspension cells (Graham et al., 1994). It is not known whether a common mechanism is responsible for differential sugar regulation.
Plants are considered to be carbon autotrophs, but they can be considered as carbon heterotrophs during some part of their life cycle or in some of their non-green organs like roots, stems, and flowers that are not involved in photosynthesis. Furthermore, carbohydrate depletion can occur and is a fact of life in most plants. For instance, variations in environmental factors, such as light, water, or temperature, and attacks by pathogens or herbivores may lead to a significant decrease in the efficiency of photosynthesis in source tissues and thus reduce the supply of carbohydrates to sink tissues. Under conditions of sugar deprivation, substantial physiological and biochemical changes occur to sustain respiration and other metabolic processes (Yu, 1999a). When Suc is omitted from the nutrient medium of cell cultures or isolated tissues, cell growth ceases and the cellular starch and sugar levels, respiration rates, and metabolic activities dramatically decline (Journet et al., 1986; Brouquisse et al., 1992; Chen et al., 1994). Sugar starvation may also trigger an autophagic process involved in the regression of cytoplasm, including organelles and in the recycling of respiratory substrates (Chen et al., 1994; Aubert et al., 1996; Yu, 1999a).
Understanding the mechanisms involved in sugar signal transduction and sugar regulation of gene expression in plants is still in its early stages. Most studies on the mechanisms of sugar activation and sugar repression in plants have emphasized regulation at the transcriptional level (Sheen, 1990; Chan et al., 1994; Graham et al., 1994; Lu et al., 1998). However, sugar repression of α-amylase gene expression involves control of both transcription and mRNA stability (Sheu et al., 1994, 1996). A sugar response complex (SRC) in the promoter region of a Suc deprivation highly inducible rice α-amylase gene, αAmy3, has been identified. This SRC contains three essential motifs for a high level of sugar starvation-induced gene expression in rice cells (Lu et al., 1998). Studies on αAmy3 mRNA have also identified essential sequences in its 3′-untranslated region (3′-UTR) that control sugar-dependent mRNA stability (Chan and Yu, 1998a, 1998b). The 3′-UTR of a cell wall invertase gene (Incw1) recently was shown to be involved in translational control of Incw1 by sugars in cultured maize suspension cells (Cheng et al., 1999).
A considerable amount of information concerning the sugar signal transduction pathway is available from research in yeast (Carlson, 1987; Entian and Barnett, 1992; Gancedo, 1992). However, very few yeast homologs in plants have been studied or shown to serve similar functions or to be regulated in a manner similar to the yeast system. Hexokinase, the enzyme that catalyzes the phosphorylation of hexose sugars at the first step of the glycolytic pathway, has been implicated as a Glc sensor in organisms as diverse as yeast (Rose et al., 1991) and mammals (Efrat et al., 1994). Recent studies suggest that hexokinase also acts as the primary sugar sensor in plants (Jang et al., 1997; Smeekens and Rook, 1997). However, multiple sugar sensing pathways have also been proposed to exist in plants (Halford et al., 1999; Sheen et al., 1999; Smeekens, 2000). A gene (SnRK1) encoding the yeast Ser/Thr protein kinase (SNF1) homolog isolated from potato was recently shown to be required for activation of Suc synthase gene expression (Purcell et al., 1998). Whether SnRK1 activity is regulated by Glc or some other hexose and whether SnRK1 plays a role in the derepression of sugar-repressible genes in plants as in yeast (Ronne, 1995) are not known.
Expression of genes regulated by sugars can also be affected by various other factors, such as light (Sheen, 1990), phosphate (Sadka et al., 1994), hormones (DeWald et al., 1994; Zhou et al., 1998), pathogen infection (Herbers et al., 1996), as well as wounding and anaerobiosis (Salanoubat and Belliard, 1989). The mechanisms underlying the crosstalk between sugar and other signal transduction pathways and gene regulation are not clear. It will be interesting to determine whether different internal and external signals are integrated to result in the coordinated regulation of gene expression. The aim of the present study was to explore whether there is a common mechanism for differential and coordinated sugar regulation and to better understand the mechanism that connects the sensing and transmission of sugar signals with the regulation of gene expression in plants. Rice suspension cell culture, which is readily amenable to exogenous metabolic manipulations, was used as a model system for these studies. The usefulness of this type of cell culture in such studies is well documented (Graham et al., 1994; Ehness et al., 1997; Cheng et al., 1999; Yu, 1999a). In the present studies, some growth- and stress-related genes up- and down-regulated by Suc, respectively, were identified and found to be differentially and coordinately regulated by Suc, and the regulation involves control of both transcription rate and mRNA stability. Hexokinase-dependent and -independent pathways were found to be involved in up- and down-regulation of gene expression. Based on several lines of experimental evidence, we propose two potential complex signal transduction networks for differential and coordinated regulation of gene expression: one which connects the sugar signal transduction pathway to the sugar metabolic pathway and one which connects the sugar starvation signal to the stress signals.
RESULTS
Sugars Have Dual Functions in Gene Regulation
To study the mechanism that switches on and off sugar-dependent gene expression, it was necessary to identify genes whose expression are up- or down-regulated by sugars. Cells were first provided with Suc for 72 h, starved of Suc for 72 h, then provided with Suc for 24 h. Total RNA was purified from these cells and subjected to gel-blot analysis. Several cDNAs that encode proteins known to be required for cell growth, e.g. actin (Act), glyceraldehyde-3-phosphate dehydrogenase (G3PD), Histone H3 (His), and Suc synthase P-2 (SSP2), as well as various proteins related to stress response, e.g. alcohol dehydrogenase (ADH2), heat shock protein 86 (HSP86), and ubiquitin precursor (Ubi), were used as probes for the gel-blot analysis. The mRNA levels of α-amylase genes (α-Amy) increased after the onset of Suc starvation and decreased as cells were provided with Suc (Fig. 1a). These observations were consistent with our previous report (Sheu et al., 1994). In contrast, the mRNA levels of Act, ADH2, G3PD, His, HSP86, and SSP2 genes were initially high in Suc-provided cells and then decreased significantly after Suc starvation. Somewhat different expression patterns were observed for His and HSP86 genes. Accumulation of the His mRNA was dramatically and transiently increased 1 h after shifting cells from Suc-free to Suc-containing medium (Fig. 1a, lane 15). Accumulation of HSP86 mRNA progressively decreased and reached the lowest level 8 h after starvation (Fig. 1a, lane 9) and progressively increased afterward while cells remained under starvation. Three hybridization signals were observed for Ubi genes: one decreased after Suc starvation, one increased after Suc starvation, and the other one increased with culture age.
Figure 1.

Two groups of genes are coordinately regulated but in an opposite manner by sugars. a, RNA gel-blot analysis of growth-related genes. Rice suspension cells were cultured in Suc-containing (+S) medium for 72 h, transferred to Suc-free (−S) medium for 72 h, and transferred to +S medium for an additional 24 h. Total RNA was purified and fractionated on two duplicated 1% (w/v) agarose gels. Five membrane blots were parallel prepared and sequentially hybridized with indicated probes. The Act probe, His and Ubi probes, G3PD probe, HSP86 and ADH2 probes, and α-Amy and SSP2 probes were respectively hybridized to each of the five blots. The α-Amy and Act cDNA probes hybridized to the mRNAs of all α-amylase and actin genes, respectively (Sheu et al., 1996). b, RNA gel-blot analysis of stress-related genes. Five membrane blots were parallel prepared. The A2 probe, B1 probe, A3 probe, D1 and F1 probes, and E1 probe were respectively hybridized to each of the five blots.
