

Abstract
Background:
Early life adversity (ELA) is a risk factor for internalizing psychopathology (IP). ELA is also linked to alterations in neural phenotypes of emotion processing and maladaptive emotion regulatory strategies, such as ruminative brooding, in adulthood. We therefore expected that ELA would predict cortical brain activation to emotional faces in trans-diagnostic IP and in turn, mediate the extent of rumination among patients with IPs and ELA (IP+ELA).
Method:
132 individuals, including 102 treatment-seeking adults with heterogeneous IPs and 30 healthy controls (HCs) performed an emotional face-matching task during fMRI. Whole brain analyses compared HC (n=30), IP (n=52), and IP+ELA (n=50) neural responses to emotional (angry, fearful, happy, and sad) faces versus shapes, controlling for depression and anxiety symptoms. Parameter estimates of activation were extracted for significant between-group differences and tested as a mediator of ruminative brooding in IP+ELA.
Results:
IP+ELA demonstrated increased activation in the superior frontal gyrus and anterior cingulate cortex (fear), superior parietal lobule, precuneus, posterior cingulate, and inferior temporal gyrus (fear only), and cuneus (fear and angry). These regions were preferentially correlated with ruminative brooding in IP+ELA, many of which mediated the link between IP+ELA and ruminative brooding.
Conclusions:
Results provide evidence that ELA history among IP patients augments engagement of brain regions involved in emotion processing, above and beyond what is accounted for by current symptoms. Though longitudinal designs are needed, alterations in the neural correlates of maladaptive processing of socio-emotional information may be a common pathway by which ELA poses risk for psychopathology.
Keywords: early childhood adversity, emotion processing, functional neuroimaging, rumination, anxiety, depression
Introduction
Childhood maltreatment is a damaging form of early life adversity (ELA) and stress broadly defined as the intentional or unintentional commission of acts (e.g. verbal, physical, or sexual abuse) or withholding of resources (emotional or physical neglect) by caregivers that adversely influence the emotional and physical health, growth, or adaptation of a child (Egeland, 2009). ELA is a particularly potent risk factor for many internalizing psychopathologies (IPs) in adulthood, such as depression, posttraumatic stress disorder (PTSD), and other anxiety disorders (Gilbert et al., 2009, Widom et al., 2007). Moreover, IP patients with a history of ELA demonstrate a particularly chronic course of illness and treatment resistance (Arnow, 2004, Wegman and Stetler, 2009). Developing a better understanding of the mechanisms by which ELA may foster a predisposition to poor long-term mental outcomes is paramount for improving targeted prevention and intervention for this vulnerable population.
From a theoretical standpoint, ELA may increase vulnerability to IPs via altered brain function underlying processing and modulation of emotion (McCrory et al., 2017, Teicher et al., 2016). Specifically, this latent vulnerability theory postulates that ELA is thought to enhance neurobiological sensitivity to sources of potential threat or negative valence (Heany et al., 2017, McCrory et al., 2017). In a childhood environment saturated with constant threats to wellbeing, heightened responsiveness to social signals of possible adverse experiences may evolve as an adaptive coping mechanism, initially facilitating early identification and avoidance of potentially harmful interactions with a caregiver (McCrory and Viding, 2015). However, if increased stress susceptibility is instantiated during a malleable period of brain development and endures over time, the success of this compensatory mechanism may break down or become maladaptive with repeated overuse (McCrory and Viding, 2015). It is this chronic “wear and tear” to regulatory and compensatory emotional processes that is suspected to increase vulnerability to emotional dysregulation and mental health problems later in life (Danese and Baldwin, 2017, Dannlowski et al., 2012).
In initial support of this hypothesis, there is an extant literature documenting structural alterations in frontal and limbic brain regions involved in the experience and regulation of emotion, such as the anterior cingulate cortex, medial and dorsolateral prefrontal cortex (Lim et al., 2014, Paquola et al., 2016), hippocampus (Riem et al., 2015), and amygdala (Ahmed-Leitao et al., 2016). Building on these structural findings, task-based functional magnetic resonance imaging (fMRI) has allowed for more direct inferences regarding the neural correlates of emotion processing via the use of socio-affective cues deliberately designed to signal threat, stimulate mentalizing, or elicit negative/positive affective states. One recent meta-analysis summarizing these studies revealed that, in whole brain analyses, adults exposed to childhood maltreatment consistently demonstrated altered activation in the superior frontal gyrus, middle temporal gyrus, hippocampus, and superior parietal lobule across a variety of socio-affective stimuli (Heany et al., 2017). Additionally, in region of interest analyses, amygdala and anterior cingulate hyper-responsivity to socioaffective cues were replicated, but marked by more heterogeneity across studies and sampling design (Heany et al., 2017). These results converge with another meta-analysis of neural response during emotional faces tasks, where across all emotions, the right but not left parahippocampal gyrus and amygdala activation was a correlate of maltreatment in whole brain-studies (Hein and Monk, 2017).
Understanding the clinical significance of these functional brain changes in ELA is limited by the fact that many existing studies are undertaken in healthy individuals or small disease-based case control samples designed around one primary diagnosis of interest (Heany et al., 2017, Hein and Monk, 2017). This approach lays the foundation for possible candidate mechanisms, but reduces explanatory power and fragments conclusions that may cut across psychological disorders, which commonly co-occur (Cuthbert, 2014). An additional caveat to the interpretation is that adults with IPs (Buff et al., 2016, Feldker et al., 2017), as well as those exposed to ELA (Heany et al., 2017, Hein and Monk, 2017), both display alterations of distributed affective corticolimbic networks in response to stimuli conveying threat or negative emotionality. Particularly, there is an extensive literature assessing the neural correlates of facial emotion processing in IPs (Etkin and Wager, 2007, Gentili et al., 2016, Hamilton et al., 2012). Illustratively, our group has previously demonstrated that both depression and anxiety symptoms are linked with activation in paralimbic, cingulate, and lateral prefrontal regions in response to negative facial expressions (MacNamara et al., 2017a). Given this overlap, it is prudent to identify whether internalizing symptoms or ELA are the primary driver of neural alterations associated with emotion processing. If a neural signature specific to individuals with IPs and ELA is identified, this level of precision could promote the early identification of individuals with ELA at highest risk for psychopathology or offer novel intervention targets for this subgroup of patients.
Seeing as ELA constitutes risk for a number of IPs (Gilbert et al., 2009, Widom et al., 2007) and in alignment with the Research Domain Criteria initiative to understand how biologically based constructs explain the core, shared features of diagnostic categories (Cuthbert, 2015), linking neural phenotypes of ELA to patterns of thought or behavior common to IPs is also of substantial clinical utility (Insel, 2014, Sharp et al., 2016). One shared feature of IPs is rumination (Nolen-Hoeksema and Watkins, 2011), a maladaptive cognitive style, defined as the tendency to constantly focus on a negative thought, problem, or mood state and on the possible causes and implications of negative feelings (Smith and Alloy, 2009). As childhood adversity is not often easily discussed and is suppressed (Levy and Anderson, 2008), internal extortion and re-production of ELA events may further increase rumination amongst individuals with IPs (Grierson et al., 2016). In fact, in a non-clinical sample, rumination was associated with depression and anxiety symptoms in individuals with ELA, (Kim et al., 2017) implicating a possible role of rumination in the maintenance of IPs. More specifically, brooding is the component of rumination most strongly associated with IP (Aldao et al., 2010, McLaughlin and Nolen-Hoeksema, 2011, Nolen-Hoeksema et al., 2008) and also involved in the maintenance of symptoms in individuals exposed to ELA and other types of adversity, such as interpersonal trauma (Allbaugh et al., 2016, Raes and Hermans, 2008). At the neural level, it is noteworthy that there is substantial overlap between the functional correlates of ruminative thought patterns and emotion processing in ELA, including in the amygdala, anterior and posterior cingulate, medial and dorsolateral prefrontal cortex, (para-) hippocampus, medial and inferior temporal gyri, and inferior parietal lobule (Burkhouse et al., 2017, Cooney et al., 2010). Additionally, rumination has been linked to activation in visual and somatosensory brain regions, such as the insula, precuneus, and cuneus, which are involved in visual cortical facial emotion processing (Burkhouse et al., 2017). Put together, ELA may heighten cortico-limbic sensitivity to negative emotional cues, in turn, promoting increased attention to symptoms and possible causes of one’s own distress. However, the integration of ELA, neural emotion processing, and rumination has yet to be examined in a clinical sample.
To address these gaps in the literature, we sought to examine the neural correlates of facial emotion processing in a large, heterogeneous, and clinically diverse population of patients with multiple and comorbid IPs, with and without exposure to ELA (IP+ELA), and healthy controls (HC). As ELA shares similar proclivity for depression, mixed anxiety disorders, and PTSD, this approach was intentionally designed to maximize generalizability. We hypothesized that IP+ELA would demonstrate the most pronounced activation in the superior frontal gyrus, lateral temporal gyri, medial temporal lobe, anterior/posterior cingulate cortex, and inferior parietal lobule. We also expected that IP+ELA patients would report a higher degree of ruminative brooding and that enhanced brain activation in these regions would account for (mediate) brooding in IP+ELA.
