

Abstract
The interplay between abscisic acid (ABA) and salicylic acid (SA) influences plant responses to various (a)biotic stresses; however, the underlying mechanism for this crosstalk is largely unknown. Here, we report that type 2C protein phosphatases (PP2Cs), some of which are negative regulators of ABA signaling, bind SA. SA binding suppressed the ABA‐enhanced interaction between these PP2Cs and various ABA receptors belonging to the PYR/PYL/RCAR protein family. Additionally, SA suppressed ABA‐enhanced degradation of PP2Cs and ABA‐induced stabilization of SnRK2s. Supporting SA's role as a negative regulator of ABA signaling, exogenous SA suppressed ABA‐induced gene expression, whereas the SA‐deficient sid2‐1 mutant displayed heightened PP2C degradation and hypersensitivity to ABA‐induced suppression of seed germination. Together, these results suggest a new molecular mechanism through which SA antagonizes ABA signaling. A better understanding of the crosstalk between these hormones is important for improving the sustainability of agriculture in the face of climate change.
Keywords: ABA–SA crosstalk, abiotic stress, abscisic acid, biotic stress, salicylic acid, type 2C protein phosphatases
1. INTRODUCTION
Elaborate hormone signaling networks allow plants to perceive and respond adaptively to various biotic and abiotic stresses (Raghavendra, Gonugunta, Christmann, & Grill, 2010; Tuteja, 2007). One of the vital hormones that plays a central role in the adaptation to abiotic stresses, particularly drought and salt stresses, is ABA. In addition, ABA is involved in regulating plant growth and developmental processes under nonstress conditions (Raghavendra et al., 2010) and modulating defense responses following pathogen attack (Denance, Sanchez‐Vallet, Goffner, & Molina, 2013; Robert‐Seilaniantz, Grant, & Jones, 2011). Because of its essential role in multiple physiological processes both under stressed and nonstress conditions, the ABA signaling pathway has been studied intensively during the last two decades. Initial attempts to identify ABA receptors were met with controversy and frustration. Several proteins were proposed to be ABA receptors, but their exact role in ABA response and their associated mechanisms were never established (Hauser, Waadt, & Schroeder, 2011).
The discovery that members of the pyrabactin resistance 1/PYR1‐like regulatory component of ABA receptor (PYR/PYL/RCAR) protein family are ABA receptors, and that they interact with members of the type 2C protein phosphatase (PP2C) protein subfamily, was a major breakthrough in dissecting the ABA signaling pathway (Fujii et al., 2009; Ma et al., 2009; Miyazono et al., 2009; Park et al., 2009; Santiago et al., 2009; Soon et al., 2012). In the absence of ABA, PP2Cs are able to bind and dephosphorylate members of the sucrose nonfermenting 1‐related subfamily 2 protein kinase (SnRK2) family. This negatively regulates ABA signaling because autophosphorylation is required for SnRK2 kinase activity, and thus their ability to transduce the ABA signal by phosphorylating downstream targets. In the presence of ABA, the ABA–receptor complex tightly binds to PP2Cs, thereby preventing PP2C‐mediated dephosphorylation of SnRK2. This, in turn, allows activated SnRK2s to relay the ABA signal.
The reversible phosphorylation of proteins by protein kinases and phosphatases is an important mechanism for regulating many biological processes. In contrast to eukaryotic protein kinases, whose primary and three‐dimensional structures are very similar, protein phosphatases are diverse. Depending on their substrate specificity, protein phosphatases can be divided into two classes, serine/threonine (Ser/Thr) or tyrosine phosphatases (Fuchs, Grill, Meskiene, & Schweighofer, 2013; Schweighofer, Hirt, & Meskiene, 2004; Singh, Pandey, Srivastava, Tran, & Pandey, 2015). The Ser/Thr phosphatases have been further organized into the phosphoprotein phosphatase (PPP) and metal‐dependent protein phosphatase (PPM) families. In plants, PP2Cs, which belong to the PPM family, represent a major portion of the phosphatase‐encoding gene family. To date, 80 or more genes have been identified in the Arabidopsis, tomato, rice, and hot pepper genomes. Phylogenetic analyses have further divided the PP2C families from these plant species into 10 or more subclades designated alphabetically from A onward (Fuchs et al., 2013; Singh et al., 2015).
Of the PP2C subclades, members of “clade A” have been studied the most extensively, as they negatively regulate ABA signaling in various plant species. In Arabidopsis, clade A proteins such as ABA‐insensitive 1 (ABI1), ABI2, hypersensitive to ABA 1 (HAB1), and PP2CA/AHG3 have been shown to mediate ABA‐induced responses to abiotic and biotic stresses via their interaction with SnRK2s and PYR/PYL/RCARs (Fujii et al., 2009; Lim, Luan, & Lee, 2014; Santiago et al., 2012; Soon et al., 2012; de Torres‐Zabala et al., 2007). Functional studies of PP2C proteins from other clades are limited, but they suggest that some of these proteins are involved in responding to (a)biotic stresses. For instance, the clade B member AP2C1 (Arabidopsis phosphatase 2C1) and its ortholog MP2C from Medicago sativa regulate the activity of stress‐induced mitogen‐activated protein kinases (MAPKs (Meskiene et al., 2003; Schweighofer et al., 2004), and the clade F member PIA1 (PP2C induced by AvrRpm1) regulates immune responses in Arabidopsis (Widjaja et al., 2010). By contrast, clades C and D contain PP2Cs that regulate developmental processes (Fuchs et al., 2013; Schweighofer et al., 2004; Singh et al., 2015). Members of clade C, including POL (Poltergeist) and PLL (POL‐like), control shoot and root meristem formation and embryo formation (Song & Clark, 2005), whereas members of clade D negatively regulate the activity of plasma membrane H+‐ATPases and thus cell expansion in the absence of auxin (Spartz et al., 2014).
Salicylic acid is another important plant hormone involved in diverse physiological and metabolic processes, including plant responses to various abiotic stresses. In addition, SA is an essential regulator of plant immune responses (Klessig, Tian, & Choi, 2016; Manohar et al., 2015; Vlot, Dempsey, & Klessig, 2009). While several recent studies have identified components of SA signaling networks and revealed some SA‐mediated signaling mechanisms, a full picture of SA‐based signaling in plants is far from complete. Indeed, the identity of the SA receptor(s) remains unclear. It was recently proposed that nonexpresser of PR1 (NPR1), which functions as a master regulator of SA‐mediated immune signaling, is an SA receptor (Wu et al., 2012). In contrast, Fu et al. (2012) suggested that NPR1's two homologs, NPR3 and NPR4, rather than NPR1, are SA receptors. As NPR3 and NPR4 are adaptors for Cullin 3 ubiquitin E3 ligase, they may regulate the SA signaling pathway by fine‐tuning NPR1 protein levels via degradation (Fu et al., 2012). In addition, nearly 30 SA‐binding proteins (SABPs) have been identified (Klessig et al., 2016). These proteins exhibit a wide range of affinities for SA, and SA binding alters their activities. Given that SA levels vary dramatically within a plant depending on the subcellular compartment, tissue type, developmental stage, and external cues, such as infection, these findings raise the possibility that SA exerts its effects by interacting with multiple targets, rather than a small number of receptors.
Although SA's role in activating disease resistance and ABA's role in signaling abiotic stress responses are well known, it is only recently becoming apparent that ABA also influences immune responses (Denance et al., 2013; Robert‐Seilaniantz et al., 2011). ABA treatment suppressed defense responses and enhanced plant susceptibility to certain bacterial and fungal pathogens (De Torres Zabala, Bennett, Truman, & Grant, 2009; McDonald & Cahill, 1999; Mohr & Cahill, 2003; Robert‐Seilaniantz et al., 2011; Thaler & Bostock, 2004; Ward, Cahill, & Bhattacharyya, 1989). Additionally, the virulence of Pseudomonas syringae in Arabidopsis was dependent on manipulation of the ABA signaling pathway by secreted bacterial effectors (de Torres‐Zabala et al., 2007). Growing evidence also indicates that there is substantial crosstalk between the ABA and SA pathways during immune signaling (de Torres‐Zabala et al., 2007; Yasuda et al., 2008). Arabidopsis mutants deficient in ABA synthesis or response not only exhibited reduced susceptibility to pathogen infection, but also showed enhanced expression of SA‐responsive genes, such as pathogenesis‐related protein‐1 (PR‐1) and PR‐4 (Audenaert, De Meyer, & Höfte, 2002; Sanchez‐Vallet et al., 2012; Thaler & Bostock, 2004). Conversely, Arabidopsis overexpressing RCAR3, which confers increased ABA sensitivity, displayed enhanced susceptibility to P. syringae DC3000 infection, which correlated with decreased expression of PR‐1 and NPR1 (Lim et al., 2014). Further demonstrating the antagonistic interaction between ABA and SA, exogenous ABA suppressed the ability of an SA functional analog to enhance pathogen resistance in Arabidopsis, while pretreatment with this analog suppressed NaCl‐induced expression of several ABA biosynthetic or ABA‐responsive genes (Yasuda et al., 2008). ABA appears to suppress immune responses by down‐regulating SA biosynthesis (de Torres‐Zabala et al., 2007; Yasuda et al., 2008); however, the mechanism through which SA inhibits ABA signaling is unknown.
