

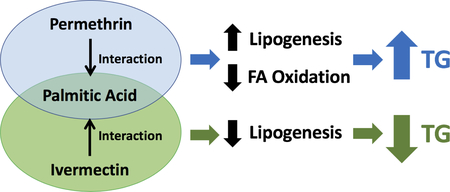
Abstract
Recent studies have reported the positive association between exposure to insecticides and increased risk of obesity and type 2 diabetes, which are closely associated with non-alcoholic fatty liver disease (NAFLD). However, it is not known if insecticide exposure can contribute to NAFLD. Thus, the goal of the current study was to determine if insecticide exposures can exacerbate the physiological conditions of NAFLD by modulating hepatic lipid metabolism. The effects of 12 insecticides on triglycerides (TG) accumulation were tested using palmitic acid (PA)-induced HepG2 hepatoma steatosis model. Results showed that among tested insecticides, permethrin and ivermectin significant interacted with palmitic acid to potentiate (permethrin) or decrease (ivermectin) TG accumulation. Further study showed that permethrin significantly promoted fatty acid synthesis, while suppressed lipid oxidation-related genes only under steatosis conditions. In comparison, ivermectin inhibited lipogenesis-related genes and promoted farnesoid X receptor, which upregulates fatty acid oxidation. Results in this study suggested that hepatic lipid metabolism may be more susceptible to insecticide exposure in the presence of excessive fatty acids, which can be associated with the development of NAFLD.
Keywords: Permethrin, Ivermectin, NAFLD, HepG2
Graphical Abstract
1. Introduction:
Recent studies have reported that insecticide exposures may disrupt lipid and glucose metabolism and contribute to the development of obesity and type 2 diabetes (Sun et al., 2017a; Swaminathan, 2013; Xiao et al., 2017a). Specifically, exposures to various insecticides induced triglyceride (TG) accumulation in adipocytes (Kim et al., 2017; Kim et al., 2014; Park et al., 2013; Sun et al., 2016a) and interacted with dietary fat, leading to disrupted lipid metabolism and insulin resistance in mice (Sun et al., 2017c; Sun et al., 2016b; Xiao et al., 2017b; Xiao et al., 2018).
Non-alcoholic fatty liver disease (NAFLD) exhibits excessive TG accumulation in the liver without routine alcohol intake and is considered the most common liver disease, with approximately 25% of adults affected worldwide (Younossi et al., 2016). There are several factors that could contribute to the development of NAFLD, including lifestyle, bodyweight and diet (Neuschwander-Tetri, 2010; Postic and Girard, 2008). NAFLD starts benignly but can progress to hepatitis, fibrosis or liver cancer due to liver injury (Argo and Caldwell, 2009). NAFLD treatments are currently geared to dietary restriction and weight loss, with limited standard treatment (Centis et al., 2013; Cheng et al., 2016). It is known that metabolic disorders, such as obesity and type 2 diabetes, are significantly correlated to the pathogenesis of NAFLD (Gaggini et al., 2013; Krishan, 2016; Marchesini et al., 2001). Additionally, it has been reported that the exposure to insecticide increased hepatic fat accumulation via altered hepatic lipid metabolism that may lead to the development of NAFLD (Yang and Park, 2018; Xiao et al., 2017a). Thus, it is important to determine how insecticides modulate hepatic lipid metabolism. In this study, 12 commonly used insecticides from the groups of avermectin (ivermectin), carbamate (aldicarb), neonicotinoid (imidacloprid), organochlorine (4,4 ‘-dichlorodiphenyldichloroethylene, DDE, and 4,4’-dichlorodiphenyltrichloroethane, DDT), organophosphorus (malathion), phenylpyrazole (fipronil), pyrethroids (cypermethrin, permethrin, and bifenthrin), and ryanoid (chlorantraniliprole) were selected based on their popularities and availabilities to evaluate their effect on TG accumulation in HepG2 hepatocytes. The insecticides used and their mode of actions have been summarized in Supplementary Table S1.
Based on the knowledge that (i) the liver is the primary organ for metabolisms of xenobiotics and (ii) insecticides may potentiate fat accumulation in the liver, the goal of this study was to determine how insecticides, particularly permethrin and ivermectin, influence hepatic lipid metabolism, and if insecticide exposure can aggravate the symptoms of NAFLD using palmitic acid (PA)-induced steatosis in HepG2 hepatocytes.
2. Materials and Methods
2.1. Materials and Chemicals
HepG2 human hepatocytes were obtained from American Type Culture Collection (Manassas, VA). Dulbecco’s modified Eagle’s medium (DMEM) and Trypan Blue stain were purchased from Thermo Fisher Scientific (Waltham, MA). Fetal bovine serum (FBS) and fatty acid free bovine serum albumin (BSA, purity ≥ 98%) were purchased from Sigma-Aldrich (St. Louis, MO). Penicillin/streptomycin was from GE Healthcare (Marlborough, MA). The insecticides, cypermethrin (> 98%), DDT (> 98.7%), DDE (> 99.7%), and permethrin (> 98.1%, a mixture of 38.7% cis and 59.4% trans isomers) were purchased from Fluka Chemical Corp. (Milwaukee, WI). Ivermectin (> 98%) was purchased from Alfa Aesar (Haverhill, MA). Deltamethrin (> 99%), fipronil (> 98%), imidacloprid (> 99.5%), and malathion (> 99.5%) were purchased from Chem Service (West Chester, PA). Aldicarb (> 98%), bifenthrin (> 98%), and chlorantraniliprole (> 99.7%) were obtained from National Pesticide Standard Repository, Environmental Protection Agency (Washington, DC). Palmitic acid (>99%) was from Nu-Chek Prep, Inc. (Elysian, MN). Primary antibodies used to detect protein expression by western blot analysis included rabbit antibodies against phosphorylated and non-phosphorylated eukaryotic initiation factor 2 α (p-eIF2 α and eIF2 α), phosphorylated and non-phosphorylated 5’ adenosine monophosphate-activated protein kinase α (p-AMPKα and AMPKα), phosphorylated and non-phosphorylated acetyl-CoA carboxylase (p-ACC and ACC), inositol-requiring enzyme 1 α (IRE1α), and binding immunoglobulin protein (BiP) were purchased from Cell Signaling Technology (Danvers, MA). Rabbit antibody against phosphorylated IRE1α (p-IRE1α) was purchased from Abcam (Cambridge, UK). Rabbit antibody against glyceraldehyde 3-phosphate dehydrogenase (GAPDH), and secondary goat horseradish peroxidase (HRP)-conjugated antibodies against rabbit were obtained from Santa Cruz Biotechnology (Dallas, TX).
2.2. Cell culture and treatment
HepG2 hepatocytes were cultured in DMEM with 10% FBS, 10,000 U/mL penicillin and 10 mg/mL streptomycin and kept at 37 °C with 5% CO2 and 95% air, as previously described (Kim et al., 2013). All insecticide samples were dissolved in dimethyl sulfoxide (DMSO) as vehicles and further diluted to achieve final concentrations of 1–50 μM. The final DMSO concentration in cell culture media was maintained at 0.1% v/v through the entire study. Cell viability was evaluated via direct viable cell count. HepG2 cells were treated with insecticides for 24 h and subjected to viable cell count using a hemocytometer (Thermo Fisher Scientific, Waltham, MA) and the dye exclusion method with 0.4% Trypan Blue stain. There was no interaction between PA and these insecticides with respect to cell viability (Suppl. Fig. S1A–S1B). PA decreased total cell count when treated with permethrin (P <0.0001, Suppl. Fig. S1A), which is consistent with a previous observation that 200 μM PA induced about 20% lipoapoptosis in HepG2 cells after 24 hour treatments (Malhi et al., 2006; Ricchi et al., 2009). By comparison, either 50 μM of permethrin or 10 μM of ivermectin had no effect on the total cell count (Suppl. Fig. S1A & S1B). Based on these results, the highest concentrations of PA, permethrin, and ivermectin used for further experiments were 200 μM, 50 μM, and 10 μM, respectively. In previous studies, the serum level of permethrin in humans was reported at 2.2 μM after oral intoxication (Gotoh et al., 1998), while the serum level of ivermectin in humans after oral administration has been recorded at 92 nM (Canga et al., 2008). Although the concentrations of permethrin and ivermectin used in this study were significantly higher than in previous human cases, insecticides with lipophilic properties can accumulate in hydrophobic areas, such as in adipose tissue (Jaga and Dharmani, 2003). In fact, DDE has been observed as high as 2,542 ng/g lipids in serum, which is equivalent to 7.3 μM (Sun et al., 2017b).
