

SUMMARY
5‐HT1A receptors have long been implicated in the pathogenesis and treatment of anxiety and depressive disorders. Recently, several lines of studies have revealed new insights into the therapeutic role of 5‐HT1A receptors in treating schizophrenia and Parkinson's disease. Specifically, 5‐HT1A receptors seem to be a promising target for alleviating antipsychotic‐induced extrapyramidal side effects (EPS) and cognitive/affective disorders in schizophrenia. In the treatment of patients with Parkinson's disease, 5‐HT1A agonists are expected to improve not only affective symptoms (e.g., anxiety and depression), but also the core parkinsonian symptoms as well as antiparkinsonian agents‐induced side effects (e.g., L‐DOPA‐induced dyskinesia). Here, the therapeutic mechanisms mediated by 5‐HT1A receptors in schizophrenia and Parkinson's disease are reviewed. This evidence should encourage discovery of new 5‐HT1A ligands, which can resolve the unmet clinical needs in the current therapy.
Keywords: Affective disorders, Cognitive impairment, Drug discovery, Movement disorders, Parkinson's disease, Schizophrenia, 5‐HT1A receptors
Introduction
The serotonergic system plays an important role in regulating various physiological functions including psychoemotional, autonomic, sensory, and motor functions [1, 2]. Serotonin (5‐HT) neurons are located in the raphe nuclei and project axons to various brain regions including the cerebral cortex, limbic areas, basal ganglia, diencephalons, and the spinal cord. Serotonergic neurotransmissions are mediated by diverse 5‐HT receptors that can be divided into seven families (5‐HT1 to 5‐HT7) encompassing 14 subtypes (5‐HT1A/1B/1D/1E/1F, 5‐HT2A/2B/2C, 5‐HT3, 5‐HT4, 5‐HT5A/5B, 5‐HT6, and 5‐HT7) according to their signal transduction pathways [1, 2]. Advances in research on 5‐HT receptors have led to the discovery of various therapeutic agents such as selective 5‐HT reuptake inhibitors (SSRI) (e.g., fluoxetine, fluvoxamine, and paroxetine), 5‐HT1A agonistic anxiolytics (e.g., buspirone, gepirone, and tandospirone), atypical antipsychotics (5‐HT2A/D2 antagonists) (e.g., risperidone, olanzapine, and quetiapine), antiemetics (5‐HT3 antagonists) (e.g., ondansetron, azasetron, and granisetron), and antimigraines (5‐HT1B/1D agonists) (e.g., sumatriptan, naratriptan, and zolmitriptan).
5‐HT1A receptors have long been implicated in the pathogenesis and treatment of anxiety and depressive disorders [3, 4, 5, 6, 7]. 5‐HT1A receptors are G‐protein coupled receptors with a 7‐transmembrane‐spanning structure (Figure 1) and are predominantly expressed in the limbic areas (e.g., hippocampus, amygdale, and lateral septum) and the raphe nuclei (e.g., raphe nuclei) [6, 8, 9]. Moderate to low levels of 5‐HT1A receptors are also expressed in the cerebral cortex, thalamus, hypothalamus, and basal ganglia (e.g., striatum) [6, 8]. 5‐HT1A receptors function both as presynaptic autoreceptors and as postsynaptic receptors. Specifically, 5‐HT1A receptors in the raphe nuclei are located on the cell body and dendrites of 5‐HT neurons, where they function as autoreceptors to negatively regulate their own activity [10]. Through this mechanism, 5‐HT1A receptors control the overall tone of serotonergic activity. On the other hand, postsynaptic 5‐HT1A receptors exist on postsynaptic membranes of neurons or nerve terminals (heteroreceptors), where 5‐HT neurons are innervated. Stimulation of postsynaptic 5‐HT1A receptors inhibits firing of target neurons through G protein‐mediated mechanisms in various regions of the brain (e.g., hippocampus, lateral septum, and cerebral cortex) (Figure 1) [11, 12, 13, 14, 15, 16].
Figure 1.
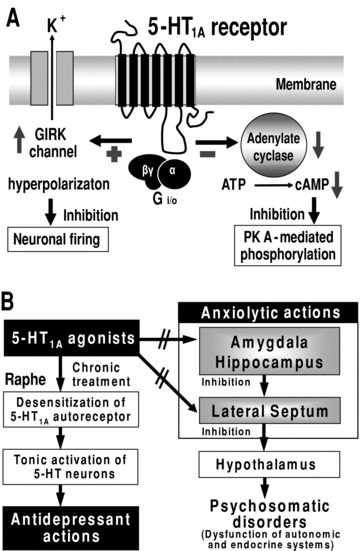
Signal transduction pathways of 5‐HT1A receptor (A) and the mechanism underlying anxiolytic and antidepressant actions of 5‐HT1A agonists (B). GIRK, G‐protein‐gated inwardly rectifying potassium channel; PK A, protein kinase A.
Recent advances in 5‐HT1A receptor research generated new insight into its therapeutic role in CNS disorders including schizophrenia, Parkinson's disease, and Alzheimer's disease [5, 17, 18, 19, 20]. Specifically, 5‐HT1A receptors seem to be a promising target for alleviating antipsychotic‐induced EPS and cognitive/affective (e.g., anxiety and depression) disorders in schizophrenia. In the treatment of patients with Parkinson's disease, 5‐HT1A agonists are expected to improve affective symptoms and core motor symptoms in Parkinson's disease. They also should ameliorate antiparkinsonian agents‐induced side effects (e.g., L‐DOPA‐induced dyskinesia and dopamine agonists‐induced emesis). In this article, the potential utility of 5‐HT1A receptors as a therapeutic target for schizophrenia and Parkinson's disease and their functional mechanisms are reviewed.
5‐HT1A Receptor and Anxiety–Depressive Disorders
Buspirone is the prototype of 5‐HT1A receptor agonist and was originally developed as an antipsychotic drug [21]. Although buspirone was not found effective for the treatment of schizophrenia, it clinically showed significant anxiolytic actions. Thereafter, numerous 5‐HT1A agonists with different intrinsic activities (e.g., full agonists: 8‐OH‐DPAT, flesinoxan, and F‐11440; partial agonists: ipsapirone, gepirone, and tandospirone) [12, 22, 23, 24] have been synthesized and shown anxiolytic activities in various animal models (e.g., Vogel's or Gellar‐Seifter conflict test and elevated‐plus maze, light–dark and conditioned‐fear paradigms) [3, 7, 24, 25, 26]. It is also known that knockout mice lacking 5‐HT1A receptors exhibit an increased anxiety status in open‐field, elevated‐plus maze, or light–dark tests [27, 28, 29]. Conversely, transgenic mice overexpressing 5‐HT1A receptors show reduced anxiety behaviors [9].
