

Abstract
Daidzein (DAZ), a dominant isoflavone in various natural products such as soybeans, has been gaining attention due to the beneficial health effects (e.g., protection against cancer and diabetes) of its metabolites. Our major hypothesis was that dietary exposure to the soy phytoestrogen DAZ could modulate the immune responses toward a protective effect and lead to improved metabolic functions (such as glucose metabolism). In this study, we applied complementary mouse models, the hybrid B6C3F1 and inbred type 1 diabetes prone non-obese diabetic (NOD) mice, to investigate if DAZ exposure modulated the immune responses. The animals were orally administered DAZ at various physiological doses (2 - 20 mg/kg body weight) during adulthood. DAZ significantly altered the relative organ weights in female B6C3F1 mice and decreased the B cell population (represented by CD3−IgM+), while the T cell populations (represented by CD3+IgM−, CD4+CD8− and CD4−CD8+) were increased. In addition, DAZ dosing produced a decrease in the percentage of late apoptotic thymocytes. However, the activities cytotoxic T cells and natural killer cells were not altered in the B6C3F1 mice. In NOD mice, the blood glucose level and glucose tolerance were not affected by DAZ exposure, but DAZ modulated the antibody production, as shown by increased levels of IgG2b in NOD females and IgG1 in NOD males. Further, DAZ increased CD8+CD25+ splenocytes in NOD females. Taken together, DAZ induced an immunomodulatory effect in both NOD and B6C3F1 mouse strains; however, minimal effects on glucose homeostasis were observed.
Keywords: Daidzein, antibody, blood glucose level, immune responses, apoptosis
Introduction
Polyphenolics are naturally occurring compounds found in various fruits, vegetables, and beverages [1]. Soy-based food is of importance to study not only because it is an intrinsic part of Asian cuisine, but also that it takes up 25% of the infant formula market [2]. Soy is a major contributor of polyphenols, including daidzein (DAZ), genistein (GEN) and glycitein [3]. Isoflavone DAZ is found in an equal amount as GEN in soy [4–6]; however, DAZ is less studied than GEN. The concentration of DAZ in soy food varies and depends on the processing procedures of soy product (e.g., 6 mg per 4 oz serving in boiled soybean, and 34 mg per 4 oz serving in meso) [7]. One case-control study demonstrated that high serum DAZ level (up to 68 ng/mL) was associated with a reduced ovarian cancer risk among Japanese women [8]. In a nested case-control study among US women, urinary DAZ level correlated with a protective effect against type 2 diabetes (T2D; odds ratio = 0.71, 95% CI 0.55-0.93)[9]. In animal studies, it has been shown that DAZ exerted a protective effect on hyperglycemia and glucose tolerance that is comparable to GEN in non-obese diabetic (NOD) mice, a model of the autoimmune type 1 diabetes (T1D), suggesting a potential immunomodulatory effect [3]. A literature review suggested that DAZ could mitigate some adverse health effects induced by GEN such as proliferation of mammary cancer cells in rodent models [10]. DAZ can be metabolized into dihydrodaidzein, equol, or O-desmethylangolensin (ODMA) [11]. Equol has a high affinity to the estrogen receptors (ERs) [11], and ODMA and equol are closely associated with obesity [12, 13] and cardiometabolic risk [14,15]. For example, in humans, ODMA non-producer phenotype is positively correlated with obesity [12]. In one systematic literature review, it was shown that high DAZ intake was associated with a 34% reduced risk of breast cancer, while the same association was not found for equol [16]. The same study concluded that the risk for T2D was reduced by 19% following high amount of DAZ intake.
It is well known that estrogenic compounds can potentially impact the immune system through both estrogen receptor-dependent and independent mechanisms [17]. Immune dysregulation not only serves as a hallmark of autoimmune disease (e.g., T1D, rheumatoid arthritis) [18], but also directly contributes to the pathogenesis of T2D [19, 20], obesity [19], ovarian cancer [21] and breast cancer [22]. With regards to the effects on immune system, equol reduced the inflammatory response and down-regulated IL-6 mRNA expression in a mouse rheumatoid arthritis model [23], and ODMA demonstrated a similar effect on immunity [24]. In another mouse study, low dose DAZ administration (20 mg/kg) did not affect experimental allergic encephalomyelitis (EAE) development while high dose DAZ (300 mg/kg) alleviated EAE symptoms by modulating T cell responses [25]. It has been reported that mean estrogen levels in female NOD mice is lower than other strains [26], such as female BALB/c mice [27]. Female B6C3F1 mice have higher and more sustained level of endogenous estradiol than female C57BL/6 and CD-1 mice [28]. We hypothesized that dietary exposure to the soy phytoestrogen DAZ could modulate the immune responses toward a beneficial effect (e.g., an-inflammatory and increasing regulatory cell subtypes) and lead to improved metabolic functions (such as glucose metabolism). Here in this study, we employed both B6C3F1 and NOD mouse strains as complementary models to study the immunomodulatory effect of DAZ.
Method
B6C3F1 and NOD mouse models
B6C3F1 mice are the offspring of a cross between C57BL/6J (B6) females and C3H/HeJ (C3) males. Adult female B6C3F1 mice (Taconic, Germantown, NY) used for the study were fed with either a soy-based diet (NTP-2000 diet, Zeigler Bros, Inc., USA) or a soy-free diet (5K96, TestDiet, St. Louis, MO), and adult male and female NOD mice (Taconic) were maintained on the soy-free diet. Mice (8-12 wks old) were randomized into vehicle control (VH) and DAZ groups based on body weight (BW) and baseline blood glucose levels (BGLs; for NOD). DAZ (CAS# 486-66-8; Mol. Wt. 254.24; Product Number: D-2946) was obtained from LC Laboratories (Woburn, MA) and contained >99% DAZ. The DAZ doses used in this study were 20 mg/kg BW or lower because they were physiologically relevant [3, 10, 26, 29]. In addition, we were able to detect a difference in BGLs at these doses for GEN in different murine diabetic models [26, 29], and it has been reported that DAZ has similar effects for BGL modulation as GEN [3]. Mice in DAZ groups were dosed with 0.1 ml DAZ solution/10 g BW (0.2-2 mg/mL DAZ dissolved in 25 mM Na2CO3 by sonication due to its low water solubility or in 0.2% methylcellulose/0.1% Tween 80) while the VH groups were dosed with the same volume of vehicle daily. A 10G gavage needle was used for dosing. Mice are given food and water ad libitum. The protocol has been approved by IACUC at the University of Georgia and Virginia Commonwealth University. The mice were housed in standard plastic cages with irradiated laboratory animal bedding (The Andersons company, Maumee, Ohio) with each cage housing 2-6 mice according to facility requirements. The animal room was kept at 22–25 °C, 50 ± 10 in relative humidity with 12/12 h light/dark cycle. As the NOD females develop T1D sooner than the males, they were kept for a shorter period of time than the males.
