

Abstract
Rapid point-of-care resistance diagnostics (POC-RD) are a key tool in the fight against antibiotic resistance. By tailoring drug choice to infection genotype, doctors can improve treatment efficacy while limiting costs of inappropriate antibiotic prescription. Here, we combine epidemiological theory and data to assess the potential of resistance diagnostics (RD) innovations in a public health context, as a means to limit or even reverse selection for antibiotic resistance. POC-RD can be used to impose a nonbiological fitness cost on resistant strains by enabling diagnostic-informed treatment and targeted interventions that reduce resistant strains’ opportunities for transmission. We assess this diagnostic-imposed fitness cost in the context of a spectrum of bacterial population biologies and find that POC-RD have a greater potential against obligate pathogens than opportunistic pathogens already subject to selection under “bystander” antibiotic exposure during asymptomatic carriage (e.g., the pneumococcus). We close by generalizing the notion of RD-informed strategies to incorporate carriage surveillance information and illustrate that coupling transmission-control interventions to the discovery of resistant strains in carriage can potentially select against resistance in a broad range of opportunistic pathogens.
Point-of-care resistance diagnostics represent an opportunity to tailor antibiotic treatment protocols to specific bacterial strains. This study shows that conditioning on both point-of-care and carriage diagnostics can produce effective patient care that also selects against resistance.
Introduction
“Because antibiotic resistance occurs as part of a natural evolution process, it can be significantly slowed but not stopped. Therefore, new antibiotics will always be needed to keep up with resistant bacteria.”—CDC, “Antibiotic Resistance Threats in the United States, 2013” [1]
The antimicrobial resistance crisis threatens to undermine key features of modern medicine at great costs in terms of patient morbidity, mortality, and treatment expense [2–6]. Current mainstream antibiotic treatment strategies sow the seeds of their own downfall by strongly selecting for resistant strains, leading some to argue that continual new antibiotic discovery is the only way to stay ahead of a “post-antibiotic future” [1,7,8]. If this bleak vision is correct, there is an urgent need to buy time by extending the lifespan of existing antibiotics while research and development for new ones takes its course. More optimistically, it may be possible to improve how we use existing antibiotics and to implement other control measures so that an endless supply of new antibiotics is not required.
Among a number of innovative approaches to improve antibiotic use [9–15], one of the most promising leverages point-of-care resistance diagnostics (POC-RD) that provide prescribers with a rapid readout of the resistance profile of an infecting organism. POC-RD allow prescribers to choose older, cheaper, and/or narrower-spectrum antibiotics when such drugs are most appropriate for patients, thereby saving newer, more expensive, and/or broader-spectrum antibiotics for situations in which they are really needed and perhaps reducing the intensity of selection for resistance to these drugs [16–18].
Less often considered is a second potential benefit of resistance diagnostics (RD): to enable “search and destroy” (S&D) tactics to combat the most dangerous resistant pathogen strains, such as methicillin-resistant Staphylococcus aureus (MRSA) and carbapenem-resistant Enterobacteriaceae (CRE) [19–24]. S&D strategies aim to identify and then isolate patients who are carrying problematic resistant strains until pathogen clearance can be confirmed. If resistant strains can be rapidly, accurately identified and their transmission curtailed by targeted infection-control measures, then S&D can create a nonbiological diagnostic-imposed fitness cost borne only by targeted resistant strains. However, the magnitude of this fitness cost is hotly debated, especially in the context of MRSA control [25,26], and, in any event, intensive medical interventions such as patient isolation are not a practical or economical option in many circumstances.
In this paper, we ask when it is possible to create net selection against resistance, even when (i) there are no biological fitness costs associated with resistance, (ii) the best available treatment cannot be withheld from any patient, and (iii) all nonantibiotic intervention options are only moderately effective compared with the efficacy of antibiotics on susceptible infections. We define “net selection against resistance” as maintaining the fitness of one or more resistant strains below that of the drug-sensitive strain, so that the frequency of these resistant strains will decline toward zero. This can be accomplished, generally speaking, when diagnostics permit medical personnel to artificially shape the pathogen fitness landscape so that resistant strains are disproportionately disadvantaged. We show that the potential to reduce or even reverse selection on resistance depends on 2 key factors.
Pathogen lifestyle: Is symptomatic disease tightly coupled to transmission (obligate pathogen), or can the pathogen also transmit from an asymptomatic carriage phase (opportunistic pathogen)?
Pan resistance: Are untreatable pan-resistant strains already in circulation?
Table 1 offers a preview of our main findings in a simple setting with 2 equally effective antibiotic treatment options, “drug 1” (first-line treatment to which resistance has already emerged in the target-pathogen population) and “drug 2” (second-line treatment to which resistance may or may not have already emerged).
Table 1. Preview of main findings.
| Lifestyle | Pan-resistant | Key findings |
|---|---|---|
| Simple obligate | No | Net selection against drug 1 resistance is possible. |
| Simple obligate | Yes | Net selection against pan resistance is possible only if either there are substantial biological fitness costs associated with pan resistance or a highly effective infection intervention (“isolation”) is available. |
| Opportunistic | No/Yes | Net selection against drug 1 resistance may be impossible, even if all those with sensitive infection are left untreated and all those with drug 1–resistant infection could be targeted for isolation—unless interventions are also conditioned on asymptomatic carriage RD |
Abbreviation: RD, resistance diagnostics.
Methods
We describe a mathematical model for a single pathogen species (“target pathogen”), with multiple strains having different antibiotic susceptibilities, in which healthcare providers and public health officials (hereafter “providers”) can shape the pathogen fitness landscape by tailoring treatment and transmission control measures informed by RD.
Pathogen strains
Two antibiotics are available to treat infections caused by the target pathogen: drug 1 (first-line treatment that would be prescribed to all patients in the absence of RD) and drug 2 (second-line treatment). Resistance to drug 1 and potentially also to drug 2 has already emerged in the target-pathogen population but not yet reached fixation. In particular, there are up to 4 resistance profiles in circulation: an untreatable “pan-resistant” strain (strain 12), a “drug 1–resistant” strain that remains sensitive to drug 2 (strain 1), a “drug 2–resistant” strain that remains sensitive to drug 1 (strain 2), and a “pan-sensitive” strain that can be effectively treated with either drug (strain 0) (Fig 1A). The model encompasses a spectrum of pathogen lifestyles, from “simple obligate pathogens” (causing disease immediately after colonization; Fig 1A and 1B) to “opportunistic pathogens” (transmitting from an asymptomatic carriage phase as well as during symptomatic disease; Fig 1C).
Fig 1.

Schematic of the obligate/SIS (A, B) and opportunistic/SCIS (C) epidemiological models. Boxes denote proportions of hosts in mutually exclusive states: S for uninfected (susceptible) hosts, I0 for hosts infected with a strain sensitive to both drugs, and I1, I2, and I12 for hosts infected with strains resistant to drugs 1, 2, or both 1 and 2, respectively. In the SCIS model (C, showing only 2 pathogen genotypes for clarity), C0 and C1 denote asymptomatic carriage of sensitive and drug 1–resistant bacteria, respectively, and d is the rate at which disease develops from carriage (when d→∞, we recover an SIS model). Box colors denote distinct clinical presentations in the absence (A) or presence (B, C) of multidrug POC-RD. Solid arrows represent flows of individuals between states, and dashed arrows represent factors influencing those flows (e.g., antibiotic treatment). Gray and black arrows denote transmission and clearance, respectively. Equations describing the system are in S1B Text. POC-RD, point-of-care resistance diagnostics; SCIS, Susceptible-Carriage-Infected-Susceptible; SIS, Susceptible-Infected-Susceptible.