Figure 1 illustrates the structure of 5‐HT1A receptors and the mechanisms underlying the anxiolytic actions of 5‐HT1A agonists. Although the possible involvement of presynaptic 5‐HT1A autoreceptors cannot be completely ruled out, the crucial role of postsynaptic 5‐HT1A receptors in regulating anxiety has been supported by the following evidences: (1) the microinjection of 5‐HT1A agonists (e.g., 8‐OH‐DPAT, buspirone, tandospirone, and flesinoxan) directly into the limbic regions (e.g., hippocampus) causes a significant anxiolytic action [30, 31, 32, 33], (2) the denervation or inactivation of 5‐HT neurons, which abolishes presynaptic 5‐HT1A autoreceptors in the raphe nuclei, does not alter the anxiolytic activity of 5‐HT1A agonists [25, 32, 34], and (3) by using a tissue‐specific conditional rescue strategy, Gross et al. (2002) [35] demonstrated that expression of postsynaptic 5‐HT1A receptors in the forebrain (e.g., hippocampus and cerebral cortex) effectively alleviate anxiety in 5‐HT1A knockout mice. As previously described, postsynaptic 5‐HT1A receptors are predominantly expressed in the limbic areas such as the hippocampus, amygdala, and lateral septum (Figure 1). The former two structures are the major sites that control psychoemotional functions including anxiogenesis and stress reactions [4, 5, 36, 37], and the latter functions as a relay nucleus that transfers the neural outputs from the limbic system to the hypothalamus, the center of the autonomic nervous system [4, 5, 12, 37]. 5‐HT1A agonists (e.g., 8‐OH‐DPAT, tandospirone, and flesinoxan) reportedly inhibit the activity of both hippocampal and lateral septal neurons probably through activating G‐protein‐gated inwardly rectifying potassium (GIRK) channels (Figure 1) [11, 12, 13, 14, 15, 16]. It is therefore conceivable that 5‐HT1A agonists alleviate anxiety and psychosomatic disorders by inhibiting anxiogenesis in the limbic areas and its propagation to the hypothalamus in the lateral septum.
Besides anxiolytic actions, 5‐HT1A agonists (e.g., 8‐OH‐DPAT, buspirone, gepirone, and tandospirone) show a significant antidepressant activity in various animal models usually following repeated treatments [38, 39, 40, 41]. Although precise mechanisms underlying the antidepressant actions of 5‐HT1A agonists are still uncertain, the desensitization (downregulation) of presynaptic 5‐HT1A autoreceptors has been implicated in the antidepressant actions of 5‐HT1A agonists (Figure 1) [10]. Namely, repeated treatment of animals with 5‐HT1A agonists (e.g., gepirone, ipsapirone, and tandospirone) desensitizes presynaptic 5‐HT1A autoreceptors in the raphe nuclei, which disengages 5‐HT neurons from the autoreceptor‐mediated self‐inhibition [10, 42, 43, 44]. Consequently, 5‐HT neurons are tonically activated by chronic treatment with 5‐HT1A agonists and this counteracts the serotonergic deficit (5‐HT reduction) in depression (Figure 1). In contrast to presynaptic 5‐HT1A autoreceptors, the sensitivity of postsynaptic 5‐HT1A responses (e.g., anxiolytic actions) are not altered even after repeated administrations of 5‐HT1A agonists [10, 42].
Therapeutic Role of 5‐HT1A Receptors in Schizophrenia
Patients with schizophrenia exhibit diverse symptoms including positive symptoms (e.g., hallucinations, delusion, and excitation), negative symptoms (e.g., apathy, social and emotional withdrawal), cognitive impairments, and affective disorders (e.g., anxiety and depression) [17, 45] (Figure 2). Classical (typical) antipsychotics (e.g., phenothiazine, butyrophenone, and benzamide derivatives) commonly possess dopamine D2 blocking activity and effectively improve positive symptoms. These agents, however, are not so effective for negative symptoms and frequently induce extrapyramidal side effects (EPS) such as parkinsonian symptoms, akathisia, and tardive dyskinesia, which significantly disrupt the quality of life in patients (Figure 2). Recently, several new atypical antipsychotics have become available as first line treatments, including 5‐HT2A and D2 antagonists (e.g., clozapine, risperidone, ziprasidone, perospirone, olanzapine, and quetiapine) and the D2 partial agonist (i.e., aripiprazole) [36, 46]. Some of the above agents (e.g., clozapine, ziprasidone, perospirone, and aripiprazole) also possess a partial agonistic activity at 5‐HT1A receptors [36, 47]. These atypical antipsychotics, unlike the typical ones, alleviate both positive and negative symptoms and have a reduced EPS liability. Nonetheless, there are still clinical demands (unmet needs) in the treatment of schizophrenia, such as (1) improving any cognitive deficits or (2) alleviating affective disorders (e.g., anxiety and depression), (3) reducing antipsychotic‐induced EPS, metabolic (e.g., weight gain and diabetes mellitus) and endocrinological (e.g., hyperprolactinemia) side effects, and (4) controlling refractory symptoms (Figure 2). Indeed, recent comprehensive clinical reports of Clinical Antipsychotic Trials of Intervention Effectiveness (CATIE) or European First‐Episode Schizophrenia Trial (EUFEST) did not show significant advantages of the second‐generation antipsychotics (e.g., risperidone, olanzapine, and quetiapine) beyond the first‐generation antipsychotics (e.g., haloperidol and perphenazine), especially in their neurocognitive effects, in the treatment of schizophrenia [48, 49, 50].
Figure 2.
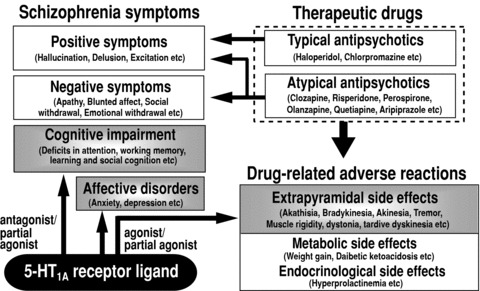
Current therapeutics, unmet clinical needs and therapeutic role of 5‐HT1A receptors in the treatment of schizophrenia.
It is known that the density of 5‐HT1A receptors is elevated in the brains of patients with chronic schizophrenia, implying a close relationship between 5‐HT1A receptors and the pathogenesis of schizophrenia [18, 36, 51, 52]. 5‐HT1A receptors are now thought to be a favored therapeutic target for schizophrenia based on the following evidences: (1) stimulation of 5‐HT1A receptors can ameliorate antipsychotic‐induced EPS [18, 53, 54, 55, 56, 57] and (2) affective disorders (e.g., anxiety and depression) [3, 7, 10], and (3) antagonism of 5‐HT1A receptors can improve cognitive impairment in schizophrenia [19, 20, 58, 59, 60] (Figure 2).
