

Abstract
The mutualistic symbiosis between flowering plants and arbuscular mycorrhizal fungi is extremely abundant in terrestrial ecosystems. In this symbiosis, obligately biotrophic fungi colonize the root of the host plants, which can benefit from these fungi by enhanced access to mineral nutrients in the soil, especially phosphorus. One of the main goals of research on this symbiosis is to find plant genes that control fungal development in the host plant. In this work, we show that mycorrhizal colonization is regulated by enod40, an early nodulin gene known to be involved in the nodule symbiosis of legumes with nitrogen-fixing bacteria. Medicago truncatula plants overexpressing enod40 exhibited stimulated mycorrhizal colonization in comparison with control plants. Overexpression of enod40 promoted fungal growth in the root cortex and increased the frequency of arbuscule formation. Transgenic lines with suppressed levels of enod40 transcripts, likely via a cosuppression phenomenon induced by the transgene, exhibited reduced mycorrhizal colonization. Hence, enod40 might be a plant regulatory gene involved in the control of the mycorrhizal symbiosis.
Most terrestrial flowering plants have the ability to establish symbiotic associations with arbuscular mycorrhizal (AM) fungi. These fungi facilitate the translocation of limiting nutrients, particularly phosphorus, from the soil to the plant root, which in exchange supplies the fungal partner with carbon (1, 2). On contact with the root surface, extraradical hyphae swell and form appressoria. Hyphal penetration of the root occurs via appressoria, and the intraradical hyphae then colonize the root cortex. An appressorium can also be formed without any accompanying infection site. After a few days, the intraradical hyphae penetrate plant cells to form highly branched haustoria, the arbuscules (3). The arbuscules are thought to be the sites where nutrient exchange occurs between the fungus and the host plant. In the late stage of symbiosis, AM fungi develop extraradical hyphae and often vesicles, which are lipid-filled storage structures (1, 2).
One important group of plants that associates with AM fungi is legumes. These plants are especially interesting for research on plant–microbe interactions because they also live in symbiosis with rhizobial bacteria to form nitrogen-fixing root nodules (4, 5). Although these two symbiotic interactions may appear rather different, several similarities have been reported during the colonization process (6, 7). For example, certain flavonoids of the host plant may act as signals promoting the establishment of the fungal as well as the rhizobial symbiosis (8). Moreover, a number of proteins of the host plant are induced in both symbiotic interactions. In Medicago sativa for example, the early nodulin gene enod40 is activated in the nodule primordia (9, 10) and in mycorrhizal roots (11). This gene has numerous short ORFs, and it has been proposed that one of them encodes a small peptide (12). The enod40 transcript is a structured RNA, which does not associate permanently to polyribosomes (10), and it was suggested that it plays a role in gene function (9). enod40 is involved in nodulation, as Medicago truncatula plants constitutively expressing enod40 (13) showed accelerated nodule formation, whereas lines cosuppressing the enod40 transcript exhibited aborted nodule development (14).
Very little is known about plant genes that control mycorrhizal development and arbuscule formation (7), and so far no plant genes have been cloned that regulate the symbiosis with AM fungi. Here, we show that modification of enod40 expression affected mycorrhizal colonization and arbuscule formation in the root cortex. Hence, enod40 might be a regulatory gene of the host plant that controls formation of the symbiosis with AM fungi.
Materials and Methods
Biological Material.
M. truncatula (Gaertner) 108R wild-type plants, the lines Se40(3) and Se40(10) overexpressing a full-length Mtenod40 cDNA from the 35S promoter and the transgenic lines Se40(2) and Se40(5) with reduced levels of enod40 transcripts, have been described previously (13, 14). In this study, the following descendants were used: T3 or T4 of line Se40(3), T2 of line Se40(10), T1 of line Se40(2), and T2 of line Se40(5). Among the T2 descendants of line Se40(5), about one-quarter of 10-day-old seedlings exhibited leaf chlorosis. These plants were excluded from the mycorrhizal assays. Wild-type plants regenerated from leaf explants and plants carrying the pG3.3 vector with a cDNA encoding a green fluorescence protein were used for control experiments. The transgenic nature of all tested plant lines was confirmed by Northern blot analysis (ref. 13 and data not shown).
Inoculum of the mycorrhizal fungus Glomus mosseae (Nicolson and Gerdemann) Gerdemann and Trappe (fungus collection Botanisches Institut der Universität Basel, Switzerland; an inoculum provided by H. Schüepp, Eidgenössische Versuchsanstalt für Obst-, Wein-, und Gartenbau, Wädenswil, Switzerland) was obtained from Zhi-Ping Xie (15) and further propagated by using Tagetes tenuifolia as host plant. Glomus intraradices Smith and Schenk (DAOM 197198; Department of Agriculture, Ottawa) was cultivated under in vitro conditions with Ri T-DNA transformed carrot (Daucus carota L.) roots, kindly provided by Yves Piché (Université Laval, Quebec). Spores were isolated from monoxenic cultures, which were cultivated at 26°C for 8–10 weeks on minimal (M) medium containing 1% gellan gum (Gel-Gro, ICN; ref. 16).
