

Abstract
Natural compounds derived from plant sources are well characterized as possessing a wide variety of remarkable anti‐tumour properties, for example modulating programmed cell death, primarily referring to apoptosis, and autophagy. Distinct from apoptosis, autophagy (an evolutionarily conserved, multi‐step lysosomal degradation process in which a cell destroys long‐lived proteins and damaged organelles) may play crucial regulatory roles in many pathological processes, most notably in cancer. In this review, we focus on highlighting several representative plant natural compounds such as curcumin, resveratrol, paclitaxel, oridonin, quercetin and plant lectin – that may lead to cancer cell death – for regulation of some core autophagic pathways, involved in Ras‐Raf signalling, Beclin‐1 interactome, BCR‐ABL, PI3KCI/Akt/mTOR, FOXO1 signalling and p53. Taken together, these findings would provide a new perspective for exploiting more plant natural compounds as potential novel anti‐tumour drugs, by targeting the pathways of autophagy, for future cancer therapeutics.
Introduction
Natural compounds originally derived from plants, have historically been regarded as an invaluable source of potential therapeutic agents, and are also are well known to play essential roles in a variety of human disease treatments, such as in cancer 1. Also, they have held (and still hold) important positions in primary health care in China, and have been recently recognized by Western countries as revealing more novel molecules for potential current cancer drug discovery 1. Cancer is a complex genetic disease with ten hallmark traits, acquired during their multistep development into human tumours, including: sustaining cell proliferative signalling, evading activity of cell population growth suppressors, enabling replicative immortality, inducing angiogenesis, activating cell invasion and metastasis, exhibiting genetic diversity, inflammation, re‐programming of energy metabolism, evading immune destruction and resisting programmed cell death (PCD) 2, 3.
Apoptosis, or type I PCD, is characterized by specific morphological and biochemical changes exhibited by dying cells, including: cell shrinkage, nuclear condensation and fragmentation, dynamic membrane blebbing and loss of cell‐cell adhesion. Biochemical changes include chromosomal DNA cleavage into inter‐nucleosomal fragments, phosphatidylserine (PS) externalization, and a number of intracellular substrate cleavages by proteolysis. As phagocytic cells can recognize and remove the apoptotic bodies, the inflammatory response is not stimulated by apoptotic cell death 4. Differing from apoptosis, autophagy, or type II PCD is an evolutionarily conserved catabolic process, which occurs when cells deliver their own cytoplasmic materials and organelles to lysosomes for degradation. In general, autophagy plays a crucial pro‐survival role in cell and tissue homeostasis, required during starvation or growth factor deprivation stress. However, accumulating evidence has revealed that autophagic cells may commit suicide by undergoing cell death and coping with excessive stress, which differs from apoptosis 5.
Hitherto, several natural compounds have been reported to modulate some autophagic signalling pathways (such as Ras/Raf/MAPK, Beclin‐1 complex, PI3KCI/Akt/mTOR, BCR‐ABL, FOXO1 signalling and p53), thus ultimately cumulating in cancer cell death 6. Here, we focus on summarizing about some selected plant natural compounds that can target autophagy in the ‘decisions’ of cancer cell death, which may provide more new clues towards exploiting them as potential anti‐tumour agents.
The autophagic process and monitoring methods
To initiate autophagy, intact organelles and/or cytoplasmic components enclosed in double‐membrane vacuoles, so‐called autophagosomes 6 mature by fusing with lysosomes, leading to the formation of autophagolysosomes 7. This process includes progressive steps for autophagic vacuole production and turnover, including autophagic induction, vesicle nucleation, vesicle elongation and completion, docking and fusion, followed by degradation and recycling (Fig. 1).
Figure 1.
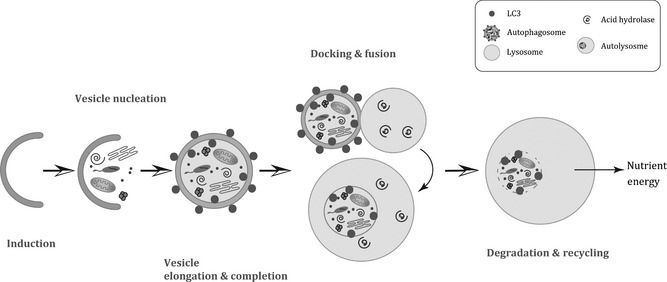
Stages of the process of autophagy. Autophagy can be stimulated by nutrient deprivation, hypoxia, cytokine activity, hormone activity and DNA damage. (a) Induction requires Atg 1 and Atg13, which in turn can be inhibited by mTOR. (b) Vesicle nucleation depends on Beclin‐1‐class III PI3K‐Vps15 core complexes and other proteins. (c) Vesicle elongation and completion mediated by Atg16L complex and LC3. (d) Docking and fusion refer to maturation of autolysosomes promoted by Rab7, LAMP1, LAMP2, SKD1, Vtil 1b and ESCRT complex. (e) In the final stage of degradation and recycling of components, autophagosomal cargoes are digested, and nutrients and inherent energy are recycled.
To date, some methods have been developed for monitoring autophagy in higher eukaryotes, of which observation by electron microscopy is the most dependable approach, by directly detecting cytoplasmic accumulation of autophagosomes 8, 9. In addition, some members of the Atg family have become general biomarkers for autophagy. Amongst them, LC3 is the mammalian homologue of Atg8, which undergoes ubiquitin‐like post‐translational modification that can target it to the isolation plasmamembrane. Localization of LC3 can easily be examined by generating chimaeric proteins fused to green fluorescent protein (GFP), and examination of GFP‐LC3 localization is simple, as it requires no more than high‐resolution fluorescence microscope 9. Moreover, the C‐terminal fragment of LC3 can be cleaved with synthesis following, to yield a cytosolic form called LC3‐I and later, a subpopulation of LC3‐I can be converted into an autophagosome‐associating form, LC3‐II 10. With immunoblotting of both LC3‐I and LC3‐II, activity of autophagy can be detected easily in mammalian cells. Beclin‐1, an orthologue of yeast Atg6, can play an important role in regulating autophagy. Levels of Beclin‐1 in mammalian cells can help determination of autophagic activity by western blotting 11 although some chemical compounds, such as 3‐MA, Wortmannin and LY294002 are found to inhibit autophagy 12.
