

Abstract
Abstract. Triploid V79 cells were established from tetraploid cells. Diploid V79 cells were polyploidized by K‐252a, an inhibitor of protein kinases, and then released from the drug for 10 days. At that time, the cell population was a mixture of diploid and tetraploid cells. Triploid cells were obtained through the cloning of tetraploid cells. They had 33 chromosomes (1.5 times the diploid number) and showed a karyotype of three homologueous chromosomes. The duration of the G1, S and G2/M phases was almost the same as for diploid cells. The cell volume of triploid V79 cells was about two times that of the diploid cells. An explanation for the diploid‐tetraploid‐triploid transition is proposed.
INTRODUCTION
V79 Chinese hamster lung cells (V79 cells) have been found to possess unusual stability in diploid chromosome pattern through subcultures. They are polyploidized by demecolcine and some of cells become tetraploid (Harris 1971). Though the re‐polyploidization of tetraploid V79 cells produced octaploid cells at a low frequency, the ploidy decreased during serial passage (Harris 1971). Chromosome instability at higher ploidy is well known and in some cases, the terminal ploidy was pseudo triploid (Moore et al. 1968) or aneuploid (Graves & McMillan 1984). It is of interest to elucidate why triploidy or aneuploidy represents the terminal stage in the ploidy degradation process.
V79 cells were polyploidized at the same rate by both K‐252a and demecolcine, inhibitors of protein kinases and spindle‐fibre formation in M phase, respectively (Fujikawa‐Yamamoto et al. 1993). The polyploidized population became a mixture of diploid and tetraploid cells after removal of K‐252a, but not demecolcine, indicating greater production of tetraploid cells with K‐252a than demecolcine (Fujikawa‐Yamamoto et al. 2000). It is therefore of interest to establish hyperploid V79 cell lines using K‐252a.
A relationship between the rate of DNA synthesis and DNA content has been reported for Chinese hamster cells by Graves & McMillan (1984), wherein the duration of S phase is almost constant regardless of the DNA content. Although several studies have reported a constant duration of the S phase in the polyploidization of cultured cells (Graves et al. 1984; Usui et al. 1991; Brenneisen et al. 1994; Watters et al. 1994; Jordan et al. 1996; Zhang et al. 1996; Fujikawa‐Yamamoto et al. 1997), the doubling time increased with the increase in ploidy (Graves et al. 1984; 2000, 1997). Triploid cells in a steady state of growth have not been well studied in terms of cell cycle parameters, i.e. the duration of the G1, S and G2/M phases.
In this study, a triploid V79 cell line was established from tetraploid cells after removal of K‐252a, and compared with diploid V79 cells. An explanation for the diploid‐tetraploid‐triploid transition is proposed. The hypothesis refers to the relative stability of the chromosomes in diploid, triploid and tetraploid cells.
MATERIALS AND METHODS
Cells
V79 cells, derived from the lung of male Chinese hamsters, were maintained in a humidified atmosphere of 5% CO2 at 37 °C as a monolayer culture in a Leibovitz's L15: Ham's F10 mixture (7 : 3) supplemented with 10% fetal calf serum (CELLect GOLD, ICN Biomedicals, Aurora, OH, USA), streptomycin (100 µg/ml) and penicillin (50 U/ml). The triploid V79 cells were cultured under the same culture‐conditions as described above.
Drug treatment and cell cloning
Exponentially growing V79 cells were plated in Petri dishes (90 mm diameter, Nunc, Naperville, IL, USA) at a density of about 2 × 105 cells/dish, and the medium was changed 24 h after seeding. Twelve hours thereafter, the cells were exposed to K‐252a (800 nm, Funakoshi, Tokyo, Japan) for 72 h. The cells were then released from the drug exposure by changing the medium. Every 3 days, the V79 cells were subcultured by 1/2 through two washes with PBS(–) (divalent cation‐free phosphate‐buffered saline) and trypsinization with 0.17% trypsin and 30 mm EDTA.