Culture System and Growth Conditions.
M. truncatula seeds were surface sterilized by treatment with diluted commercial bleach (0.35% active chlorine) containing 0.01% SDS at 28°C for 1 h on a rotary shaker at 150 rpm. After thorough washing with sterile H2O, the seeds were incubated overnight in H2O at room temperature. They were then washed again with sterile H2O, and the testa was removed with forceps. Seeds were left to germinate on water agar plates in the dark and transferred to 300-ml magenta bodies after 2 days. The upper vessel was filled with sterile loam and sand in the ratio 1:9 (wt/wt) containing the mycorrhizal inoculum in case of experiments with G. mosseae. The phosphorus content of the substrate (after extraction with 0.1 M lactic acid/0.3 M acetic acid/0.1 M ammonium acetate) was 12 mg/kg dry weight, as determined by the molybdate blue ascorbic acid method (17). The lower magenta vessel was filled with autoclaved phosphate-free Fahraeus nutrient solution, pH 6.3, containing 1 mM CaCl2, 1 mM MgSO4, 2 mM K2SO4, 1 mM KNO3, 1 mM Fe(III)-citrate, and the microelements of the Fahraeus medium (18). For experiments with G. intraradices, plants were inoculated with a spore suspension (800 spores in 5 ml of H2O per magenta vessel) 10 days after transfer of the seedlings to the magenta vessels. Plants were cultivated in a greenhouse (experiments with G. mosseae; 16 h/day period) or in a growth chamber (experiments with G. intraradices; 16-day period, photon flux of 300 μmol⋅m−2⋅s−1 at 22°C; night period at 18°C). When required, the lower magenta vessels were filled up again with autoclaved phosphate-free Fahraeus nutrient solution.
Staining of Roots and Estimation of Mycorrhizal Colonization.
At the time of harvest, the roots were washed with tap water and boiled in 20 ml of 10% (wt/vol) KOH for 5 min, washed with H2O, and placed in 20 ml of 0.3 M HCl for 2 min. Then, the HCl was poured off, and the roots were incubated for at least 12 h in 10 ml of staining solution containing 0,1% (wt/vol) trypan blue (Aldrich), 24% (vol/vol) glycerin, 31% (vol/vol) lactic acid, and 30% (wt/vol) phenol. For the estimation of mycorrhizal colonization, aliquots of the root material were washed with H2O, and the degree of mycorrhizal colonization was determined in blind tests using the gridline intersection method with a minimum of 300 line intersections per root sample (19). Data are given as percentages of root length containing blue-stained mycorrhizal structures within the root. This parameter is independent of total root length. External hyphae growing in the vicinity of the roots and sporocarps were not included into the estimation. For microscopical studies, root pieces were washed with a solution containing 24% (vol/vol) glycerin and 31% (vol/vol) lactic acid. The number of mycorrhizal structures (appressoria, arbuscules, vesicles) between two gridlines was determined, and the number of these structures was calculated by using either the total root length (colonized and noncolonized root zones) or root zones colonized by AM fungi as a basis. The size of a colonization zone in young roots was determined by measuring the root length carrying mycorrhizal structures including intraradical hyphae.
Experimental Design and Statistical Analysis.
The aim of this work was to test whether transcript levels of
enod40 influence mycorrhizal colonization. To test this,
seven different experiments were executed (see Results for
details). In each experiment, different plant lines with altered levels
of enod40 transcripts were compared with wild-type plants.
Per plant line (including the wild-type), five (or more) magenta
vessels were prepared, which thus gave at least five replicates per
treatment. Eleven (or more) replicates per treatment were used for
experiment 6. Where plants were grown together in magenta vessels,
plant material from each vessel was combined for data collection, and
pooled means from each vessel were used for statistical analysis
(S-PLUS 4.5 program from MathSoft, Bagshot, Surrey,
U.K.). Because of nonnormality within treatment and to variance
inequality among treatments in some experiments, data were analyzed by
the nonparametric Kruskal–Wallis rank sum test, which is also suitable
for unequal replication (20). The Kruskal–Wallis statistics is
approximately distributed as a
χ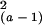 , with a being the
number of treatments that were compared. Pairwise and multiple
comparisons among plant lines were consequently executed to test
whether treatments varied from each other by the Kruskal–Wallis test.
A probability of P < 0.05 was considered as
representing a significant difference in this study. All data given are
means ± SE.
, with a being the
number of treatments that were compared. Pairwise and multiple
comparisons among plant lines were consequently executed to test
whether treatments varied from each other by the Kruskal–Wallis test.
