

Abstract
Background
Ionizing Radiation (IR) is a known pro-inflammatory agent and in the process of development of biomarkers for radiation biodosimetry, a chronic inflammatory disease condition could act as a confounding factor. Hence, it is important to develop radiation signatures that can distinguish between IR-induced inflammatory responses and pre-existing disease. In this study, we compared the gene expression response of a genetically modified mouse model of inflammatory bowel disease (Il10−/−) with that of a normal wild-type mouse to potentially develop transcriptomics-based biodosimetry markers that can predict radiation exposure in individuals regardless of pre-existing inflammatory condition.
Results
Wild-type (WT) and Il10−/− mice were exposed to whole body irradiation of 7 Gy X-rays. Gene expression responses were studied using high throughput whole genome microarrays in peripheral blood 24 h post-irradiation. Analysis resulted in identification of 1962 and 1844 genes differentially expressed (p < 0.001, FDR < 10%) after radiation exposure in Il10−/− and WT mice respectively. A set of 155 genes was also identified as differentially expressed between WT and Il10−/− mice at the baseline pre-irradiation level. Gene ontology analysis revealed that the 155 baseline differentially expressed genes were mainly involved in inflammatory response, glutathione metabolism and collagen deposition. Analysis of radiation responsive genes revealed that innate immune response and p53 signaling processes were strongly associated with up-regulated genes, whereas B-cell development process was found to be significant amongst downregulated genes in the two genotypes. However, specific immune response pathways like MHC based antigen presentation, interferon signaling and hepatic fibrosis were associated with radiation responsive genes in Il10−/− mice but not WT mice. Further analysis using the IPA prediction tool revealed significant differences in the predicted activation status of T-cell mediated signaling as well as regulators of inflammation between WT and Il10−/− after irradiation.
Conclusions
Using a mouse model we established that an inflammatory disease condition could affect the expression of many radiation responsive genes. Nevertheless, we identified a panel of genes that, regardless of disease condition, could predict radiation exposure. Our results highlight the need for consideration of pre-existing conditions in the population in the process of development of reliable biodosimetry markers.
Electronic supplementary material
The online version of this article (10.1186/s12864-019-5689-y) contains supplementary material, which is available to authorized users.
Keywords: Inflammatory bowel disease (IBD), Biodosimetry, Gene expression, Inflammation, Mouse model
Background
Chronic inflammation is a part of many human diseases [1]. Studies reveal that approximately 5–7% of the western population suffers from some sort of chronic inflammatory condition [1, 2]. Ionizing radiation is a known pro-inflammatory agent [3]. Thus, pre-existing inflammatory disease could potentially confound radiation biodosimetry approaches used for medical triage during a radiological event.
The inflammatory aspect of radiation has been studied extensively in recent years [4–7]. Radiation induced inflammation has previously been reported in the atomic bomb survivors [8]. Lately, there has been renewed interest in the role of inflammation in radiotherapy [5, 9]. On one hand, radiation-mediated immunotoxicity is one of the limiting factors for radiotherapy while on the other hand, radiation facilitated increased immunogenicity is being used to enhance tumor treatment [9–11]. Various factors govern the degree of inflammation after radiation exposure. Genetic background of an individual and preexisting disease conditions can greatly affect the outcome of a radiation exposure [12–14].
Any accidental or intentional large-scale radiological event will result in a large number of individuals requiring medical attention, which will require an effective biodosimetry tool for efficient distribution of medical resources [15, 16]. An individual with an underlying inflammatory condition could show heightened immune responses to radiation exposure and thus obscure some biodosimetry methods. Thus, it is necessary to consider the immune modulating effects of radiation and the potential impact of underlying inflammatory conditions in the development of effective biodosimetry tools for triage purposes.
Various biodosimetric methods have been proposed and evaluated for large-scale radiation exposure scenarios [16]. Gene expression represents an emerging approach to biodosimetry, and could provide estimates of absorbed dose and severity of radiation–induced injury [17–19]. It has been shown that genes involved in inflammation and immune response are among the strongest early responders following ionizing radiation exposure [18, 19]. In addition, radiation exposure triggers a long-lived inflammatory response, with up regulation of inflammatory markers and immune dysfunction that can persist for decades [3, 8].
In this study, we used Il10−/−, a mouse model of human inflammatory bowel disease (IBD), and compared its transcriptomic responses to radiation with those of normal wild-type (WT) mice. Interleukin (IL) 10 is a potent anti-inflammatory cytokine and plays a significant role in regulating the expression of Th1 cytokines, MHC mediated antigen presentation and antibody production from B-cells [20]. After radiation exposure, this cytokine plays an important role in regulating inflammation as well as free radical mediated damage in the neighboring non-exposed cells and tissues [21, 22]. In the absence of IL-10 cytokine, the Il10−/− mice become highly susceptible to enteric bacterial pathogens, and show a heightened inflammatory response to pathogen infection [23, 24]. Similar to IBD patients, the Il10−/− mice have been shown to have higher levels of IL23-producing macrophages, which through their interaction with circulating T-cells cause production of inflammatory cytokines [25]. In terms of radiation responses, Il10−/− mice have been shown to readily develop colitis following a smaller dose of radiation compared to normal WT- mice [Unpublished data]. Thus, we have used this genotype to represent those individuals in the population who have an underlying chronic inflammatory condition that could potentially skew the results from gene expression based biodosimetry during mass triage after a radiation emergency. This study was undertaken as an initial investigation into the potential impact of chronic inflammation on gene expression based radiation biodosimetry markers.
Methods
Animals and irradiation
Wild-type (WT) C57BL/6 and Il10−/− mice (B6.129P2-Il10tm1Cgn/J) were obtained from Jackson Laboratory and bred at Georgetown University as het x het. The study was conducted according to the Georgetown University Institutional Animal Care and Use Committee (IACUC). All the mice (including the WT) were housed in the same room with similar bedding, food, and water under specific pathogen- free (SPF) housing conditions. In addition, the experiment was conducted in the same time for all mice, minimizing the potential of microbiome alterations in the facility. Furthermore, no other mice besides the ones in this study were housed in that room during the time of the experiment. Male littermates between 8 and 10 weeks of age were whole body irradiated with 7 Gy X-rays using an X-Rad 320 X-ray machine (Precision X-ray Inc., Brandford, CT). Sham-irradiated control mice (0 Gy) were treated the same as the exposed animals except the source was not switched on. The dose was selected based on the LD50/30 value of WT-C57BL/6 mice (background strain for Il10−/−mice), of 8Gy for X-ray irradiation [26]. It was reduced slightly (7 Gy) as the Il10−/− mice appeared in early experiments to be more sensitive to radiation. Mice were euthanized 24 h after irradiation using carbon dioxide inhalation followed by cervical dislocation, per IACUC standard procedures. Approximately ~ 0.4 ml blood was collected from each mouse using cardiac puncture method, into a 15 ml centrifuge tube containing 1.6 ml PAXgene Blood RNA stabilization and lysis solution (PreAnalytix GmBH, catalog # 762165), and mixed thoroughly. The tubes were then frozen at − 80 °C, and shipped to Columbia University for RNA isolation. At the time of sacrifice, a drop of blood was also collected in EDTA Microtainer® tubes (BD medical, catalog #365974) for complete blood counts (CBC) using a Genesis hematology system (Oxford Science). Blood samples from 8 animals were used for CBC analysis.
RNA isolation
Blood samples in PAXgene solution were allowed to reach room temperature for 2 h before proceeding to RNA isolation. RNA was purified following the PAXgene RNA kit recommendations with on-column DNase I treatment. Globin RNA was reduced using the Ambion GLOBINclear-mouse/rat kit (Thermo Fisher Scientific). RNA yields were quantified using the NanoDrop ND1000 spectrophotometer (Thermo Fisher Scientific) and RNA quality was checked by the 2100 Bioanalyzer (Agilent). High quality RNA with an RNA integrity number of at least 7.0 was used for microarray hybridization.
Microarray hybridization
For both genotypes, RNA isolated from control and irradiated mouse blood samples (n = 5) was labeled with Cyanine-3 (Cy3) using the One-Color Low-Input Quick Amp Labeling Kit (Agilent Technologies, Santa Clara, CA) following manufacturer’s instructions. Study size (n = 5) was selected to provide 90% power to detect two fold changes, based on an analysis of our previous mouse microarray data using the methods of Lee and Whitmore [27]. A total of 100 ng of input RNA was used for each labeling process. The labeled RNA was purified using the RNAeasy mini kit (Qiagen). Purified RNA having a Cy3 specific activity more than 8 was used for microarray hybridization. A total of 1.65 μg of labeled RNA for each sample was fragmented and hybridized to mouse GE4x44 v2 microarrays (part no. G4846A; Agilent Technologies). The hybridization was carried out at 65 °C for 17 h in a hybridization oven followed by washing as recommended by the supplier (Agilent). Slides were scanned with the Agilent DNA microarray scanner (G2505B) and the images were analyzed with Feature Extraction software [v10.7] (Agilent) using default parameters for background correction and flagging non-uniform features.