CTL and NK activity
B6C3F1 mice were injected (i.p.) with 10 × 106 viable P815 tumor cells on day 18 of DAZ exposure. Ten days after P815 injection, mice were euthanized by CO2 asphyxiation, spleens were removed and splenocytes obtained. Cytotoxic T lymphocyte (CTL) activity was determined using a 4-h 51Cr release assay using Na51CrO4-labeled P815 cells as the target. The activity of natural killer (NK) cells was assayed using the NK-sensitive target, Na51CrO4-labeled YAC-1 cells, as previously described [30]. Briefly, the splenocytes at different dilutions were mixed with the target cells to obtain effector:target (E:T) ratios of 200:1, 100:1, 50:1, 25:1, 12.5:1 and 6.25:1. The spontaneous release was determined by adding 100 μL of medium to 12 replicate cultures containing the targets. The maximum release was determined by adding 100 μL of the target cells and 100 μL 0.1% Triton X-100 to each of 12 replicate wells. Specific lysis (%) of 51Cr-labeled target cells was used as the endpoint of the assay.
BW, BGL measurement, glucose tolerance test (GTT), and insulin tolerance test (ITT)
The measurements of BW and BGL were conducted as previously described [29]. GTT and ITT were performed by injecting (i.p.) the NOD mice with glucose (2 g/kg BW) following fasting for 15 hours or insulin (1.5 IU/kg BW) without fasting, respectively [29]. The BGLs at 0, 15, 30, 60 and 120 minutes following initial injection were measured subsequently.
Enzyme-linked immunosorbent assay (ELISA)
ELISA Quantitation Kits (Bethyl Laboratories Inc., Montgomery, TX) were used to measure the levels of serum antibodies (IgG2b, IgM, IgG1) in NOD mice as previously described [18, 29]. Briefly, the 96 well flat-bottom high binding microplate was coated with 100 μl/well primary antibodies (10 μg/ml) at 4 °C for overnight (16-18 h), and then blocked with 5% skim milk powder in 0.05% PBST at room temperature for 2 h. After washing with 0.05% PBST, a 100 μl sample was added. After 2 h incubation at room temperature, the plate was washed and a 100 μl HRP-conjugated secondary antibody was applied. The plate was incubated at room temperature at dark for 1 h, and then washed. Substrate 3,3’,5,5’-Tetramethylbenzidine (TMB, Sigma) and stopping solution sulfuric acid (2N) were added at 100 μL/well, and the plate was read at the wavelength of 450 nm.
Flow cytometry
The spleens were collected upon euthanasia, and the flow cytometric analysis was performed following our previous publication [29]. Briefly, the spleens were mashed in 3 mL PBS solution on ice. Leukocyte population in NOD mice were characterized with different combinations of fluorochrome-labeled antibodies (diluted 1:80-1:100; BD PharMingen, San Diego, CA) including cluster of differentiation (CD) 4-CD8-CD25 (PE-PerCP-FITC), CD40L-B220 (PE-FITC) CD40-CD44 (PE-FITC) and CD5-CD24 (PE-FITC).
For the B6C3F1 females, the antibodies included IgM-CD3 (FITC-PE), CD4-CD8-CD25 (PE-PerCP-FITC), NK1.1-CD3 (PE-FITC), and Gr-1-Mac-3 (FITC-PE). In addition, the analyses of mitochondrial transmembrane potential (ΔΨm) and reactive oxygen species (ROS) generation were performed following our previous publication [18]. Thymocytes (1 × 106 cells/ml) were stained for 15 min with 40 nM 3,3’-dihexyloxacarbocyanine (DiOC6(3); Life Technologies) and 2 μM hydroethidine (HE, Life Technologies) for assessing ΔΨm and ROS generation. Following excitation at 488 nm (250 mW), emission was monitored through a 530/30 nm bandpass filter for DiOC6(3) and 575/26 nm bandpass filter for HE, and then logarithmic amplification was used to detect the fluorescence. The late apoptosis cell population was represented by DiOC6(3)dimEthbright cells.
Statistical analysis
The data were expressed as the mean ± SEM. Dunnett’s test was used to compare the means among treatment groups when the equal variance assumption was met; otherwise, Wilcoxon test was performed to compare the means. JMP Pro 14 (SAS Inc., Cary, NC) and GraphPad Prism 7 (GraphPad Software Inc., La Jolla, CA) were used for statistical analysis and data visualization.
Results
DAZ reduced organ weights and altered the splenocyte subpopulations in female B6C3F1 mice
Although oral DAZ exposure at 2 - 20 mg/kg had no effects on the terminal body weight and absolute organ weights (Table 1). DAZ at various doses induced reductions in the relative weights of spleen (2, 6 and 20 mg/kg), liver (6 and 20 mg/kg), lungs (20 mg/kg), and kidneys (6 mg/kg). When the splenic leukocyte populations were evaluated following DAZ dosing (Table 2), the percentage of total B cells (represented by CD3−IgM+) was reduced with significant change observed at the 20 mg/kg dose (P = 0.0388), while the %total T cells (represented by CD3+IgM−) were significantly increased at this dose (P = 0.0004). CD4-CD8-CD25 marker staining revealed that both the CD4+CD8− helper T cell population (P = 0.038 at 20 mg/kg) and CD4−CD8+ cytotoxic T cell population (P < 0.05 at 6 and 20 mg/kg) were increased. In contrast, the %splenic neutrophils were significantly reduced at 20 mg/kg (P = 0.0063). No significant alteration was found for macrophage population (data not shown).
Table 1.
The body and organ weights of adult B6C3F1 females following daidzein exposure.
| Parameters | Body weight (g) | Lung (mg) (%) | Thymus (mg) (%) | Spleen (mg) (%) | Kidneys (mg) (%) | Liver (mg) (%) | |
|---|---|---|---|---|---|---|---|
| DZF | VHF | 25.40 ± 1.68 | 189.38 ± 6.63 | 54.50 ± 5.68 | 72.50 ± 5.31 | 339.25 ± 7.95 | 945.63 ± 42.58 |
| 0.76 ± 0.04 | 0.22 ± 0.02 | 0.29 ± 0.01 | 1.36 ± 0.06 | 3.77 ± 0.14 | |||
| 2 mg/kg | 29.79 ± 1.41 | 204.50 ± 7.71 | 64.00 ± 4.77 | 69.13 ± 3.72 | 358.25 ± 14.33 | 1045.88 ± 58.81 | |
| 0.69 ± 0.02 | 0.21 ± 0.01 | 0.23 ± 0.00** | 1.21 ± 0.04 | 3.51 ± 0.08 | |||
| 6 mg/kg | 30.87 ± 1.50 | 203.75 ± 10.74 | 64.00 ± 4.91 | 66.63 ± 1.76 | 356.13 ± 13.24 | 999.38 ± 45.36 | |
| 0.66 ± 0.03 | 0.21 ± 0.01 | 0.22 ± 0.01** | 1.16 ± 0.02* | 3.25 ± 0.09** | |||
| 20 mg/kg | 31.34 ± 3.06 | 174.75 ± 8.47 | 69.38 ± 4.57 | 67.25 ± 4.53 | 376.13 ± 16.09 | 1002.88 ± 75.92 | |
| 0.58 ± 0.05** | 0.23 ± 0.01 | 0.22 ± 0.01** | 1.25 ± 0.07 | 3.25 ± 0.09** | |||
Mice were maintained on a soy-based diet and administered vehicle (25 mM Na2CO3 or daidzein by gavage daily for 14 days. At study termination, the mice were necropsied and indicated organs weighed. VHF = females exposed to vehicle; DZF = females exposed to daidzein. Values represent the mean ± SE (N = 8).
p ≤ 0.05
p ≤ 0.01
Table 2.