2.3. Membrane potential determination
Cell membrane depolarization was determined by measuring the relative membrane potential using a bis-(1,3-Dibutylbarbituric Acid) trimethine oxonol (DiBAC(4)3)-based method (Miao and Joyner, 1994). DiBAC(4)3 stock solution was prepared in DMSO and diluted in a HEPES buffer to make a fresh working solution before each use. Fluorescence intensity was measured at excitation 485nm/emission 516nm to obtain the membrane potential.
2.4. Triglycerides content determination
HepG2 cells were treated with serum-free DMEM containing 200 μM palmitic acid, 0.5% BSA and insecticides (with 0.1% DMSO) for 24h. A palmitic acid (PA)-BSA complex (molar ratio=2.66:1) was prepared by mixing 40 mM palmitic acid in 0.1 M KOH and 10% BSA in a PBS buffer overnight at room temperature as previously described with slightly modification (Cousin et al., 2001). The pH of the PA-BSA complex stock (20x) was adjusted to 7.2 and sterilized by filtration before use. After treatment, cells were lysed with PBS-containing 0.1% Triton X-100 and the TG content measured using the Infinity Triglycerides reagent kit (Thermo Fisher Scientific, Waltham, MA, US) following the manual’s instructions. Protein contents were measured by the Pierce BCA protein assay (Thermo Fisher Scientific, Waltham, MA).
2.5. Immunoblotting
Immunoblotting was performed as previously described (Kim et al., 2013). HepG2 cells were homogenized in a Radioimmunoprecipitation assay (RIPA) buffer by the ultrasonic cell disruptor (Fisherbrand™ Model 50, Thermo Fisher Scientific, Waltham, MA) to extract whole-cell proteins. Protein extracts were blotted to a polyvinylidene fluoride membrane (Immobilon-P, 0.45 μM pore size, EMD Millipore, Burlington, MA) after separated by 8 or 10 % sodium dodecyl sulfate-polyacrylamide gel electrophoresis (200V, 60 min). Primary and horseradish peroxidase (HRP)-conjugated secondary antibodies were then applied to detect and visualize target proteins. A chemiluminescence signal was determined using Image Station 4000MM (Kodak, Rochester, NY) for quantification.
2.6. mRNA expression assays
The expression levels of mRNA of selected genes were determined using a real-time polymerase chain reaction (PCR). mRNA was extracted using the TRIzol Reagent kit (Thermo Fisher Scientific, Waltham, MA) according to the manufacturer’s instructions. Reverse transcriptase quantitative real-time PCR (RT-qPCR) was performed according to manufacturer’s instructions to determine the mRNA. Taqman RT-qPCR assay reagents for mRNA expressions included: 18s RNA housekeeping gene (catalog No. 4319413E) for control, sterol regulatory element-binding protein 1 (SREBP1, Hs01088679_g1), acetyl-CoA carboxylase (ACC, Hs01046047_m1), fatty acid synthase (FAS, Hs00236330_m1), peroxisome proliferator-activated receptor α (PPARα, Hs00947536_m1), Farnesoid X receptor (FXR, Hs01026590_m1), carnitine palmitoyltransferase 1α (CPT1α, Hs00912671_m1), cluster of differentiation 36 (CD36, Hs00354519_m1), diacylglycerol O-acyltransferase 2 (DGAT2, Hs01045913_m1) and DNA damage-inducible transcript 3 (DDIT3, Hs00358796_g1), which were purchased from Applied Biosystems (Foster City, CA).
2.7. Statistical Analysis
Results in Figures 1A and 1B were analyzed by one-way analysis of variance (ANOVA). Results in Figures 1C and 1D were analyzed by two-way ANOVA. Since insecticide (either permethrin or ivermectin) were not used together in the same experiment of Figures 2, 3, 4, 5, 6, and 7 and supplementary Figure S1, two separate 2-way ANOVAs (one for permethrin vs. control and the other for ivermectin vs. control) were used. All analyses were done by SAS Software (version 9.4, SAS Institute, Cay, NC). If there was no significant interaction between an insecticide and palmitic acid (PA), the overall effects of the insecticide and PA were discussed including multiple comparison for doses analyzed by Tukey’s honest significance test. If there was significant interaction between an insecticide and PA, Tukey’s honest significance test for multiple comparison was completed and letters were applied to indicate significant difference between treatments. Note that, regardless of the presence of significant interaction, all results were presented per treatment group for visual presentation purposes only. Even though the same controls were used for permethrin and ivermectin, due to two separate analyses and the absence/presence of significant interaction between PA and insecticide, trends for PA or controls may be different between permethrin and ivermectin. Comparisons with a P-value of less than 0.05 were considered statistically different.
Figure 1. Effects of insecticides on TG accumulation in non-induced and steatosis-induced HepG2 hepatocytes.

TG accumulation with insecticides in non-induced (A) and steatosis-induced HepG2 cells (B). TG accumulation with permethrin (C) and ivermectin (D). HepG2 cells were incubated with 10 μM of 12 insecticides with or without 200 μM of palmitic acid (PA). Interactions were examined following 24 h treatments of 10, 50 μM of permethrin or 1, 10 μM of ivermectin with or without 50, 100, and 200 μM of palmitic acid. Numbers are mean ± S.E.M. (n = 4–6). *Statistically different from the control group at P<0.05. Means with different letters are statistically different at P<0.05. TG; triglycerides.
Figure 2. Effects of permethrin and ivermectin on membrane potentials with palmitic acids (PA).

Membrane potential after treatment of permethrin (A) or ivermectin (B). Cells were treated with 10 and 50 μM of permethrin or 1 and 10 μM of ivermectin in combination of 0, 100 and 200 μM of palmitic acid (PA). Membrane potentials were assayed after 40 minutes by combining treatments along with 250 μM DiBAC(4)3 in HEPES buffer. Numbers are mean ± S.E.M. (n = 6–8). Means with different letters are statistically different at P<0.05. DiBAC(4)3; bis-(1,3-Dibutylbarbituric Acid) trimethine oxonol; N.S., not significant.
Figure 3. Effects permethrin and ivermectin on phosphorylation of ACC in HepG2 cells.

p-ACC (phosphorylated acetyl-CoA carboxylase) with permethrin (A) and ivermectin (B). Expression of ACC with permethrin (C) or ivermectin (D). Ratio of p-ACC/ACC with permethrin (E) or ivermectin (F). Representative pictures with permethrin (G) or ivermectin (H). Cells were treated with 50 μM of permethrin or 10 μM ivermectin with or without 200 μM palmitic acid (PA) for 24 h. Protein expressions were analyzed by western blot. Numbers are mean ± S.E.M. (n=4). Means with different letters are statistically different at P<0.05. N.S., not significant.
Figure 4. Effects permethrin and ivermectin on phosphorylation of AMPKα in HepG2 cells.

p-AMPKα (phosphorylated 5’ adenosine monophosphate-activated protein kinase α) with permethrin (A) or ivermectin (B). Expression of AMPKα with permethrin (C) or ivermectin (D). Ratio of p-AMPKα/AMPKα with permethrin (E) or ivermectin (F). Representative pictures with permethrin (G) or ivermectin (H). Cells were treated with 50 μM of permethrin or 10 μM ivermectin with or without 200 μM palmitic acid (PA) for 24 h. Protein expressions were analyzed by western blot. Numbers are mean ± S.E.M. (n=4). Means with different letters are statistically different at P<0.05. N.S., not significant.