At 10 days after K‐252a release, the cell population was a mixture of diploid and tetraploid cells, and the cells were seeded in 96‐well dishes (96‐well tissue culture clusters, 6.4 mm diameter, Corning Costar Co., Cambridge, MA, USA) at low density. The wells containing a cell were selected. The cells were incubated in a CO2 incubator with occasional changes of medium. About 10 days later, the cells were trypsinized, transferred into culture flasks (25 cm2, Corning Costar Co.) and cultured again. Cellular DNA content was checked by flow cytometry (FCM), when the cells had grown sufficiently to obtain DNA histograms. The first measurement for DNA histograms was performed at about a month after the removal of K‐252a, and the expected number of cell divisions was above 20 at that time. Triploid V79 cells were cloned.
Cell growth
Triploid V79 cells cultured continuously for 10 months were used. The cells were subcultured every day by one half or one fourth in culture flasks (25 cm2, Corning Costar Co.), and the residual cells were counted. Diploid V79 cells were prepared as a control for the measurements. Triploid V79 cells stored at −135 °C in early passage were resolved in the culture, and compared to the triploid cells cultured long‐term.
Cell preparation for FCM and cell counting
The cells were fixed with 20% ethanol, and then incubated with 0.5 ml of 0.25% RNase (Type II‐A, Sigma, St Louis, MO, USA) for 3 h at 4 °C. The cells were counted with haemocytometers at this stage. Immediately before the measurements, the cells were stained with PI (propidium iodide, 7.5 × 10−5 m) and examined for red fluorescence by FCM. Under these staining conditions, the signal due to residual double‐stranded RNA is negligible and the relative intensity of the red fluorescence corresponds to the DNA content (Krishan 1975).
Flow cytometry (FCM)
The fluorescence from individual cells was measured using a FACSORT (Becton‐Dickinson Immunocytometry Systems, Franklin Lakes, NJ, USA). The fluorescence of individual cells irradiated with a focused laser light at a wavelength of 488 nm was detected using a photomultiplier tube. The relative intensity of red fluorescence (FL2A and FL2H) was measured and DNA histograms were obtained.
Cell cycle analysis
FCM data of FL2H, signals of red‐fluorescence intensity through a logarithmic amplifier, for 10 000 cells were input to CASL, a software for cell cycle analysis of DNA histograms on a log scale, through a transfer software, FACS to ASCII (free ware), and the DNA histograms were decomposed to cell fractions depending on the DNA content (Fujikawa‐Yamamoto 1999a). CASL was written with Mathematica using a personal computer (Macintosh) and can analyse DNA histograms with a DNA content from 2 to 128 complement (c). The algorithm is similar to Fried's method (Fried et al. 1976; Fried 1977) except that normal distribution functions having the same half width instead of the same CV (coefficient of variation) value are used as components.
Chromosome analysis
Exponentially growing triploid V79 cells in Petri dishes (60 mm diameter, Nunc) were exposed to demecolcine at a concentration of 270 nm for 1 h. The cells were trypsinized and swelled by 75 mm KCl, fixed with fixing solution (CH3OH:CH3COOH = 7 : 3) and dropped onto glass slides. The cells were stained with Giemsa solution in order to photograph the chromosomes. Chromosome numbers were counted from the photographs. Karyotype analysis was performed using Karyovision (Sumitomo Kinzoku, Tokyo, Japan).
Cell volume distribution
Exponentially growing diploid and triploid V79 cells were trypsinized, fixed with 20% ethanol and resuspended in PBS(–). The distribution of cell‐volume (Coulter volume) was measured using a Coulter Counter (ZM/256, Coulter Electronics, Fullerton, CA, USA). Standard spheres (9.8 µm diameter, Coulter sphere, Coulter Electronics) were used as a control. Note that the Coulter volume depends on the material being tested, because it is calculated based on the resistance of particles.
Observation of cell morphology
Exponentially growing diploid and triploid V79 cells were washed once with PBS(–) and fixed with methanol. The cells were stained by a haematoxylin/eosin method. Photographs were taken under a microscope (BX 50, Olympus, Tokyo, Japan.) equipped with a digital camera system (DS4040, Olympus). The images were entered into a personal computer and printed out at a given magnification.