The splenic subpopulations following daidzein exposure in adult B6C3F1 females.
| Spleen | NK (%) | IgM+ (%) | CD3+(%) | Neutrophils (%) | CD4+CD25+ (%) | CD4−CD8+ (%) | CD4+CD8− (%) | |
|---|---|---|---|---|---|---|---|---|
| DZF | VHF | 1.98 ±0.48 | 38.28 ±4.19 | 16.51 ±2.68 | 2.16 ±0.36 | 3.17 ±0.54 | 4.04 ± 0.65 | 9.85 ± 1.20 |
| 2 mg/kg | 1.74 ±0.20 | 34.99 ± 1.80 | 21.53 ± 2.12 | 1.66 ±0.40 | 2.58 ±0.67 | 4.39 ± 0.44 | 10.15 ± 1.08 | |
| 6 mg/kg | 2.03 ±0.25 | 34.28 ±2.50 | 21.70 ±4.13 | 1.90 ±0.37 | 3.28 ± 0.68 | 6.83 ± 1.07* | 14.14 ± 1.89 | |
| 20 mg/kg | 2.46 ± 0.09 | 29.12 ± 1.41* | 35.00 ± 2.57** | 1.12 ±0.08* | 2.18 ± 0.12 | 9.42 ± 0.50** | 15.58 ± 1.82* | |
Mice were maintained on a soy-based diet and administered vehicle (25 mM Na2CO3) or daidzein by gavage daily for 14 days, and the percentages of splenocyte subsets were determined using flow cytometric analysis. VHF = females exposed to vehicle; DZF = females exposed to daidzein. Values represent the mean ± SE (N = 8).
p ≤ 0.05
p ≤ 0.01
DAZ affected thymocyte differentials and ΔΨm and ROS generation in female B6C3F1 mice
Two sets of cell surface markers were employed to determine thymocyte differentials following exposure to DAZ. Thymocytes were first stained for CD4 and CD8 markers. When compared to VH mice, DAZ treatment significantly decreased %CD4+CD8− thymocytes at 2 mg/kg (Table 3). The expressions of CD44 and CD25 by thymocytes were also determined by flow cytometric analysis. When compared to VH mice, DAZ treatment significantly decreased %CD44+CD25+ at low dose and %CD44−CD25+ thymocytes at all three doses (Table 3). In contrast, %CD44−CD25− was increased by DAZ with significant changes observed at the low dose (Table 3).
Table 3.
Effects of daidzein on thymocyte subsets in adult female B6C3F1 mice
| Thymus | CD4−CD8− (%) | CD4+CD8+ (%) | CD4+CD8− (%) | CD4−CD8+ (%) | |
|---|---|---|---|---|---|
| DZF | VHF | 28.91 ± 8.89 | 52.33 ± 9.16 | 14.79 ± 1.35 | 3.96 ± 0.59 |
| 2 mg/kg | 49.24 ± 6.24 | 37.41 ± 5.84 | 10.40 ± 1.05* | 2.96 ± 0.36 | |
| 6 mg/kg | 31.92 ± 6.11 | 51.37 ± 6.18 | 13.01 ± 1.34 | 3.70 ± 0.38 | |
| 20 mg/kg | 44.20 ± 4.37 | 40.71 ± 4.69 | 11.77 ± 0.72 | 3.32 ± 0.24 | |
| Thymus | CD44+CD25− (%) | CD44+CD25+ (%) | CD445−CD25+ (%) | CD445−CD255− (%) | |
| DZF | VHF | 2.21 ± 0.27 | 3.50 ± 0.74 | 3.29 ± 0.70 | 91.01 ± 1.47 |
| 2 mg/kg | 1.97 ± 0.23 | 1.73 ± 0.27* | 0.97 ± 0.18** | 95.33 ± 0.62* | |
| 6 mg/kg | 2.28 ± 0.23 | 2.37 ± 0.26 | 1.81 ± 0.20* | 93.54 ± 0.48 | |
| 20 mg/kg | 2.70 ± 0.17 | 2.53 ± 0.40 | 1.87 ± 0.28* | 92.90 ± 0.70 | |
Mice were maintained on a soy-based diet and administered vehicle (25 mM Na2CO3) or daidzein by gavage daily for 14 days, and the percentages of thymocyte subsets were determined using flow cytometric analysis. VHF = females exposed to vehicle; DZF = females exposed to daidzein. Values represent the mean ± SE (N = 8).
p ≤ 0.05
p ≤ 0.01
To shed some light on the mechanisms underlying the modulatory effects of DAZ on the development of thymocytes, the ΔΨm and ROS generation were evaluated following exposure to DAZ at the middle dose (6 mg/kg). DiOC6(3)dimEthdim cells has been shown to be the early apoptotic cells while DiOC6(3)dimEthbright cells the late apoptotic cells [18]. As shown in Figure 1, DAZ dosing induced a decrease in percentage of late apoptotic cells (P = 0.0092), while no differences were observed in early apoptotic cells (28.7 ± 3.9% vs. 28.7 ± 1.5%) and live cells (DiOC6(3) brightEthdim; 58.1 ± 4.1% vs. 64.2 ± 1.7%).
Figure 1.
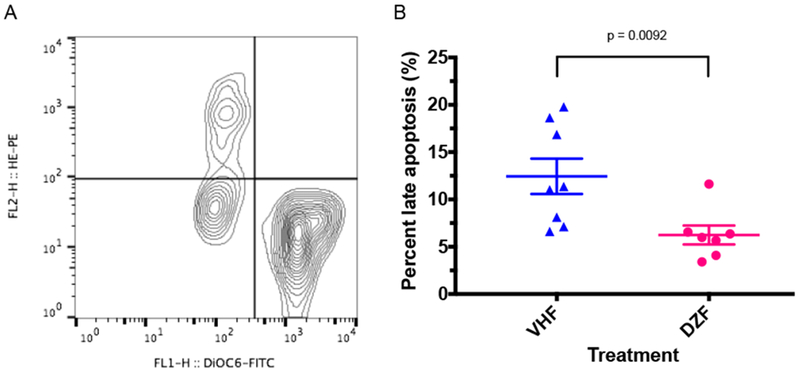
The ΔΨm and ROS generation in the thymocytes of female B6C3F1 mice maintained on a soy-based diet and dosed with daidzein (6 mg/kg). (A) A representative contour plot (DiOC6(3) vs. hydroethidine) of flow cytometric analysis; (B) %Late apoptotic cells were represented by DiOC6(3)dimEthbright. The vehicle was 0.2% methylcellulose/0.1% Tween 80. VHF = females exposed to vehicle; DZF=females exposed to daidzein; ROS = reactive oxygen species. N=7-8.