RD-informed treatment and control
We use standard reproduction-number analysis to investigate the pathogen-fitness impact of RD-informed treatment and control, depending on pathogen lifestyle (obligate versus opportunistic), what sort of RD is available, and what sort of transmission-control options can be feasibly targeted against each resistant strain, once identified. Cases considered include POC-RD with an obligate pathogen (Case 1; Fig 1B), POC-RD with an opportunistic pathogen (Case 2; Fig 1C), and carriage resistance surveillance (“carriage RD”) with an opportunistic pathogen (Case 3). See the Supporting information for omitted mathematical details as well as several extensions, including to settings with intermediate resistance, public health interventions aimed at discovering drug-resistant infections more quickly, resistance-conferring mutation, host migration, and diagnostic escape. Table 2 summarizes key notation.
Table 2. Notation.
| Notation | Details |
|---|---|
| Pathogen strains and pathogen lifestyle parameters | |
| x | Drugs x = 1,2 are available to treat infections caused by the target pathogen. |
| X | Each strain is named for the drugs (if any) X = 0, 1, 2, 12 to which it is resistant: strain 0 is susceptible to both drugs (“pan-sensitive”), strain 1 is resistant only to drug 1, strain 2 is resistant only to drug 2, and strain 12 is resistant to both drugs (“pan-resistant”). |
| βC,βI>0 | Transmission rates during carriage and during infection, if uncontrolled and if no biological fitness costs |
| fX≥0 | Biological fitness cost associated with strain X resistance, reducing transmission during carriage and infection by a factor of 1−fX |
| γC>0 | Baseline carriage clearance rate |
| Carriage clearance rate of drug x–sensitive strain due to bystander exposure to drug x | |
| d>0 | Rate at which infection develops from carriage (in the limit d→∞, we recover the SIS case) |
| Baseline infection clearance rate when untreated or ineffectively treated | |
| Diagnosis, treatment, and transmission control parameters | |
| D≥0 | Diagnostic delay, i.e., time from sample collection to result |
| rX≥0 | Rate at which resistant strain X is discovered while in the carriage state |
| Infection clearance rate of drug x–sensitive strain when treated with drug x | |
| Proportional reduction in transmission during infection () or during asymptomatic carriage () due to HTCX | |
| Key derived variables | |
| R0,X | Basic reproduction number of strain X |
| Threshold biological fitness cost for strain X not to enjoy a reproductive advantage, i.e., R0,X>R0,0 if and R0,X<R0,0 if | |
Abbreviations: HTCX, heightened transmission control measures targeted against strain X; SIS, standard Susceptible-Infected.
Results
Case 1: POC-RD and a simple obligate pathogen (SIS model)
In the limiting case when the target pathogen immediately causes disease (d = ∞), what we call a “simple obligate pathogen,” our Susceptible-Carriage-Infected-Susceptible (SCIS) epidemiological model reduces to a standard Susceptible-Infected-Susceptible (SIS) model [27–29]. Few if any real-world bacterial pathogens fit this case, but it is useful conceptually as a limiting case (Fig 1A and 1B).
Fig 2 illustrates the impact of RD-informed treatment and transmission control on whether the drug 1–resistant and/or pan-resistant strains enjoy a reproductive advantage relative to the pan-sensitive strain, for a generic pathogen with an average duration of infection of 5 days (with effective treatment) or 10 days (without treatment). The blue parameter regions are where the drug-susceptible strain has a higher reproduction number (R0) than the drug 1–resistant strain and/or the pan-resistant strain, as a function of diagnostic delay D and the biological fitness costs f1, f12 of drug 1 and pan-resistance, thereby creating net selection against these resistant strains. R0 expressions underlying Fig 2 are defined in Box 1, with further details in S1B Text.
Fig 2. Rapid RD enable conditional treatment and infection control strategies that can select against resistance for obligate pathogens even with no biological costs of resistance.
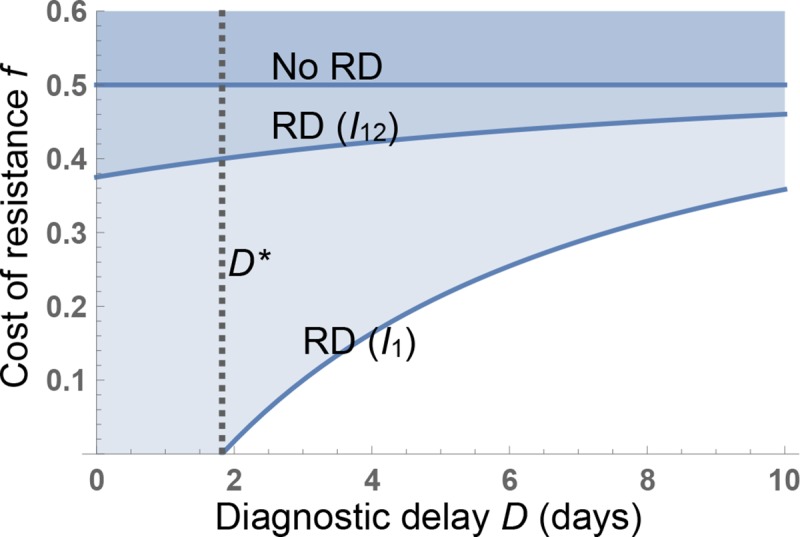
The minimal cost of resistance f*(D) that allows universal treatment without causing an increase in resistance is plotted (contour lines) against diagnostic delay D. The dashed vertical line indicates the longest diagnostic delay (D*) given which there is selection against drug 1 resistance while treating all cases. Three scenarios are shown: RD not available (No RD, contour plot of ); RD control of I1 only (RD (I1), contour plot of ); and RD control of I12 (and trivially, I1) (RD (I12), contour plot of ). Parameters (rates per day): , , . RD, resistance diagnostics.
Box 1. Reproductive numbers (R0) for strains in the SIS model
See S1B Text for more detailed derivations.
Pan-sensitive strain: Strain 0 infections are treated with drug 1 and subjected to standard transmission control, whether or not RD is available. Because strain 0 infections clear at rate under drug 1 treatment and strain 0 transmits at rate βI under standard control, strain 0’s reproduction number is
| (1) |
Drug 1–resistant strain: Because strain 1 infections clear at rate during the diagnostic delay period, a patient will remain infected long enough for RD results to become available (triggering drug 2 and transmission control) with probability . Depending on diagnostic delay D and fitness cost f1, strain 1’s reproduction number takes the form
| (2) |
Drug 2–resistant strain: Because strain 2 infections are susceptible to drug 1 and thus clear at rate during the diagnostic delay period, such infections will be diagnosed before clearance with probability . Strain 2’s resulting reproduction number is
| (3) |
Pan-resistant strain: Strain 12 infections clear at the untreated rate during and after the diagnostic delay period. However, diagnostic results (if available before infection clearance, probability ) can trigger heightened transmission control (HTC), reducing strain 12’s reproduction number to
| (4) |
The solid contours in Fig 2 illustrate the critical threshold resistance costs for the drug 1–resistant strain () and the pan-resistant strain (), the smallest fitness costs that allow universal treatment while keeping the resistant strain at a reproductive disadvantage relative to the sensitive strain. The introduction of RD has the greatest impact on the drug 1–resistant strain (Fig 2, strain I1). In the limit of no diagnostic delay (D = 0), we find that . This implies that resistance to drug 1 (given available drug 2) can be selected against even in the absence of resistance costs () so long as either drug 2 is more effective than drug 1 or drug 2 is less effective than drug 1 but transmission control is sufficiently effective that .