A line of studies has shown that selective 5‐HT1A agonists (e.g., 8‐OH‐DPAT, buspirone, and tandospirone) attenuated antipsychotic (e.g., haloperidol)‐induced catalepsy, bradykinesia, and other parkinsonian symptoms (Figure 2) [53, 54, 55, 56, 57]. In our studies, the ameliorative effects of 5‐HT1A agonists on extrapyramidal disorders were as potent as the antiparkinsonian agent trihexyphenidyl (a muscarinic acetylcholine [mACh] antagonist) or selective 5‐HT2 antagonists (e.g., ritanserin) [57, 61]. It is likely that 5‐HT1A agonists alleviate antipsychotic‐induced EPS by stimulating postsynaptic 5‐HT1A receptors (Figure 3), because the inactivation of 5‐HT neurons by p‐chlorophenylalanine did not affect the anti‐EPS actions of 5‐HT1A agonists (e.g., 8‐OH‐DPAT and buspirone) [53, 57, 62, 63]. Interestingly, analysis of the 5‐HT1A agonists’ actions (i.e., 8‐OH‐DPAT and tandospirone) on antipsychotic‐induced forebrain Fos expression revealed that they counteract the D2 blocking action of haloperidol in the striatum, which is closely associated with EPS induction [64, 65]. In contrast, the 5‐HT1A agonists did not alter the haloperidol‐induced Fos expression in the nucleus accumbens (shell region), which is responsible for the therapeutic action of antipsychotic agents. These data suggest that 5‐HT1A agonists may preferentially alleviate antipsychotic‐induced EPS without interrupting the therapeutic actions of antipsychotic agents.
Figure 3.
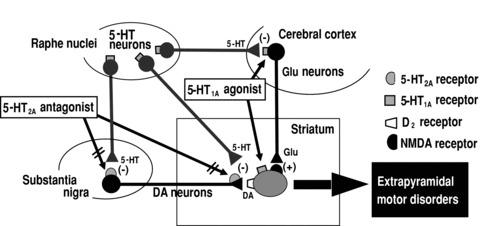
Therapeutic mechanisms and action sites of 5‐HT1A agonists and 5‐HT2A antagonists in ameliorating the extrapyramidal motor disorders. DA, dopamine; Glu, glutamate; (+), excitation; (−), inhibition.
It is postulated that blockade of 5‐HT2A receptors reduces EPS by stimulating dopaminergic neurons via blocking (a) 5‐HT2A receptor‐mediated inhibition of dopamine neuron firing in the substantia nigra and (b) dopamine release in the striatum (Figure 3) [36, 46, 66, 67, 68]. However, the action of 5‐HT1A agonists does not seem to depend on the activity of dopamine neurons because the antiparkinsonian actions of 5‐HT1A agonists (e.g., 8‐OH‐DPAT) were not altered by D2 antagonists or were apparently observed even in animals where dopamine neurons were denervated [46, 56, 62, 69]. In addition, we have recently shown that microinjections of 8‐OHDPAT into the cerebral cortex or the striatum significantly alleviate the haloperidol‐induced EPS [70]. Although systemic interactions (e.g., pharmacokinetics) between 5‐HT1A agonists and antipsychotic cannot be ruled out, these findings suggest that 5‐HT1A agonists can alleviate antipsychotic‐induced EPS by activating postsynaptic 5‐HT1A receptors both in the striatum and cerebral cortex, probably through non‐dopaminergic mechanisms (Figure 3). Because these mechanisms underlying the anti‐EPS actions of 5‐HT1A agonists are distinct from those of 5‐HT2A antagonists, it is conceivable that 5‐HT1A agonism and 5‐HT2A antagonism can work in an additive fashion in alleviating EPS and that 5‐HT1A agonists improve EPS induced not only by the classical antipsychotics (D2 antagonists), but also by the new atypical antipsychotics with 5‐HT2A blocking actions. Furthermore, although the activation of 5‐HT1A receptors did not affect dopamine release in the striatum [71], several studies [72, 73, 74] have suggested that 5‐HT1A agonist (e.g., BAY × 3702 and osemozotan) enhance dopamine release in the cerebral cortex and hippocampus, which may provide benefits in alleviating the negative symptoms of schizophrenia.
Another important role of 5‐HT1A receptors in treating schizophrenia is amelioration of cognitive deficits (Figure 2) [19, 20, 58, 59, 60]. Cognitive impairment is one of the core symptoms in schizophrenia, for which most of the currently available antipsychotics are not very effective. Recent clinical studies have shown that the partial 5‐HT1A agonist tandospirone significantly improved the cognitive deficits (assessed by Wisconsin Card Sort test or California verbal learning test) in patients with schizophrenia [58, 59]. Data from animal studies also revealed that 5‐HT1A antagonists (e.g., WAY‐100635, WAY‐101405, and lecozotan) reversed the cognitive impairment induced by mACh receptor antagonists (e.g., scopolamine) or N‐methyl‐D‐aspartate (NMDA) receptor antagonists (e.g., MK‐801) [19, 20, 75, 76]. On the other hand, full 5‐HT1A agonists (e.g., 8‐OH‐DPAT) reportedly exhibit a biphasic effects on cognitive functions, facilitation at low doses and inhibition at high doses, which is probably mediated by stimulation of presynaptic and postsynaptic 5‐HT1A receptors, respectively [19, 75]. Although further validation studies are required, the above findings suggest that antagonism of 5‐HT1A receptors provide benefits in treating cognitive impairment in schizophrenia. In this regard, the new antipsychotic lurasidone, with a 5‐HT1A antagonistic (weak partial agonistic) action [77], has been shown to improve the learning and memory impairment induced by MK‐801 [78] and to ameliorate the cognitive impairment in schizophrenia patients [79].
It is suggested that the cognitive enhancement by 5‐HT1A antagonists (e.g., WAY‐100635 and NAD‐299) is mediated by postsynaptic 5‐HT1A receptors located in the medial septum and/or diagonal band of Broca, where 5‐HT1A receptors tonically inhibit neuronal activity of acetylcholine and/or glutamate neurons [19, 20, 75, 76, 80]. Through disinhibition of this negative regulation, 5‐HT1A antagonists can enhance activities of the septo‐hippocampal/cortical acetylcholinergic and/or glutamatergic neurons and improve cognitive functions. Conversely, full 5‐HT1A agonists (e.g., 8‐OH‐DPAT) impairs cognition by inhibiting the release of glutamate and acetylcholine in various regions of the brain including the cerebral cortex [20]. A potential benefit of 5‐HT1A antagonists (e.g., WAY‐101405 and lecozotan) in the treatment of Alzheimer's disease has also been suggested [19, 20].