Figure 5. Interactions of permethrin, ivermectin and palmitic acid on lipogenesis pathways in HepG2 cells.

SREBP1 (sterol regulatory element-binding protein 1) with permethrin (A) or ivermectin (B). FAS (fatty acid synthase) with permethrin (C) or ivermectin (D). ACC (acetyl-CoA carboxylase) with permethrin (E) or ivermectin (F). DGAT2 (diglyceride acyltransferase 2) with permethrin (G) or ivermectin (H). Cells were treated with 10, 50 μM of permethrin or 1, 10 μM ivermectin with or without 200 μM palmitic acid (PA) for 24 h. mRNA expression analyzed by RT-qPCR. Numbers are mean ± S.E.M. (n=4). Means with different letters are statistically different at P<0.05. N.S., not significant.
Figure 6. Interactions of permethrin, ivermectin and palmitic acid on expression of fatty acid oxidation pathways.

FXR (farnesoid X receptor) with permethrin (A) or ivermectin (B). PPARα (peroxisome proliferator-activated receptor α) with permethrin (C) or ivermectin (D). CPT1α (carnitine palmitoyltransferase I α) with permethrin (E) or ivermectin (F). Cells were treated with 10, 50 μM of permethrin or 1, 10 μM ivermectin with or without the 200 μM palmitic acid (PA) for 24 h. mRNA transcription levels were analyzed by RT-qPCR. Numbers are mean ± S.E.M. (n=4). Means with different letters are statistically different at P<0.05. N.S., not significant.
Figure 7. Effects of permethrin and ivermectin on ER stress pathways.

CHOP (CCAAT-enhancer-binding protein homologous protein) with permethrin (A) or ivermectin (B). Protein expression of ER stress pathways with permethrin (C) or ivermectin (D); BiP (binding immunoglobulin protein); IRE1α (inositol-requiring enzyme 1 α); eIF2α (eukaryotic initiation factor 2 α); and GAPDH (Glyceraldehyde 3-phosphate dehydrogenase). Cells were treated with 10 and 50 μM permethrin or 1 and 10μM ivermectin (A and B), 50 μM permethrin or 10 μM ivermectin (C and D) with or without the 200 μM palmitic acid (PA) for 24 h. mRNA transcript levels were analyzed by RT-qPCR (A and B). Protein expressions analyzed by western blot (C and D). Numbers are mean ± S.E.M. (n=4). Means with different letters are statistically different at P < 0.05. N.S., not significant.
3. Results
3.1. Effects of insecticides on TG accumulation in non-induced and steatosis-induced HepG2 hepatocytes
To understand the role of insecticides in hepatic lipid metabolism, the effects of 12 insecticides on TG accumulation in HepG2 cells were tested in the presence or absence of 200 μM of PA. Compared to other fatty acids reported to induce steatosis, this concentration of PA was selected based on a previous report that this would result in relatively moderate induction of TG accumulation with minimum cytotoxicity in HepG2 hepatocytes (Ricchi et al., 2009). Thus, this condition would allow us to evaluate the effect of insecticides in potentially increasing TG accumulation in HepG2. Results in Fig. 1A showed that several insecticides significantly increased TG accumulation: the pyrethroid, cypermethrin (17%); the neonicotinoid, imidacloprid (30%), the phenylpyrazole, fipronil (42%), and the ryanoid, chlorantranillaprole (55%), while an avermectin, ivermectin, significantly decreased TG (38% reduction) compared with their respective controls. The organochlorines, DDT and DDE; the organophosphorus, malathion; the carbamate, aldicarb; and the pyrethroids, bifenthrin, deltamethrin, and permethrin, did not alter the TG content in non-induced HepG2 cells. When treated with 200 μM PA, the effects of certain insecticides were enhanced (as shown in Fig. 1B): permethrin (8% to 23%), cypermethrin (17% to 20%) and ivermectin (−38% to −47%) compared with their respective controls. Because of their distinctive effects on TG accumulation and enhancement in the presence of PA, permethrin, and ivermectin were selected for further study.
Results in Fig. 1C show a significant interaction between permethrin and PA (P=0.0020). Permethrin at 10 and 50 μM significantly enhanced TG accumulation when co-treated with 100 or 200 μM of PA compared to the respective controls (Fig. 1C). By contrast, 10 μM of ivermectin significantly decreased TG accumulation, without or with 50 and 200 μM of PA (Fig. 1D), with a significant interaction between ivermectin and PA (P<0.0001); There was a 38% TG reduction without PA, 42% reduction with 50 μM of PA, and 47% reduction with 200 μM of PA, respectively, compared to the respective controls.
3.2. Effects of permethrin and ivermectin on membrane potential.
Since permethrin and ivermectin both act by altering the membrane potential of cells, the effects of different concentrations of PA on membrane potential were tested with and without permethrin or ivermectin. There was no interaction between the insecticides and PA on membrane potentials (Fig. 2A and 2B). PA significantly caused hyperpolarization of HepG2 compared to the control (overall P=0.0033 with significant difference between the control and both 100μM and 200μM PA, P<0.05 for both, Fig. 2A). Permethrin significantly caused membrane depolarization (P<0.0001) regardless of the presence of PA (Fig. 2A). Ivermectin also induced membrane depolarization regardless of PA (P<0.0001, Fig. 2B). Ivermectin is generally considered a hyperpolarizing insecticide that decreases membrane potential, but several studies also reported it as a depolarizing agent since it causes the efflux of chloride ions and elevates membrane potential when it targets the glutamate-gated chloride channel (Ozkucur et al., 2015; Pemberton et al., 2001; Sabin et al., 2015).
3.3. Effects of permethrin and ivermectin on AMPK-ACC pathway in steatosis-induced HepG2 hepatocytes
Acetyl-CoA carboxylase (ACC) is one of the two rate-limiting enzymes for lipogenesis (Bechmann et al., 2012). In order to determine the mechanisms involved in permethrin and ivermectin-enhanced TG accumulation in HepG2 cells, the activation of ACC by phosphorylation was measured. There were significant effects of permethrin on p-ACC (57% reduction compared to the controls with P=0.0002) and PA (P=0.0011) without interaction between permethrin and PA (Fig. 3A). There was no effect of permethrin or PA on ACC (Fig. 3C), resulting in significant effects of permethrin and PA on the ratio of p-ACC/ACC (53% reduction by permethrin and 43% reduction by PA compared to the controls with P = 0.0003 and P=0.0024, respectively) without interaction between permethrin and PA (Fig. 3E).
Since there was significant interaction between ivermectin and PA on p-ACC (P=0.0015, Fig. 3B), all treatment groups were separately compared. Ivermectin non-significantly decreased p-ACC when treated without PA, but significantly increased it with PA (72% increase compared to the PA-treated control, P=0.0256, Fig. 3B). Neither ivermectin nor PA changed the protein expression level of ACC (Fig. 3D). There was a significant interaction between ivermectin and PA (P=0.0189) on the ratio of p-ACC/ACC without any significant differences between groups (Fig. 3F).
5’-Adenosine monophosphate-activated protein kinase α (AMPKα) is a cellular energy modulator that regulates energy expenditure. It is also one of the upstream regulators of phosphorylation of ACC. Permethrin significantly increased the protein expression level of p-AMPKα (39% increase compared to the controls with P = 0.0133) without any effects of PA or interaction between permethrin and PA (Fig. 4A). Permethrin had no significant effects on AMPK, while PA significantly reduced AMPK without interaction with permethrin (Fig. 4C), which resulted in a significant increase in the ratio of p-AMPKα/AMPKα (47% increase by permethrin compared to the controls with P = 0.0038) and significant effects of PA (P=0.0059) without interactions between permethrin and PA (Fig. 4E).