Microarray data analysis
Background corrected hybridization intensities were imported into BRB-ArrayTools, v. 4.3.2 (NCI, Biometric Research Branch, Bethesda, MD) [28] log2-transformed and median normalized. Non-uniform outliers or features not significantly above background intensity in 25% or more of the samples, as well as features not changing at least 1.5-fold in 20% or more of the samples were filtered out, giving 16,000 features that were used in subsequent analyses. The microarray data is available through the NCBI Gene Expression Omnibus (series no. GSE114142; http://www.ncbi.nlm.nih.gov/geo/query/acc.cgi?acc=GSE114142).
BRB-Array Tools was used to identify genes that were differentially expressed after radiation exposure in WT and Il10−/− mice. Genes with P < 0.001 and having a false-discovery rate of less than 10% as calculated using the Benjamini-Hochberg method were considered significantly differentially expressed.
Class prediction
We used class prediction methods in BRB-Array Tools to select genes and build predictors of radiation exposure status. The greedy pairs method [29] for feature selection was used to identify the 12 top-performing pairs of genes for discrimination between irradiated and control samples using the wild-type data as the training set. Seven classification methods (compound covariate predictor, linear discriminant analysis, 1- and 3-nearest neighbors, nearest centroid, support vector machines, and Bayesian compound covariate predictor) were used with the selected feature sets to predict the irradiation status of the remaining test samples (Il10−/−). The percentage correct classification was calculated for each approach. The same approach was then repeated using the Il10−/− data as the training set and the wild-type data as the test set.
Gene ontology and network analysis
The lists of differentially expressed genes were analyzed using the functional annotation tool of the Database for Annotation Visualization and Integrated Discovery (DAVID; v 6.7) [30]. Gene ontology terms and biological functions with a Benjamini-corrected P value < 0.05 were considered significantly over-represented within a gene list. Genes that were significantly differentially expressed in the two mouse strains were also imported into Ingenuity Pathway Analysis (IPA from QIAGEN Inc., www.qiagen.com/ingenuity) software and analyzed using IPA Core analysis and Comparative Analysis Tools. IPA uses curated information on the published relationships between gene products to predict networks and association between genes in a list. The upstream regulator analysis specifically uses information about the relationship between the activity of potential upstream regulatory factors and the expression changes of the measured genes to make predictions on the regulatory status of the upstream molecule. IPA generates a z-score for each factor in the upstream regulator analysis and for prediction of activation or inhibition state of biological functions. The IPA default cutoff of z ≥ 2 was used to predict activation and z ≤ − 2 to predict inhibition.
Quantitative real-time RT-PCR
Real-time quantitative RT-PCR (qRT-PCR) was performed for selected genes using Taqman chemistry and the ABI 7900 Real Time PCR System. The Globin cleared purified RNA from 5 control and irradiated animals was used for cDNA synthesis using the High-Capacity cDNA Archive Kit (Life Technologies). Gene expression assays (primer/probe sets) were purchased from Thermo Fisher for the following genes: Ifit3 (Mm00366278_m1), Ifit1 (Mm00365614_m1), Cdkn1a (Mm04205640_g1), Aen (Mm00471554_m1), Gstt1 (Mm00492506_m1), Il18r1 (Mm00515178_m1), Ccng1 (Mm00438084_m1), Alas1 (Mm001235914_m1), Acsbg1 (Mm00547366_m1), Cyp2b6 (Mn00456591_m1), Actb (Mm00607939_s1). The ΔΔCT method was used to calculate expression relative to controls, using normalization to Actb expression. Each reaction was run in triplicate with 5 control and 5 irradiated samples, and means were compared using an unpaired t-test.
Results
Effect of radiation on blood cell counts of WT and Il10−/− mice
In the present study, we compared gene expression in the peripheral blood of WT and Il10−/− mice with and without exposure to ionizing radiation. As different blood cell types have different susceptibility to radiation, we first analyzed the effect of radiation on blood cell counts in the two genotypes 24 h post irradiation.
The total number of white blood cells (WBC) and relative percentages of major blood cell population (i.e. lymphocytes, neutrophils and monocytes) is shown in Fig. 1. Overall, we found a decrease in WBC counts after radiation exposure, which was significant in WT, but not in Il10−/−, mice (Fig. 1a). Although post-irradiation WBC numbers were similar in both genotypes, the Il10−/− controls showed a higher degree of variability. Analysis of different blood cellular components revealed significant decreases in the lymphocyte percentage in the blood of both WT and Il10−/− mice after radiation exposure (Fig. 1b). There was an increase in neutrophil percentage 24 h after radiation exposure, which was statistically significant in WT mice (P < 0.01) but not in Il10−/− mice (Fig. 1c). There was no significant difference in the blood monocyte percentages after radiation exposure in either genotype (Fig. 1d). Interestingly, we observed significant differences in the baseline percentage of neutrophils between WT and Il10−/− mice pre-irradiation (Fig. 1c). In a separate experiment, we found that at a lower dose (2Gy) of radiation exposure, most of the decrease in the WBC counts had been recovered by Day 30 post irradiation in both genotypes (Additional file 1). We did not find any significant differences in red blood cell (RBC) and platelet counts after radiation exposure in WT or Il10−/− mice.
Fig. 1.

Total white blood cell count and percentages of different blood cell types in WT and Il10−/− mice after 24 h of radiation exposure (7Gy). a Total WBC count b Lymphocyte (%) c Neutrophils (%) d Monocytes (%). Significant differences *P < 0.05; **P < 0.01; ***P < 0.001 (n = 8). Green circles (control) and red squares (irradiated) indicate values for individual animals of the respective genotype
Baseline differences in gene expression between WT and Il10−/− mice
In order to detect baseline differences in gene expression between the two genotypes we performed class comparison analysis using BRB-ArrayTools [28]. The analysis identified 155 genes in Il10−/− mice that were differentially expressed (P < 0.001; FDR < 10%) compared to the WT strain without radiation exposure (Table 1; Additional file 2A). Of these 155 differentially expressed genes, 120 (77%) had a significantly higher expression level in Il10−/− mice, while 35 genes (23%) had a lower expression level compared to WT (Table 1) (Fig. 2a). Gene ontology analysis using DAVID [30] (Additional file 2B) showed that these genes were mainly involved in inflammatory response (Benjamini corrected P = 7.19 × 10− 7), glutathione metabolism (Benjamini corrected P = 1.38 × 10− 5) and collagen deposition (Benjamini corrected P = 2.16 × 10− 4).
Table 1.
Differentially expressed genes in blood from WT and Il10−/− mice 24 h post-irradiation (7Gy)
| Class comparison | Number of differentially expressed genes# | Up-regulated genes | Down-regulated genes |
|---|---|---|---|
| Il10−/− vs. WT* | 155 | 120 | 35 |
| Wild-type (WT) irradiated vs. control | 1844 | 1093 | 751 |
| Il10−/− irradiated vs control | 1962 | 994 | 968 |
#Statistical cut off: P-value< 0.001; FDR < 10%
*Genes differentially expressed in Il10−/− mice compared to WT at baseline (0 Gy) without irradiation
Fig. 2.

Analysis of 155 baseline differentially expressed genes between WT and Il10−/− mice. a Heat map of expression of 155 genes. Annotation of genes is provided in Additional file 2A. b IPA based network analysis of significant (P < 0.05) canonical pathways (Additional file 2C) associated with the 155 genes c LPS/IL-1 Pathway. Genes identified as differentially expressed in Il10−/− mice are highlighted in pink. The numbers below indicate fold change and p-value. LBP = LPS binding protein; Cyp2b6 = cytochrome P450 2B6; GST = Glutathione S-transferase; ALAS1 = Aminolevulinate synthase 1; CPT = Carnitine Palmitoyltransferase; ACS = Acetyl Coenzyme A synthetase
Analysis using the IPA network analysis tool [QIAGEN Inc., www.qiagen.com/ingenuity] revealed that LPS (lipopolysaccharide)/IL-1 signaling was the most significantly enriched (P = 7.23 × 10− 8) canonical pathway in this 155 gene set (Fig. 2b; Additional file 2C). This represents a broad inflammatory response, as well as a central pathway linking other responses, including inflammation and glutathione metabolism, that were most affected in the gene ontology analysis of these genes (Fig. 2c). Some of these baseline genes were validated using real-time PCR and are represented in Additional file 3.
Comparative analysis of gene expression response to radiation exposure between WT and Il10−/− mice
We next performed a comparative analysis of radiation responsive genes in WT and Il10−/− mice. We identified a total of 2075 and 2158 differentially expressed features (P < 0.001, FDR < 10%) representing 1844 and 1962 known genes in WT and Il10−/− mice respectively, after radiation exposure (Table 1; Additional file 4). In WT mice, 60% (1093 genes) of the differentially expressed genes were upregulated, and 40% (751 genes) were downregulated after radiation exposure. In Il10−/− mice the differentially expressed genes were evenly distributed with 51% (994) up-regulated genes and 49% (968) downregulated genes (Table 1). Venn diagram analysis revealed 1004 (35%) radiation responsive genes common between WT and Il10−/− mice out of which 527 were upregulated and 477 were downregulated (Fig. 3).