A probability of P < 0.05 was considered as
representing a significant difference in this study. All data given are
means ± SE.
Results
Transgenic M. truncatula Plants Overexpressing enod40 Exhibit Increased Colonization by G. mosseae.
In a first series of experiments, root colonization of M.
truncatula plants by the fungus G. mosseae was
examined. The degree of mycorrhizal colonization was significantly
higher in roots of the enod40-overexpressing line Se40(3) in
comparison with wild-type plants (n = 10,
χ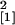 = 6.81, P = 0.009). Plants
regenerated from leaf explants and transgenic plants carrying the pG3.3
vector containing a 35S promoter–GFP construct were
used as additional controls. Mycorrhizal colonization in these plants
was as low as in the wild-type plants, indicating that increased
colonization was correlated with enod40 overexpression and
was independent of the transformation and regeneration procedure
(experiment 1; three plants per magenta vessel; Fig.
1a).
= 6.81, P = 0.009). Plants
regenerated from leaf explants and transgenic plants carrying the pG3.3
vector containing a 35S promoter–GFP construct were
used as additional controls. Mycorrhizal colonization in these plants
was as low as in the wild-type plants, indicating that increased
colonization was correlated with enod40 overexpression and
was independent of the transformation and regeneration procedure
(experiment 1; three plants per magenta vessel; Fig.
1a).
Figure 1.

Transgenic M. truncatula plants overexpressing
Mtenod40 exhibit enhanced colonization by G.
mosseae. (a) Degree of mycorrhizal colonization
(length of roots containing mycorrhizal structures in percentage of
total root length) of M. truncatula wild-type (WT)
plants, WT plants regenerated from leaf explants (Control), plants
expressing a 35S promoter-GFP fusion (GFP), and line
Se40(3) overexpressing the Mtenod40 gene harvested 21
days p.i. (experiment 1; n = 20,
χ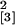 = 12.58, P = 0.006).
(b) Time course of the mycorrhizal colonization of WT
and Se40(3) plants (experiment 2). The degree of mycorrhizal
colonization was estimated at different time points after inoculation.
Differences (indicated by asterisks) were statistically significant
(P < 0.05) up to 38 days. (c and
d) Mycorrhizal colonization by G. mosseae
using low amounts of inoculum. The difference between WT and two
different enod40-overexpressing lines was significant
(P < 0.05) in both experiments. (c)
Degree of mycorrhizal colonization in roots of WT plants and line
Se40(3) harvested 35 days p.i. (experiment 3). (d)
Degree of mycorrhizal colonization in roots of WT plants and line
Se40(10) harvested 49 days p.i. (experiment 4).
= 12.58, P = 0.006).
(b) Time course of the mycorrhizal colonization of WT
and Se40(3) plants (experiment 2). The degree of mycorrhizal
colonization was estimated at different time points after inoculation.
Differences (indicated by asterisks) were statistically significant
(P < 0.05) up to 38 days. (c and
d) Mycorrhizal colonization by G. mosseae
using low amounts of inoculum. The difference between WT and two
different enod40-overexpressing lines was significant
(P < 0.05) in both experiments. (c)
Degree of mycorrhizal colonization in roots of WT plants and line
Se40(3) harvested 35 days p.i. (experiment 3). (d)
Degree of mycorrhizal colonization in roots of WT plants and line
Se40(10) harvested 49 days p.i. (experiment 4).
To test fungal colonization during the development of the root system
in wild-type and enod40-overexpressing plants, mycorrhizal
plants were harvested at various time points p.i. (experiment 2; three
plants per magenta vessel). The root system of M. truncatula
plants was progressively colonized by G. mosseae (Fig.
1b). Compared with wild-type plants, the
enod40-overexpressing line Se40(3) exhibited an accelerated
mycorrhizal colonization, and a difference was already seen 12 days
p.i. In this early stage of symbiosis, fungal colonization of line
Se40(3) was nearly 2-fold higher in comparison with wild-type plants.
At later stages, the degree of fungal colonization of line Se40(3) was
about 10% higher than that of wild-type plants. This increase
corresponded to an acceleration of mycorrhizal colonization of about 1
week. The difference in mycorrhizal colonization was statistically
different between the two lines at each of the first four time points
(12 days p.i., n = 14, χ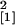 =
9.24, P = 0.002; 18 days p.i., n = 10,
χ
=
9.24, P = 0.002; 18 days p.i., n = 10,
χ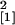 = 4.81, P = 0.028; 28 days
p.i., n = 10, χ
= 4.81, P = 0.028; 28 days
p.i., n = 10, χ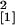 = 3.94,
P = 0.047; 36 days p.i., n = 10,
χ
= 3.94,
P = 0.047; 36 days p.i., n = 10,
χ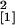 = 5.77, P = 0.016) (Fig.
1b).
