

Abstract
Background:
Mismatch negativity (MMN) is an event-related potential (ERP) component reflecting auditory predictive coding. Repeated standard tones evoke increasing positivity (“repetition positivity”; RP), reflecting strengthening of the standard’s memory trace and the prediction it will recur. Likewise, deviant tones preceded by more standard repetitions evoke greater negativity (“deviant negativity”; DN), reflecting stronger prediction error signaling. These memory trace effects are also evident in MMN difference wave. Here, we assess group differences and test-retest reliability of these indices in schizophrenia patients (SZ) and healthy controls (HC).
Methods:
Electroencephalography was recorded twice, 2-weeks apart, from 43 SZ and 30 HC, during a roving standard paradigm. We examined ERPs to the 3rd, 8th, and 33rd standards (RP), immediately subsequent deviants (DN), and the corresponding MMN. Memory trace effects were assessed by comparing amplitudes associated with the three standard repetition trains.
Results:
Compared to controls, SZ showed reduced MMNs and DNs, but normal RPs. Both groups showed memory trace effects for RP, MMN, and DN, with a trend for attenuated DNs in SZ. Intraclass correlations obtained via this paradigm indicated good to moderate reliabilities for overall MMN, DN and RP, but moderate to poor reliabilities for components associated with short, intermediate, and long standard trains, and poor reliability of their memory trace effects.
Conclusion:
MMN deficits in SZ reflected attenuated prediction error signaling (DN), with relatively intact predictive code formation (RP) and memory trace effects. This roving standard MMN paradigm requires additional development/validation to obtain suitable levels of reliability for use in clinical trials.
Keywords: mismatch negativity, predictive coding, prediction error signaling, psychotic disorders, memory trace, repetition negativity, deviant negativity
Introduction
Mismatch negativity (MMN) is an event-related potential (ERP) component that has been studied extensively in schizophrenia (SZ). MMN is elicited in response to an unexpected, low probability deviant stimulus that is presented after a repeated train of high-probability standard stimuli (e.g., an oddball paradigm) (Näätänen et al., 2007). The MMN is a negative deflection in the ERP that peaks approximately 100–150ms after stimulus onset and is maximal at fronto-central scalp sites (Garrido et al., 2009b; Näätänen et al., 2007; Todd et al., 2013). MMN is traditionally measured in the deviant-standard ERP difference wave to facilitate distinguishing it from the earlier and partially overlapping N100 component evoked by both standards and deviants. Diminished MMN amplitude is a reliable finding in SZ, with meta-analytic reviews reporting large effect sizes (Erickson et al., 2016; Umbricht and Krljes, 2005). Notably, MMN amplitude is also related to functional outcomes in patients (Friedman et al., 2012; Hamilton et al., 2017; Kiang et al., 2007; Light et al., 2015; Light and Braff, 2005; Thomas et al., 2017; Wynn et al., 2010).
MMN has been viewed within several related conceptual frameworks, including echoic memory (Winkler et al., 1993), short-term neuroplasticity (Breen et al., 2014; Garrido et al., 2009a; Näätänen, 2008; Perez et al., 2017; Stephan et al., 2006), and predictive coding (Baldeweg, 2007; Garrido et al., 2009b; Wacongne, 2016; Wacongne et al., 2012; Winkler and Czigler, 2012). Predictive coding may be an especially useful framework to understand important features of SZ, including auditory hallucinations and delusions (Adams et al., 2013; Fletcher and Frith, 2009; Horga et al., 2014; Schmack et al., 2015), and some aspects of impaired cognition and reward processing (Friston et al., 2014; Lalanne et al., 2012; Stephan et al., 2009). Predictive coding is a hierarchical information processing model that posits interactions between lower-order perceptual signals and higher-order cognitive processes in a dynamic, iterative fashion to generate predictions about the environment and compare incoming stimuli with these predictions (Fletcher and Frith, 2009; Nazimek et al., 2012). According to this model, neural responses to stimuli that match predictions are suppressed, whereas stimuli that are unexpected, violating these predictions, trigger a mismatch “prediction error” signal (Friston, 2005; Garrido et al., 2009a; Schultz and Dickinson, 2000). The prediction error signals that updating of expectations is required to accommodate the discrepant stimuli.
From a sensory echoic memory perspective, the elicitation of MMN by a “deviant” stimulus depends on formation and maintenance of a memory trace for what has been “standard” in the recent auditory stream. The predictive coding framework extended this perspective by demonstrating that there is a positive voltage deflection in the standard ERP that increases with successive repetitions of the standard stimulus. This positivity, referred to as the “repetition positivity” (RP), is hypothesized to not only reflect the strength of the memory trace for the standard stimulus, but also the associated prediction that the standard stimulus will recur (Baldeweg, 2007; Baldeweg et al., 2006, 2004, Garrido et al., 2009a, 2009b; Haenschel et al., 2005). When this prediction is violated by the appearance of a deviant stimulus, negativity is evident in the deviant ERP that is hypothesized to signal a prediction error. Importantly, the negativity evoked by the deviant stimulus increases as the number of standards preceding it increases, such that violations of stronger predictions evoke larger prediction error signals (Baldeweg et al., 2004; Baldeweg and Hirsch, 2015; Haenschel et al., 2005; Todd et al., 2013) Although this deviant-evoked negativity has traditionally been identified as the MMN, it has more recently been referred to as the “deviant negativity” (DN) (Baldeweg et al., 2006) to distinguish it from the MMN difference wave, which is now recognized to comprise two constituent parts: 1) the RP, reflecting the strength the memory trace for the standard stimulus and the associated prediction that it will recur, and 2) the DN, reflecting the strength of the prediction error signal (Garrido et al., 2009b; Heilbron and Chait, 2017). Elucidation of predictive coding features of the MMN and its constituent RP and DN components can be optimized using “roving standard” MMN paradigms (Baldeweg et al., 2004; Baldeweg, 2007; Cowan et al., 1993; Haenschel et al., 2005; Winkler et al., 1996).
In a roving standard MMN paradigm, a series of standards is presented, followed by a deviant, as in a typical oddball paradigm. However, after the first appearance of the deviant stimulus, it is repeated over successive trials, becoming the new standard. This new standard series is subsequently interrupted by a new deviant, and the process repeats. Relative to a traditional oddball sequence, the changing physical properties of each successive train of standard stimuli permits examination of the build-up of the memory trace for a new standard. By binning the standards according to their sequential position and generating standard ERPs for early (e.g., position 2), intermediate (e.g., position 6), and late (e.g., position 36) standards, a “memory trace effect” is evident in the RP component of the standard ERPs, with late standards showing a more positive RP than early standards (e.g.,(Baldeweg, 2007)). Similarly, a memory trace effect is evident in the DN, with deviants preceded by a longer train of repeating standards showing a more negative DN than those preceded by a shorter train (Baldeweg et al., 2004; Baldeweg and Hirsch, 2015; Haenschel et al., 2005; Todd et al., 2013). Finally, by calculating difference waves between the corresponding deviant and standard ERPs, a memory trace effect is also evident in the MMN (Baldeweg et al., 2004; Baldeweg and Hirsch, 2015; Haenschel et al., 2005; Todd et al., 2013), reflecting contributions from both RP and DN (Baldeweg, 2007).
Attenuation of the MMN memory trace effect, reflected by a flatter slope of MMN amplitude change across deviants preceded by shorter vs. longer trains of standards, has previously been reported in SZ compared with healthy controls (Baldeweg et al., 2004; Baldeweg and Hirsch, 2015). Moreover, greater attenuation of the MMN memory trace effect was associated with greater deficits in working memory and episodic memory (Baldeweg et al., 2004; Baldeweg and Hirsch, 2015). Beyond reporting an overall reduction in RP and DN amplitudes in SZ, one study (Baldeweg et al., 2004) found the memory trace effect to be attenuated for RP but normal for DN, suggesting that stimulus repetition failed to strengthen the predictive code but nonetheless modulated the strength of the prediction error signal.