Therapeutic Role of 5‐HT1A Receptors in Parkinson's Disease
Parkinson's disease is a major neurological disease with specific neurodegeneration of the nigrostriatal dopamine neurons [81]. Patients with Parkinson's disease show progressive extrapyramidal motor deficits such as hypokinesia (e.g., bradykinesia and akinesia), tremors, muscle rigidity, difficulty in balance, and frozen gait (Figure 4). The dopamine precursor L‐DOPA is a key drug that effectively restores the depletion of brain dopamine and improves most parkinsonian symptoms. Other antiparkinsonian agents include dopamine D2/D3 agonists (e.g., bromocriptine, cabergoline, pramipexole, and ropinirol), dopamine releasers (e.g., amantazine) and mACh receptor antagonists (e.g., trihexyphenidyl). Inhibitors of monoamine oxidase‐B (MAO‐B) (e.g., selegiline) or catechol‐O‐methyltransferase (COMT) (e.g., entacapone) are effective adjunctive drugs which potentiate the L‐DOPA's efficacy and improve the motor fluctuations in L‐DOPA therapy (Figure 4) [81].
Figure 4.
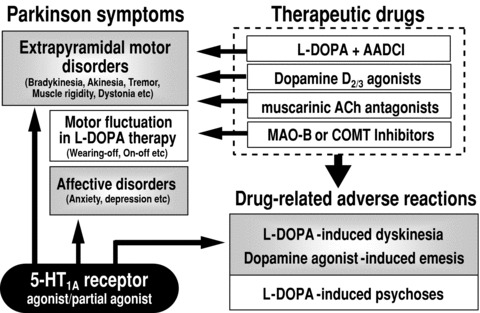
Current therapeutics, unmet clinical needs and therapeutic role of 5‐HT1A receptors in the treatment of Parkinson's disease. AADCI, aromatic L‐amino acid decarboxylase inhibitors; ACh, acetylcholine; MAO‐B, monoamine oxidase‐B; COMT, catechol‐O‐methyltransferase.
A primary concern in Parkinson's disease therapy is the lack of a drug which can prevent the development of Parkinson's disease or restore degenerated dopamine neurons. Extensive studies are now ongoing to develop neuroprotective agents or establish the gene‐, transplantation‐, or regenerative medication therapy. Other unmet needs include (1) reduction in the treatment side effects (e.g., L‐DOPA‐induced dyskinesia and psychosis and D2 agonist‐induced emesis), (2) control of motor fluctuations (e.g., wearing‐off or on‐off phenomena) in L‐DOPA therapy or (3) non‐motor symptoms including affective disorders, and (4) lack of efficacious substitutes for L‐DOPA or new antiparkinsonian agents (Figure 4).
Stimulation of 5‐HT1A receptors provides benefits in the treatment of Parkinson's disease, because 5‐HT1A agonists (e.g., 8‐OH‐DPAT) improve (1) core extrapyramidal motor disorders caused by degeneration of dopaminergic neurons [56, 62, 69, 70, 82], (2) L‐DOPA‐induced dyskinesia in animals chronically primed with L‐DOPA [70, 83, 84, 85], and (3) affective symptoms (e.g., anxiety and depressive) which are frequently observed in patients with Parkinson's disease. In addition, 5‐HT1A agonists (e.g., 8‐OH‐DPAT) reportedly possess a potent antiemetic action [86, 87, 88] (Figure 4).
As described previously, 5‐HT1A agonists alleviate antipsychotic‐induced EPS associated with the striatal D2 receptor antagonism in animals [53, 54, 55, 56, 57]. In addition, studies using animal models of Parkinson's disease have revealed that stimulation of 5‐HT1A receptors restores the motor disorders caused by lesions of dopaminergic neurons or dopamine depletion (Figure 4) [56, 62, 69, 70]. Like L‐DOPA or D2 agonists (e.g., bromocriptine), 5‐HT1A agonists (e.g., 8‐OH‐DPAT and tandospirone) induce contralateral rotation behaviors in unilaterally dopamine‐lesioned (hemiparkinsonian) animals and reverse akinesia in dopamine‐depleted animals [56, 82]. It should be noted that enhancement of locomotor activity by 5‐HT1A agonists in Parkinson's models could not be blocked by a D2 antagonist (i.e., haloperidol), but was antagonized by a 5‐HT1A antagonist (i.e., WAY‐100635) [56], indicating that the antiparkinsonian effects of 5‐HT1A agonists are independent of dopaminergic activity (Figure 3). 5‐HT1A agonists are therefore expected to exert antiparkinsonian actions in an additive fashion when combined with dopaminergic stimulants (e.g., L‐DOPA and D2/D3 agonists).
Besides the antiparkinsonian actions, several studies showed that 5‐HT1A agonists can alleviate L‐DOPA‐induced dyskinesia (Figure 4) [69, 83, 84, 85]. 8‐OH‐DPAT significantly reduced the induction of L‐DOPA‐induced dyskinesia (e.g., limb and orolingual abnormal involuntary movements) in L‐DOPA‐primed hemiparkinsonian animals. Interestingly, 8‐OH‐DPAT did not affect the antiparkinsonian actions (e.g., induction of rotation behaviors and improvement in ataxic gait) of L‐DOPA or even potentiated it [69, 85, 89]. In addition, the antidyskinetic action of 5‐HT1A agonists is mediated at least partly by striatal 5‐HT1A receptors, because the microinjection of 8‐OH‐DPAT into the striatum reverses L‐DOPA‐induced dyskinesia (Figure 3) [69]. Because orolingual dyskinesia is known to be mediated by D1 receptor activation [69, 90], interactions between 5‐HT1A and D1 receptors in the striatum may be involved in the regulation of L‐DOPA‐induced dyskinesia. Furthermore, microdialysis studies showed that local application of 5‐HT1A agonists into the cerebral cortex (primary motor areas) reduces glutamate release in the striatum [89], which could inhibit the induction of extrapyramidal disorders [91]. Thus, postsynaptic 5‐HT1A receptors in the cerebral cortex, which regulate the activity of the cortico‐striato glutamatergic neurons, are also involved in the expression of L‐DOPA‐induced dyskinesia (Figure 3). Several studies also suggest that 5‐HT1A agonists exert protective actions against dopaminergic neurotoxicity [92].
Anxiolytic and antidepressant actions of 5‐HT1A agonists are beneficial in the treatment of Parkinson's disease (Figure 4). Anxiety often coexists with depression in Parkinson's disease and the prevalence of these symptoms is reported to be very high (≈40%) in the patient population [5, 93]. 5‐HT1A agonist anxiolytics are superior to the standard benzodiazepine anxiolytics, especially in terms of their safety profiles and efficacy for depression. Specifically, anxiolytic doses of benzodiazepines (e.g., diazepam) cause diverse adverse reactions including sedation, muscle relaxation, impaired motor coordination, cognitive impairments and induction of psychophysical dependence, whereas 5‐HT1A agonistic anxiolytics (e.g., buspirone, gepirone and tandospirone) do not cause these side effects [3, 5]. The selective anxiolytic action of 5‐HT1A agonists is probably due to specific localization of 5‐HT1A receptors in brain regions (e.g., hippocampus, amygdale, and lateral septum) related to anxiogenesis and emotional regulation [6, 7]. 5‐HT1A agonists thereby appear to be more favorable in treating emotional symptoms in Parkinson's disease. Finally, a line of studies demonstrated that 5‐HT1A agonists (e.g., 8‐OH‐DPAT) show an antiemetic action against various types of emesis induced by chemicals, motion and mechanical stimulation [86, 87, 88]. Because many of the antiparkinsonian agents cause emesis as the adverse reactions, antiemetic activity of 5‐HT1A agonists also may provide a benefit in the treatment of Parkinson's disease (Figure 4).