There was a significant effect of ivermectin on p-AMPKα (133% increase compared to the controls with P<0.0001) and significant effects of PA (P=0.0262) without interaction between ivermectin and PA (Fig. 4B). This significant effect of PA is, however, not consistent with results in Fig. 4A due to two separate analyses, one for permethrin vs. control and the other for ivermectin vs. control. No significant effect of ivermectin was observed on AMPKα but a significant PA effect was observed (P=0.0365) without interaction between ivermectin and AMPK (Fig. 4D). There was a significant increase in the ratio of p-AMPKα/AMPKα by ivermectin (124% increase compared to the controls with P<0.0001) and PA (P=0.0006) without interaction between ivermectin and PA (Fig. 4F). These results show that the phosphorylation of ACC may not be regulated by AMPK in this model.
3.4. Effects of permethrin and ivermectin on lipogenesis pathways
In order to verify the roles of permethrin and ivermectin in hepatic lipid metabolism, as well as the interaction between these insecticides and PA, the mRNA expression levels of lipogenesis-related genes were tested in both non-induced and steatosis-induced HepG2 cells. SREBP1 is a nuclear transcription factor that promotes lipogenesis (Bechmann et al., 2012). Permethrin or PA treatment did not affect the transcript level of SREBP1 nor was there any interaction between permethrin and PA (Fig. 5A). However, there was significant interaction with permethrin and PA on the transcript level of FAS (P=0.0012, Fig. 5C). Without PA, 50 μM of permethrin significantly decreased the transcript level of FAS compared to the control (30% reduction, P=0.0012), while this effect of permethrin on FAS was eliminated when co-treated with PA (Fig. 5C). Expression of ACC was decreased by permethrin treatment (P=0.0003) without any effects of PA or their interaction (Fig. 5E); 50 μM permethrin treatments significantly decreased ACC expression compared to the controls (P=0.0003) and 10 μM permethrin treatments (P=0.0038, Fig. 5E). Expression of DGAT2 was not changed by permethrin, but significantly influenced by PA alone (P=0.0017) without interaction between permethrin and PA (Fig. 5G).
By contrast, ivermectin significantly decreased the transcript level of SREBP1 by 11% compared to the controls (P < 0.0001, Fig. 5B); the expression of SREBP1 by 10 μM ivermectin treatments was significantly lower than the controls and 1μM ivermectin (P<0.0001 for both, Fig. 5B). PA had significant effects on SREBP1 regardless of the effect of ivermectin (P=0.0055 for PA and P>0.05 for interaction, Fig. 5B). Ivermectin significantly decreased the transcript levels of FAS (12% reduction compared to the controls, P<0.0001) and significant PA effects were observed (P=0.0048) independent of the effect of ivermectin (Fig. 5D). There was significant interaction between ivermectin and PA on ACC expression (P=0.0068, Fig. 5F). ACC expression was significantly decreased by ivermectin at 10 μM; 39% reduction compared to the no PA control, P=0.0033, and 30% reduction compared to the PA control, P=0.0280, respectively (Fig. 5F). For expression of DGAT2, there were significant effects of ivermectin (46% reduction compared to the controls, P<0.0001) and PA (P=0.0046) without interaction between ivermectin and PA (Fig. 5H). Treatments of 10 μM ivermectin were significantly different from other treatments on DGAT2 expression (P<0.0001 for all, Fig. 5H).
3.5. Effects of permethrin and ivermectin on FXR, PPARα, and CPT1α
In addition to the lipogenesis pathway, hepatic TG accumulation also depends on energy expenditure. FXR promotes lipid utilization, including fatty acid (FA) oxidation and hepatic lipid secretion (Kalaany and Mangelsdorf, 2006), and it is also one of the upstream regulators of PPARα, which regulates FA oxidation via CPT1α, a rate-limiting enzyme in the mitochondrial FA β-oxidation pathway (Terada and Motojima, 2011). Since there was significant interaction between permethrin and PA (P=0.0008) in the expression of FXR, all groups were compared separately (Fig. 6A). Permethrin did not affect FXR without PA, but 50 μM of permethrin decreased FXR (34% decrease compared to the PA-treated control, P=0.0007, Fig. 6A). PA significantly increased the transcript level of FXR (P<0.0001, Fig. 6A). Similarly, there was significant interaction between permethrin and PA on PPARα (P=0.0014, Fig. 6C). Permethrin did not affect PPARα without PA, but decreased PPARα when co-treated with PA (28% and 33% reductions with 10 and 50 μM permethrin with P=0.0012 and P=0.0002 compared to the PA-treated control, respectively, Fig. 6C). PA treatment alone significantly increased expression of PPARα (P=0.0147, Fig. 6C). Additionally, there was significant interaction between permethrin and PA on transcription of CPT1α (P<0.0001, Fig. 6E). The transcript level of CPT1α was unchanged by permethrin without PA, but was decreased by 38% compared to the PA-treated control when co-treated with 50 μM permethrin and PA (P<0.0001, Fig. 6E). PA alone significantly increased expression of CPT1α (84% increase, P<0.0001, Fig. 6E). Overall, FA oxidation-related genes were decreased when permethrin was co-treated with PA compared to the respective controls.
There was significant interaction between ivermectin and PA on the expression of FXR (P=0.0028, Fig. 6B), PPARα (P=0.0062, Fig. 6D), and CPT1α (P=0.0264, Fig. 6F). The transcription of FXR was increased by 10 μM of ivermectin in non-PA treated cells (318% increase) but was partially attenuated when co-treated with PA (90% compared to the PA-treated control, Fig. 6B). Ivermectin also significantly altered the transcription of PPARα; without PA, only the 10 μM concentration of ivermectin decreased PPARα (34% reduction compared the control, P=0.0098), while both 1 and 10 μM of ivermectin decreased PPARα when co-treated with PA (28% and 24% reductions with 1 and 10 μM ivermectin, P=0.0037 and P=0.0137, compared to the PA-treated control, respectively, Fig. 6D). Additionally, CPT1α transcription was not affected by ivermectin without PA, but was significantly decreased by both 1 and 10 μM of ivermectin when co-treated with PA (26% reduction by 1 μM ivermectin with P=0.0022, and 19% reduction by 10 μM ivermectin with P=0.0347 compared to the PA-treated control, respectively, Fig. 6F).
3.6. Effects of permethrin and ivermectin on ER stress pathways
Endoplasmic reticulum (ER) stress results from the accumulation of unfolded or misfolded proteins and is one of the modulators in lipogenesis pathways (Lee et al., 2012). Thus, the effects of permethrin and ivermectin on ER stress in HepG2 cells were determined. CCAAT-enhancer-binding protein homologous protein (CHOP) is a biomarker for the activation of ER stress, which is initiated by BiP and then branched into 3 major pathways, including phosphorylation of IRE1α and eIF2α, where they both activate CHOP (Oslowski and Urano, 2011). There was significant interaction between permethrin and PA (P=0.0008, Fig. 7A). The transcript level of CHOP was not changed by permethrin without PA, but was suppressed by 50 μM permethrin when co-treated with PA (27% reduction compared to the PA-treated control, P=0.0024, Fig. 7A). PA also significantly increased levels of CHOP (P<0.0001, Fig. 7A). For ER stress upstream markers, there were no interactions between permethrin and PA for all markers tested (Fig. 7C). The only significant effect of permethrin was for p-eIFα, which was significantly increased (43% compared to the controls, P=0.0069, Fig. 7C). PA had significant effects on BiP, p-IRE1α/IRE1α, p-IRE1α, p-eIF2α/eIF2α, and p-eIF2α compared to no PA treatments (Fig. 7C). Overall, these findings suggest that ER stress may have minimal influence on permethrin-altered lipid metabolism in this model.
There was significant interaction between ivermectin and PA on CHOP (P<0.0001, Fig. 7B). Ivermectin significantly increased the transcription level of CHOP (>20 fold without PA, while 147% increase with PA treatments compared to the controls, P<0.0001 for both, Fig. 7B). For ER stress-related proteins, there was interaction between ivermectin and PA on BiP (P<0.0001), but not others (Fig. 7D). Without PA treatments, ivermectin increased the protein expression level of BiP, 350% increase compared to the no-PA control (P<0.0001), while the effect of ivermectin on BiP was attenuated when co-treated with PA (40% increase compared with the PA control, P=0.0118, Fig. 7D). In addition, ivermectin also increased the protein expression level of p-IRE1α (230% increase, P<0.0001) and IRE1α (92% increase, P<0.0001), and the ratio of p-IRE1α/IRE1α (72% increase, P<0.0001) compared to the respective controls without the interaction with PA (Fig. 7D). Additionally, no significant effect of ivermectin was observed on the protein expression levels of p-eIF2α or eIF2α, and on the ratio of p-eIF2α/eIF2α. These findings suggested that ER stress caused by ivermectin might be partially involved in ivermectin-altered lipid metabolism in HepG2 cells.