The aim of this study was to develop and test a roving standard MMN paradigm to optimize evaluation of predictive coding in people with SZ. We hypothesized that SZ patients, relative to HC participants, would show: 1) overall MMN amplitude reduction when ERPs were derived by traditional averaging of all standards and deviants, 2) reduced amplitudes and attenuated memory trace effects for RP, DN, and MMN associated with short, intermediate, and long trains of standard stimulus repetitions and their immediately subsequent deviants. Additional study aims were to examine cognitive and clinical correlates of the roving standard MMN indices, and to assess the test-retest reliability of the indices over a two-week interval.
Method
Participants
Clinical, cognitive, and EEG assessments were obtained from 43 SZ and 30 HC participants at baseline (Time 1). Of these participants, 43 SZ and 29 HC returned for a two-week follow-up (Time 2). After excluding participants with unusable EEG data, 40 SZ and 30 HC had useable Time 1 data, 38 SZ and 27 HC had usable Time 2 data. In total 43 SZ and 30 HC contributed usable data to at least one timepoint, and 35 SZ and 27 HC contributed usable data to both timepoints. SZ patients were recruited from local clinics and residences, and HC participants were recruited via internet advertisements. Inclusion criteria for patients were: 1) DSM-IV (American Psychiatric Association, 1994) diagnosis of schizophrenia based on SCID-I/P interview (First and Gibbon, 2004), 2) age 18–60 years, and 3) stable outpatient status and no antipsychotic medication changes in the month prior to testing. For HC participants, inclusion criteria were: 1) no history of a DSM-IV diagnosis of recurrent major depressive disorder, bipolar disorder, or schizophrenia-spectrum disorder (including schizoid, schizotypal, paranoid, and avoidant personality disorder) based on SCID-I/P and SCID-II interview (Benjamin, 1994; First et al., 2014), 2) no history of a schizophrenia-spectrum disorder among first-degree relatives, and 3) age 18–60 years. Exclusion criteria for all participants were: 1) history of a neurological disorder or head injury resulting in loss of consciousness, and 2) alcohol or substance abuse or dependence in the three months prior to testing, and 3) benzodiazepine or sedative use in the 12 hours before testing. Urine toxicology screenings were performed at each study visit. Sample characteristics are presented in Table 1.
Table 1.
Clinical and demographic information by group
| SZ (n=43) n (%) |
HC (n=30) n (%) |
χ2 (df), p | |
|---|---|---|---|
| Gender | 29 male (67.4%) | 18 male (60.0%) | χ2(1)=0.43, p=0.51 |
| Hispanic ethnicity | 5 (11.6%) | 6 (20.0%) | χ2(1)=0.97, p=0.33 |
| Race | χ2(3)=5.65, p=0.13 | ||
| Caucasian | 20 (46.5%) | 19 (63.3%) | |
| African American | 19 (44.2%) | 6 (20.0%) | |
| Asian | 2 (4.7%) | 4 (13.3%) | |
| Other | 2 (4.7%) | 1 (3.3%) | |
| Mean (s.d.) | Mean (s.d.) | t (df), p | |
| Age (years) | 47.81 (10.04) | 46.40 (8.50) | t(71)=0.63, p=0.53 |
| Personal education (years) | 12.53 (1.98) | 14.87 (1.55) | t(71)=−5.40, p<0.01 |
| Parental education (years) | 13.23 (4.25) | 15.03 (2.76) | t(68)=−2.03, p=0.05 |
| MCCB Overall Composite Score | 36.79 (13.00) | 46.97 (12.90) | t(71)=−3.30, p<0.01 |
| MCCB Speed of Processing | 43.95 (13.42) | 47.47 (11.42) | t(71)=−1.17, p=0.25 |
| MCCB Attention/Vigilance | 40.44 (12.63) | 49.07 (14.36) | t(71)=−2.71, p=0.01 |
| MCCB Working Memory | 39.91 (11.69) | 48.33 (12.53) | t(71)=−2.94, p<0.01 |
| MCCB Verbal Learning | 42.84 (12.41) | 51.40 (12.37) | t(71)=−2.91, p=0.01 |
| MCCB Visual Learning | 43.09 (10.61) | 48.87 (10.66) | t(71)=−2.28, p=0.03 |
| MCCB Reasoning and Problem Solving | 51.79 (9.17) | 48.27 (11.14) | t(71)=1.48, p=0.14 |
| MCCB Social Cognition | 31.95 (10.41) | 44.97 (11.08) | t(71)=−5.12, p<0.01 |
| RFS Work | 2.56 (1.65) | -- | |
| RFS Independent Living | 4.72 (1.49) | -- | |
| RFS Family | 5.37 (1.43) | -- | |
| RFS Social | 4.60 (1.79) | -- | |
| BPRS total | 41.51 (9.90) | -- | |
| Positive symptoms factor | 2.09 (0.96) | -- | |
| Negative symptoms factor | 2.10 (1.12) | -- | |
| Agitation/Mania symptoms factor | 1.28 (0.35) | -- | |
| Depressive/Anxious symptoms factor | 1.65 (0.56) | -- | |
| CAINS Motivation and Pleasure scale | 14.53 (5.70) | -- | |
| CAINS Expression scale | 4.63 (4.13) | -- | |
Note: SZ, schizophrenia group; HC, healthy control group; MCCB, MATRICS Consensus Cognitive Battery; RFS, Role Functioning Scale; BPRS, Brief Psychiatric Rating Scale; CAINS, Clinical Assessment Interview for Negative Symptoms.
Symptom severity and community functioning were rated by trained raters using the 24-item Brief Psychiatric Rating Scale (BPRS) (Lukoff et al., 1986) and the Clinical Assessment Interview for Negative Symptoms (CAINS) (Kring et al., 2013), and the Role Functioning Scale (RFS) (Goodman et al., 1993). Each rater achieved a median intraclass correlation coefficient (ICC) of 0.80 or higher across all BPRS, CAINS, and RFS. For the SCID, clinical raters demonstrated an overall kappa coefficient, kappa sensitivity, and kappa specificity > 0.75, and a diagnostic accuracy kappa > 0.85. The MATRICS Consensus Cognitive Battery (MCCB) (Nuechterlein and Green, 2006) was administered at Time 1 to assess cognition using age- and gender-corrected domain and overall composite scores.
Procedures
Roving Standard Mismatch Negativity Paradigm:
Prior work has shown that MMNs elicited by different types of auditory deviance (e.g., pitch versus duration) are subserved by different neural generators (Alho, 1995; Csépe, 1995; Deouell et al., 1998; Giard et al., 1990; Molholm et al., 2004; Paavilainen et al., 1991). Further, there is variability in relative sensitivity of pitch vs. duration MMN to SZ across the illness course (Avissar et al., 2017; Bodatsch et al., 2011; Erickson et al., 2016; Haigh et al., 2017; Michie et al., 2000; Näätänen and Kähkönen, 2009; Nagai et al., 2013; Perez et al., 2014; Todd et al., 2008; Umbricht and Krljes, 2005) in that paradigms that produce larger MMNs tend to show larger MMN deficits in SZ (Avissar et al., 2017; Javitt et al., 1998). Also, MMN amplitude is enhanced when multiple types of deviance are combined in a single stimulus (Levanen et al., 1993; Paavilainen et al., 2001; Schroger, 1995; Takegata et al., 1999; Wolff and Schröger, 2001). Hence, the paradigm tested here used pitch+duration “double deviants”, similar to our prior studies (Hay et al., 2015; Perez et al., 2014).