RESULTS
To establish hyperploid V79 cells, diploid V79 cells were exposed to K‐252a for 3 days and then released (Fig. 1). The V79 cells polyploidized by K‐252a, were further polyploidized after its removal. At 9–11 days after drug removal, the population was a mixture of cells with peaks of 2, 4 and 8c DNA content, suggesting a diploid/tetraploid cell mixture. At 10 days, the cells were seeded in multi‐well plates to obtain monoclonal tetraploid lines. After a month, a monoclonal cell population was obtained. We temporarily named these cells pseudo triploid V79 cells, because their DNA content measured from the fluorescence intensity of PI intercalated in double‐stranded DNA was about 1.6 times that of diploid cells (see Fig. 2).
Figure 1.
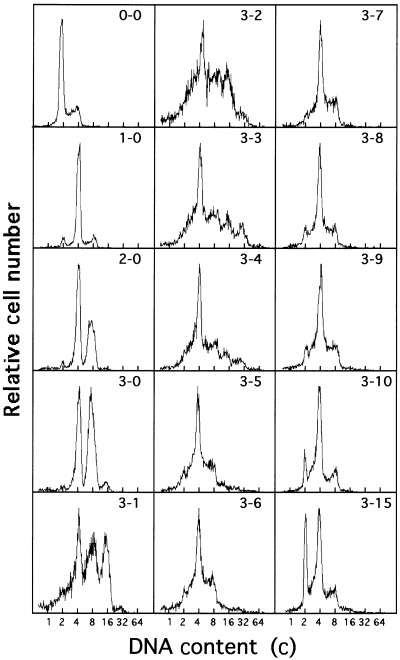
Changes in DNA fluorescence histograms of V79 cells after the addition and removal of K‐252a. Exponentially growing V79 cells were exposed to K‐252a at a concentration of 800 n m for 3 days. The cells were released from the drug and cultured again with subcultures. Paired numerals in the histogram represent the time (days) of the drug removal and the time (days) after removal, in that order. The abscissa represents the relative DNA content (c, complement).
Figure 2.
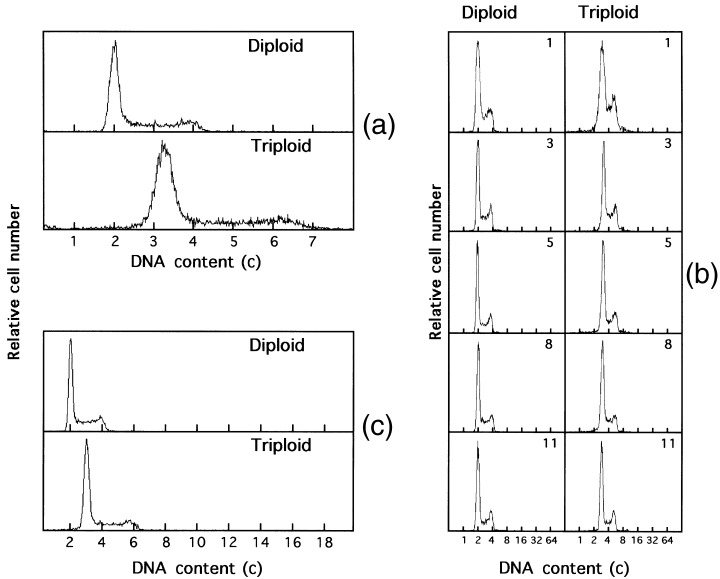
Changes in DNA fluorescence histograms of diploid and triploid V79 cells. (a) and (c) represent DNA histograms in the first (0) and the last (13th) month, respectively. (b) is of immediate times. Numerals in the histogram represent the time (month) after the first measurement. The abscissa represents the relative DNA content (c, complement).
To examine the stability of the cellular DNA content of pseudo‐triploid V79 cells, DNA histograms were measured long‐term (Fig. 2). The DNA content of the cells was about 1.6 times that of the parent diploid cells, and did not alter over 13 months with occasional subculturing. The estimated passage was above 800, indicating the establishment of a cell line.