Control of a circulating pan-resistant strain I12 is more challenging, even with POC-RD. Absent fitness costs (f12 = 0), net selection against pan-resistance can be maintained only if , meaning that transmission control of the pan-resistant strain is more effective than drug 1 treatment at speeding clearance. In the case of more modest transmission control (Fig 2; ), net selection against pan-resistance requires that there be sufficient fitness costs associated with pan-resistance. (In the POC-RD limit, when D = 0, the threshold fitness cost .)
Both thresholds and are increasing in D, because longer delays reduce the effectiveness of diagnostic-informed treatment and control, and converge in the D→∞ limit to the “no-RD fitness-cost threshold” absent any RD: . In Fig 2 with baseline clearance and drug 1–assisted clearance , the no-RD fitness-cost threshold equals 0.5, greatly exceeding typical reported costs of resistance. To give some context on empirical estimates of costs of resistance, f, a recent meta-analysis estimated f = 0.21 (±0.024) for chromosomally encoded resistance and f = 0.09 (±0.024) for plasmid-encoded resistance [30], albeit using growth rate rather than epidemiological transmission measures of fitness effects.
Case 2: An opportunistic pathogen with a carrier state (SCIS model)
Many disease-causing bacteria are opportunistic pathogens capable of transmission from an asymptomatic carriage state, while living harmlessly in a host microbial compartment such as the gut or the nasopharynx. Such pathogens face “bystander exposure” to antibiotics used to treat infections caused by other pathogens or to treat noninfectious conditions [31]. Take, for example, the pneumococcus (Streptococcus pneumoniae), one of the top bacterial causes of death globally [32] and a leading cause of antibiotic prescription. Despite the severe burden of disease, the pneumococcus is subjected in the United States to an estimated 9.1 times more courses of any antibiotic during asymptomatic carriage than during disease. Fig 3 compares the volume of bystander selection to target antibiotic exposure for several major bacterial pathogens. An alternate visualization of the proportions of bystander exposure (absent Clostridioides difficile) is presented in Tedijanto and colleagues [31].
Fig 3. Incidental antibiotic exposure during asymptomatic carriage exceeds disease-related antibiotic exposure for key human pathogens.
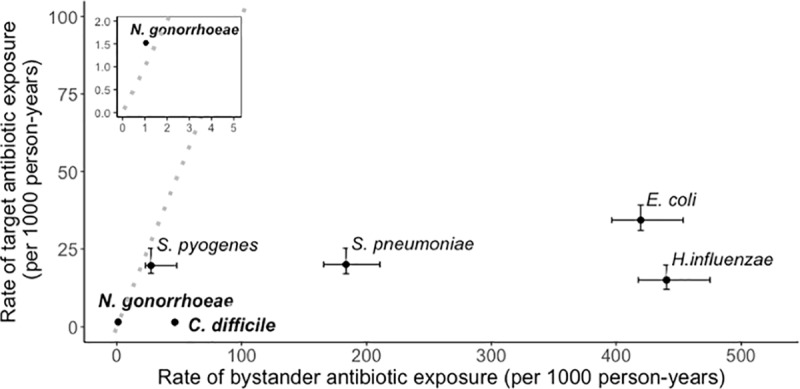
Bold font: Tier 1 urgent resistance concerns according to the CDC [1]. Standard font: The most frequent etiologic agents of the top indications for antibiotic prescription in United States ambulatory care. “Target antibiotic exposure” is defined as any antibiotic use associated with disease caused by that organism; “bystander antibiotic exposure” refers to the incidence of antibiotic exposure in asymptomatic carriage, roughly calculated as the product of the incidence of antibiotic prescription in ambulatory care and the proportion of the population in the relevant age group that carries the bacterium minus the number of target antibiotic exposures. The dotted line is where incidence of antibiotic exposure in carriage is equal to incidence of antibiotic exposure due to disease. See Tedijanto and colleagues [31] for method details, source references, and an alternate visualization of the same data on Neisseria gonorrhoeae, Streptococcus pyogenes, S. pneumoniae, Escherichia coli, and Haemophilus influenzae. Values for Clostridioides difficile were calculated using the same methodology and additional sources for disease incidence [33] and carriage prevalence [34]; see S1F Text for details. CDC, Centers for Disease Control and Prevention.
An implication of our analysis is that, for pathogens like S. pneumoniae, Escherichia coli, Haemophilus influenzae, and C. difficile that overwhelmingly face bystander exposure to antibiotics, even the strongest possible medical interventions informed by POC-RD can be insufficient to halt the rise of resistance to any drug in routine use. Mathematical details underpinning this analysis, including derivations of each strain’s reproduction number with and without POC-RD, are provided in S1C Text.
In Fig 4, we parameterize our SCIS model for the pneumococcus to show the threshold strain 1 fitness cost (contour lines) for a population with substantial bystander exposure (the I1 resistant strain is counterselected in the blue regions above the contour lines). Pneumococcal serotypes show considerable variation in carriage duration (Fig 4, arrows on x axis) [35], and our analysis illustrates that bystander selection becomes increasingly problematic for longer carriage serotypes—represented by a higher threshold resistance cost to balance selection on resistance. Our parameterized model indicates that the threshold costs even in the absence of RD are of a similar magnitude to average costs of plasmid-encoded resistance discussed earlier [30]. However, more problematic is the common observation of cost-free resistances (including single nucleotide polymorphisms [SNPs]) in the pneumococcus [36,37], indicating that these average costs are liable to decrease in response to selection.
Fig 4. POC-RD alone cannot reverse selection on cost-free pneumococcal resistance.

The minimal cost of resistance () that allows universal treatment without causing an increase in strain 1 resistance is plotted (contour lines) against the expected duration of carriage. Two POC-RD scenarios are shown: with () and without () transmission control. Arrows on the x axis are serotype-specific mean carriage duration estimates from [35] (serotypes with ≥15 recorded carriage episodes only). The remaining parameters (rates per day) are d = 0.001, = 5 × 10−4, = 0, = 1, = 0.125. We make the simplifying assumption that baseline carriage and infection transmission rates are identical (βC = βI = β) which ensures that does not depend on β. Details on parameterization are in S1G Text. POC-RD, point-of-care resistance diagnostics.
Even in the absence of pan-resistant strains, POC-RD have a weak impact due to the rarity and brevity of infection events (median infection duration in the absence of treatment is 8 days) relative to long periods of carriage and associated bystander selection. As a result, we anticipate ongoing selection for low-cost resistant strains, with or without POC-RD–informed strategies (Fig 4).
Case 3: Carriage RD
Our pessimistic conclusion concerning the public health merits of POC-RD for commensal opportunists such as S. pneumoniae (Figs 3 and 4) is based on the inevitability of bystander selection during prolonged carriage phases—but what if bystander selection could be opposed by public health interventions during carriage? For example, what if resistant-strain carriers could be identified and subjected to transmission control interventions even when they do not have active infection? Consider the South Swedish Pneumococcal Intervention Project (SSPIP) [38,39], a public health intervention launched in January 1995 in Malmöhus County, Sweden, that aimed to reduce penicillin-resistant pneumococcus (PRP) transmission, especially at preschool daycares. Any time a preschool-age child was identified with symptomatic PRP infection, providers would obtain nasopharyngeal cultures from all other children in the same daycare classroom. Children found to be carrying PRP were then required to remain home until subsequent testing proved them to be PRP negative, penalizing PRP strains by reducing their opportunities for transmission from carriage.