To identify genes whose expression is down-regulated by sugars, we performed a differential screening of a cDNA library constructed from poly(A+) mRNA prepared from 4-h Suc-starved rice cells. More than 100 cDNA clones whose expression increased after Suc starvation were isolated. Twelve cDNA clones whose signals were significantly stronger by hybridization with cDNA probes of Suc-starved cells compared with cDNA probes of Suc-provided cells were selected for further characterization. By hybridization with the αAmy8-C probe, three of the cDNAs were found to be α-amylase genes and were not further characterized. Partial 3′-end nucleotide sequence analyses of the other nine cDNAs suggested that they were derived from six different genes. To confirm preferential induction by sugar starvation, the six selected genes were used as probes to hybridize to replicated gel blots of total cellular RNA as shown in Figure 1a. mRNA levels of these genes were low or almost undetectable in Suc-provided cells and increased in Suc-starved cells (Fig. 1b). Two hybridization signals were observed for clone E1: After Suc starvation, one increased only at early stages, but the other one increased progressively with time. Nucleotide sequences of these sugar down-regulated cDNAs were compared with sequence data in GenBank. cDNA B1 is highly homologous to a Gly-rich RNA-binding protein gene from rice (Macknight et al., 1998). cDNAs A2, D1, E1, and F1 are not identical but are similar and highly homologous to a gene encoding the virus-inducible Gly-rich cell wall protein from rice (Fang et al., 1991; Lei and Wu, 1991). cDNA A3 is highly homologous to a salt and drought stress-inducible gene, salT, from rice (Claes et al., 1990).
To determine whether accumulation of proteins of the sugar-regulated genes parallel mRNA levels, protein gel-blot analyses were performed using three antibodies against α-Amy, Act, and SSP2, respectively. As shown in Figure 2, the levels of α-amylases increased in Suc-starved cells and decreased in Suc-provided cells. In contrast, the levels of Act and SSP2 decreased in Suc-starved cells and increased in Suc-provided cells.
Figure 2.

Alterations in protein levels by sugars parallel alterations in mRNA levels. Rice suspension cells were cultured in Suc-containing (+S) medium for 72 h, transferred to Suc-free (−S) medium for 72 h, and transferred to +S medium for an additional 72 h. Total proteins were extracted from rice suspension cells and subjected to protein gel-blot analysis. The amount of total proteins applied in each lane was 10 μg for α-Amy and 50 μg for Act or SSP2.
Sugars Alter Gene Transcription
To define the mechanism of sugar-dependent up- and down-regulation of gene expression, the transcription rates of individual genes were determined. Nuclear run-on transcription analyses were performed with nuclei isolated from cells provided with or without Suc for 24 h. The transcription rates of rRNA genes in Suc-provided and Suc-starved cells (Fig. 3a, slot 9 and Fig. 3b, slot 10) were kept at the same levels and the transcription rates of the remaining genes were compared. For sugar up-regulated (SU) genes, only the transcription rates of G3PD and ADH2 genes were higher in Suc-provided cells than in Suc-starved cells (Fig. 3a, slots 4 and 5). It is surprising that the transcription rates of His, HSP86, and Act genes were lower in Suc-provided cells than in Suc-starved cells (Fig. 3a, slots 1, 2, and 7). The result of Act was consistent with our previous report (Sheu et al., 1994). The transcription rate of the SSP2 gene remained unchanged (Fig. 3a, slot 3). On the other hand, for all the sugar down-regulated (SD) genes, the transcription rates were higher in Suc-starved cells than in Suc-provided cells (Fig. 3b). The transcription rate of α-Amy was significantly higher in Suc-starved cells than in Suc-provided cells (Fig. 3a, slot 6 and Fig. 3b, slot 7), which was consistent with our previous study (Sheu et al., 1994). These results demonstrate that sugar down-regulation of gene expression involves transcriptional control, whereas sugar up-regulation of gene expression may or may not involve transcriptional control.
Figure 3.

Sugars affect transcription rates of sugar-regulated genes. Rice suspension cells were grown in Suc-containing (+S) medium or Suc-free (−S) medium for 24 h. Cells were collected and nuclei were isolated. Run-on transcription analysis was performed as previously described (Sheu et al., 1994). The indicated DNA was isolated from plasmids as described in “Materials and Methods.” Equal molecules of indicated DNA were slot-blotted on the membrane. a, Transcription of SU genes. b, Transcription of SD genes. pBS, pBluescript DNA. The blots were visualized with autoradiography and quantified using phosphor-imaging software. Numbers below the autoradiographs of individual genes indicate the ratio of transcription rates between +S and −S cells. The experiment was repeated twice with similar results.
Sugars Alter mRNA Stability
To examine the effect of sugars on the mRNA half-life of individual genes, degradation of mRNA in vivo was monitored by RNA gel-blot analysis following the inhibition of transcription with actinomycin D (Act D). Addition of 10 μg mL−1 Act D to the medium inhibits total RNA transcription by more than 95% over a 12-h time-course (Sheu et al., 1994). To obtain a high starting level of mRNA for detection of changes in mRNA accumulation, cells were cultured with Suc for SU genes (Fig. 4a), or without Suc for SD genes (Fig. 4b), prior to subjecting cells to inhibition of transcription with Act D. Whereas measurement of mRNA half-lives following different growth regimes makes it difficult to directly compare the absolute mRNA half-lives between SU and SD genes, the relative mRNA half-lives of the same genes under starved or non-starved conditions can be compared. Using the latter method, we previously detected the effect of sugars on α-amylase mRNA half-lives (Sheu et al., 1994, 1996) and confirmed the role of αAmy3 3′-UTR on sugar-dependent mRNA stability (Chan and Yu, 1998a, 1998b).
Figure 4.

Sugars alter mRNA stability of sugar-regulated genes. a, Measurement of mRNA half-life of SU genes. Rice suspension cells were cultured in Suc-free (−S) medium for 24 h (RNA in lane 1) or in Suc-containing (+S) medium for 24 h to increase the starting RNA level (RNA in lane 2). ActD was then added to the +S medium to a final concentration of 10 μg/mL and cells were incubated for another 12 h and then divided in half. One-half of the cells were transferred to a +S medium containing ActD (RNAs in lanes 3–11). The other one-half were transferred to a −S medium containing ActD (RNAs in lanes 12–20). Cells were incubated and collected from 0 to 12 h and RNA was purified. RNA gel-blot analysis was performed using probes of SU genes. Ten micrograms of total RNA were loaded in each lane. Three membrane blots were parallel prepared. The Act, SSP2 and rRNA probes, ADH2 and HSP86 probes, and G3PD and His probes were respectively hybridized to each of the three blots. b, Measurement of mRNA half life of SD genes. Rice suspension cells were cultured in +S medium for 24 h (RNA in lane 1) and transferred to −S medium for 24 h to increase the starting RNA level (RNA in lane 2). ActD was then added to −S medium to a final concentration of 10 μg/mL and cells were incubated for another 12 h and then divided in half. One-half of the cells were transferred to a +S medium containing ActD (RNAs in lanes 5–12). The other one-half were transferred to a −S medium containing ActD (RNAs in lanes 13–19). Cells were incubated and collected from 0 to 9 h and RNA was purified. RNA gel-blot analysis was performed using probes of SD genes. Two membrane blots were parallel prepared and sequentially hybridized with these probes. The α-Amy probe hybridized to one blot and the rest of the probes hybridized to another blot. Ten micrograms of total RNA were loaded in each lane. Lanes 3 and 4, Cells were incubated in −S medium lacking ActD for 36 and 45 h, respectively, to serve as controls.
In the present study, mRNA levels of all the SU genes were relatively low in Suc-starved cells (Fig. 4a, lane 1) but high in Suc-provided cells (Fig. 4a, lane 2). After cells were pretreated with Act D plus Suc for 12 h, levels of Act, ADH2, G3PD, HSP86, and SSP2 mRNA decreased more slowly in cells provided with ActD plus Suc (Fig. 4a, lanes 3–11) than in cells provided with ActD minus Suc (Fig. 4a, lanes 12–20) during the next 12 h. The amounts of mRNA (Fig. 4a) were quantified using phosphor-imaging software (PhosphorImager, Molecular Dynamics, Sunnyvale, CA) and mRNA half-lives were determined (Table I). The half-lives of Act, ADH2, G3PD, and SSP2 mRNAs were approximately 1.6- to 2.6-fold longer in Suc-provided cells than in Suc-starved cells. The half-lives of His and HSP86 mRNAs were not significantly altered by varying Suc levels.