To test the integrity of pseudo triploid V79 cells, the chromosomes were examined. Figure 3 shows the distribution of chromosome numbers and karyotyping charts for the new cells. The chromosomes numbered 33 (mode) and could be divided into 11 groups of three homologueous chromosomes, proving the cells were triploid. Hereafter, the newly established cells will be called triploid V79 cells. The fact that the triploid cells had 1.6 times the DNA content of diploid cells, but only three homologueous chromosomes, will be addressed in the Discussion.
Figure 3.
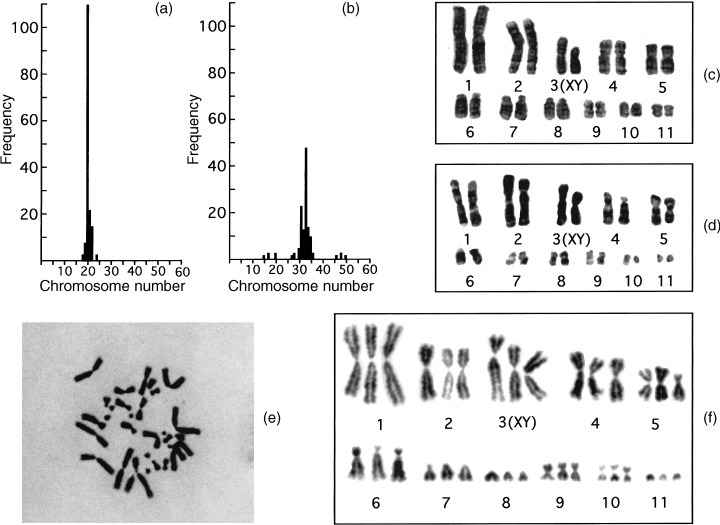
Histograms of chromosome number (a, b), karyotyping charts (c, d, f) and micrographs (e) of diploid (a, d) and triploid (b, e, f) V79 cells. Exponentially growing diploid and triploid V79 cells were exposed to demecolcine at a concentration of 270 n m for 1 h. The chromosomes were stained with Giemsa solution. About 100 cells were enumerated from enlarged photographs. (c) is for lymphocytes of a male Chinese hamster.
To examine cell‐cycle parameters, the doubling time and phase fraction of triploid V79 cells were measured compared with those of the parent diploid cells. Figure 4 shows changes in cell numbers long‐term (left panel). Doubling times of the cell population were about 20.4 h for both diploid and triploid V79 cells. We concluded that the doubling time was almost the same for diploid and triploid V79 cells under these experimental conditions. Note that frequent subculturing tends to extend the doubling time (Fujikawa‐Yamamoto 1982).
Figure 4.
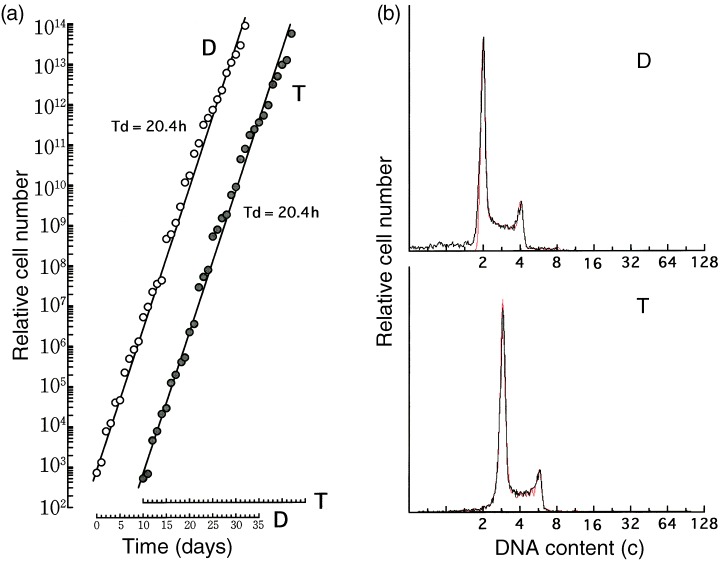
Growth curves (a) and representative CASL outputs (b) of diploid (D) and triploid (T) V79 cells. Exponentially growing diploid and triploid V79 cells were cultured long term with daily subcultures. Changes in the relative cell number of diploid (open circle) and triploid (closed circle) V79 cells were determined (a). The abscissa represents the time (day) after the drug removal. Solid lines were drawn to facilitate understanding. In b, solid (black) and dotted (red) lines represent experimental and synthesized histograms, respectively.