In S1C Text, we extend our analysis of the SCIS model to examine the potential of carriage RD-based interventions to generate selection against resistance. A key result is that, in order to select against drug 1 resistance, the drug 1–resistant strain must be discovered while in carriage at a rate r1 that exceeds the rate at which the sensitive strain is cleared from carriage due to bystander exposure to drug 1. In Fig 5, we again parameterize our SCIS model for the pneumococcus as in Fig 4, under 2 extreme scenarios of carriage duration [35]: 20 weeks (Fig 5A) and 2 weeks (Fig 5B). The blue parameter space in Fig 5 highlights the combinations of carriage discovery rate (r1, x axis) and HTC effectiveness against drug 1–resistant bacteria discovered in carriage ( y axis) that lead to a net selection against drug 1 resistance.
Fig 5. POC-RD plus carriage RD can reverse selection on pneumococcal resistance, even for long–carriage-duration serotypes.

The parameter space generating net selection against resistance is plotted in blue as a function of the rate of carriage discovery (r1) and the effectiveness of carriage HTC (). (A) Longest–carriage-duration serotype (6B, median 20 weeks). (B) Shortest–carriage-duration serotypes (1, 4, 5; approximately 2 weeks). In both (A) and (B), 2 POC-RD scenarios are shown: with (f = 0.01) and without (f = 0) biological cost of resistance. The red dashed line represents the probability of strain 1 discovery while in the carriage state (“C1 discovery”), an increasing function of the rate of carriage diagnosis. The remaining parameters (rates per day) are d = 0.001, = 5 × 10−4, = 0, = 1, = 0.125, γC = 0.006 (A), γC = 0.07 (B). We make the simplifying assumption that baseline carriage and infection transmission rates are identical (βI = βC = β), which ensures that the parameter space generating net selection against resistance does not depend on β. Details on parameterization are in S1G Text. The asterisk positions the outcome of an annual intervention with 50% efficacy in reducing C1 transmission. HTC, heightened transmission control; POC-RD, point-of-care resistance diagnostics.
Fig 5A illustrates the most problematic serotype from a POC-RD perspective, due to the dominance of bystander selection. Given the introduction of annual carriage surveillance (ensuring ), our parameterized model predicts that HTC interventions (such as removal of an infant from nursery) would need to reduce strain 1 transmission from carriage by at least 20% in order to select against the drug 1–resistant genotype in the worst case scenario of zero resistance costs (f1 = 0). Epidemiological studies of the impact of childcare attendance on pneumococcal carriage suggest that a 2-fold reduction in transmission due to removal from daycare is not unreasonable [40].
The analysis underlying Fig 5 in S1C Text implicitly assumes that all uninfected hosts are tested for drug 1–resistant carriage at a constant rate. Although this assumption is useful here from an expositional point of view to highlight ideas, we note that such an approach would be inefficient in practice because many hosts would be tested even when their likelihood of drug 1–resistant carriage is low. By contrast, in the SSPIP, only classmates of children who developed PRP infections were tested. Therefore, even if PRP were rare in the general population, each child tested through the SSPIP would have a substantial likelihood of PRP carriage. By introducing simple contact-tracing principles, the effective rate of carriage discovery can be higher for a given level of investment in patient sampling.
Discussion
The antibiotic-resistance crisis is placing increasing pressure on healthcare globally and is widely viewed as a one-way street toward a dangerous “post-antibiotic world” [1,4]. In this paper, we ask whether RD, when combined with public health interventions such as HTC for drug-resistant bacterial strains, can substantially change the trajectory of resistance evolution.
In the early decades of the antibiotic era, doctors had no choice but to treat and control infection unconditionally, creating a selective pressure favoring strains that were resistant to whatever antibiotic was being widely prescribed. In that context, once resistant strains emerge with zero biological fitness costs, all antibiotics become “exhaustible resources” whose value to society is diminished by use [2,3]. In this paper, we show that exhaustible resource antibiotics can in principle be transformed into “renewable resources” whose value to society can be maintained over time even as they are put to widespread use, so long as (i) RD is available to detect the target pathogen and determine its antibiotic-sensitivity profile, (ii) prescribers adopt RD-informed treatment strategies, (iii) identified resistant cases are subject to more stringent transmission control, and (iv) bystander selection on the target pathogen is either minimal (Fig 2) or counteracted (Fig 5). For obligate pathogens that face minimal bystander selection, POC-RD–informed treatment reduces the advantage that resistant strains would otherwise enjoy [41], but—as we show—RD-informed treatment plus heightened control can potentially be sufficient to create a net selection against resistant pathogen strains, reducing their prevalence over time in the pathogen population. However, for opportunistic pathogens that face extensive bystander selection, POC-RD alone are insufficient—identifying resistant strains when they are not yet causing infection (so-called carriage RD) is essential to reverse the rise of resistance. This note of caution is important as society seeks to allocate resources most effectively in the struggle against antibiotic resistance.
Our analysis identifies strategies to renew or maintain sensitivity to an antibiotic in a pathogen population, the most effective of which depend on the availability of other antibiotics that can be used to treat resistant infections. The potential to restore antibiotic sensitivity is therefore limited once pan-resistant strains are in circulation. Consider now the impact of the discovery of a new antibiotic (drug 3) to which these bacteria are still sensitive. Drug 3’s discovery transforms previously pan-resistant bacteria into treatable “multidrug-resistant bacteria.” Providers can now deploy targeted treatment and HTC to hold the multidrug-resistant strain at a reproductive disadvantage. In this way, introducing a new antibiotic to which disease-causing strains are not yet resistant may make it possible to reverse the rise of previously pan-resistant bacteria, restoring the effectiveness of preexisting antibiotics. Moreover, as multidrug resistance to preexisting antibiotics grows less prevalent over time, the number of patients who need the new antibiotic will itself decline over time, allowing the new antibiotic to be held in reserve for increasingly rare cases for which it is the only effective treatment. This potential decline in drug 3 use highlights the importance of having drug 3 RD available when drug 3 is introduced, when we envisage the greatest use. Indeed, if drug 3’s introduction is coupled with drug 1–3 RD, it may not be necessary to discover even more new antibiotics beyond drug 3, because diagnostics will allow S&D tactics against rare 1–3 pan-resistant strains. We note that rolling out drug 3 at the same time as a drug 3 diagnostic raises the challenge of diagnosing resistance before widespread clinical use and clinical resistance discovery, necessitating increased investment in resistance discovery in the laboratory and phenotypic resistance surveillance in the clinic.
Whether net selection against resistant strains can be maintained depends on the effectiveness of the HTC measures that can be feasibly targeted against each resistant strain. By design, HTC measures impose additional barriers to resistant-bacterial transmission by (i) identifying resistant bacteria (during infection and/or asymptomatic colonization) and (ii) deploying additional resources specifically to prevent their transmission. Many sorts of HTC measures could be relevant in different contexts. Some examples include the following: for hospital-associated infections, imposing heightened contact precautions when a hospitalized patient is found to have resistant infection [19,21]; for pneumococcal infection, requiring young children found to be infected or colonized with PRPs to stay home from daycare [38]; for sexually transmitted diseases, providing expedited partner therapy (EPT) when a patient is found to have resistant infection [42]; or, for livestock-associated infections, eradicating an entire herd when resistant infection is identified. Note that HTC effectiveness may vary depending on the resistant strain being targeted, e.g., EPT measures may be less effective against pan-resistant strains since partners’ transmissibility cannot be controlled through treatment, whereas other measures may only be economically feasible against some strains, e.g., eradicating an entire herd may only be economical when pan-resistant infection is found, because other strains can be controlled through RD-informed treatment. More research is needed to quantify the effectiveness of HTC measures in practice.