4. Discussion
The liver is one of the major organs for lipid and glucose metabolism (Scollon et al., 2009) and alteration of hepatic lipid and glucose metabolism were observed in the presence of excessive FA (Ghose et al., 2011) and NAFLD (Leung and Nieto, 2013; Woolsey et al., 2016). Base on the correlation of insecticide exposure and alteration of lipid and glucose metabolism (Sun et al., 2017a; Sun et al., 2017b; Xiao et al., 2017a), the current study investigated the significance of insecticides on hepatic lipid metabolism. This study is the first to report the effect of insecticides and FA overloads in HepG2 hepatocytes.
Permethrin is the single most used synthetic pyrethroid worldwide (Council, 1999), and is a member of one of the major classes of insecticide commercially available (Sparks, 2013). Permethrin acts as an agonist in voltage-sensitive sodium channels both in invertebrates and vertebrates, including humans. In this study, permethrin increased lipogenesis and decreased FA oxidation in HepG2 cells. The current results also suggested that the post-translational regulation of ACC might be important in permethrin-induced lipogenesis as permethrin only had minimum effects on the expression of other lipogenesis-related genes, SREBP1, ACC and DGAT2 in HepG2. The current results also suggested that permethrin regulated ACC independent of AMPK, which is one of the post-translational regulators that inactivates ACC. This result is consistent with a previous in vivo study finding that permethrin activates AMPK in mice liver (Xiao et al., 2018). However, other studies reported that permethrin induced lipogenesis, dependent on the AMPK pathway in adipocytes (Kim et al., 2014) and adipose tissues (Xiao et al., 2018), by decreasing the ratio of pAMPK to AMPK. Similarly, deltamethrin promoted adipogenesis in 3T3-L1 adipocytes and C. elegans via the AMPK-dependent pathway (Shen et al., 2017), but had no effects on TG accumulation in this study. Another pyrethroid, cypermethrin, significantly increased TG accumulation in the current study, but it is not clear if cypermethrin targeted AMPK differently in hepatocytes versus other cells as it is known to activate AMPK in neuroblastoma cells (Mishra et al., 2018). These findings suggest that the effects of pyrethroids on AMPK may be tissue-specific, and further studies are needed to elucidate the mechanism of the permethrin-induced post-translational regulation of ACC, as well as the effects of other pyrethroids on the hepatic lipogenesis pathway. Additionally, the previous study showed that permethrin-induced adipogenesis in 3T3-L1 adipocytes is associated with ER stress (Xiao et al., 2017c), while deltamethrin was reported to cause ER stress in neuroblastoma cells (Hossain and Richardson, 2011). As permethrin showed a minimum effect on the ER stress in HepG2 cells in this study, further investigations are also needed to elucidate the mechanism of the permethrin in lipogenesis independent of the ER stress pathway.
In addition to increased lipogenesis, the current results suggest that permethrin suppressed FA oxidation in part by downregulating the FXR-PPARα-CPT1α pathway. These results are consistent with a previous study, where permethrin reduced hepatic PPARα in the liver of high-fat diet-fed male mice (Xiao et al., 2018). In addition, another pyrethroid tested in the current study, bifenthrin, was reported to decrease CPT1α in HepG2 cells (Xiang et al., 2018) though it did not change TG in the current study. These findings suggest that reduced FA oxidation may be involved in the effect of permethrin on hepatic lipid metabolism.
Ivermectin is a macrocyclic lactone avermectin, which is used for controlling parasites or insects by acting on the glutamate-gated chloride channel (Bloomquist, 1993). Avermectins, including ivermectin, also are known agonists of FXR, which regulates hepatic lipid and glucose metabolism by suppressing lipogenesis and gluconeogenesis (Jin et al., 2013; Jin et al., 2015) and have been suggested as potential therapeutic agents for NAFLD (Cariou, 2008; Carr and Reid, 2015; Mudaliar et al., 2013; Trauner et al., 2010). Upregulation of FXR is known to decrease lipogenesis and increase fatty acid oxidation. The current results were consistent with the above in that ivermectin upregulated FXR, leading to the down-regulation of SREBP, FAS, and ACC, potentially acting as an agonist of FXR. However, ivermectin significantly reduced PPARα and CPT1α, which were inconsistent effects for an FXR agonist (Torra et al., 2003). Overall, these results suggest that ivermectin suppressed lipogenesis via FXR-mediated mechanisms, while the effects of ivermectin on fatty acid oxidation may be independent of FXR. Alternatively, AMPK is also known as an important regulator of lipogenesis and fatty acid oxidation, where the current results showed that activation of AMPK by ivermectin did not lead to post-translational changes of ACC, although this may lead to increased fatty acid oxidation by reducing the level of malonyl-CoA, the intermediate of lipogenesis that directly inhibits CPT1α (Lage et al., 2008). Additionally, ER stress is known to play a significant role in increased lipogenesis (Lee et al., 2012; Özcan et al., 2004). However, the current results indicate that ivermectin induced ER stress while reducing lipogenesis. Thus, further study is required to elucidate the role of ivermectin on the PPARα pathway and the correlation between ivermectin-associated ER stress and alteration of lipogenesis.
In addition to alteration of lipid metabolism, alteration of glucose metabolism, dyslipidemia, and other liver functions, such as hepatic oxidative stress and liver injury, can also contribute to the development of NAFLD (Albano et al., 2005; Argo and Caldwell, 2009; Gaggini et al., 2013). Altered glucose metabolism can result from insulin resistance, which is known to increase hepatic FA uptake, lipogenesis and blocked lipid secretion, and the degree of insulin resistance is related to the degrees of NAFLD (Bugianesi et al., 2005; Gaggini et al., 2013; Yki-Jarvinen, 2010). Several pyrethroids including permethrin, were previously reported to disturb glucose metabolism. Permethrin was reported to interrupt insulin signaling in myotubes (Kim et al., 2014; Sun et al., 2017b). Cismethrin (Cremer and Seville, 1982), cypermethrin (Veerappan et al., 2012), and deltamethrin (Ray and Cremer, 1979; Yousef et al., 2006) also increased blood glucose level in rats.
Dyslipidemia can also promote hepatic FA uptake, which results in increased hepatic TG and development of NAFLD (Gaggini et al., 2013). It has been reported previously that the pyrethroids, allethrin, prallethrin, cypermethrin, and deltamethrin, induced dyslipidemia in rodents and humans (Ince et al., 2012; Narendra et al., 2008; Veerappan et al., 2012; Yousef et al., 2006). Hepatic oxidative stress can also induce inflammation and cause liver injury, which triggers liver regeneration, leading to reduced liver function and eventually resulting in fibrosis and cirrhosis, the irreversible stages of NAFLD (Albano et al., 2005; Argo and Caldwell, 2009). As critical factors in the progression of NAFLD, several pyrethroids have been demonstrated to cause hepatic oxidative stress in rodents, including bifenthrin (Zhang et al., 2015), cyhalothrin (Al‐ Sarar et al., 2014), cypermethrin (Aldana et al., 2001; Atessahin et al., 2005; Giray et al., 2001; Yavasoglu et al., 2006), deltamethrin (Abdel-Daim et al., 2013; Abdou and Abdel-Daim, 2014; Chargui et al., 2012; Tuzmen et al., 2008; Yousef et al., 2006), and permethrin (Yang et al., 2018). In addition, avermectins were also found to induce oxidative stress in the liver (Zhu et al., 2013).