DAZ did not affect the activities of CTLs and NK cells in female B6C3F1 mice
Our flow cytometric analysis showed both %CD4+CD8− and %CD4−CD8+ splenic T cells were increased by DAZ (Table 2). Therefore, the CTL activity was examined using P815 mastocytoma cells as a target, and exposure to DAZ did not significantly affect the CTL activity at the 6 mg/kg dose (Figure 2A). In addition, when the effect of DAZ on the NK cell activity was evaluated, no significant changes were observed at any E:T ratios (Figure 2C). To determine if the phytoestrogens in the soy-based diet interacted with DAZ, adult female B6C3F1 mice were also exposed to DAZ at the dose of 20 mg/kg while on a phytoestrogen-free 5K96 diet. Similarly, DAZ had no effects on either CTL (Figure 2B) or NK (Figure 2D) activities in these mice as the adult female B6C3F1 mice on the soy-based diet.
Figure 2.
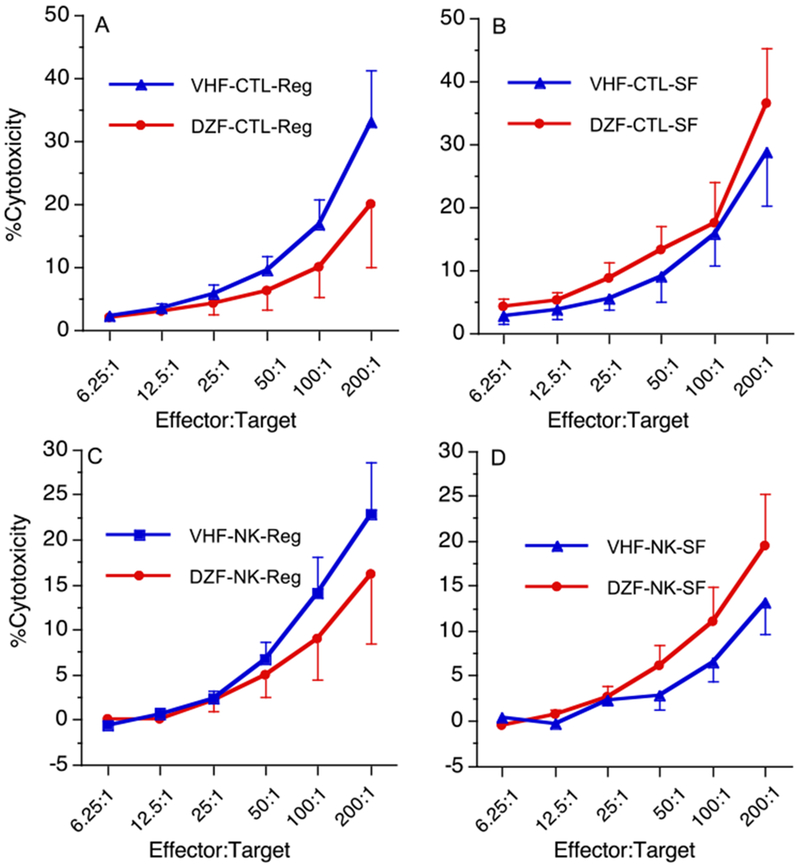
Effects of daidzein on the activities of CTLs (A, B) and NK cells (C, D) in adult female B6C3F1 mice. Values represent the mean ± SE derived from 7-8 animals. Statistical analysis was conducted as described. (A, C) Mice were maintained on a soy-based diet (Reg) and gavaged with daidzein at the 6 mg/kg dose, and (B, D) mice were maintained on a phytoestrogen-free diet (SF) and gavaged with daidzein at the 20 mg/kg dose. The vehicle was 0.2% methylcellulose/0.1% Tween 80. CTL = cytotoxic T lymphocyte; NK = natural killer cells. VHF = females exposed to vehicle; DZF = females exposed to daidzein. N=7-8.
BGL and tolerance tests did not show a strong effect in DAZ-exposed NOD mice
Our studies indicated that the 20 mg/kg dose of GEN could effectively induce immunomodulation in adult female B6C3F1 mice [18, 31] and decrease BGL in adult NOD mice [26, 29]. Thus, this dose of DAZ was studied for its effects in the NOD mice. DAZ exposure led to a reduction of BW in NOD males (Figure 3B) with statistical significances observed between days 88 - 109 following initial treatment, but not in females (Figure 3A). However, the blood glucose level with time (Figure 3C-D) did not show a strong effect on T1D protection in either sex. To further ascertain these findings, glucose tolerance test was conducted on days 76 following the first DAZ treatment for females (Figure 4A) and day 169 for males (Figure 4B). In addition, insulin tolerance test was conducted on day 165 (Figure 4C) for males. The ITT for NOD females were not performed due to a generally high T1D incidence and possible death of animals following insulin injection. All together, these data did not show a strong effect on T1D protection in either sex.
Figure 3.
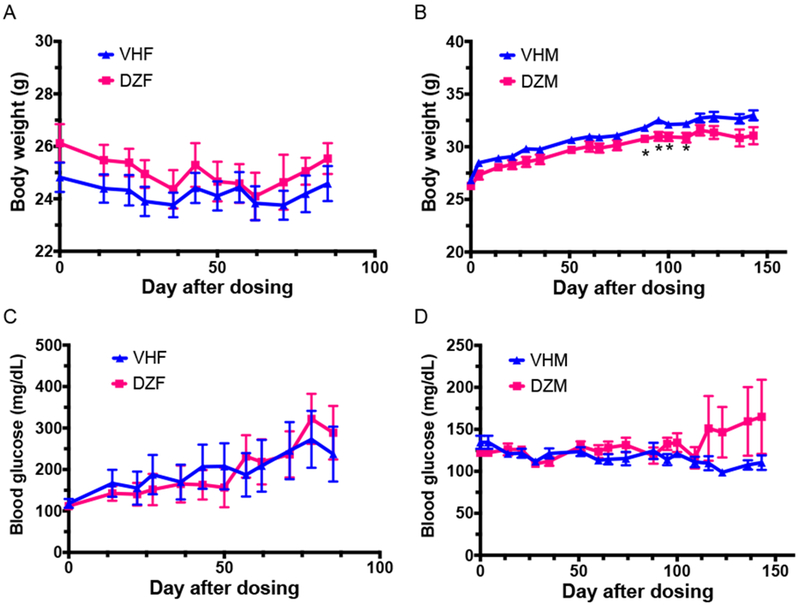
The body weight (BW) change of (A) NOD females and (B) NOD males, and the blood glucose levels (BGL) with time in NOD females (C) and males (D). Mice were maintained on the soy-free diet. VHM = males dosed with vehicle (N=12), DZM = males dosed with daidzein (N=12), VHF = females dosed with vehicle (N=10), and DZF = females dosed with daidzein (N=11)
Figure 4.