Core pathways of autophagy in cancer
Under low‐oxygen and low‐nutrient conditions, autophagy promotes tumour progression, by contributing to cancer cell survival 13, while in contrast under normal conditions, autophagy may function as a tumour suppressor, playing a cell death‐promoting role as a bona fide tumour cell death mechanism 14. These paradoxical investigation results, however, can seem to be confused, dependent on different types of tissue‐specific cancer cells, thereby regulating either pro‐survival or pro‐death signalling pathways 15, 16. At the molecular level, autophagy plays a ‘Janus role’, by regulation of oncogenes and/or tumour suppressors (Fig. 2).
Figure 2.
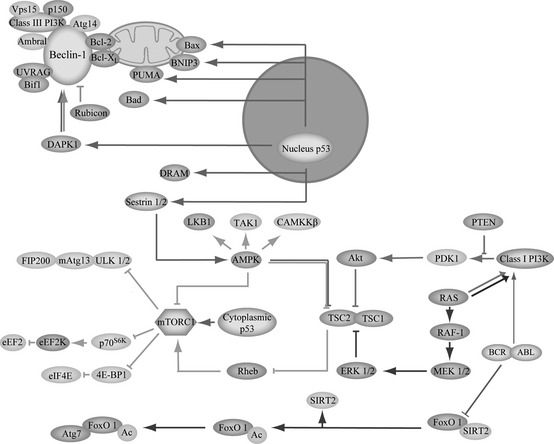
Core autophagic signalling pathways in cancer cells. Autophagy may decide the ultimate fate of cancer cells for regulation of key survival or death signalling pathways, such as Beclin‐1 complex, PI3K/Akt/mTOR, Ras/Raf/MAPK, BCR‐ABL, p53 and FOXO1 signalling in many types of cancer cells.
Oncogenic autophagic pathways
On the one hand, oncogenic pathways involved in PI3KCI, Akt, mTOR, Ras, BCR‐ABL, Bcl‐2 and Bcl‐XL can play important roles in ‘deciding’ cancer cell survival. Amongst them, the mammalian target of rapamycin (mTOR) may serve as a main negative regulator of autophagy in cancer cells. mTOR exists in two functionally different complexes in mammalian cells; of these, only mammalian target of rapamycin complex 1 (mTORC1) is sensitive to rapamycin 13. PI3KCI‐Akt, Ras‐Raf‐1‐MEK1/2‐ERK1/2 signalling pathways can activate mTORC1 yet on the contrary, the liver kinase B1 (LKB1)‐AMP‐activated protein kinase (AMPK) pathway can inhibit mTORC1 13. In addition, mTORC1 can negatively regulate autophagy when competed with ULK1 (Atg1) and mAtg13 that can be positively regulated by its upstream PI3KCI‐Akt‐1 pathway, suggesting that oncogenic action of constitutively active PI3KCI‐Akt‐mTORC1 may lead to inhibition of autophagy 17. Additionally, constitutively active Akt can inhibit induction of autophagy 15 and Ras can converge on activating mTOR for stimulating cell proliferation and inhibiting autophagy. Sustained Ras‐Raf‐MAPK may be required for tumour survival by autophagy and additionally, BCR‐ABL (product of a further oncogene), can serve as the key factor for stimulating transcription of mTOR via PI3KCI‐Akt‐FOXO signalling, in chronic myeloid leukaemia (CML), suggesting an inhibitory role of the autophagy process 18. Bcl‐2 and Bcl‐XL can play key roles by mediating autophagic signalling pathways 19 (Fig. 2).
Tumour suppressive autophagic pathways
Tumour suppressive pathways include various regulators, such as Beclin‐1 and p53 which can exert ambiguous functions for regulation of autophagy, acting as haploinsufficient tumour suppressor proteins 20, 21. Beclin‐1 can cooperate with Ambra‐1, Bif‐1 and UVRAG to activate lipid kinase Vps34, thereby inducing autophagy 17. However, it can suppress tumour function in the lysosomal degradation pathway of autophagy. p53, interacting with Bad, Noxa and PUMA, plays a distinct role in autophagy, depending on its different subcellular localizations 21. Nuclear p53 can promote autophagy through interacting with its targets, DRAM and sestrin1/2, whereas, cytoplasmic p53 can inhibit autophagy by activating Bax, and/or inhibiting Bcl‐2/Bcl‐XL (Fig. 2).
Cytosolic FOXO1, a fork‐head O family protein, is a mediator of autophagy. Under conditions of stress, FOXO1 dissociates from a complex with sirtuin‐2 (SIRT2) followed by its acetylation; acetylation status of FOXO1 is balanced by histone acetylases and HDAC. While SIRT1 is located in the nucleus, SIRT2 is primarily located in the cytosol and is the main deacetylase of cytosolic FOXO1 22. Then, acetylated FOXO1 binds to E1‐like Atg7 (Atg7 plays an important role in ubiquitin‐like conjugation (Ublc) systems for autophagosome expansion). The FOXO1‐Atg7 complex influences the autophagic process, which ultimately culminates in cell death, and FOXO1‐modulated cell death may be associated with tumour suppressor activity 22, 23 (Fig. 2). Accordingly, below we discuss in more detail, some representative natural compounds of plant origin that can result in cancer cell death by targeting the aforementioned autophagic signalling pathways, in cancer pathology and therapy.
Plant natural compound‐induced autophagy in cancer
Curcumin
Curcumin, derived from turmeric (Curcumin longa), chemically is known as diferuloylmethane (C21H20O6) 24 (Fig. 3a). As an active ingredient from the rhizome in the plant, curcumin has been shown to demonstrate activity in a wide range of biological functions, particularly against cancer 25, 26, 27, 28, 29, 30. Moreover, curcumin has been reported to regulate autophagy, resulting in inhibition of several types of cancer cell proliferation (for example, in chronic myeloid leukaemia, malignant glioma and oesophageal cancer cells) 30, 31, 32. After treatment with curcumin, levels of LC3‐II and Beclin‐1 have been detected to be elevated in K562 cells, indicating that this compound may affect autophagosome formation 30. In U87‐MG and U373‐MG cells, curcumin has been found to inhibit the Akt/p70S6 kinase pathway, as well as to activate ERK1/2, resulting in autophagic induction, whereas PI3K activity was not affected 33. Intriguingly, reactive oxygen species (ROS), upregulating Beclin‐1 and p53, have been found to be induced by curcumin in activation of autophagy, thereby culminating in concomitant cell death of HCT 116 human colon cancer cells 34. Additionally, serine/threonine protein phosphatases type‐1 (PP1) and type‐2A (PP2A) are key players in phosphorylation and dephosphorylation and curcumin can downregulate PP1, which might stimulate phosphorylation of ERK 31. Besides these autophagy‐inducing activities, curcumin has been observed to inhibit viability of K562 cells in a dose‐ and time‐dependent manner. Curcumin‐induced cell death has been found to be associated with formation of the apoptosome complex, collapse of the mitochondrial membrane potential (MMP) and caspase‐3 activation. Also, curcumin treatment has induced Bid cleavage and downregulated expression of Bcl‐2 protein in K562 cells 30. In addition, curcumin has been shown to enhance adriamycin‐induced human liver‐derived hepatoma G2 cell death through activation of mitochondria‐mediated apoptosis and autophagy 35.