Method
Participants and Procedures
The current study was designed in-line with, and funded by, the NIMH RDoC initiative (RFA-MH-13–080). The aims of the larger study sought the enrollment of a clinically representative patient population, with a full range of internalizing psychopathology and symptoms, that consented to begin treatment with pharmacotherapy (selective serotonin reuptake inhibitors) or cognitive behavioral therapy. All data used in the current study were collected prior to treatment. Eligibility criteria included: 1) age 18–65; 2) current full threshold DSM-5 depression or anxiety disorder; and 3) total score of ≥23 on the Depression, Anxiety, and Stress Scale [DASS-21; (Lovibond and Lovibond, 1995)], and ≤60 on the Global Assessment of Functioning [GAF; (Jones et al., 1995)]. Exclusion criteria included: 1) inability to provide consent and read and write in English; 2) active medical or neurological problem; 3) history of mania or psychosis; current obsessive compulsive disorder; 4) current substance dependence; 5) intellectual disability; 6) contraindication to SSRIs; 7) ongoing treatment with psychiatric medications or psychotherapy; 8) medication use (psychiatric and other), besides oral contraceptive, within the past four months; 9) history of traumatic brain injury with loss of consciousness; and 10) being pregnant. The University of Illinois at Chicago Institutional Review Board approved this study and informed consent was obtained from all participants.
Participant clinical demographic characteristics are reported in Table 1. Diagnoses were made according to the Structured Clinical Interview for DSM-5 Disorders [SCID-5; (First et al., 2015)] by trained research staff. In-line with the strategy encouraged by RDoC (Kozak and Cuthbert, 2016), individuals were not excluded for comorbid disorders, but instead classified by their clinician-determined principal diagnosis, as determined by the most severe and impairing symptoms (Table 1). Current symptom severity of depression and anxiety based on clinician-administered interviews was evaluated by trained raters using the Hamilton Depression Rating Scale [HAM-D; (Hamilton, 1960)] and Hamilton Anxiety Rating Scale [HAM-A; (Hamilton, 1959)].
Table 1.
Sample Characteristics
|
HC (n = 30) |
IP (n = 52) |
IP + ELA (n = 50) |
Omnibus Test | |||||
|---|---|---|---|---|---|---|---|---|
| M | SD | M | SD | M | SD | F | p-value | |
| Age | 25.00 | 9.98 | 26.73 | 8.89 | 25.94 | 8.34 | .36 | .698 |
| Education | 15.33 | 2.50 | 15.98 | 3.35 | 15.60 | 2.62 | .32 | .733 |
| HAM-Da,b | .50 | .78 | 12.31 | 4.44 | 12.88 | 4.55 | 107.38 | <.001 |
| HAM-Aa,b | .77 | .89 | 18.42 | 6.64 | 17.44 | 5.97 | 110.10 | <.001 |
| CTQ Totala,b,c | 27.30 | 1.91 | 32.15 | 5.50 | 50.38 | 12.31 | 91.55 | <.001 |
| Sexual Abusea,b | 5.00 | .00 | 5.00 | .00 | 7.30 | 5.18 | 8.08 | <.001 |
| Physical Abuseb,c | 5.33 | .71 | 5.67 | 1.10 | 7.44 | 2.87 | 15.15 | <.001 |
| Emotional Abusea,b,c | 5.47 | .73 | 7.27 | 2.20 | 12.60 | 4.51 | 60.33 | <.001 |
| Physical Neglecta,b,c | 5.10 | .31 | 5.98 | 1.42 | 8.52 | 2.91 | 33.37 | <.001 |
| Emotional Neglecta,b,c | 6.40 | 1.61 | 8.23 | 2.76 | 14.52 | 4.00 | 80.98 | <.001 |
| RRS Broodinga,b,c | 5.97 | .93 | 13.08 | 3.69 | 15.32 | 2.82 | 98.29 | <.001 |
| # Comorbid Diagnoses | -- | -- | 2.56 | 1.53 | 2.08 | 1.27 | 2.91 | .088 |
| N | % | N | % | N | % | χ2 | p-value | |
| Sex (% Female)a,b,c | 15 | 50.0 | 35 | 67.3 | 40 | 80.0 | 7.81 | .020 |
| Ethnicity | ||||||||
| Hispanic | 2 | 6.7 | 10 | 19.2 | 10 | 20.0 | 2.81 | .246 |
| Non-Hispanic | 28 | 93.3 | 42 | 80.8 | 40 | 80.0 | 2.81 | |
| Race | ||||||||
| Caucasian | 12 | 40.0 | 36 | 69.2 | 28 | 56.0 | 17.66 | .061 |
| African American | 6 | 20.0 | 5 | 9.6 | 11 | 22.0 | ||
| Asian | 11 | 36.7 | 6 | 11.5 | 6 | 12.0 | ||
| American Indian/Alaskan | 0 | 0.0 | 1 | 1.9 | 2 | 4.0 | ||
| Other/Unknown | 1 | 3.3 | 4 | 7.7 | 3 | 6.0 | ||
| Primary Diagnosis | ||||||||
| Major Depression | -- | -- | 11 | 21.2 | 15 | 30.0 | 3.68 | .597 |
| Dysthymia | -- | -- | 0 | 0.0 | 2 | 4.0 | ||
| Generalized Anxiety | -- | -- | 21 | 40.4 | 18 | 36.0 | ||
| Social Anxiety | -- | -- | 14 | 26.9 | 10 | 20.0 | ||
| Panic Disorder | -- | -- | 3 | 5.8 | 3 | 6.0 | ||
| Post Traumatic Stress | -- | -- | 3 | 5.8 | 2 | 4.0 | ||
HC = Healthy Control; IP = Internalizing Psychopathology Patients; IP+ELA = Internalizing Psychopathology Patients with Early life adversity; HAM-D = Hamilton Depression Rating Scale; HAM-A = Hamilton Anxiety Rating Scale; CTQ = Childhood Trauma Questionnaire; RRS = Rumination Response Scale
Significant (p < .05) post-hoc pairwise comparison, IP vs. HC
Significant (p < .05) post-hoc pairwise comparison, IP+ELA vs. HC
Significant (p < .05) post-hoc pairwise comparison, IP vs. IP+ELA
Assessment of Early Life Adversity
The Childhood Trauma Questionnaire (CTQ) is a 28-item self-report measure that provides brief, reliable, and valid screening for histories of abuse and neglect (Bernstein et al., 1997). It inquires about five subscales of maltreatment - emotional, physical, and sexual abuse, and emotional and physical neglect. The total score of each sub-scale ranges from 5 to 25, thus the total score of CTQ fluctuates from 25 to 125. Although the total score of the CTQ is intended to represent the cumulative severity of childhood adversity exposure, the distribution of the measure is often skewed by the base-rate of childhood adversity (proportion of respondents reporting little to no childhood adversity experiences). In the current sample, the skewness value was 1.52. To adjust for the degree of skewness, existence of childhood adversity exposure can be determined by a cut-off score of each CTQ subscale. Participants who score higher than the threshold of any one subscale are treated as a positive case of ELA. Consistent with previous studies (Bernstein et al., 1997, Bernstein et al., 2003, Bevilacqua et al., 2012, Gibb et al., 2009, Johnson et al., 2011, Kudinova et al., 2015, Lu et al., 2013, Walker et al., 1999), the cutoffs of each subscale for moderate exposure which best differentiate clinically-significant adversity exposure were used: 1) emotional abuse ≥13, 2) emotional neglect ≥15, 3) sexual abuse ≥ 8, 4) physical abuse ≥10, and 5) physical neglect ≥ 10. In this sample, emotional abuse α = .83, physical abuse α = .67, sexual abuse α = .95, emotional neglect α = .91, and physical neglect α = .63. Relatively low Chronbach’s alpha for physical abuse and neglect is likely influenced by particularly high skewness values for these subscales (Sheng and Sheng, 2012).
Within the patient sample, 50 participants (49.0%) had a positive history of ELA based on the cut-off scores, forming a sub-group of IPs with significant early life adversity (IP+ELA). Importantly, we also verified none of the healthy control participants reported significant ELA based on these criteria. The convergent validity of these cut-off scores was also evaluated using an exploratory cluster analysis of the study sample. This was performed with CTQ raw subscale scores as input using Wards method of minimum variance with a squared Euclidean distance measure. Ward’s method is distinct from other methods because it uses an analysis of variance approach to evaluate the distances between clusters. The cluster solution was determined from inspection of the dendrogram. The cases in the resulting cluster solution identified the same three groups, with the subset of n = 50 IP+ELA patients as most similar to each other.
Assessment of Rumination
Self-reported rumination was evaluated with the Ruminative Response Scale [RRS; (Treynor et al., 2003)], which contains subscales representing brooding and reflective pondering components of rumination. As noted earlier, brooding is the component of rumination most strongly associated with IP (Aldao et al., 2010, McLaughlin and Nolen-Hoeksema, 2011, Nolen-Hoeksema et al., 2008) and particularly relevant to the development of IP symptoms in trauma exposed individuals (Raes and Hermans, 2008). The brooding subscale consists of five Likert-type items ranging from 1 (almost never) to 4 (almost always). The measure has demonstrated good internal consistency and 1-year retest reliability (Treynor et al., 2003). In this study, brooding α = 0.87.