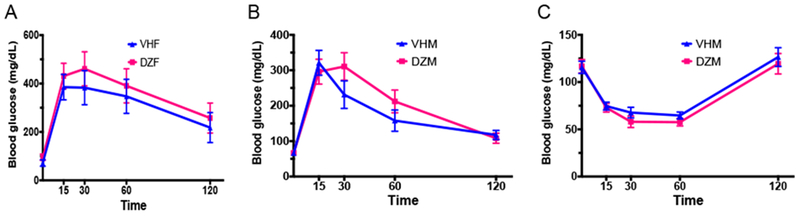
The glucose tolerance test (GTT) in NOD females (A) and NOD males (B), and the insulin tolerance test (ITT) in NOD males (C). Mice were maintained on the soy-free diet. VHM = males dosed with vehicle (N=12), DZM = males dosed with daidzein (N=10), VHF = females dosed with vehicle (N=7), and DZF = females dosed with daidzein (N=7).
DAZ exposure induced an increased level of serum IgG2b in NOD females and IgG1 in NOD males
Figure 5 demonstrates the alterations of serum antibodies, including IgG2b, IgM and IgG1, following DAZ exposure. DAZ dosing led to significant increases in IgG2b levels in females (P = 0.0211, Figure 5A) and IgG1 levels in males (P = 0.0166, Figure 5F). The other antibodies were not significantly altered by DAZ dosing (Figure 5B-E). IgG2a was not measured because the gene was deleted in NOD mice [32, 33].
Figure 5.
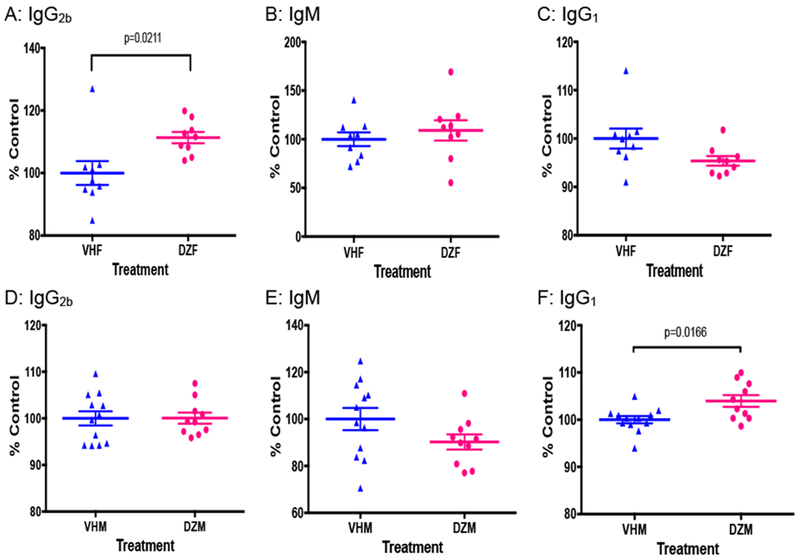
The serum levels of IgG2b (A), IgM (B), IgG1 (C) in NOD females, and IgG2b (D), IgM (E), IgG1 (F) in NOD males. Mice were maintained on the soy-free diet. VHM = males dosed with vehicle (N=12), DZM = males dosed with daidzein (N=10), VHF = females dosed with vehicle (N=9), and DZF = females dosed with daidzein (N=9).
DAZ increased CD8+CD25+ splenocytes in NOD females
Flow cytometric analysis demonstrated an increase in the percentage of CD8+CD25+ cells (Figure 6A, P = 0.0268) in spleens from NOD females following DAZ dosing (P = 0.0268). These cells are possibly regulatory T cells [34]. DAZ dosing also increased CD4−CD25+% (data not shown), possibly due to increased % CD8+CD25+ cells. In addition, the percentage of B220+CD40L+ cells in NOD females was increased (Figure 6B). No differences were observed for other splenocyte subtypes including CD40CD44 and CD5CD24 markers (data not shown).
Figure 6.
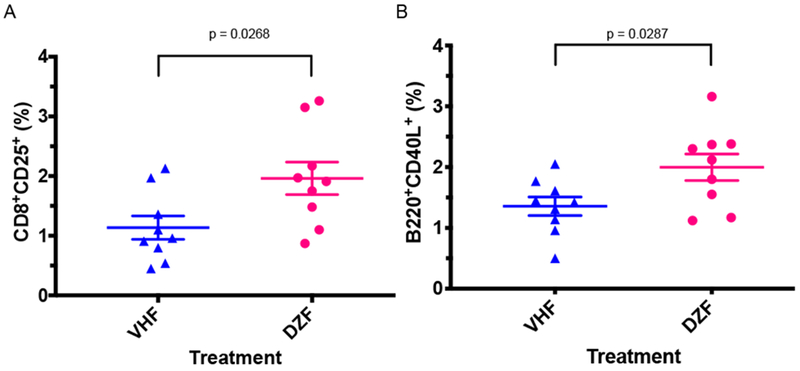
Percentage of CD8+CD25+ (A) and B220+CD40L+ (B) splenocytes following daidzein treatment in NOD females. Mice were maintained on the soy-free diet. VHF = females exposed to vehicle (N=9), DZF=females exposed to daidzein (N=9).
Discussion
DAZ and GEN are the major isoflavones in soy-based food, and they exert estrogen-like effects. In most soy products (such as boiled soybean and tofu), DAZ content is equivalent to GEN, but much less studied. The mechanisms of action for DAZ could be related to the inhibitory effects on α-glucosidase and α-amylase [35], promotion of 5’ AMP-activated protein kinase (AMPK) phosphorylation and glucose transporter type 4 (GLUT4) translocation [36], inhibition of glucose uptake in adipocytes [37], enhancement of the epithelial function and reducing oxidative stress [38], increase of peroxisome proliferator-activated receptor (PPAR)-γ transcription [39], upregulation of ER-β [40], increase of the insulin/glucagon ratio and regulation of hepatic lipid regulating enzymes (such as hepatic fatty acid synthase) [35], and/or enhancement of IL-17 gene expression [41]. Overall, the effect of DAZ is more of interest in veterinary sciences (e.g., cows, pigs and chicken) while both human epidemiological and mechanistic studies are largely missing.
The majority of studies focused on microbial-derived DAZ metabolites (e.g., ODMA and equol) because of their particular protective effect against obesity, while the parent compound has not been well studied (partly due to its low solubility and low bioavailability). In this study, we used 25 mM Na2CO3 as a vehicle to achieve better solubility in basic solution [42], and this vehicle has been consistently used in our published studies with GEN [18, 29] and by other groups [42–44]. We used two complementary models (NOD and B6C3F1) to investigate the immunological alterations following DAZ exposure and to assess the anti-diabetic effects of DAZ. Similar to our study, it has been shown that DAZ exerted a stimulatory effect on innate immunity and acquired immunity (e.g., IgM-secreting cells and peripheral T cells) in mice at the same dose used in our study [45]. Although the effects on splenic subpopulations following DAZ exposure in B6C3F1 females (Table 2) were in a dose-related manner, some low dose effects were observed in thymocytes (Table 3). This bimodal effect has been reported for hormones and endocrine disruptors [46]. However, it has also been reported that DAZ induced suppression of dendritic cells’ maturation and functions in rats [47]; therefore, further immunological studies using different species by focusing on gut microbiota are warranted.