Fig. 3.

Differentially expressed genes a) Venn diagram showing overlap of the total number of differentially expressed genes (P < 0.001, FDR < 10%; Table 1) in WT and Il10−/− mouse strains after 7Gy radiation exposure. There were 35% genes common between WT and Il10−/− mice which showed significant expression change after radiation exposure. b) Represents overlap of the number of up-regulated genes and c) Represents overlap of the number of down-regulated genes after radiation exposure in the two genotypes
Gene Ontology (GO) analysis with DAVID also revealed a level of similarity in terms of significant enrichment of radiation responsive gene ontology terms in WT and Il10−/− mice (Fig. 4). We found innate immune response and p53 signaling were significantly enriched among radiation responsive genes in both WT and Il10−/− mice (Fig. 4a). However, additional immune-related GO terms, such as MHC-I antigen presentation, interferon signaling, and hepatic fibrosis, were also enriched among up-regulated genes of Il10−/− mice but not in WT after radiation exposure. Also, down-regulated genes in WT, but not Il10−/−, mice were significantly enriched with MHC-II protein complex terms, while DNA damage response and DNA repair terms were significantly enriched among down-regulated genes of Il10−/− mice after radiation exposure (Fig. 4b).
Fig. 4.

Gene Ontology (GO) Analysis. The DAVID resource tool was used to find overrepresented GO terms. GO terms with Benjamini-corrected P < 0.05 were considered statistically significant. The color of a cell represents a P value as indicated in the key. a Significant GO categories overrepresented in up-regulated genes in WT and Il10−/− mice b Significant GO categories overrepresented among down-regulated genes in WT and Il10−/− mice in response to radiation
We then used the IPA comparison analysis tool to assess and compare categories of biological processes activated or inhibited in the two genotypes after radiation exposure, as predicted from gene expression changes. We used default z-score ≥ 2 for activation or z-score ≤ − 2 for inhibition as the significance cutoff for our analyses (Fig. 5). We found canonical pathways involved in immune response to infection and B-cell signaling showing similar trends of activity in the two genotypes after radiation exposure. However, the predicted activation status of T-cell signaling differed between the two genotypes after radiation exposure. The T-cell signaling pathway was predicted to be activated in Il10−/− mice but inhibited in WT mice in response to radiation (Additional file 5). In addition, the Th-1 response, which is associated with inflammation, appeared to be activated in Il10−/− mice but inhibited in WT mice. Hormone based signaling pathways also seemed to have different activity status after radiation exposure in the two genotypes (Fig. 5a).
Fig. 5.

Comparative analysis of a) Canonical pathways and b) Upstream regulators responding to radiation in WT and Il10−/− mice. IPA based statistical z-score was used to predict activation (z ≥ 2, orange) or suppression (z ≤ − 2, blue) of a particular pathway or regulator. Grey color indicates no significant activity
We also performed a similar predictive analysis for upstream regulators of the gene expression response to radiation in the two strains and found specific regulators of radiation response and immune function showing similar activity trends in the two genotypes. Significant activation of TP53, LPS, STAT1, IRF3 and IKBKB was indicated in the gene expression response of both mouse types. However, BNIP3L, IL27 and BCL2L1 showed opposite trends of activity in the two genotypes (Fig. 5b). We also identified a set of pathways and regulators that appeared significant in one genotype but not the other. These may suggest specific differences in the radiation response of WT and Il10−/− mice.
Impact of inflammatory response on prediction of radiation exposure
Since one of our broad objectives is the development of gene expression signatures predictive of radiation exposure, we used microarray data to build and test classifiers using a greedy pairs method with 7 different algorithms. To test the ability of a gene set selected from WT data to identify radiation exposure in the presence of chronic inflammation, a set of 24 genes was identified using a greedy pairs algorithm and the WT samples as a training data set (Table 2). The data from the Il10−/− mice were then used as a test set, and all 7 algorithms predicted the exposure status correctly with 100% accuracy (P-value 0.05). Many of the genes selected in the classifier were well-established radiation responsive genes with roles in the Trp53 pathway or in inflammation and immune response. In fact, “Immunity” was the only GO category significantly overrepresented (Benjamini corrected p-value = 0.025 by DAVID analysis) among the selected signature genes. As genes with immune functions formed such a significant part of the signature, we also used the Il10−/− data to train a classifier (Table 3) and tested it on the WT data to see what impact this would have. The new classifier included a higher proportion of immunity related genes (“Immunity” overrepresented with a Benjamini corrected p-value = 0.0019 by DAVID analysis), and performed with 100% accuracy on the Il10−/− samples during cross validation. Despite the increased contribution from immunity related genes, all 7 algorithms also predicted the exposure status of the WT test samples correctly with 100% accuracy.
Table 2.
Composition of 24-gene classifiers distinguishing radiation exposure. Features were selected using the greedy pairs method using WT mouse data as the training set
| UniqueID | Symbol | Name | Parametric p-value | Geom mean of intensities in Irradiated | Geom mean of intensities in Controls | Fold-change (Irradiated/controls) | |
|---|---|---|---|---|---|---|---|
| 1 | A_51_P391955 | Dapl1 | death associated protein-like 1 | < 1e-07 | 5.74 | 354.08 | 0.016 |
| 2 | A_55_P2136906 | Vpreb3 | pre-B lymphocyte gene 3 | < 1e-07 | 23.78 | 952.67 | 0.025 |
| 3 | A_55_P2082929 | H2-Ob | histocompatibility 2, O region beta locus | < 1e-07 | 25.11 | 544.45 | 0.046 |
| 4 | A_51_P378298 | Fcmr | Fc fragment of IgM receptor | < 1e-07 | 93.84 | 2574.17 | 0.036 |
| 5 | A_55_P2108943 | Ccr6 | chemokine (C-C motif) receptor 6 | < 1e-07 | 9.01 | 390.03 | 0.023 |
| 6 | A_55_P2117614 | Tnfrsf13c | tumor necrosis factor receptor superfamily, member 13c | < 1e-07 | 18.07 | 470.05 | 0.038 |
| 7 | A_55_P1958275 | Bcl11a | B cell CLL/lymphoma 11A (zinc finger protein) | < 1e-07 | 14.18 | 304.79 | 0.047 |
| 8 | A_55_P2332731 | D130062J21Rik | RIKEN cDNA D130062J21 gene | < 1e-07 | 23.24 | 302.25 | 0.077 |
| 9 | A_55_P1964648 | Btla | B and T lymphocyte associated | < 1e-07 | 40.07 | 445.85 | 0.09 |
| 10 | A_66_P130916 | H2-Ob | histocompatibility 2, O region beta locus | < 1e-07 | 54.33 | 623.04 | 0.087 |
| 11 | A_52_P417654 | Tcea1 | transcription elongation factor A (SII) 1 | 1.6e-06 | 54.29 | 229.5 | 0.24 |
| 12 | A_66_P103490 | Srsf1 | serine/arginine-rich splicing factor 1 | 5.8e-06 | 34 | 88.08 | 0.39 |
| 13 | A_51_P292030 | Nus1 | NUS1 dehydrodolichyl diphosphate synthase subunit | 0.0009584 | 197.92 | 116.11 | 1.7 |
| 14 | A_51_P223443 | Cd81 | CD81 antigen | 8.2e-06 | 7009.38 | 2854.95 | 2.46 |
| 15 | A_55_P2145804 | Aen | apoptosis enhancing nuclease | 4.6e-06 | 909.81 | 172.36 | 5.28 |
| 16 | A_55_P1952379 | Fkbp5 | FK506 binding protein 5 | 3.3e-06 | 678.47 | 229.64 | 2.95 |
| 17 | A_55_P1998943 | Oas1a | 2′-5′ oligoadenylate synthetase 1A | 5e-07 | 5226.79 | 408.46 | 12.8 |
| 18 | A_52_P612803 | Ccng1 | cyclin G1 | 3e-07 | 423.04 | 79.12 | 5.35 |
| 19 | A_52_P90363 | Ifi27l2a | interferon, alpha-inducible protein 27 like 2A | 1e-07 | 30,011.04 | 2971.34 | 10.1 |
| 20 | A_51_P154842 | Oas1f | 2′-5′ oligoadenylate synthetase 1F | < 1e-07 | 2053.56 | 233.96 | 8.78 |
| 21 | A_51_P363947 | Cdkn1a | cyclin-dependent kinase inhibitor 1A (P21) | < 1e-07 | 897.24 | 24.09 | 37.24 |
| 22 | A_55_P1972872 | I830012O16Rik | RIKEN cDNA I830012O16 gene | < 1e-07 | 462.01 | 18.35 | 25.17 |
| 23 | A_51_P259975 | Aspa | aspartoacylase | < 1e-07 | 159.62 | 10.1 | 15.8 |
| 24 | A_51_P329928 | Phlda3 | pleckstrin homology like domain, family A, member 3 | < 1e-07 | 2163.11 | 137.61 | 15.72 |
Table 3.