Participants completed the roving standard MMN at Time 1 and Time 2. The paradigm comprised five blocks (block duration: 317s), each separated by a 30s break, during which tones of 50 ms or 100 ms duration, varying in pitch between 700Hz - 1250Hz, were presented binaurally (80 dB, 5ms rise/fall time, 400ms SOA) through insert earphones (ER-2, Etymotic Research, Inc., Elk Grove Village, IL). The stimulus sequence comprised short (3), intermediate (8), and long (33) trains of identical standard tones followed by a deviant tone differing from the standard in both pitch (minimum change 100 Hz, maximum change 300 Hz) and duration (± 50 ms). The deviant tone then repeated, effectively becoming the new standard, with a train of 3, 8, or 33 standard repetitions preceding the next deviant (see Figure 1). A randomly permuted sequence of short, intermediate, and long trains of identical standard repetitions was generated for each block, with all 5 blocks comprising 90 occurrences of each train length. Participants were instructed to ignore the tones while they performed a tactile oddball distractor task and maintained visual fixation on a central cross on a computer screen. In the tactile oddball task, participants held a game controller with both hands that transmitted a pseudorandom sequence of three distinct vibration patterns: a frequent (p=0.8) standard (right side), an infrequent (p=0.10) target (right side), and an infrequent (p=0.10) foil (left side). Participants responded with a button press to targets only.
Figure 1.
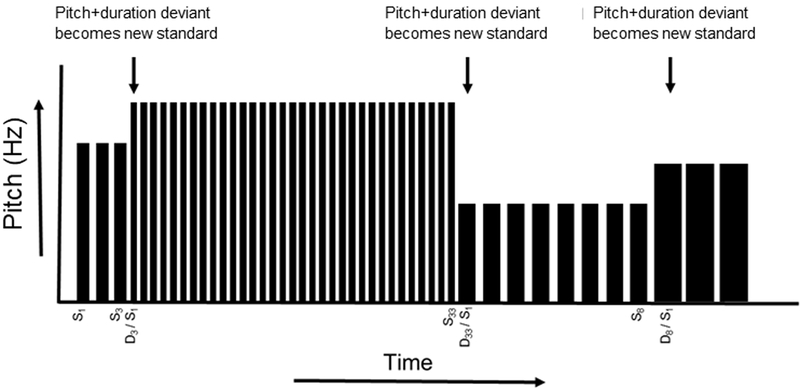
Roving standard MMN task design. For the roving standard MMN paradigm, an identical standard tone is repeated 3, 8, or 33 times within each stimulus train. For each change to a new stimulus train, the first stimulus constitutes a deviant tone, differing from the preceding train of standards in both pitch and duration. The deviant tone then becomes the new standard, repeating 3, 8, or 33 times until the next deviant appears, Note: S, standard stimulus; D, deviant stimulus.
Specific standard and deviant trials were averaged to generate ERP waveforms and corresponding deviant-standard difference waves, and all components (RP, DN, MMN) were measured by calculating the mean amplitude between 100–200ms at electrode Fz from their respective ERPs. RPs were assessed in ERPs derived from standard trials in sequence positions 3, 8, or 33 within local trains of standard repetitions, resulting in 270 trials for the 3rd standard (RP3), 180 trials for the 8th standard (RP8), and 90 trials for the 33rd standard (RP33). Likewise, DNs were assessed in ERPs derived from deviant trials occurring after 3 (DN3), 8 (DN8), or 33 (DN33) standard repetitions, with each ERP comprising 90 trials. MMNs associated with different numbers of standard repetitions preceding the deviant stimulus (MMN3, MMN8, MMN33) were assessed in corresponding deviant-standard difference waves (e.g., MMN3=DN3-RP3). “Memory trace” indices for RP, DN and MMN, were calculated as the amplitude difference between components associated with long versus short trains of standard repetitions (e.g., DNMT=DN33-DN3) for use in correlational analyses. In addition, overall mean RP, DN, and MMN were measured from ERP averages of all standards, all deviants, and their corresponding difference waves. Additional information about the EEG procedures are in Supplementary Materials.
Data analysis plan
The groups were compared on demographic, clinical, and cognitive variables using χ2 and independent sample t-tests. MMN, RP, and DN amplitudes were analyzed separately using 2 (Group; SZ, HC) x 3 (Standard Repetition; 3, 8, 33) x 2 (Time; Time 1, Time 2) mixed models. In these models, memory trace effects were assessed using two a priori orthogonal reverse Helmert contrasts for standard repetitions: 1) intermediate (8) vs. short (3), and 2) long (33) vs. mean of intermediate and short (8,3). Overall MMN amplitude was assessed using a 2 (Group; SZ, HC) x 2 (Time; Time 1, Time 2) mixed model. Using hierarchical linear regression models, MCCB cognitive measures were each regressed on a MMN index (MMNMT, RPMT, DNMT, overall MMN amplitude), Group, and the MMN index x Group interaction to test for significant regression line slope differences between the groups. Significant slope differences were followed up with bivariate correlations within each group. When the MMN index x Group interaction was not significant, it was dropped from the model, and the common slope was then tested for significance. Relationships of clinical and functioning measures with MMN indices were assessed in the SZ group with bivariate correlations. For each family of regression or correlation analyses (cognitive, clinical, functioning), correction for multiple tests were imposed using Bonferroni correction (family wise-corrected alpha set to p<0.05). Finally, ICC (3,1) coefficients (Shrout and Fleiss, 1979) were calculated to assess test-retest reliability of the roving standard indices.
Results
Demographic, clinical, cognitive, and functioning data are presented in Table 1. The groups were well matched on age, gender, race, and ethnicity. The level of personal education was significantly lower, and the level of parental education was marginally lower, in SZ compared to HC.
Means of the roving standard MMN indices are presented by group in Table 2. The number of trials included in ERP averages after artifact rejection are presented in Supplementary Materials. Grand average waveforms for RP, DN, and MMN and topographic maps across the three standard repetition train lengths for each group are presented in Figures 2 and 3. Average amplitudes for the MMN, RP, and DN associated with short, intermediate, and long trains of standard repetitions for each group are plotted in Figure 4. Descriptive data and mixed models results are presented in Tables 2 and 3.
Table 2.
Descriptive statistics for roving standard MMN indices and mixed model analyses
| Roving Standard MMN indices | SZ Time 1 n=40 Mean (S.D.) |
SZ Time 2 n=38 Mean (S.D.) |
HC Time 1 n=30 Mean (S.D.) |
HC Time 2 n=27 Mean (S.D.) |
|---|---|---|---|---|
| MMN3 | −0.19 (1.19) | 0.02 (1.54) | −0.49 (1.25) | −0.93 (1.00) |
| MMN8 | −0.64 (2.03) | −0.51 (1.63) | −0.67 (1.40) | −0.60 (1.59) |
| MMN33 | −1.64 (2.06) | −1.21 (2.15) | −1.99 (1.88) | −2.61 (2.16) |
| RP3 | −0.17 (0.98) | −0.49 (1.02) | −0.36 (0.87) | −0.36 (0.93) |
| RP8 | 0.04 (0.94) | −0.09 (1.10) | −0.11 (1.03) | −0.29 (0.96) |
| RP33 | 0.53 (1.22) | 0.41 (1.44) | 0.43 (1.01) | 0.52 (1.09) |
| DN3 | −0.36 (1.42) | −0.47 (1.46) | −0.86 (1.34) | −1.29 (1.28) |
| DN8 | −0.59 (1.78) | −0.60 (1.88) | −0.96 (1.42) | −0.89 (1.25) |
| DN33 | −1.12 (1.66) | −0.80 (1.74) | −1.72 (1.65) | −2.09 (1.80) |
| Overall MMN amplitude | −0.74 (1.12) | −0.57 (1.16) | −1.08 (1.14) | −1.38 (1.06) |
Note: SZ, schizophrenia group; HC, healthy control group; RP, repetition positivity; DN, deviant negativity.