Figure 3.
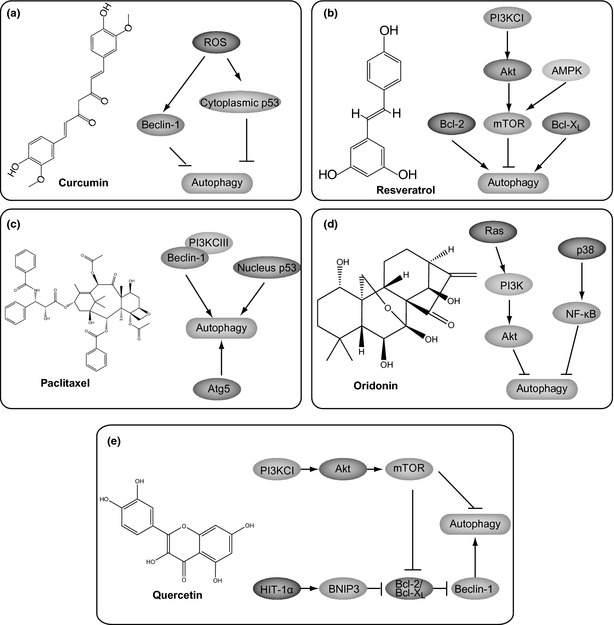
Representative natural small molecules targeting autophagic pathways. (a) Curcumin; (b) Resveratrol; (c) Paclitaxe; (d) Oridonin; (e) Quercetin.
Resveratrol
Resveratrol (Res), a phytoalexin found in grapes and root extracts of Polygonum cuspidatum, has been demonstrated to result in a wide range of biological effects including anti‐proliferative activity 36 (Fig. 3b). In a murine investigation in 1997, topical application of Res was reported to reduce numbers of skin tumours per mouse by up to 98%, which triggered research on Res 37. It is notable that Res can inhibit initiation and growth of tumours in a wide range of cancer models, such as cell population growth in leukaemias as well as in prostate, breast and further epithelial cancers 35. When treated with Res, Bcl‐2 and Bcl‐XL were found to be involved in autophagy, thus mediating a response to this agent, in ovarian A2780 cancer cells 35. Beclin‐1 promotes autophagy when overexpressed in MCF‐7 cells, and levels of typical autophagic protein Atg7 (Beclin‐1) remain unchanged after Res treatment. It has been suggested that Res, unlike many kinds of natural compound that rely on activation of Beclin‐1 for regulating autophagy, can induce Beclin‐1‐independent autophagy in breast cancer cells 38. Res‐induced autophagy has been reported to inhibit Akt/protein kinase B (PKB) phosphorylation and mTOR signalling by phosphorylation of p70S6K, a direct mTOR substrate, indicating that Res‐induced autophagy is dependent on mTOR inhibition. Moreover, N‐acetyl‐sphingosine (C2‐Cer)‐induced autophagy is characterized by inhibition of Akt/mTOR signalling in MCF‐7casp−3 and MCF‐7vc cells 39. As mTOR can upregulate Akt and AMPK, the latter can play their key roles in Res‐induced autophagy; thus Res can induce autophagy by modulating activities of Akt and AMPK in autophagy 31, 40. Res can induce autophagy in gastric cancer cells after increase in dihydroceramides, the immediate ceramide metabolic precursor in de novo sphingolipid synthesis 41. However, some studies have also suggested that resveratrol may potentially be useful in cancer chemotherapy, when used in combination with other drugs, in hepatocellular carcinoma and leukaemia cell lines, mainly due to its effect on apoptosis 42.
Paclitaxel
Paclitaxel, originally isolated from the bark of the Pacific yew tree, is a widely used agent for treatments of lung, ovarian, breast and head and neck cancers 43 (Fig. 3c). Due to its notable effects, paclitaxel‐based combination therapies have been noted as standard treatments for many patients diagnosed with non‐small cell lung carcinoma 44. As paclitaxel may generate unacceptable levels of toxicity to normal cells in clinical settings, more experiments need to be focused on how to enhance its effectiveness, as well as to reduce toxicity. During this process, paclitaxel has been found to induce autophagy 45. LC3‐II, an ingredient of the autophagosome, has been shown to be elevated in paclitaxel‐treated A549 cancer cells, suggesting a role for it in autophagy 46, 47. Atg5 and Beclin‐1 are also required for autophagosome formation, and paclitaxel treatment can lead to increase in expression levels of Atg5 and Beclin‐1 in a dose‐dependent manner in the A549 cells 46. Beclin‐1, combining with PI3KCIII, can induce autophagy at the initiation stage, suggesting the positive role of Beclin‐1 in autophagy, to inhibit tumourigenesis 48. Additionally, paclitaxel treatment has been found to induce autophagy by detection of GFP‐LC3 punctate formation in human U87 glioma cells, human PC‐3 prostate cancer and colon HT‐29 cancer cells, transfected with an expression plasmid containing GFP‐LC3 49. Moreover, paclitaxel can increase p53 expression before regulation of LC3B, thus regulating induction of autophagy 50. Paclitaxel‐caused autophagy has also been shown to play a critical role in mediating caspase‐independent cancer cell death 51, 52. Interestingly, paclitaxel resistance has been shown to be associated with profound changes in the cell death response, with deletion of multiple apoptotic factors balanced by upregulation of the autophagic pathway, and collateral sensitivity to platinum 53.