Composition of 24-gene classifiers distinguishing radiation exposure. Features were selected using the greedy pairs method using Il10−/− mouse data as the training set
| UniqueID | Symbol | Name | Parametric p-value | Geom mean of intensities in Irradiated | Geom mean of intensities in Controls | Fold-change (Irradiated/Controls) | |
|---|---|---|---|---|---|---|---|
| 1 | A_55_P2108943 | Ccr6 | chemokine (C-C motif) receptor 6 | < 1e-07 | 11.77 | 554.18 | 0.021 |
| 2 | A_51_P248122 | Bbc3 | BCL2 binding component 3 | < 1e-07 | 1019.01 | 263.9 | 3.86 |
| 3 | A_51_P378298 | Fcmr | Fc fragment of IgM receptor | < 1e-07 | 41.34 | 1536.63 | 0.027 |
| 4 | A_51_P304859 | Zfand6 | zinc finger, AN1-type domain 6 | 4e-07 | 115.94 | 422.84 | 0.27 |
| 5 | A_51_P371750 | Marco | macrophage receptor with collagenous structure | < 1e-07 | 5630.68 | 341.13 | 16.51 |
| 6 | A_52_P612803 | Ccng1 | cyclin G1 | < 1e-07 | 483.89 | 90.38 | 5.35 |
| 7 | A_51_P363947 | Cdkn1a | cyclin-dependent kinase inhibitor 1A (P21) | < 1e-07 | 1154.64 | 47.87 | 24.12 |
| 8 | A_55_P2121856 | Ier5l | immediate early response 5-like | < 1e-07 | 172.96 | 53.17 | 3.25 |
| 9 | A_55_P2137496 | Cd79a | CD79A antigen (immunoglobulin-associated alpha) | < 1e-07 | 138.15 | 1777 | 0.078 |
| 10 | A_51_P205779 | Cd5l | CD5 antigen-like | 6e-07 | 8412.39 | 1257.39 | 6.69 |
| 11 | A_51_P329928 | Phlda3 | pleckstrin homology like domain, family A, member 3 | < 1e-07 | 2635.66 | 186.27 | 14.15 |
| 12 | A_52_P77080 | Gypa | glycophorin A | < 1e-07 | 93.67 | 1314.36 | 0.071 |
| 13 | A_55_P2007964 | Cx3cr1 | chemokine (C-X3-C motif) receptor 1 | < 1e-07 | 46.72 | 374.07 | 0.12 |
| 14 | A_55_P2051254 | Pvt1 | plasmacytoma variant translocation 1 | < 1e-07 | 118.94 | 27.32 | 4.35 |
| 15 | A_51_P327751 | Ifit1 | interferon-induced protein with tetratricopeptide repeats 1 | < 1e-07 | 545.07 | 82.18 | 6.63 |
| 16 | A_51_P226429 | Rhobtb2 | Rho-related BTB domain containing 2 | 1.2e-06 | 51.68 | 128.33 | 0.4 |
| 17 | A_51_P359570 | Ifit3 | interferon-induced protein with tetratricopeptide repeats 3 | < 1e-07 | 2195.72 | 75.33 | 29.15 |
| 18 | A_52_P497625 | A630001G21Rik | RIKEN cDNA A630001G21 gene | 1.4e-06 | 40.17 | 113.97 | 0.35 |
| 19 | A_66_P120987 | Cd79a | CD79A antigen (immunoglobulin-associated alpha) | < 1e-07 | 21.65 | 180.7 | 0.12 |
| 20 | A_55_P2105256 | Trim47 | tripartite motif-containing 47 | < 1e-07 | 373.14 | 76.03 | 4.91 |
| 21 | A_55_P1958275 | Bcl11a | B cell CLL/lymphoma 11A (zinc finger protein) | < 1e-07 | 9.03 | 228.33 | 0.04 |
| 22 | A_52_P180565 | 2310061I04Rik | RIKEN cDNA 2310061I04 gene | 3.3e-06 | 158.13 | 373.06 | 0.42 |
| 23 | A_52_P390944 | Chst3 | carbohydrate (chondroitin 6/keratan) sulfotransferase 3 | < 1e-07 | 8.68 | 274.93 | 0.032 |
| 24 | A_55_P2097474 | Creb5 | cAMP responsive element binding protein 5 | 6.6e-06 | 1535.01 | 3450.09 | 0.44 |
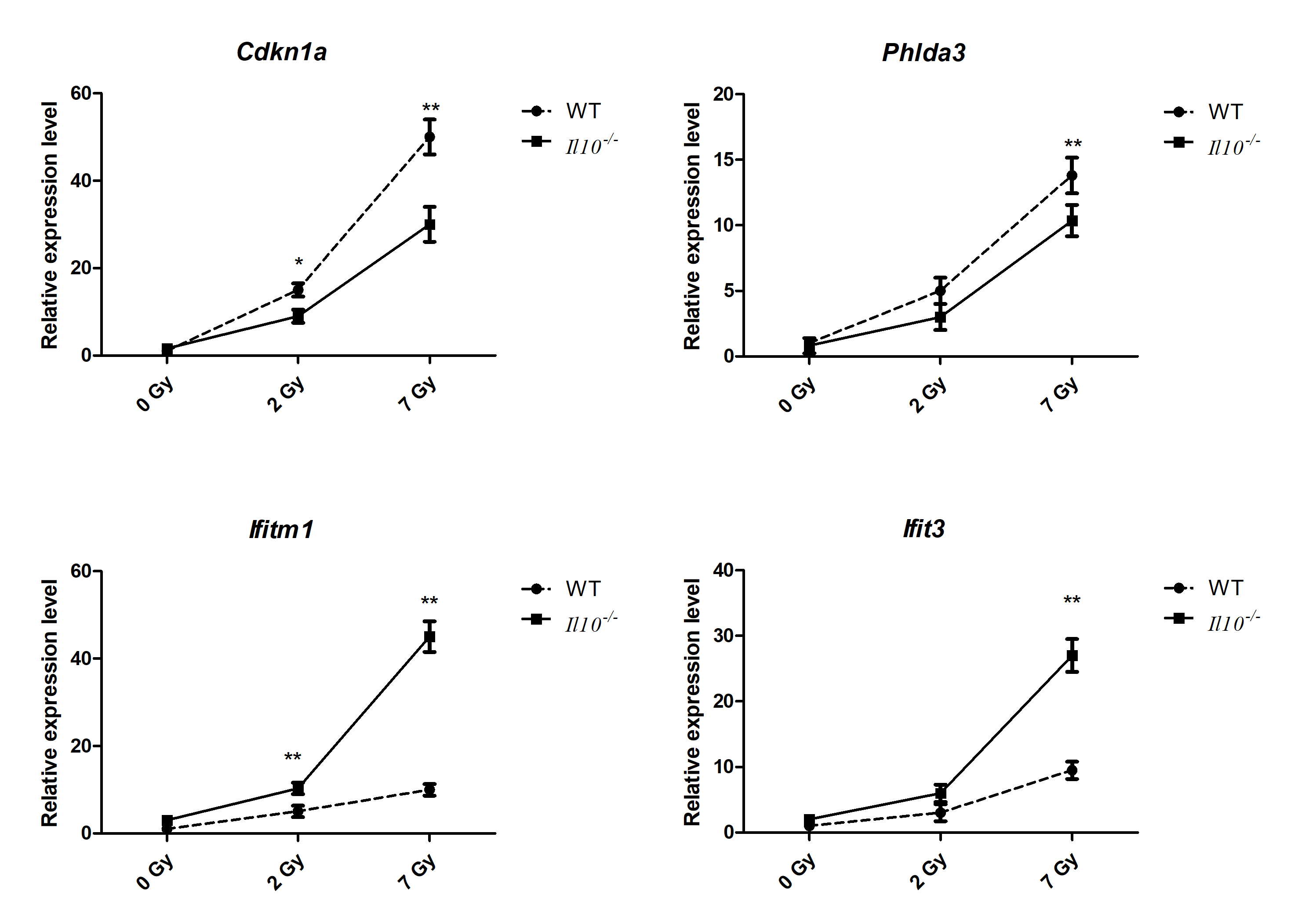
Finally, we validated expression levels of six of the classifier genes using quantitative real-time PCR. We chose four Trp53-responsive genes, Cdkn1a, Aen, Ccng1 and Phlda3 and two inflammatory genes, Ifit1 and Ifit3, for this analysis. As observed in the microarray results, we found a greater relative fold change of Tp53-responsive genes in WT compared to Il10−/− mice after radiation exposure, and a greater relative fold change of inflammatory cytokines in Il10−/− mice (Fig. 6). The inclusion of a 2 Gy dose measurement of Cdkn1a, Phlda3, Ifitm1 and Ifit3 (Additional file 6) suggests that the relative responses of the Trp53 regulated genes in the two strains are similar at low and high doses. In contrast, the heightened response of inflammatory genes in the Il10−/− mice, although still significant for Ifitm1 at 2 Gy, was much greater at the higher dose. A more detailed delineation of the interaction between radiation dose and inflammatory gene expression in these two genotypes would be of interest.