In previous studies, we have identified several SABPs that are involved in various biotic and abiotic stress responses (Manohar et al., 2015; Tian et al., 2012). Here, we identify PP2Cs from clades A and D as novel SABPs and show that SA binding to these PP2Cs suppresses their ABA‐enhanced interaction with the ABA receptors. In addition, SA suppresses ABA‐induced degradation of PP2Cs and suppresses ABA‐mediated stabilization of SnRK2s. Combined with the demonstration that SA treatment antagonizes ABA‐induced gene expression and the SA‐deficient sid2‐1 mutant is ABA hypersensitive, these results suggest that SA antagonizes ABA signaling through multiple mechanisms.
2. RESULTS
2.1. Identification of PP2Cs as novel SA‐binding proteins
To help define the SA signaling network in plants, we developed several high‐throughput screens capable of identifying SABPs on a genome‐wide scale (Choi et al., 2015; Manohar et al., 2015; Tian et al., 2012). In one screen, protein extracts prepared from Arabidopsis leaves were subjected to affinity chromatography on a Pharma‐link column to which SA was attached. After stringent washing with the biologically inactive SA analog 4‐hydroxy benzoic acid (4‐HBA), SA‐bound proteins on the column were eluted with excess SA. The eluted proteins were analyzed by mass spectroscopy and a PP2C belonging to clade D (PP2C‐D4; At3g55050) was identified along with other putative SABPs. Recombinant histidine‐tagged PP2C‐D4 was produced in Escherichia coli and the purified protein was further assessed for SA‐binding activity using three different assays, namely surface plasmon resonance (SPR), photoaffinity crosslinking, and size‐exclusion chromatography. SPR analysis was performed with a CM5 sensor chip to which the SA derivative 3‐aminoethyl SA (3AESA) was immobilized via an amide bond. Binding to 3AESA was detected when purified PP2C‐D4 was passed over the sensor chip (Figure 1a). In the presence of increasing concentrations of SA, PP2C‐D4 binding to the 3AESA‐immobilized sensor chip was modestly reduced (Figure 1a). Similar to these results, the photoaffinity labeling approach indicated that PP2C‐D4 bound and was crosslinked to 4‐azido SA (4AzSA). This binding also was suppressed by SA in a dose‐dependent manner (Figure 1b), arguing that PP2C‐D4 binding to both 3AESA and 4AzSA represents authentic SA‐binding activity. PP2C‐D4's ability to bind SA also was confirmed by size‐exclusion chromatography using [3H]SA; binding to [3H]SA was partially suppressed by excess unlabeled SA, but not by excess amount of 4‐HBA (Figure 1c). An independent, parallel screen using a SA‐derived ligand in combination with the yeast three‐hybrid technology, which relies on the in vivo interaction between the ligand (small molecule) and its protein target in the yeast nucleus, also identified PP2C‐D4 (called PP2C6 in (Cottier et al., 2011). Based on these five independent assays, we conclude that PP2C‐D4 is a true SABP.
Figure 1.
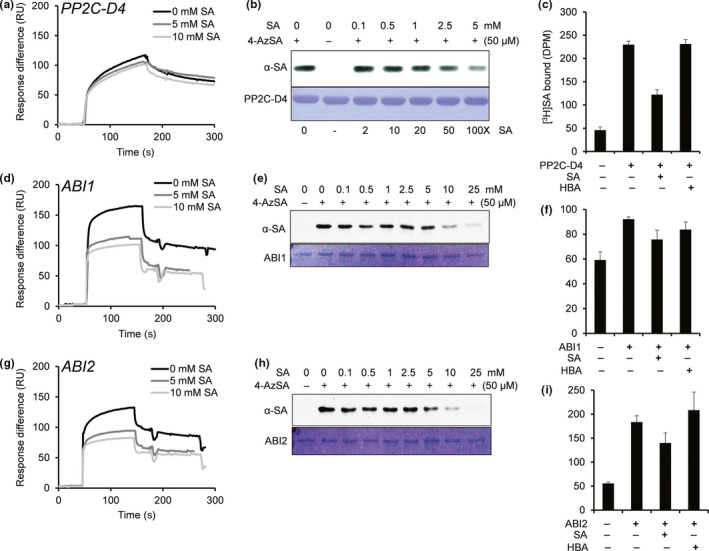
Several PP2Cs bind SA. (a, d, g) Sensorgrams for 1 μM of His6‐tagged PP2C‐D4 (At3g55050) (a), ABI1 (At4g26080) (d), and ABI2 (At5g57050) (g) in the absence (0 mM) or presence of two concentrations of SA (5 or 10 mM) using a 3AESA‐immobilized SPR sensor chip. Signals detected from a mock‐coupled control chip were subtracted. (b, e, h) Photo‐activated crosslinking of 50 ng of PP2C‐D4 (b), ABI1 (e), and ABI2 (h) to 4AzSA (50 μM) in the absence or presence of increasing amounts of SA was detected by immunoblotting using an α‐SA antibody. Reactions without 4AzSA served as negative controls. Proteins stained with Coomassie Brilliant Blue (CBB) served as the loading controls. (c, f, i) Binding of [3H]SA (200 nM) by 200 ng/μl PP2C‐D4 (c), ABI1 (f), and ABI2 (i) in the absence or presence of a 10,000‐fold excess of unlabeled SA was determined by size‐exclusion chromatography. Chromatography with [3H]SA in the absence of protein served as negative controls. Reactions with [3H]SA with excess of 4‐amino benzoic acid (4‐HBA), an inactive SA analog, served as negative controls for SA‐specific competitive inhibition. The experiments were independently repeated at least twice
Several clade A PP2C family members, including ABI1, ABI2, and HAB1, have been identified as core components of the ABA signaling network (Soon et al., 2012). This prompted us to test whether these proteins also bind SA. Like PP2C‐D4, recombinant ABI1 and ABI2 bound the 3AESA‐immobilized sensor chip and crosslinked with 4AzSA (Figure 1d,e,g,h). The ability of ABI1 and ABI2 to bind 3AESA and crosslink to 4AzSA also was partially suppressed by SA in a dose‐dependent manner. ABI2's binding to [3H]SA was comparable to that of PP2C‐D4 and this binding was partially suppressed by excess unlabeled SA but not by excess 4‐HBA, while ABI1 exhibited relatively weak binding to [3H]SA and suppression by excess unlabeled SA (Figure 1f,i). Interestingly, SA suppressed the binding of these proteins to 3AESA more effectively than that of PP2C‐D4 (Figure 1d,g). In contrast, HAB1 displayed much weaker binding to the 3AESA‐immobilized sensor chip (Fig. S1a). The ability of phosphoprotein phosphatase 2A regulatory subunit A (PP2A), a component of phosphatases belonging to the PPP family, also was tested for SA binding. This protein was previously identified during our screens, but it failed to meet the criteria as an SABP (Manohar et al., 2015). Consistent with these results, PP2A exhibited very weak binding to the 3AESA‐immobilized sensor chip (Fig. S1b). Whether other key components involved in ABA signaling also bind SA was then assessed. Size‐exclusion chromatography revealed little or no binding of [3H]SA by three members of the PYR/PYL/RCAR family of ABA receptors (PYL1, PYL2, and PYR1) or by three SnRK2s (SnRK2.2, 2.3, and 2.6) (Fig. S1c). Likewise, SnRK2.2 displayed only very low‐level binding to the 3AESA‐immobilized sensor chip (Fig. S1d). Together, these results suggest that SA preferentially interacts with specific PP2C family members, but not with other major components of the ABA signaling pathway.
As clade A PP2Cs are negative regulators of the ABA signaling pathway, we tested whether the presence of ABA affects PP2C–SA interactions by flowing the PP2Cs over 3AESA‐immobilized sensor chips in the absence or presence of ABA. Notably, binding of PP2C‐D4 to 3AESA was significantly enhanced in the presence of high μM to low mM levels of ABA (Figure 2a; Fig. S2). Similarly, binding of ABI1 and ABI2 was significantly enhanced in the presence of ABA (Figure 2b,c). This ABA‐induced enhancement of PP2Cs was suppressed nearly by 50% in the presence of excess amount of SA, further arguing that 3AESA binding by these PP2Cs represents authentic SA‐binding activity. In contrast, ABA failed to enhance the weak binding of HAB1 and PP2A or to enable SnRK2.2 to bind 3AESA (Fig. S1a,b,d). To facilitate competition analyses, high level of ABA was used to detect strong enhancement of PP2C binding to CM5 chip, which then necessitated the need to also use relatively high levels of SA.
Figure 2.