= 5.77, P = 0.016) (Fig.
1b).
The quantity of fungal inoculum is a parameter that influences
mycorrhizal root colonization. To test the effect of enod40
overexpression on M. truncatula plants with a low degree of
mycorrhizal colonization, plants were infected with another inoculum of
G. mosseae, which was added to the growth medium at 50-fold
lower amounts compared with the other experiments. As expected, fungal
colonization was delayed, and the rate of root colonization after 35
days was still low. The difference between the degree of fungal
colonization of the line Se40(3) and the wild-type plants was also seen
under these conditions (experiment 3; four plants per magenta vessel;
n = 10, χ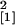 = 6.32,
P = 0.012) (Fig. 1c). A similar experiment
was performed with line Se40(10), another
enod40-overexpressing M. truncatula line, and the
root material was harvested 49 days p.i. (experiment 4; four plants per
magenta vessel). Similarly to line Se40(3), roots from line Se40(10)
exhibited enhanced mycorrhizal colonization in comparison with the
wild-type plants (n = 10, χ
= 6.32,
P = 0.012) (Fig. 1c). A similar experiment
was performed with line Se40(10), another
enod40-overexpressing M. truncatula line, and the
root material was harvested 49 days p.i. (experiment 4; four plants per
magenta vessel). Similarly to line Se40(3), roots from line Se40(10)
exhibited enhanced mycorrhizal colonization in comparison with the
wild-type plants (n = 10, χ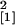 =
5.77, P = 0.016) (Fig. 1d). These data
indicate that enod40 overexpression promotes mycorrhizal
colonization in two independent enod40-overexpressing
M. truncatula lines.
=
5.77, P = 0.016) (Fig. 1d). These data
indicate that enod40 overexpression promotes mycorrhizal
colonization in two independent enod40-overexpressing
M. truncatula lines.
Overexpression of enod40 Stimulates Fungal Growth in Planta.
To determine whether enod40 overexpression affects the
initial stages of mycorrhizal colonization, comparative microscopical
studies on enod40-overexpressing and wild-type plants from
experiment 2 were performed. As shown in Fig. 1b,
mycorrhizal colonization by G. mosseae was observed as early
as 12 days p.i. At this stage, total root length of both plants was
about 30 cm, and the first small fungal colonization zones were seen.
The average number of such separated colonization zones per plant was
similar in wild-type and in the enod40-overexpressing line
Se40(3) (Table 1). The average size of a
colonization zone, however, as determined by its length, was
significantly increased in the Se40(3) line (n = 14,
χ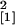 = 5.40, P = 0.020).
Consequently, the total root length colonized by G. mosseae
was higher in the Se40(3) line compared with the wild-type plants
(n = 14, χ
= 5.40, P = 0.020).
Consequently, the total root length colonized by G. mosseae
was higher in the Se40(3) line compared with the wild-type plants
(n = 14, χ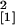 = 4.27,
P = 0.039). The number of appressoria (per cm total
root length) was similar for wild-type and
enod40-overexpressing plants. The number of appressoria per
mm colonized root zone, however, was lower in the
enod40-overexpressing line Se40(3) in comparison with the
wild-type (n = 14, χ
= 4.27,
P = 0.039). The number of appressoria (per cm total
root length) was similar for wild-type and
enod40-overexpressing plants. The number of appressoria per
mm colonized root zone, however, was lower in the
enod40-overexpressing line Se40(3) in comparison with the
wild-type (n = 14, χ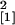 = 4.54,
P = 0.033). Taken the data of Table 1 together, they
show that overexpression of enod40 promotes fungal growth
in planta.
= 4.54,
P = 0.033). Taken the data of Table 1 together, they
show that overexpression of enod40 promotes fungal growth
in planta.
Table 1.
Mycorrhizal colonization zones and frequency of appressoria formation in roots of M. truncatula wild-type plants and line Se40(3) overexpressing the Mtenod40 gene
| Wild type | Se40(3) | |
|---|---|---|
| Number of independent colonization zones (per plant) | 4.5 ± 0.7 a | 4.4 ± 0.3 a |
| Average length of a colonization zone, mm | 4.1 ± 0.6 a | 6.3 ± 0.7 b |
| Total length, in mm, of the colonization zones (per plant) | 18.2 ± 2.9 a | 28.0 ± 2.0 b |
| Number of appressoria per cm (total root length) | 0.36 ± 0.06 a | 0.38 ± 0.05 a |
| Number of appressoria per mm (in colonized root zones) | 0.60 ± 0.07 a | 0.37 ± 0.06 b |
Plants harvested 12 days p.i. with G. mosseae were analyzed (experiment 2). Values in lines followed by different letters are significantly different (P < 0.05).
Overexpression of enod40 Promotes Development of Arbuscules.