Fig. 6.

Gene expression measured by qRT-PCR. Expression levels of 6 genes (Cdkn1a, Aen, Phlda3, Ccng1, Ifitm1, Ifit3) showing significant difference in expression in Il10−/− mice compared to WT after radiation exposure are presented here. The expression level for each gene was calculated relative to the expression levels in unexposed WT animals considered as 1. The expression was normalized against Beta-Actin gene expression. The data is represented as mean +/− SEM (n = 5). * Significant at P < 0.05, ** Significant at P < 0.01, *** Significant at P < 0.001, using unpaired t-test
Discussion
Exposure to ionizing radiation has long been known to cause production of various immune molecules like cytokines, DAMPs (Damage-associated molecular patterns) and cellular factors in the irradiated tissue, and to result in inflammation [5, 6]. The degree of radiation mediated inflammation varies in individuals and depends on various factors including genetic, life-style and pre-existing disease conditions [14]. Inflammation has been indicated as a potential confounding factor in the development of IR biomarkers in epidemiological studies [31]. Previous transcriptomic studies have shown that immune and inflammation-related genes dominate the gene expression responses seen after in vivo radiation exposure [17, 31] and therefore, inflammation related genes have been proposed as biomarkers of radiation exposure or to be used in biodosimetric panels [32]. This raises the concern that inflammation may affect radiation responses, and may impact the use of some genes in biodosimetric panels for use in triage after an accidental or deliberate radiological event. Several investigators have used treatment with LPS, a general component of the bacterial cell wall that is a potent inducer of inflammatory response [30, 31], as a model for acute inflammation or infection. LPS has been shown to induce expression of some well-established radiation responsive genes and pathways [33–35], furthering concern about the impact of inflammatory states. As an initial approach to examining the potential impact of a pre-existing chronic inflammatory condition on radiation biodosimetry, we have used a well-established mouse model (Il10−/−) of human chronic inflammatory bowel disease (IBD) and compared its gene expression with and without radiation exposure with that of wild type (WT) littermates.
First, we compared blood counts in the two genotypes before and after irradiation. Of note was the significant difference in the baseline level of neutrophils in Il10−/− compared to WT mice. Neutrophils are generally associated with inflammatory response. In addition, a higher level of neutrophils has been reported in IBD patients, which correlates with the severity of their clinical manifestations [36, 37]. The higher level of neutrophils in Il10−/− mice is similar to that in individuals suffering from IBD. The increased proportions of neutrophils contribute to a pro-inflammatory blood immune environment, which creates a concern of possible false positives or interference with radiation response for dosimetric biomarkers.
We next compared the baseline gene expression levels in un-irradiated Il10−/− mice with those in WT mice (Fig. 2). The genes that showed significant differences in expression were mainly involved in inflammatory immune response, as might be expected, as well as glutathione metabolism and collagen deposition. In a recent publication, Mak et al. [38] showed that glutathione metabolism plays an important role in the activation process of inflammatory T-cells in response to immune stimuli. Excessive deposition of collagen, a major constituent of the extracellular matrix, has also been associated with inflammation and the development of fibrosis [39], functions also associated with radiation exposure. We found LPS / IL-1 signaling, a major regulator of gene expression responses in IBD patients [40], to be a significant pathway connecting the processes of inflammation, glutathione metabolism and fibrosis (Fig. 2c). These results suggest that chronic inflammatory conditions can indeed trigger some of the same gene expression patterns indicative of radiation response in the absence of exposure.
Transcriptomic analysis of radiation responses in the two genotypes revealed broad similarity among the gene ontology terms most significantly affected by radiation exposure. Unsurprisingly, these included p53 signaling and some immune related functions (Fig. 4). Although significantly enriched among radiation responsive genes in both genotypes, the canonical radiation response of p53 signaling did appear to be slightly muted in the Il10−/− mice, as implied by a slightly higher P value compared to WT-mice. This suggests that fewer p53-regulated genes responded significantly to radiation in the mutant mice. In confirming the radiation induction of individual genes, we found that both Cdkn1a and Aen were up regulated by radiation exposure to a lesser degree in the Il10−/− mice, compared to WT mice (Fig. 6). There is some evidence in the literature of a regulatory link between IL10 and p53 in inflammatory processes. IL10-induced apoptosis in mouse bone marrow-derived mast cells has been shown to depend on activation of p53 [41]. Levels of IL10 and p53 were also found to be correlated in patients with autoimmune disease, with high levels of IL10 apparently leading to overexpression of p53 [42]. Taken together, these observations suggest that IL10 may contribute to the regulation of p53, leading to a blunted radiation response of genes downstream of p53 in the absence of IL10.
The observed enrichment of genes with functions related to B-cell development among the down-regulated genes of both genotypes could be explained by the observed overall decrease in lymphocyte levels after radiation exposure (Fig. 1), particularly as B cells are among the most radiosensitive hematopoietic cells [43].
We observed that specific immune functions like MHC based antigen presentation and interferon signaling were particularly associated with radiation responsive genes in Il10−/− mice but not WT mice. Antigen presentation mainly occurs through major histocompatibility complex (MHC) receptors on the cell surface, and is a very important process in activation of T-cells, which mount an immune response by releasing cytokines or protein mediators of apoptosis [44]. The MHC-I is present on the cell surface of all the nucleated cells while MHC-II is present only in specific antigen presenting cells (APC), like macrophages and dendritic cells. Antigen presentation through MHC-II receptors results in activation of T-helper (Th) cells, which modulate the immune environment by releasing specific cytokines [44, 45]. We found genes involved in MHC-II based antigen presentation to be specifically downregulated by radiation exposure in WT mice but upregulated in Il10−/− mice. This indicates a differential modulation of the antigen presentation mechanism and its downstream immune response in the two genotypes after radiation exposure.
It has been reported that similar to IBD patients, Il10−/− mice have higher levels IL23 producing macrophages, which through their interaction with T-cells via MHC-II receptors cause production of various inflammatory cytokines [25]. Intriguingly, IPA predicted a significant response to radiation of T-cell signaling in both genotypes, but in opposite directions. In Il10−/− mice the gene expression pattern suggested a significant radiation activation of T-cell response, while in WT mice this pathway was predicted to be inhibited. Similarly, the T-helper type-1 (Th-1) immune response pathway was predicted to follow the same response pattern, with the pathway appearing to be activated by radiation exposure in Il10−/− mice and inhibited in WT mice. Th1 mediated signaling has been reported to be the major causes of inflammation in people suffering from IBD as well as in animal models of IBD [24, 40, 45]. Production of interferons like IFN-gamma is a hallmark of Th1 immune response, and is also required for driving T-helper cell populations towards the Th1 type of immune response [46]. We found significant upregulation of interferon and its associated genes in Il10−/− mice but not in WT after radiation exposure. This type of immune response also plays an important role against viral and bacterial infection. We found a significant association of these specific gene ontology terms specifically with the radiation response in Il10−/− mice (Fig. 4; Additional file 5). Overall, we found that radiation exposure results in higher inflammatory responses in Il10−/− mice compared to WT, and this appears to be mediated by the T-cell population.
Impact on biodosimetry markers
Since our laboratory is interested in the development of biodosimetry markers for radiation exposure, we also used our data to select and test panels of genes that could distinguish radiation exposure in individual samples regardless of pre-existing inflammatory disease. The panels included p53 responsive genes like Cdkn1a and Aen as well as immune responsive genes like Ifit1 and Ifit3. Cdkn1a expression is known to respond widely to a number of stresses, and has been previously shown to respond to the inflammation inducing agent LPS [32]. In our current study, we found a significant difference between WT and Il10−/− expression levels of Cdkn1a and other p53 regulated genes after radiation exposure, with the Cdkn1a radiation response being significantly muted in Il10−/− mice compared to WT. The reverse was true of several immune related genes, which showed a larger magnitude of response to radiation exposure in the Il10−/− mice compared to WT mice. Although in our current study these differences in response did not limit our ability to detect a relatively damaging dose of radiation, these results do suggest that excessive reliance on genes in these pathways may have an impact on the ultimate goal of dose discrimination. Care must also be taken to ensure that the elevated expression of inflammatory genes noted in the unirradiated Il10−/− mice does not lead to misidentification of such inflammation-prone individuals as having received a small dose of radiation. Future studies across a range of doses will be needed to fully understand the impact of preexisting inflammation states on radiation biodosimetry using gene expression.
In a previous study, we had shown that genes selected from WT mice performed poorly in detecting exposure to an LD50/30 radiation dose in two mouse models of DNA repair deficiency (Atm−/− and Prkdcscid), but that inclusion of both WT and DNA repair deficient mice in the training set improved performance on both mutant and WT test sets to 100% [47]. We tested the ability of this earlier signature to predict radiation exposure in Il10−/− mice. Although only one gene, Cdkn1a, was common between the earlier signature and either of the signatures reported in this study, we tested the WT plus repair mutant gene set on the dataset from this study and found that it was again able to predict the radiation exposure in Il10−/− mice, and the new set of WT mice, with 100% efficiency (p < 0.005). This supports inclusion of individuals with a range of radiation sensitivities and underlying disease states in gene selection to produce more robust signatures for radiation biodosimetry.