Oridonin
Oridonin (7, 20‐epoxy‐ent‐kauranes), a diterpenoid isolated from the medicinal herb Rabdosia rubescens, was discovered in 1967 – and subsequently synthesized in 1973 54, 55, 56. Oridonin has been found to exhibit remarkable anti‐proliferative activity, particularly inhibiting tumour growth, thus leading to cancer cell death of melanoma and cervical carcinoma cells 54, 57 (Fig. 3d). Some transcriptional factors in apoptosis and autophagy can be regulated by oridonin, which inhibit tumour cell proliferation, indicating that oridonin can have both apoptosis‐ and autophagy‐inducing activities maybe in many types of cancer cell 57. The PI3KCI/Akt signalling pathway is found to switch between apoptosis and autophagy, which is frequently found to be implicated in cancer cells. In oridonin‐induced HeLa cell death, PI3KCI/Akt signalling has been found to be augmented in autophagy, as well as to suppress apoptosis, and in this context, autophagy might serve as a survival mechanism to protect cells from apoptotic death 58. When treated with oridonin, autophagy can be significantly upregulated with manumycin A, a Ras‐specific inhibitor. Treatment with a combination of oridonin and manumycin A can downregulate phosphorylation of downstream PI3KCI, and Akt 58. Thus, associated with activation of PI3KCI and Akt, Ras can negatively regulate autophagy in oridonin‐treated cancer cells, suggesting that oridonin may induce autophagy via negatively regulating Ras/PI3KCI/Akt 59. In addition, NF‐κB can function as a molecular switch in a similar way to PI3KCI/Akt, by which autophagy can protect cancer cells from death, by upregulating the p38‐NFκB survival pathway, or facilitating autophagy in oridonin‐treated human HT1080 sarcoma cells by p53 activation 60. However, under oridonin treatment, expression of Beclin‐1 (which leads to autophagy‐inducing activity), remained at the same level, suggesting that Beclin‐1 does not participate in oridonin‐induced autophagy 58.
Oridonin also has been well characterized to induce apoptosis in many types of cancer cell. In oridonin‐induced HeLa cell death, PI3K/Akt signalling has been found to be augmented in autophagy, but suppressed in apoptosis; thus autophagy may serve as a survival mechanism to protect cells from apoptotic death. In a further scenario, NF‐κB has been found to function as a molecular switch in a similar way, by which autophagy could protect cells from apoptosis through upregulating the p38‐NF‐κB survival pathway 56. A further study has demonstrated that NF‐κB facilitated both apoptosis and autophagy in oridonin‐treated human HT1080 fibrosarcoma cells by activating p53. In oridonin‐treated MCF‐7 cells, autophagy contributed to cell death by its synergic effects on apoptosis, and ERK was found to be inhibited, whereas JNK and p38 were provoked into action 56.
Quercetin
Quercetin represents the most abundant dietary flavonoid found in a broad range of fruits, vegetables and beverages, whose antioxidant and anti‐inflammatory properties have been associated with prevention of, and therapy for, cancer. It has been known to be a promising cancer chemopreventive agent that inhibits tumour promotion by inducing cell cycle arrest and promoting apoptotic cell death 61. Quercetin triggers accumulation of HIF‐1α, which represses mTOR signalling and induces expression of BNIP3/BNIP3L, to disrupt the Beclin‐1/Bcl‐2 (Bcl‐XL) complex, leading to activation of Atg7, Atg12‐Atg5 conjugate and LC3 turnover, in gastric cancer 62 (Fig. 3e). In addition, quercetin has been reported to inhibit cell viability as well as cancer cell properties such as anchorage‐independent cell population growth. It can mediate preferential degradation of oncogenic Ras, thus causing autophagy in Ha‐RAS‐transformed human colon cells 63 (Fig. 3e).
Interestingly, quercetin has been reported to bypass death receptor resistance through multiple mechanisms, in HPB‐ALL, a human acute lymphocytic leukaemia cell line. Quercetin however, can induce apoptosis only when present in combination with a CD95 agonist antibody. At the molecular level, quercetin can reduce intracellular ROS, the reduced MMP, thereby leading to expression of the CD95 receptor remaining unchanged 64.
Plant lectins
Plant lectins, a class of highly diverse non‐immune‐origin proteins, containing at least one non‐catalytic domain, can selectively recognize and reversibly bind to specific free sugars or glycans present on glycoproteins and glycolipids 8, 65. Galanthus nivalis agglutinin (GNA)‐related lectins and legume lectins have been widely reported to induce cancer cell death by targeting autophagy and apoptosis 65. Polygonatum cyrtonema lectin (PCL), a mannose/sialic acid plant lectin belonging to the GNA‐related lectin family, was first isolated from the rhizomes of Polygonatum cyrtonema Hua, a traditional Chinese medicinal herb, in 1996 66 (Fig. 4a). After its isolation, more physiological and biochemical characteristics of PCL have been revealed with respect to cancer, including breast and cervical carcinomas 67, 68, 69. Recently, PCL has been reported to induce autophagic death via a mitochondria‐mediated pathway, in human A375 melanoma cells, and such PCL‐induced autophagic death was further confirmed to be a mitochondrial‐mediated ROS–p38–p53 pathway 69. In addition, inhibition of Ras could promote L929 cell death, suggesting that the Ras–Raf signalling pathway may play the role of a key negative regulator of PCL‐induced autophagy 23. Moreover, the PI3KCI–Akt signalling pathway has been shown to play the role of a negative regulator in PCL‐induced autophagy 1. Thus, the aforementioned evidence has demonstrated that PCL can induce cancer cell autophagy by promoting the ROS–p38–p53 pathway, as well as by blocking Ras–Raf and PI3K–Akt signalling pathways, in different types of cancer cells 70. Moreover, PCL has also been reported to induce cancer cell apoptosis of HeLa, MCF‐7, A375 and L929 cells, but with concomitant low toxicity to melanocytes cells. Further studies demonstrate that PCL induces cancer cell apoptosis by activating the caspase‐dependent pathway and mitochondrial ROS‐p38‐p53 pathway, as well as through blocking Ras‐Raf and PI3K‐Akt pathways 68.
Figure 4.
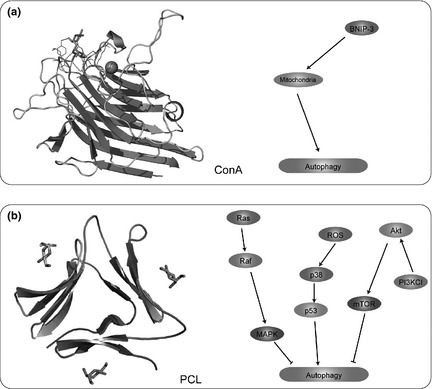
Representative natural macromolecules targeting autophagic pathways. (a) Concanavalin A (ConA); (b) Polygonatum cyrtonema lectin (PCL).