In summary, the current results suggest that permethrin promoted lipogenesis and suppressed FA oxidation, while ivermectin suppressed lipogenesis in HepG2 cells. A limitation of the current study was focusing only on the effects of insecticides on hepatic lipid metabolism. Moreover, although HepG2 cells exhibit many features of normal liver cells, it may still represent the potential implication of these chemicals in hepatocyte functions. Further studies, including testing gene-modified cells (i.e. applying RNA interference) and in vivo studies, would be needed to expand the significance of the current research, as well as to elucidate the roles of the tested insecticides in the development of NAFLD and their effects on glucose metabolism, dyslipidemia, and oxidative stress.
5. Conclusions
This study suggests that both permethrin, a pyrethroid insecticide, and ivermectin, an avermectin insecticide, can alter hepatic lipid metabolism and interact with administered palmitic acid to enhance their impacts. The results are significant in that FA overload, one of the most important factors in NAFLD development, may leave the liver more susceptible to environmental chemicals such as insecticides. However, the detailed mechanisms by which permethrin and ivermectin interact with FAs to affect hepatic lipid metabolism still require further investigation.
Supplementary Material
Permethrin and ivermectin modulated lipid metabolism in HepG2 cells.
Permethrin promoted lipogenesis and reduced FA oxidation in steatosis-induced HepG2 cells.
Ivermectin reduced lipogenesis in steatosis-induced HepG2 cells.
Acknowledgement
This project was in part supported by NIH R21ES023676.
Mr. Farias-Pereira was supported by CNPq, Conselho Nacional de Desenvolvimento Científico e Tecnológico - Brasil [National Counsel of Technological and Scientific Development - Brazil].
Footnotes
Publisher's Disclaimer: This is a PDF file of an unedited manuscript that has been accepted for publication. As a service to our customers we are providing this early version of the manuscript. The manuscript will undergo copyediting, typesetting, and review of the resulting proof before it is published in its final citable form. Please note that during the production process errors may be discovered which could affect the content, and all legal disclaimers that apply to the journal pertain.
6. Reference
- Abdel-Daim MM, Abuzead SM, Halawa SM, 2013. Protective role of Spirulina platensis against acute deltamethrin-induced toxicity in rats. PloS one 8, e72991. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Abdou RH, Abdel-Daim MM, 2014. Alpha-lipoic acid improves acute deltamethrin-induced toxicity in rats. Can J Physiol Pharm 92, 773–779. [DOI] [PubMed] [Google Scholar]
- Al‐Sarar AS, Abobakr Y, Bayoumi AE, Hussein HI, Al‐Ghothemi M, 2014. Reproductive toxicity and histopathological changes induced by lambda‐cyhalothrin in male mice. Environmental toxicology 29, 750–762. [DOI] [PubMed] [Google Scholar]
- Albano E, Mottaran E, Occhino G, Reale E, Vidali M, 2005. role of oxidative stress in the progression of non‐alcoholic steatosis. Alimentary pharmacology & therapeutics 22, 71–73. [DOI] [PubMed] [Google Scholar]
- Aldana L, Tsutsumi V, Craigmill A, Silveira MI, de Mejia EG, 2001. α-Tocopherol modulates liver toxicity of the pyrethroid cypermethrin. Toxicology letters 125, 107–116. [DOI] [PubMed] [Google Scholar]
- Argo CK, Caldwell SH, 2009. Epidemiology and natural history of non-alcoholic steatohepatitis. Clinics in liver disease 13, 511–531. [DOI] [PubMed] [Google Scholar]
- Atessahin A, Yilmaz S, Karahan I, Pirincci I, Tasdemir B, 2005. The effects of vitamin E and selenium on cypermethrin-induced oxidative stress in rats. Turk J Vet Anim Sci 29, 385–391. [Google Scholar]
- Bechmann LP, Hannivoort RA, Gerken G, Hotamisligil GS, Trauner M, Canbay A, 2012. The interaction of hepatic lipid and glucose metabolism in liver diseases. Journal of Hepatology 56, 952–964. [DOI] [PubMed] [Google Scholar]
- Bloomquist JR, 1993. Toxicology, Mode of Action and Target Site-Mediated Resistance to Insecticides Acting on Chloride Channels. Comp Biochem Phys C 106, 301–314. [DOI] [PubMed] [Google Scholar]
- Bugianesi E, Gastaldelli A, Vanni E, Gambino R, Cassader M, Baldi S, Ponti V, Pagano G, Ferrannini E, Rizzetto M, 2005. Insulin resistance in non-diabetic patients with non-alcoholic fatty liver disease: sites and mechanisms. Diabetologia 48, 634–642. [DOI] [PubMed] [Google Scholar]
- Canga AG, Prieto AMS, Liébana MJD, Martínez NF, Vega MS, Vieitez JJG, 2008. The pharmacokinetics and interactions of ivermectin in humans—a mini-review. The AAPS journal 10, 42–46. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Cariou B, 2008. The farnesoid X receptor (FXR) as a new target in non-alcoholic steatohepatitis. Diabetes & metabolism 34, 685–691. [DOI] [PubMed] [Google Scholar]
- Carr RM, Reid AE, 2015. FXR Agonists as Therapeutic Agents for Non-alcoholic Fatty Liver Disease. Curr Atheroscler Rep 17. [DOI] [PubMed] [Google Scholar]
- Centis E, Marzocchi R, Suppini A, Dalle Grave R, Villanova N, Hickman IJ, Marchesini G, 2013. The role of lifestyle change in the prevention and treatment of NAFLD. Curr Pharm Des 19, 5270–5279. [PubMed] [Google Scholar]
- Chargui I, Grissa I, Bensassi F, Hrira MY, Haouem S, Haouas Z, Bencheikh H, 2012. Oxidative stress, biochemical and histopathological alterations in the liver and kidney of female rats exposed to low doses of deltamethrin (DM): a molecular assessment. Biomed Environ Sci 25, 672–683. [DOI] [PubMed] [Google Scholar]
- Cheng Y, Zhang K, Chen Y, Li Y, Li Y, Fu K, Feng R, 2016. Associations between Dietary Nutrient Intakes and Hepatic Lipid Contents in NAFLD Patients Quantified by 1H-MRS and Dual-Echo MRI. Nutrients 8. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Council NR, 1999. California Environmental Protection Agency Department of Toxic Substances Control (DTSC) Letter of Introduction, Overview, Concept Paper, and Appendices 1–4 from DTSC Report.