Conclusions
This article reviewed the functions and therapeutic roles of 5‐HT1A receptors in treating CNS disorders. Recent studies have revealed new insights into the therapeutic role of 5‐HT1A receptors in treating CNS disorders including schizophrenia (e.g., reduction in antipsychotic‐induced EPS, control of cognitive impairment and affective disorders) and Parkinson's disease (e.g., improvement in core motor disability, L‐DOPA‐induced dyskinesia, and efficacy for affective disorders). This evidence will encourage drug discovery research into novel 5‐HT1A ligands with greater potency, higher selectivity, and favorable pharmacokinetic properties. Furthermore, designing 5‐HT1A ligands which combine multiple pharmacological actions (e.g., D2 and 5‐HT2A blocking activity for schizophrenia, D2 agonistic and 5‐HT2A blocking activity for Parkinson's disease) appears to be promising approach that may overcome unmet clinical needs in current CNS therapies.
Conflict of Interest
The author has no conflict of interest with any commercial or other associations in connection with the submitted article.
References
- 1. Roth BL. Multiple serotonin receptors: Clinical and experimental aspects. Ann Clin Psychiatry 1994;6:67–78. [DOI] [PubMed] [Google Scholar]
- 2. Baumgarten HG, Grozdanovic Z. Psychopharmacology of central serotonergic systems. Pharmacopsychiatry 1995;28:73–79. [DOI] [PubMed] [Google Scholar]
- 3. Feighner JP, Boyer WF. Serotonin‐1A anxiolytics: An overview. Psychopathology 1989;22(Suppl 1):21–26. [DOI] [PubMed] [Google Scholar]
- 4. Fuller RW. Role of serotonin in therapy of depression and related disorders. J Clin Psychiatry 1991;152(Suppl l):52–57. [PubMed] [Google Scholar]
- 5. Ohno Y. Tandospirone citrate, a new serotonergic anxiolytic agent: A potential use in Parkinson's disease In: Mizuno Y, Fisher A, Hanin I, editors. Mapping the progress of Alzheimer's and Parkinson's Disease. New York : Kluwer Academic/Plenum, 2002;423–428. [Google Scholar]
- 6. Pucadyil TJ, Kalipatnapu S, Chattopadhyay A. The serotonin1A receptor: A representative member of the serotonin receptor family. Cell Mol Neurobiol 2005;25:553–580. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 7. Akimova E, Lanzenberger R, Kasper S. The serotonin‐1A receptor in anxiety disorders. Biol Psychiatry 2009;66:627–635. [DOI] [PubMed] [Google Scholar]
- 8. Luna‐Munguia H, Manuel‐Apolinar L, Rocha L, Meneses A. 5‐HT1A receptor expression during memory formation. Psychopharmacology 2005:181:309–318. [DOI] [PubMed] [Google Scholar]
- 9. Kusserow H, Davies B, Hörtnagl H, et al Reduced anxiety‐related behaviour in transgenic mice overexpressing serotonin 1A receptors. Brain Res Mol Brain Res 2004;129:104–116. [DOI] [PubMed] [Google Scholar]
- 10. Blier P, Ward NM. Is there a role for 5‐HT1A agonists in the treatment of depression?. Biol Psychiatry 2003:53:193–203. [DOI] [PubMed] [Google Scholar]
- 11. Hirose A, Sasa M, Akaike A, Takaori S. Inhibition of hippocampal CA1 neurons by 5‐hydroxytryptamine, derived from the dorsal raphe nucleus and the 5‐hydroxytryptamine1A agonist SM‐3997. Neuropharmacology 1990;29:93–101. [DOI] [PubMed] [Google Scholar]
- 12. Van Den Hooff P, Galvan M. Actions of 5‐hydroxytryptamine and 5‐HT1A receptor ligands on rat dorso‐lateral septal neurons in vitro. Br J Pharmacol 1992;106:893–899. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 13. Hadrava V, Blier P, Dennis T, Ortemann C, de Montigny C. Characterization of 5‐hydroxytryptamine1A properties of flesinoxan: In vivo electrophysiology and hypothermia study. Neuropharmacology 1995;34:1311–1326. [DOI] [PubMed] [Google Scholar]
- 14. Ohno Y, Ishida K, Ishibashi T, Tanaka H, Shimizu H, Nakamura M. Effects of tandospirone, a selective 5‐HT1A agonist, on activities of the lateral septal nucleus neurons in cats In Takada Y, Curzon G, editors. Serotonin in the central nervous system and periphery. Amsterdam : Elsevier Science BV, 1995;159–165. [Google Scholar]
- 15. Raymond JR, Muklin YV, Gettys TW, Garnovskaya MN. The recombinant 5‐HT1A receptor: G protein coupling and signalling pathways. Brit J Pharmacol 1999;127:1751–1764. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 16. Tada K, Kasamo K, Ueda N, Suzuki T, Kojima T, Ishikawa K. Anxiolytic 5‐hydroxytryptamine1A agonists suppress firing activity of dorsal hippocampus CA1 pyramidal neurons through a postsynaptic mechanism: Single‐unit study in unanesthetized, unrestrained rats. J Pharmacol Exp Ther 1999;288:843–848. [PubMed] [Google Scholar]
- 17. Meltzer HY. The mechanism of action of novel antipsychotic drugs. Schizophrenia Bull 1991;17:263–287. [DOI] [PubMed] [Google Scholar]
- 18. Millan MJ. Improving the treatment of schizophrenia: Focus on serotonin 5‐HT1A receptors. J Pharmacol Exp Ther 2000;295:853–861. [PubMed] [Google Scholar]
- 19. King MV, Marsden CA, Fone KC. A role for the 5‐HT1A, 5‐HT4 and 5‐HT6 receptors in learning and memory. Trends Pharmacol Sci 2008;29:482–492. [DOI] [PubMed] [Google Scholar]
- 20. Ögren SO, Eriksson TM, Elvander‐Tottie E, et al The role of 5‐HT1A receptors in learning and memory. Behav Brain Res 2008;195:54–77. [DOI] [PubMed] [Google Scholar]
- 21. Jann MW. Buspirone: An update on a unique anxiolytic agent. Pharmacotherapy 1988;8:100–116. [DOI] [PubMed] [Google Scholar]