Concanavalin A (ConA) from the jack bean, was in 1936, the first legume lectin to be purified and crystallized; it is the most well‐known representative of the legume lectin family 71, 72, 73 (Fig. 4b). After associating with the mannose moiety residing on cell membrane glycoprotein, ConA is preferentially internalized and initially transported to mitochondria via clathrin‐mediated endocytosis, before autophagic cell death is initiated 74, with the observation of marked morphological changes after ConA intervention 75. ConA has been treated into Chang MHCC97‐H liver cells 75 and also, experimental data have shown that it can induce autophagic cell death in hepatoma cells using the adenovirus Bcl2/adenovirus E1B 19kDa‐interacting protein 3 (BNIP‐3)‐mediated pathway 76. In addition to its autophagy‐inducing activities, ConA has been reported to induce different types of cancer cell death by mitochondria‐mediated pathways, p73‐FOXO1a‐Bim apoptotic pathways, as well as IKK‐NF‐κB‐COX‐2, SHP‐2‐MEK‐1‐ERK and SHP‐2‐Ras‐ERK anti‐angiogenic pathways 76.
Other plant natural compounds
Recently, other natural compounds from plants have been found to induce cancer cell death by regulation of autophagy. Spermidine, a ubiquitous polycation synthesized from putrescine, has been widely accepted as a potential drug to promote cell longevity 77. Recent reports have demonstrated its crucial role for regulating autophagy as it can enhance transactivation of autophagy‐related genes such as Atg7, Atg8, Atg11 and Atg15, in various types of cancer cells 78. Genistein, a natural isoflavenoid found in soybean products, has been reported to have therapeutic potential in multiple tumour types, including prostate, breast, colon, gastric, lung and pancreatic adenocarcinomas, and in lymphoma. Genistein exerts its anti‐tumour effect by inhibition of Akt activation, suggesting that it can help cause cancer cell death through autophagocytosis 79 and chloroquine (CQ), traditionally used as an anti‐malarial medication, has been reported to induce autophagy, resulting in glioma cell death through a p53‐independent pathway 80, 81.
Concluding remarks
Cancer, the leading cause of death nowadays worldwide, attracts a great deal of attention. Accumulating lines of evidence have recently revealed that targeting autophagic signalling pathways may be a promising avenue for potential therapeutic purposes. These aforementioned oncogenic/tumour suppressive signalling pathways implicated in Beclin‐1 complex, PI3KCI/Akt/mTOR, BCR‐ABL, Ras/Raf/MAPK, FOXO1 signalling and p53 may play crucial roles in autophagy‐related cancer cell death networks. Interestingly, some natural compounds of plant origin, such as curcumin, resveratrol, paclitaxe, oridonin, quercetin and plant lectins, can induce these autophagic pathways as potential therapeutic agents to deal with carcinogenesis (Table S1).
With the accumulating exploration of molecular mechanisms of plant natural compound activity, they have been widely used as candidate anti‐tumour agents; more importantly, some have been further applied to pre‐clinical and clinical cancer therapies by using apoptotic pathways. Even though apoptosis and autophagy bear distinct morphological characteristics and physiological processes, there still exist some intricate interrelationships between them. Under some circumstances, apoptosis and autophagy can exert synergistic effects, while other times autophagy can be triggered only when apoptosis is suppressed. Moreover, recent studies have pointed out that apoptosis and autophagy may be interconnected and even simultaneously regulated by the same triggers in cancer.
Due to the complex two‐faced nature of autophagy, establishing its dual role in tumour cell survival versus death may help in determining cancer therapeutic potential. Inhibiting autophagy may enhance efficacy of currently used anti‐cancer drugs in chemo‐ and radiotherapy‐induced activation of autophagic signalling pathways, which may augment anti‐tumour activity, and thus efficacy of radiation and/or anti‐cancer drugs. On the other hand, promoting autophagy may induce autophagy‐inducing cancer cell death with high thresholds to apoptosis.
Intriguingly, there is a novel emerging viewpoint concerning disruption of protein–protein interactions (PPIs), and with increasing alacrity, more plant natural compounds that inhibit or promote the PPIs can be further screened as novel candidate anti‐cancer drugs. Thus, the best hope for cancer therapeutics may lie in the discovery of more novel natural compounds, targeting the physiological effects of core autophagy pathways, and even autophagic networks in cancer cells, rather than their individual gene or protein components. In summary, both strategies have significant potential for translation into ongoing clinical trials that may provide more valuable information on whether and how these natural plant compounds target autophagy pathways to make better sense in cancer treatments.
Supporting information
Table. S1 Representative plant natural compounds targeting autophagy as anti‐cancer agents
Acknowledgements
We are grateful to Ming‐wei Min (University of Cambridge), Qian Liu (National University of Singapore) and He‐jiao Bian (Boston University) for their critical reviews on this manuscript. We also thank Chun‐yang Li, Huai‐long Xu, Zheng Shi, Lei‐lei Fu, Xu Zhao, Cheng‐cheng Zhou, Xin Wen, Shu‐ya Wang, Shu Yao, Miao Ming, Jia‐ying Yu and Qi‐jia Yu (Sichuan University) for their good suggestions on this work.
Additionally, this work was supported in part by grants from the Young Teachers’ Fund of Sichuan University (No. 2010SCU11066), the Science Foundation for Post Doctorate Research of China (No. 20110491725) and the Major State Basic Research Development Program of China (973 Program) (No. 2010cb529900).