To investigate whether enod40 overexpression affects
formation of arbuscules, the number of arbuscules was counted on roots
harvested 12 and 18 days p.i. (experiment 2). Compared with wild-type
plants, enod40-overexpressing plants exhibited a
significantly higher number of arbuscules, when calculated per mm total
root length (12 days p.i., n = 14,
χ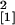 = 9.60, P = 0.002; 18 days
p.i., n = 10, χ
= 9.60, P = 0.002; 18 days
p.i., n = 10, χ = 6.32,
P = 0.012) (Fig.
2a). We also determined the
number of arbuscules per mm colonized root (Fig. 2b). In
roots harvested 12 days p.i., most arbuscules were formed in the inner
cortical cell layers that were adjacent to the endodermis. The
frequency of arbuscule formation in colonized root zones of
enod40-overexpressing plants was significantly higher in
comparison with the wild-type plants (n = 14,
χ
= 6.32,
P = 0.012) (Fig.
2a). We also determined the
number of arbuscules per mm colonized root (Fig. 2b). In
roots harvested 12 days p.i., most arbuscules were formed in the inner
cortical cell layers that were adjacent to the endodermis. The
frequency of arbuscule formation in colonized root zones of
enod40-overexpressing plants was significantly higher in
comparison with the wild-type plants (n = 14,
χ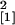 = 6.01, P = 0.014),
indicating that increased formation of arbuscules in
enod40-overexpressing plants was not simply because of an
increase in the overall length of root colonized by G.
mosseae. For roots harvested 18 days p.i., this difference was no
longer significant (Fig. 2b). Taken together, these data
show that enod40 overexpression resulted in enhanced
formation of arbuscules in young roots.
= 6.01, P = 0.014),
indicating that increased formation of arbuscules in
enod40-overexpressing plants was not simply because of an
increase in the overall length of root colonized by G.
mosseae. For roots harvested 18 days p.i., this difference was no
longer significant (Fig. 2b). Taken together, these data
show that enod40 overexpression resulted in enhanced
formation of arbuscules in young roots.
Figure 2.

Number of arbuscules in wild-type (WT) plants and line Se40(3) overexpressing enod40. Plants were inoculated with G. mosseae and harvested 12 and 18 days later (experiment 2). Asterisks indicate statistically significant differences (P < 0.05) between WT and line Se40(3). (a) Number of arbuscules (per mm root length) on the basis of the total root length (colonized and noncolonized root zones). (b) Number of arbuscules (per mm root length) in root zones colonized by G. mosseae.
We also compared the number of vesicles in
enod40-overexpressing and wild-type roots. Only few vesicles
were seen in roots harvested 12 days p.i. Because of the increased
fungal colonization rate in enod40-overexpressing plants,
the number of vesicles per mm total root length was higher in
comparison with the wild-type plants (n = 14,
χ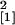 = 4.82, P = 0.028). In roots
harvested 18 days p.i., no significant differences were seen
(Fig. 3a). In contrast to the
formation of arbuscules (Fig. 2b), enod40
overexpression did not affect the frequency of vesicle formation in
colonized root zones (Fig. 3b).
= 4.82, P = 0.028). In roots
harvested 18 days p.i., no significant differences were seen
(Fig. 3a). In contrast to the
formation of arbuscules (Fig. 2b), enod40
overexpression did not affect the frequency of vesicle formation in
colonized root zones (Fig. 3b).
Figure 3.

Number of vesicles in wild-type (WT) plants and line Se40(3) overexpressing enod40. Plants were harvested 12 and 18 days after inoculation with G. mosseae (experiment 2). (a) Number of vesicles (per mm root length) on the basis of the total root length (colonized and noncolonized root zones). Asterisks indicate statistically significant differences (P < 0.05) between WT and line Se40(3) for root material harvested 12 days p.i. (b) Number of vesicles (per mm root length) in root zones colonized by G. mosseae.
M. truncatula Plants with Suppressed Levels of enod40 Transcripts Exhibit Reduced Colonization by G. intraradices.
We also tested mycorrhizal colonization of M. truncatula by
another fungus. Plants were inoculated with G. intraradices
spores and cultivated in a growth chamber. In this system, spore
germination was required and fungal colonization of young roots was
lower than in the previous experiments with G. mosseae.
Similarly to the association with G. mosseae, mycorrhizal
colonization by G. intraradices was stimulated in the
enod40-overexpressing line Se40(3) (n = 10,
χ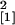 = 3.93, P = 0.047) or in the
line Se40(10) (n = 11, χ
= 3.93, P = 0.047) or in the
line Se40(10) (n = 11, χ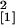 =
4.80, P = 0.029) when compared with wild-type plants.
Transgenic control plants carrying the pG3.3 vector with a 35S
promoter-GFP did not exhibit enhanced fungal colonization
(experiment 5; three plants per magenta vessel; Fig.