Task
Participants completed a version of the Emotional Face-Matching Task (Hariri et al., 2002) previously validated for use with fMRI of blood-oxygen-level dependent (BOLD) signal (Gorka et al., 2015, Hariri et al., 2002, MacNamara et al., 2017b, Phan et al., 2013). Angry, fearful, happy, and sad faces were selected from the Gur emotional faces set (Gur et al., 2002). There were three angry, three fearful, three happy, and three sad blocks of trials, interspersed with shape-matching blocks. Each block lasted 20s and consisted of four back-to-back 5-s trials. Shapes were used as control stimuli instead of neutral faces because they may provide a more truly neutral baseline for comparison, particularly when patients are involved (Filkowski and Haas, 2017).
fMRI Data Acquisition and Processing
fMRI based on BOLD contrast was performed on a three 3T GE (General Electric Healthcare, Waukesha, WI) MR750 scanner at the University of Illinois at Chicago (UIC). Scanning was performed using an eight-channel phased-array radio frequency head coil, using either a gradient-echo echo planar imaging sequence, with the following parameters: repetition time 2s, echo time 25 ms, flip angle 90°, field of view 22 cm, acquisition matrix 64 × 64, 3 mm slices with no gap, 44 axial slices per volume.
All data met our criteria for image quality with minimal motion correction (movements ≤3 mm in any direction across the run). The first four volumes were discarded to allow for the magnetization to reach equilibrium. Statistical Parametric Mapping (SPM8) software (Wellcome Trust Centre for Neuroimaging, London, www.fil.ion.ucl.ac.uk/spm) was used to perform conventional image pre-processing steps. In brief, slice-time correction was performed to account for temporal differences between slice collection order and images were spatially realigned to the first image of the run, normalized to a Montreal Neurological Institute (MNI) template using the EPI template, resampled to 2 mm3 voxels, and smoothed with an 8-mm isotropic Gaussian kernel.
The time series data were subjected to a general linear model, convolved with the canonical hemodynamic response function, and filtered with a 128-s high-pass filter. Because of the heterogeneous IP sample and that we did not have any particular predictions about specific emotional expressions, Angry, Fearful, Happy, Sad, and Shapes conditions were modeled separately, with effects estimated for each voxel for each participant. Individual motion parameters were entered in the model as covariates of no interest. Angry>Shapes, Fearful>Shapes, Sad>Shapes and Happy>Shapes, contrasts, created separately for each participant, were included as random effects for the second level analysis. Each emotion contrast was modeled as a separate dependent variable due to violations of generalized linear model assumptions that occur in repeated measures fMRI designs, particularly when covariates of non-interest are included. Therefore, this approach allowed us to maximize explanatory power by covarying for brain activation related to symptoms of depression and anxiety.
Data Analytic Approach
Clinical Analyses:
Clinical and demographic measures comparing IP+ELA, IP, and HC participants were inspected for normal distribution and analyzed using analysis of variance and chi-square tests, as appropriate.
fMRI Analyses.
Whole-brain group (IP+ELA, IP, and HC) differences were modeled using a one-way ANOVA in SPM8, with follow-up t-contrasts for all possible pairwise differences. Symptoms of depression (HAM-D scores) and anxiety (HAM-A scores) were included in the model as covariates of no interest. To threshold results, we used a whole-brain mask encompassing all gray matter regions excluding cerebellum owing to limited coverage [created with MARINA; (Walter et al., 2003)]. Clusters of activation were identified using an uncorrected voxel threshold of p <.001 and then subjected to correction for multiple comparisons via simulation using 3dClustSim utility (December 16, 2015, updated release; 10,000 iterations; http://afni.nimh.nih.gov/pub/dist/doc/program_help/3dClustSim.html). Given the smoothness estimates of the data, family-wise error (FWE) correction at α<.05 was achieved using a voxel threshold of p<.001, with minimum cluster sizes of 93 (angry>shapes), 91 (fearful>shapes), 76 (sad>shapes), and 92 (happy>shapes) voxels. To clarify the direction of significant between-group differences, we extracted parameter estimates of BOLD signal responses (beta-weights, in arbitrary units of activation) averaged across voxels within a 5-mm radius sphere surrounding each peak maxima.
Mediation Analyses:
After identifying foci of between-group differences in brain activation, a series of mediation analyses tested whether brain activation to emotions versus shapes (extracted using the 5mm-radius sphere surrounding each peak maxima from group contrasts) was, in turn, associated with ruminative brooding in IP+ELA (relative to HC and IP). A separate mediation model was run for each cluster of between group differences in brain activation. Mediation analyses were conducted using the SPSS macro PROCESS (Hayes, 2012), including covariates of no interest for depression (HAM-D) and anxiety (HAM-A) symptoms, age, and sex. Multiple parallel tests were controlled using a false-discovery rate adjusted alpha threshold of p < .036 (Benjamini and Hochberg, 1995).Tests of mediation employed a bootstrapping approach with N = 5,000 bootstrap resamples and a 95% confidence interval to assess indirect effects using PROCESS (Preacher and Hayes, 2004). Bootstrapping is a nonparametric resampling procedure that generates an approximation of the sampling distribution of a statistic from the available data. Sampling distributions of indirect effects are generated by taking a sample (with replacement) of size N from the full data set and calculating the indirect effects (i.e. conducting mediation analyses) in each of the resamples. Thus, the 95% confidence interval represents that of each of those 5,000 resample analyses, 95% of the generated indirect effects fall between the given two estimates.
Results
Demographic and Clinical Characteristics
IP+ELA, (n=50), IP (n=52), and healthy controls (HC, n=30) were equivalent in terms of age, education, and distribution of race/ethnicity (Table 1). IP+ELA, IP, and HC groups did differ in sex distribution, with a greater proportion of females in the IP+ELA group relative to both IP and HC (Table 1). HAM-A scores were higher in IP+ELA (t = -19.38, p <.001) and IP (t = -18.88, p < .001) relative to HC, but IP+ELA and IP did not differ (t = .79, p = .433). HAM-D scores were higher in IP+ELA (t = -18.78, p <.001) and IP (t = -18.67, p <.001 relative to HC, but IP+ELA and IP did not differ (t = -.64, p = .522). IP+ELA reported higher levels of ruminative brooding than IP (t = -3.46, p = .001) and HC participants (t = -21.56, p <.001). Ruminative brooding was also higher in IP relative to HC (t = -13.20, p <.001).
ELA and Brain Activation to Emotional Faces
Overall activation for each emotion condition is presented in Supplemental Figure 1. There were no differences in brain activation between IP and HC groups for any of the emotion contrasts. All findings below pertain to increased activation in IP+ELA relative to IP and HC; there were no significant foci of decreased activation in IP+ELA.
Angry>Shapes: IP+ELA demonstrated greater activation (Figure 1c, Table 2) in the right cuneus [BA 19]).
Figure 1.
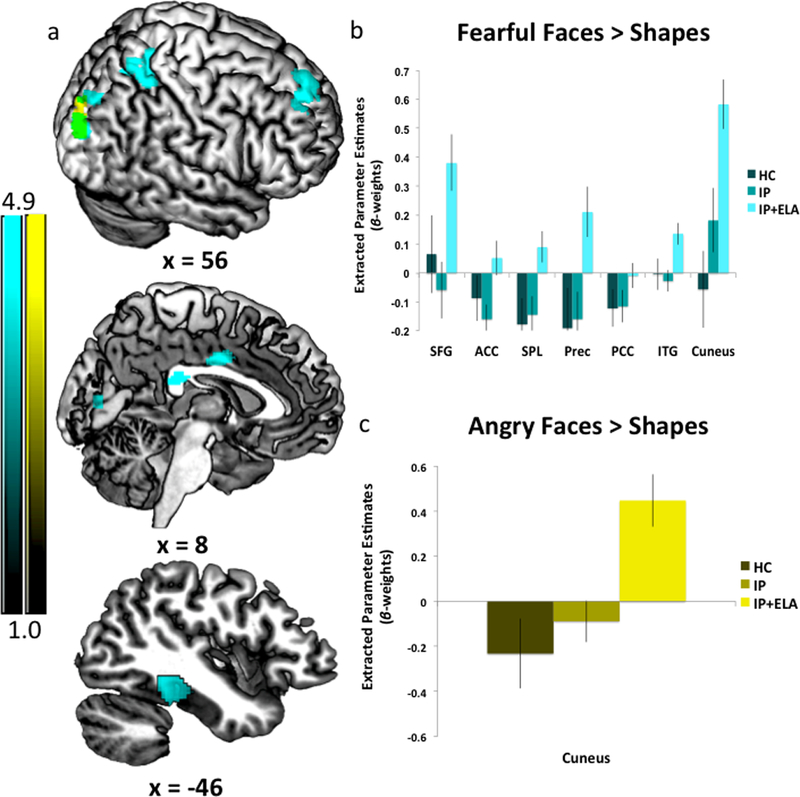
a) Foci of increased neural response for Fearful Faces > Shapes (cyan) and Angry Faces > Shapes (yellow). Areas of anatomical overlap between Fear > Shapes and Angry > Shapes (green). IP+ELA, IP, and HC extracted parameter estimates of activation (beta-weights) for b) Fearful > Shapes, c) Sad > Shapes, and d) Angry > Shapes. SFG = Superior Frontal Gyrus; ACC = Anterior Cingulate Cortex; PCC = Posterior Cingulate Cortex; Prec = Precuneus; SPL = Superior Parietal Lobule; ITG = Inferior Temporal Gyrus
Table 2.