References
- 1. Liu B, Cheng Y, Liu Q, Bao JK, Yang JM (2010) Autophagic pathways as new targets for cancer drug development. Acta Pharmacol. Sin. 31, 1154–1164. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 2. Jemal A, Bray F, Center MM, Ferlay J, Ward E, Forman D (2011) Global cancer statistics. CA Cancer J. Clin. 61, 69–90. [DOI] [PubMed] [Google Scholar]
- 3. Hanahan D, Weinberg RA (2011) Hallmarks of cancer: the next generation. Cell 144, 646–674. [DOI] [PubMed] [Google Scholar]
- 4. Liu JJ, Lin M, Yu JY, Liu B, Bao JK (2011) Targeting apoptotic and autophagic pathways for cancer therapeutics. Cancer Lett. 300, 105–114. [DOI] [PubMed] [Google Scholar]
- 5. Amaravadi RK, Lippincott‐Schwartz J, Yin XM, Weiss WA, Takebe N, Timmer W et al (2011) Principles and current strategies for targeting autophagy for cancer treatment. Clin. Cancer Res. 17, 654–666. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 6. Wen X, Lin ZQ, Liu B, Wei YQ (2012) Caspase‐mediated programmed cell death pathways as potential therapeutic targets in cancer. Cell Prolif. 45, 217–224. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 7. Eskelinen EL (2011) The dual role of autophagy in cancer. Curr. Opin. Pharmacol. 11, 294–300. [DOI] [PubMed] [Google Scholar]
- 8. Mizushima N (2004) Methods for monitoring autophagy. Int. J. Biochem. Cell Biol. 36, 2491–2502. [DOI] [PubMed] [Google Scholar]
- 9. Galluzzi L, Aaronson SA, Abrams J, Alnemri ES, Andrews DW, Baehrecke EH et al (2009) Guidelines for the use and interpretation of assays for monitoring cell death in higher eukaryotes. Cell Death Differ. 16, 1093–1107. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 10. Kabeya Y, Mizushima N, Yamamoto A, Oshitani‐Okamoto S, Ohsumi Y, Yoshimori T (2004) LC3, GABARAP and GATE16 localize to autophagosomal membrane depending on form‐II formation. J. Cell Sci. 117, 2805–2812. [DOI] [PubMed] [Google Scholar]
- 11. Kepp O, Galluzzi L, Lipinski M, Yuan J, Kroemer G (2011) Cell death assays for drug discovery. Nat. Rev. Drug Discov. 10, 221–237. [DOI] [PubMed] [Google Scholar]
- 12. Klionsky DJ, Fabio CA, Hagai A, Robert TA, Abraham AA, Khosrow A et al (2012) Guidelines for the use and interpretation of assays for monitoring autophagy. Autophagy 8, 1–100.22082964 [Google Scholar]
- 13. Wang SY, Yu QJ, Zhang RD, Liu B (2011) Core signaling pathways of survival/death in autophagy‐related cancer networks. Int. J. Biochem. Cell Biol. 43, 1263–1266. [DOI] [PubMed] [Google Scholar]
- 14. Kondo Y, Kanzawa T, Sawaya R, Kondo S (2005) The role of autophagy in cancer development and response to therapy. Nat. Rev. Cancer 5, 726–734. [DOI] [PubMed] [Google Scholar]
- 15. Mathew R, Karantza‐Wadsworth V, White E (2007) Role of autophagy in cancer. Nat. Rev. Cancer 7, 961–967. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 16. Gozuacik D, Kimchi A (2004) Autophagy as a cell death and tumor suppressor mechanism. Oncogene 23, 2891–2906. [DOI] [PubMed] [Google Scholar]
- 17. Brech A, Ahlquist T, Lothe RA, Stenmark H (2009) Autophagy in tumour suppression and promotion. Mol. Oncol. 3, 366–375. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 18. Kang R, Zeh HJ, Lotze MT, Tang D (2011) The Beclin 1 network regulates autophagy and apoptosis. Cell Death Differ. 18, 571–580. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 19. Maiuri MC, Tasdemir E, Criollo A, Morselli E, Vicencio JM, Carnuccio R et al (2009) Control of autophagy by oncogenes and tumor suppressor genes. Cell Death Differ. 16, 87–93. [DOI] [PubMed] [Google Scholar]
- 20. Miracco C, Cosci E, Oliveri G, Luzi P, Pacenti L, Monciatti I et al (2007) Protein and mRNA expression of autophagy gene Beclin 1 in human brain tumours. Int. J. Oncol. 30, 429–436. [PubMed] [Google Scholar]
- 21. Ryan KM (2011) p53 and autophagy in cancer: guardian of the genome meets guardian of the proteome. Eur. J. Cancer 47, 44–50. [DOI] [PubMed] [Google Scholar]
- 22. Ling LU, Tan KB, Lin H, Chiu GN (2011) The role of reactive oxygen species and autophagy in safingol‐induced cell death. Cell Death Dis. 2, e129. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 23. Zhao Y, Wang L, Yang J, Zhang P, Ma K, Zhou JY (2010) Anti‐neoplastic activity of the cytosolic FoxO1 results from autophagic cell death. Autophagy 6, 988–990. [DOI] [PubMed] [Google Scholar]
- 24. Campbell FC, Collett GP (2005) Chemopreventive properties of curcumin. Future Oncol. 1, 405–414. [DOI] [PubMed] [Google Scholar]
- 25. Oyama Y, Masuda T, Nakata M, Chikahisa L, Yamazaki Y, Miura K et al (1998) Protective actions of 5′‐n‐alkylated curcumins on living cells suffering from oxidative stress. Eur. J. Pharmacol. 360, 65–71. [DOI] [PubMed] [Google Scholar]
- 26. Pal S, Choudhuri T, Chattopadhyay S, Bhattacharya A, Datta GK, Das T et al (2001) Mechanisms of curcumin‐induced apoptosis of Ehrlich's ascites carcinoma cells. Biochem. Biophys. Res. Commun. 288, 658–665. [DOI] [PubMed] [Google Scholar]
- 27. Pal S, Bhattacharyya S, Choudhuri T, Datta GK, Das T, Sa G (2005) Amelioration of immune cell number depletion and potentiation of depressed detoxification system of tumor‐bearing mice by curcumin. Cancer Detect. Prev. 29, 470–478. [DOI] [PubMed] [Google Scholar]
- 28. Sharma RA, Gescher AJ, Steward WP (2005) Curcumin: the story so far. Eur. J. Cancer 41, 1955–1968. [DOI] [PubMed] [Google Scholar]
- 29. Sugimoto K, Hanai H, Tozawa K, Aoshi T, Uchijima M, Nagata T et al (2002) Curcumin prevents and ameliorates trinitrobenzene sulfonic acid‐induced colitis in mice. Gastroenterology 123, 1912–1922. [DOI] [PubMed] [Google Scholar]