Table I.
mRNA half lives of Suc up-regulated genes
| Gene/Enzyme | Half Life (h)
|
Fold of Change in Half Life (+S/−S) | |
|---|---|---|---|
| +S | −S | ||
| Act | 13.2 | 5.0 | 2.6 |
| ADH2 | 9.9 | 5.5 | 1.8 |
| His | 14.0 | 12.0 | 1.2 |
| G3PD | 14.6 | 9.2 | 1.6 |
| HSP86 | 10.6 | 9.1 | 1.2 |
| SSP2 | 9.2 | 4.9 | 1.9 |
For all the tested SD genes, the mRNA levels were relatively low in Suc-provided cells (Fig. 4b, lane 1) and significantly higher in Suc-starved cells (Fig. 4b, lanes 2–4). After cells were pretreated with Act D minus Suc for 12 h, the mRNA levels decreased more rapidly in cells provided with ActD plus Suc (Fig. 4b, lanes 5–12) than in cells provided with ActD minus Suc (Fig. 4b, lanes 13–19) during the next 9 h. The half-life of α-Amy mRNA was 11.0-fold longer in Suc-starved cells than in Suc-provided cells (Table II), which was similar to our previous findings (Sheu et al., 1994). The half-lives of other mRNAs were 2.5- to 7.4-fold longer in Suc-starved cells than in Suc-provided cells (Table II). The above results demonstrate that mRNA stability plays an important role in the expression of some SU and all SD genes.
Table II.
mRNA half lives of Suc down-regulated genes
| Gene/Enzyme | Half Life (h)
|
Fold of Change in Half Life (−S/+S) | |
|---|---|---|---|
| −S | +S | ||
| α-Amy | 13.2 | 1.2 | 11.0 |
| A2 | 10.2 | 3.4 | 3.0 |
| A3 | 32.0 | 4.3 | 7.4 |
| B1 | 23.6 | 5.2 | 4.5 |
| D1 | 12.2 | 4.8 | 2.5 |
| E1 | 14.2 | 2.9 | 4.9 |
| F1 | 11.9 | 4.0 | 3.0 |
Phosphorylation of Hexose Activates or Suppresses Sugar-Regulated Gene Expression
We previously showed that expression of αamylase genes in rice suspension cells was suppressed by the presence of Glc, Fru, or Suc in the culture medium, and Suc was hydrolyzed to Glc and Fru by the cell wall invertase prior to uptake by rice cells (Yu et al., 1991). The phosphorylation of hexose sugars by hexokinase has been shown to be critical for repression of photosynthetic gene expression in maize protoplasts (Jang and Sheen, 1994) and glyoxylate cycle gene expression in cucumber cell culture (Graham et al., 1994). To investigate whether phosphorylation of hexose sugars activates or suppresses sugar-regulated gene expression, we conducted experiments using Glc analogs. Rice suspension cells were cultured in 3-O-methyl-Glc (3-OMG) and 6-deoxy-Glc (6-dG), which are taken up by cells but are not phosphorylated by hexokinase, and also 2-deoxy-Glc (2-dG), which is phosphorylated by hexokinase but is not further metabolized by cells (Dixon and Webb, 1979). Application of 2-dG at various effective concentrations (as indicated by suppression of α-amylase gene expression) between 0.1 to 0.5 mm, which were below the commonly used concentrations for plant cells (Graham et al., 1994; Jang and Sheen, 1994), resulted in a toxic effect on cell growth (as indicated by abnormal morphology of suspension cells and increased degradation of rRNA) within 12 h. The data of treatment with 2-dG were therefore not presented in this report. The effects of 40 mm each of Suc, Glc, 3-OMG, and 6-dG on steady-state mRNA levels of SU and SD genes are shown in Figure 5. mRNA levels of all the SU genes were relatively high in cells grown in medium containing Suc or Glc (Fig. 5a, lanes 2–5), but were significantly lower in cells grown in medium lacking sugar or containing 3-OMG or 6-dG for 12 h (Fig. 5a, lanes 7, 9, and 11). In contrast, mRNA levels of all the SD genes as well as αAmy3 were relatively low or barely detectable in cells grown in medium containing Suc or Glc (Fig. 5b, lanes 2–5) but were significantly higher in cells grown in medium lacking sugar or containing 3-OMG or 6-dG for 12 h (Fig. 5b, lanes 7, 9, and 11).
Figure 5.

Non-phosphorylatable Glc analogs do not activate or repress sugar-responsive gene expression. Rice suspension cells were cultured in Suc-containing medium for 5 d and transferred to medium containing 40 mm Suc (S), 40 mm Glc (G), no sugar (−), 40 mm 3-OMG, or 10 mm 6-dG for 2 and 12 h. Total RNA was purified and subjected to RNA gel-blot analysis. a, SU genes as probes. b, SD genes as probes. Four membrane blots were parallel prepared. The SSP2, rDNA and E1 probes, ADH2, HSP86, αAmy3, and D1 probes, Act, G3PD, A3, and B1 probes, A2 and F1 probes were respectively hybridized to each of the four blots.
Because 2-dG was toxic to plant cells, rice suspension cells were cultured in another Glc analog, Man. Upon entry into the plant cell, Man is phosphorylated by hexokinase, yielding Man-6-P. Man-6-P is then slowly processed by plant cells, since the enzymes required for this process are either absent or exist in very low concentrations (Walder and Sivak, 1986). Man has been shown to inhibit germination of Arabidopsis seeds due to phosphorylation by hexokinase and not due to ATP or P depletion (Peto et al., 1999). As shown in Figure 6, the effect of Glc on mRNA accumulation of SU and SD genes was similar as in Figure 5. Man slightly activated the expression of ADH2, G3PD, and HSP86 genes but not that of Act and SSP2 genes in sugar-starved cells (Fig. 6a, compare lane 4 with lane 6). In contrast, Man suppressed the expression of αAmy3, αAmy8, and A3 but not that of A2, B1, D1, E1, and F1 in sugar-starved cells (Fig. 6b, compare lane 4 with lane 6).
Figure 6.
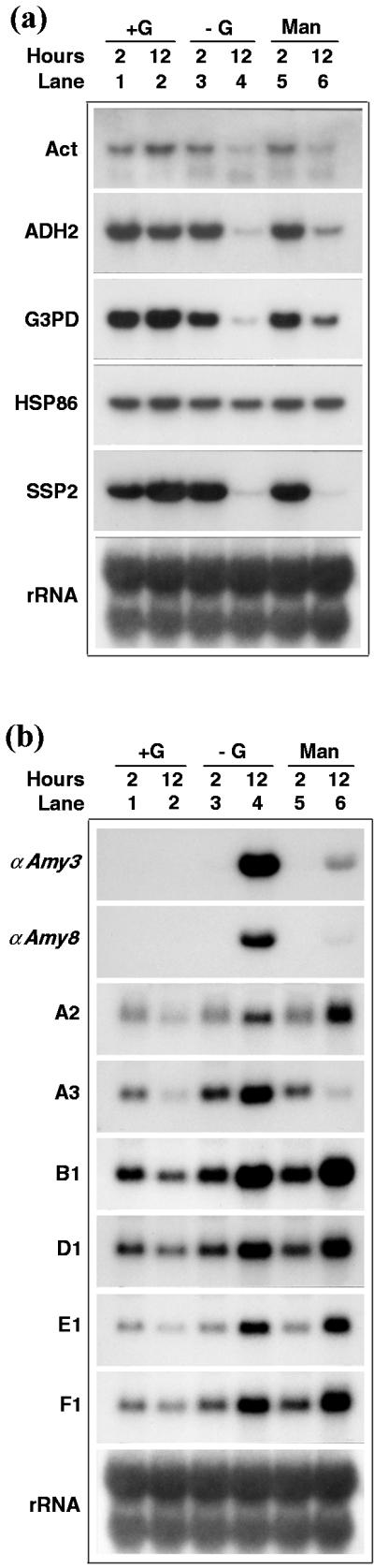
Phosphorylatable but slowly metabolized Glc analog activates or represses certain gene expression. Rice suspension cells were cultured in Suc-containing medium for 5 d and transferred to medium containing 40 mm Glc (+G), no Glc (−G), or 2 mm Man for 2 and 12 h. Total RNA was purified and subjected to RNA gel-blot analysis. a, SU genes as probes. b, SD genes as probes. Six membrane blots were parallel prepared. The Act, HSP86 and rDNA probes, ADH2, G3PD, and SSP2 probes, αAmy3 and D1 probes, αAmy8 and B1 probes, A3 and F1 probes, and A2 probe, were respectively hybridized to each of the six blots.