4a). In another experiment, we
tested the transgenic M. truncatula lines Se40(2) and
Se40(5), which show reduced enod40 transcript accumulation
(13). This reduction in transcript levels is probably as a result of
cosuppression induced by the transgene (21). Compared with wild-type
plants, mycorrhizal colonization of line Se40(2) (n =
36, χ
=
4.80, P = 0.029) when compared with wild-type plants.
Transgenic control plants carrying the pG3.3 vector with a 35S
promoter-GFP did not exhibit enhanced fungal colonization
(experiment 5; three plants per magenta vessel; Fig.
4a). In another experiment, we
tested the transgenic M. truncatula lines Se40(2) and
Se40(5), which show reduced enod40 transcript accumulation
(13). This reduction in transcript levels is probably as a result of
cosuppression induced by the transgene (21). Compared with wild-type
plants, mycorrhizal colonization of line Se40(2) (n =
36, χ = 4.85, P = 0.027) and
line Se40(5) (n = 28, χ
= 4.85, P = 0.027) and
line Se40(5) (n = 28, χ = 6.34,
P = 0.012) was significantly lower, whereas the
enod40-overexpressing Se40(3) line exhibited stimulated
fungal colonization (n = 32, χ
= 6.34,
P = 0.012) was significantly lower, whereas the
enod40-overexpressing Se40(3) line exhibited stimulated
fungal colonization (n = 32, χ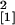 = 7.60, P = 0.006) (experiment 6; one plant per magenta
vessel) (Fig. 4b). These results indicate that the amount of
enod40 transcript accumulation alters the degree of
mycorrhizal colonization.
= 7.60, P = 0.006) (experiment 6; one plant per magenta
vessel) (Fig. 4b). These results indicate that the amount of
enod40 transcript accumulation alters the degree of
mycorrhizal colonization.
Figure 4.

Mycorrhizal colonization by G. intraradices.
(a) Degree of mycorrhizal colonization of M.
truncatula wild-type (WT) plants, plants expressing a 35S
promoter-GFP fusion (GFP) and overexpressing the
Mtenod40 gene [lines Se40(3) and Se40(10)] harvested
56 days p.i. (experiment 5; n = 22,
χ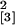 = 12.39, P = 0.006).
(b) Degree of mycorrhizal colonization of M.
truncatula wild-type (WT) plants, two lines with a
cosuppression phenomenon [lines Se40(2) and Se40(5)] and line Se40(3)
overexpressing enod40 harvested 49 days p.i.
(experiment 6; n = 62,
χ
= 12.39, P = 0.006).
(b) Degree of mycorrhizal colonization of M.
truncatula wild-type (WT) plants, two lines with a
cosuppression phenomenon [lines Se40(2) and Se40(5)] and line Se40(3)
overexpressing enod40 harvested 49 days p.i.
(experiment 6; n = 62,
χ = 27.17, P < 0.001).
= 27.17, P < 0.001).
Symbiotic Efficiency.
Mycorrhizal colonization of legumes can promote plant growth responses, indicating a benefit of the host plant from the symbiosis (22, 23). To examine the mycorrhizal responsiveness of M. truncatula under our growth conditions, noninoculated plants and mycorrhizal plants colonized by G. mosseae were harvested 42 days p.i., and their dry weight was determined (experiment 7). Independently of the inoculation with the fungus, wild-type plants and the enod40-overexpressing Se40(3) line showed similar values for total dry weight per plant [noninoculated wild type, 32.4 ± 5.1 mg; inoculated wild type, 32.3 ± 4.0 mg; noninoculated line Se40(3), 36.7 ± 7.6 mg; inoculated line Se40(3), 41.1 ± 8.7 mg]. Statistical analysis revealed no significant differences, indicating that the amount of produced plant biomass was independent of mycorrhizal colonization under our test conditions. No significant differences were also seen when dry weight values for roots and shoots were compared (data not shown). As expected, mycorrhizal plants exhibited a ≈2-fold and significant increase in their phosphorus content in comparison with noninoculated control plants, but no significant difference was found between line Se40(3) and wild-type plants (data not shown). These data indicate that enod40 transcript levels in M. truncatula plants did not affect symbiotic efficiency, even though alteration of enod40 expression significantly influenced the degree of root colonization by AM fungi.
Discussion
This paper shows that alteration in the transcript level of the enod40 gene has an effect on the growth of mycorrhizal fungi in the root and on the development of cells containing arbuscules. Effects of enod40 transcript levels on nodule formation of M. truncatula have been reported (14). These findings indicate that enod40 may have a general role in mutualistic endosymbioses. This is in line with the genetic evidence, obtained from symbiosis-defective mutants, that certain sym genes of the host plant control both rhizobial and mycorrhizal symbioses (6, 7).