Representative outputs of CASL for DNA histograms of exponentially growing diploid and tetraploid V79 cells are also shown in Fig. 4 (right panel). The phase fractions obtained through CASL are listed in Table 1. No remarkable difference was observed in the cell cycle parameters of diploid and tetraploid V79 cells, although a 0.9‐h elongation of the S phase in triploid cells was observed. No difference was observed between triploid V79 cells re‐cultured after storage at −135 °C and cultured continuously (data not shown). The cell cycle parameters of triploid and diploid V79 cells are the same, at least under our experimental conditions.
Table 1.
Cell cycle parameters of diploid and triploid V79 cells
| Diploid | Triploid | |||||
|---|---|---|---|---|---|---|
| Doubling time (h) | 20.4 | 20.4 | ||||
| Phase | G1 | S | G2/M | G1 | S | G2/M |
| Fraction* | 0.508 (0.485) | 0.375 (0.359) | 0.116 (0.111) | 0.480 (0.388) | 0.415 (0.336) | 0.105 (0.085) |
| Duration (h)** | 8.6 | 8.5 | 3.2 | 8.1 | 9.4 | 2.9 |
Phase fractions of diploid and triploid cells were determined omitting those for other ploidies in the V79 cell population. Numbers in parentheses represent the fraction in the total cell population.
Phase duration was calculated through conventional equations ( Watanabe & Okada 1967 ) using doubling time instead of cycle time.
To examine the morphological characteristics of triploid V79 cells, micrographs and the cell distribution in terms of volume were compared with those of diploid cells (Fig. 5). Most triploid V79 cells were mononuclear and exhibited normal cell division. The cell volume (coulter volume) was about two times that of the diploid cells, suggesting a tetraploid‐triploid transition in the establishment of triploid V79 cells.
Figure 5.
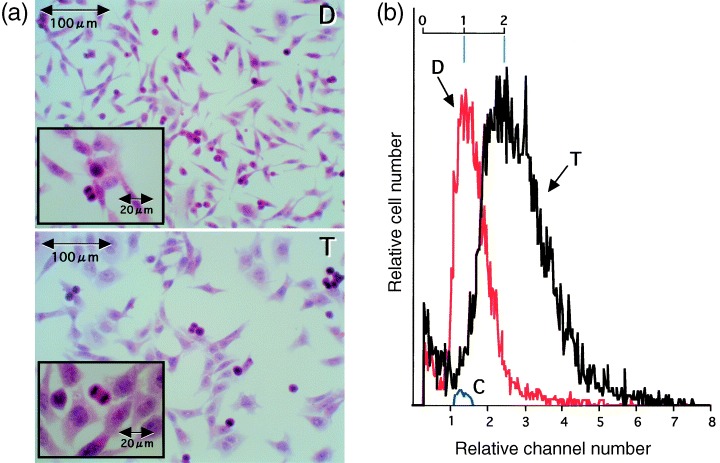
Micrographs (a) and volume distribution (b) of diploid (D) and triploid (T) V79 cells. Exponentially growing diploid and triploid V79 cells were prepared for measurements. In a, the cells were stained with HE. In b, longitudinal lines (blue) were drawn to facilitate understanding. C is of standard spheres 9.8 µm in diameter.
DISCUSSION
K‐252a, a potent inhibitor of protein kinases C and I (Zollner 1993), polyploidizes cultured cells in G2‐G1 transition (Usui et al. 1991), although its role in polyploidization is still unclear (1995, 1994). V79 cells were polyploidized by K‐252a exhibiting a mainly multilobed mononuclear morphology (Fujikawa‐Yamamoto et al. 1999b). Seven days after K‐252a removal, the cell population was a mixture of diploid and tetraploid cells (Fujikawa‐Yamamoto et al. 2000). Although we aimed to establish a tetraploid cell line, the resultant ploidy was triploid. It seems that the transformation from tetraploidy to triploidy occurred within 20 passages.