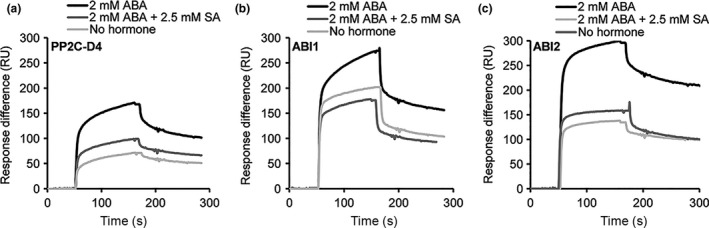
SA‐binding activities of PP2Cs are enhanced by ABA and this binding is partially suppressed by SA. (a–c) Sensorgrams obtained with recombinant, purified 1 μM of His6‐tagged PP2C‐D4 (a), ABI1 (b), and ABI2 (c) using a 3AESA‐immobilized sensor chip in the absence or in the presence of 2 mM ABA or 2 mM ABA plus 2.5 mM SA. Signals detected from a mock‐coupled control chip were subtracted. The experiments were independently repeated at least twice
2.2. SA suppresses the ABA‐enhanced interaction between PP2Cs and ABA receptors
To assess whether SA binding by PP2Cs alters their ability to interact with ABA receptors, SPR was performed. Purified PYL1 was immobilized on the CM5 sensor chip via an amide bond and interactions were detected by flowing purified PP2C‐D4, ABI1, or ABI2 over the sensor chip. Dose‐dependent binding responses were obtained with all three PP2Cs tested, and this binding was enhanced several fold in the presence of ABA (Fig. S3). Notably, SA suppressed the ABA‐enhanced interaction between PYL1 and all three PP2Cs by 35%–55% (Figure 3). SPR analysis with PYL2 and PYR1 revealed similarly dose‐dependent binding to the PP2Cs, which was further enhanced in the presence of ABA (Figs. S4 and S5). Furthermore, this ABA‐enhanced binding was suppressed by SA, although the level of suppression varied depending on the identity of the interacting proteins. For example, SA only weakly suppressed the ABA‐enhanced interactions between PYR1 and all three PP2Cs, while interactions between these PP2Cs and either PYL1 or PYL2 were more strongly suppressed by SA (Figs. S4 and S5). The ABA‐enhanced interactions between PP2C‐D4 and PYL1 or PYL2 also were suppressed less effectively by SA than the interactions between these ABA receptors and other PP2Cs (Figure 3; Fig. S4). Interestingly, SA alone consistently enhanced the interaction between PP2C‐D4 and all three ABA receptors (Figure 3; Figs. S4 and S5). Note that due to the low amount of PYL1 and the other ABA receptor that could be covalently attached to the CM5 sensor chips, relatively high, nonphysiological levels of ABA were needed to detect changes in the binding of the PP2Cs to PYL1 and other receptors, which then necessitated the need to also use relatively high levels of SA.
Figure 3.
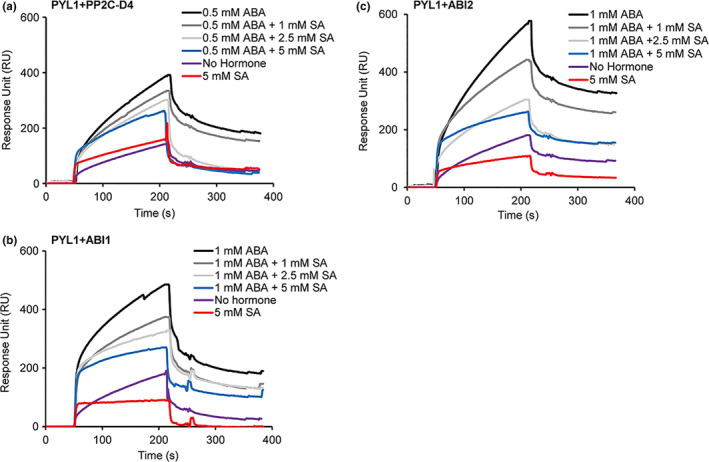
SA disrupts the ABA‐induced interaction between PP2Cs and PYL1. (a–c) Sensorgrams obtained with recombinant, purified 10 μM of His6‐tagged PP2C‐D4 (a), 1 μM of ABI1 (b) or 2.5 μM of ABI2 (c) using a His6‐SUMO‐tagged PYL1‐immobilized sensor chip in the absence or presence of the indicated concentrations of ABA or SA. Signals detected from a mock‐coupled control chip were subtracted. The experiments were independently repeated at least twice
2.3. SA suppresses ABA‐enhanced degradation of PP2Cs and ABA‐induced stabilization of SnRK2s
Proteolysis plays an important role in regulating plant responses to various stresses by fine‐tuning the turnover of key signaling components (Vierstra, 2009). Recent studies in Arabidopsis and rice have demonstrated that all three key components of ABA signaling, namely PYR/PYL/RCARs, PP2Cs, and SnRK2s, are regulated by controlled proteolysis (Irigoyen et al., 2014; Kong et al., 2015; Lin et al., 2015). For example, ABA promotes degradation of ABI1, but it suppresses degradation of certain PYR/PYL/RCARs and SnRK2s (Kong et al., 2015; Lin et al., 2015). Furthermore, the plant hormone gibberellic acid (GA) antagonizes ABA signaling, in part, by stimulating degradation of PYR/PYL/RCARs and SnRK2s (Lin et al., 2015). To determine whether SA affects protein turnover, we analyze the stability of purified recombinant PP2Cs and SnRK2s in a cell‐free degradation assay. Following incubation in protein extracts prepared from Arabidopsis seedlings supplemented with ABA and/or SA, immunoblot analyses indicated that the levels of His6‐tagged PP2C‐D4, ABI1, and ABI2 decreased in extracts supplemented with 10 μM ABA (Figure 4a). By contrast, the levels of these proteins remained fairly stable in extracts supplemented with both ABA and SA. SA alone had little effect on ABI1 or ABI2 levels but a modest decrease in PP2C‐D4 levels was detected (Figure 4a). Together, these results suggest that ABA enhances PP2C degradation and that this heightened turnover is suppressed by SA.
Figure 4.
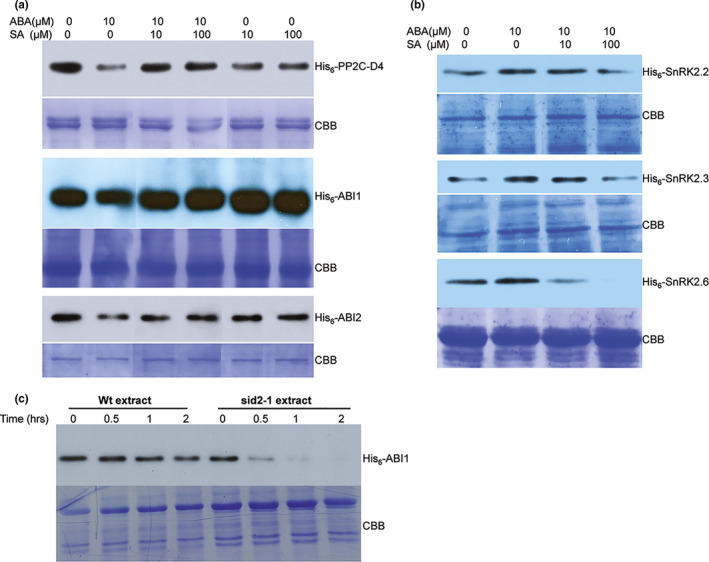
SA alters ABA‐mediated turnover of PP2Cs and SnRK2s. (a) Cell‐free degradation assay using approximately 100 μg of total protein extracts prepared from 10‐day‐old Arabidopsis seedlings supplemented with 500 ng of His6‐tagged PP2Cs (PP2C‐D4, ABI1, and ABI2) and indicated concentrations of ABA, SA, or ABA+SA. (b) Cell‐free degradation assay using approximately 100 μg of total protein extracts prepared from 10‐day‐old Arabidopsis seedlings supplemented with 500 ng of His6‐Sumo‐tagged SnRK2s (SnRK2.2, 2.3, and 2.6) and indicated concentrations of ABA or ABA+SA. For a and b, the degradation assay was carried out at 30°C for 3 hr. All lanes shown are from the same experiment; some lanes unrelated to this study have been removed and lanes were then merged for clarity of presentation. (c) Cell‐free degradation assay using approximately 100 μg of total protein extracts prepared from 10‐day‐old wild‐type or sid2‐1 Arabidopsis seedlings supplemented with 500 ng of His6‐tagged ABI1. Samples were taken after 0, 0.5, 1, or 2 hr of incubation; proteolysis was stopped by the addition of SDS‐PAGE buffer. Proteins were detected by immunoblotting using an α‐His6‐HRP antibody. Staining with Coomassie Brilliant Blue (CBB) staining of the gel served as a loading control. The experiments were independently repeated at least twice
The stability of three SnRK2s, SnRK2.2, SnRK2.3, and SnRK2.6, was then assessed using the cell‐free protein degradation assay. The levels of all three recombinant SnRK2s were slightly greater in extracts supplemented with 10 μM ABA as compared with unsupplemented extracts (Figure 4b). By contrast, SnRK2 levels were reduced in extracts containing both ABA and SA, with the greatest decrease detected after supplementation with ABA and 100 μM SA. Thus, ABA appears to stabilize SnRK2s, while SA suppresses ABA's effect. Analysis of PYL1 did not reveal any change in protein levels regardless of supplementation with ABA and/or SA, suggesting that these hormones do not affect PYL1 stability (Fig. S6).