Conclusions
Using a mouse model of inflammatory bowel disease, we have found that a chronic inflammatory state can impact both the basal expression levels and the relative radiation response of genes involved in immune and inflammatory pathways, as well as other key radiation response pathways, such as p53. While this did not impact the ability of gene expression signatures derived from either WT or pro-inflammatory Il10−/− mice to correctly distinguish between unexposed and irradiated mice, it does raise concerns for the accurate determination of doses across a broad dose range. Future experiments employing multiple radiation doses will enable selection of genes with expression levels unaffected by underlying inflammation, and will help to avoid confounding of radiation biodosimetric gene expression signatures by underlying chronic inflammation.
Additional files
{kind=link}
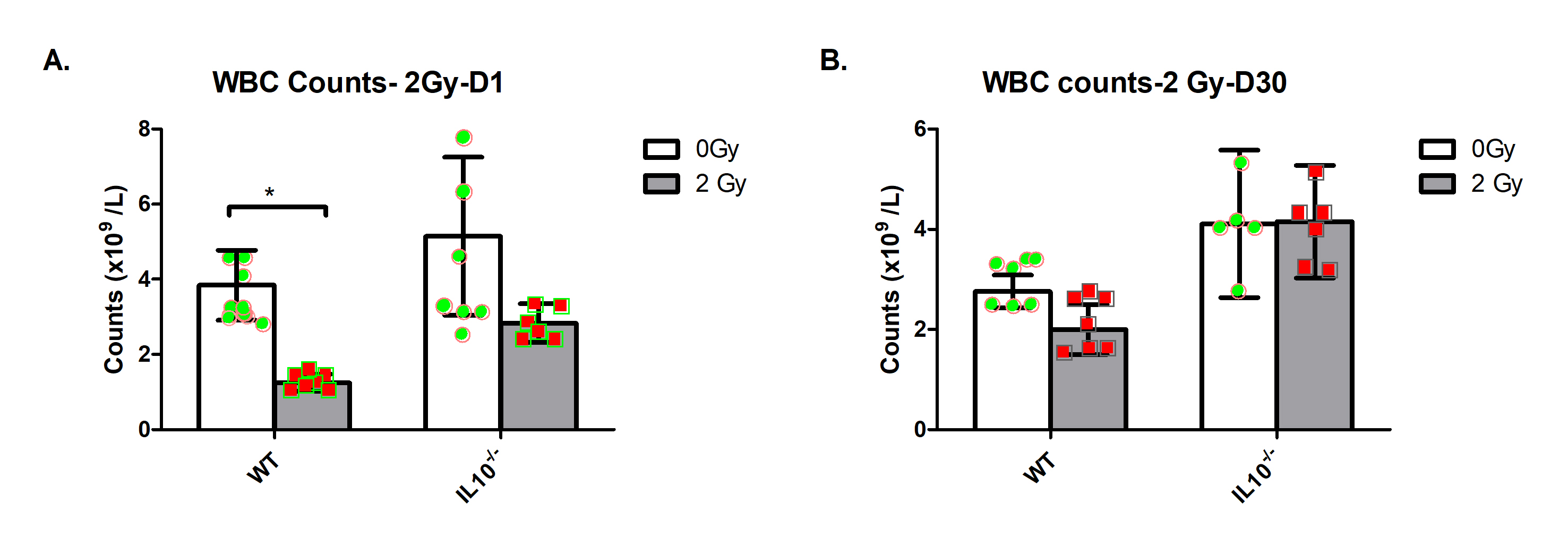
Total white blood cell counts in WT and Il10−/− mice after 2 Gy of radiation exposure A) Counts at Day1 post exposure B) Counts at Day 30 post exposure. Green circles (control) and red squares (irradiated) indicate values for individual animals of the respective genotype. Significant differences *P < 0.05 (n = 8). (JPG 360 kb)
A. Genes showing significant differences in expression levels between WT and Il10−/− mice prior to radiation exposure. B. Gene ontology (GO) analysis genes using DAVID [30]. Top 5 significant (Benjamini corrected P-value< 0.05) GO terms are shown in the bar diagram. C. IPA based network analysis. Top 10 significant (P < 0.05) Canonical pathways are shown in bar diagrams. (XLSX 323 kb)
{kind=link}
Real-time qRT-PCR validation of 5 genes (Gstt1, Il18r1, Alas1, Acsbg1, Cyp2b6) relevant to the current study, showing significant difference in expression at baseline level between WT and Il10−/− mice prior to radiation exposure. The data is represented as mean +/− SEM (n = 5) for each genotype. * Significant at P < 0.05 using unpaired t-test. (JPG 398 kb)
BRB-array tools output of differentially expressed genes after radiation exposure in the two genotypes. Results of each genotype are in a separate tab. (XLSX 708 kb)
{kind=link}
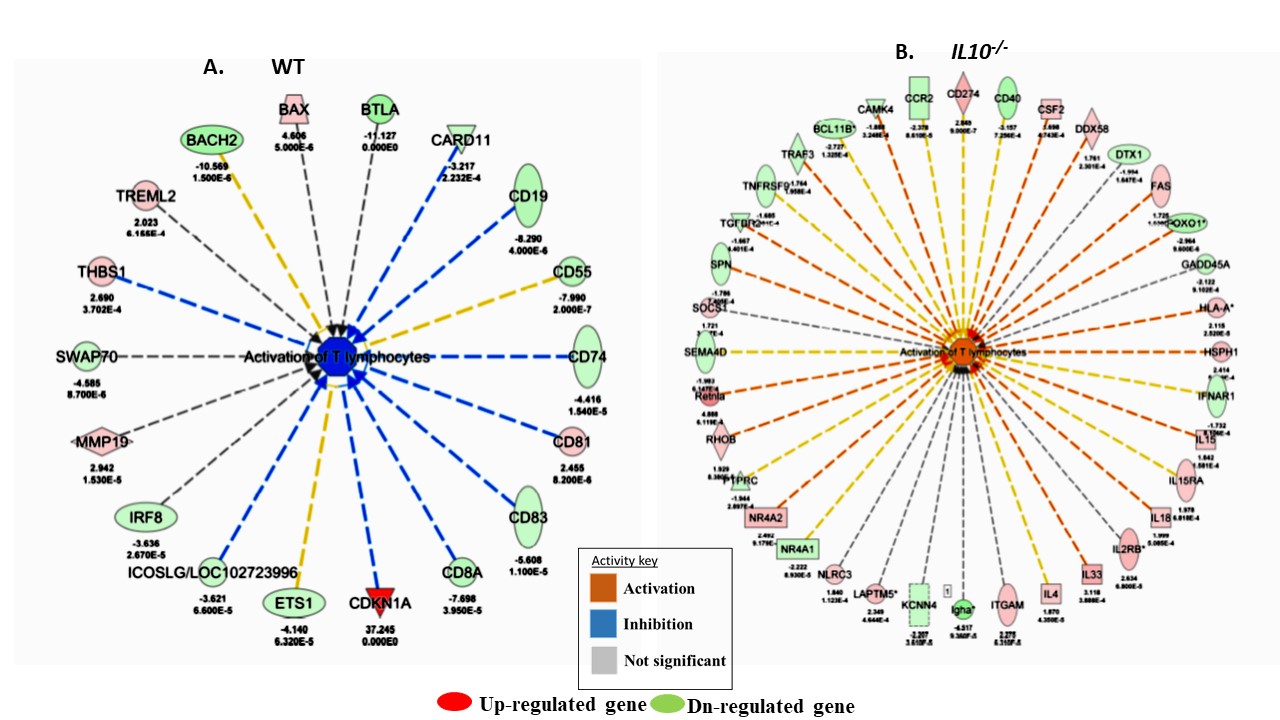
IPA network analysis of genes involved in T-cell signaling in A. WT and B. Il10−/− mice in response to radiation. Statistical z-score (> 2 or < − 2) was used for prediction of the activation status of the signaling pathway. Orange lines indicate activation (z ≥ 2) and blue indicates inhibition (z ≤ − 2), Grey lines indicate no significant contribution to activity. Up-regulated genes are shown in red and down-regulated genes in green. (JPG 165 kb)
{kind=link}
Dose response curves of expression of 4 genes (Cdkn1a, Phlda3, Ifitm1, Ifit3) based on qRT-PCR analysis. Relative gene expression at 0, 2 and 7 Gy of radiation exposure is shown for WT (dotted line) and Il10−/− mice (Solid line). Relative expression was calculated compared to the expression level at 0 Gy (unexposed-sham controls) of WT considered as 1. The expression was normalized against Actin-Beta gene expression. The data is represented as mean +/− SD (n = 5) for each genotype. * Significant at P < 0.05, **Significant at P < 0.01, *** Significant at P < 0.001, using unpaired t-test. (JPG 598 kb)
Acknowledgements
The authors would like to thank Ms. Pelagie Ake, Ms. Linze Song and Mr. Jerry Angdisen for their help in animal breeding and sample collection. Analyses were performed using BRB-ArrayTools developed by Dr. Richard Simon and the BRB-ArrayTools Development Team at the National Cancer Institute.