Figure 2.


a) MMN averaged waveforms (electrode Fz), and b) MMN topographic maps, collapsed across Time 1 and Time 2. Note: HC, healthy control group; SZ, schizophrenia group; MMN, mismatch negativity. Shaded box denotes 100–200ms time window.
Figure 3.

RP and DN averaged waveforms (electrode Fz) collapsed across Time 1 and Time 2 Note: HC, healthy control group; SZ, schizophrenia group; RP, repetition positivity; DN, deviant negativity. Shaded box denotes 100–200ms time window.
Figure 4.

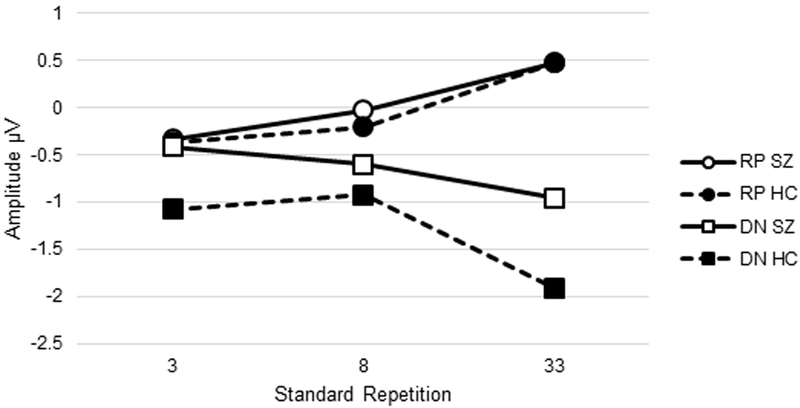
Average amplitude for a) MMN and b) RP to standards and DN to deviants associated with different numbers of standard repetitions in SZ and HC. Note: MMN, mismatch negativity; RP, repetition positivity; DN, deviant negativity; HC, healthy control group; SZ, schizophrenia group.
Table 3.
Mixed model analyses
| Roving Standard MMN indices | Contrast | F/t (df), p |
|---|---|---|
| MMN3,8,33 | Effect of Group: | F(1,66.62)=4.14, p=0.05 |
| Effect of Standard Repetition: | F(2,327.00)=37.44, p<0.001 | |
| Effect of Time: | F(1,347.64)=0.03, p=0.87 | |
| Group × Time: | F(1,347.64)=4.82, p=0.03 | |
| Reverse Helmert Contrasts | ||
| MMN3 vs MMN8 | t(327.00)=−1.13, p=0.26 | |
| MMN3 vs. MMN8 × Group: | t(327.00)=1.54, p=0.12 | |
| MMN3 vs. MMN8 × Time: | t(327.00)=0.59, p=0.56 | |
| MMN3 vs. MMN8 × Group × Time: | t(327.00)=−0.82, p=0.42 | |
| MMN3,8 vs. MMN33: | t(327.00)=−8.58, p<0.001 | |
| MMN3,8 vs. MMN33 × Group: | t(327.00)=−1.67, p=0.10 | |
| MMN3,8 vs. MMN33 × Time: | t(327.00)=−0.27, p=0.79 | |
| MMN3,8 vs. MMN33 × Group × Time: | t(327.00)=1.10, p=0.27 | |
| RP3,8,33 | Effect of Group: | F(1,73.12)=0.06, p=0.81 |
| Effect of Standard Repetition: | F(2,332.88)=35.27, p<0.001 | |
| Effect of Time: | F(1,346.82)=2.00, p=0.16 | |
| Group × Time: | F(1,346.82)=1.75, p=0.19 | |
| Reverse Helmert Contrasts | ||
| RP3 vs. RP8: | t(332.88)=2.34, p=0.02 | |
| RP3 vs. RP8 × Group: | t(332.88)=−0.72, p=0.47 | |
| RP3 vs. RP8 × Time: | t(332.88)=−0.01, p=0.99 | |
| RP3 vs. RP8 × Group × Time: | t(332.88)=0.93, p=0.36 | |
| RP3,8 vs. RP33: | t(332.88)=8.07, p<0.001 | |
| RP3,8 vs. RP33 × Group: | t(332.88)=0.62, p=0.54 | |
| RP3,8 vs. RP33 × Time: | t(332.88)=0.84, p=0.40 | |
| RP3,8 vs. RP33 × Group × Time: | t(332.88)=−0.19, p=0.85 | |
| DN3,8,33 | Effect of Group: | F(1,70.86)=4.39, p=0.04 |
| Effect of Standard Repetition: | F(2,330.38)=15.13, p<0.001 | |
| Effect of Time: | F(1,342.79)=0.38, p=0.51 | |
| Group × Time: | F(1,342.79)=2.28, p=0.13 | |
| Reverse Helmert Contrasts | ||
| DN3 vs. DN8: | t(330.38)=−0.12, p=0.91 | |
| DN3 vs. DN8 × Group: | t(330.38)=1.17, p=0.24 | |
| DN3 vs. DN8 × Time: | t(330.38)=1.06, p=0.29 | |
| DN3 vs. DN8 × Group × Time: | t(330.38)=−0.70, p=0.48 | |
| DN3,8 vs. DN33: | t(330.38)=−5.50, p<0.001 | |
| DN3,8 vs. DN33 × Group: | t(330.38)=−1.84, p=0.07 | |
| DN3,8 vs. DN33 × Time: | t(330.38)=0.37, p=0.71 | |
| DN3,8 vs. DN33 × Group × Time: | t(330.38)=1.14, p=0.25 | |
| Overall MMN amplitude | Effect of Group: | F(1,68.51)=4.49, p=0.04 |
| Effect of Time: | F(1,61,21)=0.10, p=0.75 | |
| Group × Time: | F(1,61.21)=4.60, p=0.04 | |
Note: SZ, schizophrenia group; HC, healthy control group; RP, repetition positivity; DN, deviant negativity.
Mixed model analyses of roving standard ERP indices.
The mixed model analysis of MMN amplitudes revealed a significant main effect of Group, with SZ patients showing smaller (i.e., less negative) MMN than HC participants. This effect interacted significantly with Time, with the MMN amplitude deficit in SZ patients, relative to HC, being larger at Time 2 than at Time 1. Despite this, neither the increase in MMN amplitude over time in HC [F(1,147.25)=2.81, p=0.10], nor the decrease in MMN amplitude over time in SZ [F(1,203.29)=2.20, p=0.14], were significant. Two reverse Helmert contrasts among levels of the Standard Repetition factor were used to assess the MMN memory trace effect. The MMN3 vs MMN8 contrast was not significant, nor did it significantly interact with Group, Time, or with Group x Time. However, the MMN3,8 vs. MMN33 contrast was highly significant (p<0.001) reflecting a marked increase in MMN amplitude (i.e., increased negativity) when deviants were preceded by long trains of standard repetitions, relative to the average MMN associated with short and intermediate trains of standard repetitions. While this MMN memory trace effect appeared to be larger in the HC than in the SZ group (see Figure 3a), the difference was not significant (p=0.10). Furthermore, the MMN3,8 vs. MMN33 contrast effect did not significantly interact with Time or with Group x Time. No other main effects or interactions were significant.