Additional strategic options can be used to reduce the prevalence of bacterial strains with intermediate resistance, if RD is available that provides quantitative (e.g., sequence-based inference of minimal inhibitory concentration [43]) information on the degree of intermediate resistance. Details are provided in S1B Text but, to see the point, imagine that POC-RD had been available when penicillin was first introduced that could quantitatively determine the penicillin sensitivity of gonorrhea infections. Neisseria gonorrhoeae strains that were less sensitive to penicillin emerged in the 1940s and 1950s, but, at the time, these strains could still be effectively treated with a higher dose [44]. Armed with quantitative POC-RD, providers would have been able to target intermediate-resistant gonococci with a higher penicillin dose—taking away the treatment survival advantage that intermediate-resistant gonococci would otherwise enjoy—and could also have deployed additional public health resources to find and treat others who might still be spreading intermediate-resistant gonococci. Such a policy of RD-informed treatment and heightened discovery could have potentially held intermediate-resistant gonorrhea strains at an overall reproductive disadvantage relative to highly sensitive strains, though only with HTC would it have been possible to avoid rapid selection of the highly resistant strains [45]. See S1B Text for mathematical details.
The example of the gonococcus raises the key challenge of bystander exposure to antibiotics, because gonorrhea infection is initially (and in some carriers, entirely) asymptomatic and therefore does not drive immediate medical attention and exposure to POC-RD. During the asymptomatic phase of infection, drug-sensitive gonococcal genotypes are at risk of being cleared due to antibiotics taken for other medical concerns [46]. In Fig 3, we outline how the extent of the bystander challenge is even greater for commensal opportunistic pathogens [47,48] that spend proportionately longer in asymptomatic carriage states. Parameterizing our SCIS model (incorporating a carriage/asymptomatic stage, prone to bystander selection) for the key commensal opportunist S. pneumoniae illustrates that selecting against resistance via POC-RD–informed strategies alone is not a plausible outcome for this particular pathogen (Fig 4), given the lengthy duration of carriage and relatively rare and brief infection events caused by this species.
We explore a strategic response to this concern: conditional interventions in response to diagnostic information during asymptomatic carriage. Fig 5 illustrates that coupling differential transmission control to carriage RD can drive net selection against resistance, even for the most carriage-prone serotypes of the pneumococcus. The SSPIP offers a concrete example of using carriage RD to drive public health interventions. Although the SSPIP targeted pneumococcal strains that remained treatable by other antibiotics, similar programs could target pan-resistant strains and, if sufficiently intensive and comprehensive, potentially select against these pan-resistant strains even as those with sensitive infection continue to receive antibiotic treatment.
We note that, in theory, our logic of conditional interventions during carriage could be extended to incorporate broader microbiome resistance diagnostics (M-RD) and M-RD–informed interventions. Although simple in outline, implementation presents technical challenges on several fronts, not least in establishing meaningful sampling protocols, designing appropriate narrow-spectrum interventions [10,49–54], and designing appropriate strategic rules for intervention choice given potentially conflicting microbiome and infection-site resistance profiles. We also note that, independent of any M-RD innovations, the widespread uptake of POC-RD–informed antibiotic use will likely reduce bystander selection due to an overall reduction in antibiotic use (e.g., in the context of viral infections) and a potential shift toward narrower-spectrum antibiotics being prescribed against known pathogen targets.
Our focus on homogenous, closed populations ensures that R0 maximization is always favored by selection, simplifying the evolutionary analysis [55]. However, the basic idea underlying our analysis of modulating strain-fitness landscapes applies more broadly to models in which R0 is not a sufficient proxy for fitness, such as in cases of multiple carriage [55], host population structure [56], or an open population [28].
In the Supporting information, we extend our SIS model analysis to consider the effect of antibiotic rationing whereby some patients are left untreated (S1B Text) and show that our findings are robust to (i) environmental reservoirs of resistant bacteria (S1D Text), (ii) host migration from high-resistance regions (S1D Text), (iii) resistance-conferring mutation (S1D Text), (iv) competitive release (S1D Text), (v) diagnostic errors (S1E Text), and (vi) diagnostic escape (S1E Text). Inflows of resistant cases (mutation, migration) together with diagnostic errors weigh on the scale in favor of resistant strains but can all be counteracted by sufficiently high reproductive penalties to correctly targeted resistant strains. Diagnostic escape [57] presents a qualitatively distinct challenge in which diagnostic tests themselves become obsolete due to evolutionary responses in the pathogen (e.g., loss or modification of resistance marker). The risk of diagnostic escape highlights the importance of active resistance surveillance and rapid new-diagnostic development.
POC-RD are already a top public health priority, with a major emphasis on rapidity (<1 hour) [58]. POC-RD are critical for early effective treatment of life-threatening infections when treatment cannot be delayed. However, most antibiotic prescriptions are for less severe infections in which patients can wait longer to benefit from more complete diagnostic information [13]. Our analyses illustrate that, for the public health goal of selecting against resistance in pathogens with minimal bystander selection, we have more time to act—delays until treatment on the order of hours or even days following initial infection may still allow for selection against resistance (Fig 2). However, our conclusions depend critically on the life history of the target pathogen, with the message that reversing resistance in opportunistic pathogens subject to bystander selection is not generally plausible with POC-RD information alone (Fig 4) and will require additional interventions conditioned on carriage RD (Fig 5). Our conclusions also depend on the ability of RD to distinguish multiple resistances in a multidrug context, highlighting the importance of diagnostic breadth as well as rapidity. Diagnostic-informed approaches to reversing resistance face another time constraint—our proposed strategies for resistance-targeted intervention are most effective when pan-resistant strains are still rare (S1B Text). If we fail to act decisively while bacteria that are resistant to all antibiotics remain rare [59,60], we may then be unable to reverse the continued rise of untreatable bacterial disease.
Supporting information
This supplement provides supporting discussion (Part A) and mathematical detail. Part B provides detailed derivations of the mathematical expressions in Case 1 on an obligate pathogen whereas Part C provides details for Cases 2–3 on an opportunistic pathogen. Parts D–E explore the robustness of our main findings in a variety of extensions. Part F provides details on data sources for Fig 3. Part G provides details supporting the parameter values used to construct Figs 4 and 5.
(DOCX)
Reproduction numbers of sensitive strain (I0), drug 1–resistant strain (I1), and pan-resistant strain (I12), under various medical interventions: (a) no treatment, I0 enjoys advantage due to fitness costs associated with resistance; (b) all treated with drug 1, I1 and I12 enjoy advantage due to drug 1 resistance; (c) resistance-targeted treatment, I1 now at disadvantage unless (as shown) drug 2 is sufficiently less effective than drug 1; and (d) resistance-targeted accelerated treatment, whereby strain-I1 infections come more quickly to medical attention due to heightened discovery.
(TIF)
Reproduction numbers of sensitive strain (I0) and pan-resistant strain (I12), under various medical interventions: (a) fraction α≤1 treated with drug 1, I12 enjoys advantage due to surviving treatment unless α is sufficiently small; (b) PRT isolation, I12 now at disadvantage even if all sensitive infections treated with drug 1; (c) PRT HTC, with HTC shown here as much less effective than isolation; (d) PRT accelerated HTC, whereby strain-I12 infections come more quickly under control due to heightened discovery. HTC, heightened transmission control; PRT, pan-resistance targeted.
(TIF)
Reproduction numbers of sensitive strain (I0) and pan-resistant strain (I12) of an opportunistic pathogen, under various medical interventions: (a) no treatment or control, I12 enjoys advantage due to surviving incidental antibiotic exposure; (b) PRT isolation during infection, I12 may still enjoy advantage if the pathogen dwells mainly in carriage; (c) PRT isolation during infection plus PRT carriage intervention, I12 now at overall disadvantage so long as carriage intervention more effective at clearing pan-resistant bacteria than incidental exposure is at clearing sensitive bacteria (see Part C for details). PRT, pan-resistance targeted.