Although DNA degradation in polyploid Chinese hamster cells has been reported (Harris 1971; Graves et al. 1984), the mechanism behind the loss of chromosome has not been interpreted. It is of note that the tetraploid V79 cells became triploid within 20 passages, suggesting that a haploid set was missed in the short time. A new structural model that allows for a tetraploid‐triploid transition may be required to explain the loss of a haploid set of chromosomes.
1987, 1989a, 1989b, 1996 ) presented a set of models for the DNA structure in cells. In one model, the double‐stranded DNA in cells has a fractal‐like structure with an S‐shaped unit and homologueous chromosomes arranged to point symmetrically ( Takahashi 1996 ). To interpret the change from tetraploidy to triploidy in V79 cells, Takahashi's model was extended to polyploid cells ( Fig. 6 ). As shown in Fig. 6 , homologueous chromosomes cannot close in on each other in the folded structure (Rabl orientation) at G 1 phase in diploid cells, but can in tetraploid cells. DNA replication may be bypassed at paired double‐stranded DNA, resulting in chromosomal degradation. Whole pairing of DNA may induce a tetraploid‐diploid transition through cell division without DNA replication ( Fig. 6 , arrow 4–2). If pairing of double‐stranded DNA occurs in half part of the S loop in tetraploid cells, triploid cells will be produced through cell division ( Fig. 6 , arrow 4–3). Note that the pairing (hybridization) is difficult in diploid and triploid cells, suggesting a stable ploidy.
Figure 6.
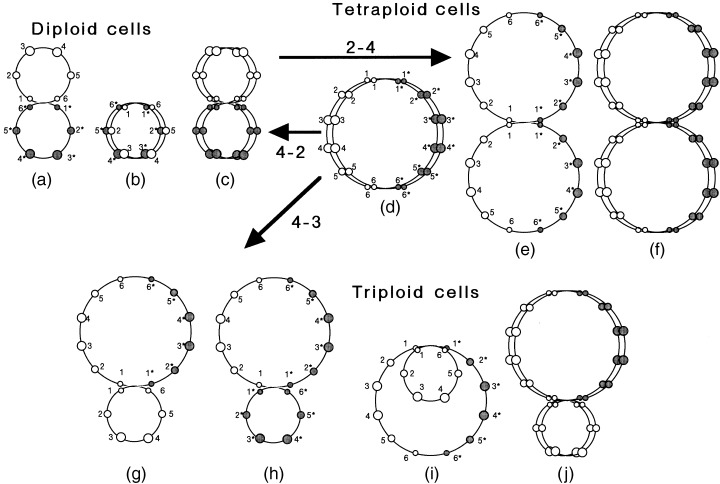
A model of the DNA structure of diploid, triploid and tetraploid cells. In diploid cells (a, b and c), DNA is linked as homologueous chromosomes arranged to point symmetrically (a). Circles 1 and 6, 2 and 5, and 3 and 4 represent sets of short, medium‐sized and long chromosomes, respectively. 1–6 and 1* to 6* represent paternal and maternal haploid sets, respectively. Homologous chromosomes cannot close each other in the folded structure (Rabl orientation) in G 0 /G 1 phase (b). (c) is the replicated DNA structure in the G 2 phase. Tetraploid cells are made through the linking of maternal 1* and paternal 1 DNA between two double‐stranded chains (arrow 2–4, e). Homologous chromosomes can approach each other allowing DNA hybridization in the Rabl orientation of tetraploid cells (d), producing a possible tetraploid‐diploid transition (arrow 4–2). (f) is the replicated DNA structure in the G 2 phase of tetraploid cells. When two haploid sets of 1–6 (or 1*‐6*) hybridize and escape DNA replication, triploid cells are produced through cell division (arrow 4–3, g or h). Homologous chromosomes cannot close each other in the Rabl orientation of triploid cells (i). (j) is the replicated DNA structure at G 2 in triploid cells.