The above results raised the possibility that endogenous SA antagonizes ABA signaling, at least in part, by stabilizing PP2Cs. To further assess this, the rate of ABI1 degradation was compared in protein extracts prepared from wild‐type (WT) plants and the SA biosynthesis‐deficient mutant sid2‐1. ABI1 levels in the extract from sid2‐1 plants decreased substantially by 30 min and were barely detectable after 1 hr, whereas those in the WT extract decreased gradually over time (Figure 4c). Surprisingly, the enhanced degradation observed in sid2‐1 extracts was not reversed by (i) adding SA to the extract, (ii) spraying SA on sid2‐1 plants, or (iii) supplementing sid2‐1 growth media with SA (Fig. S7). Thus, while these results suggest that SA stabilizes ABI1, the failure of exogenous SA to slow ABI1 degradation in sid2‐1 extracts suggests that an additional factor might be involved in this process.
2.4. SA antagonizes ABA‐induced gene expression in vivo
Previous studies have demonstrated that exogenously supplied ABA induces the accumulation of ABI1 and ABI2 transcripts (Hoth et al., 2002). To determine whether SA antagonizes the expression of these ABA signaling components, transcript levels for ABI1 and ABI2, as well as PP2C‐D4, were monitored in ABA‐ and/or SA‐treated Arabidopsis. Quantitative reverse transcriptase PCR (qRT‐PCR) analyses showed that transcripts for ABI1 and ABI2 accumulated after ABA, but not SA, treatment (Figure 5a). An intermediate level of transcripts was detected in plants treated with SA and ABA, suggesting that the ABA‐induced expression of these genes was partially suppressed by SA (Figure 5a). In comparison with the clade A PP2Cs, transcript accumulation for PP2C‐D4 was reduced in plants treated with either ABA or SA; an even greater reduction was observed in plants treated with both hormones. The expression of two well‐known ABA‐responsive genes, response to desiccation 29A (RD29A) and ABA‐responsive element binding protein 2 (AREB2), also was analyzed. Consistent with previous studies, the expression of RD29A and AREB2 was induced by ABA (Figure 5b) (Nakashima et al., 2006; Uno et al., 2000). Importantly, plants treated with ABA and SA accumulated reduced levels of RD29A and AREB2 transcripts, indicating that the ABA‐induced expression of these genes is partially suppressed by SA. By contrast, SA alone did not affect the expression of either gene.
Figure 5.
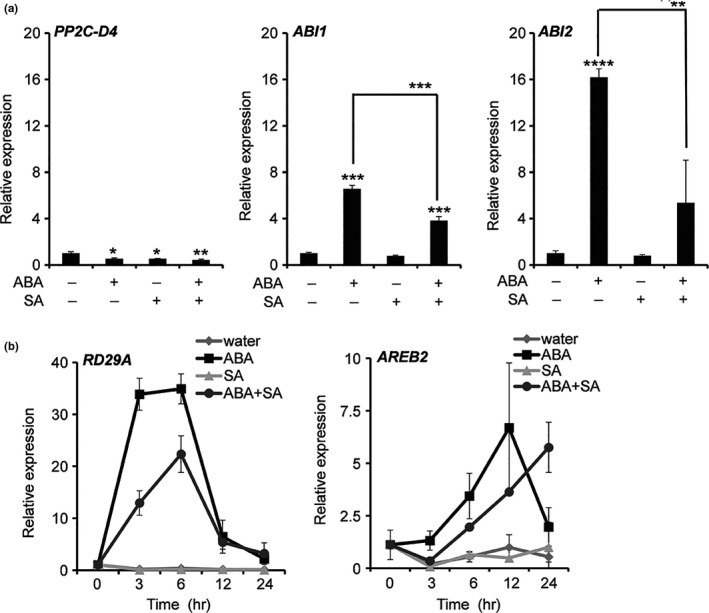
SA suppresses ABA‐induced gene expression. (a) Transcript levels, as measured by qRT‐PCR, in seedlings pretreated with either water, 100 μM ABA, 100 μM SA, or 100 μM ABA plus 100 μM SA. Transcript levels of PP2C‐D4, ABI1, and ABI2 were determined at 3 hr post‐treatment (hpt). Data are averaged ±SD (n = 3). (b) Transcript levels as measured by qRT‐PCR of ABA‐responsive marker genes in seedlings pretreated with either water, 100 μM ABA, 100 μM SA, or 100 μM ABA plus 100 μM SA. Transcript levels of RD29A (RESPONSIVE TO DESICCATION 29A) and AREB2 (ABRE BINDING FACTOR 2) were determined at 0, 3, 6, 12, and 24 hpt. The relative expression levels were quantified by normalizing to ubiquitin expression level. Data are averaged ±SD (n = 4). *p ≤ .05; **p ≤ .005; ***p ≤ .0005; ****p ≤ .00005; two‐tailed t test. The experiments were independently repeated at least twice
2.5. An SA‐deficient mutant is more sensitive to ABA‐mediated seed dormancy
In addition to (a)biotic stress responses, ABA is involved in growth and developmental processes, including maintaining seed dormancy to prevent untimely germination (Hubbard et al., 2010; Kermode, 2005). To investigate whether SA antagonizes ABA's ability to suppress germination, we monitored Arabidopsis seed germination on plates containing Murashige and Skoog (MS) medium in the presence or absence of ABA and/or SA. In the presence of 1 μM ABA, germination was dramatically reduced at all times monitored (Figure 6a). By contrast, 10 μM SA reduced germination slightly at 36 hr, but from 48 hr onward the germination percentage of SA‐treated and control seeds was comparable. Plates containing both SA and ABA displayed an intermediate level of germination. Thus, SA appears to suppress ABA‐mediated inhibition of germination. Whether endogenous SA levels also affect ABA‐mediated suppression of germination was then tested by comparing the germination of WT and sid2‐1 seeds. The germination rate for sid2‐1 seeds grown on ABA‐containing plates was consistently lower than that of comparably grown WT seeds; by 72 hr 15% of the sid2‐1 seeds had germinated in the presence of 1 μM ABA, in contrast to 40% of WT seeds (Figure 6b). While SA completely overcame ABA suppression of seed germination in WT by 96 hr postplating, it only partially reversed ABA's effect in sid2‐1. Based on the ABA‐hypersensitive phenotype displayed by sid2‐1 seeds, endogenous SA appears to play an important role in antagonizing ABA‐mediated suppression of seed germination in planta.
Figure 6.
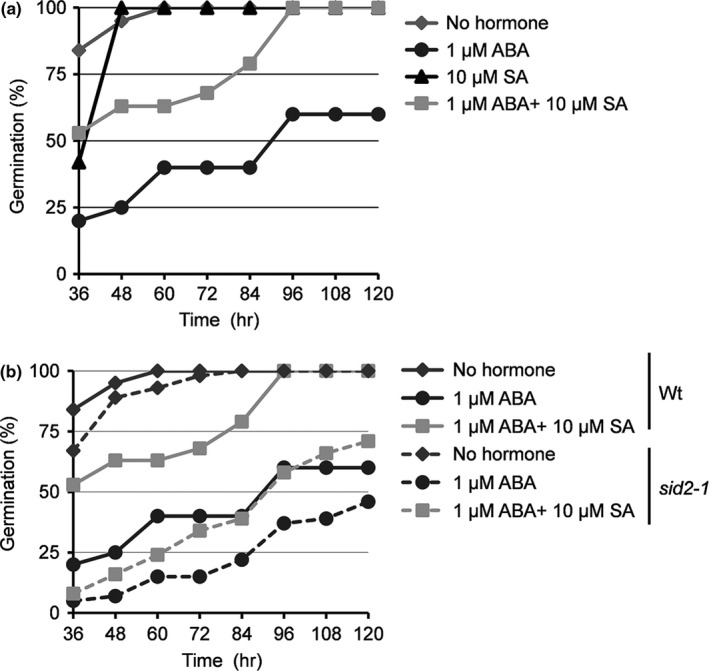
SA antagonizes ABA‐mediated suppression of seed germination. (a) Germination rate of Arabidopsis wild‐type seeds on MS medium containing no hormone or in the presence of 1 μM ABA, 10 μM SA, or 1 μM ABA plus 10 μM SA. (b) Comparison of germination rate between wild‐type and sid2‐1 on MS medium containing no hormone or in the presence of 1 μM ABA or 1 μM ABA plus 10 μM SA. The germination time course for wild‐type seeds is shown with a solid line, while for sid2‐1 it is shown with a broken line (n = 40 seeds). The percentage of germinated seeds was determined at 36, 48, 60, 72, 84, 96, 108, and 120 hr postplating. The experiments were independently repeated at least twice
3. DISCUSSION
Elucidating the crosstalk between biotic and abiotic stress signaling pathways in plants is a rapidly expanding area of research. There is a growing recognition that ABA not only regulates abiotic stress responses and developmental processes, but also impacts plant–pathogen interactions (Denance et al., 2013; Robert‐Seilaniantz et al., 2011). Likewise, SA not only signals plant immunity (Klessig et al., 2016; Manohar et al., 2015; Vlot et al., 2009), but also regulates responses to abiotic stresses and various aspects of growth and development (Hayat, Hayat, Irfan, & Ahmad, 2010; Khan, Fatma, Per, Anjum, & Khan, 2015). To gain insights into how SA exerts its myriad effects, we previously developed several high‐throughput screens for identifying SABPs (Choi et al., 2015; Manohar et al., 2015; Tian et al., 2012). Here, we report that several PP2Cs, including PP2C‐D4, a member of clade D, and ABI1 and ABI2, members of clade A, are novel SABPs. By contrast, SA binding was not detected for two other phosphatases, HAB1 or PP2A, or for other components of the ABA signaling pathway, including various PYR/PYL/RCARs and SnRK2s. SPR analysis also revealed that binding of PP2C‐D4, ABI1, and ABI2 to the SA analog 3AESA was enhanced in the presence of ABA. This finding suggests that PP2Cs from both clade A and clade D bind both ABA and SA and that they do so in a cooperative manner. Although a previous report failed to detect binding between ABI1 and ABA (Ma et al., 2009), this discrepancy may be due to the very high sensitivity of SPR. Indeed, Ma et al. (2009) noted that ABI1 phosphatase activity was reduced up to 20% in presence of ABA.