The above results demonstrate that Glc by itself or Glc and Fru hydrolyzed from Suc all can activate the expression of SU genes and suppress the expression of SD genes. In contrast, the Glc analogs, 3-OMG and 6-dG, which cannot be phosphorylated and metabolized by cells, do not activate the expression of SU genes or suppress the expression of SD genes. The Glc analog, Man, which can be phosphorylated but slow metabolized by cells, partially activates the expression of only certain SU genes. Man behaves in a mixed fashion with respect to SD genes by suppressing or activating expression of different SD genes.
The Metabolic Intermediate Pyruvate Suppresses Gene Expression
Pyruvate has been shown to prevent autophage in carbohydrate-starved sycamore cells, and since pyruvate is the main mitochondrial substrate derived from glycolysis, it was suggested that pyruvate prevents autophage by supplying the mitochondria with a respiratory substrate (Aubert et al., 1996). Pyruvate was tested for its ability to activate or suppress sugar-regulated gene expression. Pyruvate did not activate the expression of all the SU genes in Suc-starved cells (Fig. 7a, compare lane 4 with lane 8), however, it enhanced the expression of G3PD and SSP2 in Suc-provided cells (Fig. 7a, compare lane 2 with lane 6). On the other hand, pyruvate suppressed the expression of all the SD genes, except αAmy3 and A3, in Suc-starved cell (Fig. 7b, compare lane 4 with lane 8). It is interesting that pyruvate enhanced the expression of αAmy3 and A3 in Suc-starved cells (Fig. 7b, compare lane 4 with lane 8). These results demonstrate that expression of certain SD genes can be suppressed by the metabolic intermediate pyruvate.
Figure 7.

Pyruvate represses SD gene expression. Rice suspension cells were cultured in Suc-containing (+S) medium for 5 d and transferred to +S medium or Suc-free (−S) medium with or without 50 mm pyruvate. Total RNA was purified and subjected to RNA gel-blot analysis. a, SU genes as probes. b, SD genes as probes. Six membrane-blots were parallel prepared. The HSP86, αAmy3 and αAmy8 probes, SSP2 and A3 probes, Act and F1 probes, G3PD, B1, and E1 probes, rRNA and D1 probes, ADH2 and A2 probes were respectively hybridized to each of the six blots.
DISCUSSION
Growth-Related Genes Are Positively But May or May Not Be Coordinately Regulated by Sugars
In the present study, preferential expression of growth-related genes in Suc-provided growing cells was as expected (Fig. 1). The parallel change in levels of α-Amy, Act, and SSP2 proteins (Fig. 2) with their mRNAs in response to sugar availability suggests that these sugar-responsive proteins may be physiologically relevant. The expression of most SU genes is coordinately up-regulated by sugars. However, expression of His, HSP86, and Ubi genes may also be non-coordinately regulated, probably depending on the special needs of different members of these protein families under various growth conditions. The present study did not focus on monitoring the expression of various genes in response to varying sugar levels based on gene-specific probes; a single gene or distinct groups of genes may contribute to the overall expression pattern. Histones are required in large quantities at the time of cell division, and in many eukaryotic systems histones are synthesized primarily during the S phase of the cell cycle (Gething and Sambrook, 1992). The quick and dramatic but transient increase in the expression of His 1 h after shifting cells from −S to +S medium (Fig. 1a, lane 15) suggests a burst in DNA replication after resumption of sugar supply. Heat shock proteins (HSPs) are assumed to participate in the maintenance of cellular structure during heat stress as well as to act as molecular chaperones in normally growing cells (Viestra, 1996). This may explain the phenomenon that the expression of HSPs was high in Suc-provided cells (Fig. 1a, lanes 1–4), decreased initially after Suc starvation (Fig. 1a, lanes 5–9), but increased again at later stages of starvation (Fig. 1a, lanes 10–14). Ubiquitin is responsible for cellular housekeeping as well as stress response by removing abnormal or mis-folded proteins and recycling amino acids (Mikami and Iwabuchi, 1993). This may explain the contrasting expression patterns of different members of ubiquitin genes in response to the availability of sugars (Fig. 1a).
Stress-Related Genes Are Coordinately and Negatively Regulated by Sugars
The Suc-repressible genes identified in this study, except for α-amylase genes, all encode stress-related proteins and represent a group of genes distinct from those previously identified as being involved in photosynthesis and reserve remobilization and catabolism (Koch, 1996). cDNA B1 encodes a Gly-rich protein (GRP) that shows homology to RNA-binding proteins containing the ribonucleoprotein consensus sequence. A class of RNA-binding GRP genes responsive to wounding and cold were identified in plants (Sturm, 1992; Horvath and Olson, 1998). cDNAs A2, D1, E1, and F1 encode GRPs that show homology to a group of Gly-rich cell wall or periplasmic space-associated proteins (Fang et al., 1991; Lei and Wu, 1991). GRPs are widely distributed in plants and are developmentally regulated (Cheng et al., 1996) and responsive to wounding, virus infection, drought, and flooding (Condit and Meagher, 1987; Gómez et al., 1988; de Oliveira et al., 1990; Fang et al., 1991). cDNA A3 is almost identical to the salt-responsive rice gene salT with only minor nucleotide variation. Expression of salT is significantly induced in sheaths and roots after plants are osmotically challenged (Claes et al., 1990) and in cultured rice suspension cells by abscisic acid and NaCl (Garcia et al., 1998).
Suc starvation is a type of nutritional stress. Coordinated expression of GRP genes and salT in Suc-starved cells suggests that these genes may play roles in protecting cells against nutritional stress. In addition, inducibility of expression of these genes in rice cell culture by sugar starvation also suggests that different stress conditions might trigger a general response by creating the same intracellular signal. The inducibility of expression of various stress-related genes in a rice cell culture by sugar starvation makes the rice cell culture an ideal model system for studying the signal transduction pathway and regulatory mechanism of plant stress response.
Dual Regulation by Sugars Operates at Transcriptional and Posttranscriptional Levels
We found that sugars not only regulates the transcription but also alters the stability of mRNAs. Among the six SU genes tested, under the experimental condition in which the transcription rate was measured and compared 24 h after cell culture with or without Suc, an increase in transcription rate in Suc-provided cells was observed only for ADH2 and G3PD (Fig. 3a). At the same time, accumulation of HSP86 mRNA in Suc-starved cells began to rise (Fig. 1a, lane 11). This may explain why at this time point the transcription rate of HSP86 gene was higher in Suc-starved cells than in Suc-provided cells (Fig. 1a, slot 2). Since the mRNA half-life of HSP86 mRNA was not significantly altered by Suc level (Table I), transcriptional activity may play a more dominant role than mRNA stability in the accumulation of HSP86 mRNA. The transcription rate of the SSP2 gene was similar in either Suc-provided or Suc-starved cells and the transcription rate of the Act gene was lower in Suc-provided cells than in Suc-starved cells (Fig. 3a). These findings suggest that mRNA stability may play a more dominant role than transcriptional activity in accumulation of these two mRNAs in Suc-provided cells. This notion is supported by the observation that the half lives of Act and SSP2 mRNAs are 2- to 3-fold longer in Suc-provided cells than in Suc-starved cells (Table I). The massive and transient accumulation of His mRNA 1 h after shifting cells from −S to +S medium (Fig. 1a, lane 15) suggests a sugar hypersensitive but short half-lived increase in transcriptional activity and/or mRNA stability coupled with His DNA synthesis during a short time window. The half-lives of ADH2 and G3PD are 1.6- to 1.8-fold longer in Suc-provided cells than in Suc-starved cells. Therefore, in the present study, sugar up-regulation of transcription and mRNA stability are coupled only for ADH2, G3PD, and His. On the other hand, sugar down-regulation of transcription and mRNA stability are coupled for all of the SD genes.