In mycorrhizal M. sativa roots, in situ hybridization studies showed accumulation of enod40 transcripts in the pericycle, in the epidermis, as well as in young arbuscules (11). In general, enod40 transcripts were found in cells that transport carbohydrates to strong sinks or become those sinks, such as the nodule and lateral root primordium or the peripheral cells of vascular bundles (9, 10, 24, 25). Indeed, in developing rice leaves, enod40 expression changed during the source/sink transition (26). One possibility is that enod40 transcripts in mycorrhizal roots are involved in the control of carbohydrate supply or partitioning into specific cells. Fungal growth in the root and formation of arbuscules are two closely linked processes that depend on carbon availability. Increased carbohydrate supply to the fungus could be a reason why M. truncatula plants overexpressing enod40 exhibited accelerated mycorrhizal colonization. Alternatively, enod40 transcripts might stimulate fungal growth in the root cortex by affecting targets that interfere with plant hormone action and unknown cell-to-cell communication processes between vascular and cortical tissues. It is worth noting in this context that the amount of enod40 transcripts in M. sativa can be enhanced by treatment of roots with cytokinins and rhizobial Nod factors as well (9, 11, 27). It is possible therefore that enod40 in mycorrhizal roots is activated via hormonal imbalances. We suggest that the reported promoting effects of purified Nod factors (15) and auxin transport inhibitors (28) on mycorrhizal colonization are connected to activation and action of enod40.
During rhizobial infection, a transient increase in the root length of enod40-overexpressing plants was observed (14). In the experiments of this study, we did not find any significant difference in root length of plants colonized by AM fungi. It should be considered that large root systems developed under our culture conditions for mycorrhizae. Initial AM colonization 12 days p.i. was detected on 30-cm-long root systems, where slight differences in root length could not be easily measured. Under our test conditions, M. truncatula plants also did not show increased biomass acquisition compared with noncolonized plants, indicating a neutral mycorrhizal dependency of this species. Future studies on plant species with high mycorrhizal dependency are required to find a possible benefit from enhanced enod40 transcript levels on plant growth and crop yield.
Future studies will also focus on the question whether enod40 per se is an essential element for mycorrhizal colonization. An approach would be to identify enod40 genes in various host and nonhost plants of AM fungi. Interestingly, enod40 genes are not restricted to legumes. Homologs of enod40 are known from tobacco (12) and rice (26), host plants of AM fungi, but were not found in the genome of the nonhost Arabidopsis thaliana. A search for enod40 homologs from various nonlegumes could result in a correlation between the finding of enod40 sequences and host plants. Further efforts are also required to study mycorrhizal colonization in enod40-transformed nonlegumes, including nonhost plants. Our recent observations indicate, however, that expression of Mtenod40 in the nonhost A. thaliana does not promote growth of G. intraradices in the rhizosphere.
Taken together, this study shows that transcript levels of a plant gene affect root colonization by AM fungi and formation of arbuscules. This increases our knowledge in understanding the molecular mechanisms and how plants regulate the symbiotic interaction with mycorrhizal fungi.
Acknowledgments
We express our gratitude to Yves Piché (Centre de Recherche en Biologie Forestière, Université Laval, Quebec) for the monoxenic G. intraradices/carrot culture. We thank Eva Kondorosi (Centre National de la Recherche Scientifique, Institut des Sciences du Végétal, Gif-sur-Yvette, France), Andres Wiemken (Botanisches Institut der Universität Basel, Switzerland), and William J. Broughton (Université de Genève, Switzerland) for helpful discussions. We are indebted to Yamama Naciri-Graven (Conservatoire et Jardin Botaniques de Genève, Switzerland) for help in statistical analysis. We are grateful to Ann M. Hirsch (University of California, Los Angeles) and Marcel G. A. van der Heijden (Free Universiteit, Amsterdam) for helpful comments on the manuscript. This work was supported in part by a grant from the Novartis Foundation Basel (to C.S.), the Max-Planck-Gesellschaft research award (to A.K.), and the Ministère Français de l'Enseignement Supérieur et de la Recherche (to C.C.).