Whole brain activation during facial emotion processing in IP+ELA relative to IP and HC
| MNI Coordinates | ||||||
|---|---|---|---|---|---|---|
| Contrast/Region^* | BA | cluster k | z-score | x | y | z |
| Angry > Shapes | ||||||
| R Cuneus | 19 | 269 | 4.86 | 30 | −90 | 24 |
| 3.59 | 26 | −90 | 38 | |||
| Fearful > Shapes | ||||||
| R Superior Frontal | 6/8/9 | 448 | 4.17 | 28 | 48 | 40 |
| 3.90 | 10 | 48 | 46 | |||
| 3.18 | 12 | 56 | 32 | |||
| R-L Anterior/mid cingulate | 24 | 150 | 3.79 | 2 | −4 | 36 |
| R Superior Parietal | 7 | 678 | 4.01 | 22 | −48 | 60 |
| 3.82 | 38 | −48 | 60 | |||
| 3.81 | 18 | −62 | 52 | |||
| R Precuneus | 7 | 194 | 3.66 | 28 | −74 | 40 |
| 3.32 | 22 | −90 | 38 | |||
| R Posterior Cingulate | 23 | 104 | 3.54 | 4 | −32 | 22 |
| 3.23 | 6 | −24 | 24 | |||
| L Inferior Temporal | 20/36/37 | 219 | 3.69 | −42 | −34 | −20 |
| 3.48 | −48 | −22 | −22 | |||
| R Cuneus | 17/18/19 | 576 | 4.73 | 28 | −90 | 24 |
| 3.82 | 12 | −84 | 6 | |||
| 3.45 | 16 | −88 | 16 | |||
| Happy > Shapes | ||||||
| No significant findings | ||||||
| Sad > Shapes | ||||||
| No significant findings | ||||||
MNI = Montreal Neurological Institute; BA = Brodmann’s area; k = number of contiguous voxels; L= Left; R = Right;
Results are unchanged when re-performed without HAM-D and HAM-A as covariates. Results are also maintained when including sex as a covariate of non-interest in the SPM8 analysis.
Fearful>Shapes: IP+ELA demonstrated greater activation (Figure 1a, Table 2) in the right superior frontal gyrus (encompassing aspects of the dorsolateral and dorsomedial prefrontal cortex; [BA 6, 8, 9]), bilateral anterior/mid cingulate [BA 24], and right posterior cingulate [BA 23]. Greater activation was also observed in the left temporal lobe [BA 20, 36, 37], including the inferior temporal, fusiform, and parahippocampal gyri, right precuneus [BA 7], right superior parietal lobule [BA 7], and right cuneus/lingual gyrus [BA 17, 18, 19).
Sad>Shapes: No significant findings.
Happy>Shapes: No significant findings.
A post-hoc multivariate general linear model of extracted peak BOLD signal indicated there were no significant main effects of sex [F(11, 132) .14, p = .999] or sex by group interactions [F(22, 132) = .978, p = .494] in relation to BOLD activation of any of these clusters. Findings were also maintained when including sex as a covariate of non-interest in the SPM8 analysis.
Brain Activation to Emotional Faces as a Mediator of Rumination in IP+ELA
Table 3 reports BOLD response to emotional faces in relation to ruminative brooding
Table 3.
Mediation analyses evaluating brain activation to fearful and angry faces as a mechanism of increased rumination in IP+ELA
|
Path Ba,b,c, d Relative Direct Effects of Brain Activation on Ruminative Brooding |
Path Ca,e Brain Activation as a Mediator of Ruminative Brooding |
|||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Contrast | β | SE | p | LLCI | ULCI | R2 | Effect |
SE
(boot) |
LLCI | ULCI | ||
| Angry > Shapes | R Cuneus | R Cuneus | ||||||||||
| 1) | Cuneus | .87 | .36 | .018 | 0.15 | 1.58 | .17 | Full Sample | .08 | .06 | 0.009 | 0.27 |
| 2) | Cuneus x IP+ELA | 2.06 | .59 | <.001 | 0.88 | 3.23 | IP+ELA | .46 | .25 | 0.08 | 1.09 | |
| Fearful > Shapes | R SpFrnt | R SpFrnt | ||||||||||
| 1) | SpFrnt | .72 | .40 | .076 | −0.08 | 1.51 | .15 | Full Sample | .06 | .05 | 0.003 | 0.19 |
| 2) | SpFrnt x IP+ELA | 2.12 | .59 | <.001 | 1.03 | 3.37 | IP+ELA | .31 | .18 | 0.03 | 0.74 | |
| R-L ACC | R-L ACC | |||||||||||
| 1) | ACC | .96 | .71 | .176 | −0.44 | 2.35 | .14 | Full Sample | .04 | .07 | −0.01 | 0.28 |
| 2) | ACC x IP+ELA | 2.32 | .58 | <.001 | 1.16 | 3.48 | IP+ELA | .20 | .17 | −0.04 | 0.64 | |
| R PCC | R PCC | |||||||||||
| 1) | PCC | 1.32 | .75 | .081 | −0.16 | 2.80 | .15 | Full Sample | .02 | .04 | −0.01 | 0.16 |
| 2) | PCC x IP+ELA | 2.36 | .57 | <.001 | 1.23 | 3.49 | IP+ELA | .15 | .12 | −0.01 | 0.16 | |
| R Prec | R Prec | |||||||||||
| 1) | Prec | 1.07 | .43 | .014 | 0.22 | 1.91 | .17 | Full Sample | .08 | .06 | 0.01 | 0.23 |
| 2) | Prec x IP+ELA | 2.12 | .58 | <.001 | 0.95 | 3.26 | IP+ELA | .41 | .19 | 0.12 | 0.87 | |
| R SPL | R SPL | |||||||||||
| 1) | SPL | 1.79 | .65 | .006 | 0.51 | 3.07 | .18 | Full Sample | .12 | .08 | 0.01 | 0.35 |
| 2) | SPL x IP+ELA | 2.09 | .58 | <.001 | 0.94 | 3.23 | IP+ELA | .47 | .24 | 0.12 | 1.12 | |
| R Cuneus | R Cuneus | |||||||||||
| 1) | Cuneus | .76 | .39 | .058 | −0.03 | 1.55 | .17 | Full Sample | .06 | .05 | −0.001 | 0.21 |
| 2) | Cuneus x IP+ELA | 2.74 | .75 | <.001 | 1.25 | 4.24 | IP+ELA | .43 | .20 | 0.13 | 0.93 | |
| L ITG | L ITG | |||||||||||
| 1) | ITG | 1.72 | 1.02 | .094 | −0.29 | 3.73 | .15 | Full Sample | .12 | .11 | −0.01 | 0.45 |
| 2) | ITG x IP+ELA | 2.24 | .59 | <.001 | 1.08 | 3.41 | IP+ELA | .27 | .20 | −0.03 | 0.80 | |
Covariates: age, sex, HAM-D, HAM-A
Estimates 1) association between extracted parameter estimates of brain activation and brooding across the entire sample and 2) association between extracted brain activation and brooding in IP+ELA, with HC+IP as a combined reference group. There are no significant associations between brain activation and brooding in IP alone with HC as the reference group.
All correlations between extracted parameter estimates of brain activation and brooding are non-significant in HC.
Exploratory, post-hoc analyses also evaluated whether brain activation to fearful and angry faces was significantly related to the other component of rumination, self-reflection; there were no significant associations.
Estimates 1) mediation in the full sample and 2) mediation in IP+ELA relative to HC+IP. There are no significant tests of mediation in IP alone with HC as the reference group.
SE = Standard Error; LLCI = Lower Limit Confidence Interval; ULCI = Upper Limit Confidence Interval; R = Right; L = Left; SpFrnt = Superior Frontal Gyrus; ACC = Anterior Cingulate Cortex; PCC = Posterior Cingulate Cortex; Prec = Precuneus; SPL = Superior Parietal Lobule; ITG = Inferior Temporal Gyrus;
(Path B: Illustratively, Figure 2) and tests of brain activation to emotional faces as a mediator of ruminative brooding in IP+ELA participants (Path C: For a conceptual model, Figure 3). Eight separate mediation models were conducted (1 for angry > shapes, 7 for fearful > shapes).
Figure 2.