The possibility that triploid V79 cells were directly derived from diploid cells by DNA reduplication of a haploid set of chromosomes should be mentioned. The cell volume of triploid V79 cells was about two times that of the diploid cells. This fact suggests that the triploid cells were derived from tetraploid cells, because the signal intensity of forward scatter for tetraploid cells at 10 days after K‐252a release was about two times that of diploid cells (data not shown). It has been reported that the diploid‐tetraploid cell mixture became a diploid population 28 days after K‐252a release (Fujikawa‐Yamamoto et al. 2000). It seems that K‐252a derived‐tetraploid V79 cells tend to be transformed.
It is of interest that the cell cycle parameters were almost the same in diploid and triploid V79 cells. Though two rates of DNA synthesis, constant and double‐increase, have been reported for polyploid cells, the doubling time increased with the ploidy in both cases (Graves et al. 1984; 2000, 1997). The relative abundance of cellular content in triploid cells as tetraploid cells may be responsible for the similarity in cell cycle parameters (Fujikawa‐Yamamoto 1982).
The DNA content of triploid cells was measured as 1.6 times that of diploid cells, inconsistent with the presence of three homologueous chromosomes. It is possible that the double‐stranded DNA of triploid V79 cells was relatively loose which increased the intercalation of PI, or that part of a small chromosome near the symmetrical centre did not hybridize during the tetraploid‐triploid transition, resulting in an increase in the DNA content of triploid V79 cells. At this stage, we have no evidence to explain the contradiction. A precise karyotyping of several triploid V79 cells may be required, particularly regarding the number of X and Y chromosomes.
ACKNOWLEDGEMENTS
This study was supported in part by a Grant‐in‐Aid for Scientific Research from the Ministry of Education, Culture, Sports, Science and Technology (13670228), and by a grant for High‐Technology Research Center Project by Kanazawa Medical University (H‐01). Two male Chinese hamsters were donated by the animal experimental centre of Kanazawa University.
REFERENCES
- Brenneisen P, Gogol J, Bayreuther K (1994) DNA synthesis and Fos and Jun protein expression in mitotic and postmitotic WI‐38 fibroblasts in vitro . Exp. Cell Res. 211, 219. [DOI] [PubMed] [Google Scholar]
- Fried J (1977) Analysis of deoxyribonucleic acid histograms from flowcytofluorometry. Estimation of distribution of cells within S phase. J. Histochem. Cytochem. 25, 942. [DOI] [PubMed] [Google Scholar]
- Fried J, Perez AG, Clarkson BD (1976) Flowcytofluorometric analysis of cell cycle distribution using propidium iodide. Properties of the method and mathematical analysis of the data. J. Cell Biol. 71, 172. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Fujikawa‐Yamamoto K (1982) RNA dependence in the cell cycle of V79 cells. J. Cell. Physiol. 112, 60. [DOI] [PubMed] [Google Scholar]
- Fujikawa‐Yamamoto K (1999a) Cell cycle analysis of DNA histograms in logarithmic scale. Cytometry Res. 9, 73. [Google Scholar]
- Fujikawa‐Yamamoto K, Ohdoi C, Yamagishi H, Zong Z, Murakami M, Yamaguchi N (1999b) Lack of synchrony among multiple nuclei induces partial DNA fragmentation in V79 cells polyploidized by demecolcine. Cell Prolifer. 32, 337. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Fujikawa‐Yamamoto K, Teraoka K, Odashima S (1993) Hyperploidization of V79 cells by K‐252a in comparison with demecolcine. Cell Struct. Funct. 18, 449. [DOI] [PubMed] [Google Scholar]