Sugars Regulate Gene Expression through Common Control Mechanisms
The coordinated activation or repression of sugar-responsive genes suggests that common mechanisms might be responsible for regulation of these genes. The mechanisms would include interaction between transacting factors (transcription or RNA degradation factors) and cis-regulatory elements (promoter or mRNA sequences) of the individual genes. Specific cis-regulatory elements in the promoters or mRNAs are probably shared by these sugar-regulated genes. The SRC of αAmy3 promoter contains a TATCCA element that has been shown to serve as an enhancer for sugar starvation-induced expression (Lu et al., 1998). The TATCCA element along with its variants are present at a proximity upstream of the transcription start sites of 18 α-amylase genes isolated from various plant species (Yu, 1999b) and other sugar-repressible genes, e.g. the cucumber malate synthase and isocitrate lyase genes (Graham et al., 1989; Reynolds and Smith, 1995) and the maize Suc synthase gene (Sh1) (Koch et al., 1992). The promoter region of Sh1 containing the TATCCA element also confers sugar inhibition of downstream reporter gene expression (Maas et al., 1990). These findings suggests that the TATCCA element or its variants could be a common cis-regulatory element responsible for sugar repression. Computer analysis found that nucleotide sequences similar to the essential motifs in the αAmy3 3′-UTR for sugar-dependent mRNA stability (Chan and Yu, 1998a, 1998b) were also present in the 3′-UTRs of the SD genes examined in the present study. Whether these mRNA sequences are functional remain to be determined. Promoter analysis of the sugar up-regulated patatin class-I gene identified two separate cis-sequences responsible for sugar induction (Grierson et al., 1994). The cis-sequences contain two conserved 9-bp Suc response elements (SURE), which are similar to some motifs present in the promoters of some SU genes, e.g. the β-amylase and sporamin genes of sweet potato (Ohta et al., 1991; Ishiguro and Nakamura, 1992) and the proteinase inhibitor II gene of potato (Kim et al., 1991). In future studies identification of genes encoding the transacting transcription factors would allow us to understand whether a common or multiple transcription factors interact with the cis-regulatory elements and are involved in the coordinated transcription of SU or SD genes.
Hexokinase-Dependent and -Independent Pathways Regulate Gene Expression and the Sugar Signal Transduction Pathway May Overlap the Glycolytic Pathway for Gene Repression
Various glycolytic intermediates downstream of Glc have been shown to have no repressive effect on photosynthetic gene expression (Jang and Sheen, 1994). In the present study, pyruvate, which is produced at the final step of glycolysis, suppressed the expression of all the SD genes except αAmy3 and A3 in Suc-starved cells. This study provides the first evidence that the sugar signal transduction pathway may overlap the downstream sugar metabolic pathway for gene regulation.
Recent study has indicated that sugar signaling in plants occurs by hexokinase-dependent pathway, by hexose-dependent but hexokinase-independent pathway, and by Suc-dependent pathway (Jang and Sheen, 1997; Smeekens and Rook, 1997; Halford et al., 1999; Sheen et al., 1999; Smeekens, 2000). Our observation is that Glc and Suc are equally effective in up- and down-regulation of SU and SD gene expression, respectively, suggesting that in the present study the first two pathways may play more important roles in the activation and repression of the SU and SD genes, respectively. Man slightly activates the expression of ADH2, G3PD, and HSP86 genes but not that of Act and SSP2 genes (Fig. 6a). Pyruvate did not activate the expression of all the SU genes (Fig. 7a). These results suggest that hexokinase may play a signaling role in activation of ADH2, G3PD, and HSP86 gene expression, and hexose-dependent but hexokinase-independent pathway activates the expression of Act and SSP2 genes. On the other hand, Man suppressed the expression of αAmy3, αAmy8, and A3 but not that of A2, B1, D1, E1, and F1 (Fig. 6b). Pyruvate suppresses the expression of αAmy8, A2, B1, D1, E1, and F1 but not that of αAmy3 and A3. These results suggest that hexokinase may play a signaling role in suppression of αAmy3, αAmy8, and A3, and the metabolic pathway delivers signal to suppress expression of αAmy8, A2, B1, D1, E1, and F1. Figure 8a summarizes the results of Figures 6 and 7 and shows that although coordinately regulated by sugars, different genes are up-regulated or down-regulated through hexokinase-dependent and/or hexokinase-independent pathways. In addition, the sugar signal transduction pathway may overlap the glycolytic pathway for gene repression.
Figure 8.

Complex sugar signal transduction networks for differential and coordinated regulation of gene expression. a, Signal transduction in sugar-provided cells. Suc may be sensed by the receptor directly, or it may be converted to Glc and Fru and sensed by hexokinase-dependent or hexokinase-independent pathway. The sugar signal transduction pathway may overlap or be independent of the sugar metabolic pathway. b, Signal transduction in Suc-starved cells. The signals of sugar starvation and environmental stresses may be perceived by the same receptor or by different receptors but the signal pathways converge downstream. R, Receptor; INV, invertase. The solid arrows indicate pathways proposed based upon currently available information. +, −; Positive and negative regulation, respectively.
Convergence of Nutritional and Environmental Stress Signal Transduction Pathways
Complex regulatory circuitries may also link the sugar starvation signal to the stress signals (Fig. 8b) for differential and coordinated regulation of gene expression. In sugar-starved cells, as expression of α-amylase and stress-related genes are coordinately regulated, the signals of sugar starvation and environmental stresses may be perceived by the same receptor, or by different receptors while the signal pathways converge downstream (Fig. 8b). The latter pathways have been proposed in a study on sugar- and stress-regulated sink-specific and defense-related gene expression in Chenopodium rubrum (Ehness et al., 1997). Under either condition, the exact points where the stimulating and inhibitory pathways diverge from the same signal transduction chains are not known. Whether the signals of sugar and sugar starvation are transduced through independent pathways or through the opposite action of the same components in the signal transduction chain is also not known.
Although in the present study the stress-related genes are randomly selected and the mode of their regulation by sugars may not be applied to all stress-related gene, the requirement of convergence of sugar starvation and stress signal transduction pathways could be of physiological significance. In stress situations, cells have a high demand for sugars to fulfil the energy and carbon requirements needed for appropriate response to stresses. This may explain why expression of α-amylases, which hydrolyze starch to provide sugars for metabolic activity, couples with the expression of stress-related proteins. The notion is supported by observations that expression of α-amylases is induced in barley leaves by water stress (Jacobsen et al., 1986), in tobacco leaves by virus infection (Heitz et al., 1991), and in mung bean cotyledons by wounding (Koizuka et al., 1995). Similar phenomena are found for other hydrolytic enzymes. For example, expression of β-fructosidase (invertase), which hydrolyzes stored Suc to hexose, is enhanced in carrot roots in response to wounding or pathogen infection (Sturm and Chrispeels, 1990). The increase in expression of these carbohydrate hydrolytic enzymes suggests a role of these enzymes in defense responses to biotic or abiotic stress.
Among the SD genes, A2, B1, D1, E1, and F1 seem to be controlled by sugar via a similar or identical pathway, whereas A3 is controlled via somewhat different pathways (Fig. 8a). The notion that expression of αAmy3 and αAmy8 are also differentially regulated via somewhat different pathways is supported by the observation that pyruvate suppressed the expression of αAmy8 but not αAmy3 (Figs. 7b and 8a).
In conclusion, sugars can modulate gene expression at the transcriptional and posttranscriptional levels through a complex signal transduction network and some common mechanisms. Additional work is necessary to identify the various components of the signal transduction chains and to explore how the interaction between the cis-acting elements in the promoters and/or mRNAs with the trans-acting factors are involved in the coordinated but differential regulation of diverse gene expression in response to the sugar status in plant cells. Such future studies should lead to a better understanding of the mechanisms that underlie the global sugar regulation of gene expression in plants.
MATERIALS AND METHODS
Rice (Oryza sativa cv Tainan 5) Cell Culture
A suspension cell culture of rice was established as previously described (Yu et al., 1991). Cells were subcultured every 7 d by transferring approximately 0.5 mL of cells into 25 mL of fresh liquid Murashige and Skoog medium (Murashige and Skoog, 1962) containing 3% (w/v) Suc in a 125-mL flask. Cells were cultured on a reciprocal shaker at 120 rpm and incubated at 26°C in the dark.