- 22. Schoeffter P, Hoyer D. Centrally acting hypotensive agents with affinity for 5‐HT1A binding sites inhibit forskolin‐stimulated adenylate cyclase activity in calf hippocampus. Br J Pharmacol 1988;95:975–985. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 23. Tanaka H, Tatsuno T, Shimizu , Hirose A, Kumasaka Y, Nakamura M. Effects of tandospirone on second messenger systems and neurotransmitter release in the rat brain. Gen Pharmacol 1995;26:1765–1772. [DOI] [PubMed] [Google Scholar]
- 24. Koek W, Patoiseau J‐F, Assie M‐B, et al F11440, a potent, selective, high efficacy 5‐HT1A receptor agonist with marked anxiolytic and antidepressant potential. J Pharmacol Exp Ther 1998;287:266–283. [PubMed] [Google Scholar]
- 25. Shimizu H, Tatsuno T, Tanaka H, Hirose A, Araki Y, Nakamura M. Serotonergic mechanisms in anxiolytic effect of tandospirone in the Vogel conflict test. Jpn J Pharmacol 1992;59:105–112. [DOI] [PubMed] [Google Scholar]
- 26. Stefański R, Pałejko W, Kostowski W, Płaźnik A. The comparison of benzodiazepine derivatives and serotonergic agonists and antagonists in two animal models of anxiety. Neuropharmacology 1992;31:1251–1258. [DOI] [PubMed] [Google Scholar]
- 27. Parks CL, Robinson PS, Sibille E, Shenk T, Toth M. Increased anxiety of mice lacking the serotonin1A receptor. Proc Natl Acad Sci USA 1998;95:10734–10739. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 28. Ramboz S, Oosting R, Amara DA, et al Serotonin receptor 1A knockout: An animal model of anxiety‐related disorder. Proc Natl Acad Sci USA 1998;95:14476–14481. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 29. Klemenhagen KC, Gordon JA, David DJ, Hen R, Gross CT. Increased fear response to contextual cues in mice lacking the 5‐HT1A receptor. Neuropsychopharmacology 2006;31:101–111. [DOI] [PubMed] [Google Scholar]
- 30. Kostowski W, Plaźnik A, Stefański R. Intra‐hippocampal buspirone in animal models of anxiety. Eur J Pharmacol 1989;168:393–396. [DOI] [PubMed] [Google Scholar]
- 31. Kataoka Y, Shibata K, Miyazaki A, et al Involvement of the dorsal hippocampus in mediation of the antianxiety action of tandospirone, a 5‐hydroxytryptamine1A agonistic anxiolytic. Neuropharmacology 1991;30:475–480. [DOI] [PubMed] [Google Scholar]
- 32. Schreiber R, De Vry J. Neural circuits involved in the anxiolytic effects of the 5‐HT1A receptor agonists 8‐OH‐DPAT, ipsapirone and buspirone in the rat. Eur J Pharmacol 1993;249:341–351. [DOI] [PubMed] [Google Scholar]
- 33. Li X, Inoue T, Abekawa T, et al 5‐HT1A receptor agonist affects fear conditioning through stimulations of the postsynaptic 5‐HT1A receptors in the hippocampus and amygdala. Eur J Pharmacol 2006;532:74–80. [DOI] [PubMed] [Google Scholar]
- 34. Przegalinski E, Chojnacka‐Wojcik E, Filip M. Stimulation of postsynaptic 5‐HT1A receptors is responsible for the anticonflict effect of ipsapirone in rats. J Pharm Pharmacol 1992;44:780–782. [DOI] [PubMed] [Google Scholar]
- 35. Gross C, Zhuang X, Stark K, et al Serotonin1A receptor acts during development to establish normal anxiety‐like behaviour in the adult. Nature 2002;416:396–400. [DOI] [PubMed] [Google Scholar]
- 36. Meltzer HY, Li Z, Kaneda Y, Ichikawa J. Serotonin receptors: Their key role in drugs to treat schizophrenia. Prog Neuro-Psychopharmacol Biol Psychiat 2003;27:1159–1172. [DOI] [PubMed] [Google Scholar]
- 37. Morilak DA, Barrera G, Echevarria DJ, et al Role of brain norepinephrine in the behavioral response to stress. Prog Neuro-Psychopharmacol Biol Psychiat 2005;29:1214–1224. [DOI] [PubMed] [Google Scholar]
- 38. Wieland S, Lucki I. Antidepressant‐like activity of 5‐HT1A agonists measured with the forced swim test. Psychopharmacology (Berl) 1990;101:497–504. [DOI] [PubMed] [Google Scholar]
- 39. Schreiber R, Brocco M, Gobert A, Veiga S, Millan MJ. The potent activity of the 5‐HT1A receptor agonists, S 14506 and S 14671, in the rat forced swim test is blocked by novel 5‐HT1A receptor antagonists. Eur J Pharmacol 1994;271:537–541. [DOI] [PubMed] [Google Scholar]
- 40. Matsuda T, Somboonthum P, Suzuki M, Asano S, Baba A. Antidepressant‐like effect by postsynaptic 5‐HT1A receptor activation in mice. Eur J Pharmacol 1995;280:235–238. [DOI] [PubMed] [Google Scholar]
- 41. Koek W, Vacher B, Cosi C, Assié MB, Patoiseau JF, Pauwels PJ, et al 5‐HT1A receptor activation and antidepressant‐like effects: F 13714 has high efficacy and marked antidepressant potential. Eur J Pharmacol 2001;420:103–112. [DOI] [PubMed] [Google Scholar]
- 42. Blier P, de Montigny C, Modification of 5‐HT neuron properties by sustained administration of the 5‐HT1A agonist gepirone: Electrophysiological studies in the rat brain. Synapse 1987;1:470–480. [DOI] [PubMed] [Google Scholar]
- 43. Schechter LE, Bolaños FJ, Gozlan H, et al Alterations of central serotoninergic and dopaminergic neurotransmission in rats chronically treated with ipsapirone: Biochemical and electrophysiological studies. J Pharmacol Exp Ther 1990;255:1335–1347. [PubMed] [Google Scholar]
- 44. Godbout R, Chaput Y, Blier P, de Montigny C. Tandospirone and its metabolite, 1‐(2‐pyrimidinyl)‐piperazine—I. Effects of acute and long‐term administration of tandospirone on serotonin neurotransmission. Neuropharmacology 1991;30:679–690. [DOI] [PubMed] [Google Scholar]
- 45. Kapur S, Remington G. Atypical antipsychotics: New directions and new challenges in the treatment of schizophrenia. Ann Rev Med 2001;52:503–517. [DOI] [PubMed] [Google Scholar]