Next, RP and DN across the three Standard Repetition train lengths and two Time points in SZ and HC were tested in two separate mixed model analyses. For RP, memory trace effects were significant for both reverse Helmert contrasts, indicating that RP amplitude modestly (p=0.02) increased in positivity for the 8th standard relative to the 3rd standard in repeating standard stimulus trains, and strongly (p<0.001) increased for the 33rd standard relative to the mean of the 3rd and 8th standards. These memory trace effects on the RP did not significantly interact with Group, Time, or Group x Time.
For DN, there was a significant main effect of Group (p=0.04), with HC exhibiting a larger (i.e., more negative) DN amplitude than SZ. This effect did not significantly interact with Time, nor was the main effect of Time significant. Memory trace effects on DN were not significant for the DN3 vs. DN8 contrast, nor did this contrast significantly interact with Group, Time, or Group x Time. However, when the mean of DN3 and DN8 were contrasted with DN33, a strong memory trace effect was evident (p<0.001). This significant memory trace effect on DN showed a trend interaction with Group (p=0.07), with the increase in amplitude (i.e., increased negativity) of DN33, relative to DN3,8, being larger in HC [t(141.54)=−4.87, p<0.001] than in SZ [t(189.67)=−2.80, p=0.06].
For overall MMN amplitude (i.e., based on ERPs derived from all deviants and standards), there was a significant main effect of Group (p=0.04), with HC exhibiting larger (i.e., more negative) MMN amplitude compared to SZ. In addition, a significant Group x Time interaction (p=0.04) was present, indicating that the SZ deficit in overall MMN amplitude relative to HC was greater at Time 2 than at Time 1. Despite this interaction, neither the slight increase in MMN amplitude in HC over time [F(1,27.98)=2.89, p=0.10], nor the slight decrease in MMN amplitude in SZ over time [F(1,33.61)=1.89, p=0.19], was significant.
Cognitive, clinical, and functioning relationships with roving standard MMN indices.
The regression analyses and correlation matrices are presented in Supplementary Materials. Although a few differential associations between the memory trace indices and MCCB variables were found in SZ and HC, none survived correction for multiple tests. Similarly, correlations of clinical or community functioning with MMN indices in the SZ group were not significant after correction for multiple tests.
Test-retest reliability of roving standard ERP indices.
Test-retest reliability coefficients (ICCs) for the roving standard ERP indices are presented in Table 4. In brief, the ICCs indicated good to moderate reliabilities for overall MMN, DN and RP. However, when looking at the values for the separate short, intermediate, and long standard trains, the reliabilities were moderate to poor. Finally, the reliabilities were poor for memory trace effects.
Table 4.
Test-retest reliability coefficients for roving standard MMN indices.
| ICC | ||
|---|---|---|
| Roving standard MMN Measure | SZ n=35 |
HC n=27 |
| MMN3 | 0.00 | 0.01 |
| MMN8 | 0.35* | 0.34* |
| MMN33 | 0.30* | 0.56** |
| RP3 | 0.59** | 0.53** |
| RP8 | 0.58** | 0.59** |
| RP33 | 0.20 | 0.56** |
| DN3 | 0.30* | 0.37* |
| DN8 | 0.48** | 0.07 |
| DN33 | 0.57** | 0.59** |
| MMNMT | 0.04 | 0.37* |
| RPMT | 0.05 | 0.30 |
| DNMT | 0.14 | 0.22 |
| Overall MMN | 0.55** | 0.64** |
| Overall RP | 0.74** | 0.86** |
| Overall DN | 0.72** | 0.67** |
Note:
p<0.05,
p<0.01;
ICC, intra-class correlation, using two-way mixed single measures consistency formula (ICC 3, 1 from Shrout and Fleiss, 1979); SZ, schizophrenia group; HC, healthy control group; MMN, mismatch negativity; RP, repetition positivity; DN, deviant negativity. MT, memory trace effect, defined for each component by subtracting value associated with repeated standard 3 from value associated with repeated standard 33. Overall MMN, RP, and DN measures are based on ERP averages of all standards, all deviants, and the resulting ERP deviant-standard difference waves.
Discussion
Using a roving standard MMN paradigm, with trains of 3, 8, or 33 standards before a deviant that heralded the onset of a new standard train, a significant memory trace effect was evident for MMN in both HC and SZ groups, with MMN amplitude being substantially larger (i.e., more negative) when deviants were preceded by 33 versus 3 or 8 standards. This memory trace effect is thought to reflect the build-up of a stronger memory trace for the standard, and a stronger prediction that it will recur, as the number of standard repetitions increase, and a correspondingly larger prediction error signal to deviants preceded by longer trains of standards. Although this memory trace effect tended to be smaller in the SZ patients than in the HC controls, this difference was not significant. Moreover, MMN did not show a significant memory trace effect in either group when deviants were preceded by 8 standards relative to 3 standards.
An advantage of the roving standard MMN paradigm is its ability to disentangle the two processes that contribute to MMN: 1) the build-up of the memory trace for the standard and the development of the prior expectation that it will recur, reflected by the RP response to repeated standard sounds, and 2) the prediction error signal elicited by stimuli that violate prior expectations, reflected by the DN response to infrequent deviant sounds. A corresponding limitation of the roving standard paradigm is that it does not equate the number of trials for the various repetition sequence lengths for RP. Our results showed the predicted memory trace effect on the RP, with its amplitude modestly but significantly increasing to the 8th repeated standard relative to the 3rd, and a more prominent increase to the 33rd standard. These effects were equally evident in both groups. While the standard repetition train lengths used in this study (i.e., 3, 8, and 33) were similar to those used in prior roving standard MMN paradigms (e.g., (Baldeweg et al., 2004)), a memory trace effect on the DN and the MMN was not evident in either group when comparing deviants preceded by 3 versus 8 standards. However, a memory trace effect was seen in increased negativity for deviants preceded by 33 standards relative to 8 or 3 standards, and this effect was attenuated at a trend level in SZ relative to HC participants.
In addition to these memory trace effects, our results replicated the expected reduction in overall MMN amplitude in SZ patients, relative to HC, both in the overall MMN measured from ERP averages of all standards and deviants, and in the standard repetition analysis. Based on the separate analyses of MMN’s constituents (i.e., DN and RP), the SZ patient MMN deficit can be attributed to a significant overall reduction in DN, but not RP, amplitude. Overall, our findings support the idea that SZ patients show aberrant prediction error signaling, and they show a trend toward reduced strengthening of this prediction error signal with more standard repetitions. Furthermore, our findings with RP suggest that SZ patients do not have deficits in forming memory traces of, or establishing and building expectations for, recurring auditory stimuli. The overall reduction in amplitude of the DN and MMN in this SZ sample are congruent with the findings of Baldeweg et al. (2004). However, the studies differ in that Baldeweg et al. (2004) found that the memory trace was diminished for RP and relatively intact for DN in SZ, whereas we found the opposite pattern. The roving standard paradigm used by Baldeweg et al. (2004) differed from the one used in the current study with regard to the type of deviant and the number of repetitions of the standard, which may contribute to the different pattern of findings. It is also possible that the poor reliability of the roving standard indices may contribute to the different findings across studies.
Previous studies have reported significant relationships between MMN and symptoms, cognitive performance, and functional outcome in schizophrenia (Baldeweg et al., 2004; Baldeweg and Hirsch, 2015; Friedman et al., 2012; Hamilton et al., 2017; Kiang et al., 2007; Light et al., 2015; Light and Braff, 2005; Wynn et al., 2010). We expected to find similar correlations with the predictive coding indices derived from this roving standard paradigm. However, while we found some moderately-sized associations within groups, none of the correlations survived correction for multiple tests. One could speculate that predictive coding-related measures from the roving standard MMN paradigm might be more relevant to shorter term mechanisms of neural plasticity that support new learning, rather than stable cognitive abilities that may be learned over a lifetime and may be more closely related to daily functioning.