Plasmids
Plasmid αAmy8-C carries a 1.4-kb rice α-amylase cDNA insert in pBluescript KS+ (Stratagene, La Jolla, CA) (Yu et al., 1992). Plasmid pcRAc1.3 contains a 1.4-kb rice actin 1 cDNA insert in pBluescript II-KS (McElroy et al., 1990). Plasmid pRY18 carries a 3.8-kb DNA fragment that contains a rice genomic rDNA cluster, including the 3′-half portion of 18S rRNA gene, the complete 5.8S rRNA gene, and the 5′-half portion of the 25S rRNA gene in pUC13 (Sano and Sano, 1990). cDNAs encoding ADH2, G3PD, HSP86, His, SSP2, and Ubi were isolated from rice callus cDNA libraries and inserted in SalI(5′) and NotI(3′) sites of pBluescript SKII+ (Stratagene) by the Japan Rice Genome Research Program (Uchimiya et al., 1992).
RNA Gel-Blot Analysis
Total RNA was purified from rice suspension cells using TRIZOL reagent (Life Technologies/Gibco-BRL, Cleveland). α-32P-labeled DNA probes were prepared and RNA gel-blot analysis was performed as described (Sheu et al., 1996). In cases when the membrane blot was sequentially hybridized with various probes, each probe on the membrane was stripped and rehybridized as described (Sheu et al., 1994). Plasmid DNAs of αAmy8-C and pcRAc1.3 were digested with EcoRI. Plasmid DNAs of Act, ADH2, G3PD, His, HSP86, and SSP2 were digested with SalI and NotI. Plasmid DNAs of A1, A3, B1, D1, E1, and F1 were digested with EcoRI. αAmy3 and αAmy8 gene-specific DNAs were prepared as described (Sheu et al., 1996). The insert DNAs were individually isolated, labeled with 32P, and used as probes. A DNA fragment containing 25S, 18S, and 5.8S rDNAs was excised from pRY18 using BamHI, labeled with α-32P, and used as a probe for equalizing RNA loading.
Differential Screening of cDNA Library
Rice suspension cells were cultured in Suc-containing medium for 5 d and transferred to Suc-containing (+S) or Suc-free (−S) medium for 4 h. Cells were collected and total RNA was purified. Poly(A)+ RNA was further purified using an oligo(dT)-cellulose spin column (5′ → 3′). The poly(A)+ RNA isolated from −S cells was used to construct a cDNA library in λGEM-2 vector (Promega, Madison, WI). The 32P-labeled single-stranded cDNA probe was prepared from poly(A)+ RNA of +S or −S cells using an oligo(dT) primer and avian myeloblastosis virus reverse transcriptase. Duplicated filter lifts from high-density platings of the cDNA library were then differentially screened with the cDNA probes. The phage plaques that hybridized strongly with the cDNA probe of −S cells and only weakly or not at all with the cDNA probe of +S cells were isolated. The cDNA in λGEM-2 was then cleaved with EcoRI, subcloned into the EcoRI site of pBluescript vector, and nucleotide sequenced.
Protein Gel-Blot Analysis
Total proteins were extracted from cultured suspension cells with an extraction buffer (50 mm Tris [tris(hydroxymethyl)aminomethane]-HCl, pH 8.8, 1 mm EDTA, 10% [v/v] glycerol, 1% [v/v] Triton X-100, 10 mm β-mercaptoethanol, and 0.1% [w/v] sarkosyl). The culture medium was collected and centrifuged at 18,000g at 4°C for 15 min to remove cell debris. Protein gel-blot analysis was performed as described (Yu et al., 1991). The anti-α-amylase polyclonal antibodies raised against rice α-amylases were previously produced in our laboratory (Chen et al., 1994). The anti-Suc synthase polyclonal antibodies were raised in rabbit against Suc synthase purified from developing rice grains and were a gift from Ai-Yu Wang (National Taiwan University, Taipei). The mouse anti-actin monoclonal antibody raised against chicken gizzard actin was purchased from Chemicon International, Inc.
ACKNOWLEDGMENTS
We thank Dr. Jun-Jei Sheu and Ms. Lin-Tze Yu for technical assistance and Mr. Douglas Platt for help in preparing the manuscript. We also thank the Japan Rice Genome Research Program for providing us the rice cDNAs.
Footnotes
This work was supported by the Academia Sinica, the National Science Council (grant no. NSC 89–2311–B–001–023), and the Biomedical Research Foundation of the Republic of China.
LITERATURE CITED
- Aubert S, Gout E, Bligny R, Marty-Mazars D, Barrieu F, Alabouvette J, Marty F, Douce R. Ultrastructural and biochemical characterization of autophagy in higher plant cells subjected to carbon deprivation: control by the supply of mitochondria with respiratory substrates. J Cell Biol. 1996;133:1251–1263. doi: 10.1083/jcb.133.6.1251. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Brouquisse R, James F, Pradet A, Raymond P. Asparagine metabolism and nitrogen distribution during protein degradation in sugar-starved maize root tips. Planta. 1992;188:384–395. doi: 10.1007/BF00192806. [DOI] [PubMed] [Google Scholar]
- Carlson M. Regulation of sugar utilization in Saccharomyces species. J Bacteriol. 1987;169:4873–4877. doi: 10.1128/jb.169.11.4873-4877.1987. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Chan M-T, Chao Y-C, Yu S-M. Novel gene expression system for plant cells based on induction of α-amylase promoter by carbohydrate starvation. J Biol Chem. 1994;269:17635–17641. [PubMed] [Google Scholar]
- Chan M-T, Yu S-M. The 3′-untranslated region of a rice α-amylase gene mediates sugar-dependent abundance of mRNA. Plant J. 1998a;15:685–696. doi: 10.1046/j.1365-313x.1998.00244.x. [DOI] [PubMed] [Google Scholar]
- Chan M-T, Yu S-M. The 3′-untranslated region of a rice α-amylase gene functions as a sugar-dependent mRNA stability determinant. Proc Natl Acad Sci USA. 1998b;95:6543–6547. doi: 10.1073/pnas.95.11.6543. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Chen M-H, Liu L-F, Chen Y-R, Wu H-K, Yu S-M. Expression of α-amylases, carbohydrate metabolism, and autophagy in cultured rice cells are coordinately regulated by sugar nutrient. Plant J. 1994;6:625–636. doi: 10.1046/j.1365-313x.1994.6050625.x. [DOI] [PubMed] [Google Scholar]
- Cheng S-H, Keller B, Condit CM. Common occurrence of homologues of petunia glycine-rich protein-1 among plants. Plant Mol Biol. 1996;31:163–168. doi: 10.1007/BF00020616. [DOI] [PubMed] [Google Scholar]
- Cheng W-H, Taliercio EW, Chourey PS. Sugars modulate an unusual mode of control of the cell-wall invertase gene (Incw1) through its 3′-untranslated region in a cell suspension culture of maize. Proc Natl Acad Sci USA. 1999;96:10512–10517. doi: 10.1073/pnas.96.18.10512. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Chevalier C, Bourgeois E, Pradet A, Raymond P. Molecular cloning and characterization of six cDNAs expressed during glucose starvation in excised maize (Zea Mays L.) root tips. Plant Mol Biol. 1995;28:473–485. doi: 10.1007/BF00020395. [DOI] [PubMed] [Google Scholar]
- Claes B, Dekeyser R, Villarroel R, Van den Bulcke M, Bauw G, Van Montagu M, Caplan A. Characterization of a rice gene showing organ-specific expression in response to salt stress and drought. Plant Cell. 1990;2:19–27. doi: 10.1105/tpc.2.1.19. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Condit CM, Meagher RB. Expression of a gene encoding a glycine-rich protein in petunia. Mol Cell Biol. 1987;7:4273–4279. doi: 10.1128/mcb.7.12.4273. [DOI] [PMC free article] [PubMed] [Google Scholar]