Funding
This work was supported by the Center for High-Throughput Minimally-Invasive Radiation Biodosimetry, National Institute of Allergy and Infectious Diseases grant number U19AI067773. The funding body did not play any role in the design of the study and collection, analysis, and interpretation of data and in writing the manuscript.
Availability of data and materials
The microarray data generated in this study have been deposited in the National Center for Biotechnology Information (NCBI) Gene Expression Omnibus (GEO) database with accession number GSE114142 (http://www.ncbi.nlm.nih.gov/geo/query/acc.cgi?acc=GSE114142).
Abbreviations
- DAVID
Database for annotation, visualization and integrated discovery
- FDR
False discovery rate
- GO
Gene ontology
- IBD
Inflammatory bowel disease
- IPA
Ingenuity pathway analysis
- qRT-PCR
quantitative Real-Time Polymerase Chain Reaction
Authors’ contributions
SAA, ECL and AJF designed the study. SM and ECL managed the study and performed the experiments. SM did the data analysis, result interpretation and wrote the manuscript. SAA and AJF provided critical feedback on the manuscript. All authors read and approved the final manuscript.
Ethics approval and consent to participate
This study was approved by the Georgetown University Institutional Animal Care and Use Committee (IACUC) (approval number 2016–1152) and conducted in accordance with applicable federal and state guidelines.
Consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing interests.
Publisher’s Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Contributor Information
Sanjay Mukherjee, Email: sm4312@cumc.columbia.edu.
Evagelia C. Laiakis, Email: ecl28@georgetown.edu
Albert J. Fornace, Jr, Email: af294@georgetown.edu.
Sally A. Amundson, Email: saa2108@cumc.columbia.edu
References
- 1.El-Gabalawy H, Guenther LC, Bernstein CN. Epidemiology of immune-mediated inflammatory diseases: incidence, prevalence, natural history, and comorbidities. J Rheumatol Suppl. 2010;85(0):2–10. doi: 10.3899/jrheum.091461. [DOI] [PubMed] [Google Scholar]
- 2.Okin D, Medzhitov R. Evolution of inflammatory diseases. Curr Biol. 2012;22(17):R733–R740. doi: 10.1016/j.cub.2012.07.029. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 3.Mukherjee D, Coates PJ, Lorimore SA, Wright EG. Responses to ionizing radiation mediated by inflammatory mechanisms: tissue responses in radiation pathology. J Pathol. 2014;232(3):289–299. doi: 10.1002/path.4299. [DOI] [PubMed] [Google Scholar]
- 4.Hekim N, Cetin Z, Nikitaki Z, Cort A, Saygili EI. Radiation triggering immune response and inflammation. Cancer Lett. 2015;368(2):156–163. doi: 10.1016/j.canlet.2015.04.016. [DOI] [PubMed] [Google Scholar]
- 5.Schaue D, Micewicz ED, Ratikan JA, Xie MW, Cheng G, McBride WH. Radiation and inflammation. Semin Radiat Oncol. 2015;25(1):4–10. doi: 10.1016/j.semradonc.2014.07.007. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 6.Candéias SM, Testard I. The many interactions between the innate immune system and the response to radiation. Cancer Lett. 2015;368(2):173–178. doi: 10.1016/j.canlet.2015.02.007. [DOI] [PubMed] [Google Scholar]
- 7.Gandhi S, Chandna S. Radiation-induced inflammatory cascade and its reverberating crosstalks as potential cause of post-radiotherapy second malignancies. Cancer Metastasis Rev. 2017;36(2):375–393. doi: 10.1007/s10555-017-9669-x. [DOI] [PubMed] [Google Scholar]
- 8.Neriishi K, Nakashima E, Delongchamp RR. Persistent subclinical inflammation among A-bomb survivors. Int J Radiat Biol. 2001;77(4):475–482. doi: 10.1080/09553000010024911. [DOI] [PubMed] [Google Scholar]
- 9.Demaria S, Coleman CN, Formenti SC. Radiotherapy: changing the game in immunotherapy. Trends Cancer. 2016;2(6):286–294. doi: 10.1016/j.trecan.2016.05.002. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 10.Derer A, Frey B, Fietkau R, Gaipl US. Immune-modulating properties of ionizing radiation: rationale for the treatment of cancer by combination radiotherapy and immune checkpoint inhibitors. Cancer Immunol Immunother CII. 2016;65(7):779–786. doi: 10.1007/s00262-015-1771-8. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 11.Hodge JW, Ardiani A, Farsaci B, Kwilas AR, Gameiro SR. The tipping point for combination therapy: cancer vaccines with radiation, chemotherapy, or targeted small molecule inhibitors. Semin Oncol. 2012;39(3):323–339. doi: 10.1053/j.seminoncol.2012.02.006. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 12.van de Vosse E, van Dissel JT, Ottenhoff TH. Genetic deficiencies of innate immune signalling in human infectious disease. Lancet Infect Dis. 2009;9(11):688–698. doi: 10.1016/S1473-3099(09)70255-5. [DOI] [PubMed] [Google Scholar]
- 13.Jin P, Wang E. Polymorphism in clinical immunology – from HLA typing to immunogenetic profiling. J Transl Med. 2003;1:8. doi: 10.1186/1479-5876-1-8. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 14.Multhoff G, Radons J. Radiation, inflammation, and immune responses in Cancer. Front Oncol. 2012;2:58. doi: 10.3389/fonc.2012.00058. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 15.Coleman CN, Koerner JF. Biodosimetry: medicine, science, and systems to support the medical decision-maker following a large scale nuclear or radiation incident. Radiat Prot Dosim. 2016;172(1–3):38–46. doi: 10.1093/rpd/ncw155. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 16.Sullivan JM, Prasanna PGS, Grace MB, Wathen LK, Wallace RL, Koerner JF, et al. Assessment of biodosimetry methods for a mass-casualty radiological incident: medical response and management considerations. Health Phys. 2013;105(6):540–554. doi: 10.1097/HP.0b013e31829cf221. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 17.Paul S, Barker CA, Turner HC, McLane A, Wolden SL, Amundson SA. Prediction of In Vivo radiation dose status in radiotherapy patients using Ex Vivo and In Vivo gene expression signatures. Radiat Res. 2011;175(3):257–265. doi: 10.1667/RR2420.1. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 18.Amundson SA, Grace MB, McLeland CB, Epperly MW, Yeager A, Zhan Q, et al. Human in vivo radiation-induced biomarkers: gene expression changes in radiotherapy patients. Cancer Res. 2004;64(18):6368–6371. doi: 10.1158/0008-5472.CAN-04-1883. [DOI] [PubMed] [Google Scholar]
- 19.Dressman HK, Muramoto GG, Chao NJ, Meadows S, Marshall D, Ginsburg GS, et al. Gene expression signatures that predict radiation exposure in mice and humans. PLoS Med. 2007;4(4):e106. doi: 10.1371/journal.pmed.0040106. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 20.Kühn R, Löhler J, Rennick D, Rajewsky K, Müller W. Interleukin-10-deficient mice develop chronic enterocolitis. Cell. 1993;75(2):263–274. doi: 10.1016/0092-8674(93)80068-P. [DOI] [PubMed] [Google Scholar]
- 21.Schaue D, Kachikwu EL, McBride WH. Cytokines in radiobiological responses: a review. Radiat Res. 2012;178(6):505–523. doi: 10.1667/RR3031.1. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 22.Di Maggio FM, Minafra L, Forte GI, Cammarata FP, Lio D, Messa C, et al. Portrait of inflammatory response to ionizing radiation treatment. J Inflamm. 2015;12:14. doi: 10.1186/s12950-015-0058-3. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 23.Kennedy RJ, Hoper M, Deodhar K, Erwin PJ, Kirk SJ, Gardiner KR. Interleukin 10-deficient colitis: new similarities to human inflammatory bowel disease. Br J Surg. 2000;87(10):1346–1351. doi: 10.1046/j.1365-2168.2000.01615.x. [DOI] [PubMed] [Google Scholar]
- 24.Wirtz S, Neurath M. Mouse models of inflammatory bowel disease☆. Adv Drug Deliv Rev. 2007;59(11):1073–1083. doi: 10.1016/j.addr.2007.07.003. [DOI] [PubMed] [Google Scholar]