(TIF)
The maximal proportion of sensitive infections that can be treated without causing an increase in resistance (α*) is plotted against diagnostic delay D, assuming that all infections are immediately discovered and there are no biological fitness costs. The dashed vertical line indicates the longest diagnostic delay (D*) consistent with selection against drug 1 resistance while treating all cases. Three scenarios are shown: RD not available (No RD), for which α* = 0; RD available with delay and pan-resistance not yet emerged (RD, no I12); RD available with delay and pan-resistance widespread (RD, I12). Parameters (rates per day): , . RD, resistance diagnostics.
(TIF)
The maximal proportion of sensitive infections that can be treated without causing an increase in drug 1 resistance (α*) is plotted against the expected duration of carriage. Two POC-RD scenarios are shown: with () and without () transmission control. Vertical arrows represent pneumococcal serotypes with below-average carriage duration (see main text Fig 4 for broader range of serotype carriage durations). The remaining parameters (rates per day) are d = 0.001, = 5 × 10−4, = 0, = 1, = 0.125. We make the simplifying assumption that baseline carriage and infection transmission rates are identical (βC = βI = β), ensuring that α* does not depend on β. Details on parameterization are in Part G. POC-RD, point-of-care resistance diagnostics.
(TIF)
Acknowledgments
We thank Luke McNally, Dan Cornforth, Steve Diggle, Arjun Srinivasan, Cliff McDonald, Ben Metcalf, and Alison Halpin for discussion and comments on earlier drafts.
Abbreviations
- CDC
Centers for Disease Control and Prevention
- CRE
carbapenem-resistant Enterobacteriaceae
- EPT
expedited partner therapy
- HTC
heightened transmission control
- M-RD
microbiome resistance diagnostics
- MRSA
methicillin-resistant Staphylococcus aureus
- POC-RD
point-of-care resistance diagnostics
- PRP
penicillin-resistant pneumococcus
- SCIS
Susceptible-Carriage-Infected-Susceptible
- S&D
search and destroy
- SIS
Susceptible-Infected-Susceptible; SSPIP, South Swedish Pneumococcal Intervention Project
Data Availability
All relevant data are within the paper and its Supporting Information files.
Funding Statement
The project described was supported by the Centers for Disease Control (OADS BAA 2016-N-17812), the National Institute of General Medical Sciences (U54GM088558), the National Heart Lung Blood Institute (R56HL142857), the Simons Foundation (396001), the Wenner-Gren Foundations, and the Royal Physiographic Society of Lund. The funders had no role in study design, data collection and analysis, decision to publish, or preparation of the manuscript
References
- 1.Centers for Disease Control and Prevention. Antibiotic resistance threats in the United States, 2013. U.S. Department of Health and Human Services, Centers for Disease Control and Prevention; 2013. [Google Scholar]
- 2.Laxminarayan R, Duse A, Wattal C, Zaidi AKM, Wertheim HFL, Sumpradit N, et al. Antibiotic resistance—the need for global solutions. Lancet Infect Dis. 2013. December;13:1057–98. 10.1016/S1473-3099(13)70318-9 [DOI] [PubMed] [Google Scholar]
- 3.Center for Disease Dynamics Economics & Policy. The state of the world’s antibiotics 2015. Washington, D.C.: The Center; 2015. [Google Scholar]
- 4.World Health Organization. Antimicrobial resistance Fact Sheet 194 [Internet]. Geneva: The Organization; 2014. April Available from: http://www.wpro.who.int/mediacentre/releases/2014/AMR_factsheet_FINAL.pdf. [cited 2019 Apr 3]. [Google Scholar]
- 5.European Centre for Disease Prevention and Control. Antimicrobial resistance surveillance in Europe 2014. Solna (Sweden): The Centre; 2015. [Google Scholar]
- 6.Review on Antimicrobial Resistance. Tackling Drug-Resistant Infections Globally: Final Report and Recommendations. London: The Review; 2016. May. [Google Scholar]
- 7.Holmes AH, Moore LSP, Sundsfjord A, Steinbakk M, Regmi S, Karkey A, et al. Understanding the mechanisms and drivers of antimicrobial resistance. Lancet. 2016;387(10014):176–87. 10.1016/S0140-6736(15)00473-0 [DOI] [PubMed] [Google Scholar]
- 8.Bell G, Maclean C. The Search for ‘Evolution-Proof ‘ Antibiotics. Trends Microbiol. 2018;26(6):471–483. 10.1016/j.tim.2017.11.005 [DOI] [PubMed] [Google Scholar]
- 9.Baym M, Stone LK, Kishony R. Multidrug evolutionary strategies to reverse antibiotic resistance. Science. 2016. January 1;351(6268):aad3292 10.1126/science.aad3292 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 10.Chan BK, Sistrom M, Wertz JE, Kortright KE, Narayan D, Turner PE. Phage selection restores antibiotic sensitivity in MDR Pseudomonas aeruginosa. Sci Rep. 2016. July 26;6(1):26717. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 11.Kouyos RD, Abel zur Wiesch P, Bonhoeffer S. Informed Switching Strongly Decreases the Prevalence of Antibiotic Resistance in Hospital Wards. PLoS Comput Biol. 2011;7(3):e1001094 10.1371/journal.pcbi.1001094 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 12.Beardmore RE, Peña-Miller R. Rotating antibiotics selects optimally against antibiotic resistance, in theory. Math Biosci Eng. 2010. July;7(3):527–52. 10.3934/mbe.2010.7.527 [DOI] [PubMed] [Google Scholar]
- 13.Wollein Waldetoft K, Brown SP. Alternative therapeutics for self-limiting infections—An indirect approach to the antibiotic resistance challenge. PLoS Biol. 2017;15(12):e2003533 10.1371/journal.pbio.2003533 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 14.Ross-Gillespie A, Weigert M, Brown SP, Kümmerli R. Gallium-mediated siderophore quenching as an evolutionarily robust antibacterial treatment. Evol Med Public Heal. 2014. January 1;2014(1):18–29. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 15.Allen RC, Popat R, Diggle SP, Brown SP. Targeting virulence: can we make evolution-proof drugs? Nat Rev Microbiol. 2014. April 1;12(4):300–8. 10.1038/nrmicro3232 [DOI] [PubMed] [Google Scholar]
- 16.Tuite AR, Gift TL, Chesson HW, Hsu K, Salomon JA, Grad YH. Impact of Rapid Susceptibility Testing and Antibiotic Selection Strategy on the Emergence and Spread of Antibiotic Resistance in Gonorrhea. J Infect Dis. 2017. November 27;216(9):1141–9. 10.1093/infdis/jix450 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 17.Turner KM, Christensen H, Adams EJ, McAdams D, Fifer H, McDonnell A, et al. Analysis of the potential for point-of-care test to enable individualised treatment of infections caused by antimicrobial-resistant and susceptible strains of Neisseria gonorrhoeae: a modelling study. BMJ Open. 2017. June 14;7(6):e015447 10.1136/bmjopen-2016-015447 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 18.Menzies NA, Cohen T, Lin H-H, Murray M, Salomon JA. Population health impact and cost-effectiveness of tuberculosis diagnosis with Xpert MTB/RIF: a dynamic simulation and economic evaluation. PLoS Med. 2012;9(11):e1001347 10.1371/journal.pmed.1001347 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 19.Bootsma MCJ, Diekmann O, Bonten MJM. Controlling methicillin-resistant Staphylococcus aureus: Quantifying the effects of interventions. Proc Natl Acad Sci. 2006;103(14):5620–5. 10.1073/pnas.0510077103 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 20.Schwaber MJ, Lev B, Israeli A, Solter E, Smollan G, Rubinovitch B, et al. Containment of a Country-wide Outbreak of Carbapenem-Resistant Klebsiella pneumoniae in Israeli Hospitals via a Nationally Implemented Intervention. Clin Infect Dis. 2011;52(7):848–855. 10.1093/cid/cir025 [DOI] [PubMed] [Google Scholar]