- 46. Ohno Y, Ishida‐Tokuda K, Ishibashi T, et al Potential role of 5‐HT2 and D2 receptor interaction in the atypical antipsychotic action of the novel succimide derivative, perospirone. Pol J Pharmacol 1997;49:213–219. [PubMed] [Google Scholar]
- 47. Ishibashi T, Ohno Y. Perospirone hydrochloride: The novel atypical antipsychotic agent with high affinities for 5‐HT2, D2 and 5‐HT1A receptors. Biog Amines 2004;18:307–317. [Google Scholar]
- 48. Lieberman JA, Stroup TS, McEvoy JP, Swartz MS, Rosenheck RA, Perkins DO. Effectiveness of antipsychotic drugs in patients with chronic schizophrenia. N Engl J Med 2005;353:1209–1223. [DOI] [PubMed] [Google Scholar]
- 49. Keefe RS, Bilder RM, Davis SM, et al Neurocognitive effects of antipsychotic medications in patients with chronic schizophrenia in the CATIE Trial. Arch Gen Psychiatry 2007;64:633–647. [DOI] [PubMed] [Google Scholar]
- 50. Davidson M, Galderisi S, Weiser M, et al Cognitive effects of antipsychotic drugs in first‐episode schizophrenia and schizophreniform disorder: A randomized, open‐label clinical trial (EUFEST). Am J Psychiatry 2009;166:675–682. [DOI] [PubMed] [Google Scholar]
- 51. Hashimoto T, Nishino N, Nakai H, Tanaka C. Increase in serotonin 5‐HT1A receptors in prefrontal and temporal cortices of brains from patients with chronic schizophrenia. Life Sci 1991;48:355–363. [DOI] [PubMed] [Google Scholar]
- 52. Gurevich EV, Joyce JN. Alterations in the cortical serotonergic system in schizophrenia: A postmortem study. Biol Psychiatry 1997;42:529–545. [DOI] [PubMed] [Google Scholar]
- 53. Neal‐Beliveau BS, Joyce JN, Lucki I. Serotonergic involvement in haloperidol‐induced catalepsy. J Pharmacol Exp Ther 1993;265:207–217. [PubMed] [Google Scholar]
- 54. Wadenberg ML, Young KA, Richter JT, Hicks PB. Effects of local application of 5‐hydroxytryptamine into the dorsal or median raphe nuclei on haloperidol‐induced catalepsy in the rat. Neuropharmacology 1999;38:151–156. [DOI] [PubMed] [Google Scholar]
- 55. Prinssen EP, Colpaert FC, Koek W. 5‐HT1A receptor activation and anti‐cataleptic effects: High‐efficacy agonists maximally inhibit haloperidol‐induced catalepsy. Eur J Pharmacol 2002;453:217–221. [DOI] [PubMed] [Google Scholar]
- 56. Ishibashi T, Ohno Y. Antiparkinsonian actions of a selective 5‐HT1A agonist, tandospirone, in rats. Biog. Amines 2004;8:329–338. [Google Scholar]
- 57. Ohno Y, Shimizu S, Imaki J, et al Evaluation of the antibradykinetic actions of 5‐HT1A agonists using the mouse pole test. Prog Neuro-Psychopharmacol Biol Psychiat 2008;32:1302–1307. [DOI] [PubMed] [Google Scholar]
- 58. Sumiyoshi T, Matsui M, Nohara S, et al Enhancement of cognitive performance in schizophrenia by addition of tandospirone to neuroleptic treatment. Am J Psychiatry 2001;158:1722–1725. [DOI] [PubMed] [Google Scholar]
- 59. Sumiyoshi T, Park S, Jayathilake K, Roy A, Ertugrul A, Meltzer HY. Effect of buspirone, a serotonin1A partial agonist, on cognitive function in schizophrenia: A randomized, double‐blind, placebo‐controlled study. Schizophrenia Res 2007;95:158–168. [DOI] [PubMed] [Google Scholar]
- 60. Meltzer HY, Sumiyoshi T. Does stimulation of 5‐HT1A receptors improve cognition in schizophrenia? Behav Brain Res 2008;195:98–102. [DOI] [PubMed] [Google Scholar]
- 61. Ohno Y, Ishida K, Ikeda K, Ishibashi T, Okada K, Nakamura M. Evaluation of bradykinesia induction by SM‐9018, a novel 5‐HT2 and D2 receptor antagonist, using the mouse pole test. Pharmacol Biochem Behav 1994;49:19–23. [DOI] [PubMed] [Google Scholar]
- 62. Mignon L, Wolf WA. Postsynaptic 5‐HT1A receptors mediate an increase in locomotor activity in the monoamine‐depleted rat. Psychopharmacology 2002;163:85–94. [DOI] [PubMed] [Google Scholar]
- 63. Zazpe A, Artaiz I, Innerarity A, et al In vitro and in vivo characterization of F‐97013‐GD, a partial 5‐HT1A agonist with antipsychotic‐ and antiparkinsonian‐like properties. Neuropharmacology 2006;51:129–140. [DOI] [PubMed] [Google Scholar]
- 64. Ohno Y, Shimizu S, Imaki J, et al. (Anticataleptic 8‐OH‐DPAT preferentially counteracts with haloperidol‐induced Fos expression in the dorsolateral striatum and the core region of the nucleus accumbens. Neuropharmacology 2008;55:717–723. [DOI] [PubMed] [Google Scholar]
- 65. Ohno Y, Shimizu S, Imaki J. Effects of tandospirone, a 5‐HT1A agonistic anxiolytic agent, on haloperidol‐induced catalepsy and forebrain Fos expression in mice. J Pharmacol Sci 2009;109:593–599. [DOI] [PubMed] [Google Scholar]
- 66. Kapur S, Remington G. Serotonin‐dopamine interaction and its relevance to schizophrenia. Am J Psychiatry 1996;153:466–476. [DOI] [PubMed] [Google Scholar]
- 67. Remington G, Kapur S. D2 and 5‐HT2 receptor effects of antipsychotics: Bridging basic and clinical findings using PET. J Clin Psychiatry 1999;60(Suppl 10):15–19. [PubMed] [Google Scholar]
- 68. Horacek J, Bubenikova‐Valesova V, Kopecek M, et al Mechanism of action of atypical antipsychotic drugs and the neurobiology of schizophrenia. CNS Drugs 2006;20:389–409. [DOI] [PubMed] [Google Scholar]
- 69. Dupre KB, Eskow KL, Barnum CJ, Bishop C. Striatal 5‐HT1A receptor stimulation reduces D1 receptor‐induced dyskinesia and improves movement in the hemiparkinsonian rat. Neuropharmacology 2008;55:1321–1328. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 70. Shimizu, S , Tatara, A , Imaki, J , Ohno, Y , Role of cortical and striatal 5‐HT1A receptors in alleviating antipsychotic‐induced extrapyramidal disorders. Prog Neuro-Psychopharmacol Biol Psychiat 2010;34:877–881. [DOI] [PubMed] [Google Scholar]