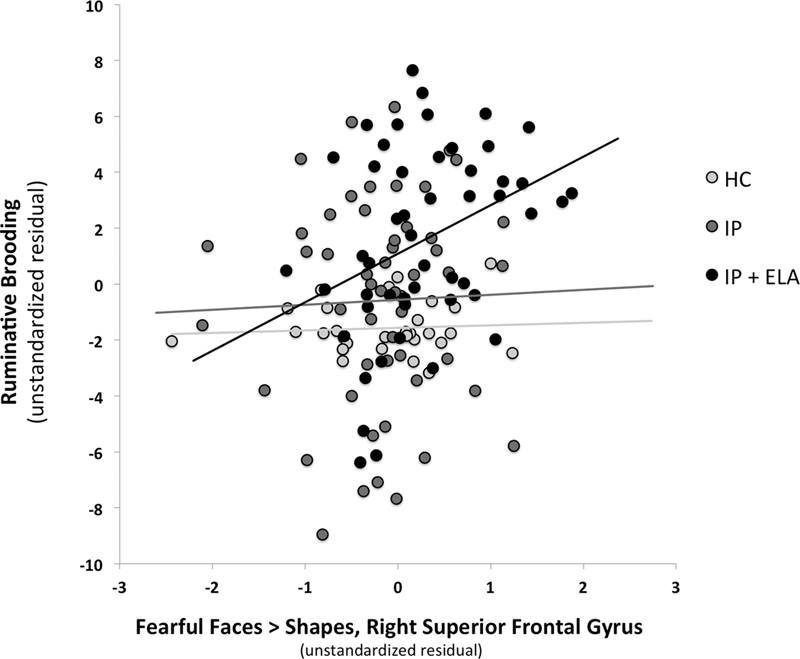
Scatterplot of association between fear-related brain activation in the right superior frontal gyrus and ruminative brooding (as unstandardized residuals after controlling for covariates). HC = Healthy Control; IP = Internalizing Psychopathology Patients; IP+ELA = Internalizing Psychopathology Patients with Early Life Adversity history.
Figure 3.
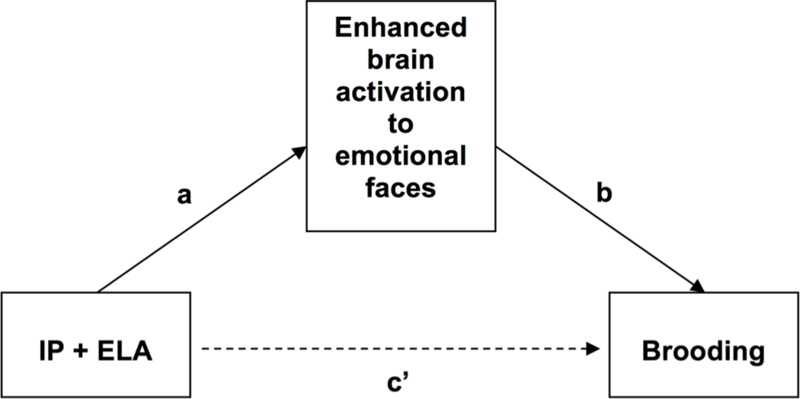
Conceptual mediation model evaluating enhanced brain activation to emotional faces as a mediator of increased ruminative brooding in IP+ELA. IP+ELA = Internalizing Psychopathology Patients with Early Life Adversity history.
Angry>Shapes: The indirect pathway (test of mediation) was significant in IP+ELA for the right cuneus; the total direct effect also remained significant indicating partial mediation.
Fearful>Shapes: Indirect pathways were significant in IP+ELA for the right superior frontal gyrus, right precuneus, right superior parietal lobule, and right cuneus. Tests of mediation for the bilateral anterior cingulate, right posterior cingulate, and left inferior temporal gyrus did not yield significant indirect pathways in IP+ELA; the total direct effects also remained significant indicating partial mediation.
Discussion
Recent initiatives underscore the importance of examining neurobiological signatures that span diagnostic boundaries (Cuthbert, 2014, 2015), particularly those that might be amenable to early detection and prevention (Garvey et al., 2016), yet few studies have comprehensively accomplished this task. In this study, we utilized functional neural activation patterns to facial emotions across a large, heterogeneous, and clinically diverse sample of IP diagnoses with and without exposure to ELA and HCs, in effort to understand whether ELA is a correlate of aberrant neural activity relevant to behavioral constructs involved in internalizing psychopathology. Specifically, we tested the hypothesis that enhanced neural activity in response to negative facial emotions would characterize IP+ELA and that these patterns of brain activity would in turn, relate to brooding in IP+ELA. The present findings, which were most pronounced for fearful faces, implicate hyper-activation of superior frontal gyrus, cingulate cortex, inferior temporal gyrus, and the inferior and superior aspects of the parietal and occipital lobes, respectively, as a correlate of ELA in IPs. We speculate that ELA is candidate etiological mechanism of these neural alterations such that the observed brain changes may be due to the effects of ELA exposure on the brain. Moreover, no group differences were observed between HC and IP only, further implicating ELA exposure as a possible driver of cortico-limbic hyperactivity to negative emotions; however, future studies are needed to demonstrate this among an ELA-exposed control group. Interestingly, the aforementioned brain regions were also consistently correlated with brooding rumination, and many of these neural regions mediated the link between IP+ELA and brooding. Taken together, these findings provide preliminary evidence that greater cortico-limbic reactivity to negative affective stimuli may explain the relationship between ELA and rumination in IPs.
On the whole, we observed cortico-limbic responsivity to especially fearful facial expressions, with some key regions that warrant attention. Namely, fearful faces elicited activation of the right middle anterior cingulate and superior frontal gyrus amongst IP+ELA. These findings are broadly in accordance with existing emotional regulatory theories implicating the anterior cingulate in the recognition, experience, and appraisal of emotional and the dorsolateral and medial prefrontal cortex in regulating emotional reactivity (Etkin et al., 2011). However, as there are some opposing findings regarding activation directionality in anxiety versus depression (MacNamara et al., 2017a), our design offers some confidence that enhanced anterior cingulate and dorsolateral prefrontal engagement may transcend internalizing diagnoses and symptoms when ELA is present. Additionally, and consistent with recent meta-analytic work (Heany et al., 2017), we observed increased activation in the left inferior temporal gyrus, including the fusiform gyrus and extending to the parahippocampal gyrus in IP+ELA. This cluster of regions is typically activated by a broad array of social cognitive tasks requiring abstract reasoning and perspective taking (Schurz et al., 2014), implicating their involvement with an excessive focus on negative social judgment. Collectively, these results underscore convergent neurobiological alterations among IP+ELA related to emotion perception and social reasoning, particularly to fearful stimuli.
Processing of fearful faces was also associated with activation in the posterior cingulate, aspects of the medial prefrontal cortex, and superior parietal lobule amongst IP+ELA. Although these regions have not been extensively reported in relation to ELA amongst healthy samples (Dannlowski et al., 2012, Holz et al., 2015), they are linked to rumination (Burkhouse et al., 2017, Cooney et al., 2010) in IPs. The observed correlations with rumination may suggest that individuals with ELA who develop IPs are particularly prone to engage in affect-congruent self-focus when faced with external cues of fear (Waters and Craske, 2016). It is also noteworthy that ELA patients demonstrated greater activation in visual processing and somatosensory areas during fear (and to a lesser extent anger) processing, such as the cuneus and precuneus. Since IP+ELA were more likely to engage in ruminative brooding relative to IPs and HCs, rumination may also be a more elaborative process for these individuals, particularly when confronted with social signals of threat (Burkhouse et al., 2017). That is, IP+ELA may be more likely to ruminate on social signals of threat more extensively and vividly, as supported by superior occipital and inferior parietal recruitment (Vuilleumier and Pourtois, 2007). Indeed, these regions, in addition to the superior frontal gyrus, superior parietal lobule, partially mediated the extent of rumination present amongst IP+ELA patients. Notably, the observed patterns of brain activation accounted for 13–18% of the variance in rumination, consistent with a moderate to large effect size, suggesting that targeting this neurocircuitry among IP+ELA could have measureable effects on maladaptive coping styles known to increase proneness to or persistence of internalizing mental health problems (Kim et al., 2017). Of particular interest, in light of the emotional faces eliciting the neural response is to evaluate whether cognitive therapy emphasizing appraisals of other’s emotional responses and interpretation bias, would have effects on these brain alterations. Additionally, another possibility would be to target regulatory neural circuits (e.g. cognitive control network) with brain stimulation (e.g. trans-cranial magnetic stimulation) as a top-down approach to reduce the observed cortico-limbic reactivity to negative affective stimuli.
There are aspects of the present findings that were inconsistent with our hypotheses. Namely, in contrast to the expectation that IP+ELA would exhibit similar neural alterations to all negative facial emotions, the overwhelming pattern of results showed dominance for fearful faces; ELA-specific neural correlates were surprisingly sparse for angry faces. One possible reason for this discrepancy relates prior work implicating neural sensitivity to angry faces as a mediator of aggressive behavior (Shackman and Pollak, 2014), of which base rates in this internalizing psychopathology sample were very low. It may be the case that maltreated children who tend to respond to threat with reactive aggression are more likely to develop forms of externalizing psychopathology (Lee and Hoaken, 2007) and hyper-vigilance to facial expressions of anger is therefore more relevant in that context. Equally, sensitivity to angry faces is particularly pronounced in victims of physical, as opposed to other forms of abuse (Pollak et al., 2000, Pollak and Tolley-Schell, 2003). Although we were not powered to undertake within- and between-group comparisons of specific types of abuse and neglect, higher levels of emotional than physical maltreatment characterized the current sample and the emotion-specificity of our findings may reflect this variability. On the other hand, we must also consider that cortico-limbic hyper-responsivity to angry faces has nevertheless been reported in both depression and anxiety disorders (MacNamara et al., 2017a); this may be a broader marker of psychological distress that simply is not a specific etiological pathway related to ELA.