- Cousin SP, Hugl SR, Wrede CE, Kajio H, Myers MG, Rhodes CJ, 2001. Free fatty acid-induced inhibition of glucose and insulin-like growth factor I-induced deoxyribonucleic acid synthesis in the pancreatic beta-cell line INS-1. Endocrinology 142, 229–240. [DOI] [PubMed] [Google Scholar]
- Cremer JE, Seville MP, 1982. Comparative effects of two pyrethroids, deltamethrin and cismethrin, on plasma catecholamines and on blood glucose and lactate. Toxicol Appl Pharm 66, 124–133. [DOI] [PubMed] [Google Scholar]
- Gaggini M, Morelli M, Buzzigoli E, DeFronzo RA, Bugianesi E, Gastaldelli A, 2013. Non-alcoholic fatty liver disease (NAFLD) and its connection with insulin resistance, dyslipidemia, atherosclerosis and coronary heart disease. Nutrients 5, 1544–1560. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Ghose R, Omoluabi O, Gandhi A, Shah P, Strohacker K, Carpenter KC, McFarlin B, Guo T, 2011. Role of high-fat diet in regulation of gene expression of drug metabolizing enzymes and transporters. Life sciences 89, 57–64. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Giray B, Gurbay A, Hincal F, 2001. Cypermethrin-induced oxidative stress in rat brain and liver is prevented by Vitamin E or allopurinol. Toxicology letters 118, 139–146. [DOI] [PubMed] [Google Scholar]
- Gotoh Y, Kawakami M, Matsumoto N, Okada Y, 1998. Permethrin emulsion ingestion: clinical manifestations and clearance of isomers. Journal of Toxicology: Clinical Toxicology 36, 57–61. [DOI] [PubMed] [Google Scholar]
- Hossain MM, Richardson JR, 2011. Mechanism of Pyrethroid Pesticide-Induced Apoptosis: Role of Calpain and the ER Stress Pathway. Toxicological Sciences 122, 512–525. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Ince S, Kucukkurt I, Demirel HH, Turkmen R, Sever E, 2012. Thymoquinone attenuates cypermethrin induced oxidative stress in Swiss albino mice. Pesticide Biochemistry and Physiology 104, 229–235. [Google Scholar]
- Jaga K, Dharmani C, 2003. Global surveillance of DDT and DDE levels in human tissues. International Journal of Occupational Medicine and Environmental Health 16, 7–20. [PubMed] [Google Scholar]
- Jin LH, Feng XH, Rong H, Pan ZF, Inaba Y, Qiu L, Zheng WL, Lin SC, Wang R, Wang Z, Wang SS, Liu HY, Li S, Xie W, Li Y, 2013. The antiparasitic drug ivermectin is a novel FXR ligand that regulates metabolism. Nature Communications 4. [DOI] [PubMed] [Google Scholar]
- Jin LH, Wang R, Zhu YL, Zheng WL, Han YP, Guo FS, Bin Ye F, Li Y, 2015. Selective targeting of nuclear receptor FXR by avermectin analogues with therapeutic effects on nonalcoholic fatty liver disease. Sci Rep-Uk 5. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kalaany NY, Mangelsdorf DJ, 2006. LXRS and FXR: the yin and yang of cholesterol and fat metabolism. Annu Rev Physiol 68, 159–191. [DOI] [PubMed] [Google Scholar]
- Kim J, Park MY, Kim Y, Yoon KS, Clark JM, Park Y, Whang KY, 2017. 4,4 ‘-Dichlorodiphenyltrichloroethane (DDT) and 4,4 ‘-dichlorodiphenyldichloroethylene (DDE) inhibit myogenesis in C2C12 myoblasts. J Sci Food Agr 97, 5176–5185. [DOI] [PubMed] [Google Scholar]
- Kim J, Park Y, Yoon KS, Clark JM, Park Y, 2013. Imidacloprid, a neonicotinoid insecticide, induces insulin resistance. Journal of Toxicological Sciences 38, 655–660. [DOI] [PubMed] [Google Scholar]
- Kim J, Park Y, Yoon KS, Clark JM, Park Y, 2014. Permethrin AltersAdipogenesis in 3T3-L1 Adipocytes and Causes Insulin Resistance in C2C12 Myotubes. Journal of Biochemical and Molecular Toxicology 28, 418–424. [DOI] [PubMed] [Google Scholar]
- Krishan S, 2016. Correlation between non-alcoholic fatty liver disease (NAFLD) and dyslipidemia in type 2 diabetes. Diabetes Metab Syndr 10, S77–81. [DOI] [PubMed] [Google Scholar]
- Lage R, Diéguez C, Vidal-Puig A, López M, 2008. AMPK: a metabolic gauge regulating whole-body energy homeostasis. Trends in molecular medicine 14, 539–549. [DOI] [PubMed] [Google Scholar]
- Lee JS, Mendez R, Heng HH, Yang ZQ, Zhang K, 2012. Pharmacological ER stress promotes hepatic lipogenesis and lipid droplet formation. Am J Transl Res 4, 102–113. [PMC free article] [PubMed] [Google Scholar]
- Leung TM, Nieto N, 2013. CYP2E1 and oxidant stress in alcoholic and non-alcoholic fatty liver disease. J Hepatol 58, 395–398. [DOI] [PubMed] [Google Scholar]
- Malhi H, Bronk SF, Werneburg NW,Gores GJ, 2006. Free fatty acids induce JNK-dependent hepatocyte lipoapoptosis, J Biol Chem 281, 12093–12101. [DOI] [PubMed] [Google Scholar]
- Marchesini G, Brizi M, Bianchi G, Tomassetti S, Bugianesi E, Lenzi M, McCullough AJ, Natale S, Forlani G, Melchionda N, 2001. Nonalcoholic fatty liver disease: a feature of the metabolic syndrome. Diabetes 50, 1844–1850. [DOI] [PubMed] [Google Scholar]
- Miao K, Joyner WL, 1994. In-Situ Study of the Membrane-Potential in Microvascular Endothelial-Cells Using a Fluorescent-Probe. Microvasc Res 48, 135–142. [DOI] [PubMed] [Google Scholar]
- Mishra AK, Mishra S, Rajput C, ur Rasheed MS, Patel DK, Singh MP, 2018. Cypermethrin Activates Autophagosome Formation Albeit Inhibits Autophagy Owing to Poor Lysosome Quality: Relevance to Parkinson’s Disease. Neurotoxicity research 33, 377–387. [DOI] [PubMed] [Google Scholar]
- Mudaliar S, Henry RR, Sanyal AJ, Morrow L, Marschall HU, Kipnes M, Adorini L, Sciacca CI, Clopton P, Castelloe E, 2013. Efficacy and safety of the farnesoid X receptor agonist obeticholic acid in patients with type 2 diabetes and nonalcoholic fatty liver disease. Gastroenterology 145, 574–582. e571. [DOI] [PubMed] [Google Scholar]
- Narendra M, Kavitha G, Kiranmai AH, Rao NR, Varadacharyulu NC, 2008. Chronic exposure to pyrethroid-based allethrin and prallethrin mosquito repellents alters plasma biochemical profile. Chemosphere 73, 360–364. [DOI] [PubMed] [Google Scholar]
- Neuschwander-Tetri BA, 2010. Hepatic Lipotoxicity and the Pathogenesis of Nonalcoholic Steatohepatitis: the Central Role of Nontriglyceride Fatty Acid Metabolites. Hepatology 52, 774–788. [DOI] [PubMed] [Google Scholar]
- Oslowski CM, Urano F, 2011. Measuring Er Stress and the Unfolded Protein Response Using Mammalian Tissue Culture System. Method Enzymol 490, 71–92. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Özcan U, Cao Q, Yilmaz E, Lee A-H, Iwakoshi NN, Özdelen E, Tuncman G, Görgün C, Glimcher LH, Hotamisligil GS, 2004. Endoplasmic reticulum stress links obesity, insulin action, and type 2 diabetes. Science 306, 457–461. [DOI] [PubMed] [Google Scholar]
- Ozkucur N, Quinn KP, Pang JC, Du C, Georgakoudi I, Miller E, Levin M, Kaplan DL, 2015. Membrane potential depolarization causes alterations in neuron arrangement and connectivity in cocultures. Brain Behav 5. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Park Y, Kim Y, Kim J, Yoon KS, Clark J, Lee J, 2013. Imidacloprid, a neonicotinoid insecticide, potentiates adipogenesis in 3T3-L1 adipocytes. Journal of agricultural and food chemistry 61, 255–259. [DOI] [PubMed] [Google Scholar]
- Pemberton DJ, Franks CJ, Walker RJ, Holden-Dye L, 2001. Characterization of glutamate-gated chloride channels in the pharynx of wild-type and mutant Caenorhabditis elegans delineates the role of the subunit GluCl-alpha2 in the function of the native receptor. Mol Pharmacol 59, 1037–1043. [DOI] [PubMed] [Google Scholar]
- Postic C, Girard J, 2008. The role of the lipogenic pathway in the development of hepatic steatosis. Diabetes Metab 34, 643–648. [DOI] [PubMed] [Google Scholar]