In male rats, it was reported that DAZ could mitigate lipopolysaccharide-induced immune responses represented by decreased neutrophils, tumor necrosis factor (TNF)-α, macrophages and NF-κB markers [48], suggesting an anti-inflammatory response. Similar findings that DAZ exposure suppressed pro-inflammatory cytokines such as IL-12p40, IL-6 and TNF-α has also been reported in vitro[47]. Importantly, we observed an increase in IgG1 in NOD males following DAZ exposure, indicating a stronger Th2 response. Thus, it will be important to further study the Th1/Th2 balance following DAZ exposure. In NOD females following DAZ exposure, a numeric decrease in IgG1 corresponding to a decreased Th2 response was observed, which was in agreement with a decreased B cell population in B6C3F1 females. Fan and colleagues [49] observed an increase in IgG following exposure to DAZ at 20 mg/kg BW in chicken breeders. In lactating cows, both 300 mg/kg and 400 mg/kg DAZ increased serum IgG [50]. Consistently, we observed an increase in IgG2b in NOD females. It should be noted that the IgG2b level was not altered by GEN in NOD females following a similar exposure. Interestingly, similar differential effects were observed in NOD males in which GEN exposure produced a decreased IgG2b, while IgG1 level was not changed [29]. Therefore, the phytoestrogen GEN and DAZ could have different mechanisms of immune regulation, which depended on sex.
B cells are essential in maintaining the immune homeostasis against pro-inflammatory responses. It has been speculated by Hampe [51] that T1D development is initiated by B cell and followed by T cell response. Ectopic CD40 ligand expression on B cells triggers intestinal inflammation [52]. In NOD mice, some insulin-binding B cells are B220-low or -negative [53]. Our study suggested a reduced total B cell population in female B6C3F1 mice, while the percentage of B220+CD40L+ cells in NOD females was increased. Moreover, our studies suggested that the T cells (total, cytotoxic and helper T cells) were significantly increased in female B6C3F1 mice by DAZ, which were further supported by our findings in NOD females that the CD8+CD25+ population was greatly increased. In this study, the lack of diabetes development and T1D exacerbation in the presence of T cell increase following DAZ exposure might be due to the up-regulated CD8+CD25+ and B220+CD40L+ cells, which possess regulatory functions [34, 54]. In addition, it is believed that late apoptosis, with compromised cell membranes and delayed clearance by phagocytes, can lead to autoimmunity [55], and our study showed a reduced late apoptotic cell population. A down-regulated expression of genes related to lymphocyte apoptosis was also found [49]. Indeed, one study has shown that DAZ (added at 0.2 g/kg diet) possessed a similar protective effect against T1D (for both blood glucose level and glucose tolerance) as GEN in NOD mice [3]. The amount used here would be similar to our dose at 20 mg/kg BW [18, 56]. However, we did not observe any improvements in blood glucose and tolerance in the current study, which might be due to different routes of exposure (gavage vs food intake).
In conclusion, DAZ dosing reduced the B cell population as well as IgG1 production, while increased the helper and cytotoxic T cell population in females. The lack of protection or exacerbation of T1D can be explained by reduced B cell population and increased regulatory cells, revealing a completely different mechanism compared to GEN exposure. As for food intake, animals are exposed to a mix of compounds. Combinatorial exposure of GEN and DAZ in female BALB/c mice demonstrated a decreased IgG1 for mucosal immune response [57]. Therefore, future studies of the interaction between DAZ and GEN are warranted. Furthermore, to improve the bioavailability of DAZ and to enhance the potential beneficial effect, we may consider the use of DAZ metabolites in modulating T1D risk, which may require an in-depth understanding of host-microbiota interaction and a thorough investigation on the mode of action following DAZ exposure in both sexes.
Highlights:
Daidzein decreases B cells while increases T cells in female B6C3F1 mice
Daidzein decreases late apoptotic thymocytes in female B6C3F1 mice
Daidzein increases IgG2b in NOD females and IgG1 in NOD males
Daidzein increases CD8+CD25+ and B220+CD40L+ splenocytes in NOD females
Acknowledgments
This study was supported by the United Soybean Board (USB), and in part by the NIH R21ES24487, R41AT009523 and R21ES012286. The authors declare no conflict of interest.
Footnotes
Publisher's Disclaimer: This is a PDF file of an unedited manuscript that has been accepted for publication. As a service to our customers we are providing this early version of the manuscript. The manuscript will undergo copyediting, typesetting, and review of the resulting proof before it is published in its final citable form. Please note that during the production process errors may be discovered which could affect the content, and all legal disclaimers that apply to the journal pertain.
Reference:
- [1].Pandey KB, Rizvi SI. Plant polyphenols as dietary antioxidants in human health and disease. Oxidative medicine and cellular longevity. 2009;2(5):270–278. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [2].Cao Y, Calafat AM, Doerge DR, Umbach DM, Bernbaum JC, Twaddle NC, et al. Isoflavones in urine, saliva, and blood of infants: data from a pilot study on the estrogenic activity of soy formula. J Expo Sci Environ Epidemiol. 2009;19(2):223–234. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [3].Choi MS, Jung UJ, Yeo J, Kim MJ, Lee MK. Genistein and daidzein prevent diabetes onset by elevating insulin level and altering hepatic gluconeogenic and lipogenic enzyme activities in non-obese diabetic (NOD) mice. Diabetes-Metabolism Research and Reviews. 2008;24(1):74–81. [DOI] [PubMed] [Google Scholar]
- [4].Gilbert ER, Liu D. Anti-diabetic functions of soy isoflavone genistein: mechanisms underlying its effects on pancreatic beta-cell function. Food Funct. 2013;4(2):200–212. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [5].Ding M, Pan A, Manson J, Willett W, Malik V, Rosner B, et al. Consumption of soy foods and isoflavones and risk of type 2 diabetes: a pooled analysis of three US cohorts. European Journal of Clinical Nutrition. 2016. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [6].Nanri A, Mizoue T, Takahashi Y, Kirii K, Inoue M, Noda M, et al. Soy product and isoflavone intakes are associated with a lower risk of type 2 diabetes in overweight Japanese women. J Nutr. 2010;140(3):580–586. [DOI] [PubMed] [Google Scholar]