MMN from traditional (i.e., non-roving) paradigms have previously been shown to have moderate to good test-retest reliability in SZ (Biagianti et al., 2017; Hall et al., 2006; Lew et al., 2007; Light et al., 2012; Light and Braff, 2005). The ICCs for the overall MMN derived in our study is comparable to some of these estimates. This paper represents the first report of test-retest reliability of MMN, RP, and DN components, as well as the corresponding memory trace effects, derived from a roving standard MMN paradigm in SZ and HC. Within the two groups, the roving ERP indices for RP, DN, and MMN at each of the three repetition sequences showed poor to moderate test-retest reliability. Notably, there was a general tendency for reliability to improve as the number of standard presentations in a sequence increased, and correspondingly, as the amplitudes of the components increased. The memory trace effects, i.e., the difference in ERP amplitude between the longest and shortest repetition sequences for MMN, RP, and DN, showed particularly poor reliability. Possibly, the memory trace effects are more susceptible to state-related influences and noise than the component amplitudes from which they are derived.
Even though the paradigm included more trials of the short sequences (i.e. 3 and 8), even this might not have been enough to establish high levels of reliability because shorter sequences would be expected to have more noise than longer sequences. This potential benefit of increasing trial numbers is consistent with the observation that the overall MMN, RP, and DN, derived from all available standard and deviant trials, achieved moderate to strong reliability. However, the potential impact on testing session length and subject burden does present a limitation for the number of long sequence trials that can be included in a testing session. Moreover, it is not possible to equate the number of trials presented to the subject for the various repetition sequence lengths for RP. It is also possible that the tactile distractor task used in this study may have impacted the reliability estimates of the MMN indices, as MMN latency and amplitude can be influenced by distractor task attentional demands and modality (Rissling et al., 2013). Further study of the impact of altering task parameters, including the number of standard repetitions in the sequence, the type of deviants, and the type of distractor task, on psychometric properties is warranted.
In conclusion, our results demonstrate the potential utility of roving standard MMN paradigms and applications of the predictive coding framework to the study of SZ. These data demonstrate that MMN deficits in SZ patients can be understood to reflect deficient prediction error signaling, rather than reduced memory trace formation for the standard. The reliability analyses suggest that further refinements to roving standard paradigms will be needed to yield adequate reliability for roving standard ERP indices if they are to be used in longitudinal studies or clinical trials.
Supplementary Material
Acknowledgement
We thank the research subjects for participating in this study. We gratefully acknowledge Ana Cecilia Myers, M.S. for coordinating the study, Michelle Dolinsky, B.A. for subject recruitment efforts, Courtney Fazli, B.S., Gabrielle Pascual, B.S., and Nora Polon, B.A. for their assistance with data collection.
Financial Support
This study was supported by funding from FORUM Pharmaceuticals, Inc. (PIs: Green & Marder), the Veterans Administration VISN 22 Mental Illness Research, Education, and Clinical Center (MIRECC), the Veterans Administration Research Enhancement Award Program (REAP) on Enhancing Community Integration for Homeless Veterans. Dr. McCleery is currently supported by a career development award from NIH (K23MH108829), and previously by an NIH institutional training fellowship (T32MH096682).
Footnotes
Conflict of Interest
Dr. McCleery has received compensation from MedAvante-Prophase, Inc. for clinical assessment services. Dr. Mathalon serves as a consultant for Boehringer Ingelheim and Takeda. Dr. Marder has been a consultant for FORUM, Allergan, Lundbeck, Roche, Otsuka, Merck, Teva, Takeda, and Newron. He has received research support from FORUM and Neurocrine. Dr. Green has been a consultant to AiCure, Lundbeck, and Takeda, and he is on the scientific board of Cadent. He has received research funds from FORUM. Dr. Wynn, Dr. Hellemann, and Mr. Roach report no disclosures.
Ethical Standards
The authors assert that all procedures contributing to this work comply with the ethical standards of the relevant national and institutional committees on human experimentation and with the Helsinki Declaration of 1975, as revised in 2008.
References
- Adams RA, Stephan KE, Brown HR, Frith CD, Friston KJ, 2013. The Computational Anatomy of Psychosis. Frontiers in Psychiatry 4, 1–26. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Alho K, 1995. Cerebral generators of mismatch negativity (MMN) and its magnetic counterpart (MMNm) elicited by sound changes. Ear and Hearing 16, 38–51. [DOI] [PubMed] [Google Scholar]
- American Psychiatric Association, 1994. Diagnostic and statistical manual of mental disorders, 4th ed. (DSM-IV), Washington, DC: American Psychiatric Association. [Google Scholar]
- Avissar M, Xie S, Vail B, Lopez-Calderon J, Wang Y, Javitt DC, 2017. Meta-analysis of mismatch negativity to simple versus complex deviants in schizophrenia. Schizophrenia Research 191, 25–34. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Baldeweg T, 2007. ERP repetition effects and mismatch negativity generation: A predictive coding perspective. Journal of Psychophysiology 21, 204–213. [Google Scholar]
- Baldeweg T, Hirsch SR, 2015. Mismatch negativity indexes illness-specific impairments of cortical plasticity in schizophrenia: a comparison with bipolar disorder and Alzheimer’s disease. International Journal of Psychophysiology 95, 145–155. [DOI] [PubMed] [Google Scholar]
- Baldeweg T, Klugman A, Gruzelier J, Hirsch SR, 2004. Mismatch negativity potentials and cognitive impairment in schizophrenia. Schizophrenia Research 69, 203–217. [DOI] [PubMed] [Google Scholar]
- Baldeweg T, Wong D, Stephan KE, 2006. Nicotinic modulation of human auditory sensory memory: Evidence from mismatch negativity potentials. International Journal of Psychophysiology 59, 49–58. [DOI] [PubMed] [Google Scholar]
- Benjamin L, 1994. Structured Clinical Interview for DSM-IV Axis II personality disorders (SCID II). New York, NY: Biometric Research Department, New York State Psychiatric Institute. [Google Scholar]
- Biagianti B, Roach BJ, Fisher M, Loewy R, Ford JM, Vinogradov S, Mathalon DH, 2017. Trait aspects of auditory mismatch negativity predict response to auditory training in individuals with early illness schizophrenia. Neuropsychiatric Electrophysiology 3, 2. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Bodatsch M, Ruhrmann S, Wagner M, Müller R, Schultze-Lutter F, Frommann I, Brinkmeyer J, Gaebel W, Maier W, Klosterkötter J, Brockhaus-Dumke A, 2011. Prediction of Psychosis by Mismatch Negativity. Biological Psychiatry 69, 959–966. [DOI] [PubMed] [Google Scholar]
- Breen EC, Perez C, Olmstead R, Eisenberger N, Irwin MR, 2014. 135. Comparison of multiplex immunoassays and ELISAs for the determination of circulating levels of inflammatory cytokines. Brain, Behavior, and Immunity 40, e39. [Google Scholar]