- 30. Jia YL, Li J, Qin ZH, Liang ZQ (2009) Autophagic and apoptotic mechanisms of curcumin‐induced death in K562 cells. J. Asian Nat. Prod. Res. 11, 918–928. [DOI] [PubMed] [Google Scholar]
- 31. Aoki H, Takada Y, Kondo S, Sawaya R, Aggarwal BB, Kondo Y (2007) Evidence that curcumin suppresses the growth of malignant gliomas in vitro and in vivo through induction of autophagy: role of Akt and extracellular signal‐regulated kinase signaling pathways. Mol. Pharmacol. 72, 29–39. [DOI] [PubMed] [Google Scholar]
- 32. O'Sullivan‐Coyne G, O'Sullivan GC, O'Donovan TR, Piwocka K, McKenna SL (2009) Curcumin induces apoptosis‐independent death in oesophageal cancer cells. Br. J. Cancer 101, 1585–1595. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 33. Shinojima N, Yokoyama T, Kondo Y, Kondo S (2007) Roles of the Akt/mTOR/p70S6K and ERK1/2 signaling pathways in curcumin‐induced autophagy. Autophagy 3, 635–637. [DOI] [PubMed] [Google Scholar]
- 34. Lee YJ, Kim NY, Suh YA, Lee C (2011) Involvement of ROS in curcumin‐induced autophagic cell death. Korean J. Physiol. Pharmacol. 15, 1–7. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 35. Qian H, Yang Y, Wang XF (2011) Curcumin enhanced adriamycin‐induced human liver‐derived Hepatoma G2 cell death through activation of mitochondria‐mediated apoptosis and autophagy. Eur. J. Pharm. Sci. 43, 125–131. [DOI] [PubMed] [Google Scholar]
- 36. Opipari AW Jr, Tan L, Boitano AE, Sorenson DR, Aurora A, Liu JR (2004) Resveratrol‐induced autophagocytosis in ovarian cancer cells. Cancer Res. 64, 696–703. [DOI] [PubMed] [Google Scholar]
- 37. Jang M, Cai L, Udeani GO, Slowing KV, Thomas CF, Beecher CW et al (1997) Cancer chemopreventive activity of resveratrol, a natural product derived from grapes. Science 275, 218–220. [DOI] [PubMed] [Google Scholar]
- 38. Scarlatti F, Maffei R, Beau I, Codogno P, Ghidoni R (2008) Role of non‐canonical Beclin 1‐independent autophagy in cell death induced by resveratrol in human breast cancer cells. Cell Death Differ. 15, 1318–1329. [DOI] [PubMed] [Google Scholar]
- 39. Scarlatti F, Bauvy C, Ventruti A, Sala G, Cluzeaud F, Vandewalle A et al (2004) Ceramide‐mediated macroautophagy involves inhibition of protein kinase B and up‐regulation of beclin 1. J. Biol. Chem. 279, 18384–18391. [DOI] [PubMed] [Google Scholar]
- 40. Ohshiro K, Rayala SK, El‐Naggar AK, Kumar R (2008) Delivery of cytoplasmic proteins to autophagosomes. Autophagy 4, 104–106. [DOI] [PubMed] [Google Scholar]
- 41. Signorelli P, Munoz‐Olaya JM, Gagliostro V, Casas J, Ghidoni R, Fabrias G (2009) Dihydroceramide intracellular increase in response to resveratrol treatment mediates autophagy in gastric cancer cells. Cancer Lett. 282, 238–243. [DOI] [PubMed] [Google Scholar]
- 42. Wu SL, Sun ZJ, Yu L, Meng KW, Qin XL, Pan CE (2004) Effect of resveratrol and in combination with 5‐FU on murine liver cancer. World J. Gastroenterol. 10, 3048–3052. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 43. Wessely R, Schomig A, Kastrati A (2006) Sirolimus and Paclitaxel on polymer‐based drug‐eluting stents: similar but different. J. Am. Coll. Cardiol. 47, 708–714. [DOI] [PubMed] [Google Scholar]
- 44. Rowinsky EK, Donehower RC (1995) Paclitaxel (taxol). N. Engl. J. Med. 332, 1004–1014. [DOI] [PubMed] [Google Scholar]
- 45. Horwitz SB (1992) Mechanism of action of taxol. Trends Pharmacol. Sci. 13, 134–136. [DOI] [PubMed] [Google Scholar]
- 46. Kabeya Y, Mizushima N, Ueno T, Yamamoto A, Kirisako T, Noda T et al (2000) LC3, a mammalian homologue of yeast Apg8p, is localized in autophagosome membranes after processing. EMBO J. 19, 5720–5728. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 47. Xi G, Hu X, Wu B, Jiang H, Young CY, Pang Y et al (2011) Autophagy inhibition promotes paclitaxel‐induced apoptosis in cancer cells. Cancer Lett. 307, 141–148. [DOI] [PubMed] [Google Scholar]
- 48. Levy JM, Thorburn A (2011) Targeting autophagy during cancer therapy to improve clinical outcomes. Pharmacol. Ther. 131, 130–141. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 49. Klionsky DJ, Codogno P, Cuervo AM, Deretic V, Elazar Z, Fueyo‐Margareto J et al (2010) A comprehensive glossary of autophagy‐related molecules and processes. Autophagy 6, 438–448. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 50. Hayashi S, Yamamoto A, You F, Yamashita K, Ikegame Y, Tawada M et al (2009) The stent‐eluting drugs sirolimus and paclitaxel suppress healing of the endothelium by induction of autophagy. Am. J. Pathol. 175, 2226–2234. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 51. Apel A, Zentgraf H, Buchler MW, Herr I (2009) Autophagy – a double‐edged sword in oncology. Int. J. Cancer 125, 991–995. [DOI] [PubMed] [Google Scholar]
- 52. Yu L, Lenardo MJ, Baehrecke EH (2004) Autophagy and caspases: a new cell death program. Cell Cycle 3, 1124–1126. [PubMed] [Google Scholar]
- 53. Ajabnoor GMA, Crook T, Coley HM (2012) Paclitaxel resistance is associated with switch from apoptotic to autophagic cell death in MCF‐7 breast cancer cells. Cell Death Dis. 3, e260; doi: 10.1038/cddis.2011.139. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 54. Abelson PH (1990) Medicine from plants. Science 247, 513. [DOI] [PubMed] [Google Scholar]
- 55. Fujita E, Nagao Y, Node M, Kaneko K, Nakazawa S, Kuroda H (1976) Antitumor activity of the Isodon diterpenoids: structural requirements for the activity. Experientia 32, 203–206. [DOI] [PubMed] [Google Scholar]
- 56. Li CY, Wang EQ, Cheng Y, Bao JK (2011) Oridonin: an active diterpenoid targeting cell cycle arrest, apoptotic and autophagic pathways for cancer therapeutics. Int. J. Biochem. Cell Biol. 43, 701–704. [DOI] [PubMed] [Google Scholar]