- [7].Huang G, Xu J, Guo TL. Exposure to Polyphenolic Compounds Modulates Type 1 Diabetes: The Case of Genistein Polyphenols: Mechanisms of Action in Human Health and Disease: Elsevier; 2018. p. 193–203. [Google Scholar]
- [8].Otokozawa S, Tanaka R, Akasaka H, Ito E, Asakura S, Ohnishi H, et al. Associations of Serum Isoflavone, Adiponectin and Insulin Levels with Risk for Epithelial Ovarian Cancer: Results of a Case-control Study. Asian Pac J Cancer Prev. 2015;16(12):4987–4991. [DOI] [PubMed] [Google Scholar]
- [9].Ding M, Franke AA, Rosner BA, Giovannucci E, van Dam RM, Tworoger SS, et al. Urinary isoflavonoids and risk of type 2 diabetes: a prospective investigation in US women. Br J Nutr. 2015;114(10):1694–1701. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [10].Yang Z, Kulkarni K, Zhu W, Hu M. Bioavailability and Pharmacokinetics of Genistein: Mechanistic Studies on its ADME. Anti-Cancer Agent Me. 2012;12(10):1264–1280. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [11].Hwang CS, Kwak HS, Lim HJ, Lee SH, Kang YS, Choe TB, et al. Isoflavone metabolites and their in vitro dual functions: they can act as an estrogenic agonist or antagonist depending on the estrogen concentration. The Journal of steroid biochemistry and molecular biology. 2006;101(4):246–253. [DOI] [PubMed] [Google Scholar]
- [12].Miller LM, Lampe JW, Newton KM, Gundersen G, Fuller S, Reed SD, et al. Being overweight or obese is associated with harboring a gut microbial community not capable of metabolizing the soy isoflavone daidzein to O-desmethylangolensin in peri-and post-menopausal women. Maturitas. 2017;99:37–42. [DOI] [PubMed] [Google Scholar]
- [13].Frankenfeld CL, Atkinson C, Wahala K, Lampe JW. Obesity prevalence in relation to gut microbial environments capable of producing equol or O-desmethylangolensin from the isoflavone daidzein. Eur J Clin Nutr. 2014;68(4):526–530. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [14].Reverri EJ, Slupsky CM, Mishchuk DO, Steinberg FM. Metabolomics reveals differences between three daidzein metabolizing phenotypes in adults with cardiometabolic risk factors. Molecular nutrition & food research. 2017;61(1). [DOI] [PubMed] [Google Scholar]
- [15].Hazim S, Curtis PJ, Schar MY, Ostertag LM, Kay CD, Minihane AM, et al. Acute benefits of the microbial-derived isoflavone metabolite equol on arterial stiffness in men prospectively recruited according to equol producer phenotype: a double-blind randomized controlled trial. Am J Clin Nutr. 2016;103(3):694–702. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [16].Rienks J, Barbaresko J, Nothlings U. Association of isoflavone biomarkers with risk of chronic disease and mortality: a systematic review and meta-analysis of observational studies. Nutr Rev. 2017;75(8):616–641. [DOI] [PubMed] [Google Scholar]
- [17].Khan D, Ansar Ahmed S. The immune system is a natural target for estrogen action: opposing effects of estrogen in two prototypical autoimmune diseases. Frontiers in immunology. 2016;6:635. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [18].Guo TL, Wang YB, Xiong T, Ling X, Zheng JF. Genistein modulation of streptozotocin diabetes in male B6C3F1 mice can be induced by diet. Toxicology and Applied Pharmacology. 2014;280(3):455–466. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [19].Dandona P, Aljada A, Bandyopadhyay A. Inflammation: the link between insulin resistance, obesity and diabetes. Trends in immunology. 2004;25(1):4–7. [DOI] [PubMed] [Google Scholar]
- [20].Kristiansen OP, Mandrup-Poulsen T. Interleukin-6 and diabetes: the good, the bad, or the indifferent? Diabetes. 2005;54 Suppl 2:S114–124. [DOI] [PubMed] [Google Scholar]
- [21].Curiel TJ, Coukos G, Zou L, Alvarez X, Cheng P, Mottram P, et al. Specific recruitment of regulatory T cells in ovarian carcinoma fosters immune privilege and predicts reduced survival. Nat Med. 2004;10(9):942–949. [DOI] [PubMed] [Google Scholar]
- [22].Liyanage UK, Moore TT, Joo H-G, Tanaka Y, Herrmann V, Doherty G, et al. Prevalence of regulatory T cells is increased in peripheral blood and tumor microenvironment of patients with pancreas or breast adenocarcinoma. The Journal of Immunology. 2002;169(5):2756–2761. [DOI] [PubMed] [Google Scholar]
- [23].Lin IC, Yamashita S, Murata M, Kumazoe M, Tachibana H. Equol suppresses inflammatory response and bone erosion due to rheumatoid arthritis in mice. Journal of Nutritional Biochemistry. 2016;32:101–106. [DOI] [PubMed] [Google Scholar]
- [24].Medjakovic S, Mueller M, Jungbauer A. Potential health-modulating effects of isoflavones and metabolites via activation of PPAR and AhR. Nutrients. 2010;2(3):241–279. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [25].Jahromi SR, Arrefhosseini SR, Ghaemi A, Alizadeh A, Tabriz HM, Togha M. Alleviation of Experimental Allergic Encephalomyelitis in C57BL/6 Mice by Soy Daidzein. Iran J Allergy Asthm. 2014;13(4):256–264. [PubMed] [Google Scholar]
- [26].Guo TL, Germolec DR, Zheng JF, Kooistra L, Auttachoat W, Smith MJ, et al. Genistein Protects Female Nonobese Diabetic Mice from Developing Type 1 Diabetes When Fed a Soy- and Alfalfa-free Diet. Toxicologic Pathology. 2015;43(3):435–448. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [27].Yashida MH, Da Silva Faria AL, Caldeira EJ. Estrogen and insulin replacement therapy modulates the expression of insulin-like growth factor-I receptors in the salivary glands of diabetic mice. Anat Rec (Hoboken). 2011;294(11):1930–1938. [DOI] [PubMed] [Google Scholar]
- [28].Gervais F, Attia MA. Fibro-osseous proliferation in the sternums and femurs of female B6C3F1, C57black and CD-1 mice: a comparative study. Dtsch Tierarztl Wochenschr. 2005;112(9):323–326. [PubMed] [Google Scholar]
- [29].Huang G, Xu J, Lefever DE, Glenn TC, Nagy T, Guo TL. Genistein prevention of hyperglycemia and improvement of glucose tolerance in adult non-obese diabetic mice are associated with alterations of gut microbiome and immune homeostasis. Toxicol Appl Pharmacol. 2017. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [30].Guo TL, Chi RP, Hernandez DM, Auttachoat W, Zheng JF. Decreased 7, 12-dimethylbenz [a] anthracene-induced carcinogenesis coincides with the induction of antitumor immunities in adult female B6C3F1 mice pretreated with genistein. Carcinogenesis. 2007;28(12):2560–2566. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [31].Guo TL, McCay JA, Zhang LX, Brown RD, You L, Karrow NA, et al. Genistein modulates immune responses and increases host resistance to B16F10 tumor in adult female B6C3F1 mice. The Journal of nutrition. 2001;131(12):3251–3258. [DOI] [PubMed] [Google Scholar]