- Cowan N, Winkler I, Teder W, Naatanen R, 1993. Memory prerequisites of mismatch negativity in the auditory event-related potential (ERP). Journal of Experimental Psychology: Learning, Memory, and Cognition 19, 909–921. [DOI] [PubMed] [Google Scholar]
- Csépe V, 1995. On the origin and development of the mismatch negativity. Ear and Hearing 16, 91–104. [DOI] [PubMed] [Google Scholar]
- Deouell LY, Bentin S, Giard M-H, 1998. Mismatch negativity in dichotic listening: evidence for interhemispheric differences and multiple generators. Psychophysiology 35, 355–365. [PubMed] [Google Scholar]
- Erickson MA, Ruffle A, Gold JM, 2016. A Meta-Analysis of Mismatch Negativity in Schizophrenia: From Clinical Risk to Disease Specificity and Progression. Biological Psychiatry 79, 980–987. [DOI] [PMC free article] [PubMed] [Google Scholar]
- First M, Gibbon M, 2004. The Structured Clinical Interview for DSM-IV Axis I Disorders (SCID-I) and the Structured Clinical Interview for DSM-IV Axis II Disorders (SCID-II) In Hilsenroth MJ & Segal DL (Eds.), Comprehensive handbook of psychological assessment, Vol. 2 Personality assessment (pp. 134–143). Hoboken, NJ, US: John Wiley & Sons Inc. [Google Scholar]
- First M, Williams J, Karg R, Spitzer R, 2014. Structured Clinical Interview for DSM-5 Disorders (SCID-5), Research Version. Arlington, VA: American Psychiatric Association. [Google Scholar]
- Fletcher PC, Frith CD, 2009. Perceiving is believing: A Bayesian approach to explaining the positive symptoms of schizophrenia. Nature Reviews Neuroscience 10, 48–58. [DOI] [PubMed] [Google Scholar]
- Friedman T, Sehatpour P, Dias E, Perrin M, Javitt DC, 2012. Differential Relationships of Mismatch Negativity and Visual P1 Deficits to Premorbid Characteristics and Functional Outcome in Schizophrenia. Biological Psychiatry 71, 521–529. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Friston K, 2005. A theory of cortical responses. Philosophical Transactions of the Royal Society B: Biological Sciences 360, 815–36. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Friston KJ, Stephan KE, Montague R, Dolan RJ, 2014. Computational psychiatry: The brain as a phantastic organ. The Lancet Psychiatry 1, 148–158. [DOI] [PubMed] [Google Scholar]
- Garrido MI, Kilner JM, Kiebel SJ, Stephan KE, Baldeweg T, Friston KJ, 2009a. Repetition suppression and plasticity in the human brain. Neuroimage 48, 269–279. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Garrido MI, Kilner JM, Stephan KE, Friston KJ, 2009b. The mismatch negativity: A review of underlying mechanisms. Clinical Neurophysiology 120, 453–463. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Giard M, Perrin F, Pernier J, Bouchet P, 1990. Brain generators implicated in the processing of auditory stimulus deviance: a topographic event‐related potential study. Psychophysiology 27, 627–640. [DOI] [PubMed] [Google Scholar]
- Goodman SH, Sewell DR, Cooley EL, Leavitt N, 1993. Assessing levels of adaptive functioning: the Role Functioning Scale. Community Mental Health Journal 29, 119–131. [DOI] [PubMed] [Google Scholar]
- Haenschel C, Vernon DJ, Dwivedi P, Gruzelier JH, Baldeweg T, 2005. Event-related brain potential correlates of human auditory sensory memory-trace formation. Journal of Neuroscience 25, 10494–10501. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Haigh SM, Coffman BA, Salisbury DF, 2017. Mismatch Negativity in First-Episode Schizophrenia. Clinical EEG and Neuroscience 48, 3–10. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Hall MH, Schulze K, Rijsdijk F, Picchioni M, Ettinger U, Bramon E, Freedman R, Murray RM, Sham P, 2006. Heritability and Reliability of P300, P50 and Duration Mismatch Negativity. Behavior Genetics 36, 845–857. [DOI] [PubMed] [Google Scholar]
- Hamilton HK, Perez VB, Ford JM, Roach BJ, Jaeger J, Mathalon DH, 2017. Mismatch Negativity But Not P300 Is Associated With Functional Disability in Schizophrenia. Schizophrenia Bulletin 44, 492–504. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Hay RA, Roach BJ, Srihari VH, Woods SW, Ford JM, Mathalon DH, 2015. Equivalent mismatch negativity deficits across deviant types in early illness schizophrenia-spectrum patients. Biological Psychology 105C, 130–137. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Heilbron M, Chait M, 2017. Great expectations: Is there evidence for predictive coding in auditory cortex? Neuroscience 389, 54–73. [DOI] [PubMed] [Google Scholar]
- Horga G, Schatz KC, Abi-Dargham A, Peterson BS, 2014. Deficits in Predictive Coding Underlie Hallucinations in Schizophrenia. Journal of Neuroscience 34, 8072–8082. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Javitt DC, Grochowski S, Shelley AM, Ritter W, 1998. Impaired mismatch negativity (MMN) generation in schizophrenia as a function of stimulus deviance, probability, and interstimulus/interdeviant interval. Electroencephalography and Clinical Neurophysiology 108, 143–53. [DOI] [PubMed] [Google Scholar]
- Kiang M, Light GA, Prugh J, Coulson S, Braff DL, Kutas M, 2007. Cognitive, neurophysiological, and functional correlates of proverb interpretation abnormalities in schizophrenia. Journal of the International Neuropsychological Society 13, 653–663. [DOI] [PubMed] [Google Scholar]
- Kring AM, Gur RE, Blanchard JJ, Horan WP, Reise SP, 2013. The Clinical Assessment Interview for Negative Symptoms (CAINS): Final Development and Validation. American Journal of Psychiatry 170, 165–172. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Lalanne L, Van Assche M, Giersch A, 2012. When predictive mechanisms go wrong: Disordered visual synchrony thresholds in schizophrenia. Schizophrenia Bulletin 38, 506–513. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Levanen S, Hari R, McEvoy L, Sams M, 1993. Responses of the human auditory cortex to changes in one versus two stimulus features. Experimental Brain Research 97, 177–183. [DOI] [PubMed] [Google Scholar]
- Lew HL, Gray M, Poole JH, 2007. Temporal stability of auditory event-related potentials in healthy individuals and patients with traumatic brain injury. Journal of Clinical Neurophysiology 24, 392–397. [DOI] [PubMed] [Google Scholar]
- Light GA, Braff DL, 2005. Mismatch negativity deficits are associated with poor functioning in schizophrenia patients. Archives of General Psychiatry 62, 127–136. [DOI] [PubMed] [Google Scholar]
- Light GA, Swerdlow NR, Rissling AJ, Radant A, Sugar CA, Sprock J, Pela M, Geyer MA, Braff DL, 2012. Characterization of Neurophysiologic and Neurocognitive Biomarkers for Use in Genomic and Clinical Outcome Studies of Schizophrenia. PLoS One 7, e39434. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Light GA, Swerdlow NR, Thomas ML, Calkins ME, Green MF, Greenwood TA, Gur RE, Gur RC, Lazzeroni LC, Nuechterlein KH, Pela M, Radant AD, Seidman LJ, Sharp RF, Siever LJ, Silverman JM, Sprock J, Stone WS, Sugar CA, Tsuang DW, Tsuang MT, Braff DL, Turetsky BI, 2015. Validation of mismatch negativity and P3a for use in multi-site studies of schizophrenia: Characterization of demographic, clinical, cognitive, and functional correlates in COGS-2. Schizophrenia Research 163, 63–72. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Lukoff D, Nuechterlein KH, Ventura J, 1986. Manual for the expanded brief psychiatric rating scale. Schizophrenia Bulletin 12, 594–602. [Google Scholar]