As clade A PP2Cs are critical negative regulators of ABA signaling, the discovery that they bind SA suggested that they play a role in modulating SA/ABA crosstalk. Several studies have documented an antagonistic relationship between SA and the ABA signaling pathway. For example, SA suppressed ABA‐mediated inhibition of shoot growth and expression of cell cycle‐related genes in rice (Meguro & Sato, 2014). Likewise, pretreating Arabidopsis with a compound that activates SA‐dependent defense signaling antagonized the induction of ABA biosynthesis‐related and ABA‐responsive genes after NaCl treatment (Yasuda et al., 2008). Expanding on these findings, we demonstrated that SA treatment suppresses ABA‐induced expression of the ABA signaling components ABI1 and ABI2 and the ABA‐responsive genes RD29A and AREB2. In addition, SA antagonized ABA's ability to suppress seed germination. The combined observations that (i) SA‐deficient sid2‐1 seeds germinated more slowly than WT seeds and (ii) sid2‐1 seeds were hypersensitive to exogenously supplied ABA argue that endogenous SA plays an important role in counteracting the effects of both endogenously and exogenously supplied ABA.
To investigate the mechanism through which SA antagonizes ABA signaling, we monitored the interaction between several ABA receptors and PP2Cs. SPR analyses revealed that the clade A PP2Cs, ABI1 and ABI2, bind PYL1, PYL2, and PYR1 even in the absence of ABA (Figure 7a); however, these interactions were strongly enhanced in the presence of ABA (Figure 7b). Strikingly, SA suppressed the ABA‐enhanced interaction between these proteins, albeit to varying extents depending on the identity of the interacting partners (Figure 7c). Consistent with these results, both in vitro and in vivo analyses have previously demonstrated that ABA strongly enhances binding between clade A PP2Cs and certain ABA receptors, including PYL1, PYL2, and PYR1 (Park et al., 2009). Crystal structure analyses further demonstrated that this interaction inhibits PP2C activity by occluding PP2C's active site (Melcher et al., 2009; Miyazono et al., 2009). As PP2Cs repress ABA signaling by preventing autophosphorylation‐dependent activation of SnRK2s (Soon et al., 2012; Umezawa et al., 2009), ABA‐induced binding of PP2Cs by ABA receptors is a critical step in activating ABA signaling (Fujii et al., 2009). SA's ability to suppress the interaction between ABI1 or ABI2 and the ABA receptors therefore provides one mechanism through which SA can antagonize ABA signaling. In addition, our cell‐free degradation assay revealed that SA suppresses the ABA‐enhanced turnover of PP2Cs and stabilization of SnRK2s. Given that ABI1 was degraded substantially more rapidly in extracts from sid2‐1 mutants than from WT plants, endogenous SA appears to play an important role in regulating cellular PP2C levels. Taken together, these results suggest that SA antagonizes ABA signaling via multiple mechanisms that both promote the enzymatic activity and/or protein stability of negative regulators and decrease the stability of downstream effectors.
Figure 7.
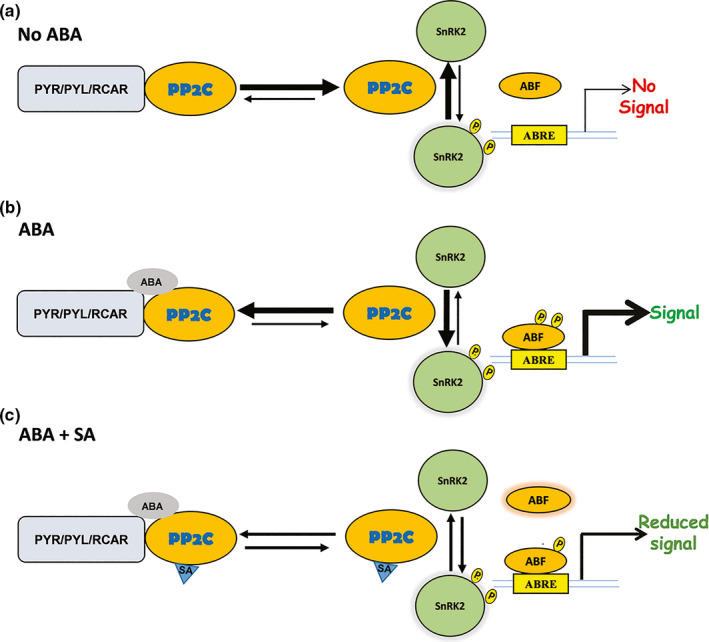
Schematic illustrating part of SA's antagonistic effects on the ABA signaling module. (a) In the absence of ABA, free PP2Cs prevents autophosphorylation‐dependent activation of SnRK2s by dephosphorylating them. (b) In the presence of ABA, PYR/PYL receptors tightly bind to PP2Cs, thereby preventing free PP2C‐mediated dephosphorylation of SnRK2s. Receptor‐mediated occlusion of PP2Cs allows autophosphorylation‐dependent activation of SnRK2s to relay the ABA signaling by phosphorylating downstream targets such as abscisic acid‐responsive element‐binding factor 2 (ABF2), which enables its binding to ABA‐responsive elements (ABRE) in the promoter region of ABA‐responsive genes. (c) SA suppresses ABA's enhancement of the interaction of PP2Cs with the PYR/PYL/RCAR receptor and the resulting autophosphorylation‐dependent activation of SnRK2s, which results in reduced ABA signaling. The length and thickness of the arrows indicate the equilibrium between free and receptor‐bound PP2Cs and between inactive, nonphosphorylated and active, autophosphorylated SnRK2s
Within this overall framework, differences among the binding specificities and affinities, protein–protein interactions and/or stability of various ABA signaling components may further influence SA/ABA crosstalk. For example, while ABI1 and ABI2 bound SA, another clade A member, HAB1 did not. ABI1 differs from ABI2 as it displayed substantially greater affinity for all three ABA receptors in the absence of ABA; it also was the most stable PP2C in our in vitro degradation assay. SA's ability to disrupt the ABA‐enhanced interactions between ABA receptors and PP2Cs also varied, depending on the proteins involved. In particular, the interaction between PYR1 and ABI1 or ABI2 was suppressed less effectively by SA than the interactions between these PP2Cs and the other ABA receptors. Similar to these findings, reconstitution of the ABA signaling pathway in Arabidopsis protoplasts using different combinations of ABA receptors, PP2Cs, and SnRK2s previously revealed that the intensity of interactions varied significantly depending on which members of the protein families were involved (Fujii et al., 2009). Combined with our findings, these results suggest that while various members of the PP2C, ABA receptor, and SnRK2 families serve overlapping functions, differences in their temporal and/or spatial expression patterns, as well as their affinity for specific interacting partners and/or SA and ABA, could fine‐tune ABA signaling and regulate crosstalk with the SA pathway.
In comparison with the clade A PP2Cs, members of clade D were recently shown to negatively regulate cell expansion by dephosphorylating and thereby inactivating plasma membrane H+‐ATPases (Spartz et al., 2014). In the presence of auxin, this suppression is relieved by members of the SAUR (small auxin up‐regulated) protein family, which bind and inhibit PP2C‐Ds. Interestingly, while different SAURs inhibited the activity of several PP2C‐D family members (including PP2C‐D4) to varying extents, they did not inhibit the activity of a clade A PP2Cs, ABI1, a clade E member, or a phosphatase belonging to the PPP family. Our studies provide additional insights into PP2C‐D function, as we demonstrate that PP2C‐D4 is an SABP, whose binding to 3AESA is enhanced by ABA. PP2C‐D4 also bound several ABA receptors, and this interaction was enhanced by the presence of ABA. In comparison with the clade A PP2C–ABA receptor interactions, however, the binding between PP2C‐D4 and the ABA receptors was substantially weaker and its suppression by SA was less effective. In addition, SA consistently stimulated this interaction in the absence of ABA. The expression pattern of PP2C‐D4 in the presence of ABA and/or SA also differed significantly from that of the clade A PP2Cs. Thus, while PP2C‐D4 and the clade A PP2Cs share some common features, many differences between them are consistent with the previous demonstration that clade D and clade A PP2Cs serve distinct functions.