- 21.Cohen MJ, Block C, Levin PD, Schwartz C, Gross I, Weiss Y, et al. Institutional Control Measures to Curtail the Epidemic Spread of Carbapenem-Resistant Klebsiella pneumoniae: A 4-Year Perspective. Infect Control Hosp Epidemiol. 2011;32(7):673 10.1086/660358 [DOI] [PubMed] [Google Scholar]
- 22.Jain R, Kralovic SM, Evans ME, Ambrose M, Simbartl LA, Obrosky DS, et al. Veterans Affairs Initiative to Prevent Methicillin-Resistant Staphylococcus aureus Infections. N Engl J Med. 2011;364(15):1419–30. 10.1056/NEJMoa1007474 [DOI] [PubMed] [Google Scholar]
- 23.Wertheim HFL, Vos MC, Boelens HAM, Voss A, Vandenbroucke-Grauls CMJE, Meester MHM, et al. Low prevalence of methicillin-resistant Staphylococcus aureus (MRSA) at hospital admission in the Netherlands: the value of search and destroy and restrictive antibiotic use. J Hosp Infect. 2004;56:321–5. 10.1016/j.jhin.2004.01.026 [DOI] [PubMed] [Google Scholar]
- 24.Vos MC, Ott A, Verbrugh HA. Successful Search-and-Destroy Policy for Methicillin-Resistant Staphylococcus aureus in The Netherlands. J Clin Microbiol. 2005;43(4):2034–5. 10.1128/JCM.43.4.2034-2035.2005 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 25.Gurieva T, Bootsma MCJ, Bonten MJM. Successful Veterans Affairs Initiative to Prevent Methicillin-Resistant Staphylococcus aureus Infections Revisited. Clin Infect Dis. 2012. February;54:1618–20. 10.1093/cid/cis272 [DOI] [PubMed] [Google Scholar]
- 26.Edmond MB, Wenzel RP. Screening Inpatients for MRSA—Case Closed. N Engl J Med. 2013;368(24):2314–5. 10.1056/NEJMe1304831 [DOI] [PubMed] [Google Scholar]
- 27.Spicknall IH, Foxman B, Marrs CF, Eisenberg JNS. A Modeling Framework for the Evolution and Spread of Antibiotic Resistance: Literature Review and Model Categorization. Am J Epidemiol. 2013;178(14):508–20. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 28.Lipsitch M, Bergstrom CT, Levin BR. The epidemiology of antibiotic resistance in hospitals: Paradoxes and prescriptions. Proc Natl Acad Sci. 2000;97(4):1938–43. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 29.Laxminarayan R, Brown G. Economics of Antibiotic Resistance: A Theory of Optimal Use. J Environ Econ Manage. 2001;42:183–206. [Google Scholar]
- 30.Vogwill T, MacLean RC. The genetic basis of the fitness costs of antimicrobial resistance: a meta-analysis approach. Evol Appl. 2015. March;8(3):284–95. 10.1111/eva.12202 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 31.Tedijanto C, Olesen S, Grad Y, Lipsitch M. Estimating the proportion of bystander selection for antibiotic resistance among potentially pathogenic bacterial flora. Proc Natl Acad Sci. 2018;115(51):E11988–E11995. 10.1073/pnas.1810840115 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 32.Carapetis JR, Steer AC, Mulholland EK, Weber M. The global burden of group A streptococcal diseases. Lancet Infect Dis. 2005. November;5(11):685–94. 10.1016/S1473-3099(05)70267-X [DOI] [PubMed] [Google Scholar]
- 33.Lessa FC, Mu Y, Bamberg WM, Beldavs ZG, Dumyati GK, Dunn JR, et al. Burden of Clostridium difficile Infection in the United States. N Engl J Med. 2015. February 26;372(9):825–34. 10.1056/NEJMoa1408913 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 34.Furuya-Kanamori L, Marquess J, Yakob L, Riley TV, Paterson DL, Foster NF, et al. Asymptomatic Clostridium difficile colonization: epidemiology and clinical implications. BMC Infect Dis. 2015. December 14;15(1):516. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 35.Sleeman KL, Griffiths D, Shackley F, Diggle L, Gupta S, Maiden MC, et al. Capsular Serotype–Specific Attack Rates and Duration of Carriage of Streptococcus pneumoniae in a Population of Children. J Infect Dis. 2006. September 1;194(5):682–8. 10.1086/505710 [DOI] [PubMed] [Google Scholar]
- 36.Rozen DE, McGee L, Levin BR, Klugman KP. Fitness Costs of Fluoroquinolone Resistance in Streptococcus pneumoniae. Antimicrob Agents Chemother. 2007. February 1;51(2):412–6. 10.1128/AAC.01161-06 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 37.Trzcinski K, Thompson CM, Gilbey AM, Dowson CG, Lipsitch M. Incremental increase in fitness cost with increased beta-lactam resistance in pneumococci evaluated by competition in an infant rat nasal colonization model. J Infect Dis. 2006. May 1;193(9):1296–303. 10.1086/501367 [DOI] [PubMed] [Google Scholar]
- 38.Ekdahl K, Hansson HB, Mölstad S, Söderström M, Walder M, Persson K. Limiting the spread of penicillin-resistant Streptococcus pneumoniae: experiences from the South Swedish Pneumococcal Intervention Project. Microb Drug Resist. 1998. January 29;4(2):99–105. 10.1089/mdr.1998.4.99 [DOI] [PubMed] [Google Scholar]
- 39.Melander E, Hansson H-B, Mölstad S, Persson K, Ringberg H. Limited spread of penicillin-nonsusceptible pneumococci, Skåne County, Sweden. Emerg Infect Dis. Centers for Disease Control and Prevention; 2004. June;10(6):1082–7. 10.3201/eid1006.030488 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 40.Huang SS, Finkelstein JA, Lipsitch M. Modeling Community- and Individual-Level Effects of Child-Care Center Attendance on Pneumococcal Carriage. Clin Infect Dis. 2005. May 1;40(9):1215–22. 10.1086/428580 [DOI] [PubMed] [Google Scholar]
- 41.McAdams D. Resistance diagnosis and the changing epidemiology of antibiotic resistance. Ann N Y Acad Sci. 2017. January 1;1388(1):5–17. 10.1111/nyas.13300 [DOI] [PubMed] [Google Scholar]
- 42.Hogben M, Collins D, Hoots B, OʼConnor K. Partner Services in Sexually Transmitted Disease Prevention Programs: A Review. Sex Transm Dis. 2016. February;43(2 Suppl 1):S53–62. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 43.Li Y, Metcalf BJ, Chochua S, Li Z, Gertz RE, Walker H, et al. Penicillin-Binding Protein Transpeptidase Signatures for Tracking and Predicting Beta-Lactam Resistance Levels in Streptococcus pneumoniae. MBio. 2016;7(3):1–9. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 44.Amies CR. Development of resistance of gonococci to penicillin: an eight-year study. Can Med Assoc J. 1967. January;96(1):33–5. [PMC free article] [PubMed] [Google Scholar]
- 45.Wang YC, Lipsitch M. Upgrading antibiotic use within a class: Tradeoff between resistance and treatment success. Proc Natl Acad Sci. 2006. June 20;103(25):9655–60. 10.1073/pnas.0600636103 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 46.Olesen SW, Torrone EA, Papp JR, Kirkcaldy RD, Lipsitch M, Grad YH. Azithromycin Susceptibility Among Neisseria gonorrhoeae Isolates and Seasonal Macrolide Use. J Infect Dis. 2019. January 29;219(4):619–23. 10.1093/infdis/jiy551 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 47.Brown SP, Cornforth DM, Mideo N. Evolution of virulence in opportunistic pathogens: generalism, plasticity, and control. Trends Microbiol. 2012. July 1;20(7):336–42. 10.1016/j.tim.2012.04.005 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 48.Levin BR, Bull JJ. Short-sighted evolution and the virulence of pathogenic microorganisms. Trends Microbiol. 1994. March 1;2(3):76–81. [DOI] [PubMed] [Google Scholar]