- Michie PT, Budd TW, Todd J, Rock D, Wichmann H, Box J, Jablensky AV, 2000. Duration and frequency mismatch negativity in schizophrenia. Clinical Neurophysiology 111, 1054–1065. [DOI] [PubMed] [Google Scholar]
- Molholm S, Martinez A, Ritter W, Javitt DC, Foxe JJ, 2004. The neural circuitry of pre-attentive auditory change-detection: an fMRI study of pitch and duration mismatch negativity generators. Cerebral Cortex 15, 545–551. [DOI] [PubMed] [Google Scholar]
- Näätänen R, 2008. Mismatch negativity (MMN) as an index of central auditory system plasticity. International Journal of Audiology 47, S16–S20. [DOI] [PubMed] [Google Scholar]
- Näätänen R, Kähkönen S, 2009. Central auditory dysfunction in schizophrenia as revealed by the mismatch negativity (MMN) and its magnetic equivalent MMNm: a review. International Journal of Neuropsychopharmacology 12, 125–135. [DOI] [PubMed] [Google Scholar]
- Näätänen R, Paavilainen P, Rinne T, Alho K, 2007. The mismatch negativity (MMN) in basic research of central auditory processing: A review. Clinical Neurophysiology 118, 2544–2590. [DOI] [PubMed] [Google Scholar]
- Nagai T, Tada M, Kirihara K, Yahata N, Hashimoto R, Araki T, Kasai K, 2013. Auditory mismatch negativity and P3a in response to duration and frequency changes in the early stages of psychosis. Schizophrenia Research 150, 547–554. [DOI] [PubMed] [Google Scholar]
- Nazimek JM, Hunter MD, Woodruff PWR, 2012. Auditory hallucinations: Expectation-perception model. Medical Hypotheses 78, 802–810. [DOI] [PubMed] [Google Scholar]
- Nuechterlein K, Green M, 2006. MCCB: matrics consensus cognitive battery. Los Angeles, CA: MATRICS Assessment Inc. [Google Scholar]
- Paavilainen P, Alho K, Reinikainen K, Sams M, Näätänen R, 1991. Right hemisphere dominance of different mismatch negativities. Electroencephalography and Clinical Neurophysiology 78, 466–479. [DOI] [PubMed] [Google Scholar]
- Paavilainen P, Valppu S, Naatanen R, Näätänen R, 2001. The additivity of the auditory feature analysis in the human brain as indexed by the mismatch negativity: 1+1 ≈ 2 but 1+1+1<3. Neuroscience Letters 301, 179–182. [DOI] [PubMed] [Google Scholar]
- Perez VB, Tarasenko M, Miyakoshi M, Pianka ST, Makeig SD, Braff DL, Swerdlow NR, Light GA, 2017. Mismatch Negativity is a Sensitive and Predictive Biomarker of Perceptual Learning During Auditory Cognitive Training in Schizophrenia. Neuropsychopharmacology 42, 2206–2213. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Perez VB, Woods SW, Roach BJ, Ford JM, McGlashan TH, Srihari VH, Mathalon DH, 2014. Automatic auditory processing deficits in schizophrenia and clinical high-risk patients: forecasting psychosis risk with mismatch negativity. Biological Psychiatry 75, 459–469. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Rissling AJ, Park S-H, Young JW, Rissling MB, Sugar CA, Sprock J, Mathias DJ, Pela M, Sharp RF, Braff DL, Light GA, 2013. Demand and modality of directed attention modulate “pre-attentive” sensory processes in schizophrenia patients and nonpsychiatric controls. Schizophrenia Research 146, 326–335. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Schmack K, Schnack A, Priller J, Sterzer P, 2015. Perceptual instability in schizophrenia: Probing predictive coding accounts of delusions with ambiguous stimuli. Schizophrenia Research Cognition 2, 72–77. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Schroger E, 1995. Processing of auditory deviants with changes in one versus two stimulus dimensions. Psychophysiology 32, 55–65. [DOI] [PubMed] [Google Scholar]
- Schultz W, Dickinson A, 2000. Neuronal Coding of Prediction Errors. Annual Review Of Neuroscience 23, 473–500. [DOI] [PubMed] [Google Scholar]
- Shrout PE, Fleiss JL, 1979. Intraclass correlations: uses in assessing rater reliability. Psychological Bulletin 86, 420–428. [DOI] [PubMed] [Google Scholar]
- Stephan KE, Baldeweg T, Friston KJ, 2006. Synaptic plasticity and dysconnection in schizophrenia. Biological Psychiatry 59, 929–939. [DOI] [PubMed] [Google Scholar]
- Stephan KE, Friston KJ, Frith CD, 2009. Dysconnection in Schizophrenia: From abnormal synaptic plasticity to failures of self-monitoring. Schizophrenia Bulletin 35, 509–527. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Takegata R, Paavilainen P, Naatanen R, Winkler I, Näätänen R, Winkler I, 1999. Independent processing of changes in auditory single features and feature conjunctions in humans as indexed by the mismatch negativity. Neuroscience Letters 266, 109–112. [DOI] [PubMed] [Google Scholar]
- Thomas ML, Green MF, Hellemann G, Sugar CA, Tarasenko M, Calkins ME, Greenwood TA, Gur RE, Gur RC, Lazzeroni LC, Nuechterlein KH, Radant AD, Seidman LJ, Shiluk AL, Siever LJ, Silverman JM, Sprock J, Stone WS, Swerdlow NR, Tsuang DW, Tsuang MT, Turetsky BI, Braff DL, Light GA, 2017. Modeling Deficits From Early Auditory Information Processing to Psychosocial Functioning in Schizophrenia. JAMA Psychiatry 74, 37–46. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Todd J, Harms L, Schall U, Michie PT, 2013. Mismatch negativity: Translating the potential. Frontiers in Psychiatry 4, 1–22. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Todd J, Michie PT, Schall U, Karayanidis F, Yabe H, Näätänen R, 2008. Deviant matters: duration, frequency, and intensity deviants reveal different patterns of mismatch negativity reduction in early and late schizophrenia. Biological Psychiatry 63, 58–64. [DOI] [PubMed] [Google Scholar]
- Umbricht D, Krljes S, 2005. Mismatch negativity in schizophrenia: a meta-analysis. Schizophrenia Research 76, 1–23. [DOI] [PubMed] [Google Scholar]
- Wacongne C, 2016. A predictive coding account of MMN reduction in schizophrenia. Biological Psychology 116, 68–74. [DOI] [PubMed] [Google Scholar]
- Wacongne C, Changeux J-P, Dehaene S, 2012. A neuronal model of predictive coding accounting for the mismatch negativity. Journal of Neuroscience 32, 3665–3678. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Winkler I, Cowan N, Csépe V, Czigler I, Näätänen R, 1996. Interactions between Transient and Long-Term Auditory Memory as Reflected by the Mismatch Negativity. Journal of Cognitive Neuroscience 8, 403–415. [DOI] [PubMed] [Google Scholar]
- Winkler I, Czigler I, 2012. Evidence from auditory and visual event-related potential (ERP) studies of deviance detection (MMN and vMMN) linking predictive coding theories and perceptual object representations. Int. Journal of Psychophysiology 83, 132–143. [DOI] [PubMed] [Google Scholar]
- Winkler I, Reinikainen K, Näätänen R, 1993. Event-related brain potentials reflect traces of echoic memory in humans. Perception & Psychophysics 53, 443–449. [DOI] [PubMed] [Google Scholar]
- Wolff C, Schröger E, 2001. Human pre-attentive auditory change-detection with single, double, and triple deviations as revealed by mismatch negativity additivity. Neuroscience Letters 311, 37–40. [DOI] [PubMed] [Google Scholar]
- Wynn JK, Sugar C, Horan WP, Kern R, Green MF, 2010. Mismatch Negativity, Social Cognition, and Functioning in Schizophrenia Patients. Biological Psychiatry 67, 940–947. [DOI] [PMC free article] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.