In response to stress, plants maximize their chances of survival and reproduction by redistributing cellular resources from growth and developmental processes to defensive responses (Asselbergh, Achuo, Höfte, & Van Gijsegem, 2008; Atkinson & Urwin, 2012). Many studies have assessed plant responses to individual stresses, but there is a growing recognition that plants in the field contend with multiple stresses simultaneously and that, depending on the specific stresses, the responses may be additive or antagonistic. Thus, the hormones responsible for mediating developmental processes and stress responses are involved in complex crosstalk that ultimately allows the plant to tailor its response to the environmental conditions. A previous study demonstrated that ABA can interfere with SA‐mediated innate immune responses by down‐regulating SA biosynthesis (De Torres Zabala et al., 2009). By contrast, our results provide a novel mechanism through which SA can antagonize ABA by interfering with multiple aspects of the ABA signaling pathway. The discovery that PP2C‐D4 binds SA and several ABA receptors, and that this binding is enhanced in the presence of ABA, suggests an additional mechanism through which SA and ABA can negatively regulate auxin‐mediated growth and developmental processes. Indeed, ABA was shown to suppress hypocotyl elongation, and this correlated with dephosphorylation of H+‐ATPases (Hayashi, Takahashi, Inoue, & Kinoshita, 2014). These ABA‐induced responses were suppressed in the abi1‐1 mutant, suggesting that clade A PP2Cs are involved in auxin‐mediated physiological processes. Future studies will be required to determine whether PP2C‐D4 and/or other clade D PP2Cs also mediate ABA antagonism of cell expansion and whether SA binding by PP2C‐D4 affects its ability to interact with SAURs and thereby impact auxin signaling.
In summary, we have elucidated SA mechanisms of action in negatively regulating ABA signaling, which likely serves to properly balance the plant response to multiple stresses. As plants confront a changing climate, this balance and our understanding of how it is regulated and might be beneficially altered take on increased significance.
4. EXPERIMENTAL PROCEDURES
4.1. Plant materials and growth conditions
The wild‐type and sid2‐1 Arabidopsis thaliana ecotype Col‐0 plants were grown on standard Murashige and Skoog (MS) media containing half‐strength of MS with pH adjusted to 6.0 by KOH and supplemented with 10 g/L sucrose. Arabidopsis seeds were first surface‐sterilized by soaking in a solution of 30% bleach with 0.1% Triton X‐100 for 5–10 min and then rinsed five times with sterile water. The surface‐sterilized seeds were incubated at 4°C for 2 days for stratification before planting on the MS media. For the seed germination assay (±) abscisic acid (Caisson labs) or salicylic acid (Sigma) was added directly into the MS media. The plates with seed were placed vertically in the growth chamber with 16/8‐hr light/dark cycle, 22°C, and 70% humidity. The germination rate was measured. For spray treatment, one‐week‐old seedlings were subjected to water, ABA, SA, or ABA+SA spray treatment and whole seedlings were collected for RNA analysis. For cell‐free degradation assay, 10‐day‐old wild‐type or sid2‐1 seedlings were subjected to water, ABA, SA, or ABA+SA spray treatment to compare the effects of protein extracts on the stability of ABI1.
4.2. Cloning and plasmid constructs
All oligonucleotides used for cloning and plasmid construction are listed in Table S1. ORFs of PP2CD, ABI1, ABI2, and HAB1 were amplified from an Arabidopsis cDNA library. The resulting PCR products were digested with NdeI and BamHI for ABI1, NdeI and SacI for ABI2, and HAB1 and cloned into the expression vector pET28a (EMD Millipore, MA, USA) for expression. PP2CD was cloned into pET42a (EMD Millipore, MA, USA) using NdeI and XhoI cloning sites. Cloning of PYL1, PYL2, PYR1, SnRK2.2, SnRK2.3, and SnRK2.6 into pSUMO‐H6SUMO vector was described previously (Soon et al., 2012).
4.3. Protein purifications
Two‐step protein purifications were performed as described previously (Manohar et al., 2017). Briefly, the Rosetta 2 (DE3) (EMD, Millipore, MA, USA) bacterial cells were grown at 37οC in LB medium containing 50 μg/ml kanamycin and 34 μg/ml chloramphenicol until the OD600 of the culture reached approximately 0.6 before the addition of isopropyl‐β‐d‐thiogalactoside (IPTG) to a final concentration of 1 mM to induce expression. Induced culture was incubated overnight at 20οC. The cells were then harvested by centrifugation and the pellet was resuspended in the lysis buffer (50 mM Tris pH 7.5, 500 mM NaCl, 10% glycerol, 20 mM imidazole, 0.5% Triton X‐100, and 1 mM phenylmethylsulphonyl fluoride) and disrupted by sonication. The clarified supernatant obtained after centrifugation was incubated with Ni‐NTA His resin (Novagen, MA, USA), and the bound protein was eluted in lysis buffer supplemented with 250 mM imidazole. The eluted proteins were then subjected to gel filtration chromatography on a HiLoad 16/600 Superdex 200 prep grade column (GE Healthcare, PA, USA), using gel filtration buffer (50 mM Tris pH 7.5, 150 mM NaCl, and 10% glycerol). The two‐step purified proteins were stored at −80°C.
4.4. Assessment of 3AESA‐binding activities by surface plasmon resonance
Surface plasmon resonance (SPR) analyses of 3AESA binding and competition by SA were performed with a Biacore 3000 instrument (GE Healthcare) as described previously (Manohar et al., 2015). Immobilization of 3AESA on the CM5 sensor chip was performed as described previously (Tian et al., 2012). To test SA‐binding activity, proteins were diluted in HBS‐EP buffer (GE Healthcare) and passed over the sensor surface of the 3AESA‐immobilized/coupled or mock‐coupled flow cells. The specific binding signal was determined by subtracting the signal generated with the mock‐coupled flow cell from the signal generated by the 3AESA‐immobilized cell. To re‐use the sensor chips, bound proteins were stripped off by injecting NaOH solution (pH 12).
4.5. Assessment of protein–protein interactions by SPR
Protein interaction analyses were performed by SPR using Biacore 3000 instrument. His‐SUMO‐tagged PYL1, PYL2, and PYR1 were immobilized on a CM5 sensor chip by amine coupling (GE healthcare), essentially by following the manufacturer's instructions. Briefly, proteins were diluted in 10 mM sodium acetate, pH 5.0 buffer at a concentration of 50 μg/ml. CM5 sensor chip surface was activated by injecting 85 μl of EDC/NHS solution with a flow rate of 10 μl/min. After activation, protein solution was injected for 42 min with a flow rate of 10 μl/min. Finally, 85 μl of ethanolamine was flowed over the surface to deactivate remaining active groups and remove noncovalently bound protein with a flow rate of 10 μl/min. The protein immobilization level was stabilized for 12 hr by flowing HBS‐EP buffer with a flow rate of 10 μl/min. To test protein–protein interactions, protein analytes (PP2Cs and SnRK2s) were diluted to desirable concentration in HBS‐EP buffer in the presence or absence of various concentrations of ABA, SA, or ABA+SA, and then passed over the protein‐immobilized sensor surface and mock‐coupled flow cells with a flow rate of 30 μl/min. The higher flow rate was used to avoid mass transfer as recommended by the manufacturer. The binding signal was generated by subtracting the signal for mock‐coupled flow cells from that for the protein‐immobilized flow cells. To re‐use the chip, bound proteins were stripped off by injecting 8 μl of 10 mM glycine‐HCl solution (pH 3) with a flow rate of 30 μl/min.
4.6. RNA analyses
Unless stated otherwise, at least three biological replicates were used for all RNA analyses. For each replicate, total RNA from one‐week‐old Arabidopsis seedlings was isolated from a pool of five seedlings. Total RNA was isolated using Qiagen RNeasy plant mini kit (Qiagen) according to the manufacturer's instructions. DNAse treatment was performed using DNA‐free kit (Ambion) following the manufacturer's instructions. First‐strand cDNA was synthesized from 1 mg of RNA using M‐MLV reverse transcriptase (Promega). For quantitative real‐time PCR, transcripts were amplified using SYBR premix Ex Taq II (Takara) with gene‐specific primers listed in Table S1. Reactions were performed using a CFX96 touch Bio‐Rad Real‐time PCR system (Bio‐Rad). The PCR conditions were 95°C for 3 min (initial denaturation) followed by 44 cycles of amplifications (95°C for 10 s, 60°C for 30 s), followed by generation of a dissociation curve. The relative fold change was calculated according to the 2−▲▲Ct method (Manosalva et al., 2015). Ubiquitin was used as endogenous reference gene. The paired t test with an α‐level of 0.05 was used to compare transcript level in the ABA, SA, ABA+SA, and mock‐treated plant samples.
4.7. Cell‐free degradation assay
The tissue samples were collected from 10‐day‐old seedlings of wild‐type and sid2‐1 and finely grounded using liquid nitrogen. The total protein extracts were then prepared using protein extraction buffer (25 mM Tris pH 7.5, 10 mM NaCl, 10 mM MgCl2, and 4 mM PMSF). The sample was vortexed to mix and centrifuged twice at 17,000 g for 10 min at 4°C to remove debris. The clarified supernatant was pretreated with 1 mM cycloheximide (MP Biomedicals) for 1 hr to inhibit de novo protein biosynthesis. The extracts were then adjusted to equal protein concentrations in degradation buffer (25 mM Tris pH 7.5, 10 mM NaCl, 10 mM MgCl2, 4 mM PMSF, 5 mM DTT, and 10 mM ATP). For degradation assay, an equal amount (approximately 500 ng) of purified PP2Cs, SnRK2s, and PYL1 were incubated in 50 μl of Arabidopsis total protein extract (containing approximately 100 μg total proteins) at 28 °C for 3 h, unless otherwise indicated. For ABI1 and SnRK2.6, twice as much total protein extract (approximately 200 μg) was used to more clearly visualize the effect of SA (Figure 4a,b). Immunoblot analyses were performed to detect protein levels by using an α‐His6‐HRP polyclonal antibody (QED Biosciences).