- Fujikawa‐Yamamoto K, Yamagishi H, Zong Z, Ohdoi C, Wang S (2000) Different responses of polyploidized V79 cells after removal of two drugs, demecolcine and K‐252a. Cell Struct. Funct. 25, 41. [DOI] [PubMed] [Google Scholar]
- Fujikawa‐Yamamoto K, Zong Z, Murakami M, Odashima S (1997) Different manner of DNA synthesis in polyploidizations of Meth‐A and B16F10 cell lines. Cell Struct. Funct. 22, 527. [DOI] [PubMed] [Google Scholar]
- Graves JA, McMillan J (1984) Control of DNA synthesis in polyploid mammalian cells. J. Cell Physiol. 121, 409. [DOI] [PubMed] [Google Scholar]
- Harris M (1971) Polyploid series of mammalian cells. Exp. Cell Res. 66, 329. [DOI] [PubMed] [Google Scholar]
- Jordan MA, Wendell K, Gardiner S, Derry WB, Copp H, Wilson L (1996) Mitotic block induced in HeLa cells by low concentrations of paclitaxel (Taxol) results in abnormal mitotic exit and apoptotic cell death. Cancer Res. 56, 816. [PubMed] [Google Scholar]
- Krishan A (1975) Rapid flow cytofluorometric analysis of mammalian cell cycle by propidium iodide staining. J. Cell Biol. 66, 188. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Moore J, Kieler J, Biczowa G (1968) Comparative studies of a near‐tetraploid and a near‐diploid line of Ehrlich's ascites tumor propagate in vivo and in vitro II. Cytology and transplantability. Eur. J. Cancer 4, 81. [DOI] [PubMed] [Google Scholar]
- Takahashi M (1987) A model for the spatio‐temporal organization of DNA replication in mammalian cells. J. Theol. Biol. 129, 91. [DOI] [PubMed] [Google Scholar]
- Takahashi M (1989a) A fractal model of chromosomes and chromosomal DNA replication. J. Theol. Biol. 141, 117. [DOI] [PubMed] [Google Scholar]
- Takahashi M (1989b) A model of chromatin‐dependent DNA replication sequences based on the decondensation units hypothesis. J. Theol. Biol. 136, 427. [DOI] [PubMed] [Google Scholar]
- Takahashi M (1996) A fractal‐like geometry of the nucleus and its reproduction: a hypothesis In: Takahashi M. ed. A genomic theory of the tumor cell cycle. 1st edn, pp. 1 – 265. Ube: Private Book. [Google Scholar]
- Usui T, Yoshida M, Abe K, Osada M, Isono K, Beppu T (1991) Uncoupled cell cycle without mitosis induced by a protein kinase inhibitor, K‐252a. J. Cell. Biol. 115, 1275. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Watanabe I, Okada S (1967) Effects of temperature on growth rate of cultured mammalian cells (L1578Y). J. Cell Biol. 32, 309. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Watters DJ, Beamish HJ, Marshall KA, Gardiner RA, Seymour GJ, Lavin MF (1994) Accumulation of HL‐60 leukemia cells in G2/M and inhibition of cytokinesis caused by two marine compounds, bistraten A and cycloxazoline. Cancer Chemother. Pharmacol. 33, 399. [DOI] [PubMed] [Google Scholar]
- Zhang Y, Wang Z, Ravid K (1996) The cell cycle in polyploid megakaryocytes is associated with reduced activity of cyclin B1‐dependent cdc2 kinase. J. Biol. Chem. 271, 4266. [DOI] [PubMed] [Google Scholar]
- Zollner H (1993) Handbook of Enzyme Inhibitors, 2nd edn, p. 813 Weinheim: VCH‐Verlagsgesellschaft Mbh. [Google Scholar]
- Zong Z, Fujikawa‐Yamamoto K, Tanino M, Yamagishi H, Gai X, Odashima S (1995) The important role of PKC in controlling polyploidy formation in cultured fibrosarcoma cell line. Biochem. Mol. Biol. Intern. 35, 1009. [PubMed] [Google Scholar]
- Zong Z, Fujikawa‐Yamamoto K, Teraoka K, Yamagishi H, Tanino M, Odashima S (1994) Potentiation of K‐252a, a protein kinase inhibitor‐induced polyploidization, by cAMP in cultured fibrosarcoma cell line. Biochem. Biophys. Res. Commun. 205, 746. [DOI] [PubMed] [Google Scholar]