- 25.Peluso I, Pallone F, Monteleone G. Interleukin-12 and Th1 immune response in Crohn’s disease: Pathogenetic relevance and therapeutic inplication. World J Gastroenterol WJG. 2006;12(35):5606–5610. doi: 10.3748/wjg.v12.i35.5606. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 26.Laiakis EC, Mak TD, Strawn SJ, Wang YW, Moon BH, Ake P, Fornace AJ., Jr Global metabolomic responses in urine from atm deficient mice in response to LD50/30 gamma irradiation doses. Environ Mol Mutagen. 2018;59(7):576–585. doi: 10.1002/em.22202. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 27.Lee M-LT, Whitmore GA. Power and sample size for DNA microarray studies. Stat Med. 2002;21(23):3543–3570. doi: 10.1002/sim.1335. [DOI] [PubMed] [Google Scholar]
- 28.Simon R, Lam A, Li M-C, Ngan M, Menenzes S, Zhao Y. Analysis of gene expression data using BRB-Array tools. Cancer Inform. 2007;3:11–17. doi: 10.1177/117693510700300022. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 29.Bø TH, Jonassen I. New feature subset selection procedures for classification of expression profiles. Genome Biol. 2002;3(4):research0017. doi: 10.1186/gb-2002-3-4-research0017. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 30.Huang DW, Sherman BT, Lempicki RA. Systematic and integrative analysis of large gene lists using DAVID bioinformatics resources. Nat Protoc. 2009;4(1):44–57. doi: 10.1038/nprot.2008.211. [DOI] [PubMed] [Google Scholar]
- 31.Hall J, Jeggo PA, West C, Gomolka M, Quintens R, Badie C, et al. Ionizing radiation biomarkers in epidemiological studies – an update. Mutat Res Mutat Res. 2017;771:59–84. doi: 10.1016/j.mrrev.2017.01.001. [DOI] [PubMed] [Google Scholar]
- 32.Tucker JD, Joiner MC, Thomas RA, Grever WE, Bakhmutsky MV, Chinkhota CN, et al. Accurate gene expression-based biodosimetry using a minimal set of human gene transcripts. Int J Radiat Oncol. 2014;88(4):933–939. doi: 10.1016/j.ijrobp.2013.11.248. [DOI] [PubMed] [Google Scholar]
- 33.Lucas J, Dressman HK, Suchindran S, Nakamura M, Chao NJ, Himburg H, et al. A translatable predictor of human radiation exposure. Cordes N, editor. PLoS One 2014; 9(9):e107897. [DOI] [PMC free article] [PubMed]
- 34.Tucker JD, Grever WE, Joiner MC, Konski AA, Thomas RA, Smolinski JM, et al. Gene expression-based detection of radiation exposure in mice after treatment with granulocyte Colony-stimulating factor and lipopolysaccharide. Radiat Res. 2012;177(2):209–219. doi: 10.1667/RR2749.1. [DOI] [PubMed] [Google Scholar]
- 35.Budworth H, Snijders AM, Marchetti F, Mannion B, Bhatnagar S, Kwoh E, et al. DNA repair and cell cycle biomarkers of radiation exposure and inflammation stress in human blood. Borgmann K, editor. PLoS One 2012; 7(11):e48619. [DOI] [PMC free article] [PubMed]
- 36.Gao S-Q, Huang L-D, Dai R-J, Chen D-D, Hu W-J, Shan Y-F. Neutrophil-lymphocyte ratio: a controversial marker in predicting Crohn’s disease severity. Int J Clin Exp Pathol. 2015;8(11):14779–14785. [PMC free article] [PubMed] [Google Scholar]
- 37.Wright HL, Moots RJ, Bucknall RC, Edwards SW. Neutrophil function in inflammation and inflammatory diseases. Rheumatology. 2010;49(9):1618–1631. doi: 10.1093/rheumatology/keq045. [DOI] [PubMed] [Google Scholar]
- 38.Mak TW, Grusdat M, Duncan GS, Dostert C, Nonnenmacher Y, Cox M, et al. Glutathione primes T cell metabolism for inflammation. Immunity. 2017;46(6):1089–1090. doi: 10.1016/j.immuni.2017.06.009. [DOI] [PubMed] [Google Scholar]
- 39.White ES, Mantovani AR. Inflammation, wound repair, and fibrosis: reassessing the spectrum of tissue injury and resolution. J Pathol. 2013;229(2):141–144. doi: 10.1002/path.4126. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 40.Xavier RJ, Podolsky DK. Unravelling the pathogenesis of inflammatory bowel disease. Nature. 2007;448(7152):427–434. doi: 10.1038/nature06005. [DOI] [PubMed] [Google Scholar]
- 41.Bouton LA, Ramirez CD, Bailey DP, Yeatman CF, Yue J, Wright HV, et al. Costimulation with interleukin-4 and interleukin-10 induces mast cell apoptosis and cell-cycle arrest: the role of p53 and the mitochondrion. Exp Hematol. 2004;32(12):1137–1145. doi: 10.1016/j.exphem.2004.09.002. [DOI] [PubMed] [Google Scholar]
- 42.Miret C, Molina R, Filella X, García-Carrasco M, Claver G, Ingelmo M, et al. Relationship of p53 with other oncogenes, cytokines and systemic lupus erythematosus activity. Tumor Biol Lond. 2003;24(4):185–188. doi: 10.1159/000074428. [DOI] [PubMed] [Google Scholar]
- 43.Heylmann D, Rödel F, Kindler T, Kaina B. Radiation sensitivity of human and murine peripheral blood lymphocytes, stem and progenitor cells. Biochim Biophys Acta BBA Rev Cancer. 2014;1846(1):121–129. doi: 10.1016/j.bbcan.2014.04.009. [DOI] [PubMed] [Google Scholar]
- 44.Wieczorek M, Abualrous ET, Sticht J, Álvaro-Benito M, Stolzenberg S, Noé F, et al. Major histocompatibility complex (MHC) class I and MHC class II proteins: conformational plasticity in antigen presentation. Front Immunol. 2017;8:292. doi: 10.3389/fimmu.2017.00292. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 45.Skapenko A, Leipe J, Lipsky PE, Schulze-Koops H. The role of the T cell in autoimmune inflammation. Arthritis Res. 2005;7:11. doi: 10.1186/ar1703. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 46.Bradley LM, Dalton DK, Croft M. A direct role for IFN-gamma in regulation of Th1 cell development. J Immunol. 1996;157(4):1350–1358. [PubMed] [Google Scholar]
- 47.Rudqvist N, Laiakis EC, Ghandhi SA, Kumar S, Knotts JD, Chowdhury M, et al. Global gene expression response in mouse models of DNA repair deficiency after gamma irradiation. Radiat Res. 2018;189(4):337–344. doi: 10.1667/RR14862.1. [DOI] [PMC free article] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.
Supplementary Materials
Total white blood cell counts in WT and Il10−/− mice after 2 Gy of radiation exposure A) Counts at Day1 post exposure B) Counts at Day 30 post exposure. Green circles (control) and red squares (irradiated) indicate values for individual animals of the respective genotype. Significant differences *P < 0.05 (n = 8). (JPG 360 kb)
A. Genes showing significant differences in expression levels between WT and Il10−/− mice prior to radiation exposure. B. Gene ontology (GO) analysis genes using DAVID [30]. Top 5 significant (Benjamini corrected P-value< 0.05) GO terms are shown in the bar diagram. C. IPA based network analysis. Top 10 significant (P < 0.05) Canonical pathways are shown in bar diagrams. (XLSX 323 kb)
Real-time qRT-PCR validation of 5 genes (Gstt1, Il18r1, Alas1, Acsbg1, Cyp2b6) relevant to the current study, showing significant difference in expression at baseline level between WT and Il10−/− mice prior to radiation exposure. The data is represented as mean +/− SEM (n = 5) for each genotype. * Significant at P < 0.05 using unpaired t-test. (JPG 398 kb)
BRB-array tools output of differentially expressed genes after radiation exposure in the two genotypes. Results of each genotype are in a separate tab. (XLSX 708 kb)
IPA network analysis of genes involved in T-cell signaling in A. WT and B. Il10−/− mice in response to radiation. Statistical z-score (> 2 or < − 2) was used for prediction of the activation status of the signaling pathway. Orange lines indicate activation (z ≥ 2) and blue indicates inhibition (z ≤ − 2), Grey lines indicate no significant contribution to activity. Up-regulated genes are shown in red and down-regulated genes in green. (JPG 165 kb)
Dose response curves of expression of 4 genes (Cdkn1a, Phlda3, Ifitm1, Ifit3) based on qRT-PCR analysis. Relative gene expression at 0, 2 and 7 Gy of radiation exposure is shown for WT (dotted line) and Il10−/− mice (Solid line). Relative expression was calculated compared to the expression level at 0 Gy (unexposed-sham controls) of WT considered as 1. The expression was normalized against Actin-Beta gene expression. The data is represented as mean +/− SD (n = 5) for each genotype. * Significant at P < 0.05, **Significant at P < 0.01, *** Significant at P < 0.001, using unpaired t-test. (JPG 598 kb)
Data Availability Statement
The microarray data generated in this study have been deposited in the National Center for Biotechnology Information (NCBI) Gene Expression Omnibus (GEO) database with accession number GSE114142 (http://www.ncbi.nlm.nih.gov/geo/query/acc.cgi?acc=GSE114142).