- 57. Cui Q, Tashiro S, Onodera S, Ikejima T (2006) Augmentation of oridonin‐induced apoptosis observed with reduced autophagy. J. Pharmacol. Sci. 101, 230–239. [DOI] [PubMed] [Google Scholar]
- 58. Hu HZ, Yang YB, Xu XD, Shen HW, Shu YM, Ren Z et al (2007) Oridonin induces apoptosis via PI3K/Akt pathway in cervical carcinoma HeLa cell line. Acta Pharmacol. Sin. 28, 1819–1826. [DOI] [PubMed] [Google Scholar]
- 59. Cheng Y, Qiu F, Ikejima T (2009) Molecular mechanisms of oridonin‐induced apoptosis and autophagy in murine fibrosarcoma L929 cells. Autophagy 5, 430–431. [DOI] [PubMed] [Google Scholar]
- 60. Cheng Y, Qiu F, Ye YC, Guo ZM, Tashiro S, Onodera S et al (2009) Autophagy inhibits reactive oxygen species‐mediated apoptosis via activating p38‐nuclear factor‐kappa B survival pathways in oridonin‐treated murine fibrosarcoma L929 cells. FEBS J. 276, 1291–1306. [DOI] [PubMed] [Google Scholar]
- 61. Indo HP, Sriburee S, Tomita K, Matsui H, Yen HC, Ozawa T et al (2010) Quercetin induced autophagy in rat gastric mucosal cells RGM1 and its tumor cells RGK1. Free Radical Biol. Med. 49, S185–S186. [Google Scholar]
- 62. Wang K, Liu R, Li JY, Mao JL, Lei YL, Wu JH et al (2011) Quercetin induces protective autophagy in gastric cancer cells: involvement of Akt‐mTOR‐ and hypoxia‐induced factor 1α‐mediated signaling. Autophagy 7, 966–978. [DOI] [PubMed] [Google Scholar]
- 63. Psahoulia FH, Moumtzi S, Roberts ML, Sasazuki T, Shirasawa S, Pintzas A (2007) Quercetin mediates preferential degradation of oncogenic Ras and causes autophagy in Ha‐RAS‐transformed human colon cells. Carcinogenesis 28, 1021–1031. [DOI] [PubMed] [Google Scholar]
- 64. Maria R, Carmela S, Idolo T, Stefania B, Gian LR (2012) The flavonoid quercetin in disease prevention and therapy: facts and fancies. Biochem. Pharm. 83, 6–15. [DOI] [PubMed] [Google Scholar]
- 65. Fu LL, Zhou CC, Yao S, Yu JY, Liu B, Bao JK (2011) Plant lectins: targeting programmed cell death pathways as antitumor agents. Int. J. Biochem. Cell Biol. 43, 1442–1449. [DOI] [PubMed] [Google Scholar]
- 66. Liu B, Cheng Y, Bian HJ, Bao JK (2009) Molecular mechanisms of Polygonatum cyrtonema lectin‐induced apoptosis and autophagy in cancer cells. Autophagy 5, 253–255. [DOI] [PubMed] [Google Scholar]
- 67. Liu B, Cheng Y, Zhang B, Bian HJ, Bao JK (2009) Polygonatum cyrtonema lectin induces apoptosis and autophagy in human melanoma A375 cells through a mitochondria‐mediated ROS‐p38‐p53 pathway. Cancer Lett. 275, 54–60. [DOI] [PubMed] [Google Scholar]
- 68. Wang SY, Yu QJ, Bao JK, Liu B (2011) Polygonatum cyrtonema lectin, a potential antineoplastic drug targeting programmed cell death pathways. Biochem. Biophys. Res. Commun. 406, 497–500. [DOI] [PubMed] [Google Scholar]
- 69. Liu B, Bian HJ, Bao JK (2010) Plant lectins: potential antineoplastic drugs from bench to clinic. Cancer Lett. 287, 1–12. [DOI] [PubMed] [Google Scholar]
- 70. Johns CO, Jones DB (1917) The Proteins of the Peanut, Arachis Hypogaea. Proc. Natl. Acad. Sci. USA 3, 365–369. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 71. Loris R, Hamelryck T, Bouckaert J, Wyns L (1998) Legume lectin structure. Biochim. Biophys. Acta 1383, 9–36. [DOI] [PubMed] [Google Scholar]
- 72. Sinha S, Gupta G, Vijayan M, Surolia A (2007) Subunit assembly of plant lectins. Curr. Opin. Struct. Biol. 17, 498–505. [DOI] [PubMed] [Google Scholar]
- 73. Lei HY, Chang CP (2009) Lectin of Concanavalin A as an anti‐hepatoma therapeutic agent. J. Biomed. Sci. 16, 10. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 74. Xu Z, Zhou X, Lu H, Wu N, Zhao H, Zhang L et al (2007) Comparative glycoproteomics based on lectins affinity capture of N‐linked glycoproteins from human Chang liver cells and MHCC97‐H cells. Proteomics 7, 2358–2370. [DOI] [PubMed] [Google Scholar]
- 75. Chang CP, Yang MC, Liu HS, Lin YS, Lei HY (2007) Concanavalin A induces autophagy in hepatoma cells and has a therapeutic effect in a murine in situ hepatoma model. Hepatology 45, 286–296. [DOI] [PubMed] [Google Scholar]
- 76. Li WW, Yu JY, Xu HL, Bao JK (2011) Concanavalin A: a potential anti‐neoplastic agent targeting apoptosis, autophagy and anti‐angiogenesis for cancer therapeutics. Biochem. Biophys. Res. Commun. 414, 282–286. [DOI] [PubMed] [Google Scholar]
- 77. Dalby KN, Tekedereli I, Lopez‐Berestein G, Ozpolat B (2010) Targeting the prodeath and prosurvival functions of autophagy as novel therapeutic strategies in cancer. Autophagy 6, 322–329. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 78. Madeo F, Eisenberg T, Buttner S, Ruckenstuhl C, Kroemer G (2010) Spermidine: a novel autophagy inducer and longevity elixir. Autophagy 6, 160–162. [DOI] [PubMed] [Google Scholar]
- 79. Gossner G, Choi M, Tan L, Fogoros S, Griffith KA, Kuenker M et al (2007) Genistein‐induced apoptosis and autophagocytosis in ovarian cancer cells. Gynecol. Oncol. 105, 23–30. [DOI] [PubMed] [Google Scholar]
- 80. Ying G, Latika K, Barbara JK, Kevin AR (2010) Chloroquine‐induced autophagic vacuole accumulation and cell death in glioma cells is p53 independent. Neuro Oncol. 12, 473–481. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 81. Solomon VR, Lee HY (2012) Chloroquine and its analogs: a new promise of an old drug for effective and safe cancer therapies. Eur. J. Pharmacol. 625, 220–233. [DOI] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.
Supplementary Materials
Table. S1 Representative plant natural compounds targeting autophagy as anti‐cancer agents