- Ray DE, Cremer JE, 1979. Action of Decamethrin (a Synthetic Pyrethroid) on the Rat. Pesticide Biochemistry and Physiology 10, 333–340. [Google Scholar]
- Ricchi M, Odoardi MR, Carulli L, Anzivino C, Ballestri S, Pinetti A, Fantoni LI, Marra F, Bertolotti M, Banni S, Lonardo A, Carulli N, Loria P, 2009. Differential effect of oleic and palmitic acid on lipid accumulation and apoptosis in cultured hepatocytes. J Gastroenterol Hepatol 24, 830–840. [DOI] [PubMed] [Google Scholar]
- Sabin K, Santos-Ferreira T, Essig J, Rudasill S, Echeverri K, 2015. Dynamic membrane depolarization is an early regulator of ependymoglial cell response to spinal cord injury in axolotl. Developmental Biology 408, 14–25. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Scollon EJ, Starr JM, Godin SJ, DeVito MJ, Hughes MF, 2009. In Vitro Metabolism of Pyrethroid Pesticides by Rat and Human Hepatic Microsomes and Cytochrome P450 Isoforms. Drug Metabolism and Disposition 37, 221–228. [DOI] [PubMed] [Google Scholar]
- Shen PY, Hsieh TH, Yue YR, Sun QC, Clark JM, Park Y, 2017. Deltamethrin increases the fat accumulation in 3T3-L1 adipocytes and Caenorhabditis elegans. Food and Chemical Toxicology 101, 149–156. [DOI] [PubMed] [Google Scholar]
- Sparks TC, 2013. Insecticide discovery: An evaluation and analysis. Pesticide Biochemistry and Physiology 107, 8–17. [DOI] [PubMed] [Google Scholar]
- Sun Q, Qi W, Yang JJ, Yoon KS, Clark JM, Park Y, 2016a. Fipronil promotes adipogenesis via AMPKalpha-mediated pathway in 3T3-L1 adipocytes. Food and Chemical Toxicology 92, 217–223. [DOI] [PubMed] [Google Scholar]
- Sun QC, Clark JM, Park Y, 2017a. Environmental pollutants and type 2 diabetes: a review of human studies. Toxicol Environ Chem 99, 1283–1303. [Google Scholar]
- Sun QC, Peng Y, Qi WP, Kim Y, Clark JM, Kim D, Park Y, 2017b. Permethrin decreased insulin-stimulated AKT phosphorylation dependent on extracellular signal-regulated kinase-1 (ERK), but not AMP-activated protein kinase alpha (AMPK alpha), in C2C12 myotubes. Food and Chemical Toxicology 109, 95–101. [DOI] [PubMed] [Google Scholar]
- Sun QC, Qi WP, Xiao X, Yang SH, Kim D, Yoon KS, Clark JM, Park Y, 2017c. Imidacloprid Promotes High Fat Diet-Induced Adiposity in Female C57BL/6J Mice and Enhances Adipogenesis in 3T3-L1 Adipocytes via the AMPK alpha-Mediated Pathway. Journal of agricultural and food chemistry 65, 6572–6581. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Sun QC, Xiao X, Kim Y, Kim D, Yoon KS, Clark JM, Park Y, 2016b. Imidacloprid Promotes High Fat Diet-Induced Adiposity and Insulin Resistance in Male C57BL/6J Mice. Journal of agricultural and food chemistry 64, 9293–9306. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Swaminathan K, 2013. Pesticides and human diabetes: a link worth exploring? Diabetic Med. 30, 1268–1271. [DOI] [PubMed] [Google Scholar]
- Terada M, Motojima K, 2011. PPAR alpha plays a key role in whole-body lipid homeostasis by regulating transport and metabolism of unsaturated fatty acids in the liver. Chem Phys Lipids 164, S7–S7. [Google Scholar]
- Torra IP, Claudel T, Duval C, Kosykh V, Fruchart JC, Staels B, 2003. Bile acids induce the expression of the human peroxisome proliferator-activated receptor alpha gene via activation of the farnesoid X receptor. Mol Endocrinol 17, 259–272. [DOI] [PubMed] [Google Scholar]
- Trauner M, Claudel T, Fickert P, Moustafa T, Wagner M, 2010. Bile acids as regulators of hepatic lipid and glucose metabolism. Digestive diseases 28, 220–224. [DOI] [PubMed] [Google Scholar]
- Tuzmen N, Candan N, Kaya E, Demiryas N, 2008. Biochemical effects of chlorpyrifos and deltamethrin on altered antioxidative defense mechanisms and lipid peroxidation in rat liver. Cell Biochem Funct 26, 119–124. [DOI] [PubMed] [Google Scholar]
- Veerappan M, Hwang I, Pandurangan M, 2012. Effect of cypermethrin, carbendazim and their combination on male albino rat serum. Int J Exp Pathol 93, 361–369. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Woolsey SJ, Beaton MD, Mansell SE, Leon-Ponte M, Yu J, Pin CL, Adams PC, Kim RB, Tirona RG, 2016. A Fibroblast Growth Factor 21-Pregnane X Receptor Pathway Downregulates Hepatic CYP3A4 in Nonalcoholic Fatty Liver Disease. Mol Pharmacol 90, 437–446. [DOI] [PubMed] [Google Scholar]
- Xiang D, Chu T, Li M, Wang Q, Zhu G, 2018. Effects of pyrethroid pesticide cis-bifenthrin on lipogenesis in hepatic cell line. Chemosphere 201, 840–849. [DOI] [PubMed] [Google Scholar]
- Xiao X, Clark JM, Park Y, 2017a. Potential contribution of insecticide exposure and development of obesity and type 2 diabetes. Food and Chemical Toxicology 105, 456–474. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Xiao X, Kim Y, Kim D, Yoon KS, Clark JM, Park Y, 2017b. Permethrin alters glucose metabolism in conjunction with high fat diet by potentiating insulin resistance and decreases voluntary activities in female C57BL/6J mice. Food and Chemical Toxicology 108, 161–170. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Xiao X, Qi WP, Clark JM, Park Y, 2017c. Permethrin potentiates adipogenesis via intracellular calcium and endoplasmic reticulum stress-mediated mechanisms in 3T3-L1 adipocytes. Food and Chemical Toxicology 109, 123–129. [DOI] [PubMed] [Google Scholar]
- Xiao X, Sun Q, Kim Y, Yang SH, Qi W, Kim D, Yoon KS, Clark JM, Park Y, 2018. Exposure to permethrin promotes high fat diet-induced weight gain and insulin resistance in male C57BL/6J mice. Food and Chemical Toxicology 111, 405–416. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Yang JS, Park Y, 2018. Insecticide Exposure and Development of Nonalcoholic Fatty Liver Disease. Journal of agricultural and food chemistry 66, 10132–10138. [DOI] [PubMed] [Google Scholar]
- Yang JS, Symington S, Clark JM, Park Y, 2018. Permethrin, a pyrethroid insecticide, regulates ERK1/2 activation through membrane depolarization-mediated pathway in HepG2 hepatocytes. Food and Chemical Toxicology 121, 387–395. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Yavasoglu A, Sayim F, Uyanikgil Y, Turgut M, Karabay-Yavasoglu NÜ, 2006. The pyrethroid cypermethrin-induced biochemical and histological alterations in rat liver. J Health Sci 52, 774–780. [Google Scholar]
- Yki-Jarvinen H, 2010. Liver Fat in the Pathogenesis of Insulin Resistance and Type 2 Diabetes. Digestive Diseases 28, 203–209. [DOI] [PubMed] [Google Scholar]
- Younossi ZM, Koenig AB, Abdelatif D, Fazel Y, Henry L, Wymer M, 2016. Global epidemiology of nonalcoholic fatty liver disease-Meta-analytic assessment of prevalence, incidence, and outcomes. Hepatology 64, 73–84. [DOI] [PubMed] [Google Scholar]
- Yousef MI, Awad TI, Mohamed EH, 2006. Deltamethrin-induced oxidative damage and biochemical alterations in rat and its attenuation by Vitamin E. Toxicology 227, 240–247. [DOI] [PubMed] [Google Scholar]
- Zhang Y, Lu M, Zhou P, Wang C, Zhang Q, Zhao M, 2015. Multilevel evaluations of potential liver injury of bifenthrin. Pesticide biochemistry and physiology 122, 29–37. [DOI] [PubMed] [Google Scholar]
- Zhu W-J, Li M, Liu C, Qu J-P, Min Y-H, Xu S-W, Li S, 2013. Avermectin induced liver injury in pigeon: mechanisms of apoptosis and oxidative stress. Ecotox Environ Safe 98, 74–81. [DOI] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.