- 71. Bantick RA, De Vries MH, Grasby PM. The effect of a 5‐HT1A receptor agonist on striatal dopamine release. Synapse 2005;57:67–75. [DOI] [PubMed] [Google Scholar]
- 72. Ago Y, Koyama Y, Baba A, Matsuda T. Regulation by 5‐HT1A receptors of the in vivo release of 5‐HT and DA in mouse frontal cortex. Neuropharmacology 2003;45:1050–1056. [DOI] [PubMed] [Google Scholar]
- 73. Li Z, Ichikawa J, Dai J, Meltzer HY. Aripiprazole, a novel antipsychotic drug, preferentially increases dopamine release in the prefrontal cortex and hippocampus in rat brain. Eur J Pharmacol 2004;493:75–83. [DOI] [PubMed] [Google Scholar]
- 74. Díaz‐Mataix L, Scorza MC, Bortolozzi A, Toth M, Celada P, Artigas F. Involvement of 5‐HT1A receptors in prefrontal cortex in the modulation of dopaminergic activity: Role in atypical antipsychotic action. J Neurosci 2005;25:10831–10843. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 75. Lüttgen M, Elvander E, Madjid N, Ögren SO. Analysis of the role of 5‐HT1A receptors in spatial and aversive learning in the rat. Neuropharmacology 2005;48:830–852. [DOI] [PubMed] [Google Scholar]
- 76. Madjid N, Tottie EE, Lüttgen M, et al 5‐Hydroxytryptamine 1A receptor blockade facilitates aversive learning in mice: Interactions with cholinergic and glutamatergic mechanisms. J Pharmacol Exp Ther 2006;316:581–591. [DOI] [PubMed] [Google Scholar]
- 77. Ishibashi T, Horisawa T, Tokuda K, et al Pharmacological profile of lurasidone, a novel antipsychotic agent with potent 5‐HT7 and 5‐HT1A receptor activity. J Pharmacol Exp Ther 2010;334:171–181. [DOI] [PubMed] [Google Scholar]
- 78. Ishiyama T, Tokuda K, Ishibashi T, Ito A, Toma S, Ohno, Y . Lurasidone (SM‐13496), a novel atypical antipsychotic drug, reverses MK‐801‐induced impairment of learning and memory in the rat passive‐avoidance test. Eur J Pharmacol 2007;572:160–170. [DOI] [PubMed] [Google Scholar]
- 79. Nakamura M, Ogasa M, Guarino J, et al Lurasidone in the treatment of acute schizophrenia: A double‐blind, placebo‐controlled trial. J Clin Psychiatry 2009;70:829–836. [DOI] [PubMed] [Google Scholar]
- 80. Jeltsch H, Bertand F, Galani R, Lazarus C, Schimchowitsch S, Cassel J‐C. Intraseptal injection of the 5‐HT1A/5‐HT7 agonist 8‐OH‐DPAT and working memory in rats. Psychopharmacology 2004;175:37–46. [DOI] [PubMed] [Google Scholar]
- 81. Samii A, Nutt JG, Ransom B. Parkinson's disease. Lancet 2004;363:1783–1793. [DOI] [PubMed] [Google Scholar]
- 82. Matsubara K, Shimizu K, Suno M, et al Tandospirone, a 5‐HT1A agonist, ameliorates movement disorder via non‐dopaminergic systems in rats with unilateral 6‐hydroxydopamine‐ generated lesions. Brain Res 2006;1112:126–133. [DOI] [PubMed] [Google Scholar]
- 83. Tomiyama M, Kimura T, Maeda T, Kannari K, Matsunaga M, Baba M. A serotonin 5‐HT1A receptor agonist prevents behavioral sensitization to L‐DOPA in a rodent model of Parkinson's disease. Neurosci Res 2005;52:185–194. [DOI] [PubMed] [Google Scholar]
- 84. Dupre KB, Eskow KL, Steiniger A, et al Effects of coincident 5‐HT1A receptor stimulation and NMDA receptor antagonism on L‐DOPA‐induced dyskinesia and rotational behaviors in the hemi‐parkinsonian rat. Psychopharmacology (Berl) 2008;199:99–108. [DOI] [PubMed] [Google Scholar]
- 85. Bishop C, Krolewski DM, Eskow KL, et al Contribution of the striatum to the effects of 5‐HT1A receptor stimulation in L‐DOPA‐treated Hemiparkinsonian rats. J Neurosci Res 2009;87:1645–1658. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 86. Wolff MC, Leander JD. Comparison of the antiemetic effects of a 5‐HT1A agonist, LY228729, and 5‐HT3 antagonists in the pigeon. Pharmacol Biochem Behav 1995;52:571–575. [DOI] [PubMed] [Google Scholar]
- 87. Andrews P, Torii Y, Saito H, Matsuki N. The pharmacology of the emetic response to upper gastrointestinal tract stimulation in Suncus murinus. Eur J Pharmacol 1996;307:305–313. [DOI] [PubMed] [Google Scholar]
- 88. Javid FA, Naylor RJ. The effect of the 5‐HT1A receptor agonist, 8‐OH‐DPAT, on motion‐induced emesis in Suncus murinus. Pharmacol Biochem Behav 2006;85:820–826. [DOI] [PubMed] [Google Scholar]
- 89. Antonelli T, Fuxe K, Tomasini MC, et al Effects of sarizotan on the corticostriatal glutamate pathways. Synapse 2005;58:193–199. [DOI] [PubMed] [Google Scholar]
- 90. Ohno Y, Ishida‐Tokuda K, Ishibashi T, Nakamura M. Effects of perospirone (SM‐9018), a potential atypical neuroleptic, on dopamine D1 receptor‐mediated vacuous chewing movement in rats: A rple of 5‐HT2 receptor blocking activity. Pharmacol Biochem Behav. 1997;57:889–895. [DOI] [PubMed] [Google Scholar]
- 91. Chartoff EH, Ward RP, Dorsa DM. Role of adenosine and N‐methyl‐D‐aspartate receptors in mediating haloperidol‐induced gene expression and catalepsy. J Pharmacol Exp Ther. 1999;291:531–537. [PubMed] [Google Scholar]
- 92. Bezard E, Gerlach I, Moratalla R, Gross CE, Jork R. 5‐HT1A receptor agonist‐mediated protection from MPTP toxicity in mice and macaque models of Parkinson's disease. Neurobiol Disease, 2006;23:77–86. [DOI] [PubMed] [Google Scholar]
- 93. Richard IH, Schiffer RB, Kurlan R. Anxiety and Parkinson's disease. J Neuropsychiatry Clin Neurosci 1996;8:383–392. [DOI] [PubMed] [Google Scholar]