- de Oliveira DE, Seurinck J, Inzé D, Van Montagu M, Botterman J. Differential expression of five Arabidopsis genes encoding glycine-rich proteins. Plant Cell. 1990;2:427–436. doi: 10.1105/tpc.2.5.427. [DOI] [PMC free article] [PubMed] [Google Scholar]
- DeWald DB, Sadka A, Mullet JE. Sucrose modulation of soybean Vsp gene expression is inhibited by auxin. Plant Physiol. 1994;104:439–444. doi: 10.1104/pp.104.2.439. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Dixon M, Webb EC, editors. Enzymes. London: Longman Scientific & Technical; 1979. pp. 248–251. [Google Scholar]
- Ehness R, Ecker M, Godt DE, Roitsch T. Glucose and stress independently regulate source and sink metabolism and defense mechanisms via signal transduction pathways involving protein phosphorylation. Plant Cell. 1997;9:1825–1841. doi: 10.1105/tpc.9.10.1825. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Efrat S, Tal M, Lodish HF. The pancreatic β-cell glucose sensor. Trends Biochem Sci. 1994;19:535–538. doi: 10.1016/0968-0004(94)90056-6. [DOI] [PubMed] [Google Scholar]
- Entian K-D, Barnett JA. Regulation of sugar utilization by Saccharomyces cerevisiae. Trends Biochem Sci. 1992;17:506–510. doi: 10.1016/0968-0004(92)90341-6. [DOI] [PubMed] [Google Scholar]
- Fang R-X, Pang Z, Gao D-M, Mang K-Q, Chua N-H. cDNA sequence of a virus-inducible, glycine-rich protein gene from rice. Plant Mol Biol. 1991;17:1255–1257. doi: 10.1007/BF00028742. [DOI] [PubMed] [Google Scholar]
- Gancedo JM. Carbon catabolite repression in yeast. Eur J Biochem. 1992;206:297–313. doi: 10.1111/j.1432-1033.1992.tb16928.x. [DOI] [PubMed] [Google Scholar]
- Garcia AB, de Almeida Engler J, Claes B, Villarroel R, Van Montagu M, Gerats T, Caplan A. The expression of the salt-responsive gene salT from rice is regulated by hormonal and developmental cues. Planta. 1998;207:172–180. doi: 10.1007/s004250050470. [DOI] [PubMed] [Google Scholar]
- Gething MJ, Sambrook J. Protein folding in the cell. Nature. 1992;355:33–45. doi: 10.1038/355033a0. [DOI] [PubMed] [Google Scholar]
- Gómez J, Sanchez-Martinez D, Stiefel R, Rigau J, Puigdomènech P, Pagès M. A gene induced by the plant hormone abscisic acid in response to water stress encodes a glycine-rich protein. Nature. 1988;334:262–264. doi: 10.1038/334262a0. [DOI] [PubMed] [Google Scholar]
- Graham IA, Denby KJ, Leaver CJ. Carbon catabolite repression regulates glyoxylate cycle gene expression in cucumber. Plant Cell. 1994;6:761–772. doi: 10.1105/tpc.6.5.761. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Graham IA, Smith LM, Brown JW, Leaver DJ, Smith SM. The malate synthase gene of cucumber. Plant Mol Biol. 1989;13:673–684. doi: 10.1007/BF00016022. [DOI] [PubMed] [Google Scholar]
- Grierson C, Du J-S, de Torres Zabala M, Beggs K, Smith C, Holdsworth M, Bevan M. Separate cis sequences and trans factors direct metabolic and developmental regulation of a potato tuber storage protein gene. Plant J. 1994;5:815–826. doi: 10.1046/j.1365-313x.1994.5060815.x. [DOI] [PubMed] [Google Scholar]
- Halford NG, Purcell PC, Grahame Hardie D. Is hexokinase really a sugar sensor in plants. Trends Plant Sci. 1999;4:117–120. doi: 10.1016/s1360-1385(99)01377-1. [DOI] [PubMed] [Google Scholar]
- Hattori T, Nakagawa S, Nakamura K. High level expression of tuberous root storage protein genes of sweet potato in stems of plantlets grown in vitro on sucrose medium. Plant Mol Biol. 1990;14:595–604. doi: 10.1007/BF00027505. [DOI] [PubMed] [Google Scholar]
- Heitz T, Gioffroy P, Fritig B, Legrand M. Two apoplastic α-amylases are induced in tobacco by virus infection. Plant Physiol. 1991;97:651–656. doi: 10.1104/pp.97.2.651. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Herbers K, Meuwly P, Frommer WB, Métraux J-P, Sonnewald U. Systemic acquired resistance mediated by the ectopic expression of invertase: possible hexose sensing in the secretory pathway. Plant Cell. 1996;8:793–803. doi: 10.1105/tpc.8.5.793. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Horvath DP, Olson PA. Cloning and characterization of cold-regulated glycine-rich RNA-binding protein genes from leafy spurge (Euphorbia esula L.) and comparison to heterologous genomic clones. Plant Mol Biol. 1998;38:531–538. doi: 10.1023/a:1006050208670. [DOI] [PubMed] [Google Scholar]
- Ishiguro S, Nakamura K. The nuclear factor SP8BF binds to the 5′-upstream regions of three different genes coding for major proteins of sweet potato tuberous roots. Plant Mol Biol. 1992;14:995–1006. doi: 10.1007/BF00018460. [DOI] [PubMed] [Google Scholar]
- Jacobsen JV, Hanson AD, Chandler PC. Water stress enhances expression of an α-amylase gene in barley leaves. Plant Physiol. 1986;80:350–359. doi: 10.1104/pp.80.2.350. [DOI] [PMC free article] [PubMed] [Google Scholar]
- James F, Brouquisse R, Pradet A, Raymond P. Changes in proteolytic activities in glucose-starved maize root tips: regulation by sugars. Plant Physiol Biochem. 1993;31:845–856. [Google Scholar]
- Jang J-C, Leó P, Zhou L, Sheen J. Hexokinase as a sugar sensor in higher plants. Plant Cell. 1997;9:5–19. doi: 10.1105/tpc.9.1.5. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Jang J-C, Sheen J. Sugar sensing in higher plants. Plant Cell. 1994;6:1665–1679. doi: 10.1105/tpc.6.11.1665. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Jang J-C, Sheen J. Sugar sensing in higher plants. Trends Plant Sci. 1997;2:208–213. [Google Scholar]
- Jefferson R, Goldsbrough A, Bevan M. Transcriptional regulation of a patatin-1 gene in potato. Plant Mol Biol. 1990;14:995–1006. doi: 10.1007/BF00019396. [DOI] [PubMed] [Google Scholar]
- Journet EP, Bligny R, Douce R. Biochemical changes during sucrose deprivation in higher plant cells. J Biol Chem. 1986;261:3193–3199. [PubMed] [Google Scholar]
- Kim S-R, Costa MA, An G. Sugar response element enhances wound response of potato proteinase inhibitor II in transgenic tobacco. Plant Mol Biol. 1991;17:973–983. doi: 10.1007/BF00037137. [DOI] [PubMed] [Google Scholar]
- Koch KE. Carbohydrate-modulated gene expression in plants. Annu Rev Plant Physiol Plant Mol Biol. 1996;47:509–540. doi: 10.1146/annurev.arplant.47.1.509. [DOI] [PubMed] [Google Scholar]
- Koch KE, Nolte KD, Duke ER, McCarty DR, Avigne WT. Sugar levels modulate differential expression of maize sucrose synthase genes. Plant Cell. 1992;4:59–69. doi: 10.1105/tpc.4.1.59. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Koizuka N, Tanaka Y, Morohashi Y. Expression of α-amylase in response to wounding in mung bean. Planta. 1995;195:530–534. [Google Scholar]
- Lei M, Wu R. A novel glycine-rich cell wall protein gene in rice. Plant Mol Biol. 1991;16:187–198. doi: 10.1007/BF00020551. [DOI] [PubMed] [Google Scholar]
- Lu C-A, Lim E-K, Yu S-M. Sugar response sequence in the promoter of a rice α-amylase gene serves as a transcriptional enhancer. J Biol Chem. 1998;273:10120–10131. doi: 10.1074/jbc.273.17.10120. [DOI] [PubMed] [Google Scholar]