Additionally, it was also somewhat surprising that activation in the amygdala was not enhanced in ELA. Amygdala hyperactivity has been reported in association with ELA (McCrory et al., 2012, Teicher et al., 2002) and in the pathophysiology of IPs (Heller, 2016, Shin and Liberzon, 2010); nonetheless, many of these studies are small, single disorder case control designs (Heany et al., 2017, Hein and Monk, 2017). In fact, whole-brain evidence in support of amygdala hyperactivity is actually somewhat inconsistent, identified in certain meta-analyses (Hein and Monk, 2017), but not others (Heany et al., 2017, Hein and Monk, 2017). Both recent meta-analyses identified increased amygdala activation using a region of analysis (ROI) approach, lending credence to the need for small volume correction of this small anatomical region. However, an ROI-driven approach to threshold results can also increase the likelihood of identifying significant areas of activation; thus, amygdala activation may be preferentially present in experiments that opt to report ROI versus whole brain analyses. Alternatively, as there is great heterogeneity in social-affective stimuli and tasks reporting amygdala hyperactivation in ELA and IPs (Etkin and Wager, 2007, Gentili et al., 2016, Hamilton et al., 2012, Heany et al., 2017), we also consider the possibility that faces are less evocative, depersonalized elicitors of emotion for some patients. For instance, facial expressions of others are likely to have less emotional significance compared to a personal narrative of adversity experiences (MacNamara et al., 2017a). Particularly in the case of ELA-exposed IPs, recruitment of the amygdala for emotional learning of facial expressions could be superseded by the salience of ongoing personal fear memories (Clark and Mackay, 2015).
We are also cautious not to over interpret null findings for the amygdala, as amygdala activation in response to the emotional stimuli in this task was present, but across all participants and not specific to the IP or IP+ELA groups (Supplemental Figure 1). Likewise, it is also noteworthy that IP and HC groups did not demonstrate other differences in brain activation. One explanation relates to the fact that most case-control designs have compared single disorders to controls rather than a heterogeneous group of different internalizing diagnoses. Accordingly, the lack of group differences identified could simply represent there are not shared neural alterations across different anxiety disorders and depression diagnoses relative to controls. Alternatively, as demonstrated in our prior work (MacNamara et al., 2017a) and in line with the RDoC initiative, trans-diagnostic neural alterations may be more likely to co-vary with depression and anxiety symptoms dimensionally, rather than discrete diagnostic categories.
There were several limitations to the current study. First, ELA assessment was retrospective by means of a self-report measure. This kind of reporting could be subject to inaccuracies or mood congruent recall, which might be particularly relevant in IP+ELA(Gaddy and Ingram, 2014, Ono et al., 2016, Schonfeld and Ehlers, 2017). Second, while the present analysis benefited from an HC comparison and IP reference group without ELA equivalent in anxiety and depression symptoms, this design could be further strengthened by inclusion of an ELA-exposed HC group. That is, although ELA substantially increases the risk for IPs, not all individuals exposed to ELA develop IPs and this kind of comparison could elucidate key determinants of adaptive coping and resilience (Bowes and Jaffee, 2013, Kim-Cohen and Turkewitz, 2012). Moreover, there may also be unique markers involved the propensity for ELA to develop into other forms of psychopathology, such as substance use (Puetz and McCrory, 2015) or externalizing disorders (Busso et al., 2017), that were not the emphasis of the current study. A third limitation is that our IP sample, and particularly the IP+ELA group, constituted a greater proportion of female participants relative to controls. Although this is reflective of the sex differences that characterizes sensitivity to stress and prevalence of internalizing disorders (Bekhbat and Neigh, 2018), these group differences called for careful post-hoc analysis to ensure sex effects did not drive our significant findings (footnote, Table 2). The null results of such post-hoc analyses notwithstanding, additional confidence in these findings would be procured in a design matched on sex across groups. Additionally, the patients in this study were seeking treatment and we do not know if these findings are generalizable to what might be observed in naturalistic, community settings. Last, the present study was cross-sectional; therefore the results are correlational in nature and we are unable to draw definitive conclusions regarding causal effects of ELA on brain function or how any such effects may influence susceptibility to rumination. Although the analytic approach of the current study was informed by theory and prior evidence (Hart and Rubia, 2012, Heany et al., 2017), they are hypothesis generating and future longitudinal designs are needed to establish temporal precedence of these constructs.
To close, this study provides evidence supporting a neural mechanism (i.e., cortico-limbic hyper-responsivity) linking ELA to rumination among adults with current IPs. Findings may suggest that for individuals experiencing childhood adversity, enhanced cortico-limbic reactivity to negative socio-emotional stimuli may enhance vulnerability to psychological characteristics involved in the maintenance of internalizing symptoms, such as rumination. Therefore, targeting aberrant emotion neurocircuitry in IP+ELA, possibly throughout psychological interventions, could be a novel target for modifying ruminative habits known to increase persistence of internalizing mental health problems. These results also help to close the gap between the segregated sampling designs of earlier studies (Hart and Rubia, 2012, Heany et al., 2017), affirming that ELA is associated with trans-diagnostic alterations to cortico-limbic emotional processing, above and beyond what can be accounted for by current internalizing symptoms. This is an important step towards identifying a common pathway to prevalent and co-occurring forms of internalizing psychopathology in adulthood. We hope that the work herein will provide further impetus for the assessment of longitudinal, developmental trajectories of risk and resilience to adversity within systems neuroscience models; it is these models that are most likely to bring the field closer to early detection of etiological factors involved in IP across the lifespan.
Supplementary Material
Acknowledgments
Financial Support
This work was supported by the National Institute of Mental Health (R01-MH101497–04) to KLP. ATP is supported by National Institute of Mental Health T32 MH112485. KLB is supported by National Institute of Mental Health Grant K23-MH113793–01.
Footnotes
Conflict of Interest
None.
References
- Ahmed-Leitao F, Spies G, van den Heuvel L & Seedat S (2016). Hippocampal and amygdala volumes in adults with posttraumatic stress disorder secondary to childhood abuse or maltreatment: A systematic review. Psychiatry Research 256, 33–43. [DOI] [PubMed] [Google Scholar]
- Aldao A, Nolen-Hoeksema S & Schweizer S (2010). Emotion-regulation strategies across psychopathology: A meta-analytic review. Clinical psychology review 30, 217–237. [DOI] [PubMed] [Google Scholar]
- Allbaugh LJ, Wright MO & Folger SF (2016). The role of repetitive thought in determining posttraumatic growth and distress following interpersonal trauma. Anxiety Stress Coping 29, 21–37. [DOI] [PubMed] [Google Scholar]
- Arnow BA (2004). Relationships between childhood maltreatment, adult health and psychiatric outcomes, and medical utilization. Journal of Clinical Psychiatry 65 Suppl 12, 10–5. [PubMed] [Google Scholar]
- Bekhbat M & Neigh GN (2018). Sex differences in the neuro-immune consequences of stress: Focus on depression and anxiety. Brain Behavior and Immunity 67, 1–12. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Benjamini Y & Hochberg Y (1995). Controlling the false discovery rate: a practical and powerful approach to multiple testing. Journal of the royal statistical society. Series B (Methodological), 289–300. [Google Scholar]
- Bernstein DP, Ahluvalia T, Pogge D & Handelsman L (1997). Validity of the Childhood Trauma Questionnaire in an adolescent psychiatric population. Journal of the American Academy of Child & Adolescent Psychiatry 36, 340–348. [DOI] [PubMed] [Google Scholar]
- Bernstein DP, Stein JA, Newcomb MD, Walker E, Pogge D, Ahluvalia T, Stokes J, Handelsman L, Medrano M, Desmond D & Zule W (2003). Development and validation of a brief screening version of the Childhood Trauma Questionnaire. Child Abuse Neglect 27, 169–90. [DOI] [PubMed] [Google Scholar]