ACKNOWLEDGMENT
We thank D'Maris Dempsey for editing and the US National Science Foundation for financial support via Grant IOS‐0820405 to D.F.K. We thank Prof. Karsten Melcher (Van Andel Research Institute), for the E. coli expression vectors corresponding to PYL1, PYL2, PYR1, SnRK2.2, SnRK2.3, and SnRK2.6.
AUTHORS' CONTRIBUTIONS
M.M., P.M., and D.F.K. conceived the research. M.M., D.W., E. K., and D.F.K. designed the research. M.M, D.W, P.M., and H.W.C. performed the research. M.M., D.W., E.K., and D.F.K. analyzed the data. M.M. and D.F.K. wrote the manuscript. All authors have read and approved the final manuscript.
Supporting information
Manohar M, Wang D, Manosalva PM, Choi HW, Kombrink E, Klessig DF. Members of the abscisic acid co‐receptor PP2C protein family mediate salicylic acid–abscisic acid crosstalk. Plant Direct. 2017;1:1–14. 10.1002/pld3.20
Link of manuscript published on the biorxiv beta (the preprint server for biology) https://www.biorxiv.org/content/early/2017/03/31/123059.
Significance Statement: The phytohormone salicylic acid (SA) antagonizes phytohormone abscisic acid (ABA) signaling by binding to type 2C protein phosphatases, which suppresses their ABA‐enhanced turnover as well as their interaction with the ABA receptors.
This manuscript was previously deposited as a preprint at doi: https://doi.org/10.1101/123059
REFERENCES
- Asselbergh, B. , Achuo, A. E. , Höfte, M. , & Van Gijsegem, F. (2008). Abscisic acid deficiency leads to rapid activation of tomato defence responses upon infection with Erwinia chrysanthemi . Molecular Plant Pathology, 9, 11–24. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Atkinson, N. J. , & Urwin, P. E. (2012). The interaction of plant biotic and abiotic stresses: From genes to the field. Journal of Experimental Botany, 63, 3523–3544. [DOI] [PubMed] [Google Scholar]
- Audenaert, K. , De Meyer, G. B. , & Höfte, M. M. (2002). Abscisic acid determines basal susceptibility of tomato to Botrytis cinerea and suppresses salicylic acid‐dependent signaling mechanisms. Plant Physiology, 128, 491–501. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Choi, H. W. , Tian, M. , Song, F. , Venereau, E. , Preti, A. , Park, S.‐W. , … Klessig, D. F. (2015). Aspirin's active metabolite salicylic acid targets high mobility group Box 1 to modulate inflammatory responses. Molecular Medicine, 21, 526–535. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Cottier, S. , Monig, T. , Wang, Z. , Svoboda, J. , Boland, W. , Kaiser, M. , & Kombrink, E. (2011). The yeast three‐hybrid system as an experimental platform to identify proteins interacting with small signaling molecules in plant cells: Potential and limitations. Frontiers in Plant Science, 2, 101. [DOI] [PMC free article] [PubMed] [Google Scholar]
- De Torres Zabala, M. , Bennett, M. H. , Truman, W. H. , & Grant, M. R. (2009). Antagonism between salicylic and abscisic acid reflects early host‐pathogen conflict and moulds plant defence responses. Plant Journal, 59, 375–386. [DOI] [PubMed] [Google Scholar]
- de Torres‐Zabala, M. , Truman, W. , Bennett, M. H. , Lafforgue, G. , Mansfield, J. W. , Rodriguez Egea, P. , … Grant, M. (2007). Pseudomonas syringae pv. tomato hijacks the Arabidopsis abscisic acid signalling pathway to cause disease. The EMBO Journal, 26, 1434–1443. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Denance, N. , Sanchez‐Vallet, A. , Goffner, D. , & Molina, A. (2013). Disease resistance or growth: The role of plant hormones in balancing immune responses and fitness costs. Frontiers in Plant Science, 4, 155. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Fu, Z. Q. , Yan, S. , Saleh, A. , Wang, W. , Ruble, J. , Oka, N. , … Dong, X. (2012). NPR3 and NPR4 are receptors for the immune signal salicylic acid in plants. Nature, 486, 228–232. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Fuchs, S. , Grill, E. , Meskiene, I. , & Schweighofer, A. (2013). Type 2C protein phosphatases in plants. FEBS Journal, 280, 681–693. [DOI] [PubMed] [Google Scholar]
- Fujii, H. , Chinnusamy, V. , Rodrigues, A. , Rubio, S. , Antoni, R. , Park, S. , … Zhu, J. (2009). In vitro reconstitution of an abscisic acid signalling pathway. Nature, 462, 660–664. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Hauser, F. , Waadt, R. , & Schroeder, J. I. (2011). Evolution of abscisic acid synthesis and signaling mechanisms. Current Biology, 21, R346–R355. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Hayashi, Y. , Takahashi, K. , Inoue, S. I. , & Kinoshita, T. (2014). Abscisic acid suppresses hypocotyl elongation by dephosphorylating plasma membrane H + ‐ATPase in Arabidopsis thaliana . Plant and Cell Physiology, 55, 845–853. [DOI] [PubMed] [Google Scholar]
- Hayat, Q. , Hayat, S. , Irfan, M. , & Ahmad, A. (2010). Effect of exogenous salicylic acid under changing environment: A review. Environmental and Experimental Botany, 68, 14–25. [Google Scholar]
- Hoth, S. , Morgante, M. , Sanchez, J. P. , Hanafey, M. K. , Tingey, S. V. , & Chua, N. H. (2002). Genome‐wide gene expression profiling in Arabidopsis thaliana reveals new targets of abscisic acid and largely impaired gene regulation in the abi1‐1 mutant. Journal of Cell Science, 15, 4891–4900. [DOI] [PubMed] [Google Scholar]
- Hubbard, K. E. , Nishimura, N. , Hitomi, K. , Hubbard, K. E. , Nishimura, N. , Hitomi, K. , … Schroeder, J. I. (2010). Early abscisic acid signal transduction mechanisms: Newly discovered components and newly emerging questions. Genes and Development, 24, 1695–1708. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Irigoyen, M. L. , Iniesto, E. , Rodriguez, L. , Puga, M. I. , Yanagawa, Y. , Pick, E. , … Rubio, V. (2014). Targeted degradation of abscisic acid receptors is mediated by the ubiquitin ligase substrate adaptor DDA1 in Arabidopsis. The Plant Cell, 26, 712–728. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kermode, A. R. (2005). Role of abscisic acid in seed dormancy. Journal of Plant Growth Regulation, 24, 319–344. [Google Scholar]
- Khan, M. I. R. , Fatma, M. , Per, T. S. , Anjum, N. A. , & Khan, N. A. (2015). Salicylic acid‐induced abiotic stress tolerance and underlying mechanisms in plants. Frontiers in Plant Science, 6, 462. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Klessig, D. F. , Tian, M. , & Choi, H. W. (2016). Multiple targets of salicylic acid and its derivatives in plants and animals. Frontiers in Immunology, 7, 206. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kong, L. , Cheng, J. , Zhu, Y. , Ding, Y. , Meng, J. , Chen, Z. , … Gong, Z. (2015). Degradation of the ABA co‐receptor ABI1 by PUB12/13 U‐box E3 ligases. Nature Communications, 6, 8630. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Lim, C. W. , Luan, S. , & Lee, S. C. (2014). A prominent role for RCAR3‐mediated ABA signaling in response to Pseudomonas syringae pv. tomato DC3000 infection in Arabidopsis . Plant and Cell Physiology, 55, 1691–1703. [DOI] [PubMed] [Google Scholar]
- Lin, Q. , Wu, F. , Sheng, P. , Zhang, Z. , Zhang, X. , Guo, X. , … Wan, J. (2015). The SnRK2‐APC/C(TE) regulatory module mediates the antagonistic action of gibberellic acid and abscisic acid pathways. Nature Communications, 6, 7981. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Ma, Y. , Szostkiewicz, I. , Korte, A. , Moes, D. , Yang, Y. , Christmann, A. , & Grill, E. (2009). Regulators of PP2C phosphatase activity function as abscisic acid sensors. Science, 324, 1064–1068. [DOI] [PubMed] [Google Scholar]
- Manohar, M. , Choi, H. W. , Manosalva, P. M. , Austin, C. , Peters, J. , & Klessig, D. F. (2017). Plant and human MORC proteins have DNA modifying activities similar to type II topoisomerases, but require additional factor(s) for full activity. Molecular Plant‐Microbe Interactions, 30, 87–100. [DOI] [PubMed] [Google Scholar]