- Maas C, Schaal S, Werr W. A feedback control element near the transcription start site of the maize Shrunken gene determines promoter activity. EMBO J. 1990;9:3447–3452. doi: 10.1002/j.1460-2075.1990.tb07552.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Macknight R, Lister C, Dean C. Rice cDNA clones OsGRP1 and OsGRP2 define two classes of glycine-rich RNA binding proteins. Plant Physiol. 1998;117:1527–1527. [Google Scholar]
- McElroy D, Rothenberg M, Wu R. Structural characterization of a rice actin gene. Plant Mol Biol. 1990;14:163–171. doi: 10.1007/BF00018557. [DOI] [PubMed] [Google Scholar]
- Mikami K, Iwabuchi M. Regulation of cell cycle-dependent gene expression. In: Verma DPS, editor. Control of Plant Gene Expression. Boca Raton, FL: CRC Press; 1993. pp. 51–68. [Google Scholar]
- Müller-Röber BT, Koβmann J, Hannah LC, Willmitzer L, Sonnewald U. One of two different ADP-glucose pyrophosphorylase genes from potato responds strongly to elevated levels of sucrose. Mol Gen Genet. 1990;224:136–146. doi: 10.1007/BF00259460. [DOI] [PubMed] [Google Scholar]
- Murashige T, Skoog F. A revised medium for rapid growth and bioassays with tobacco tissue cultures. Physiol Plant. 1962;15:473–497. [Google Scholar]
- Ohta S, Hattori T, Morikami A, Nakamura K. High-level expression of a sweet potato sporamin gene promoter: β-glucuronidase (GUS) fusion gene in the stems of transgenic tobacco plants is conferred by multiple cell type-specific regulatory elements. Mol Gen Genet. 1991;225:369–378. doi: 10.1007/BF00261676. [DOI] [PubMed] [Google Scholar]
- Peto JV, Weisbeek PJ, Smeekens SCM. Mannose inhibits Arabidopsis germination via a hexokinase-mediated step. Plant Physiol. 1999;119:1017–1023. doi: 10.1104/pp.119.3.1017. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Purcell PC, Smith AM, Halford NG. Antisense expression of a sucrose non-fermenting-1-related protein kinase sequence in potato results in decreased expression of sucrose synthase in tubers and loss of sucrose-inducibility of sucrose synthase transcripts in leaves. Plant J. 1998;14:195–202. [Google Scholar]
- Reynolds SJ, Smith SM. The isocitrate lyase gene of cucumber: isolation, characterization and expression in cotyledons following seed germination. Plant Mol Biol. 1995;27:487–497. doi: 10.1007/BF00019316. [DOI] [PubMed] [Google Scholar]
- Roitsch T, Bittner M, Godt DE. Induction of apoplastic invertase of Chenopodium rubrum by d-glucose and a glucose analog and tissue-specific expression suggest a role in sink-source regulation. Plant Physiol. 1995;108:285–294. doi: 10.1104/pp.108.1.285. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Ronne H. Glucose repression in fungi. Trends Genet. 1995;11:12–17. doi: 10.1016/s0168-9525(00)88980-5. [DOI] [PubMed] [Google Scholar]
- Rose M, Albig W, Entian KD. Glucose repression in Saccharomyces cerevisiae is directly associated with hexose phosphorylation by hexokinase PI and PII. Eur J Biochem. 1991;199:511–518. doi: 10.1111/j.1432-1033.1991.tb16149.x. [DOI] [PubMed] [Google Scholar]
- Sadka A, DeWald DB, May GD, Park WD, Mullet JE. Phosphate modulates transcription of soybean VspB and other sugar-inducible genes. Plant Cell. 1994;6:737–749. doi: 10.1105/tpc.6.5.737. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Salanoubat M, Belliard G. The steady-state level of potato sucrose synthase mRNA is dependent on wounding, anaerobiosis and sucrose concentration. Gene. 1989;84:181–185. doi: 10.1016/0378-1119(89)90153-4. [DOI] [PubMed] [Google Scholar]
- Sano Y, Sano R. Variation of the intergenic spacer region of ribosomal DNA in cultivated and wild rice species. Genome. 1990;33:209–218. [Google Scholar]
- Sheen J. Metabolic repression of transcription in higher plants. Plant Cell. 1990;2:1027–1038. doi: 10.1105/tpc.2.10.1027. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Sheen J, Zhou L, Jang J-C. Sugars as signaling molecules. Curr Opin Plant Biol. 1999;2:410–418. doi: 10.1016/s1369-5266(99)00014-x. [DOI] [PubMed] [Google Scholar]
- Sheu J-J, Jan S-P, Lee H-T, Yu S-M. Control of transcription and mRNA turnover as mechanisms of metabolic repression of α-amylase gene expression. Plant J. 1994;5:655–664. [Google Scholar]
- Sheu J-J, Yu T-S, Tong W-F, Yu S-M. Carbohydrate starvation stimulates differential expression of rice α-amylase genes that is modulated through complicated transcriptional and posttranscriptional processes. J Biol Chem. 1996;271:26998–27004. doi: 10.1074/jbc.271.43.26998. [DOI] [PubMed] [Google Scholar]
- Smeekens S. Sugar-induced signal transduction in plants. Annu Rev Plant Physiol Plant Mol Biol. 2000;51:49–81. doi: 10.1146/annurev.arplant.51.1.49. [DOI] [PubMed] [Google Scholar]
- Smeekens S, Rook F. Sugar sensing and sugar-mediated signal transduction in plants. Plant Physiol. 1997;115:7–13. doi: 10.1104/pp.115.1.7. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Sturm A. A wound-inducible glycine-rich protein from Daucus carota with homology to single-stranded nucleic acid-binding proteins. Plant Physiol. 1992;99:1689–1692. doi: 10.1104/pp.99.4.1689. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Sturm A, Chrispeels MJ. cDNA cloning of carrot extracellular β-fructosidase and its expression in response to wounding and bacterial infection. Plant Cell. 1990;2:1107–1119. doi: 10.1105/tpc.2.11.1107. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Uchimiya H, Kidou S-I, Shimazaki T, Aotsuka S, Takamatsu S, Nishi R, Hashimoto H, Matsubayashi Y, Kidou N, Umeda M, Kato A. Random sequencing of cDNA libraries reveals a variety of expressed genes in cultured cells of rice (Oryza sativa L.) Plant J. 1992;2:1005–1009. [Google Scholar]
- Viestra RD. Proteolysis in plants: mechanisms and functions. Plant Mol Biol. 1996;32:275–302. doi: 10.1007/BF00039386. [DOI] [PubMed] [Google Scholar]
- Walker DA, Sivak MN. Photosynthesis and phosphate: a cellular affair? Trends Biochem Sci. 1986;11:176–179. [Google Scholar]
- Yu S-M. Cellular and genetic responses of plants to sugar starvation. Plant Physiol. 1999a;121:687–693. doi: 10.1104/pp.121.3.687. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Yu S-M. Regulation of α-amylase gene expression. In: Shimamoto K, editor. Molecular Biology of Rice, Chapter 9. Tokyo: Springer-Verlag; 1999b. pp. 161–178. [Google Scholar]
- Yu S-M, Kuo Y-H, Sheu G, Sheu Y-J, Liu L-F. Metabolic derepression of α-amylase gene expression in suspension-cultured cells of rice. J Biol Chem. 1991;266:21131–21137. [PubMed] [Google Scholar]
- Yu S-M, Lee Y-C, Fang S-C, Chan M-T, Hwa S-F, Liu L-F. Sugars act as signal molecules and osmotica to regulate the expression of α-amylase genes and metabolic activities in germinating cereal grains. Plant Mol Biol. 1996;30:1277–1289. doi: 10.1007/BF00019558. [DOI] [PubMed] [Google Scholar]
- Yu S-M, Tzou W-S, Lo W-S, Kuo Y-H, Lee H-T, Wu R. Regulation of α-amylase-encoding gene expression in germinating seeds and cultured cells of rice. Gene. 1992;122:247–253. doi: 10.1016/0378-1119(92)90212-8. [DOI] [PubMed] [Google Scholar]
- Zhou L, Jang J-C, Jones TL, Sheen J. Glucose and ethylene signal transduction crosstalk revealed by an Arabidopsis glucose-insensitive mutant. Proc Natl Acad Sci USA. 1998;95:10294–10299. doi: 10.1073/pnas.95.17.10294. [DOI] [PMC free article] [PubMed] [Google Scholar]