- Bevilacqua L, Carli V, Sarchiapone M, George DK, Goldman D, Roy A & Enoch MA (2012). Interaction between FKBP5 and childhood trauma and risk of aggressive behavior. Archives of General Psychiatry 69, 62–70. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Bowes L & Jaffee SR (2013). Biology, genes, and resilience: toward a multidisciplinary approach. Trauma Violence Abuse 14, 195–208. [DOI] [PubMed] [Google Scholar]
- Buff C, Brinkmann L, Neumeister P, Feldker K, Heitmann C, Gathmann B, Andor T & Straube T (2016). Specifically altered brain responses to threat in generalized anxiety disorder relative to social anxiety disorder and panic disorder. Neuroimage Clinical 12, 698–706. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Burkhouse KL, Jacobs RH, Peters AT, Ajilore O, Watkins ER & Langenecker SA (2017). Neural correlates of rumination in adolescents with remitted major depressive disorder and healthy controls. Cognitive Affective & Behavioral Neuroscience 17, 394–405. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Busso DS, McLaughlin KA & Sheridan MA (2017). Dimensions of Adversity, Physiological Reactivity, and Externalizing Psychopathology in Adolescence: Deprivation and Threat. Psychosomatic Medicine 79, 162–171. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Clark IA & Mackay CE (2015). Mental Imagery and Post-Traumatic Stress Disorder: A Neuroimaging and Experimental Psychopathology Approach to Intrusive Memories of Trauma. Frontiers in Psychiatry 6, 104. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Cooney RE, Joormann J, Eugene F, Dennis EL & Gotlib IH (2010). Neural correlates of rumination in depression. Cognitive Affective & Behavioral Neuroscience 10, 470–8. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Cuthbert BN (2014). The RDoC framework: facilitating transition from ICD/DSM to dimensional approaches that integrate neuroscience and psychopathology. World Psychiatry 13, 28–35. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Cuthbert BN (2015). Research Domain Criteria: toward future psychiatric nosologies. Dialogues in Clinical Neuroscience 17, 89–97. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Danese A & Baldwin JR (2017). Hidden Wounds? Inflammatory Links Between Childhood Trauma and Psychopathology. Annual Review of Psychology 68, 517–544. [DOI] [PubMed] [Google Scholar]
- Dannlowski U, Stuhrmann A, Beutelmann V, Zwanzger P, Lenzen T, Grotegerd D, Domschke K, Hohoff C, Ohrmann P, Bauer J, Lindner C, Postert C, Konrad C, Arolt V, Heindel W, Suslow T & Kugel H (2012). Limbic scars: long-term consequences of childhood maltreatment revealed by functional and structural magnetic resonance imaging. Biological Psychiatry 71, 286–93. [DOI] [PubMed] [Google Scholar]
- Egeland B (2009). Taking stock: childhood emotional maltreatment and developmental psychopathology. Child Abuse & Neglect 33, 22–6. [DOI] [PubMed] [Google Scholar]
- Etkin A, Egner T & Kalisch R (2011). Emotional processing in anterior cingulate and medial prefrontal cortex. Trends in Cognitive Science 15, 85–93. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Etkin A & Wager TD (2007). Functional neuroimaging of anxiety: a meta-analysis of emotional processing in PTSD, social anxiety disorder, and specific phobia. American Journal of Psychiatry 164, 1476–88. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Feldker K, Heitmann CY, Neumeister P, Tupak SV, Schrammen E, Moeck R, Zwitserlood P, Bruchmann M & Straube T (2017). Transdiagnostic brain responses to disorder-related threat across four psychiatric disorders. Psychological Medicine 47, 730–743. [DOI] [PubMed] [Google Scholar]
- Filkowski MM & Haas BW (2017). Rethinking the Use of Neutral Faces as a Baseline in fMRI Neuroimaging Studies of Axis‐I Psychiatric Disorders. Journal of Neuroimaging 27, 281–291. [DOI] [PubMed] [Google Scholar]
- First M, Williams J, Karg R & Spitzer R (2015). Structured clinical interview for DSM-5—Research version (SCID-5 for DSM-5, research version; SCID-5-RV). Arlington, VA: American Psychiatric Association. [Google Scholar]
- Gaddy MA & Ingram RE (2014). A meta-analytic review of mood-congruent implicit memory in depressed mood. Clinical Psychology Review 34, 402–16. [DOI] [PubMed] [Google Scholar]
- Garvey M, Avenevoli S & Anderson K (2016). The National Institute of Mental Health Research Domain Criteria and Clinical Research in Child and Adolescent Psychiatry. Journal of the American Academy of Child and Adolescent Psychiatry 55, 93–8. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Gentili C, Cristea IA, Angstadt M, Klumpp H, Tozzi L, Phan KL & Pietrini P (2016). Beyond emotions: A meta-analysis of neural response within face processing system in social anxiety. Experimental Biology & Medicine (Maywood) 241, 225–37. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Gibb BE, Schofield CA & Coles ME (2009). Reported history of childhood abuse and young adults’ information-processing biases for facial displays of emotion. Child Maltreatment 14, 148–56. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Gilbert R, Widom CS, Browne K, Fergusson D, Webb E & Janson S (2009). Burden and consequences of child maltreatment in high-income countries. Lancet 373, 68–81. [DOI] [PubMed] [Google Scholar]
- Gorka SM, Fitzgerald DA, Labuschagne I, Hosanagar A, Wood AG, Nathan PJ & Phan KL (2015). Oxytocin modulation of amygdala functional connectivity to fearful faces in generalized social anxiety disorder. Neuropsychopharmacology 40, 278. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Grierson AB, Hickie IB, Naismith SL & Scott J (2016). The role of rumination in illness trajectories in youth: linking trans-diagnostic processes with clinical staging models. Psychological Medicine 46, 2467–84. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Gur RC, Schroeder L, Turner T, McGrath C, Chan RM, Turetsky BI, Alsop D, Maldjian J & Gur RE (2002). Brain activation during facial emotion processing. Neuroimage 16, 651–662. [DOI] [PubMed] [Google Scholar]
- Hamilton JP, Etkin A, Furman DJ, Lemus MG, Johnson RF & Gotlib IH (2012). Functional neuroimaging of major depressive disorder: a meta-analysis and new integration of base line activation and neural response data. American Journal of Psychiatry 169, 693–703. [DOI] [PubMed] [Google Scholar]
- Hamilton M (1959). The assessment of anxiety states by rating. Psychology and Psychotherapy: Theory, Research and Practice 32, 50–55. [DOI] [PubMed] [Google Scholar]
- Hamilton M (1960). A rating scale for depression. Journal of neurology, neurosurgery, and psychiatry 23, 56. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Hariri AR, Tessitore A, Mattay VS, Fera F & Weinberger DR (2002). The amygdala response to emotional stimuli: a comparison of faces and scenes. Neuroimage 17, 317–323. [DOI] [PubMed] [Google Scholar]
- Hart H & Rubia K (2012). Neuroimaging of child abuse: a critical review. Frontiers in Human Neuroscience 6, 52. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Hayes AF (2012). PROCESS: A versatile computational tool for observed variable mediation, moderation, and conditional process modeling. University of Kansas, KS. [Google Scholar]
- Heany SJ, Groenewold NA, Uhlmann A, Dalvie S, Stein DJ & Brooks SJ (2017). The neural correlates of Childhood Trauma Questionnaire scores in adults: A meta-analysis and review of functional magnetic resonance imaging studies. Developmental Psychopatholy, 1–11. [DOI] [PubMed] [Google Scholar]
- Hein TC & Monk CS (2017). Research Review: Neural response to threat in children, adolescents, and adults after child maltreatment - a quantitative meta-analysis. Journal of Child Psychology and Psychiatry 58, 222–230. [DOI] [PubMed] [Google Scholar]
- Heller AS (2016). Cortical-Subcortical Interactions in Depression: From Animal Models to Human Psychopathology. Frontiers in Systems Neuroscience 10, 20. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Holz NE, Buchmann AF, Boecker R, Blomeyer D, Baumeister S, Wolf I, Rietschel M, Witt SH, Plichta MM, Meyer-Lindenberg A, Banaschewski T, Brandeis D & Laucht M (2015). Role of FKBP5 in emotion processing: results on amygdala activity, connectivity and volume. Brain Structure & Function 220, 1355–68. [DOI] [PubMed] [Google Scholar]
- Insel TR (2014). The NIMH Research Domain Criteria (RDoC) Project: precision medicine for psychiatry. American Journal of Psychiatry 171, 395–7. [DOI] [PubMed] [Google Scholar]
- Johnson AL, Benas JS & Gibb BE (2011). Depressive implicit associations and adults’ reports of childhood abuse. Cognition & Emotion 25, 328–33. [DOI] [PubMed] [Google Scholar]
- Jones SH, Thornicroft G, Coffey M & Dunn G (1995). A brief mental health outcome scale-reliability and validity of the Global Assessment of Functioning (GAF). The British Journal of Psychiatry 166, 654–659. [DOI] [PubMed] [Google Scholar]
- Kim JS, Jin MJ, Jung W, Hahn SW & Lee SH (2017). Rumination as a Mediator between Childhood Trauma and Adulthood Depression/Anxiety in Non-clinical Participants. Frontiers in Psychology 8, 1597. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kim-Cohen J & Turkewitz R (2012). Resilience and measured gene-environment interactions. Developmental Psychopathol 24, 1297–306. [DOI] [PubMed] [Google Scholar]