- Manohar, M. , Tian, M. , Moreau, M. , Park, S.‐W. , Choi, H. W. , Fei, Z. , … Klessig, D. F. (2015). Identification of multiple salicylic acid‐binding proteins using two high throughput screens. Frontiers in Plant Science, 5, 1–14. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Manosalva, P. , Manohar, M. , von Reuss, S. H. , Chen, S. , Koch, A. , Kaplan, F. , … Klessig, D. F. (2015). Conserved nematode signalling molecules elicit plant defenses and pathogen resistance. Nature Communications, 6, 7795. [DOI] [PMC free article] [PubMed] [Google Scholar]
- McDonald, K. L. , & Cahill, D. M. (1999). Influence of abscisic acid and the abscisic acid biosynthesis inhibitor, norflurazon, on interactions between Phytophthora sojae and soybean (Glycine max). European Journal of Plant Pathology, 105, 651–658. [Google Scholar]
- Meguro, A. , & Sato, Y. (2014). Salicylic acid antagonizes abscisic acid inhibition of shoot growth and cell cycle progression in rice. Scientific Reports, 4, 4555. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Melcher, K. , Ng, L.‐M. , Zhou, X. E. , Soon, F.‐F. , Xu, Y. , Suino‐Powell, K. M. , … Xu, H. E. (2009). A gate‐latch‐lock mechanism for hormone signalling by abscisic acid receptors. Nature, 462, 602–608. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Meskiene, I. , Baudouin, E. , Schweighofer, A. , Liwosz, A. , Jonak, C. , Rodriguez, P. L. , … Hirt, H. (2003). Stress‐induced protein phosphatase 2C is a negative regulator of a mitogen‐activated protein kinase. Journal of Biological Chemistry, 278, 18945–18952. [DOI] [PubMed] [Google Scholar]
- Miyazono, K.‐I. , Miyakawa, T. , Sawano, Y. , Kubota, K. , Kang, H.‐J. , Asano, A. , … Tanokura, M. (2009). Structural basis of abscisic acid signalling. Nature, 462, 609–614. [DOI] [PubMed] [Google Scholar]
- Mohr, P. G. , & Cahill, D. M. (2003). Abscisic acid influences the susceptibility of Arabidopsis thaliana to Pseudomonas syringae pv. tomato and Peronospora parasitica . Functional Plant Biology, 30, 461–469. [DOI] [PubMed] [Google Scholar]
- Nakashima, K. , Fujita, Y. , Katsura, K. , Maruyama, K. , Narusaka, Y. , Seki, M. , … Yamaguchi‐Shinozaki, K. (2006). Transcriptional regulation of ABI3‐ and ABA‐responsive genes including RD29B and RD29A in seeds, germinating embryos, and seedlings of Arabidopsis. Plant Molecular Biology, 60, 51–68. [DOI] [PubMed] [Google Scholar]
- Park, S.‐Y. , Fung, P. , Nishimura, N. , Jensen, D. R. , Fujii, H. , Zhao, Y. , … Cutler, S. R. (2009). Abscisic acid inhibits type 2C protein phosphatases via the PYR/PYL family of START proteins. Science, 324, 1068–1071. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Raghavendra, A. S. , Gonugunta, V. K. , Christmann, A. , & Grill, E. (2010). ABA perception and signalling. Trends in Plant Science, 15, 395–401. [DOI] [PubMed] [Google Scholar]
- Robert‐Seilaniantz, A. , Grant, M. , & Jones, J. D. G. (2011). Hormone crosstalk in plant disease and defense: More than just JASMONATE‐SALICYLATE antagonism. Annual Review of Phytopathology, 49, 317–343. [DOI] [PubMed] [Google Scholar]
- Sanchez‐Vallet, A. , Lopez, G. , Ramos, B. , Delgado‐Cerezo, M. , Riviere, M.‐P. , Liorente, F. , … Molina, A. (2012). Disruption of abscisic acid signalling constitutively activates Arabidopsis resistance to the necrotrophic fungus Plectosphaerella cucumerina . Plant Physiology, 160, 2109–2124. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Santiago, J. , Dupeux, F. , Betz, K. , Antoni, R. , Gonzalez‐Guzman, M. , Rodriguez, L. , … Rodriguez, P. L. (2012). Structural insights into PYR/PYL/RCAR ABA receptors and PP2Cs. Plant Science, 182, 3–11. [DOI] [PubMed] [Google Scholar]
- Santiago, J. , Dupeux, F. , Round, A. , Antoni, R. , Park, S.‐Y. , Jamin, M. , … Márquez, J. A. (2009). The abscisic acid receptor PYR1 in complex with abscisic acid. Nature, 462, 665–668. [DOI] [PubMed] [Google Scholar]
- Schweighofer, A. , Hirt, H. , & Meskiene, I. (2004). Plant PP2C phosphatases: Emerging functions in stress signaling. Trends in Plant Science, 9, 236–243. [DOI] [PubMed] [Google Scholar]
- Singh, A. , Pandey, A. , Srivastava, A. K. , Tran, L.‐S. P. , & Pandey, G. K. (2015). Plant protein phosphatases 2C: From genomic diversity to functional multiplicity and importance in stress management. Critical Reviews in Biotechnology, 8551, 1–13. [DOI] [PubMed] [Google Scholar]
- Song, S. K. , & Clark, S. E. (2005). POL and related phosphatases are dosage‐sensitive regulators of meristem and organ development in Arabidopsis. Developmental Biology, 285, 272–284. [DOI] [PubMed] [Google Scholar]
- Soon, F.‐F. , Ng, L.‐M. , Zhou, X. E. , West, G. M. , Kovach, A. , Tan, M. H. E. , … Xu, H. E. (2012). Molecular mimicry regulates ABA signaling by SnRK2 kinases and PP2C phosphatases. Science, 335, 85–88. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Spartz, A. K. , Ren, H. , Park, M. Y. , Grandt, K. N. , Lee, S. H. , Murphy, A. S. , … Gray, W. M. (2014). SAUR inhibition of PP2C‐D phosphatases activates plasma membrane H + ‐ATPases to promote cell expansion in Arabidopsis. The Plant Cell, 26, 2129–2142. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Thaler, J. S. , & Bostock, R. M. (2004). Interactions between abscisic‐acid‐mediated responses and plant resistance to pathogens and insects. Ecology, 85, 48–58. [Google Scholar]
- Tian, M. , Caroline, C. V. D. , Liu, P. P. , Friso, G. , Van Wijk, K. J. , & Klessig, D. F. (2012). The combined use of photoaffinity labeling and surface plasmon resonance‐based technology identifies multiple salicylic acid‐binding proteins. Plant Journal, 72, 1027–1038. [DOI] [PubMed] [Google Scholar]
- Tuteja, N. (2007). Abscisic acid and abiotic stress signaling. Plant Signaling & Behavior, 2, 135–138. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Umezawa, T. , Sugiyama, N. , Mizoguchi, M. , Hayashi, S. , Myouga, F. , Yamaguchi‐Shinozaki, K. , … Shinozaki, K. (2009). Type 2C protein phosphatases directly regulate abscisic acid‐activated protein kinases in Arabidopsis. Proceedings of the National Academy of Sciences of the United States of America, 106, 17588–17593. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Uno, Y. , Furihata, T. , Abe, H. , Yoshida, R. , Shinozaki, K. , & Yamaguchi‐Shinozaki, K. (2000). Arabidopsis basic leucine zipper transcription factors involved in an abscisic acid‐dependent signal transduction pathway under drought and high‐salinity conditions. Proceedings of the National Academy of Sciences of the United States of America, 97, 11632–11637. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Vierstra, R. D. (2009). The ubiquitin‐26S proteasome system at the nexus of plant biology. Nature Reviews. Molecular Cell Biology, 10, 385–397. [DOI] [PubMed] [Google Scholar]
- Vlot, A. C. , Dempsey, D. A. , & Klessig, D. F. (2009). Salicylic acid, a multifaceted hormone to combat disease. Annual Review of Phytopathology, 47, 177–206. [DOI] [PubMed] [Google Scholar]
- Ward, E. W. B. , Cahill, D. M. , & Bhattacharyya, M. K. (1989). Abscisic acid suppression of phenylalanine ammonia‐lyase activity and mRNA, and resistance of soybeans to Phytophthora megasperma f.sp. glycinea . Plant Physiology, 91, 23–27. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Widjaja, I. , Lassowskat, I. , Bethke, G. , Eschen‐Lippold, L. , Long, H. H. , Naumann, K. , … Lee, J. (2010). A protein phosphatase 2C, responsive to the bacterial effector AvrRpm1 but not to the AvrB effector, regulates defense responses in Arabidopsis. Plant Journal, 61, 249–258. [DOI] [PubMed] [Google Scholar]
- Wu, Y. , Zhang, D. , Chu, J. Y. , Boyle, P. , Wang, Y. , Brindle, I. D. , … Després, C. (2012). The Arabidopsis NPR1 protein is a receptor for the plant defense hormone salicylic acid. Cell Reports, 1, 639–647. [DOI] [PubMed] [Google Scholar]
- Yasuda, M. , Ishikawa, A. , Jikumaru, Y. , Seki, M. , Umezawa, T. , Asami, T. , … Nakashita, H. (2008). Antagonistic interaction between systemic acquired resistance and the abscisic acid‐mediated abiotic stress response in Arabidopsis. The Plant Cell, 20, 1678–1692. [DOI] [PMC free article] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.
Supplementary Materials