- 49.Santagati M, Scillato M, Patanè F, Aiello C, Stefani S. Bacteriocin-producing oral streptococci and inhibition of respiratory pathogens. FEMS Immunol Med Microbiol. 2012. June 1;65(1):23–31. 10.1111/j.1574-695X.2012.00928.x [DOI] [PubMed] [Google Scholar]
- 50.Nakatsuji T, Chen TH, Narala S, Chun KA, Two AM, Yun T, et al. Antimicrobials from human skin commensal bacteria protect against Staphylococcus aureus and are deficient in atopic dermatitis. Sci Transl Med. 2017. February 22;9(378):eaah4680 10.1126/scitranslmed.aah4680 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 51.Saeidi N, Wong CK, Lo T-M, Nguyen HX, Ling H, Leong SSJ, et al. Engineering microbes to sense and eradicate Pseudomonas aeruginosa, a human pathogen. Mol Syst Biol. 2014. April 16;7(1):521–521. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 52.Hwang IY, Tan MH, Koh E, Ho CL, Poh CL, Chang MW. Reprogramming Microbes to Be Pathogen-Seeking Killers. ACS Synth Biol. 2014. April 18;3(4):228–37. 10.1021/sb400077j [DOI] [PubMed] [Google Scholar]
- 53.Citorik RJ, Mimee M, Lu TK. Sequence-specific antimicrobials using efficiently delivered RNA-guided nucleases. Nat Biotechnol. 2014. November 21;32(11):1141–5. 10.1038/nbt.3011 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 54.Bikard D, Euler CW, Jiang W, Nussenzweig PM, Goldberg GW, Duportet X, et al. Exploiting CRISPR-Cas nucleases to produce sequence-specific antimicrobials. Nat Biotechnol. 2014. November 5;32(11):1146–50. 10.1038/nbt.3043 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 55.Lipsitch M, Colijn C, Cohen T, Hanage WP, Fraser C. No coexistence for free: Neutral null models for multistrain pathogens. Epidemics. 2009. March;1(1):2–13. 10.1016/j.epidem.2008.07.001 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 56.Blanquart F, Lehtinen S, Lipsitch M, Fraser C. The evolution of antibiotic resistance in a structured host population. J R Soc Interface. 2018. June;15:20180040 10.1098/rsif.2018.0040 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 57.Hill-Cawthorne GA, Hudson LO, El Ghany MFA, Piepenburg O, Nair M, Dodgson A, et al. Recombinations in Staphylococcal Cassette Chromosome mec Elements Compromise the Molecular Detection of Methicillin Resistance in Staphylococcus aureus. PLoS ONE. 2014. June 27;9(6):e101419 10.1371/journal.pone.0101419 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 58.Longitude Prize. What Kind of Test Could Win the Longitude Prize 2017. July 26 In: Longitude Prize Blog [Internet]. London;The Prize; Available from: https://longitudeprize.org/blog-post/what-kind-test-could-win-longitude-prize. [cited 2019 Apr 3] [Google Scholar]
- 59.Elemam A, Rahimian J, Mandell W. Infection with Panresistant Klebsiella pneumoniae: A Report of 2 Cases and a Brief Review of the Literature. Clin Infect Dis. 2009;49:271–4. 10.1086/600042 [DOI] [PubMed] [Google Scholar]
- 60.Lobo LJ, Tulu Z, Aris RM, Noone PG. Pan-Resistant Achromobacter xylosoxidans and Stenotrophomonas maltophilia Infection in Cystic Fibrosis Does Not Reduce Survival After Lung Transplantation. Transplantation. 2015;99(10):2196–202. 10.1097/TP.0000000000000709 [DOI] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.
Supplementary Materials
This supplement provides supporting discussion (Part A) and mathematical detail. Part B provides detailed derivations of the mathematical expressions in Case 1 on an obligate pathogen whereas Part C provides details for Cases 2–3 on an opportunistic pathogen. Parts D–E explore the robustness of our main findings in a variety of extensions. Part F provides details on data sources for Fig 3. Part G provides details supporting the parameter values used to construct Figs 4 and 5.
(DOCX)
Reproduction numbers of sensitive strain (I0), drug 1–resistant strain (I1), and pan-resistant strain (I12), under various medical interventions: (a) no treatment, I0 enjoys advantage due to fitness costs associated with resistance; (b) all treated with drug 1, I1 and I12 enjoy advantage due to drug 1 resistance; (c) resistance-targeted treatment, I1 now at disadvantage unless (as shown) drug 2 is sufficiently less effective than drug 1; and (d) resistance-targeted accelerated treatment, whereby strain-I1 infections come more quickly to medical attention due to heightened discovery.
(TIF)
Reproduction numbers of sensitive strain (I0) and pan-resistant strain (I12), under various medical interventions: (a) fraction α≤1 treated with drug 1, I12 enjoys advantage due to surviving treatment unless α is sufficiently small; (b) PRT isolation, I12 now at disadvantage even if all sensitive infections treated with drug 1; (c) PRT HTC, with HTC shown here as much less effective than isolation; (d) PRT accelerated HTC, whereby strain-I12 infections come more quickly under control due to heightened discovery. HTC, heightened transmission control; PRT, pan-resistance targeted.
(TIF)
Reproduction numbers of sensitive strain (I0) and pan-resistant strain (I12) of an opportunistic pathogen, under various medical interventions: (a) no treatment or control, I12 enjoys advantage due to surviving incidental antibiotic exposure; (b) PRT isolation during infection, I12 may still enjoy advantage if the pathogen dwells mainly in carriage; (c) PRT isolation during infection plus PRT carriage intervention, I12 now at overall disadvantage so long as carriage intervention more effective at clearing pan-resistant bacteria than incidental exposure is at clearing sensitive bacteria (see Part C for details). PRT, pan-resistance targeted.
(TIF)
The maximal proportion of sensitive infections that can be treated without causing an increase in resistance (α*) is plotted against diagnostic delay D, assuming that all infections are immediately discovered and there are no biological fitness costs. The dashed vertical line indicates the longest diagnostic delay (D*) consistent with selection against drug 1 resistance while treating all cases. Three scenarios are shown: RD not available (No RD), for which α* = 0; RD available with delay and pan-resistance not yet emerged (RD, no I12); RD available with delay and pan-resistance widespread (RD, I12). Parameters (rates per day): , . RD, resistance diagnostics.
(TIF)
The maximal proportion of sensitive infections that can be treated without causing an increase in drug 1 resistance (α*) is plotted against the expected duration of carriage. Two POC-RD scenarios are shown: with () and without () transmission control. Vertical arrows represent pneumococcal serotypes with below-average carriage duration (see main text Fig 4 for broader range of serotype carriage durations). The remaining parameters (rates per day) are d = 0.001, = 5 × 10−4, = 0, = 1, = 0.125. We make the simplifying assumption that baseline carriage and infection transmission rates are identical (βC = βI = β), ensuring that α* does not depend on β. Details on parameterization are in Part G. POC-RD, point-of-care resistance diagnostics.
(TIF)
Data Availability Statement
All relevant data are within the paper and its Supporting Information files.