- Kozak MJ & Cuthbert BN (2016). The NIMH research domain criteria initiative: background, issues, and pragmatics. Psychophysiology 53, 286–297. [DOI] [PubMed] [Google Scholar]
- Kudinova AY, Gibb BE, McGeary JE & Knopik VS (2015). Brain derived neurotrophic factor (BDNF) polymorphism moderates the interactive effect of 5-HTTLPR polymorphism and childhood abuse on diagnoses of major depression in women. Psychiatry Research 225, 746–7. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Lee V & Hoaken PN (2007). Cognition, emotion, and neurobiological development: mediating the relation between maltreatment and aggression. Childhood Maltreatreant 12, 281–98. [DOI] [PubMed] [Google Scholar]
- Levy BJ & Anderson MC (2008). Individual differences in the suppression of unwanted memories: the executive deficit hypothesis. Acta Psychologica (Amst) 127, 623–35. [DOI] [PubMed] [Google Scholar]
- Lim L, Radua J & Rubia K (2014). Gray matter abnormalities in childhood maltreatment: a voxel-wise meta-analysis. American Journal of Psychiatry 171, 854–63. [DOI] [PubMed] [Google Scholar]
- Lovibond PF & Lovibond SH (1995). The structure of negative emotional states: Comparison of the Depression Anxiety Stress Scales (DASS) with the Beck Depression and Anxiety Inventories. Behaviour research and therapy 33, 335–343. [DOI] [PubMed] [Google Scholar]
- Lu S, Peng H, Wang L, Vasish S, Zhang Y, Gao W, Wu W, Liao M, Wang M, Tang H, Li W, Li W, Li Z, Zhou J, Zhang Z & Li L (2013). Elevated specific peripheral cytokines found in major depressive disorder patients with childhood trauma exposure: a cytokine antibody array analysis. Comprehensive Psychiatry 54, 953–61. [DOI] [PubMed] [Google Scholar]
- MacNamara A, Klumpp H, Kennedy AE, Langenecker SA & Phan KL (2017a). Transdiagnostic neural correlates of affective face processing in anxiety and depression. Depression and Anxiety 34, 621–631. [DOI] [PMC free article] [PubMed] [Google Scholar]
- MacNamara A, Klumpp H, Kennedy AE, Langenecker SA & Phan KL (2017b). Transdiagnostic neural correlates of affective face processing in anxiety and depression. Depression and anxiety 34, 621–631. [DOI] [PMC free article] [PubMed] [Google Scholar]
- McCrory E, De Brito SA & Viding E (2012). The link between child abuse and psychopathology: a review of neurobiological and genetic research. Journal of the Royal Society of Medicine 105, 151–6. [DOI] [PMC free article] [PubMed] [Google Scholar]
- McCrory EJ, Gerin MI & Viding E (2017). Annual Research Review: Childhood maltreatment, latent vulnerability and the shift to preventative psychiatry - the contribution of functional brain imaging. Journal of Child Psychology and Psychiatry 58, 338–357. [DOI] [PMC free article] [PubMed] [Google Scholar]
- McCrory EJ & Viding E (2015). The theory of latent vulnerability: Reconceptualizing the link between childhood maltreatment and psychiatric disorder. Developmental Psychopathology 27, 493–505. [DOI] [PubMed] [Google Scholar]
- McLaughlin KA & Nolen-Hoeksema S (2011). Rumination as a transdiagnostic factor in depression and anxiety. Behaviour research and therapy 49, 186–193. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Nolen-Hoeksema S & Watkins ER (2011). A Heuristic for Developing Transdiagnostic Models of Psychopathology: Explaining Multifinality and Divergent Trajectories. Perspectives on Psycholigcal Science 6, 589–609. [DOI] [PubMed] [Google Scholar]
- Nolen-Hoeksema S, Wisco BE & Lyubomirsky S (2008). Rethinking rumination. Perspectives on psychological science 3, 400–424. [DOI] [PubMed] [Google Scholar]
- Ono M, Devilly GJ & Shum DH (2016). A meta-analytic review of overgeneral memory: The role of trauma history, mood, and the presence of posttraumatic stress disorder. Psychological Trauma 8, 157–64. [DOI] [PubMed] [Google Scholar]
- Paquola C, Bennett MR & Lagopoulos J (2016). Understanding heterogeneity in grey matter research of adults with childhood maltreatment-A meta-analysis and review. Neuroscience Biobehavioral Reviews 69, 299–312. [DOI] [PubMed] [Google Scholar]
- Phan KL, Coccaro EF, Angstadt M, Kreger KJ, Mayberg HS, Liberzon I & Stein MB (2013). Corticolimbic brain reactivity to social signals of threat before and after sertraline treatment in generalized social phobia. Biological Psychiatry 73, 329–336. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Pollak SD, Cicchetti D, Hornung K & Reed A (2000). Recognizing emotion in faces: developmental effects of child abuse and neglect. Developmental Psychology 36, 679–688. [DOI] [PubMed] [Google Scholar]
- Pollak SD & Tolley-Schell SA (2003). Selective attention to facial emotion in physically abused children. Journal of Abnormal Psychology 112, 323–38. [DOI] [PubMed] [Google Scholar]
- Preacher KJ & Hayes AF (2004). SPSS and SAS procedures for estimating indirect effects in simple mediation models. Behavior research methods, instruments, & computers 36, 717–731. [DOI] [PubMed] [Google Scholar]
- Puetz VB & McCrory E (2015). Exploring the Relationship Between Childhood Maltreatment and Addiction: A Review of the Neurocognitive Evidence. Current Addiction Reports 2, 318–325. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Raes F & Hermans D (2008). On the mediating role of subtypes of rumination in the relationship between childhood emotional abuse and depressed mood: brooding versus reflection. Depression and Anxiety 25, 1067–70. [DOI] [PubMed] [Google Scholar]
- Riem MM, Alink LR, Out D, Van Ijzendoorn MH & Bakermans-Kranenburg MJ (2015). Beating the brain about abuse: Empirical and meta-analytic studies of the association between maltreatment and hippocampal volume across childhood and adolescence. Developmental Psychopathology 27, 507–20. [DOI] [PubMed] [Google Scholar]
- Schonfeld S & Ehlers A (2017). Posttraumatic Stress Disorder and Autobiographical Memories in Everyday Life. Clinical Psychological Science 5, 325–340. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Schurz M, Radua J, Aichhorn M, Richlan F & Perner J (2014). Fractionating theory of mind: a meta-analysis of functional brain imaging studies. Neuroscience Biobehavioral Reviews 42, 9–34. [DOI] [PubMed] [Google Scholar]
- Shackman JE & Pollak SD (2014). Impact of physical maltreatment on the regulation of negative affect and aggression. Developmental Psychopathology 26, 1021–33. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Sharp C, Fowler JC, Salas R, Nielsen D, Allen J, Oldham J, Kosten T, Mathew S, Madan A, Frueh BC & Fonagy P (2016). Operationalizing NIMH Research Domain Criteria (RDoC) in naturalistic clinical settings. Bulletine of the Menninger Clinic 80, 187–212. [DOI] [PubMed] [Google Scholar]
- Sheng Y & Sheng Z (2012). Is coefficient alpha robust to non-normal data? Frontiers in Psychology 3, 34. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Shin LM & Liberzon I (2010). The neurocircuitry of fear, stress, and anxiety disorders. Neuropsychopharmacology 35, 169–91. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Smith JM & Alloy LB (2009). A roadmap to rumination: a review of the definition, assessment, and conceptualization of this multifaceted construct. Clinical Psychology Review 29, 116–28. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Teicher MH, Andersen SL, Polcari A, Anderson CM & Navalta CP (2002). Developmental neurobiology of childhood stress and trauma. Psychiatric Clinics of North America 25, 397–426, vii-viii. [DOI] [PubMed] [Google Scholar]
- Teicher MH, Samson JA, Anderson CM & Ohashi K (2016). The effects of childhood maltreatment on brain structure, function and connectivity. Nature Reviews Neuroscience 17, 652–66. [DOI] [PubMed] [Google Scholar]
- Treynor W, Gonzalez R & Nolen-Hoeksema S (2003). Rumination reconsidered: A psychometric analysis. Cognitive therapy and research 27, 247–259. [Google Scholar]
- Vuilleumier P & Pourtois G (2007). Distributed and interactive brain mechanisms during emotion face perception: evidence from functional neuroimaging. Neuropsychologia 45, 174–94. [DOI] [PubMed] [Google Scholar]
- Walker EA, Unutzer J, Rutter C, Gelfand A, Saunders K, VonKorff M, Koss MP & Katon W (1999). Costs of health care use by women HMO members with a history of childhood abuse and neglect. Archives of General Psychiatry 56, 609–13. [DOI] [PubMed] [Google Scholar]
- Walter B, Blecker C, Kirsch P, Sammer G, Schienle A, Stark R & Vaitl D (2003). MARINA: an easy to use tool for the creation of MAsks for Region of INterest Analyes. Neuroimage 19, e1899. [Google Scholar]
- Waters AM & Craske MG (2016). Towards a cognitive-learning formulation of youth anxiety: A narrative review of theory and evidence and implications for treatment. Clinical Psychology Review 50, 50–66. [DOI] [PubMed] [Google Scholar]
- Wegman HL & Stetler C (2009). A meta-analytic review of the effects of childhood abuse on medical outcomes in adulthood. Psychosomatic Medicine 71, 805–12. [DOI] [PubMed] [Google Scholar]
- Widom CS, DuMont K & Czaja SJ (2007). A prospective investigation of major depressive disorder and comorbidity in abused and neglected children grown up. Archives of General Psychiatry 64, 49–56. [DOI] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.