- [32].Martin RM, Brady JL, Lew AM. The need for IgG2c specific antiserum when isotyping antibodies from C57BL/6 and NOD mice. Journal of immunological methods. 1998;212(2):187–192. [DOI] [PubMed] [Google Scholar]
- [33].Quintana FJ, Cohen IR. Autoantibody patterns in diabetes-prone NOD mice and in standard C57BL/6 mice. J Autoimmun. 2001;17(3):191–197. [DOI] [PubMed] [Google Scholar]
- [34].Churlaud G, Pitoiset F, Jebbawi F, Lorenzon R, Bellier B, Rosenzwajg M, et al. Human and mouse CD8+ CD25+ FOXP3+ regulatory T cells at steady state and during interleukin-2 therapy. Frontiers in immunology. 2015;6:171. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [35].Park MH, Ju JW, Park MJ, Han JS. Daidzein inhibits carbohydrate digestive enzymes in vitro and alleviates postprandial hyperglycemia in diabetic mice. Eur J Pharmacol. 2013;712(1-3):48–52. [DOI] [PubMed] [Google Scholar]
- [36].Cheong SH, Furuhashi K, Ito K, Nagaoka M, Yonezawa T, Miura Y, et al. Daidzein promotes glucose uptake through glucose transporter 4 translocation to plasma membrane in L6 myocytes and improves glucose homeostasis in Type 2 diabetic model mice. The Journal of nutritional biochemistry. 2014;25(2): 136–143. [DOI] [PubMed] [Google Scholar]
- [37].Claussnitzer M, Skurk T, Hauner H, Daniel H, Rist MJ. Effect of flavonoids on basal and insulin-stimulated 2-deoxyglucose uptake in adipocytes. Molecular Nutrition & Food Research. 2011;55:S26–S34. [DOI] [PubMed] [Google Scholar]
- [38].Roghani M, Vaez Mahdavi MR, Jalali-Nadoushan MR, Baluchnejadmojarad T, Naderi G, Roghani-Dehkordi F, et al. Chronic Administration of Daidzein, a Soybean Isoflavone, Improves Endothelial Dysfunction and Attenuates Oxidative Stress in Streptozotocin-induced Diabetic Rats. Phytotherapy Research. 2013;27(1): 112–117. [DOI] [PubMed] [Google Scholar]
- [39].Cho KW, Lee OH, Banz WJ, Moustaid-Moussa N, Shay NF, Kim YC. Daidzein and the daidzein metabolite, equol, enhance adipocyte differentiation and PPARgamma transcriptional activity. J Nutr Biochem. 2010;21(9):841–847. [DOI] [PubMed] [Google Scholar]
- [40].Xu SZ, Zhong W, Ghavideldarestani M, Saurabh R, Lindow SW, Atkin SL. Multiple mechanisms of soy isoflavones against oxidative stress-induced endothelium injury. Free Radic Biol Med. 2009;47(2): 167–175. [DOI] [PubMed] [Google Scholar]
- [41].Kojima H, Takeda Y, Muromoto R, Takahashi M, Hirao T, Takeuchi S, et al. Isoflavones enhance interleukin-17 gene expression via retinoic acid receptor-related orphan receptors alpha and gamma. Toxicology. 2015;329:32–39. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [42].Piskula MK, Yamakoshi J, Iwai Y. Daidzein and genistein but not their glucosides are absorbed from the rat stomach. FEBS letters. 1999;447(2-3):287–291. [DOI] [PubMed] [Google Scholar]
- [43].Hillman GG, Wang Y, Che MX, Raffoul JJ, Yudelev M, Kucuk O, et al. Progression of renal cell carcinoma is inhibited by genistein and radiation in an orthotopic model. Bmc Cancer. 2007;7. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [44].King RA, Broadbent JL, Head RJ. Absorption and excretion of the soy isoflavone genistein in rats. The Journal of nutrition. 1996; 126(1): 176–182. [DOI] [PubMed] [Google Scholar]
- [45].Zhang R, Li Y, Wang W. Enhancement of immune function in mice fed high doses of soy daidzein. Nutr Cancer. 1997;29(l):24–28. [DOI] [PubMed] [Google Scholar]
- [46].Vandenberg LN. Low-dose effects of hormones and endocrine disruptors Vitamins & Hormones. 94: Elsevier; 2014. p. 129–165. [DOI] [PubMed] [Google Scholar]
- [47].Yum MK, Jung MY, Cho D, Kim TS. Suppression of dendritic cells’ maturation and functions by daidzein, a phytoestrogen. Toxicology and applied pharmacology. 2011;257(2): 174–181. [DOI] [PubMed] [Google Scholar]
- [48].Feng G, Sun B, Li TZ. Daidzein attenuates lipopolysaccharide-induced acute lung injury via toll-like receptor 4/NF-kappaB pathway. Int Immunopharmacol. 2015;26(2):392–400. [DOI] [PubMed] [Google Scholar]
- [49].Fan H, Lv Z, Gan L, Guo Y. Transcriptomics-Related Mechanisms of Supplementing Laying Broiler Breeder Hens with Dietary Daidzein to Improve the Immune Function and Growth Performance of Offspring. J Agric Food Chem. 2018;66(8):2049–2060. [DOI] [PubMed] [Google Scholar]
- [50].Liu DY, He SJ, Liu SQ, Tang YG, Jin EH, Chen HL, et al. Daidzein enhances immune function in late lactation cows under heat stress. Animal Science Journal. 2014;85(l):85–89. [DOI] [PubMed] [Google Scholar]
- [51].Hampe CS. B cells in autoimmune diseases. Scientifica. 2012;2012. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [52].Kawamura T, Kanai T, Dohi T, Uraushihara K, Totsuka T, Iiyama R, et al. Ectopic CD40 ligand expression on B cells triggers intestinal inflammation. J Immunol. 2004;172(10):6388–6397. [DOI] [PubMed] [Google Scholar]
- [53].Hulbert C, Riseili B, Rojas M, Thomas JW. B cell specificity contributes to the outcome of diabetes in nonobese diabetic mice. J Immunol. 2001;167(10):5535–5538. [DOI] [PubMed] [Google Scholar]
- [54].Tachikawa S, Kawamura T, Kawamura H, Kanda Y, Fujii Y, Matsumoto H, et al. Appearance of B2201ow autoantibody-producing B-1 cells at neonatal and older stages in mice. Clinical & Experimental Immunology. 2008;153(3):448–455. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [55].Patel VA, Longacre A, Hsiao K, Fan HL, Meng F, Mitchell JE, et al. Apoptotic cells, at all stages of the death process, trigger characteristic signaling events that are divergent from and dominant over those triggered by necrotic cells - Implications for the delayed clearance model of autoimmunity. Journal of Biological Chemistry. 2006;281(8):4663–4670. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [56].Sepehr E, Cooke G, Robertson P, Gilani GS. Bioavailability of soy isoflavones in rats Part I: application of accurate methodology for studying the effects of gender and source of isoflavones. Molecular nutrition & food research. 2007;51(7):799–812. [DOI] [PubMed] [Google Scholar]
- [57].Wei J, Bhatt S, Chang LM, Sampson HA, Masilamani M. Isoflavones, Genistein and Daidzein, Regulate Mucosal Immune Response by Suppressing Dendritic Cell Function. Plos One. 2012;7(10). [DOI] [PMC free article] [PubMed] [Google Scholar]