Abbreviation
- AM
arbuscular mycorrhizal
References
- 1.Harrison M J. Annu Rev Plant Physiol Plant Mol Biol. 1999;50:361–389. doi: 10.1146/annurev.arplant.50.1.361. [DOI] [PubMed] [Google Scholar]
- 2.Smith S E, Read D J. Mycorrhizal Symbioses. 2nd Ed. London: Academic; 1997. [Google Scholar]
- 3.Blee K A, Anderson A J. Plant J. 1998;16:523–530. [Google Scholar]
- 4.Schultze M, Kondorosi A. Annu Rev Genet. 1998;32:33–57. doi: 10.1146/annurev.genet.32.1.33. [DOI] [PubMed] [Google Scholar]
- 5.Perret X, Staehelin C, Broughton W J. Microbiol Mol Biol Rev. 2000;64:180–201. doi: 10.1128/mmbr.64.1.180-201.2000. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 6.Hirsch A M, Kapulnik Y. Fungal Genet Biol. 1998;23:205–212. doi: 10.1006/fgbi.1998.1046. [DOI] [PubMed] [Google Scholar]
- 7.Marsh S F, Schultze M. New Phytol. 2001;150:525–532. [Google Scholar]
- 8.Vierheilig H, Bago B, Albrecht C, Poulin M J, Piché Y. In: Flavonoids in the Living System. Manthey J, Buslig B, editors. New York: Plenum; 1998. pp. 9–33. [Google Scholar]
- 9.Crespi M D, Jurkevitch E, Poiret M, D'Aubenton-Carafa Y, Petrovics G, Kondorosi E, Kondorosi A. EMBO J. 1994;13:5099–5112. doi: 10.1002/j.1460-2075.1994.tb06839.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 10.Asad S, Fang Y, Wycoff K L, Hirsch A M. Protoplasma. 1994;183:10–23. [Google Scholar]
- 11.Van Rhijn P, Fang Y, Galili S, Shaul O, Atzmon N, Wininger S, Eshed Y, Lum M, Li Y, To V, et al. Proc Natl Acad Sci USA. 1997;94:5467–5472. doi: 10.1073/pnas.94.10.5467. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 12.Van de Sande K, Pawlowski K, Czaja I, Wieneke U, Schell J, Schmidt J, Walden R, Matvienko M, Wellink J, Van Kammen A, et al. Science. 1996;273:370–373. doi: 10.1126/science.273.5273.370. [DOI] [PubMed] [Google Scholar]
- 13.Charon C, Johansson C, Kondorosi E, Kondorosi A, Crespi M. Proc Natl Acad Sci USA. 1997;94:8901–8906. doi: 10.1073/pnas.94.16.8901. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 14.Charon C, Sousa C, Crespi M, Kondorosi A. Plant Cell. 1999;11:1953–1965. doi: 10.1105/tpc.11.10.1953. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 15.Xie Z-P, Staehelin C, Vierheilig H, Wiemken A, Jabbouri S, Broughton W J, Vögeli-Lange R, Boller T. Plant Physiol. 1995;108:1519–1525. doi: 10.1104/pp.108.4.1519. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 16.Bécard G, Fortin J A. New Phytol. 1988;108:211–218. doi: 10.1111/j.1469-8137.1988.tb03698.x. [DOI] [PubMed] [Google Scholar]
- 17.Watanabe F S, Olsen S R. Soil Sci Soc Proc. 1965;29:677–678. [Google Scholar]
- 18.Fahraeus G. J Gen Microbiol. 1957;16:374–381. doi: 10.1099/00221287-16-2-374. [DOI] [PubMed] [Google Scholar]
- 19.Giovannetti M, Mosse B. New Phytol. 1980;84:489–500. [Google Scholar]
- 20.Sokal R R, Rohlf F J. Biometry: The Principles and Practice of Statistics in Biological Research. 3rd Ed. New York: Freeman; 1995. [Google Scholar]
- 21.Vaucheret H, Béclin C, Elmayan T, Feuerbach F, Godon C, Morel J-B, Mourrain P, Palauqui J-C, Vernhettes S. Plant J. 1998;16:651–659. doi: 10.1046/j.1365-313x.1998.00337.x. [DOI] [PubMed] [Google Scholar]
- 22.van der Heijden M G A, Klironomos J N, Ursic M, Moutoglis P, Streitwolf-Engel R, Boller T, Wiemken A, Sanders I R. Nature (London) 1998;396:69–72. [Google Scholar]
- 23.Wilson G W T, Hartnett D C. Am J Bot. 1998;85:1732–1738. [PubMed] [Google Scholar]
- 24.Yang W C, Katinakis P, Hendriks P, Smolders A, de Vries F, Spee J, van Kammen A, Bisseling T. Plant J. 1993;3:573–585. doi: 10.1046/j.1365-313x.1993.03040573.x. [DOI] [PubMed] [Google Scholar]
- 25.Minami E, Kouchi H, Cohn J R, Ogawa T, Stacey G. Plant J. 1996;10:23–32. doi: 10.1046/j.1365-313x.1996.10010023.x. [DOI] [PubMed] [Google Scholar]
- 26.Kouchi H, Takane K-I, So R B, Ladha J K, Reddy P M. Plant J. 1999;18:121–129. doi: 10.1046/j.1365-313x.1999.00432.x. [DOI] [PubMed] [Google Scholar]
- 27.Fang Y W, Hirsch A M. Plant Physiol. 1998;116:53–68. doi: 10.1104/pp.116.1.53. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 28.Xie Z-P, Müller J, Wiemken A, Broughton W J, Boller T. New Phytol. 1998;139:361–366. [Google Scholar]